

Base Excision DNA Repair in Plants: Arabidopsis and Beyond
by
 ,
,
Inga R. Grin
1,2,*,
Daria V. Petrova
1,
Anton V. Endutkin
1,
Chunquan Ma
3,4,5,
Bing Yu
3,4,5,
Haiying Li
3,4,5 and
Dmitry O. Zharkov
1,2,* 1
Siberian Branch of the Russian Academy of Sciences Institute of Chemical Biology and Fundamental Medicine, 8 Lavrentieva Ave., Novosibirsk 630090, Russia
2
Department of Natural Sciences, Novosibirsk State University, 2 Pirogova St., Novosibirsk 630090, Russia
3
Engineering Research Center of Agricultural Microbiology Technology, Ministry of Education, Harbin 150080, China
4
Heilongjiang Provincial Key Laboratory of Plant Genetic Engineering and Biological Fermentation Engineering for Cold Region, Harbin 150080, China
5
School of Life Sciences, Heilongjiang University, Harbin 150080, China
*
Authors to whom correspondence should be addressed.
Int. J. Mol. Sci. 2023, 24(19), 14746; https://doi.org/10.3390/ijms241914746
Submission received: 4 September 2023
/
Revised: 27 September 2023
/
Accepted: 27 September 2023
/
Published: 29 September 2023
(This article belongs to the Special Issue Molecular Mechanism of DNA Replication and Repair, 2nd Edition )
Abstract
:Base excision DNA repair (BER) is a key pathway safeguarding the genome of all living organisms from damage caused by both intrinsic and environmental factors. Most present knowledge about BER comes from studies of human cells, E. coli, and yeast. Plants may be under an even heavier DNA damage threat from abiotic stress, reactive oxygen species leaking from the photosynthetic system, and reactive secondary metabolites. In general, BER in plant species is similar to that in humans and model organisms, but several important details are specific to plants. Here, we review the current state of knowledge about BER in plants, with special attention paid to its unique features, such as the existence of active epigenetic demethylation based on the BER machinery, the unexplained diversity of alkylation damage repair enzymes, and the differences in the processing of abasic sites that appear either spontaneously or are generated as BER intermediates. Understanding the biochemistry of plant DNA repair, especially in species other than the Arabidopsis model, is important for future efforts to develop new crop varieties.
1. Introduction: BER Essentials
DNA repair is a vital process in all living organisms [1]. Repair is required to restore the integrity of DNA after damage that, in human cells, occurs at the level of tens to hundreds of thousands of events per cell, per day [2,3]. The most important sources of DNA damage are endogenous, including oxidative metabolism, spontaneous hydrolytic reactions, and replication machinery errors, whereas environmental factors, such as ionizing and UV radiation, and genotoxic biogenic or industrial chemicals, also contribute significantly [1]. In humans and higher animals, inactivation of repair systems is lethal or causes severe morbidity [1]. The accumulation of damage not eliminated by DNA repair, in many cases, leads to mutations and is among the primary causes of aging, carcinogenesis, and some neurodegenerative and autoimmune diseases [4,5].
The vast majority of intrinsic DNA lesions (products of spontaneous deamination, hydrolysis, oxidation, and metabolic-related alkylation) are dealt with by a system known as base excision repair (BER) [6,7,8]. The current model of BER (Figure 1) is mostly derived from studies in mammalian cells, with significant input from E. coli and yeast systems. If base damage is present in the genome, it is recognized and excised by DNA glycosylases, lesion-specific enzymes that hydrolyze the N-glycosidic bond in nucleotides. Eleven DNA glycosylases with different substrate specificities are presently known in humans, and eight in E. coli (Table 1), while some bacteria and viruses possess a few additional DNA glycosylases absent from these two species. Some DNA glycosylases only cleave the N-glycosidic bond, leaving a baseless 2′-deoxyribose (apurinic/apyrimidinic site, or AP site) in DNA (Figure 2). Others, termed DNA glycosylases/AP lyases, can nick DNA 3′ to the nascent AP site by β-elimination, leaving an α,β-unsaturated aldehyde at the 3′-end of DNA at the break. A subset of those (β,δ-lyases) proceeds even further, catalyzing the removal of the whole sugar fragment and leaving a single-nucleotide gap flanked by two phosphate groups (Figure 2). All DNA glycosylases/AP lyases share a common reaction mechanism and form a covalent Schiff base intermediate with C1′ of the damaged nucleotide in the reaction. Both β- and β,δ-elimination products at the 3′-end cannot be extended by DNA polymerases and need additional processing to expose a 3′-OH group. Thus, a second BER step involves hydrolytic enzymes that nick DNA at a phosphate 5′ to an AP site, or an unsaturated aldehyde, or a 3′-phosphate (Figure 2). The respective activities are referred to as AP endonuclease, 3′-phosphodiesterase, or 3′-phosphatase, although they usually reside within the same protein, which therefore can continue the process after any DNA glycosylase (Table 1). A notable exception is human APE1, which lacks a 3′-phosphatase activity and, thus, polynucleotide kinase/3′-phosphatase (PNKP) substitutes for APE1 after the action of human β,δ-lyases (NEIL1, NEIL2, and NEIL3).
After a free 3′-OH end is formed, a DNA polymerase can incorporate an undamaged dNTP complementary to the base in the template (Figure 1). In human cells, DNA polymerase β (POLβ) is the principal BER polymerase, although DNA polymerase λ (POLλ), the main non-homologous end joining polymerase, can be involved in specific cases, and DNA polymerase γ (POLγ) does the job in mitochondria. If the repair started with a monofunctional DNA glycosylase followed by an AP endonuclease, a 2′-deoxyribo-5′-phosphate (dRP) remnant lingers at this point at the 5′-end of DNA downstream of the nick, and POLβ polymerase activity is coupled with a dRP lyase activity that removes this moiety. One or several nucleotides can be incorporated (short-patch or long-patch BER, respectively; Figure 1), with a possible switch from POLβ to DNA polymerases δ or ε assisted by the PCNA processivity factor. If several nucleotides are replaced, the hanging flap is clipped by flap endonuclease, FEN1. In E. coli, DNA polymerase I is the major BER polymerase, and its 5′→3′ exonuclease activity degrades the downstream fragment. Finally, DNA ligase (LigI or LigIIIα in human cells, LigA in E. coli) seals the last nick restoring the integrity of DNA (Figure 1).
2. DNA Damage Specifics in Plants
Compared with animals, and probably more in common with bacteria, plants are exposed to an even wider variety of genotoxic stresses. Abiotic factors, such as light regime, temperature, and water availability, have a stronger impact on the biochemistry of plant cells compared with multicellular animals. While mitochondria are the sole major source of reactive oxygen species (ROS) in animal cells, chloroplasts generate an abundant additional ROS flow in plants [9,10]. Last, but not least, plants produce a wide variety of genotoxic secondary metabolites, of which psoralens, topoisomerase inhibitors (camptothecins, podophyllotoxins), and aristolochic acid are probably the best-studied examples [11,12,13,14,15]. Moreover, some widespread plant protein toxins, such as ribosome-inactivating proteins, also damage DNA on top of their primary mode of action [16]. To protect against abiotic stress associated with environmental factors, different repair systems operate in different plant organs, and the consequences caused by their deficiency can vary from a decrease in the stability of the seed genome to a decrease in the yield of adult plants [17,18].
Abiotic stress plays a critical role in plant life and is one of the main factors limiting crop productivity. Different types of abiotic stress can have a damaging effect on the genome of plant cells, both directly and through the production of reactive secondary metabolites and signaling molecules [19]. BER activation, double-strand break response, and nucleotide pool sanitization are essential elements of the abiotic stress response in plants [20,21,22]. Information on the role of plant BER in response to abiotic stresses other than known DNA damaging factors (such as ionizing radiation, UV radiation, or exposure to genotoxic compounds) is extremely limited, with the exception of BER-dependent epigenetic regulation [23,24]. However, some observations indicate that, even apart from the epigenetic mechanism, BER is indeed involved in protecting plants from thermal and osmotic stress [25,26,27,28]. This may be due, in part, to the fact that many types of abiotic stress lead to increased leakage of DNA-damaging reactive oxygen species from chloroplasts, and DNA repair may be needed to protect cells from this damage [29,30]. Characterization of the role of BER and other types of DNA repair in plant responses to abiotic stress is important for understanding the mechanisms of plant adaptation to adverse environmental conditions.
3. Plant DNA Glycosylases: The Universal Themes
3.1. Uracil–DNA Glycosylases
Uracil–DNA glycosylase (Ung in bacteria, UNG in humans) was the first DNA glycosylase discovered and studied with respect to its mechanism [31,32]. Several other DNA glycosylases have U as their target in DNA, namely mismatched uracil–DNA glycosylase (Mug) in E. coli, G/T mismatch-specific thymine DNA glycosylase (TDG), single-strand-selective monofunctional uracil-DNA glycosylase 1 (SMUG1), and methyl-CpG binding domain 4-containing DNA glycosylase (MBD4) in humans. Of these, Ung/UNG and SMUG1 are general-purpose repair glycosylases removing U from any DNA context, MBD4 is specialized for U repair in damaged CpG dinucleotides, and TDG mainly participates in active demethylation rather than in DNA repair [33,34]. With the exception of MBD4, all uracil–DNA glycosylases belong to the same structural superfamily, sharing the α/β-fold and a common evolutionary origin [35].
The first plant protein with the uracil–DNA glycosylase (or, indeed, any DNA glycosylase) activity was purified in 1983 from wheat germ and showed properties fairly typical for E. coli and human UNG [36]. Enzymatic activity removing U from DNA was detected in living cells or cell extracts from a number of phylogenetically diverged plant species [37,38,39,40]. A. thaliana cell extracts support the full BER cycle for U with base excision, AP site processing, dNMP incorporation, and nick ligation [41,42]. The same capability was demonstrated for a single-cell green alga, Chlamydomonas reinhardtii [43].
Cloned recombinant AtUNG removes U from both single- and double-stranded DNA, with a moderate preference for U:G pairs [42]. AtUNG seems to be the only enzyme capable of U excision in Arabidopsis since the UNG gene knockout totally eliminates the activity in cell extracts and enhances resistance to 5-fluorouracil, a chemical that promotes DNA instability via excessive U accumulation and excision [42]. Paradoxically, however, ung plants do not accumulate the expected C→T mutations, suggesting the presence of alternative repair mechanisms [44].
Genome sequencing reveals no orthologs of human TDG and SMUG1 in flowering plants, although a TDG homolog is found in green algae and a SMUG1 homolog is present in mosses, ferns, horsetails, and gymnosperms. At the sequence level, they are more closely related to bacterial rather than to metazoan homologs, indicating possible horizontal transfer events. None of these homologs are functionally characterized.
MBD4 in vertebrates, first discovered as a protein containing an MeCP2-like mC-binding domain, is thought to counteract deamination specifically in CpG dinucleotides [45,46,47]. The name of the plant homolog is somewhat of a misnomer, since plant MBD4 contains no methyl-binding domain [48,49]. However, the overall organization of AtMBD4 and hMBD4 is similar, with the C-terminal catalytic domain and an extended N-terminal part, with the catalytic residues fully conserved [48,49]. AtMBD4 excises U and, less efficiently, T mispaired with G, and also processes 5-BrU, 5-FU, 5-hydroxymethyluracil, and 5-hydroxyuracil, with a modest twofold preference for a 5′-XG-3′ context and no methylation status preference [48]. Since CG, CHG, and CHH sites are all widely used for methylation in plants, these properties are consistent with a view of AtMBD4 as a guardian of methylation sites against C deamination. Interestingly, truncation of the N-terminal part, leaving only the catalytic domain, improves the kinetics of AtMBD4 [48]. This finding may be of biological relevance, since of several functional AtMBD4 isoforms detected in vivo at the mRNA level, one splices out a cryptic intron and lacks a 116 amino acid fragment from the N-terminal part [49,50]. This protein isoform is directed to the nucleolus, whereas the longer one is evenly distributed in the nucleus, and the alternative splicing and relocalization is heat induced, underscoring the role of plant MBD4 in the abiotic stress response [50].
3.2. Oxidative Damage Repair
Oxidative DNA base damage presents the largest group of chemically diverse lesions, unavoidable in living organisms with an oxygen-based metabolism [51,52]. Most of them are cytotoxic, but some are miscoding; a textbook example of the latter is 8-oxoguanine (8-oxoG), which can pair with either C or A during replication and cause G→T transversions in the latter case [53]. High frequency and mutagenicity 8-oxoG necessitate a dedicated system for its repair, the so-called GO system, which consists of three enzymes, namely 8-oxodGTPase MutT/MTH1 (in E. coli and humans, respectively) that sanitizes the dNTP pool and prevent 8-oxoG incorporation by DNA polymerases, and two DNA glycosylases, Fpg/OGG1 and MutY/MUTYH (Table 1). Fpg and OGG1 are entirely different with respect to their sequence and structure, yet their substrate specificities are almost identical: they remove 8-oxoG from pairs with C and strongly discriminate against 8-oxoG:A substrates. As such, mispairs are most often formed when replicative DNA polymerases incorporate A opposite to 8-oxoG, an excision of 8-oxoG would replace it with T and finalize the mutation. So instead of Fpg/OGG1, 8-oxoG:A is recognized by MutY/MUTYH, which remove A and give repair polymerases (PolI in E. coli, POLλ in humans) a chance to incorporate C opposite to 8-oxoG and hand the baton to Fpg/OGG1 for the second round of repair (reviewed in [53,54]). In addition to 8-oxoG, Fpg and OGG1 excise formamidopyrimidines (Fapy), the products of hydroxyl radical attack at A or G, followed by reduction. The repair of oxidatively damaged pyrimidines is less complicated and proceeds essentially according to the general BER scheme; the involved enzymes are Nth and Nei in E. coli and NTHL1, NEIL1, NEIL2, and NEIL3 in human cells. One important distinction regarding the repair mechanism is that MutY/MUTYH, OGG1, and Nth/NTHL1 belong to the helix–hairpin–helix (HhH) structural superfamily and are either monofunctional (MutY/MUTYH) or catalyze β-elimination (OGG1 and Nth/NTHL1), whereas Fpg, Nei, and NEIL1–NEIL3 come from the helix–two-turn–helix (H2TH) structural superfamily and catalyze β,δ-elimination [55,56].
Plants possess a collection of all glycosylases for oxidative damage repair. A homolog of bacterial Fpg was cloned from an A. thaliana cDNA library based on sequence homology and termed MMH-1 (MutM Homolog 1; historically, MutM is an alternative name for Fpg) [57]. The primary transcript is alternatively spliced producing two major and two minor protein isoforms that apparently keep all catalytic residues in common but diverge in the C-terminal part [57,58,59]. The substrate preference of AtMMH-1, as characterized initially, is reminiscent of Fpg and OGG1 with its ability to excise 8-oxoG and discriminate against A opposite to the lesion [57], and both AtOGG1 (see below) and AtMMH-1 (isoform 1) suppress characteristic 8-oxoG-induced G→T transition in a reporter fpg E. coli strain [60]. Interestingly, when compared side by side, purified AtMMH-1 is much less active in the removal of 8-oxoG than AtOGG1 [61]; however, 8-oxoG repair activities in MMH-1 and OGG1 knockout cell extracts are affected to about an equal degree [26]. Further studies with oligonucleotide and irradiated DNA substrates showed excellent activity by AtMMH-1 against Fapy bases and products of four-electron G oxidation (guanidinohydantoin and spiroiminodihydantoin), but only minor capability for 8-oxoG removal [62]. The structural reason for this is, apparently, the lack of the “capping loop” in the C-terminal domain, which partly forms an 8-oxoG recognition pocket in bacterial Fpg but is much shorter in AtMMH-1 [63]. Notably, the shorter major AtMMH-1 isoform deviates from the longer isoform, starting immediately before this loop and does not rescue the fpg phenotype when expressed in E. coli, raising questions about the substrate specificity of this protein variant [60].
Besides A. thaliana, MMH homologs were partly characterized in sugarcane [64,65] and barrel clover (Medicago truncatula) [66]. The sugarcane genome encodes two MMH homologs that are highly conserved (but not identical) in their N-terminal domain and in the C-terminal domain up to the H2TH motif but diverge thereafter; both are ubiquitously expressed and were shown to decrease mutations in the fpg mutY E. coli reporter strain [65]. The barrel clover protein was not characterized but, together with OGG1, is upregulated in response to heavy metals and osmotic stress and during seed imbibition, all known to induce oxidative stress [66]. In a related species, alfalfa (M. sativa), MMH is upregulated in response to cell exposure to quantum dots, which apparently also cause oxidative stress [67].
After a long history of failures to find eukaryotic orthologs of bacterial Fpg in a pre-genomic era, in 1996, a non-homologous 8-oxoguanine glycosylase, OGG1, was cloned from yeast based on the ability to complement the mutator phenotype of fpg mutY-deficient E. coli [68] or cross-link specifically to an 8-oxoG:C duplex [69]. The human [70,71,72] and A. thaliana orthologs [73,74] followed soon. As the yeast and human proteins, AtOGG1 excises 8-oxoG via a Schiff base intermediate, preferring 8-oxoG:C over 8-oxoG:A pairs, and is capable of rescuing the mutator phenotype of fpg or fpg mutY-deficient E. coli [60,73,74,75]. Besides 8-oxoG, AtOGG1 can remove FapyG, but not other oxidative lesions generated by γ-irradiation of DNA [75]. The AtOGG1 gene is universally expressed and is induced neither by oxidative stress nor increased 8-oxodGTP caused by a deficiency of AtNUDX1, the only 8-oxodGTPase in A. thaliana [73,74,76]. Interestingly, unlike human OGG1, which undergoes extensive alternative splicing and generates 13 protein isoforms [77], only one mRNA and protein are known for AtOGG1.
Transgenic A. thaliana plants overexpressing OGG1 show increased seed longevity and resistance to osmotic and heat stress [25]. OGG1 is upregulated in response to the conditions known to induce oxidative stress, such as heavy metals and osmotic stress, and histone deacetylase inhibitor trichostatin A in barrel clover, as well as during seed imbibition in A. thaliana, barrel clover, and eggplant [25,66,78,79].
Together, MMH-1 and OGG1 account for the majority of 8-oxoG repair in A. thaliana, but some residual activity is still observed in the double knockouts [26]. Its nature remains to be established. The gene that is the last member of the GO system, MutY, is present in plants, but has not been characterized so far.
As for oxidized pyrimidine repair, A. thaliana possesses two Nth homologs, NTH1 and NTH2 [80,81], which are rather poorly studied at present. Recombinant NTH1 and NTH2 are bifunctional DNA glycosylases that efficiently release thymine glycol and urea residues from DNA and cleave the backbone by β-elimination. The knockout of both genes has no overt phenotype [81].
4. Epigenetic Glycosylases, the Plants’ Unique Toolkit
Gene expression in higher eukaryotes is controlled through several mechanisms, of which epigenetic DNA methylation is especially prominent. In most animals and plants, it relies on a “fifth nucleobase”, 5-methylcytosine (mC), serving as a reversible mark that plays an important role in transcription regulation and genome protection from mobile elements [82,83]. Vertebrates overwhelmingly use CpG dinucleotides for this purpose, whereas plants are more flexible, methylating C in CG, CHG, and CHH contexts (H = A, C, or T). In all species that use mC residues as epigenetic marks, they are set on by DNA (cytosine-5)-methyltransferases that either methylate DNA de novo or reinstall mC in hemimethylated DNA. However, the opposite-direction process, active demethylation, employs entirely different mechanisms in animals and plants, albeit both based on BER. In animals, mC is oxidized by TET family dioxygenases first to 5-hydroxymethylcytosine, an epigenetic mark itself, and further to 5-formylcytosine and 5-carboxycytosine; all these derivatives can also be actively deaminated to 5-hydroxymethyluracil, 5-formyluracil, and 5-carboxyuracil, respectively. The oxidized/deaminated bases are excised by TDG and replaced with C through the BER pathway [84,85]. Plants, in contrast, possess DNA glycosylases that directly recognize and excise mC. In 2002, two paralogous genes involved in the epigenetic control in A. thaliana were found to encode DNA glycosylases: DEMETER (DME), a gene responsible for maternal allele derepression of MEDEA Polycomb group chromatin remodeler during embryo and endosperm development, and REPRESSOR OF SILENCING 1 (ROS1, also known as DEMETER-LIKE 1, DML1), an activator of transcription of several endogenous genes acting throughout the life cycle [86,87]. Two more paralogs, termed DEMETER-LIKE 2 (DML2) and DEMETER-LIKE 3 (DML3), were characterized shortly thereafter [86,88,89]. There is a fifth paralogous gene residing on chromosome 3 (At3g47830), which remains uncharacterized, but its predicted protein product lacks several critical functional domains (see below) and may be non-functional. Finally, the analysis of sequenced plant genomes reveals another cluster of homologs (labeled DMLX in Figure 3), distributed in the major groups of dicots. They keep all functional domains of DML proteins but differ in their long, disordered N-terminal tails. No representative of this branch has been studied so far.
A word about the nomenclature is prudent here, since DME-like genes/proteins from different species often bear the name “DML” or “ROS” followed by a number, but are most similar to differently numbered DME-like proteins from A. thaliana. DME is the first discovered member of this small family. ROS1 is systematically known as DML1 and can be found under this name in some papers; however, here we keep the name ROS1 to be consistent with the bulk of the literature. DML2 and DML3 in A. thaliana have been so named from the moment of their discovery [86]. However, in tobacco, for example, four demethylating glycosylases were described as ROS1, ROS2a, ROS2b, and ROS3, all being closer to AtROS1 than to other DML homologs from A. thaliana [95], and of three predicted genes more similar to DME, DML2, and DML3, two are designated “DEMETER-like”, and one, “ROS1-LIKE”. Also, with the expanded number of sequenced plant genomes, it became apparent that true DME homologs are present only in dicots, whereas all homologs from monocots should be grouped with DML3, which appears to be ancestral to the whole family ([96,97] and Figure 3). Therefore, “DME” and “ROS1” proteins in wheat, barley, and rice [98,99,100,101,102] should actually be DML3, but we will keep the original names (in quotation marks) to be consistent with the other publications. We will further refer to the whole group as DML proteins or DML glycosylases.
DME and ROS1 are 5-methylcytosine–DNA glycosylases with a strong preference for mCG dinucleotides [103,104]. Some activity in mCHG and mCHH contexts is also observed, consistent with the known methylation pattern in plants. They can excise T with much lower efficiency, but have no activity on U bases [103]. Both DME and ROS1 possess a Lys–Asp catalytic dyad characteristic of bifunctional HhH superfamily DNA glycosylases and cleave DNA by β- and β,δ-elimination forming a Schiff base intermediate in the process [103,104,105]. Besides DME and ROS1 from A. thaliana, recombinant ROS1 from tobacco was partially biochemically characterized [95,106,107].
All DML proteins possess a unique split HhH catalytic domain, and an iron–sulfur cluster, a solitary zinc finger, and a putative RNA recognition motif-containing (RRM) domain [108,109]. They are all grouped in the C-terminal part of the protein, while the N-terminal part shows no homology to other known proteins and is highly variable even within DML proteins. The catalytic domain carries a large insertion of variable length (several hundred residues) between the αB and αC helices of the six-helix barrel domain, but the arrangement of the residues important for catalysis is conserved between DMLs and other HhH glycosylases, such as Nth [109,110]. Interestingly, the N-terminal domain of AtDME is dispensable for correct demethylation of canonical functional gene targets, but its removal impedes demethylation in heterochromatin and causes demethylation of non-specific gene body targets [111].
Of all DML proteins, ROS1 is probably the best investigated in terms of its enzymatic properties. In addition to mC and T, it efficiently removes 5-hydroxyuracil and 5-hydroxymethylcytosine, has moderate activity on 5-fluorouracil, 5-bromocytosine, and 5-bromouracil, and possesses only marginal ability to excise 5-hydroxymethyluracil [105,107,112]. The high activity against 5-hmC is especially intriguing since plants have long been believed to lack this modification in their genomes, but recent advances in mass spectroscopic detection and single-base resolution sequencing revealed its presence in DNA from several plant species [113,114,115]. AtDME and AtDML3 can also excise 5-hmC [116]. With respect to the strand methylation status, ROS1 utilizes both hemimethylated and bimethylated substrates but prefers the former, which may be important to avoid double-strand breaks [105,107]. Damage to the opposite G base adversely affects mC excision, which again may be important to prevent mutagenesis by misincorporation during BER [106]. The enzyme forcibly opens mC:G pairs using a trio of residues (Gln607, Arg903, and Met905) that intercalate into the DNA helix [117]. The iron–sulfur cluster is also necessary for ROS1 and DME activity, and cluster-disrupting mutations, chemical oxidation, or inactivation of the cellular cluster assembly pathway render these enzymes non-functional [118,119,120,121,122]. The RRM motif is essential for DNA binding and mC removal in vitro and male fertility in vivo, despite being missing in all other HhH glycosylases [97,123]. Like many HhH glycosylases, ROS1 is a slow-turnover enzyme that binds its AP site product with high affinity [105]. The search for mC by ROS1 in long DNA targets proceeds through non-specific DNA binding and one-dimensional diffusion along DNA, a behavior characteristic of many DNA glycosylases, and depends on the presence of the long N-terminal domain [117,124,125]. ROS1 is sumoylated by SIZ1 SUMO E3 ligase, which stabilizes ROS1 and enhances demethylation of many of its targets [126]. In vivo, ROS1 appears to function as part of a multiprotein complex, also involving a WD40 domain-containing protein RWD40, a methyl-DNA binding protein RMB1, and a zinc finger and homeobox domain protein RHD1 [127]. ROS1 is anchored to the complex through the interaction of its N-terminal tail with RWD40, whereas RMB1 acts as a mC reader targeting the complex to methylated genome regions [127].
Despite the presence of RRM domains, it is not clear whether DML proteins are guided to their site of action by any RNA. In vivo, the proper targeting of ROS1 depends on ROS3, a small RNA-binding protein, but physical interaction between ROS1 and ROS3 has not been demonstrated [128]. Demethylation by DMLs requires the glycosylase interaction with the H1 linker histone and prior chromatin opening by the MBD7 methyl-CpG-binding protein and IDM1/IDM2 histone H3 acetyltransferases [129,130,131,132]. ROS1 directly binds H3 histone through the RRM and this interaction is regulated by specific histone tail modifications in vitro [123], whereas in chromatin, ROS1 targets are enriched in H3K18ac and H3K27me3, and depleted of H3K9me2 and H3K27me1 [133].
The biochemistry of DML2 and DML3 has received much less attention, but both enzymes seem to be similar to DME and ROS1 in terms of the substrate specificity and reaction mechanism, although DML2 lacks thymine-excising activity [88,89]. It remains to be seen whether they have any specialized roles.
DME, so far, seems to act only in the regulation of plant development. Unlike mammals, in which a global wave of demethylation occurs immediately after fertilization and then new mC marks are laid out, plants experience more local methylation reconfiguration in the early zygote [134,135]. DME supports temporally and spatially restricted expression of maternal origin alleles normally silenced in the rest of the plant and in other developmental phases, such as MEA, FLOWERING WAGENINGEN (FWA), FERTILIZATION INDEPENDENT SEED 2 (FIS2), or FLOWERING LOCUS C (FLC) in A. thaliana [88,136,137,138,139]. DME also accounts for development-associated demethylation and activation of many mobile elements [140]. Both DME and ROS1 are required for proper pollen tube growth and orientation regulating a network of pollen tube signaling through active demethylation and gene activation [141]. In vegetative tissues, the only known role of DME is in regulation of pathogen defense genes [142], which is in common with ROS1, DML2, and DML3 [143].
Other DML proteins are also partially involved in development regulation. For example, in wheat (eleven homologs), barley (three homologs), and rice (six homologs), “DME” and “ROS1a” derepress genes coding for prolamins, the main storage proteins, during seed development [99,102]. In rice, both paternal and maternal loss-of-function alleles of “ROS1a” are lethal [98], and at least three DML glycosylases participate in demethylation in the gametes and the zygote [101]. In sorghum, two DML homologs are markedly upregulated in callus cultures, indicating the involvement of active demethylation in cell (de)differentiation processes [144]. The efficiency of organ regeneration from explants was shown to correlate with DME, ROS1, and DML2 levels in rock violet (Boea hygrometrica) [145].
ROS1, DML2, and DML3 primarily regulate gene expression in somatic tissues. Mutant ros1 A. thaliana plants show a dramatic increase in methylation of some endogenous promoters, such as RD29A, FWA, or EPF2, while overexpression of ROS1 induces their demethylation and activation [104,146,147]. Importantly, ROS1 activity is required for methylation and suppression of transposons of several types (GP1, LINE1-4, SN1, MU1) in plant cells [146], and ROS1/DML2/DML3-targeted regions are enriched in repeats that are considered the remnants of transposons [148]. One of the DML proteins in rice has been shown to demethylate and activate the Tos17 retrotransposon, a Copia family element widely used for experimental insertion mutagenesis in rice [149]. Rice DMLs also participate in differential methylation during generative cells production and early plant development [150] and are upregulated by heavy metals stress [151]. The tomato genome encodes four DML proteins, one of which, most homologous to ROS1, is upregulated in ripening fruit, and its suppression delays ripening due to the hypermethylation of promoters of several genes involved in the process [152,153]. The upregulation of DML proteins correlating with methylation changes has been demonstrated in rice for spaceflight [154]. A. thaliana requires ROS1, DML2, and DML3 to mount an immune response to Pseudomonas syringae infection [143], while in potato, ROS1 is elevated in response to β-aminobutyric acid, a pathogen signaling molecule that primes plant cells to fungal infection [155].
Despite an apparently wider range of processes and targets in comparison with DME, the methylation control by ROS1 seems to be limited to a few loci. Although ROS1/DML2/DML3-targeted regions contain many methylation-directing siRNA targets, at least ROS1 seems to remove methyl labels whose origin is not limited to RNAi-dependent pathways [148,156]. Strikingly, the global methylation level is not changed even in triple ros1 dml2 dml3 mutants [88]. Bisulfite sequencing reveals that ROS1, DML2, and DML3 display a degree of specificity for the demethylated loci, but in most cases it is not absolute [88,139]. Quadruple dme ros1 dml2 dml3 mutants lacking all DML proteins in somatic tissues can be obtained by central cell-specific ectopic expression of DME and display global hypermethylation and loss of tissue-specific differential methylation [139].
Interestingly, expression of ROS1 itself is positively controlled by methylation: a helitron transposon is present in front of the ROS1 promoter and suppresses transcription, whereas a specific “methylation monitoring sequence” in between can be methylated to uncouple the helitron from the promoter and boost ROS1 expression [157,158]. DML genes are also regulated by miRNA in development and during abiotic stress, as documented in A. thaliana, barley, and red sage (Salvia miltiorrhiza) [100,159,160].
Although the full spectrum of plant cell responses involving DML glycosylases has not yet been elucidated, it is clear that these proteins orchestrate gene activity in development and integrate many responses to various kinds of abiotic stress. In dicots, DME seems to be strictly dedicated to developmental regulation, while other DMLs participate both in development and in the stress response. In monocots, such specialization is not yet clear. A concept on “stress memory” has been proposed, in which the methylation status changes in response to abiotic stress can be imprinted and stably maintained for some time in plant development [161], or even in a trans-generational manner [162], through the balanced action of DML proteins and the methylation apparatus.
On a more practical side, the expression of transgenes in plants is often regulated by methylation-sensitive elements, such as cauliflower mosaic virus 35S promoter, and regulation of ROS1 was proposed as a way to drive or silence the expression of transgenes [163]. DML glycosylases are also promising tools for epigenetic status manipulation in non-plant cells: transfecting DME or ROS1 into human cells causes global loss or the mC epigenetic label [107,164,165], and fusions of ROS1 with specific DNA-addressing domains induce targeted DNA demethylation and gene activation [166,167]. However, DME overexpression in human cells induces DNA damage, the stress response, interferon production, S phase arrest, and ultimately cell death [164]. Interestingly, non-targeted demethylation by ROS1 in cancer cells also causes methylation gain, partially reversing the methylome pattern to that characteristic of normal cells [165].
5. Unexplained Diversity of Alkylation Damage Glycosylases
The major products of electrophilic addition removed by BER are purines alkylated at N3 and N7 positions, whereas products of alkylation at exocyclic groups and other ring nitrogens (N1 of purines, N3 of pyrimidines) are repaired by Ada-like alkyltransferases and AlkB-like dioxygenases without base excision [168,169,170]. DNA glycosylases from several structural superfamilies are capable of alkylation damage removal. In E. coli, the AlkA protein, which belongs to a wide, functionally diverse HhH superfamily, is induced by alkylation stress, and removes N3- and N7-alkylpurines and 1,N6-ethenoadenine (εA). Another E. coli alkylpurine–DNA glycosylase, Tag, is constitutive, and its substrate specificity is limited to N3-alkyladenine; structurally, it is sometimes also grouped with HhH proteins, but outside of the namesake helix–hairpin–helix motif, it is very different from the rest of the superfamily. In human cells, MPG protein (also known as AAG or APNG) removes 3-methylpurines, hypoxanthine, and εA and belongs to a completely different structural family, resembling the C-terminal domain of methionyl-tRNAfMet formyltransferase [171,172]. Moreover, some bacteria (but not E. coli) possess DNA glycosylases belonging to the HEAT repeat domain and winged-helix domain superfamilies and specific for alkylated purines or purine adducts with some natural genotoxic antibiotics [173,174].
In vertebrates, the primary endogenous source of DNA alkylation is stray reactions with metabolic methyl group donors, mainly S-adenosyl methionine [3]. Many plants, however, synthesize DNA-alkylating secondary metabolites that are either electrophiles or are metabolized to electrophilic species and, thus, may put the plant’s own genome under threat [12,14]. Thus, the problem of alkylation damage repair can be more acute in plants. Admittedly, many bulky adducts with electrophilic species may be removed by nucleotide excision repair [175] or mismatch repair [176], and ring-alkylated purine nucleotides can decay spontaneously [177], yet BER still plays an important role in fixing DNA damage from these lesions.
Plant cells excise both 3-mA and 7-mG from methylated DNA [178]. Most unfortunately, the mechanisms of this repair received very little attention, and only today, when many plant genomes have been sequenced, do we realize that they are exquisitely enriched in homologs of alkylpurine glycosylases. Gene duplication in plants followed by specialization of paralogs are not unusual [179], and some BER genes have apparently undergone such events, as the DML proteins discussed above can attest. Among other DNA glycosylases, a UNG pseudogene (At2g10550) arising from duplication and then disrupted by two transposon insertion events is found in the A. thaliana genome [42]. Compared with other branches of life, TAG genes are highly overrepresented in plant genomes [180]; A. thaliana possesses seven TAG homologs, some of which appear to result from ancestral duplications, while others appeared recently (Figure 4). On top of that, the A. thaliana genome codes for three AlkA homologs and one MPG homolog. The last one is actually the only plant alkylpurine–DNA glycosylase characterized in any detail: a cDNA from an A. thaliana library was cloned based on its functional ability to complement the methyl methanesulfonate sensitivity in an alkA tag E. coli strain [181]. Although the protein was not purified, extracts of E. coli cells harboring the plasmid with the gene insert excised 3-mA, but not 7-mG [181]. The expression of maize MPG is induced by UV and zeocin (a radiomimetic drug) treatments [182], but the significance of this finding is uncertain since neither agent is alkylating.
Both targeted and high-throughput experiments show that all alkylation damage glycosylases are expressed at some stages in the plant life cycle, so it is unlikely that the homologs detected at the genetic level represent pseudogenes [183,184]. It remains to be seen what their biologically relevant substrates are and whether some of them play organ- and development-specialized roles.
6. AP Site Cleavage and 3′-End Processing
All three types of DNA glycosylase products, namely AP site, 3′-terminal unsaturated aldehyde, and 3′-terminal phosphate, need further processing to expose a 3′-OH end that can be extended by DNA polymerases (Figure 1 and Figure 2). In E. coli, all these reactions are catalyzed by either of two AP endonucleases, exonuclease III (Xth, belonging to the Exonuclease–Endonuclease–Phosphatase (EEP) structural superfamily) and endonuclease IV (Nfo, triose phosphate isomerase barrel structural superfamily) [185]. Xth is a constitutive protein, while Nfo is induced by oxidative stress but is still present at the level of ~10% of Xth in uninduced cells. Higher eukaryotes possess two Xth homologs, APE1 and APE2. The former is the major AP endonuclease in human cells and can also remove 3′-terminal sugar remnants, but lacks the 3′-phosphatase activity, and these ends are cleaned by PNKP [186,187]. APE2 is predominantly a 3′→5′ exonuclease, rather than an AP endonuclease, and is involved in end resection at single-strand breaks in several DNA repair pathways, including BER [188].
Three A. thaliana enzymes belonging to the EEP superfamily, AtAPE1L, AtARP, and AtAPE2, show an AP endonuclease activity [189,190]. Additionally, there is a functional equivalent of PNKP, AtZDP (see below). APE2 is a closer homolog to human APE2 and, similar to its human counterpart, possesses only a modest AP endonuclease activity, but strong 3′→5′-exonuclease activity [190,191]. Of the remaining two, APE1L is apparently more important, since ape1L ape2 and ape1L zdp double knockout seeds are inviable, whereas those of the arp ape1L and arp ape2 genotype survive [190,192,193]. APE1L also eagerly removes 3′-terminal unsaturated aldehyde moieties generated by DNA glycosylases that carry out β-elimination and the 3′-terminal phosphate, the product of β,δ-elimination [190].
APE1L interacts with ROS1 forming a ternary complex on DNA, but does not displace ROS1 from the reaction product [190]. This distinguishes APE1L from hAPE1, which stimulates many DNA glycosylases by disrupting the tight enzyme–product complexes [194]. Methylation patterns are severely disrupted in ape1L and ape1L zdp knockout cells [190].
ARP proteins from A. thaliana, wheat, and sugarcane demonstrate the AP endonuclease activity [190,195,196,197] and, in addition, can cleave DNA 5′ of some damaged nucleotides, such as α-deoxyadenosine [196]. The 3′→5′-exonuclease, 3′-phosphodiesterase, and 3′-phosphatase activities are somewhat controversial; AtARP was reported to lack [190] and to possess them [196]. ARP is involved in the post-incision steps during U and 8-oxoG repair [195,198]. A. thaliana arp plants show sensitivity to methyl methanesulfonate and tert-butyl hydroperoxide, but not to H2O2 [196].
In addition to their AP endonuclease activity, mammalian APE1 proteins possess so-called “redox activity”, reactivating many oxidized transcription factors [199]. ARP has been reported to possess redox activity [189]; however, its mechanism is unclear since no equivalent of the redox-active Cys65 residue of hAPE1 is found in AtARP. Recently, the redox nature of transcription factor reactivation has been questioned [200,201].
The APE-less BER branch in plants relies on the zinc finger DNA 3′-phosphatase (ZDP), rather than on PNKP as in mammals [198]. AtZDP efficiently removes the 3′-phosphate from a gapped substrate, but lacks the kinase domain present in human PNKP and, thus, has no 5′-kinase activity [198,202]. In addition, instead of the forkhead-associated domain of PNKP responsible for XRCC1 binding [203], ZDP is equipped with three PARP-like zinc fingers, which may serve as a nick sensor [202,204,205]. In addition to MMH-1, an efficient catalyst of β,δ-elimination, ZDP assists the processing of intermediates formed by OGG1 and DMLs, which also produce a fraction of product with a 3′-terminal phosphate [26,103,105,198,206]. Genetic data show that APE2 is epistatic with ZDP in DML-associated demethylation, but the phosphatase activity of APE2 is weak, and the repair probably depends on its 3′→5′-exonuclease activity [191]. In zdp ape2 double knockouts, 3′-blocked BER intermediates accumulate and cause proliferation and development defects partially rescued by abrogation of MMH glycosylase/β,δ-lyase or of SOG1, a plant-specific transcriptional regulator acting in a break-sensing ATR pathway [28]. Thus, APE1L, APE2, and ZDP all appear to participate in 3′-end cleaning in plants.
DNA glycosylases/AP lyases can also process AP sites, formed either by spontaneous depurination or by other DNA glycosylases. Until recently, this reaction was considered rather irrelevant biologically because the processing of the 3′-ends would still require AP endonucleases or 3′-phosphatases. However, recently the repair of damage induced by methyl methanesulfonate (MMS) in A. thaliana was shown to depend on the sequential action of MMH-1 and ZDP [177]. MMS predominantly produces N7-methylguanine, which either depurinates spontaneously or is excised by the strictly monofunctional alkylpurine glycosylases, producing an AP site. It has been suggested that, at least in plants, ARP and/or APE1L may be predominantly involved in the repair of AP sites formed by DNA glycosylases, whereas AP lyases contribute significantly to the repair of spontaneously formed AP sites [177,207].
7. Downstream Events in Plant BER
7.1. DNA Polymerases
A. thaliana cell extracts are proficient in both short- and long-patch BER [41,195,208]. In a major departure from the human paradigm, plants lack homologs of POLβ, which initiates repair DNA synthesis in both short- and long-patch BER. Nevertheless, POLλ is present and functional, possessing both polymerase and dRP lyase activities [202,209,210]. Its expression is induced by MMS, UV and white light, and salt stress [210,211,212]. As in human cells, AtPOLλ is involved in non-homologous end joining, and likely participates in nucleotide excision repair of UV-B damage [211,213]. Moreover, similar to the human system [214], AtPOLλ shows the ability to incorporate a correct C opposite to 8-oxoG, making it a good candidate for the GO system polymerase [215]. All this said, direct evidence of POLλ participation in plant BER, such as reconstitution experiments or the establishment of the full BER cycle in knockout plants, is lacking at the moment.
There definitely is a possibility that other plant DNA polymerases, whose main role is in replication, translesion synthesis, or other DNA repair pathways, also participate in BER. In addition to POLλ, plant genomes code for homologs of human DNA polymerases γ and θ (Family A), α, δ, ε, ζ (Family B), η, κ, and REV1 (Family Y). In the literature, for historical reasons, two A. thaliana POLγ homologs are often referred to as homologs of E. coli DNA polymerase I (PolI), albeit they are actually closer to POLγ. Mutations in the small subunit of POLδ, POLD2, are hypersensitive to MMS, which may suggest POLδ involvement in BER [216]. The POLγ/PolI homologs have been shown to participate in BER in mitochondria and chloroplasts (see below).
7.2. Flap Endonuclease
Flap endonuclease, FEN1, is a key enzyme in long-patch BER in vertebrates, removing the strand displaced by a DNA polymerase once it progresses beyond the replacement of a single nucleotide [217,218]. FEN1 is not a specific BER enzyme, its main role being the removal of Okazaki fragments during lagging-strand replication [218]. A. thaliana FEN1 was independently cloned by two groups in 2016, as a suppressor of transgene silencing and a regulator of light-dependent root and hypocotyl elongation [219,220]. Biochemically, AtFEN1 is similar to hFEN1, but lacks a 5′→3′-exonuclease activity, and the mutant plants with a splicing defect show sensitivity to MMS and UV, but are still viable, unlike complete knockouts [219,220]. FEN1 homologs from cauliflower and rice (two paralogs) were purified or cloned and characterized, showing essentially the same properties, except the presence of the 5′→3′-exonuclease activity in one of the rice proteins [221,222,223,224]. The second rice homolog might be non-functional, since it does not complement the rad27 yeast strain deficient in FEN1 [224].
7.3. DNA Ligases
In vertebrates, two DNA ligases participate in different BER branches: Lig IIIα completes short-patch BER, and Lig I works in the long-patch subpathway [217]. As with FEN1, Lig I’s main role is in replication. Lig IIIα is absent from plants, and Lig I (AtLIG1 in A. thaliana) seems to be the only DNA ligase acting in plant BER [195,225]. Genetic evidence suggests that LIG1 works in the same pathway as MBD4, DME, and ROS1 [49,226,227]. A. thaliana also possesses a second homolog of Lig I (At1g49250), which remains uncharacterized at the moment.
7.4. XRCC1
XRCC1, the adaptor protein, orchestrates the coordinated transfer of repair intermediates between sequential BER enzymes in higher eukaryotes [228,229]. Plant XRCC1 homologs have been characterized from A. thaliana [230] and rice [231]. Notably, AtXRCC1 is about twice as short as the human protein and lacks both the N-terminal domain and the second BRCT domain that forms the dimerization interface [232,233]. Nevertheless, plant XRCC1 proteins seem to be functionally equivalent to hXRCC1, at least with respect to BER: they bind DNA forming a scaffold that brings together the upstream actors (at least ROS1 and ZDP) and stimulate their activity both as purified proteins and in cell extracts [230]. Co-localization of APE1L, ZDP, and ROS1 observed in living cells [190] may be due to their assembly on the XRCC1 scaffold. A. thaliana plants deficient in XRCC1 are viable but radiosensitive [234].
7.5. Poly(ADP-Ribose) Polymerases
Poly(ADP-ribose) polymerases are multifunctional proteins sensing single- and double-strand breaks and initiating post-translational modification of many proteins, including themselves by mono- and poly(ADP-ribose) [235,236]. In humans, the PARP family comprises 17 members, the best studied of which is PARP1, and the role in DNA repair has also been shown for PARP2 and PARP3. The main function of PARPs in BER seems to be the reorganization of chromatin at single-strand breaks formed after AP endonuclease action (or spontaneously), clearing the site of action for further BER steps. In contrast, A. thaliana possesses only three PARPs; in the literature, two of them are somewhat non-discriminately named either PARP1 or PARP2. AtPARP1 (At2g31320) seems to be the primary poly(ADP-ribose) polymerase in DNA damage response, while AtPARP2 (At4g02390) is more active as a recombinant protein in vitro, but apparently plays a back-up role when assayed in cell-free extracts [237,238]. AtPARP1 and AtPARP2 knockouts are hypersensitive to MMS, and the expression of both genes is induced by zeocin [239]. AtPARP3 (At5g22470) is highly expressed in seeds and is required for seed storability, yet it is not induced by genotoxic stress and purified recombinant AtPARP3 shows no activity [238,239,240]. Overall, PARP1 and PARP2 seem to participate in plant BER.
8. BER in Plant Mitochondria and Chloroplasts
BER in organelles is organized along with the general BER scheme, but has some important details. The best-studied example of organellar BER is provided by mammalian mitochondria [241,242]. The majority of DNA glycosylases, as well as APE1, have been experimentally shown to localize in mitochondria, or their mitochondrial localization signals can target reporter proteins to mitochondria, or at least have predicted mitochondrial isoforms. POLγ is the main BER polymerase in mitochondria, and the ligation step depends on the mitochondrial isoform of Lig IIIα.
A. thaliana and potato tuber mitochondria can support the full BER cycle for U [243,244], and complete BER of 5-hydroxyuracil was detected in mitochondrial extracts from potato [244]. Unlike human UNG gene, which produces mitochondrial UNG1 and nuclear UNG2 isoforms with different N-terminal tails via alternative transcription initiation and splicing, only one AtUNG mRNA and protein exist; however, the first 174 residues of AtUNG have been shown to target an EGFP reporter to mitochondria [243]. Interestingly, UNG and, to a lesser degree, AP endonuclease activity is partly localized in the mitochondrial membrane fraction [243]. This is reminiscent of the membrane localization of the mitochondrial fraction of human UNG and OGG1 [245,246]. An interesting but still untested possibility is that the inner membrane, which both hosts the electron transport chain and is tightly associated with mitochondrial DNA [247], is the principal site of both damage and its immediate repair in this organelle. However, as with the nuclear genome, ung A. thaliana plants do not accumulate mutations in mitochondrial DNA, and the same holds for mmh and ogg1 plants, suggesting the presence of alternative repair mechanisms, most likely mismatch repair [44,248]. UNG activity was also partially purified from maize mitochondria [38], and mitochondria from Brazilian pine (Araucaria angustifolia) contain activities of UNG, MMH, and AP endonuclease [249]. Pol I/POLγ homologs, characterized from A. thaliana, rice, and tobacco [250,251,252,253,254], carry out the repair synthesis step. Notably, one of these homologs, PolIB, carries an insertion in the polymerase domain providing it with the dRP lyase activity, which is absent from other Family A DNA polymerases [254]. Based on the observed error rate, which is ~10-fold higher for AtPolIB than for AtPolIA, and drawing parallels with the low fidelity of POLβ [255], one can surmise that PolIB is a repair polymerase, whereas PolIA is better suited for a role in organellar replication [256]. The mitochondrial isoform of Lig I completes the repair.
In comparison, BER in chloroplasts has received less attention. Regarding the initial BER stages, UNG activity was detected in maize chloroplasts [257] and AtNTH1, AtNTH2, and AtARP were found to be localized in chloroplasts [81]. Both short- and long-patch BER has been observed in chloroplasts, and the same Pol I/POLγ-like DNA polymerases participate in it [251,253,254]. The nature of the ligase remains a mystery, since, at least in A. thaliana, Lig I is targeted to nuclei and mitochondria, but not to chloroplasts [258,259]. The overall organization of BER in chloroplasts is most likely similar to nuclear and mitochondrial BER, but many crucial details are still missing.
9. Conclusions
Although the principal mechanism of base excision DNA repair in plants is the same as in vertebrates, bacteria, and yeast, the evolutionary history and specific lifestyle endowed plants with their unique repair features. The most obvious and seemingly the most important of those is the existence of epigenetic DNA glycosylases, subverting the genome protection machinery for the ends of gene activity regulation. No equivalent of this system is known outside of the Viridiplantae kingdom; the animal TETs/TDG active demethylation pathway apparently evolved independently. Another striking feature of plant BER, still virtually unstudied, is the high diversity of DNA glycosylases homologous to proteins repairing alkylation damage in bacteria. Elucidating their specificity and biological roles might shed light on the unique kinds of genotoxic stress to which plants are exposed as compared to other living things. Processing of DNA glycosylase reaction products, namely AP sites and dirty 3′-ends, also seems to vary between plants on the one hand and humans and bacteria on the other hand, and relies more on AP lyases and 3′-phosphatases in plants, although there is still no clear picture of distinct subpathways following the action of different glycosylases. Finally, chloroplasts, the most distinctive plant organelle, are an exceptionally important source of DNA damage, yet the repair of the chloroplast genome has been barely addressed so far. Understanding the biochemistry of plant DNA repair, especially in the key crop species, such as rice, wheat, vegetable cultures, etc., should advance our ability to breed or engineer new varieties with increased robustness to abiotic stress.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/ijms241914746/s1.
Author Contributions
Conceptualization, I.R.G., H.L. and D.O.Z.; writing—original draft preparation, I.R.G., D.V.P., A.V.E., C.M. and B.Y.; writing—review and editing, I.R.G., H.L. and D.O.Z.; visualization, D.V.P. and D.O.Z.; supervision, I.R.G. and D.O.Z.; project administration, I.R.G., H.L. and D.O.Z.; funding acquisition, I.R.G., H.L. and D.O.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by a joint grant from the Russian Science Foundation (23-44-00050) and the International Cooperation and Exchange of the National Science Foundation of China Project (32261133530). Sequence analysis of DML and TAG protein families was supported by the Russian Ministry of Science and Education (project 12103130056-8).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data are contained in the paper and Supplementary Materials.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Friedberg, E.C.; Walker, G.C.; Siede, W.; Wood, R.D.; Schultz, R.A.; Ellenberger, T. DNA Repair and Mutagenesis; ASM Press: Washington, DC, USA, 2006; p. 1118. [Google Scholar]
- Lindahl, T. Instability and decay of the primary structure of DNA. Nature 1993, 362, 709–715. [Google Scholar] [CrossRef] [PubMed]
- De Bont, R.; van Larebeke, N. Endogenous DNA damage in humans: A review of quantitative data. Mutagenesis 2004, 19, 169–185. [Google Scholar] [CrossRef] [PubMed]
- Vijg, J.; Suh, Y. Genome instability and aging. Annu. Rev. Physiol. 2013, 75, 645–668. [Google Scholar] [CrossRef] [PubMed]
- Behrens, A.; van Deursen, J.M.; Rudolph, K.L.; Schumacher, B. Impact of genomic damage and ageing on stem cell function. Nat. Cell Biol. 2014, 16, 201–207. [Google Scholar] [CrossRef] [PubMed]
- Caldecott, K.W. Mammalian DNA base excision repair: Dancing in the moonlight. DNA Repair 2020, 93, 102921. [Google Scholar] [CrossRef] [PubMed]
- Hindi, N.N.; Elsakrmy, N.; Ramotar, D. The base excision repair process: Comparison between higher and lower eukaryotes. Cell. Mol. Life Sci. 2021, 78, 7943–7965. [Google Scholar] [CrossRef] [PubMed]
- Wozniak, K.J.; Simmons, L.A. Bacterial DNA excision repair pathways. Nat. Rev. Microbiol. 2022, 20, 465–477. [Google Scholar] [CrossRef]
- Schmitt, F.-J.; Renger, G.; Friedrich, T.; Kreslavski, V.D.; Zharmukhamedov, S.K.; Los, D.A.; Kuznetsov, V.V.; Allakhverdiev, S.I. Reactive oxygen species: Re-evaluation of generation, monitoring and role in stress-signaling in phototrophic organisms. Biochim. Biophys. Acta 2014, 1837, 835–848. [Google Scholar] [CrossRef]
- Czarnocka, W.; Karpiński, S. Friend or foe? Reactive oxygen species production, scavenging and signaling in plant response to environmental stresses. Free Radic. Biol. Med. 2018, 122, 4–20. [Google Scholar] [CrossRef]
- Lorence, A.; Nessler, C.L. Camptothecin, over four decades of surprising findings. Phytochemistry 2004, 65, 2735–2749. [Google Scholar] [CrossRef]
- Rietjens, I.M.C.M.; Martena, M.J.; Boersma, M.G.; Spiegelenberg, W.; Alink, G.M. Molecular mechanisms of toxicity of important food-borne phytotoxins. Mol. Nutr. Food Res. 2005, 49, 131–158. [Google Scholar] [CrossRef] [PubMed]
- Pommier, Y.; Leo, E.; Zhang, H.; Marchand, C. DNA topoisomerases and their poisoning by anticancer and antibacterial drugs. Chem. Biol. 2010, 17, 421–433. [Google Scholar] [CrossRef] [PubMed]
- Prinsloo, G.; Nogemane, N.; Street, R. The use of plants containing genotoxic carcinogens as foods and medicine. Food Chem. Toxicol. 2018, 116, 27–39. [Google Scholar] [CrossRef] [PubMed]
- Sidorenko, V.S. Biotransformation and toxicities of aristolochic acids. Adv. Exp. Med. Biol. 2020, 1241, 139–166. [Google Scholar] [CrossRef] [PubMed]
- Stirpe, F.; Battelli, M.G. Ribosome-inactivating proteins: Progress and problems. Cell. Mol. Life Sci. 2006, 63, 1850–1866. [Google Scholar] [CrossRef] [PubMed]
- Spampinato, C.P. Protecting DNA from errors and damage: An overview of DNA repair mechanisms in plants compared to mammals. Cell. Mol. Life Sci. 2017, 74, 1693–1709. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zhu, J.; Gong, Z.; Zhu, J.-K. Abiotic stress responses in plants. Nat. Rev. Genet. 2022, 23, 104–119. [Google Scholar] [CrossRef] [PubMed]
- Møller, I.M.; Jensen, P.E.; Hansson, A. Oxidative modifications to cellular components in plants. Annu. Rev. Plant Biol. 2007, 58, 459–481. [Google Scholar] [CrossRef]
- Briggs, A.G.; Bent, A.F. Poly(ADP-ribosyl)ation in plants. Trends Plant Sci. 2011, 16, 372–380. [Google Scholar] [CrossRef]
- Mannuss, A.; Trapp, O.; Puchta, H. Gene regulation in response to DNA damage. Biochim. Biophys. Acta 2012, 1819, 154–165. [Google Scholar] [CrossRef]
- Rissel, D.; Peiter, E. Poly(ADP-ribose) polymerases in plants and their human counterparts: Parallels and peculiarities. Int. J. Mol. Sci. 2019, 20, 1638. [Google Scholar] [CrossRef]
- Parrilla-Doblas, J.T.; Roldán-Arjona, T.; Ariza, R.R.; Córdoba-Cañero, D. Active DNA demethylation in plants. Int. J. Mol. Sci. 2019, 20, 4683. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Lang, Z. The mechanism and function of active DNA demethylation in plants. J. Integr. Plant Biol. 2020, 62, 148–159. [Google Scholar] [CrossRef]
- Chen, H.; Chu, P.; Zhou, Y.; Li, Y.; Liu, J.; Ding, Y.; Tsang, E.W.T.; Jiang, L.; Wu, K.; Huang, S. Overexpression of AtOGG1, a DNA glycosylase/AP lyase, enhances seed longevity and abiotic stress tolerance in Arabidopsis. J. Exp. Bot. 2012, 63, 4107–4121. [Google Scholar] [CrossRef] [PubMed]
- Córdoba-Cañero, D.; Roldán-Arjona, T.; Ariza, R.R. Arabidopsis ZDP DNA 3′-phosphatase and ARP endonuclease function in 8-oxoG repair initiated by FPG and OGG1 DNA glycosylases. Plant J. 2014, 79, 824–834. [Google Scholar] [CrossRef] [PubMed]
- Forti, C.; Shankar, A.; Singh, A.; Balestrazzi, A.; Prasad, V.; Macovei, A. Hydropriming and biopriming improve Medicago truncatula seed germination and upregulate DNA repair and antioxidant genes. Genes 2020, 11, 242. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Liang, W.; Liu, Y.; Ren, Z.; Ci, D.; Chang, J.; Qian, W. The Arabidopsis ATR-SOG1 signaling module regulates pleiotropic developmental adjustments in response to 3′-blocked DNA repair intermediates. Plant Cell 2022, 34, 852–866. [Google Scholar] [CrossRef] [PubMed]
- Nisa, M.-U.; Huang, Y.; Benhamed, M.; Raynaud, C. The plant DNA damage response: Signaling pathways leading to growth inhibition and putative role in response to stress conditions. Front. Plant Sci. 2019, 10, 653. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Lu, Z.; Wang, L.; Jin, B. Plant responses to heat stress: Physiology, transcription, noncoding RNAs, and epigenetics. Int. J. Mol. Sci. 2021, 22, 117. [Google Scholar] [CrossRef]
- Lindahl, T. An N-glycosidase from Escherichia coli that releases free uracil from DNA containing deaminated cytosine residues. Proc. Natl. Acad. Sci. USA 1974, 71, 3649–3653. [Google Scholar] [CrossRef]
- Krokan, H.; Wittwer, C.U. Uracil DNA-glycosylase from HeLa cells: General properties, substrate specificity and effect of uracil analogs. Nucleic Acids Res. 1981, 9, 2599–2613. [Google Scholar] [CrossRef] [PubMed]
- Kavli, B.; Slupphaug, G.; Krokan, H.E. Genomic uracil in biology, immunity and cancer. In DNA Damage, DNA Repair and Disease; Dizdaroglu, M., Lloyd, R.S., Eds.; Royal Society of Chemistry: London, UK, 2021; Volume 1, pp. 220–248. [Google Scholar] [CrossRef]
- Endutkin, A.V.; Zharkov, D.O. Substrate specificities of DNA glycosylases in vitro and in vivo. In DNA Damage, DNA Repair and Disease; Dizdaroglu, M., Lloyd, R.S., Eds.; Royal Society of Chemistry: London, UK, 2021; Volume 1, pp. 175–203. [Google Scholar] [CrossRef]
- Aravind, L.; Koonin, E.V. The α/β fold uracil DNA glycosylases: A common origin with diverse fates. Genome Biol. 2000, 1, research0007. [Google Scholar] [CrossRef] [PubMed]
- Blaisdell, P.; Warner, H. Partial purification and characterization of a uracil-DNA glycosylase from wheat germ. J. Biol. Chem. 1983, 258, 1603–1609. [Google Scholar] [CrossRef] [PubMed]
- Talpaert-Borlé, M.; Liuzzi, M. Base-excision repair in carrot cells: Partial purification and characterization of uracil-DNA glycosylase and apurinic/apyrimidinic endodeoxyribonuclease. Eur. J. Biochem. 1982, 124, 435–440. [Google Scholar] [CrossRef] [PubMed]
- Bensen, R.J.; Warner, H.R. The partial purification and characterization of nuclear and mitochondrial uracil-DNA glycosylase activities from Zea mays seedlings. Plant Physiol. 1987, 83, 149–154. [Google Scholar] [CrossRef] [PubMed]
- Inamdar, N.M.; Zhang, X.-Y.; Brough, C.L.; Gardiner, W.E.; Bisaro, D.M.; Ehrlich, M. Transfection of heteroduplexes containing uracil·guanine or thymine·guanine mispairs into plant cells. Plant Mol. Biol. 1992, 20, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Bones, A.M. Expression and occurrence of uracil-DNA glycosylase in higher plants. Physiol. Plant. 1993, 88, 682–688. [Google Scholar] [CrossRef] [PubMed]
- Córdoba-Cañero, D.; Morales-Ruiz, T.; Roldán-Arjona, T.; Ariza, R.R. Single-nucleotide and long-patch base excision repair of DNA damage in plants. Plant J. 2009, 60, 716–728. [Google Scholar] [CrossRef]
- Córdoba-Cañero, D.; Dubois, E.; Ariza, R.R.; Doutriaux, M.-P.; Roldán-Arjona, T. Arabidopsis uracil DNA glycosylase (UNG) is required for base excision repair of uracil and increases plant sensitivity to 5-fluorouracil. J. Biol. Chem. 2010, 285, 7475–7483. [Google Scholar] [CrossRef]
- Morales-Ruiz, T.; Romero-Valenzuela, Á.C.; Vázquez-Grande, V.M.; Roldán-Arjona, T.; Ariza, R.R.; Córdoba-Cañero, D. Monitoring base excision repair in Chlamydomonas reinhardtii cell extracts. DNA Repair 2018, 65, 34–41. [Google Scholar] [CrossRef]
- Wynn, E.; Purfeerst, E.; Christensen, A. Mitochondrial DNA repair in an Arabidopsis thaliana uracil N-glycosylase mutant. Plants 2020, 9, 261. [Google Scholar] [CrossRef] [PubMed]
- Hendrich, B.; Bird, A. Identification and characterization of a family of mammalian methyl-CpG binding proteins. Mol. Cell. Biol. 1998, 18, 6538–6547. [Google Scholar] [CrossRef] [PubMed]
- Bellacosa, A.; Cicchillitti, L.; Schepis, F.; Riccio, A.; Yeung, A.T.; Matsumoto, Y.; Golemis, E.A.; Genuardi, M.; Neri, G. MED1, a novel human methyl-CpG-binding endonuclease, interacts with DNA mismatch repair protein MLH1. Proc. Natl. Acad. Sci. USA 1999, 96, 3969–3974. [Google Scholar] [CrossRef] [PubMed]
- Sjolund, A.B.; Senejani, A.G.; Sweasy, J.B. MBD4 and TDG: Multifaceted DNA glycosylases with ever expanding biological roles. Mutat. Res. 2013, 743–744, 12–25. [Google Scholar] [CrossRef] [PubMed]
- Ramiro-Merina, Á.; Ariza, R.R.; Roldán-Arjona, T. Molecular characterization of a putative plant homolog of MBD4 DNA glycosylase. DNA Repair 2013, 12, 890–898. [Google Scholar] [CrossRef] [PubMed]
- Nota, F.; Cambiagno, D.A.; Ribone, P.; Alvarez, M.E. Expression and function of AtMBD4L, the single gene encoding the nuclear DNA glycosylase MBD4L in Arabidopsis. Plant Sci. 2015, 235, 122–129. [Google Scholar] [CrossRef]
- Cecchini, N.M.; Torres, J.R.; López, I.L.; Cobo, S.; Nota, F.; Alvarez, M.E. Alternative splicing of an exitron determines the subnuclear localization of the Arabidopsis DNA glycosylase MBD4L under heat stress. Plant J. 2022, 110, 377–388. [Google Scholar] [CrossRef] [PubMed]
- Halliwell, B.; Gutteridge, J.M.C. Free Radicals in Biology and Medicine, 5th ed.; Oxford University Press: Oxford, UK, 2015; p. 905. [Google Scholar]
- Cadet, J.; Davies, K.J.A.; Medeiros, M.H.G.; Di Mascio, P.; Wagner, J.R. Formation and repair of oxidatively generated damage in cellular DNA. Free Radic. Biol. Med. 2017, 107, 13–34. [Google Scholar] [CrossRef]
- Yudkina, A.V.; Shilkin, E.S.; Endutkin, A.V.; Makarova, A.V.; Zharkov, D.O. Reading and misreading 8-oxoguanine, a paradigmatic ambiguous nucleobase. Crystals 2019, 9, 269. [Google Scholar] [CrossRef]
- Endutkin, A.V.; Zharkov, D.O. GO system, a DNA repair pathway to cope with oxidative damage. Mol. Biol. 2021, 55, 193–210. [Google Scholar] [CrossRef]
- Huffman, J.L.; Sundheim, O.; Tainer, J.A. DNA base damage recognition and removal: New twists and grooves. Mutat. Res. 2005, 577, 55–76. [Google Scholar] [CrossRef] [PubMed]
- Hitomi, K.; Iwai, S.; Tainer, J.A. The intricate structural chemistry of base excision repair machinery: Implications for DNA damage recognition, removal, and repair. DNA Repair 2007, 6, 410–428. [Google Scholar] [CrossRef] [PubMed]
- Ohtsubo, T.; Matsuda, O.; Iba, K.; Terashima, I.; Sekiguchi, M.; Nakabeppu, Y. Molecular cloning of AtMMH, an Arabidopsis thaliana ortholog of the Escherichia coli mutM gene, and analysis of functional domains of its product. Mol. Gen. Genet. 1998, 259, 577–590. [Google Scholar] [CrossRef] [PubMed]
- Gao, M.-J.; Murphy, T.M. Alternative forms of formamidopyrimidine-DNA glycosylase from Arabidopsis thaliana. Photochem. Photobiol. 2001, 73, 128–134. [Google Scholar] [CrossRef] [PubMed]
- Murphy, T.M.; Gao, M.-J. Multiple forms of formamidopyrimidine-DNA glycosylase produced by alternative splicing in Arabidopsis thaliana. J. Photochem. Photobiol. B 2001, 61, 87–93. [Google Scholar] [CrossRef] [PubMed]
- Murphy, T.M.; Guo, Y.-Y. Antimutagenic specificities of two plant glycosylases, oxoguanine glycosylase and formamidopyrimidine glycosylase, assayed in vivo. Biochem. Biophys. Res. Commun. 2010, 392, 335–339. [Google Scholar] [CrossRef] [PubMed]
- Murphy, T.M.; George, A. A comparison of two DNA base excision repair glycosylases from Arabidopsis thaliana. Biochem. Biophys. Res. Commun. 2005, 329, 869–872. [Google Scholar] [CrossRef]
- Kathe, S.D.; Barrantes-Reynolds, R.; Jaruga, P.; Newton, M.R.; Burrows, C.J.; Bandaru, V.; Dizdaroglu, M.; Bond, J.P.; Wallace, S.S. Plant and fungal Fpg homologs are formamidopyrimidine DNA glycosylases but not 8-oxoguanine DNA glycosylases. DNA Repair 2009, 8, 643–653. [Google Scholar] [CrossRef]
- Duclos, S.; Aller, P.; Jaruga, P.; Dizdaroglu, M.; Wallace, S.S.; Doublié, S. Structural and biochemical studies of a plant formamidopyrimidine-DNA glycosylase reveal why eukaryotic Fpg glycosylases do not excise 8-oxoguanine. DNA Repair 2012, 11, 714–725. [Google Scholar] [CrossRef]
- Agnez-Lima, L.F.; Batistuzzo de Medeiros, S.R.; Maggi, B.S.; Quaresma, G.A.S. Base excision repair in sugarcane. Genet. Mol. Biol. 2001, 24, 123–129. [Google Scholar] [CrossRef]
- Scortecci, K.C.; Lima, A.F.O.; Carvalho, F.M.; Silva, U.B.; Agnez-Lima, L.F.; Batistuzzo de Medeiros, S.R. A characterization of a MutM/Fpg ortholog in sugarcane—A monocot plant. Biochem. Biophys. Res. Commun. 2007, 361, 1054–1060. [Google Scholar] [CrossRef] [PubMed]
- Macovei, A.; Balestrazzi, A.; Confalonieri, M.; Faé, M.; Carbonera, D. New insights on the barrel medic MtOGG1 and MtFPG functions in relation to oxidative stress response in planta and during seed imbibition. Plant Physiol. Biochem. 2011, 49, 1040–1050. [Google Scholar] [CrossRef] [PubMed]
- Santos, A.R.; Miguel, A.S.; Macovei, A.; Maycock, C.; Balestrazzi, A.; Oliva, A.; Fevereiro, P. CdSe/ZnS quantum dots trigger DNA repair and antioxidant enzyme systems in Medicago sativa cells in suspension culture. BMC Biotechnol. 2013, 13, 111. [Google Scholar] [CrossRef] [PubMed]
- Auffret van der Kemp, P.; Thomas, D.; Barbey, R.; de Oliveira, R.; Boiteux, S. Cloning and expression in Escherichia coli of the OGG1 gene of Saccharomyces cerevisiae, which codes for a DNA glycosylase that excises 7,8-dihydro-8-oxoguanine and 2,6-diamino-4-hydroxy-5-N-methylformamidopyrimidine. Proc. Natl. Acad. Sci. USA 1996, 93, 5197–5202. [Google Scholar] [CrossRef] [PubMed]
- Nash, H.M.; Bruner, S.D.; Schärer, O.D.; Kawate, T.; Addona, T.A.; Spooner, E.; Lane, W.S.; Verdine, G.L. Cloning of a yeast 8-oxoguanine DNA glycosylase reveals the existence of a base-excision DNA-repair protein superfamily. Curr. Biol. 1996, 6, 968–980. [Google Scholar] [CrossRef] [PubMed]
- Rosenquist, T.A.; Zharkov, D.O.; Grollman, A.P. Cloning and characterization of a mammalian 8-oxoguanine DNA glycosylase. Proc. Natl. Acad. Sci. USA 1997, 94, 7429–7434. [Google Scholar] [CrossRef] [PubMed]
- Radicella, J.P.; Dherin, C.; Desmaze, C.; Fox, M.S.; Boiteux, S. Cloning and characterization of hOGG1, a human homolog of the OGG1 gene of Saccharomyces cerevisiae. Proc. Natl. Acad. Sci. USA 1997, 94, 8010–8015. [Google Scholar] [CrossRef] [PubMed]
- Roldán-Arjona, T.; Wei, Y.-F.; Carter, K.C.; Klungland, A.; Anselmino, C.; Wang, R.-P.; Augustus, M.; Lindahl, T. Molecular cloning and functional expression of a human cDNA encoding the antimutator enzyme 8-hydroxyguanine-DNA glycosylase. Proc. Natl. Acad. Sci. USA 1997, 94, 8016–8020. [Google Scholar] [CrossRef]
- Dany, A.L.; Tissier, A. A functional OGG1 homologue from Arabidopsis thaliana. Mol. Genet. Genom. 2001, 265, 293–301. [Google Scholar] [CrossRef]
- García-Ortiz, M.-V.; Ariza, R.R.; Roldán-Arjona, T. An OGG1 orthologue encoding a functional 8-oxoguanine DNA glycosylase/lyase in Arabidopsis thaliana. Plant Mol. Biol. 2001, 47, 795–804. [Google Scholar] [CrossRef]
- Morales-Ruiz, T.; Birincioglu, M.; Jaruga, P.; Rodriguez, H.; Roldan-Arjona, T.; Dizdaroglu, M. Arabidopsis thaliana Ogg1 protein excises 8-hydroxyguanine and 2,6-diamino-4-hydroxy-5-formamidopyrimidine from oxidatively damaged DNA containing multiple lesions. Biochemistry 2003, 42, 3089–3095. [Google Scholar] [CrossRef] [PubMed]
- Yoshimura, K.; Ogawa, T.; Ueda, Y.; Shigeoka, S. AtNUDX1, an 8-oxo-7,8-dihydro-2′-deoxyguanosine 5′-triphosphate pyrophosphohydrolase, is responsible for eliminating oxidized nucleotides in Arabidopsis. Plant Cell Physiol. 2007, 48, 1438–1449. [Google Scholar] [CrossRef] [PubMed]
- Boldinova, E.O.; Khairullin, R.F.; Makarova, A.V.; Zharkov, D.O. Isoforms of base excision repair enzymes produced by alternative splicing. Int. J. Mol. Sci. 2019, 20, 3279. [Google Scholar] [CrossRef] [PubMed]
- Pagano, A.; de Sousa Araújo, S.; Macovei, A.; Leonetti, P.; Balestrazzi, A. The seed repair response during germination: Disclosing correlations between DNA repair, antioxidant response, and chromatin remodeling in Medicago truncatula. Front. Plant Sci. 2017, 8, 1972. [Google Scholar] [CrossRef] [PubMed]
- Forti, C.; Ottobrino, V.; Bassolino, L.; Toppino, L.; Rotino, G.L.; Pagano, A.; Macovei, A.; Balestrazzi, A. Molecular dynamics of pre-germinative metabolism in primed eggplant (Solanum melongena L.) seeds. Hortic. Res. 2020, 7, 87. [Google Scholar] [CrossRef] [PubMed]
- Roldán-Arjona, T.; Garcia-Ortiz, M.-V.; Ruiz-Rubio, M.; Ariza, R.R. cDNA cloning, expression and functional characterization of an Arabidopsis thaliana homologue of the Escherichia coli DNA repair enzyme endonuclease III. Plant Mol. Biol. 2000, 44, 43–52. [Google Scholar] [CrossRef] [PubMed]
- Gutman, B.L.; Niyogi, K.K. Evidence for base excision repair of oxidative DNA damage in chloroplasts of Arabidopsis thaliana. J. Biol. Chem. 2009, 284, 17006–17012. [Google Scholar] [CrossRef]
- Lee, T.-f.; Zhai, J.; Meyers, B.C. Conservation and divergence in eukaryotic DNA methylation. Proc. Natl. Acad. Sci. USA 2010, 107, 9027–9028. [Google Scholar] [CrossRef]
- Zemach, A.; Zilberman, D. Evolution of eukaryotic DNA methylation and the pursuit of safer sex. Curr. Biol. 2010, 20, R780–R785. [Google Scholar] [CrossRef]
- Bochtler, M.; Kolano, A.; Xu, G.-L. DNA demethylation pathways: Additional players and regulators. Bioessays 2017, 39, 1600178. [Google Scholar] [CrossRef]
- Wu, X.; Zhang, Y. TET-mediated active DNA demethylation: Mechanism, function and beyond. Nat. Rev. Genet. 2017, 18, 517–534. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.; Gehring, M.; Johnson, L.; Hannon, M.; Harada, J.J.; Goldberg, R.B.; Jacobsen, S.E.; Fischer, R.L. DEMETER, a DNA glycosylase domain protein, is required for endosperm gene imprinting and seed viability in Arabidopsis. Cell 2002, 110, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Gong, Z.; Morales-Ruiz, T.; Ariza, R.R.; Roldán-Arjona, T.; David, L.; Zhu, J.-K. ROS1, a repressor of transcriptional gene silencing in Arabidopsis, encodes a DNA glycosylase/lyase. Cell 2002, 111, 803–814. [Google Scholar] [CrossRef] [PubMed]
- Penterman, J.; Zilberman, D.; Huh, J.H.; Ballinger, T.; Henikoff, S.; Fischer, R.L. DNA demethylation in the Arabidopsis genome. Proc. Natl. Acad. Sci. USA 2007, 104, 6752–6757. [Google Scholar] [CrossRef] [PubMed]
- Ortega-Galisteo, A.P.; Morales-Ruiz, T.; Ariza, R.R.; Roldán-Arjona, T. Arabidopsis DEMETER-LIKE proteins DML2 and DML3 are required for appropriate distribution of DNA methylation marks. Plant Mol. Biol. 2008, 67, 671–681. [Google Scholar] [CrossRef] [PubMed]
- O’Leary, N.A.; Wright, M.W.; Brister, J.R.; Ciufo, S.; Haddad, D.; McVeigh, R.; Rajput, B.; Robbertse, B.; Smith-White, B.; Ako-Adjei, D.; et al. Reference sequence (RefSeq) database at NCBI: Current status, taxonomic expansion, and functional annotation. Nucleic Acids Res. 2016, 44, D733–D745. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. High-accuracy alignment ensembles enable unbiased assessments of sequence homology and phylogeny. BioRxiv 2022. [Google Scholar] [CrossRef]
- Felsenstein, J. PHYLIP—Phylogeny Inference Package (Version 3.2). Cladistics 1989, 5, 164–166. [Google Scholar] [CrossRef]
- Okonechnikov, K.; Golosova, O.; Fursov, M.; The UGENE Team. Unipro UGENE: A unified bioinformatics toolkit. Bioinformatics 2012, 28, 1166–1167. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive tree of life (iTOL) v3: An online tool for the display and annotation of phylogenetic and other trees. Nucleic Acids Res. 2016, 44, W242–W245. [Google Scholar] [CrossRef]
- Choi, C.-S.; Sano, H. Identification of tobacco genes encoding proteins possessing removal activity of 5-methylcytosines from intact tobacco DNA. Plant Biotechnol. 2007, 24, 339–344. [Google Scholar] [CrossRef]
- Zemach, A.; Kim, M.Y.; Silva, P.; Rodrigues, J.A.; Dotson, B.; Brooks, M.D.; Zilberman, D. Local DNA hypomethylation activates genes in rice endosperm. Proc. Natl. Acad. Sci. USA 2010, 107, 18729–18734. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.-H.; Irshad, F.; Yan, Y.; Li, C. Loss of function of the RRMF domain in OsROS1a causes sterility in rice (Oryza sativa L.). Int. J. Mol. Sci. 2022, 23, 11349. [Google Scholar] [CrossRef] [PubMed]
- Ono, A.; Yamaguchi, K.; Fukada-Tanaka, S.; Terada, R.; Mitsui, T.; Iida, S. A null mutation of ROS1a for DNA demethylation in rice is not transmittable to progeny. Plant J. 2012, 71, 564–574. [Google Scholar] [CrossRef] [PubMed]
- Wen, S.; Wen, N.; Pang, J.; Langen, G.; Brew-Appiah, R.A.T.; Mejias, J.H.; Osorio, C.; Yang, M.; Gemini, R.; Moehs, C.P.; et al. Structural genes of wheat and barley 5-methylcytosine DNA glycosylases and their potential applications for human health. Proc. Natl. Acad. Sci. USA 2012, 109, 20543–20548. [Google Scholar] [CrossRef] [PubMed]
- Kapazoglou, A.; Drosou, V.; Argiriou, A.; Tsaftaris, A.S. The study of a barley epigenetic regulator, HvDME, in seed development and under drought. BMC Plant Biol. 2013, 13, 172. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.; Li, X.; Liu, Q.; Zhao, Y.; Jiang, W.; Wu, A.; Zhou, D.-X. DNA demethylases remodel DNA methylation in rice gametes and zygote and are required for reproduction. Mol. Plant 2021, 14, 1569–1583. [Google Scholar] [CrossRef] [PubMed]
- Irshad, F.; Li, C.; Wu, H.-Y.; Yan, Y.; Xu, J.-H. The function of DNA demethylase gene ROS1a null mutant on seed development in rice (Oryza sativa) using the CRISPR/Cas9 system. Int. J. Mol. Sci. 2022, 23, 6357. [Google Scholar] [CrossRef]
- Morales-Ruiz, T.; Ortega-Galisteo, A.P.; Ponferrada-Marín, M.I.; Martínez-Macías, M.I.; Ariza, R.R.; Roldán-Arjona, T. DEMETER and REPRESSOR OF SILENCING 1 encode 5-methylcytosine DNA glycosylases. Proc. Natl. Acad. Sci. USA 2006, 103, 6853–6858. [Google Scholar] [CrossRef]
- Agius, F.; Kapoor, A.; Zhu, J.-K. Role of the Arabidopsis DNA glycosylase/lyase ROS1 in active DNA demethylation. Proc. Natl. Acad. Sci. USA 2006, 103, 11796–11801. [Google Scholar] [CrossRef]
- Ponferrada-Marín, M.I.; Roldán-Arjona, T.; Ariza, R.R. ROS1 5-methylcytosine DNA glycosylase is a slow-turnover catalyst that initiates DNA demethylation in a distributive fashion. Nucleic Acids Res. 2009, 37, 4264–4274. [Google Scholar] [CrossRef] [PubMed]
- Gruber, D.R.; Toner, J.J.; Miears, H.L.; Shernyukov, A.V.; Kiryutin, A.S.; Lomzov, A.A.; Endutkin, A.V.; Grin, I.R.; Petrova, D.V.; Kupryushkin, M.S.; et al. Oxidative damage to epigenetically methylated sites affects DNA stability, dynamics, and enzymatic demethylation. Nucleic Acids Res. 2018, 46, 10827–10839. [Google Scholar] [CrossRef] [PubMed]
- Petrova, D.V.; Permyakova, N.V.; Grin, I.R.; Zharkov, D.O. Characterization of demethylating DNA glycosylase ROS1 from Nicotiana tabacum L. Vavilov J. Genet. Breed. 2022, 26, 341–348. [Google Scholar] [CrossRef] [PubMed]
- Mok, Y.G.; Uzawa, R.; Lee, J.; Weiner, G.M.; Eichman, B.F.; Fischer, R.L.; Huh, J.H. Domain structure of the DEMETER 5-methylcytosine DNA glycosylase. Proc. Natl. Acad. Sci. USA 2010, 107, 19225–19230. [Google Scholar] [CrossRef] [PubMed]
- Ponferrada-Marín, M.I.; Parrilla-Doblas, J.T.; Roldán-Arjona, T.; Ariza, R.R. A discontinuous DNA glycosylase domain in a family of enzymes that excise 5-methylcytosine. Nucleic Acids Res. 2011, 39, 1473–1484. [Google Scholar] [CrossRef] [PubMed]
- Du, X.; Yang, Z.; Xie, G.; Wang, C.; Zhang, L.; Yan, K.; Yang, M.; Li, S.; Zhu, J.-K.; Du, J. Molecular basis of the plant ROS1-mediated active DNA demethylation. Nat. Plants 2023, 9, 271–279. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Hung, Y.-H.; Rim, H.J.; Zhang, D.; Frost, J.M.; Shin, H.; Jang, H.; Liu, F.; Xiao, W.; Iyer, L.M.; et al. The catalytic core of DEMETER guides active DNA demethylation in Arabidopsis. Proc. Natl. Acad. Sci. USA 2019, 116, 17563–17571. [Google Scholar] [CrossRef]
- Jang, H.; Shin, H.; Eichman, B.F.; Huh, J.H. Excision of 5-hydroxymethylcytosine by DEMETER family DNA glycosylases. Biochem. Biophys. Res. Commun. 2014, 446, 1067–1072. [Google Scholar] [CrossRef]
- Wang, X.-l.; Song, S.-h.; Wu, Y.-S.; Li, Y.-L.; Chen, T.-t.; Huang, Z.-y.; Liu, S.; Dunwell, T.L.; Pfeifer, G.P.; Dunwell, J.M.; et al. Genome-wide mapping of 5-hydroxymethylcytosine in three rice cultivars reveals its preferential localization in transcriptionally silent transposable element genes. J. Exp. Bot. 2015, 66, 6651–6663. [Google Scholar] [CrossRef]
- Varma, S.J.; Calvani, E.; Grüning, N.-M.; Messner, C.B.; Grayson, N.; Capuano, F.; Mülleder, M.; Ralser, M. Global analysis of cytosine and adenine DNA modifications across the tree of life. eLife 2022, 11, e81002. [Google Scholar] [CrossRef]
- Kalinka, A.; Starczak, M.; Gackowski, D.; Stępień, E.; Achrem, M. Global DNA 5-hydroxymethylcytosine level and its chromosomal distribution in four rye species. J. Exp. Bot. 2023, 74, 3488–3502. [Google Scholar] [CrossRef] [PubMed]
- Brooks, S.C.; Fischer, R.L.; Huh, J.H.; Eichman, B.F. 5-methylcytosine recognition by Arabidopsis thaliana DNA glycosylases DEMETER and DML3. Biochemistry 2014, 53, 2525–2532. [Google Scholar] [CrossRef]
- Parrilla-Doblas, J.T.; Ponferrada-Marín, M.I.; Roldán-Arjona, T.; Ariza, R.R. Early steps of active DNA demethylation initiated by ROS1 glycosylase require three putative helix-invading residues. Nucleic Acids Res. 2013, 41, 8654–8664. [Google Scholar] [CrossRef]
- Luo, D.; Bernard, D.G.; Balk, J.; Hai, H.; Cui, X. The DUF59 family gene AE7 acts in the cytosolic iron-sulfur cluster assembly pathway to maintain nuclear genome integrity in Arabidopsis. Plant Cell 2012, 24, 4135–4148. [Google Scholar] [CrossRef] [PubMed]
- Buzas, D.M.; Nakamura, M.; Kinoshita, T. Epigenetic role for the conserved Fe-S cluster biogenesis protein AtDRE2 in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2014, 111, 13565–13570. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Li, Q.; Yuan, W.; Cao, Z.; Qi, B.; Kumar, S.; Li, Y.; Qian, W. The cytosolic Fe-S cluster assembly component MET18 is required for the full enzymatic activity of ROS1 in active DNA demethylation. Sci. Rep. 2016, 6, 26443. [Google Scholar] [CrossRef] [PubMed]
- Bastow, E.L.; Bych, K.; Crack, J.C.; Le Brun, N.E.; Balk, J. NBP35 interacts with DRE2 in the maturation of cytosolic iron-sulphur proteins in Arabidopsis thaliana. Plant J. 2017, 89, 590–600. [Google Scholar] [CrossRef] [PubMed]
- Mikhailov, A.A.; Khantakova, D.V.; Nichiporenko, V.A.; Glebov, E.M.; Grivin, V.P.; Plyusnin, V.F.; Yanshole, V.V.; Petrova, D.V.; Kostin, G.A.; Grin, I.R. Photoinduced inhibition of DNA repair enzymes and the possible mechanism of photochemical transformations of the ruthenium nitrosyl complex [RuNO(β-Pic)2(NO2)2OH]. Metallomics 2019, 11, 1999–2009. [Google Scholar] [CrossRef]
- Parrilla-Doblas, J.T.; Morales-Ruiz, T.; Ariza, R.R.; Martínez-Macías, M.I.; Roldán-Arjona, T. The C-terminal domain of Arabidopsis ROS1 DNA demethylase interacts with histone H3 and is required for DNA binding and catalytic activity. DNA Repair 2022, 115, 103341. [Google Scholar] [CrossRef]
- Ponferrada-Marín, M.I.; Martínez-Macías, M.I.; Morales-Ruiz, T.; Roldán-Arjona, T.; Ariza, R.R. Methylation-independent DNA binding modulates specificity of Repressor of Silencing 1 (ROS1) and facilitates demethylation in long substrates. J. Biol. Chem. 2010, 285, 23032–23039. [Google Scholar] [CrossRef]
- Ponferrada-Marín, M.I.; Roldán-Arjona, T.; Ariza, R.R. Demethylation initiated by ROS1 glycosylase involves random sliding along DNA. Nucleic Acids Res. 2012, 40, 11554–11562. [Google Scholar] [CrossRef]
- Kong, X.; Hong, Y.; Hsu, Y.-F.; Huang, H.; Liu, X.; Song, Z.; Zhu, J.-K. SIZ1-mediated SUMOylation of ROS1 enhances its stability and positively regulates active DNA demethylation in Arabidopsis. Mol. Plant 2020, 13, 1816–1824. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Nie, W.-F.; Xiong, X.; Wang, Y.; Jiang, Y.; Huang, P.; Lin, X.; Qin, G.; Huang, H.; Niu, Q.; et al. A novel protein complex that regulates active DNA demethylation in Arabidopsis. J. Integr. Plant Biol. 2021, 63, 772–786. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Pontes, O.; Zhu, J.; Miki, D.; Zhang, F.; Li, W.-X.; Iida, K.; Kapoor, A.; Pikaard, C.S.; Zhu, J.-K. ROS3 is an RNA-binding protein required for DNA demethylation in Arabidopsis. Nature 2008, 455, 1259–1262. [Google Scholar] [CrossRef] [PubMed]
- Qian, W.; Miki, D.; Zhang, H.; Liu, Y.; Zhang, X.; Tang, K.; Kan, Y.; La, H.; Li, X.; Li, S.; et al. A histone acetyltransferase regulates active DNA demethylation in Arabidopsis. Science 2012, 336, 1445–1448. [Google Scholar] [CrossRef]
- Rea, M.; Zheng, W.; Chen, M.; Braud, C.; Bhangu, D.; Rognan, T.N.; Xiao, W. Histone H1 affects gene imprinting and DNA methylation in Arabidopsis. Plant J. 2012, 71, 776–786. [Google Scholar] [CrossRef] [PubMed]
- Lang, Z.; Lei, M.; Wang, X.; Tang, K.; Miki, D.; Zhang, H.; Mangrauthia, S.K.; Liu, W.; Nie, W.; Ma, G.; et al. The methyl-CpG-binding protein MBD7 facilitates active DNA demethylation to limit DNA hyper-methylation and transcriptional gene silencing. Mol. Cell 2015, 57, 971–983. [Google Scholar] [CrossRef]
- Li, Q.; Wang, X.; Sun, H.; Zeng, J.; Cao, Z.; Li, Y.; Qian, W. Regulation of active DNA demethylation by a methyl-CpG-binding domain protein in Arabidopsis thaliana. PLoS Genet. 2015, 11, e1005210. [Google Scholar] [CrossRef]
- Tang, K.; Lang, Z.; Zhang, H.; Zhu, J.-K. The DNA demethylase ROS1 targets genomic regions with distinct chromatin modifications. Nat. Plants 2016, 2, 16169. [Google Scholar] [CrossRef]
- Scott, R.J.; Spielman, M. Genomic imprinting in plants and mammals: How life history constrains convergence. Cytogenet. Genome Res. 2006, 113, 53–67. [Google Scholar] [CrossRef]
- Eckersley-Maslin, M.A.; Alda-Catalinas, C.; Reik, W. Dynamics of the epigenetic landscape during the maternal-to-zygotic transition. Nat. Rev. Mol. Cell Biol. 2018, 19, 436–450. [Google Scholar] [CrossRef] [PubMed]
- Kinoshita, T.; Miura, A.; Choi, Y.; Kinoshita, Y.; Cao, X.; Jacobsen, S.E.; Fischer, R.L.; Kakutani, T. One-way control of FWA imprinting in Arabidopsis endosperm by DNA methylation. Science 2004, 303, 521–523. [Google Scholar] [CrossRef] [PubMed]
- Ohr, H.; Bui, A.Q.; Le, B.H.; Fischer, R.L.; Choi, Y. Identification of putative Arabidopsis DEMETER target genes by GeneChip analysis. Biochem. Biophys. Res. Commun. 2007, 364, 856–860. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, T.-F.; Shin, J.; Uzawa, R.; Silva, P.; Cohen, S.; Bauer, M.J.; Hashimoto, M.; Kirkbride, R.C.; Harada, J.J.; Zilberman, D.; et al. Regulation of imprinted gene expression in Arabidopsis endosperm. Proc. Natl. Acad. Sci. USA 2011, 108, 1755–1762. [Google Scholar] [CrossRef] [PubMed]
- Williams, B.P.; Bechen, L.L.; Pohlmann, D.A.; Gehring, M. Somatic DNA demethylation generates tissue-specific methylation states and impacts flowering time. Plant Cell 2022, 34, 1189–1206. [Google Scholar] [CrossRef] [PubMed]
- Ibarra, C.A.; Feng, X.; Schoft, V.K.; Hsieh, T.-F.; Uzawa, R.; Rodrigues, J.A.; Zemach, A.; Chumak, N.; Machlicova, A.; Nishimura, T.; et al. Active DNA demethylation in plant companion cells reinforces transposon methylation in gametes. Science 2012, 337, 1360–1364. [Google Scholar] [CrossRef] [PubMed]
- Khouider, S.; Borges, F.; LeBlanc, C.; Ungru, A.; Schnittger, A.; Martienssen, R.; Colot, V.; Bouyer, D. Male fertility in Arabidopsis requires active DNA demethylation of genes that control pollen tube function. Nat. Commun. 2021, 12, 410. [Google Scholar] [CrossRef]
- Zeng, W.; Huang, H.; Lin, X.; Zhu, C.; Kosami, K.-I.; Huang, C.; Zhang, H.; Duan, C.-G.; Zhu, J.-K.; Miki, D. Roles of DEMETER in regulating DNA methylation in vegetative tissues and pathogen resistance. J. Integr. Plant Biol. 2021, 63, 691–706. [Google Scholar] [CrossRef]
- Huang, M.; Zhang, Y.; Wang, Y.; Xie, J.; Cheng, J.; Fu, Y.; Jiang, D.; Yu, X.; Li, B. Active DNA demethylation regulates MAMP-triggered immune priming in Arabidopsis. J. Genet. Genom. 2022, 49, 796–809. [Google Scholar] [CrossRef]
- Zhang, M.; Xu, C.; Yan, H.; Zhao, N.; von Wettstein, D.; Liu, B. Limited tissue culture-induced mutations and linked epigenetic modifications in F1 hybrids of sorghum pure lines are accompanied by increased transcription of DNA methyltransferases and 5-methylcytosine glycosylases. Plant J. 2009, 57, 666–679. [Google Scholar] [CrossRef]
- Sun, R.-Z.; Zuo, E.-H.; Qi, J.-F.; Liu, Y.; Lin, C.-T.; Deng, X. A role of age-dependent DNA methylation reprogramming in regulating the regeneration capacity of Boea hygrometrica leaves. Funct. Integr. Genom. 2020, 20, 133–149. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Kapoor, A.; Sridhar, V.V.; Agius, F.; Zhu, J.-K. The DNA glycosylase/lyase ROS1 functions in pruning DNA methylation patterns in Arabidopsis. Curr. Biol. 2007, 17, 54–59. [Google Scholar] [CrossRef] [PubMed]
- Yamamuro, C.; Miki, D.; Zheng, Z.; Ma, J.; Wang, J.; Yang, Z.; Dong, J.; Zhu, J.-K. Overproduction of stomatal lineage cells in Arabidopsis mutants defective in active DNA demethylation. Nat. Commun. 2014, 5, 4062. [Google Scholar] [CrossRef] [PubMed]
- Penterman, J.; Uzawa, R.; Fischer, R.L. Genetic interactions between DNA demethylation and methylation in Arabidopsis. Plant Physiol. 2007, 145, 1549–1557. [Google Scholar] [CrossRef] [PubMed]
- La, H.; Ding, B.; Mishra, G.P.; Zhou, B.; Yang, H.; del Rosario Bellizzi, M.; Chen, S.; Meyers, B.C.; Peng, Z.; Zhu, J.-K.; et al. A 5-methylcytosine DNA glycosylase/lyase demethylates the retrotransposon Tos17 and promotes its transposition in rice. Proc. Natl. Acad. Sci. USA 2011, 108, 15498–15503. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.Y.; Ono, A.; Scholten, S.; Kinoshita, T.; Zilberman, D.; Okamoto, T.; Fischer, R.L. DNA demethylation by ROS1a in rice vegetative cells promotes methylation in sperm. Proc. Natl. Acad. Sci. USA 2019, 116, 9652–9657. [Google Scholar] [CrossRef] [PubMed]
- Ou, X.; Zhang, Y.; Xu, C.; Lin, X.; Zang, Q.; Zhuang, T.; Jiang, L.; von Wettstein, D.; Liu, B. Transgenerational inheritance of modified DNA methylation patterns and enhanced tolerance induced by heavy metal stress in rice (Oryza sativa L.). PLoS ONE 2012, 7, e41143. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; How-Kit, A.; Stammitti, L.; Teyssier, E.; Rolin, D.; Mortain-Bertrand, A.; Halle, S.; Liu, M.; Kong, J.; Wu, C.; et al. A DEMETER-like DNA demethylase governs tomato fruit ripening. Proc. Natl. Acad. Sci. USA 2015, 112, 10804–10809. [Google Scholar] [CrossRef]
- Lang, Z.; Wang, Y.; Tang, K.; Tang, D.; Datsenka, T.; Cheng, J.; Zhang, Y.; Handa, A.K.; Zhu, J.-K. Critical roles of DNA demethylation in the activation of ripening-induced genes and inhibition of ripening-repressed genes in tomato fruit. Proc. Natl. Acad. Sci. USA 2017, 114, E4511–E4519. [Google Scholar] [CrossRef]
- Ou, X.; Long, L.; Zhang, Y.; Xue, Y.; Liu, J.; Lin, X.; Liu, B. Spaceflight induces both transient and heritable alterations in DNA methylation and gene expression in rice (Oryza sativa L.). Mutat. Res. 2009, 662, 44–53. [Google Scholar] [CrossRef]
- Kuźnicki, D.; Meller, B.; Arasimowicz-Jelonek, M.; Braszewska-Zalewska, A.; Drozda, A.; Floryszak-Wieczorek, J. BABA-induced DNA methylome adjustment to intergenerational defense priming in potato to Phytophthora infestans. Front. Plant Sci. 2019, 10, 650. [Google Scholar] [CrossRef] [PubMed]
- Mosher, R.A.; Tan, E.H.; Shin, J.; Fischer, R.L.; Pikaard, C.S.; Baulcombe, D.C. An atypical epigenetic mechanism affects uniparental expression of Pol IV-dependent siRNAs. PLoS ONE 2011, 6, e25756. [Google Scholar] [CrossRef] [PubMed]
- Lei, M.; Zhang, H.; Julian, R.; Tang, K.; Xie, S.; Zhu, J.-K. Regulatory link between DNA methylation and active demethylation in Arabidopsis. Proc. Natl. Acad. Sci. USA 2015, 112, 3553–3557. [Google Scholar] [CrossRef] [PubMed]
- Williams, B.P.; Pignatta, D.; Henikoff, S.; Gehring, M. Methylation-sensitive expression of a DNA demethylase gene serves as an epigenetic rheostat. PLoS Genet. 2015, 11, e1005142. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Kwak, K.J.; Jung, H.J.; Lee, H.J.; Kang, H. MicroRNA402 affects seed germination of Arabidopsis thaliana under stress conditions via targeting DEMETER-LIKE Protein3 mRNA. Plant Cell Physiol. 2010, 51, 1079–1083. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Li, C.; Lu, S. Systematic analysis of DEMETER-like DNA glycosylase genes shows lineage-specific Smi-miR7972 involved in SmDML1 regulation in Salvia miltiorrhiza. Sci. Rep. 2018, 8, 7143. [Google Scholar] [CrossRef] [PubMed]
- Wibowo, A.; Becker, C.; Marconi, G.; Durr, J.; Price, J.; Hagmann, J.; Papareddy, R.; Putra, H.; Kageyama, J.; Becker, J.; et al. Hyperosmotic stress memory in Arabidopsis is mediated by distinct epigenetically labile sites in the genome and is restricted in the male germline by DNA glycosylase activity. eLife 2016, 5, e13546. [Google Scholar] [CrossRef] [PubMed]
- Williams, B.P.; Gehring, M. Stable transgenerational epigenetic inheritance requires a DNA methylation-sensing circuit. Nat. Commun. 2017, 8, 2124. [Google Scholar] [CrossRef]
- Otagaki, S.; Kasai, M.; Masuta, C.; Kanazawa, A. Enhancement of RNA-directed DNA methylation of a transgene by simultaneously downregulating a ROS1 ortholog using a virus vector in Nicotiana benthamiana. Front. Genet. 2013, 4, 44. [Google Scholar] [CrossRef]
- Mok, Y.G.; Choi, K.Y.; Hong, S.H.; Huh, J.H. DEMETER plant DNA demethylase induces antiviral response by interferon signalling in animal cells. Sci. Rep. 2017, 7, 9160. [Google Scholar] [CrossRef]
- Morales-Ruiz, T.; García-Ortiz, M.V.; Devesa-Guerra, I.; Raya-Ruiz, L.; Tejedor, J.R.; Bayón, G.F.; Sierra, M.I.; Fraga, M.F.; Ariza, R.R.; Roldán-Arjona, T. DNA methylation reprogramming of human cancer cells by expression of a plant 5-methylcytosine DNA glycosylase. Epigenetics 2018, 13, 95–107. [Google Scholar] [CrossRef] [PubMed]
- Parrilla-Doblas, J.T.; Ariza, R.R.; Roldán-Arjona, T. Targeted DNA demethylation in human cells by fusion of a plant 5-methylcytosine DNA glycosylase to a sequence-specific DNA binding domain. Epigenetics 2017, 12, 296–303. [Google Scholar] [CrossRef] [PubMed]
- Devesa-Guerra, I.; Morales-Ruiz, T.; Pérez-Roldán, J.; Parrilla-Doblas, J.T.; Dorado-León, M.; García-Ortiz, M.V.; Ariza, R.R.; Roldán-Arjona, T. DNA methylation editing by CRISPR-guided excision of 5-methylcytosine. J. Mol. Biol. 2020, 432, 2204–2216. [Google Scholar] [CrossRef] [PubMed]
- Drabløs, F.; Feyzi, E.; Aas, P.A.; Vaagbø, C.B.; Kavli, B.; Bratlie, M.S.; Peña-Diaz, J.; Otterlei, M.; Slupphaug, G.; Krokan, H.E. Alkylation damage in DNA and RNA—Repair mechanisms and medical significance. DNA Repair 2004, 3, 1389–1407. [Google Scholar] [CrossRef] [PubMed]
- Soll, J.M.; Sobol, R.W.; Mosammaparast, N. Regulation of DNA alkylation damage repair: Lessons and therapeutic opportunities. Trends Biochem. Sci. 2017, 42, 206–218. [Google Scholar] [CrossRef] [PubMed]
- Tsao, N.; Schärer, O.D.; Mosammaparast, N. The complexity and regulation of repair of alkylation damage to nucleic acids. Crit. Rev. Biochem. Mol. Biol. 2021, 56, 125–136. [Google Scholar] [CrossRef] [PubMed]
- Wyatt, M.D.; Allan, J.M.; Lau, A.Y.; Ellenberger, T.E.; Samson, L.D. 3-methyladenine DNA glycosylases: Structure, function, and biological importance. Bioessays 1999, 21, 668–676. [Google Scholar] [CrossRef]
- Hollis, T.; Lau, A.; Ellenberger, T. Structural studies of human alkyladenine glycosylase and E. coli 3-methyladenine glycosylase. Mutat. Res. 2000, 460, 201–210. [Google Scholar] [CrossRef]
- Shi, R.; Shen, X.-X.; Rokas, A.; Eichman, B.F. Structural biology of the HEAT-like repeat family of DNA glycosylases. Bioessays 2018, 40, 1800133. [Google Scholar] [CrossRef]
- Mullins, E.A.; Rodriguez, A.A.; Bradley, N.P.; Eichman, B.F. Emerging roles of DNA glycosylases and the base excision repair pathway. Trends Biochem. Sci. 2019, 44, 765–781. [Google Scholar] [CrossRef]
- Kunz, B.A.; Anderson, H.J.; Osmond, M.J.; Vonarx, E.J. Components of nucleotide excision repair and DNA damage tolerance in Arabidopsis thaliana. Environ. Mol. Mutagen. 2005, 45, 115–127. [Google Scholar] [CrossRef] [PubMed]
- Greene, E.A.; Codomo, C.A.; Taylor, N.E.; Henikoff, J.G.; Till, B.J.; Reynolds, S.H.; Enns, L.C.; Burtner, C.; Johnson, J.E.; Odden, A.R.; et al. Spectrum of chemically induced mutations from a large-scale reverse-genetic screen in Arabidopsis. Genetics 2003, 164, 731–740. [Google Scholar] [CrossRef]
- Barbado, C.; Córdoba-Cañero, D.; Ariza, R.R.; Roldán-Arjona, T. Nonenzymatic release of N7-methylguanine channels repair of abasic sites into an AP endonuclease-independent pathway in Arabidopsis. Proc. Natl. Acad. Sci. USA 2018, 115, E916–E924. [Google Scholar] [CrossRef]
- Kraszewska, E.; Dobrzańska, M.; Tudek, B. Tobacco BY-2 cells excise both 3-methyladenine and 7-methylguanine from methylated DNA. Mutat. Res. 1998, 409, 91–95. [Google Scholar] [CrossRef] [PubMed]
- Panchy, N.; Lehti-Shiu, M.; Shiu, S.-H. Evolution of gene duplication in plants. Plant Physiol. 2016, 171, 2294–2316. [Google Scholar] [CrossRef]
- Malhotra, S.; Sowdhamini, R. Genome-wide survey of DNA-binding proteins in Arabidopsis thaliana: Analysis of distribution and functions. Nucleic Acids Res. 2013, 41, 7212–7219. [Google Scholar] [CrossRef] [PubMed]
- Santerre, A.; Britt, A.B. Cloning of a 3-methyladenine-DNA glycosylase from Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 1994, 91, 2240–2244. [Google Scholar] [CrossRef] [PubMed]
- Fang, H.; Huangfu, L.; Chen, R.; Li, P.; Xu, S.; Zhang, E.; Cao, W.; Liu, L.; Yao, Y.; Liang, G.; et al. Ancestor of land plants acquired the DNA-3-methyladenine glycosylase (MAG) gene from bacteria through horizontal gene transfer. Sci. Rep. 2017, 7, 9324. [Google Scholar] [CrossRef]
- Shi, L.; Kent, R.; Bence, N.; Britt, A.B. Developmental expression of a DNA repair gene in Arabidopsis. Mutat. Res. 1997, 384, 145–156. [Google Scholar] [CrossRef]
- Klepikova, A.V.; Kasianov, A.S.; Gerasimov, E.S.; Logacheva, M.D.; Penin, A.A. A high resolution map of the Arabidopsis thaliana developmental transcriptome based on RNA-seq profiling. Plant J. 2016, 88, 1058–1070. [Google Scholar] [CrossRef]
- Mol, C.D.; Hosfield, D.J.; Tainer, J.A. Abasic site recognition by two apurinic/apyrimidinic endonuclease families in DNA base excision repair: The 3′ ends justify the means. Mutat. Res. 2000, 460, 211–229. [Google Scholar] [CrossRef] [PubMed]
- Wiederhold, L.; Leppard, J.B.; Kedar, P.; Karimi-Busheri, F.; Rasouli-Nia, A.; Weinfeld, M.; Tomkinson, A.E.; Izumi, T.; Prasad, R.; Wilson, S.H.; et al. AP endonuclease-independent DNA base excision repair in human cells. Mol. Cell 2004, 15, 209–220. [Google Scholar] [CrossRef] [PubMed]
- Weinfeld, M.; Mani, R.S.; Abdou, I.; Aceytuno, R.D.; Glover, J.N.M. Tidying up loose ends: The role of polynucleotide kinase/phosphatase in DNA strand break repair. Trends Biochem. Sci. 2011, 36, 262–271. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.A.; Lin, Y.; Yan, S. Single-strand break end resection in genome integrity: Mechanism and regulation by APE2. Int. J. Mol. Sci. 2018, 19, 2389. [Google Scholar] [CrossRef] [PubMed]
- Babiychuk, E.; Kushnir, S.; Van Montagu, M.; Inzé, D. The Arabidopsis thaliana apurinic endonuclease Arp reduces human transcription factors Fos and Jun. Proc. Natl. Acad. Sci. USA 1994, 91, 3299–3303. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Córdoba-Cañero, D.; Qian, W.; Zhu, X.; Tang, K.; Zhang, H.; Ariza, R.R.; Roldán-Arjona, T.; Zhu, J.-K. An AP endonuclease functions in active DNA demethylation and gene imprinting in Arabidopsis. PLoS Genet. 2015, 11, e1004905. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Liang, W.; Li, Y.; Qian, W. APURINIC/APYRIMIDINIC ENDONUCLEASE2 and ZINC FINGER DNA 3′-PHOSPHOESTERASE play overlapping roles in the maintenance of epigenome and genome stability. Plant Cell 2018, 30, 1954–1970. [Google Scholar] [CrossRef]
- Murphy, T.M.; Belmonte, M.; Shu, S.; Britt, A.B.; Hatteroth, J. Requirement for abasic endonuclease gene homologues in Arabidopsis seed development. PLoS ONE 2009, 4, e4297. [Google Scholar] [CrossRef]
- Li, J.; Wang, C.; Liang, W.; Zhang, J.; Jiang, C.-K.; Liu, Y.; Ren, Z.; Ci, D.; Chang, J.; Han, S.; et al. Functional importance and divergence of plant apurinic/apyrimidinic endonucleases in somatic and meiotic DNA repair. Plant Cell 2023, 35, 2316–2331. [Google Scholar] [CrossRef]
- Endutkin, A.V.; Yudkina, A.V.; Sidorenko, V.S.; Zharkov, D.O. Transient protein–protein complexes in base excision repair. J. Biomol. Struct. Dyn. 2019, 37, 4407–4418. [Google Scholar] [CrossRef]
- Córdoba-Cañero, D.; Roldán-Arjona, T.; Ariza, R.R. Arabidopsis ARP endonuclease functions in a branched base excision DNA repair pathway completed by LIG1. Plant J. 2011, 68, 693–702. [Google Scholar] [CrossRef] [PubMed]
- Akishev, Z.; Taipakova, S.; Joldybayeva, B.; Zutterling, C.; Smekenov, I.; Ishchenko, A.A.; Zharkov, D.O.; Bissenbaev, A.K.; Saparbaev, M. The major Arabidopsis thaliana apurinic/apyrimidinic endonuclease, ARP is involved in the plant nucleotide incision repair pathway. DNA Repair 2016, 48, 30–42. [Google Scholar] [CrossRef] [PubMed]
- Cabral Medeiros, N.M.; Córdoba-Cañero, D.; García-Gil, C.B.; Ariza, R.R.; Roldán-Arjona, T.; Scortecci, K.C. Characterization of an AP endonuclease from sugarcane—ScARP1. Biochem. Biophys. Res. Commun. 2019, 514, 926–932. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Macías, M.I.; Qian, W.; Miki, D.; Pontes, O.; Liu, Y.; Tang, K.; Liu, R.; Morales-Ruiz, T.; Ariza, R.R.; Roldán-Arjona, T.; et al. A DNA 3′ phosphatase functions in active DNA demethylation in Arabidopsis. Mol. Cell 2012, 45, 357–370. [Google Scholar] [CrossRef] [PubMed]
- Tell, G.; Fantini, D.; Quadrifoglio, F. Understanding different functions of mammalian AP endonuclease (APE1) as a promising tool for cancer treatment. Cell. Mol. Life Sci. 2010, 67, 3589–3608. [Google Scholar] [CrossRef] [PubMed]
- Bazlekowa-Karaban, M.; Prorok, P.; Baconnais, S.; Taipakova, S.; Akishev, Z.; Zembrzuska, D.; Popov, A.V.; Endutkin, A.V.; Groisman, R.; Ishchenko, A.A.; et al. Mechanism of stimulation of DNA binding of the transcription factors by human apurinic/apyrimidinic endonuclease 1, APE1. DNA Repair 2019, 82, 102698. [Google Scholar] [CrossRef] [PubMed]
- Popov, A.V.; Grin, I.R.; Dvornikova, A.P.; Matkarimov, B.T.; Groisman, R.; Saparbaev, M.; Zharkov, D.O. Reading targeted DNA damage in the active demethylation pathway: Role of accessory domains of eukaryotic AP endonucleases and thymine-DNA glycosylases. J. Mol. Biol. 2020, 432, 1747–1768. [Google Scholar] [CrossRef] [PubMed]
- Petrucco, S.; Volpi, G.; Bolchi, A.; Rivetti, C.; Ottonello, S. A nick-sensing DNA 3′-repair enzyme from Arabidopsis. J. Biol. Chem. 2002, 277, 23675–23683. [Google Scholar] [CrossRef]
- Ali, A.A.E.; Jukes, R.M.; Pearl, L.H.; Oliver, A.W. Specific recognition of a multiply phosphorylated motif in the DNA repair scaffold XRCC1 by the FHA domain of human PNK. Nucleic Acids Res. 2009, 37, 1701–1712. [Google Scholar] [CrossRef]
- Ali, A.A.E.; Timinszky, G.; Arribas-Bosacoma, R.; Kozlowski, M.; Hassa, P.O.; Hassler, M.; Ladurner, A.G.; Pearl, L.H.; Oliver, A.W. The zinc-finger domains of PARP1 cooperate to recognize DNA strand breaks. Nat. Struct. Mol. Biol. 2012, 19, 685–692. [Google Scholar] [CrossRef]
- Eustermann, S.; Wu, W.-F.; Langelier, M.-F.; Yang, J.-C.; Easton, L.E.; Riccio, A.A.; Pascal, J.M.; Neuhaus, D. Structural basis of detection and signaling of DNA single-strand breaks by human PARP-1. Mol. Cell 2015, 60, 742–754. [Google Scholar] [CrossRef] [PubMed]
- Gehring, M.; Huh, J.H.; Hsieh, T.-F.; Penterman, J.; Choi, Y.; Harada, J.J.; Goldberg, R.B.; Fischer, R.L. DEMETER DNA glycosylase establishes MEDEA polycomb gene self-imprinting by allele-specific demethylation. Cell 2006, 124, 495–506. [Google Scholar] [CrossRef] [PubMed]
- Jordano-Raya, M.; Beltrán-Melero, C.; Moreno-Recio, M.D.; Martínez-Macías, M.I.; Ariza, R.R.; Roldán-Arjona, T.; Córdoba-Cañero, D. Complementary functions of plant AP endonucleases and AP lyases during DNA repair of abasic sites arising from C:G base pairs. Int. J. Mol. Sci. 2021, 22, 8763. [Google Scholar] [CrossRef] [PubMed]
- Li, A.; Schuermann, D.; Gallego, F.; Kovalchuk, I.; Tinland, B. Repair of damaged DNA by Arabidopsis cell extract. Plant Cell 2002, 14, 263–273. [Google Scholar] [CrossRef] [PubMed]
- García-Díaz, M.; Domínguez, O.; López-Fernández, L.A.; de Lera, L.T.; Saníger, M.L.; Ruiz, J.F.; Párraga, M.; García-Ortiz, M.J.; Kirchhoff, T.; del Mazo, J.; et al. DNA polymerase lambda (Pol λ), a novel eukaryotic DNA polymerase with a potential role in meiosis. J. Mol. Biol. 2000, 301, 851–867. [Google Scholar] [CrossRef] [PubMed]
- Uchiyama, Y.; Kimura, S.; Yamamoto, T.; Ishibashi, T.; Sakaguchi, K. Plant DNA polymerase λ, a DNA repair enzyme that functions in plant meristematic and meiotic tissues. Eur. J. Biochem. 2004, 271, 2799–2807. [Google Scholar] [CrossRef] [PubMed]
- Roy, S.; Choudhury, S.R.; Singh, S.K.; Das, K.P. AtPolλ, a homolog of mammalian DNA polymerase λ in Arabidopsis thaliana, is involved in the repair of UV-B induced DNA damage through the dark repair pathway. Plant Cell Physiol. 2011, 52, 448–467. [Google Scholar] [CrossRef] [PubMed]
- Roy, S.; Choudhury, S.R.; Singh, S.K.; Das, K.P. Functional analysis of light-regulated promoter region of AtPolλ gene. Planta 2012, 235, 411–432. [Google Scholar] [CrossRef]
- Roy, S.; Choudhury, S.R.; Sengupta, D.N.; Das, K.P. Involvement of AtPolλ in the repair of high salt- and DNA cross-linking agent-induced double strand breaks in Arabidopsis. Plant Physiol. 2013, 162, 1195–1210. [Google Scholar] [CrossRef]
- van Loon, B.; Hübscher, U. An 8-oxo-guanine repair pathway coordinated by MUTYH glycosylase and DNA polymerase λ. Proc. Natl. Acad. Sci. USA 2009, 106, 18201–18206. [Google Scholar] [CrossRef]
- Amoroso, A.; Concia, L.; Maggio, C.; Raynaud, C.; Bergounioux, C.; Crespan, E.; Cella, R.; Maga, G. Oxidative DNA damage bypass in Arabidopsis thaliana requires DNA polymerase λ and proliferating cell nuclear antigen 2. Plant Cell 2011, 23, 806–822. [Google Scholar] [CrossRef]
- Zhang, J.; Xie, S.; Cheng, J.; Lai, J.; Zhu, J.-K.; Gong, Z. The second subunit of DNA polymerase delta is required for genomic stability and epigenetic regulation. Plant Physiol. 2016, 171, 1192–1208. [Google Scholar] [CrossRef] [PubMed]
- Fortini, P.; Dogliotti, E. Base damage and single-strand break repair: Mechanisms and functional significance of short- and long-patch repair subpathways. DNA Repair 2007, 6, 398–409. [Google Scholar] [CrossRef] [PubMed]
- Finger, L.D.; Atack, J.M.; Tsutakawa, S.; Classen, S.; Tainer, J.; Grasby, J.; Shen, B. The wonders of flap endonucleases: Structure, function, mechanism and regulation. Subcell. Biochem. 2012, 62, 301–326. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wen, C.; Liu, S.; Zheng, L.; Shen, B.; Tao, Y. Shade avoidance 6 encodes an Arabidopsis flap endonuclease required for maintenance of genome integrity and development. Nucleic Acids Res. 2016, 44, 1271–1284. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Xie, S.; Zhu, J.-K.; Gong, Z. Requirement for flap endonuclease 1 (FEN1) to maintain genomic stability and transcriptional gene silencing in Arabidopsis. Plant J. 2016, 87, 629–640. [Google Scholar] [CrossRef] [PubMed]
- Kimura, S.; Kai, M.; Kobayashi, H.; Suzuki, A.; Morioka, H.; Otsuka, E.; Sakaguchi, K. A structure-specific endonuclease from cauliflower (Brassica oleracea var. botrytis) inflorescence. Nucleic Acids Res. 1997, 25, 4970–4976. [Google Scholar] [CrossRef] [PubMed]
- Kimura, S.; Ueda, T.; Hatanaka, M.; Takenouchi, M.; Hashimoto, J.; Sakaguchi, K. Plant homologue of flap endonuclease-1: Molecular cloning, characterization, and evidence of expression in meristematic tissues. Plant Mol. Biol. 2000, 42, 415–427. [Google Scholar] [CrossRef]
- Kimura, S.; Suzuki, T.; Yanagawa, Y.; Yamamoto, T.; Nakagawa, H.; Tanaka, I.; Hashimoto, J.; Sakaguchi, K. Characterization of plant proliferating cell nuclear antigen (PCNA) and flap endonuclease-1 (FEN-1), and their distribution in mitotic and meiotic cell cycles. Plant J. 2001, 28, 643–653. [Google Scholar] [CrossRef]
- Kimura, S.; Furukawa, T.; Kasai, N.; Mori, Y.; Kitamoto, H.K.; Sugawara, F.; Hashimoto, J.; Sakaguchi, K. Functional characterization of two flap endonuclease-1 homologues in rice. Gene 2003, 314, 63–71. [Google Scholar] [CrossRef]
- Waterworth, W.M.; Kozak, J.; Provost, C.M.; Bray, C.M.; Angelis, K.J.; West, C.E. DNA ligase 1 deficient plants display severe growth defects and delayed repair of both DNA single and double strand breaks. BMC Plant Biol. 2009, 9, 79. [Google Scholar] [CrossRef] [PubMed]
- Andreuzza, S.; Li, J.; Guitton, A.-E.; Faure, J.-E.; Casanova, S.; Park, J.-S.; Choi, Y.; Chen, Z.; Berger, F. DNA LIGASE I exerts a maternal effect on seed development in Arabidopsis thaliana. Development 2010, 137, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Duan, C.-G.; Zhu, X.; Qian, W.; Zhu, J.-K. A DNA ligase required for active DNA demethylation and genomic imprinting in Arabidopsis. Cell Res. 2015, 25, 757–760. [Google Scholar] [CrossRef] [PubMed]
- Caldecott, K.W. XRCC1 protein; Form and function. DNA Repair 2019, 81, 102664. [Google Scholar] [CrossRef] [PubMed]
- London, R.E. XRCC1—Strategies for coordinating and assembling a versatile DNA damage response. DNA Repair 2020, 93, 102917. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Macías, M.I.; Córdoba-Cañero, D.; Ariza, R.R.; Roldán-Arjona, T. The DNA repair protein XRCC1 functions in the plant DNA demethylation pathway by stimulating cytosine methylation (5-meC) excision, gap tailoring, and DNA ligation. J. Biol. Chem. 2013, 288, 5496–5505. [Google Scholar] [CrossRef] [PubMed]
- Uchiyama, Y.; Suzuki, Y.; Sakaguchi, K. Characterization of plant XRCC1 and its interaction with proliferating cell nuclear antigen. Planta 2008, 227, 1233–1241. [Google Scholar] [CrossRef] [PubMed]
- Taylor, R.M.; Thistlethwaite, A.; Caldecott, K.W. Central role for the XRCC1 BRCT I domain in mammalian DNA single-strand break repair. Mol. Cell. Biol. 2002, 22, 2556–2563. [Google Scholar] [CrossRef]
- Cuneo, M.J.; Gabel, S.A.; Krahn, J.M.; Ricker, M.A.; London, R.E. The structural basis for partitioning of the XRCC1/DNA ligase III-α BRCT-mediated dimer complexes. Nucleic Acids Res. 2011, 39, 7816–7827. [Google Scholar] [CrossRef]
- Charbonnel, C.; Gallego, M.E.; White, C.I. Xrcc1-dependent and Ku-dependent DNA double-strand break repair kinetics in Arabidopsis plants. Plant J. 2010, 64, 280–290. [Google Scholar] [CrossRef]
- Alemasova, E.E.; Lavrik, O.I. Poly(ADP-ribosyl)ation by PARP1: Reaction mechanism and regulatory proteins. Nucleic Acids Res. 2019, 47, 3811–3827. [Google Scholar] [CrossRef] [PubMed]
- Suskiewicz, M.J.; Palazzo, L.; Hughes, R.; Ahel, I. Progress and outlook in studying the substrate specificities of PARPs and related enzymes. FEBS J. 2021, 288, 2131–2142. [Google Scholar] [CrossRef] [PubMed]
- Feng, B.; Liu, C.; de Oliveira, M.V.V.; Intorne, A.C.; Li, B.; Babilonia, K.; de Souza Filho, G.A.; Shan, L.; He, P. Protein poly(ADP-ribosyl)ation regulates Arabidopsis immune gene expression and defense responses. PLoS Genet. 2015, 11, e1004936. [Google Scholar] [CrossRef] [PubMed]
- Gu, Z.; Pan, W.; Chen, W.; Lian, Q.; Wu, Q.; Lv, Z.; Cheng, X.; Ge, X. New perspectives on the plant PARP family: Arabidopsis PARP3 is inactive, and PARP1 exhibits predominant poly (ADP-ribose) polymerase activity in response to DNA damage. BMC Plant Biol. 2019, 19, 364. [Google Scholar] [CrossRef] [PubMed]
- Boltz, K.A.; Jasti, M.; Townley, J.M.; Shippen, D.E. Analysis of poly(ADP-Ribose) polymerases in Arabidopsis telomere biology. PLoS ONE 2014, 9, e88872. [Google Scholar] [CrossRef] [PubMed]
- Rissel, D.; Losch, J.; Peiter, E. The nuclear protein Poly(ADP-ribose) polymerase 3 (AtPARP3) is required for seed storability in Arabidopsis thaliana. Plant Biol. 2014, 16, 1058–1064. [Google Scholar] [CrossRef] [PubMed]
- Prakash, A.; Doublié, S. Base excision repair in the mitochondria. J. Cell. Biochem. 2015, 116, 1490–1499. [Google Scholar] [CrossRef] [PubMed]
- Akbari, M.; Nilsen, H.L.; Montaldo, N.P. Dynamic features of human mitochondrial DNA maintenance and transcription. Front. Cell Dev. Biol. 2022, 10, 984245. [Google Scholar] [CrossRef]
- Boesch, P.; Ibrahim, N.; Paulus, F.; Cosset, A.; Tarasenko, V.; Dietrich, A. Plant mitochondria possess a short-patch base excision DNA repair pathway. Nucleic Acids Res. 2009, 37, 5690–5700. [Google Scholar] [CrossRef]
- Ferrando, B.; Furlanetto, A.L.D.M.; Gredilla, R.; Havelund, J.F.; Hebelstrup, K.H.; Møller, I.M.; Stevnsner, T. DNA repair in plant mitochondria—A complete base excision repair pathway in potato tuber mitochondria. Physiol. Plant. 2019, 166, 494–512. [Google Scholar] [CrossRef]
- Szczesny, B.; Hazra, T.K.; Papaconstantinou, J.; Mitra, S.; Boldogh, I. Age-dependent deficiency in import of mitochondrial DNA glycosylases required for repair of oxidatively damaged bases. Proc. Natl. Acad. Sci. USA 2003, 100, 10670–10675. [Google Scholar] [CrossRef] [PubMed]
- Panduri, V.; Liu, G.; Surapureddi, S.; Kondapalli, J.; Soberanes, S.; de Souza-Pinto, N.C.; Bohr, V.A.; Budinger, G.R.S.; Schumacker, P.T.; Weitzman, S.A.; et al. Role of mitochondrial hOGG1 and aconitase in oxidant-induced lung epithelial cell apoptosis. Free Radic. Biol. Med. 2009, 47, 750–759. [Google Scholar] [CrossRef]
- Chapman, J.; Ng, Y.S.; Nicholls, T.J. The maintenance of mitochondrial DNA integrity and dynamics by mitochondrial membranes. Life 2020, 10, 164. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Waneka, G.; Broz, A.K.; King, C.R.; Sloan, D.B. MSH1 is required for maintenance of the low mutation rates in plant mitochondrial and plastid genomes. Proc. Natl. Acad. Sci. USA 2020, 117, 16448–16455. [Google Scholar] [CrossRef] [PubMed]
- Furlanetto, A.L.D.M.; Cadena, S.M.S.C.; Martinez, G.R.; Ferrando, B.; Stevnsner, T.; Møller, I.M. Short-term high temperature treatment reduces viability and inhibits respiration and DNA repair enzymes in Araucaria angustifolia cells. Physiol. Plant. 2019, 166, 513–524. [Google Scholar] [CrossRef] [PubMed]
- Kimura, S.; Uchiyama, Y.; Kasai, N.; Namekawa, S.; Saotome, A.; Ueda, T.; Ando, T.; Ishibashi, T.; Oshige, M.; Furukawa, T.; et al. A novel DNA polymerase homologous to Escherichia coli DNA polymerase I from a higher plant, rice (Oryza sativa L.). Nucleic Acids Res. 2002, 30, 1585–1592. [Google Scholar] [CrossRef]
- Mori, Y.; Kimura, S.; Saotome, A.; Kasai, N.; Sakaguchi, N.; Uchiyama, Y.; Ishibashi, T.; Yamamoto, T.; Chiku, H.; Sakaguchi, K. Plastid DNA polymerases from higher plants, Arabidopsis thaliana. Biochem. Biophys. Res. Commun. 2005, 334, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Ono, Y.; Sakai, A.; Takechi, K.; Takio, S.; Takusagawa, M.; Takano, H. NtPolI-like1 and NtPolI-like2, bacterial DNA polymerase I homologs isolated from BY-2 cultured tobacco cells, encode DNA polymerases engaged in DNA replication in both plastids and mitochondria. Plant Cell Physiol. 2007, 48, 1679–1692. [Google Scholar] [CrossRef]
- Parent, J.-S.; Lepage, E.; Brisson, N. Divergent roles for the two PolI-like organelle DNA polymerases of Arabidopsis. Plant Physiol. 2011, 156, 254–262. [Google Scholar] [CrossRef]
- Trasviña-Arenas, C.H.; Baruch-Torres, N.; Cordoba-Andrade, F.J.; Ayala-García, V.M.; García-Medel, P.L.; Díaz-Quezada, C.; Peralta-Castro, A.; Ordaz-Ortiz, J.J.; Brieba, L.G. Identification of a unique insertion in plant organellar DNA polymerases responsible for 5′-dRP lyase and strand-displacement activities: Implications for Base Excision Repair. DNA Repair 2018, 65, 1–10. [Google Scholar] [CrossRef]
- Chan, K.K.L.; Zhang, Q.-M.; Dianov, G.L. Base excision repair fidelity in normal and cancer cells. Mutagenesis 2006, 21, 173–178. [Google Scholar] [CrossRef] [PubMed]
- Ayala-García, V.M.; Baruch-Torres, N.; García-Medel, P.L.; Brieba, L.G. Plant organellar DNA polymerases paralogs exhibit dissimilar nucleotide incorporation fidelity. FEBS J. 2018, 285, 4005–4018. [Google Scholar] [CrossRef]
- Bensen, R.J.; Warner, H.R. Partial purification and characterization of uracil-DNA glycosylase activity from chloroplasts of Zea mays seedlings. Plant Physiol. 1987, 84, 1102–1106. [Google Scholar] [CrossRef]
- Sunderland, P.A.; West, C.E.; Waterworth, W.M.; Bray, C.M. An evolutionarily conserved translation initiation mechanism regulates nuclear or mitochondrial targeting of DNA ligase 1 in Arabidopsis thaliana. Plant J. 2006, 47, 356–367. [Google Scholar] [CrossRef]
- Ferro, M.; Brugière, S.; Salvi, D.; Seigneurin-Berny, D.; Court, M.; Moyet, L.; Ramus, C.; Miras, S.; Mellal, M.; Le Gall, S.; et al. AT_CHLORO, a comprehensive chloroplast proteome database with subplastidial localization and curated information on envelope proteins. Mol. Cell. Proteom. 2010, 9, 1063–1084. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
General scheme of short- and long-patch BER.

Figure 2.
Chemistry of the initial BER steps. Only one DNA strand is shown for clarity. Monofunctional DNA glycosylases only hydrolyze the N-glycosidic bond (i), whereas bifunctional DNA glycosylases possess the AP lyase activity that catalyzes β-elimination (ii) or β,δ-elimination (iii). After the action of monofunctional glycosylases, AP endonucleases cleave the phosphodiester bond 5′ to the AP site (iv). After β-elimination, the 3′-phosphodiesterase activity of AP endonucleases removes the 3′-terminal α,β-unsaturated aldehyde (v). After β,δ-elimination, the 3′-phosphatase activity, either standalone or residing in AP endonucleases, removes the 3′-phosphate (vi).
Figure 2.
Chemistry of the initial BER steps. Only one DNA strand is shown for clarity. Monofunctional DNA glycosylases only hydrolyze the N-glycosidic bond (i), whereas bifunctional DNA glycosylases possess the AP lyase activity that catalyzes β-elimination (ii) or β,δ-elimination (iii). After the action of monofunctional glycosylases, AP endonucleases cleave the phosphodiester bond 5′ to the AP site (iv). After β-elimination, the 3′-phosphodiesterase activity of AP endonucleases removes the 3′-terminal α,β-unsaturated aldehyde (v). After β,δ-elimination, the 3′-phosphatase activity, either standalone or residing in AP endonucleases, removes the 3′-phosphate (vi).

Figure 3.
Unrooted tree with 989 plant DML protein sequences. NCBI RefSeq protein database [90] was searched using AtDME, AtROS1, AtDML2, and AtDML3 as queries, limited to the Viridiplantae kingdom. Only the sequences containing the HhH motif, the iron–sulfur cluster, the zinc finger, and the RRM motif were retained. Nineteen plant NTH proteins were taken as an outgroup. The sequences were aligned with MUSCLE [91], and the tree was built with the PHYLIP neighbor-joining method [92], as implemented in UGENE v48 [93]. The tree was visualized using iTOL [94]; the Newick file can be found in the Supplementary Materials (Grin_et_al_Fig_3.nwk).
Figure 3.
Unrooted tree with 989 plant DML protein sequences. NCBI RefSeq protein database [90] was searched using AtDME, AtROS1, AtDML2, and AtDML3 as queries, limited to the Viridiplantae kingdom. Only the sequences containing the HhH motif, the iron–sulfur cluster, the zinc finger, and the RRM motif were retained. Nineteen plant NTH proteins were taken as an outgroup. The sequences were aligned with MUSCLE [91], and the tree was built with the PHYLIP neighbor-joining method [92], as implemented in UGENE v48 [93]. The tree was visualized using iTOL [94]; the Newick file can be found in the Supplementary Materials (Grin_et_al_Fig_3.nwk).

Figure 4.
Unrooted tree with 1064 plant TAG protein sequences. NCBI RefSeq protein database [90] was searched using E. coli Tag and seven A. thaliana TAG homologs (marked red, Table 1) as queries, limited to the Viridiplantae kingdom. The sequences were aligned with MUSCLE [91], and the tree was built with the PHYLIP neighbor-joining method [92], as implemented in UGENE v48 [93]. The tree was visualized using iTOL [94]; the Newick file can be found in the Supplementary Materials (Grin_et_al_Fig_4.nwk).
Figure 4.
Unrooted tree with 1064 plant TAG protein sequences. NCBI RefSeq protein database [90] was searched using E. coli Tag and seven A. thaliana TAG homologs (marked red, Table 1) as queries, limited to the Viridiplantae kingdom. The sequences were aligned with MUSCLE [91], and the tree was built with the PHYLIP neighbor-joining method [92], as implemented in UGENE v48 [93]. The tree was visualized using iTOL [94]; the Newick file can be found in the Supplementary Materials (Grin_et_al_Fig_4.nwk).


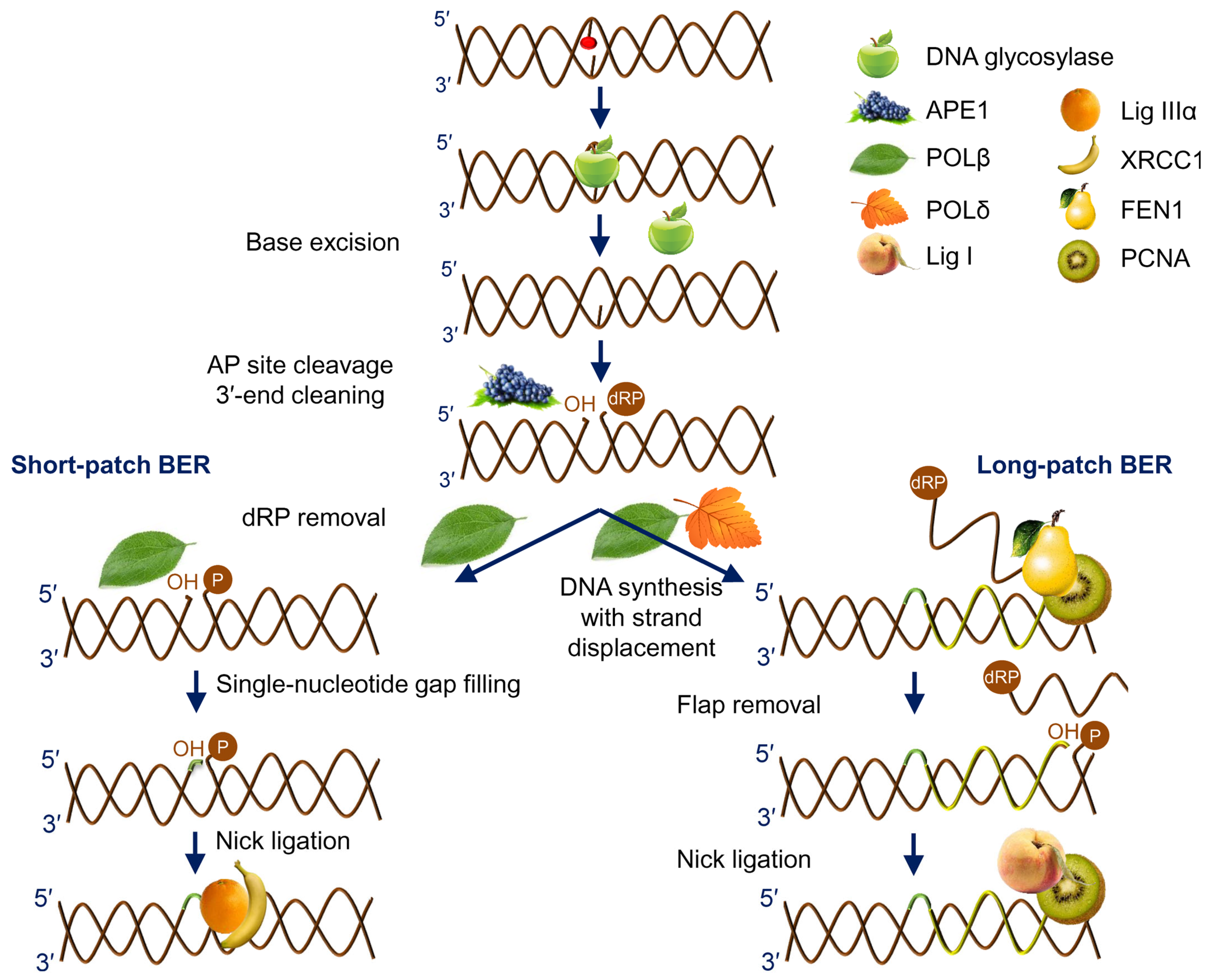{kind=link}
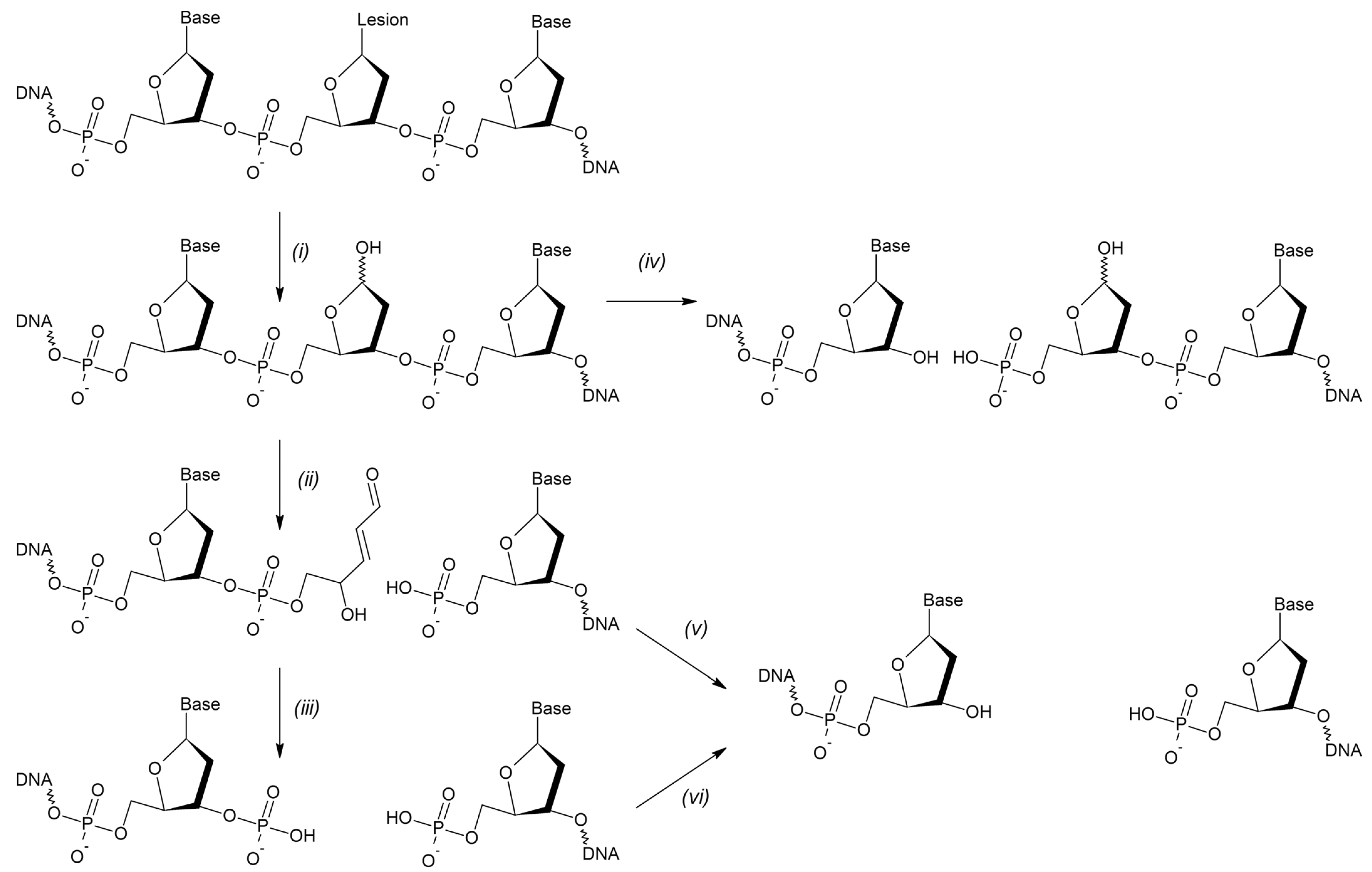{kind=link}
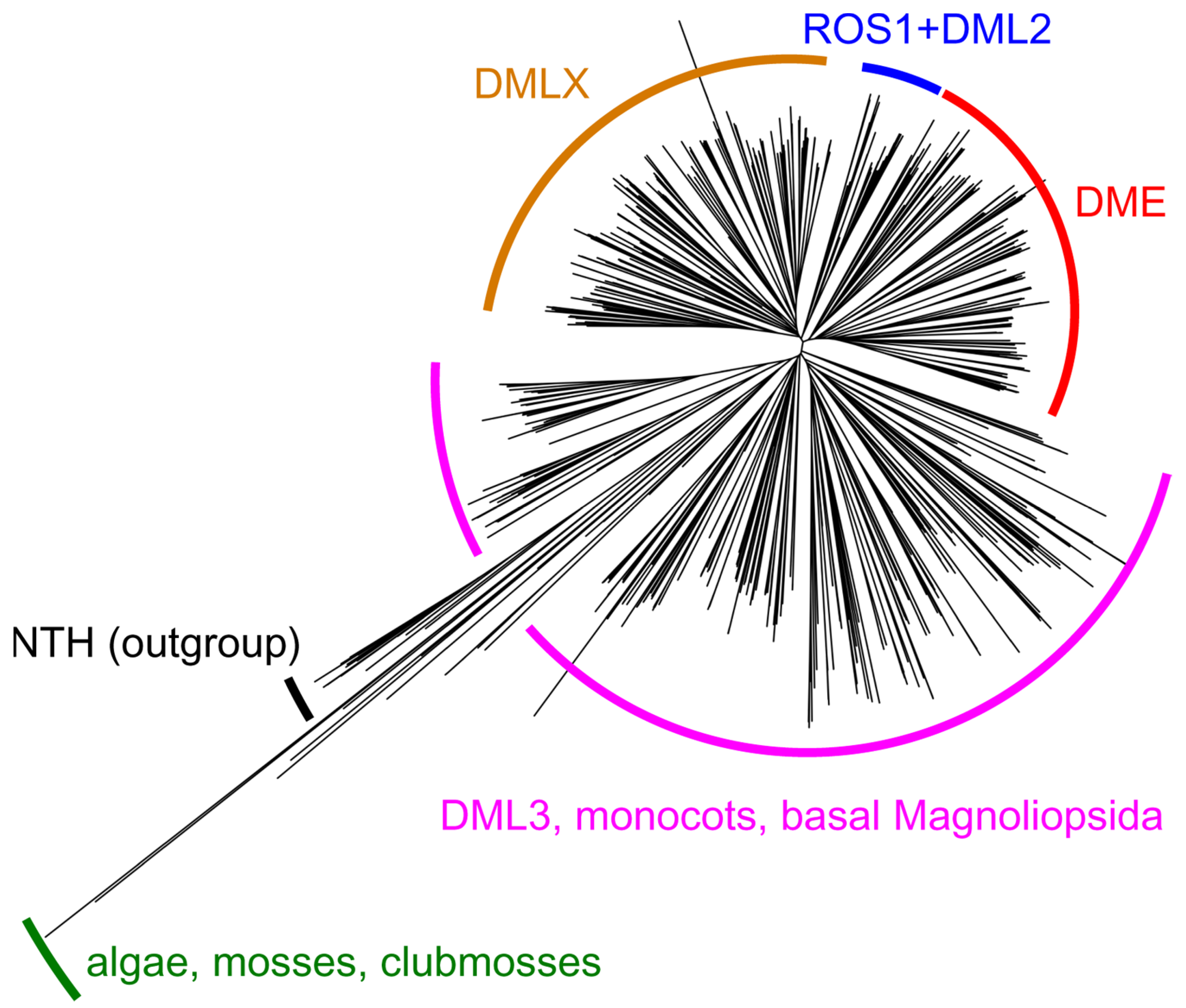{kind=link}
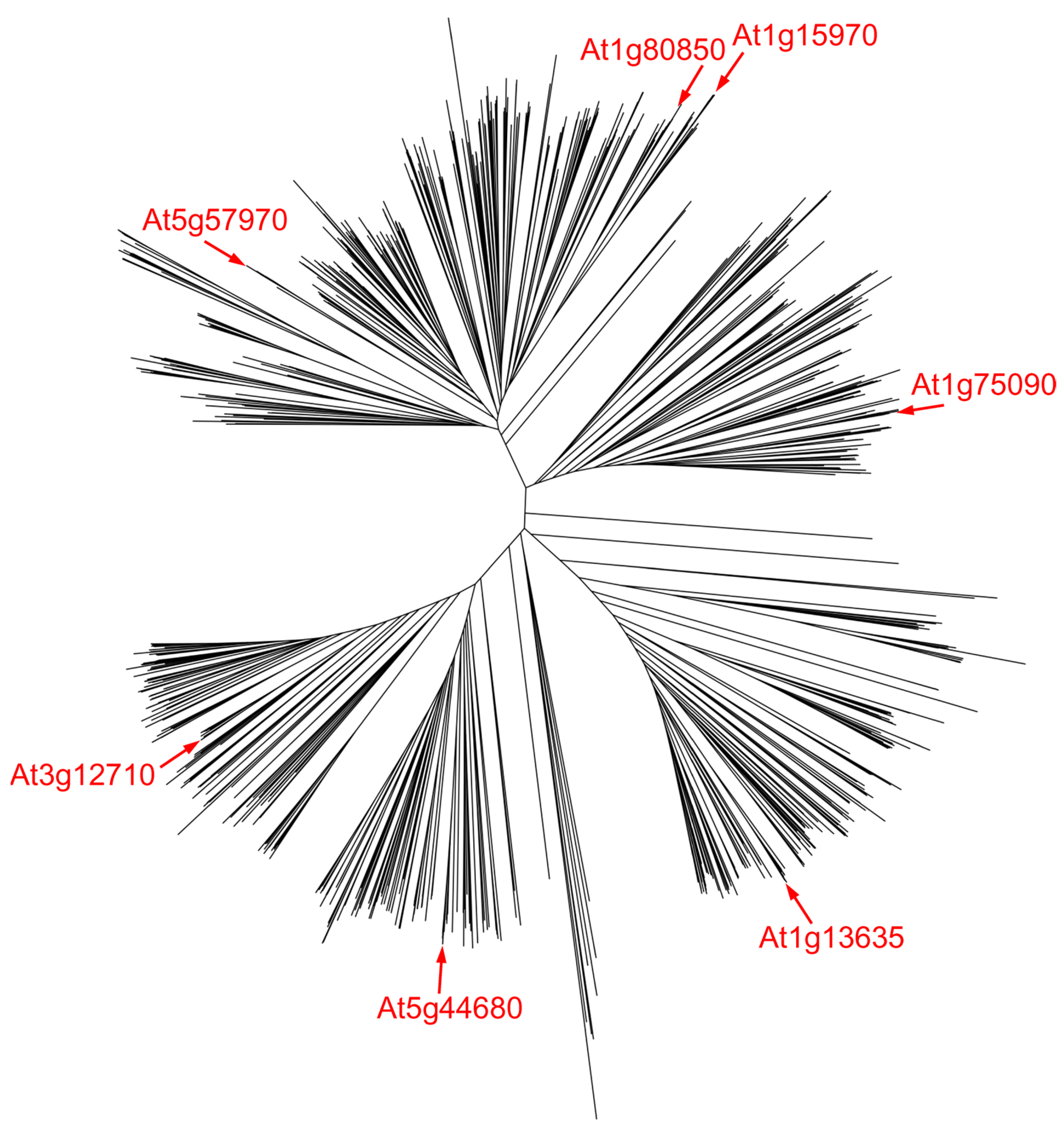{kind=link}
Table 1.
Principal BER proteins in E. coli, humans, and Arabidopsis.
| Enzyme | E. coli | Human | A. thaliana |
|---|---|---|---|
| Uracil–DNA glycosylases | Ung | UNG | At3g18630 (AtUNG) |
| Mug | TDG | ||
| SMUG1 | |||
| Endonuclease III | Nth | NTHL1 | At2g31450 (AtNTH1) |
| At1g05900 (AtNTH2) | |||
| A:oxoG-specific adenine–DNA glycosylase | MutY | MUTYH | At4g12740 (AtMYH) |
| 8-oxoguanine–DNA glycosylase | – | OGG1 | At1g21710 (AtOGG1) |
| Bacterial alkylpurine–DNA glycosylase I | Tag | – | At1g13635 |
| At1g15970 | |||
| At1g75090 | |||
| At1g80850 | |||
| At3g12710 | |||
| At5g44680 | |||
| At5g57970 | |||
| Bacterial alkylpurine–DNA glycosylase II | AlkA | – | At1g19480 |
| At1g75230 | |||
| At3g50880 | |||
| Human alkylpurine–DNA glycosylase | – | MPG | At3g12040 (AtMAG) |
| Methyl-CpG-specific uracil–DNA glycosylase | – | MBD4 | At3g07930 (AtMBD4) |
| Formamidopyrimidine–DNA glycosylase/endonuclease VIII | Fpg | At3g07930 (AtMMH) | |
| Nei | NEIL1 | ||
| NEIL2 | |||
| NEIL3 | |||
| Epigenetic 5-methylcytosine–DNA glycosylases | – | – | At5g04560 (AtDME) |
| At2g36490 (AtROS1) | |||
| At3g10010 (AtDML2) | |||
| At4g34060 (AtDML3) | |||
| At3g47830 | |||
| AP endonucleases, EEP superfamily | Xth | APE1 | At2g41460 (AtARP) |
| At3g48425 (AtAPE1L) | |||
| APE2 | At4g36050 (AtAPE2) | ||
| AP endonucleases, TIM barrel superfamily | Nfo | – | – |
| 3′-phosphatase | – | PNKP | At3g14890 (AtZDP) |
| DNA polymerases, Family A | PolI | POLγ | At1g50840 (AtPolIA) |
| At3g20540 (AtPolIB) | |||
| DNA polymerases, Family X | – | POLβ | |
| POLλ | At1g10520 (AtPOLλ) | ||
| Flap endonuclease | PolI | FEN1 | At5g26680 (AtFEN1) |
| DNA ligase | LigA | Lig I | At1g08130 (AtLIG1) |
| At1g49250 | |||
| Lig IIIα | |||
| BRCT domain scaffold protein | – | XRCC1 | At1g80420 (AtXRCC1) |
| Poly(ADP-ribose) polymerases | – | PARP1 | At2g31320 (AtPARP1) |
| PARP2 | At4g02390 (AtPARP2) | ||
| PARP3 | At5g22470 (AtPARP3) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Grin, I.R.; Petrova, D.V.; Endutkin, A.V.; Ma, C.; Yu, B.; Li, H.; Zharkov, D.O. Base Excision DNA Repair in Plants: Arabidopsis and Beyond. Int. J. Mol. Sci. 2023, 24, 14746. https://doi.org/10.3390/ijms241914746
AMA Style
Grin IR, Petrova DV, Endutkin AV, Ma C, Yu B, Li H, Zharkov DO. Base Excision DNA Repair in Plants: Arabidopsis and Beyond. International Journal of Molecular Sciences. 2023; 24(19):14746. https://doi.org/10.3390/ijms241914746
Chicago/Turabian StyleGrin, Inga R., Daria V. Petrova, Anton V. Endutkin, Chunquan Ma, Bing Yu, Haiying Li, and Dmitry O. Zharkov. 2023. "Base Excision DNA Repair in Plants: Arabidopsis and Beyond" International Journal of Molecular Sciences 24, no. 19: 14746. https://doi.org/10.3390/ijms241914746
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.