
Natural Killer T Cell Diversity and Immunotherapy
 ,
,  , , , and
, , , and
Abstract
:Simple Summary
Abstract
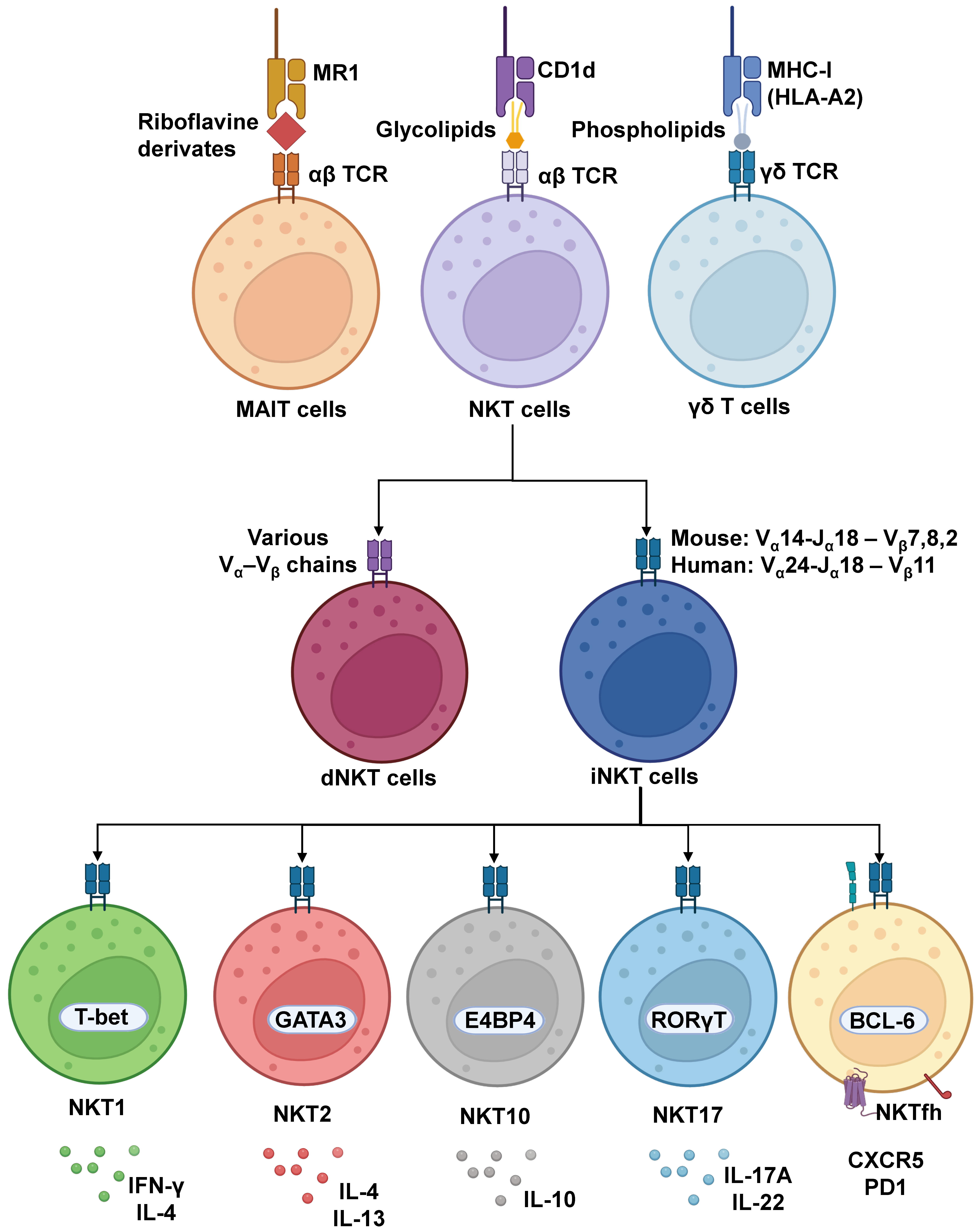
1. Introduction to Unconventional T Cell Subtypes, iNKTs, and CD1d Interactions
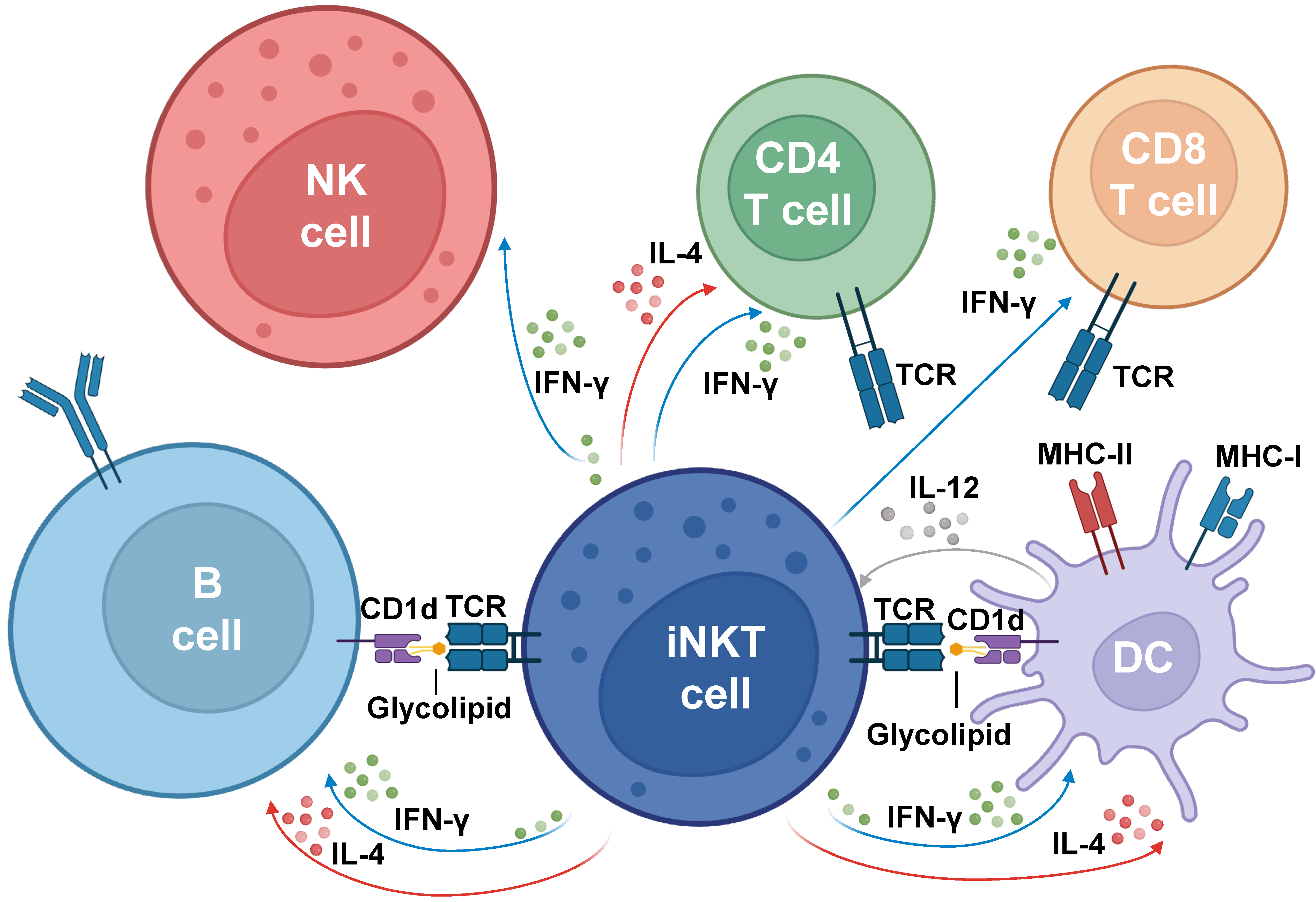
2. Role of iNKTs in Innate and Adaptive Immune Responses
3. α-Galactosylceramide and Its Derivatives as Modulators of iNKT-Dependent T Helper-like Responses
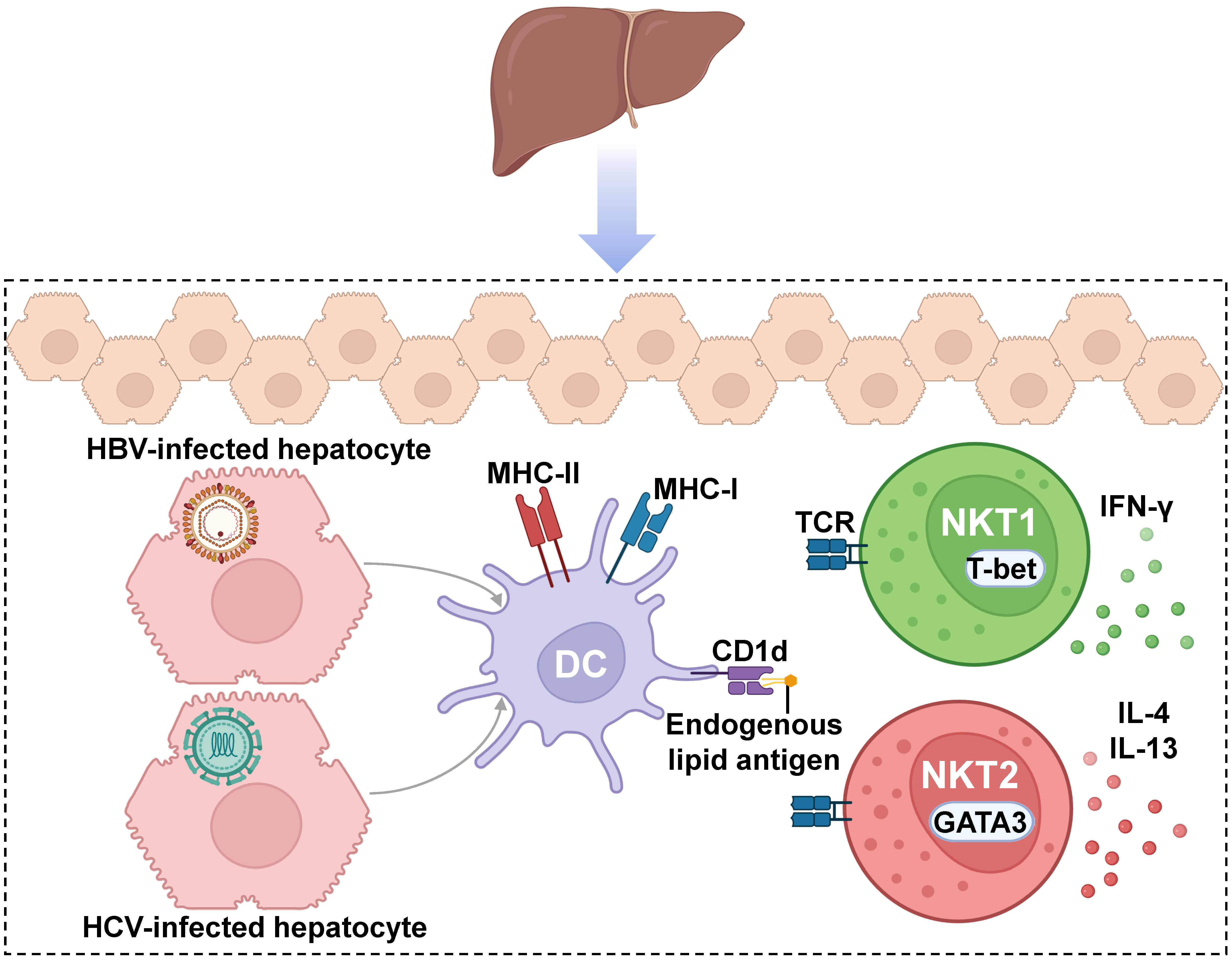
4. Role and iNKT Modulation Strategies in Infectious Diseases and Hepatocarcinoma
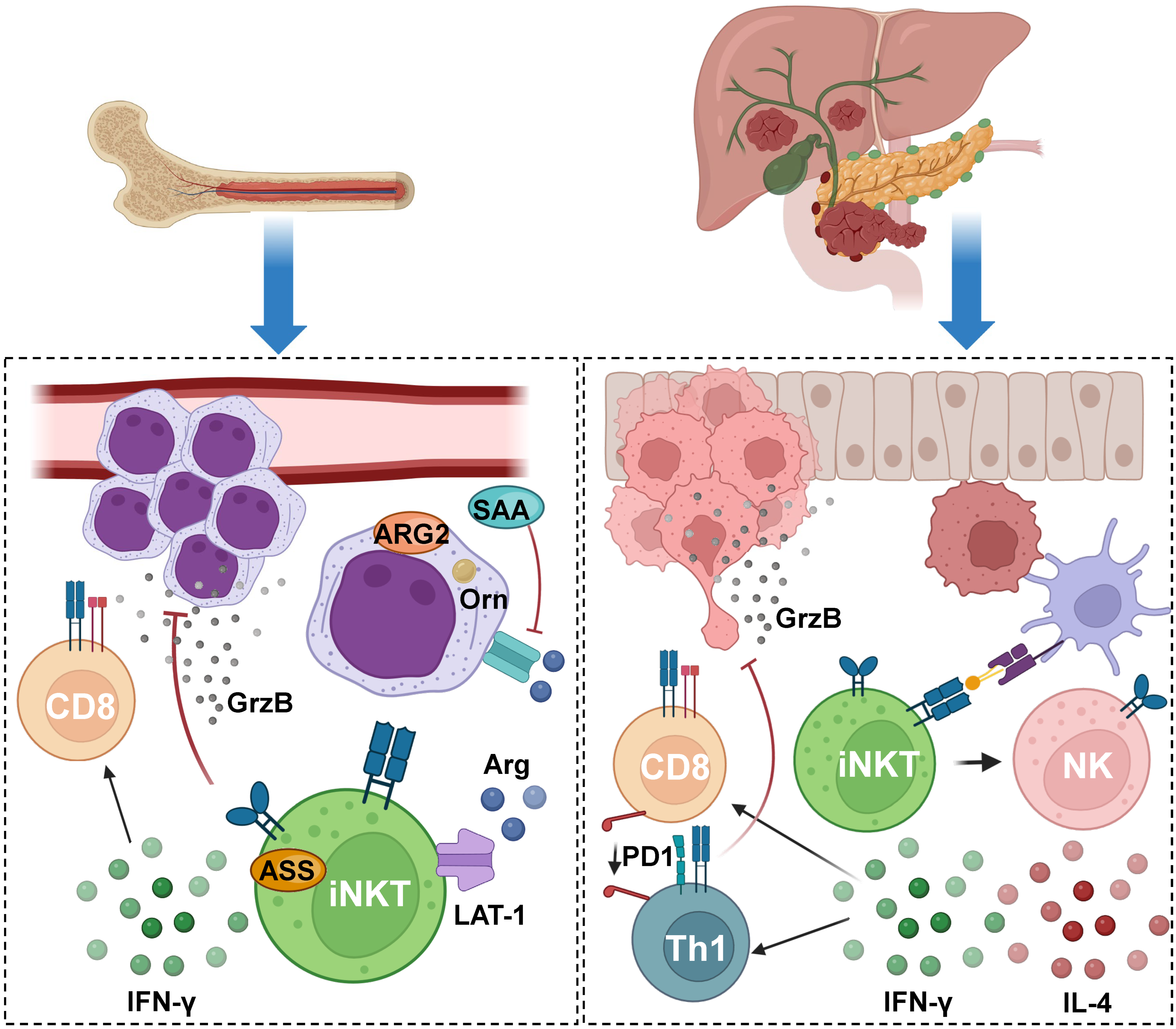
5. Impact of iNKT during the Establishment of Cancer and Progression
6. Current Advances in iNKT-Mediated Cancer Immunotherapy
7. Challenges and Future Prospects of iNKT-Based Immunotherapy
8. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Marshall, J.S.; Warrington, R.; Watson, W.; Kim, H.L. An introduction to immunology and immunopathology. Allergy Asthma Clin. Immunol. 2018, 14, 449. [Google Scholar] [CrossRef]
- Li, X.; Dai, H.; Wang, H.; Han, W. Exploring innate immunity in cancer immunotherapy: Opportunities and challenges. Cell Mol. Immunol. 2021, 18, 1607–1609. [Google Scholar] [CrossRef]
- Diamond, M.S.; Kanneganti, T.-D. Innate immunity: The first line of defense against SARS-CoV-2. Nat. Immunol. 2022, 23, 165–176. [Google Scholar] [CrossRef] [PubMed]
- Labrie, M.; Brugge, J.S.; Mills, G.B.; Zervantonakis, I.K. Therapy resistance: Opportunities created by adaptive responses to targeted therapies in cancer. Nat. Rev. Cancer 2022, 22, 323–339. [Google Scholar] [CrossRef] [PubMed]
- Chang, R.B.; Beatty, G.L. The interplay between innate and adaptive immunity in cancer shapes the productivity of cancer immunosurveillance. J. Leukoc. Biol. 2020, 108, 363–376. [Google Scholar] [CrossRef] [PubMed]
- Kałużna, A.; Olczyk, P.; Komosińska-Vassev, K. The Role of Innate and Adaptive Immune Cells in the Pathogenesis and Development of the Inflammatory Response in Ulcerative Colitis. J. Clin. Med. 2022, 11, 400. [Google Scholar] [CrossRef] [PubMed]
- Ma, W.-T.; Gao, F.; Gu, K.; Chen, D.-K. The Role of Monocytes and Macrophages in Autoimmune Diseases: A Comprehensive Review. Front. Immunol. 2019, 10, 1140. [Google Scholar] [CrossRef] [PubMed]
- Razizadeh, M.H.; Zafarani, A.; Taghavi-Farahabadi, M.; Khorramdelazad, H.; Minaeian, S.; Mahmoudi, M. Natural killer cells and their exosomes in viral infections and related therapeutic approaches: Where are we? Cell Commun. Signal 2023, 21, 261. [Google Scholar] [CrossRef] [PubMed]
- Molfetta, R.; Lecce, M.; Milito, N.D.; Putro, E.; Pietropaolo, G.; Marangio, C.; Scarno, G.; Moretti, M.; De Smaele, E.; Santini, T.; et al. SCF and IL-33 regulate mouse mast cell phenotypic and functional plasticity supporting a pro-inflammatory microenvironment. Cell Death Dis. 2023, 14, 616. [Google Scholar] [CrossRef]
- Li, X.; Xiao, S.; Filipczak, N.; Yalamarty, S.S.K.; Shang, H.; Zhang, J.; Zheng, Q. Role and Therapeutic Targeting Strategies of Neutrophil Extracellular Traps in Inflammation. Int. J. Nanomed. 2023, 18, 5265–5287. [Google Scholar] [CrossRef]
- Santalla Méndez, R.; Rodgers Furones, A.; Classens, R.; Fedorova, K.; Haverdil, M.; Canela Capdevila, M.M.; van Duffelen, A.; Spruijt, C.G.; Vermeulen, M.; ter Beest, M.; et al. Galectin-9 interacts with Vamp-3 to regulate cytokine secretion in dendritic cells. Cell Mol. Life Sci. 2023, 80, 306. [Google Scholar] [CrossRef] [PubMed]
- Fan, X.; Rudensky, A.Y. Hallmarks of Tissue-Resident Lymphocytes. Cell 2016, 164, 1198–1211. [Google Scholar] [CrossRef] [PubMed]
- Artis, D.; Spits, H. The biology of innate lymphoid cells. Nature 2015, 517, 293–301. [Google Scholar] [CrossRef] [PubMed]
- Colonna, M. Innate Lymphoid Cells: Diversity, Plasticity and Unique Functions in Immunity. Immunity 2018, 48, 1104–1117. [Google Scholar] [CrossRef]
- Villarino, A.V.; Sciumè, G.; Davis, F.P.; Iwata, S.; Zitti, B.; Robinson, G.W.; Hennighausen, L.; Kanno, Y.; O’Shea, J.J. Subset- and tissue-defined STAT5 thresholds control homeostasis and function of innate lymphoid cells. J. Exp. Med. 2017, 214, 2999–3014. [Google Scholar] [CrossRef]
- Bennstein, S.B.; Scherenschlich, N.; Weinhold, S.; Manser, A.R.; Noll, A.; Raba, K.; Kögler, G.; Walter, L.; Uhrberg, M. Transcriptional and functional characterization of neonatal circulating Innate Lymphoid Cells. Stem Cells Transl. Med. 2021, 10, 867–882. [Google Scholar] [CrossRef]
- Vivier, E.; Artis, D.; Colonna, M.; Diefenbach, A.; Di Santo, J.P.; Eberl, G.; Koyasu, S.; Locksley, R.M.; McKenzie, A.N.J.; Mebius, R.E.; et al. Innate Lymphoid Cells: 10 Years On. Cell 2018, 174, 1054–1066. [Google Scholar] [CrossRef]
- Korchagina, A.A.; Koroleva, E.; Tumanov, A.V. Innate Lymphoid Cells in Response to Intracellular Pathogens: Protection Versus Immunopathology. Front. Cell. Infect. Microbiol. 2021, 11, 1168. [Google Scholar] [CrossRef]
- Gordon, S.M.; Chaix, J.; Rupp, L.J.; Wu, J.; Madera, S.; Sun, J.C.; Lindsten, T.; Reiner, S.L. The transcription factors T-bet and Eomes control key checkpoints of natural killer cell maturation. Immunity 2012, 36, 55–67. [Google Scholar] [CrossRef]
- Spits, H.; Artis, D.; Colonna, M.; Diefenbach, A.; Di Santo, J.P.; Eberl, G.; Koyasu, S.; Locksley, R.M.; McKenzie, A.N.J.; Mebius, R.E.; et al. Innate lymphoid cells—A proposal for uniform nomenclature. Nat. Rev. Immunol. 2013, 13, 145–149. [Google Scholar] [CrossRef]
- Walker, J.A.; McKenzie, A.N. Development and function of group 2 innate lymphoid cells. Curr. Opin. Immunol. 2013, 25, 148–155. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, A.C.F.; Szeto, A.C.H.; Heycock, M.W.D.; Clark, P.A.; Walker, J.A.; Crisp, A.; Barlow, J.L.; Kitching, S.; Lim, A.; Gogoi, M.; et al. RORα is a critical checkpoint for T cell and ILC2 commitment in the embryonic thymus. Nat. Immunol. 2021, 22, 166–178. [Google Scholar] [CrossRef] [PubMed]
- Buonocore, S.; Ahern, P.P.; Uhlig, H.H.; Ivanov, I.I.; Littman, D.R.; Maloy, K.J.; Powrie, F. Innate lymphoid cells drive interleukin-23-dependent innate intestinal pathology. Nature 2010, 464, 1371–1375. [Google Scholar] [CrossRef] [PubMed]
- Elemam, N.M.; Hannawi, S.; Maghazachi, A.A. Innate Lymphoid Cells (ILCs) as Mediators of Inflammation, Release of Cytokines and Lytic Molecules. Toxins 2017, 9, 398. [Google Scholar] [CrossRef]
- Ruf, B.; Greten, T.F.; Korangy, F. Innate lymphoid cells and innate-like T cells in cancer—At the crossroads of innate and adaptive immunity. Nat. Rev. Cancer 2023, 23, 351–371. [Google Scholar] [CrossRef] [PubMed]
- Park, J.-E.; Botting, R.A.; Conde, C.D.; Popescu, D.-M.; Lavaert, M.; Kunz, D.J.; Goh, I.; Stephenson, E.; Ragazzini, R.; Tuck, E.; et al. A cell atlas of human thymic development defines T cell repertoire formation. Science 2020, 367, eaay3224. [Google Scholar] [CrossRef]
- Pellicci, D.G.; Koay, H.-F.; Berzins, S.P. Thymic development of unconventional T cells: How NKT cells, MAIT cells and γδ T cells emerge. Nat. Rev. Immunol. 2020, 20, 756–770. [Google Scholar] [CrossRef]
- Huang, S. Targeting Innate-Like T Cells in Tuberculosis. Front. Immunol. 2016, 7, 594. [Google Scholar] [CrossRef]
- Mayassi, T.; Barreiro, L.B.; Rossjohn, J.; Jabri, B. A multilayered immune system through the lens of unconventional T cells. Nature 2021, 595, 501–510. [Google Scholar] [CrossRef]
- Deseke, M.; Prinz, I. Ligand recognition by the γδ TCR and discrimination between homeostasis and stress conditions. Cell Mol. Immunol. 2020, 17, 914–924. [Google Scholar] [CrossRef]
- Rhost, S.; Sedimbi, S.; Kadri, N.; Cardell, S.L. Immunomodulatory Type II Natural Killer T Lymphocytes in Health and Disease. Scand. J. Immunol. 2012, 76, 246–255. [Google Scholar] [CrossRef] [PubMed]
- Godfrey, D.I.; Koay, H.-F.; McCluskey, J.; Gherardin, N.A. The biology and functional importance of MAIT cells. Nat. Immunol. 2019, 20, 1110–1128. [Google Scholar] [CrossRef] [PubMed]
- Godfrey, D.I.; Uldrich, A.P.; McCluskey, J.; Rossjohn, J.; Moody, D.B. The burgeoning family of unconventional T cells. Nat. Immunol. 2015, 16, 1114–1123. [Google Scholar] [CrossRef] [PubMed]
- Yazdanifar, M.; Barbarito, G.; Bertaina, A.; Airoldi, I. γδ T Cells: The Ideal Tool for Cancer Immunotherapy. Cells 2020, 9, 1305. [Google Scholar] [CrossRef] [PubMed]
- Karunathilaka, A.; Halstrom, S.; Price, P.; Holt, M.; Lutzky, V.P.; Doolan, D.L.; Kupz, A.; Bell, S.C.; Thomson, R.M.; Miles, J.J.; et al. CD161 expression defines new human γδ T cell subsets. Immun. Ageing 2022, 19, 11. [Google Scholar] [CrossRef]
- Kent, A.; Crump, L.S.; Davila, E. Beyond αβ T cells: NK, iNKT, and γδT cell biology in leukemic patients and potential for off-the-shelf adoptive cell therapies for AML. Front. Immunol. 2023, 14, 1202950. [Google Scholar] [CrossRef] [PubMed]
- Franciszkiewicz, K.; Salou, M.; Legoux, F.; Zhou, Q.; Cui, Y.; Bessoles, S.; Lantz, O. MHC class I-related molecule, MR1, and mucosal-associated invariant T cells. Immunol. Rev. 2016, 272, 120–138. [Google Scholar] [CrossRef]
- Ussher, J.E.; Bilton, M.; Attwod, E.; Shadwell, J.; Richardson, R.; de Lara, C.; Mettke, E.; Kurioka, A.; Hansen, T.H.; Klenerman, P.; et al. CD161++ CD8+ T cells, including the MAIT cell subset, are specifically activated by IL-12+IL-18 in a TCR-independent manner. Eur. J. Immunol. 2014, 44, 195–203. [Google Scholar] [CrossRef]
- Le Bourhis, L.; Guerri, L.; Dusseaux, M.; Martin, E.; Soudais, C.; Lantz, O. Mucosal-associated invariant T cells: Unconventional development and function. Trends Immunol. 2011, 32, 212–218. [Google Scholar] [CrossRef]
- Leeansyah, E.; Boulouis, C.; Kwa, A.L.H.; Sandberg, J.K. Emerging Role for MAIT Cells in Control of Antimicrobial Resistance. Trends Microbiol. 2021, 29, 504–516. [Google Scholar] [CrossRef]
- Meierovics, A.; Yankelevich, W.-J.C.; Cowley, S.C. MAIT cells are critical for optimal mucosal immune responses during in vivo pulmonary bacterial infection. Proc. Natl. Acad. Sci. USA 2013, 110, E3119–E3128. [Google Scholar] [CrossRef]
- Rudak, P.T.; Choi, J.; Haeryfar, S.M.M. MAIT cell-mediated cytotoxicity: Roles in host defense and therapeutic potentials in infectious diseases and cancer. J. Leukoc. Biol. 2018, 104, 473–486. [Google Scholar] [CrossRef] [PubMed]
- Sharma, M.; Zhang, S.; Niu, L.; Lewinsohn, D.M.; Zhang, X.; Huang, S. Mucosal-Associated Invariant T Cells Develop an Innate-Like Transcriptomic Program in Anti-mycobacterial Responses. Front. Immunol. 2020, 11, 1136. [Google Scholar] [CrossRef] [PubMed]
- Leng, T.; Akther, H.D.; Hackstein, C.-P.; Powell, K.; King, T.; Friedrich, M.; Christoforidou, Z.; McCuaig, S.; Neyazi, M.; Arancibia-Cárcamo, C.V.; et al. TCR and Inflammatory Signals Tune Human MAIT Cells to Exert Specific Tissue Repair and Effector Functions. Cell Rep. 2019, 28, 3077–3091.e5. [Google Scholar] [CrossRef]
- Constantinides, M.G.; Link, V.M.; Tamoutounour, S.; Wong, A.C.; Perez-Chaparro, P.J.; Han, S.-J.; Chen, E.Y.; Li, K.; Farhat, S.; Weckel, A.; et al. MAIT cells are imprinted by the microbiota in early life and promote tissue repair. Science 2019, 366, eaax6624. [Google Scholar] [CrossRef]
- Hinks, T.S.C.; Marchi, E.; Jabeen, M.; Olshansky, M.; Kurioka, A.; Pediongco, T.J.; Meehan, B.S.; Kostenko, L.; Turner, S.J.; Corbett, A.J.; et al. Activation and In Vivo Evolution of the MAIT Cell Transcriptome in Mice and Humans Reveals Tissue Repair Functionality. Cell Rep. 2019, 28, 3249–3262.e5. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.I.; Ninkov, M.; Haeryfar, S.M.M. Classic costimulatory interactions in MAIT cell responses: From gene expression to immune regulation. Clin. Exp. Immunol. 2023, 213, 50–66. [Google Scholar] [CrossRef] [PubMed]
- Su, B.; Kong, D.; Yang, X.; Zhang, T.; Kuang, Y.-Q. Mucosal-associated invariant T cells: A cryptic coordinator in HIV-infected immune reconstitution. J. Med. Virol. 2022, 94, 3043–3053. [Google Scholar] [CrossRef]
- Emgård, J.; Bergsten, H.; McCormick, J.K.; Barrantes, I.; Skrede, S.; Sandberg, J.K.; Norrby-Teglund, A. MAIT Cells Are Major Contributors to the Cytokine Response in Group A Streptococcal Toxic Shock Syndrome. Proc. Natl. Acad. Sci. USA 2019, 116, 25923–25931. [Google Scholar] [CrossRef]
- Hegde, P.; Weiss, E.; Paradis, V.; Wan, J.; Mabire, M.; Sukriti, S.; Rautou, P.-E.; Albuquerque, M.; Picq, O.; Chandra Gupta, A.; et al. Mucosal-associated invariant T cells are a profibrogenic immune cell population in the liver. Nat. Commun. 2018, 9, 2146. [Google Scholar] [CrossRef]
- Yan, C.; Luo, Z.; Li, W.; Li, X.; Dallmann, R.; Kurihara, H.; Li, Y.-F.; He, R.-R. Disturbed Yin–Yang balance: Stress increases the susceptibility to primary and recurrent infections of herpes simplex virus type 1. Acta Pharm. Sin. B 2020, 10, 383–398. [Google Scholar] [CrossRef]
- Petley, E.V.; Koay, H.-F.; Henderson, M.A.; Sek, K.; Todd, K.L.; Keam, S.P.; Lai, J.; House, I.G.; Li, J.; Zethoven, M.; et al. MAIT cells regulate NK cell-mediated tumor immunity. Nat. Commun. 2021, 12, 4746. [Google Scholar] [CrossRef]
- Ussher, J.E.; Willberg, C.B.; Klenerman, P. MAIT cells and viruses. Immunol. Cell Biol. 2018, 96, 630–641. [Google Scholar] [CrossRef]
- Rouxel, O.; Lehuen, A. Mucosal-associated invariant T cells in autoimmune and immune-mediated diseases. Immunol. Cell Biol. 2018, 96, 618–629. [Google Scholar] [CrossRef] [PubMed]
- Chien, Y.; Meyer, C.; Bonneville, M. γδ T cells: First line of defense and beyond. Annu. Rev. Immunol. 2014, 32, 121–155. [Google Scholar] [CrossRef] [PubMed]
- Wilcox, D.R.; Muller, W.J.; Longnecker, R. HSV targeting of the host phosphatase PP1α is required for disseminated disease in the neonate and contributes to pathogenesis in the brain. Proc. Natl. Acad. Sci. USA 2015, 112, E6937–E6944. [Google Scholar] [CrossRef] [PubMed]
- Melandri, D.; Zlatareva, I.; Chaleil, R.A.G.; Dart, R.J.; Chancellor, A.; Nussbaumer, O.; Polyakova, O.; Roberts, N.A.; Wesch, D.; Kabelitz, D.; et al. The γδTCR combines innate immunity with adaptive immunity by utilizing spatially distinct regions for agonist selection and antigen responsiveness. Nat. Immunol. 2018, 19, 1352–1365. [Google Scholar] [CrossRef] [PubMed]
- Park, J.H.; Lee, H.K. Function of γδ T cells in tumor immunology and their application to cancer therapy. Exp. Mol. Med. 2021, 53, 318–327. [Google Scholar] [CrossRef]
- De Libero, G.; Lau, S.-Y.; Mori, L. Phosphoantigen Presentation to TCR γδ Cells, a Conundrum Getting Less Gray Zones. Front. Immunol. 2014, 5, 679. [Google Scholar] [CrossRef] [PubMed]
- Uldrich, A.P.; Le Nours, J.; Pellicci, D.G.; Gherardin, N.A.; McPherson, K.G.; Lim, R.T.; Patel, O.; Beddoe, T.; Gras, S.; Rossjohn, J.; et al. CD1d-lipid antigen recognition by the γδ TCR. Nat. Immunol. 2013, 14, 1137–1145. [Google Scholar] [CrossRef]
- Coffelt, S.B.; Kersten, K.; Doornebal, C.W.; Weiden, J.; Vrijland, K.; Hau, C.-S.; Verstegen, N.J.M.; Ciampricotti, M.; Hawinkels, L.J.A.C.; Jonkers, J.; et al. IL-17-producing γδ T cells and neutrophils conspire to promote breast cancer metastasis. Nature 2015, 522, 345–348. [Google Scholar] [CrossRef]
- Kuen, D.-S.; Kim, B.-S.; Chung, Y. IL-17-Producing Cells in Tumor Immunity: Friends or Foes? Immune Netw. 2020, 20, e6. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Dong, P.; He, W.; Zhang, J.; Chen, H. γδ T cells: Major advances in basic and clinical research in tumor immunotherapy. Chin. Med. J. 2023, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Baena, A.; Gómez-Giraldo, L.; Carreño, L.J. Mecanismos de activación de las células T asesinas naturales invariantes (iNKT). Iatreia 2016, 29, 51–64. [Google Scholar] [CrossRef]
- Tan, A.H.-M.; Chong, W.P.-K.; Ng, S.-W.; Basri, N.; Xu, S.; Lam, K.-P. Aberrant presentation of self-lipids by autoimmune B cells depletes peripheral iNKT cells. Cell Rep. 2014, 9, 24–31. [Google Scholar] [CrossRef]
- Brennan, P.J.; Brigl, M.; Brenner, M.B. Invariant natural killer T cells: An innate activation scheme linked to diverse effector functions. Nat. Rev. Immunol. 2013, 13, 101–117. [Google Scholar] [CrossRef]
- Krovi, S.H.; Gapin, L. Invariant Natural Killer T Cell Subsets—More Than Just Developmental Intermediates. Front. Immunol. 2018, 9, 1393. [Google Scholar] [CrossRef]
- Crosby, C.M.; Kronenberg, M. Tissue-specific functions of invariant natural killer T cells. Nat. Rev. Immunol. 2018, 18, 559–574. [Google Scholar] [CrossRef] [PubMed]
- Kawano, T.; Cui, J.; Koezuka, Y.; Toura, I.; Kaneko, Y.; Motoki, K.; Ueno, H.; Nakagawa, R.; Sato, H.; Kondo, E.; et al. CD1d-Restricted and TCR-Mediated Activation of Vα14 NKT Cells by Glycosylceramides. Science 1997, 278, 1626–1629. [Google Scholar] [CrossRef]
- Patel, O.; Pellicci, D.G.; Uldrich, A.P.; Sullivan, L.C.; Bhati, M.; McKnight, M.; Richardson, S.K.; Howell, A.R.; Mallevaey, T.; Zhang, J.; et al. Vβ2 natural killer T cell antigen receptor-mediated recognition of CD1d-glycolipid antigen. Proc. Natl. Acad. Sci. USA 2011, 108, 19007–19012. [Google Scholar] [CrossRef]
- Macho-Fernandez, E.; Brigl, M. The Extended Family of CD1d-Restricted NKT Cells: Sifting through a Mixed Bag of TCRs, Antigens, and Functions. Front. Immunol. 2015, 6, 362. [Google Scholar] [CrossRef]
- Pellicci, D.G.; Uldrich, A.P. Unappreciated diversity within the pool of CD1d-restricted T cells. Semin. Cell Dev. Biol. 2018, 84, 42–47. [Google Scholar] [CrossRef]
- Montalvillo, E.; Garrote, J.A.; Bernardo, D.; Arranz, E. Innate lymphoid cells and natural killer T cells in the gastrointestinal tract immune system. Rev. Esp. Enferm. Dig. 2014, 106, 334–345. [Google Scholar] [PubMed]
- Calabi, F.; Jarvis, J.M.; Martin, L.; Milstein, C. Two classes of CD1 genes. Eur. J. Immunol. 1989, 19, 285–292. [Google Scholar] [CrossRef] [PubMed]
- Kaczmarek, R.; Pasciak, M.; Szymczak-Kulus, K.; Czerwinski, M. CD1: A Singed Cat of the Three Antigen Presentation Systems. Arch. Immunol. Ther. Exp. 2017, 65, 201–214. [Google Scholar] [CrossRef] [PubMed]
- Chaudhry, M.S.; Karadimitris, A. Role and regulation of CD1d in normal and pathological B cells. J. Immunol. 2014, 193, 4761–4768. [Google Scholar] [CrossRef] [PubMed]
- Lee, G.K.; Park, H.J.; Macleod, M.; Chandler, P.; Munn, D.H.; Mellor, A.L. Tryptophan deprivation sensitizes activated T cells to apoptosis prior to cell division. Immunology 2002, 107, 452–460. [Google Scholar] [CrossRef]
- Calabi, F.; Belt, K.T.; Yu, C.Y.; Bradbury, A.; Mandy, W.J.; Milstein, C. The rabbit CD1 and the evolutionary conservation of the CD1 gene family. Immunogenetics 1989, 30, 370–377. [Google Scholar] [CrossRef]
- Dougan, S.K.; Kaser, A.; Blumberg, R.S. CD1 expression on antigen-presenting cells. Curr. Top Microbiol. Immunol. 2007, 314, 113–141. [Google Scholar] [CrossRef]
- Barral, D.C.; Brenner, M.B. CD1 antigen presentation: How it works. Nat. Rev. Immunol. 2007, 7, 929–941. [Google Scholar] [CrossRef]
- Berzins, S.P.; Ritchie, D.S. Natural killer T cells: Drivers or passengers in preventing human disease? Nat. Rev. Immunol. 2014, 14, 640–646. [Google Scholar] [CrossRef]
- Godfrey, D.I.; Kronenberg, M. Going both ways: Immune regulation via CD1d-dependent NKT cells. J. Clin. Investig. 2004, 114, 1379–1388. [Google Scholar] [CrossRef]
- Fujii, S.; Shimizu, K.; Smith, C.; Bonifaz, L.; Steinman, R.M. Activation of natural killer T cells by alpha-galactosylceramide rapidly induces the full maturation of dendritic cells in vivo and thereby acts as an adjuvant for combined CD4 and CD8 T cell immunity to a coadministered protein. J. Exp. Med. 2003, 198, 267–279. [Google Scholar] [CrossRef]
- Chennamadhavuni, D.; Saavedra-Avila, N.A.; Carreño, L.J.; Guberman-Pfeffer, M.J.; Arora, P.; Yongqing, T.; Pryce, R.; Koay, H.-F.; Godfrey, D.I.; Keshipeddy, S.; et al. Dual Modifications of α-Galactosylceramide Synergize to Promote Activation of Human Invariant Natural Killer T Cells and Stimulate Anti-tumor Immunity. Cell Chem. Biol. 2018, 25, 571–584.e8. [Google Scholar] [CrossRef]
- Mak, T.W.; Jett, B.D. NK, γδ T and NKT Cells. In Primer to the Immune Response, 2nd ed.; Saunders, M.E., Mak, T.W., Jett, B.D., Eds.; Academic Cell: Burlington, ON, Canada, 2014; pp. 247–268. [Google Scholar]
- Bandyopadhyay, K.; Marrero, I.; Kumar, V. NKT cell subsets as key participants in liver physiology and pathology. Cell Mol. Immunol. 2016, 13, 337–346. [Google Scholar] [CrossRef]
- Werner, J.M.; Lang, C.; Scherer, M.N.; Farkas, S.A.; Geissler, E.K.; Schlitt, H.J.; Hornung, M. Distribution of intrahepatic T, NK and CD3+CD56+NKT cells alters after liver transplantation: Shift from innate to adaptive immunity? Transpl. Immunol. 2011, 25, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Guarín, C.J.M.; López, M.T.R. Las Células iNKT Invariantes: Una Vía Promisoria para la Terapia de la Infección por el VIH. Salud UIS 2003, 35, 80–89. [Google Scholar]
- Sepúlveda, C.; Puente, J. Natural killer cells and the innate immune system in infectious pathology. Rev. Med. Chil. 2000, 128, 1361–1370. [Google Scholar] [CrossRef]
- Tupin, E.; Kinjo, Y.; Kronenberg, M. The unique role of natural killer T cells in the response to microorganisms. Nat. Rev. Microbiol. 2007, 5, 405–417. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Paul, W.E. Cultured NK1.1+ CD4+ T cells produce large amounts of IL-4 and IFN-gamma upon activation by anti-CD3 or CD1. J. Immunol. 1997, 159, 2240–2249. [Google Scholar] [CrossRef]
- Coquet, J.M.; Chakravarti, S.; Kyparissoudis, K.; McNab, F.W.; Pitt, L.A.; McKenzie, B.S.; Berzins, S.P.; Smyth, M.J.; Godfrey, D.I. Diverse cytokine production by NKT cell subsets and identification of an IL-17-producing CD4-NK1.1- NKT cell population. Proc. Natl. Acad. Sci. USA 2008, 105, 11287–11292. [Google Scholar] [CrossRef] [PubMed]
- Sag, D.; Krause, P.; Hedrick, C.C.; Kronenberg, M.; Wingender, G. IL-10-producing NKT10 cells are a distinct regulatory invariant NKT cell subset. J. Clin. Investig. 2014, 124, 3725–3740. [Google Scholar] [CrossRef] [PubMed]
- Grasseau, A.; Boudigou, M.; Le Pottier, L.; Chriti, N.; Cornec, D.; Pers, J.-O.; Renaudineau, Y.; Hillion, S. Innate B Cells: The Archetype of Protective Immune Cells. Clin. Rev. Allerg. Immunol. 2020, 58, 92–106. [Google Scholar] [CrossRef] [PubMed]
- Chang, P.-P.; Barral, P.; Fitch, J.; Pratama, A.; Ma, C.S.; Kallies, A.; Hogan, J.J.; Cerundolo, V.; Tangye, S.G.; Bittman, R.; et al. Identification of Bcl-6-dependent follicular helper NKT cells that provide cognate help for B cell responses. Nat. Immunol. 2012, 13, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Bai, L.; Deng, S.; Reboulet, R.; Mathew, R.; Teyton, L.; Savage, P.B.; Bendelac, A. Natural killer T (NKT)-B-cell interactions promote prolonged antibody responses and long-term memory to pneumococcal capsular polysaccharides. Proc. Natl. Acad. Sci. USA 2013, 110, 16097–16102. [Google Scholar] [CrossRef] [PubMed]
- Zimmer, M.I.; Nguyen, H.P.; Wang, B.; Xu, H.; Colmone, A.; Felio, K.; Choi, H.-J.; Zhou, P.; Alegre, M.L.; Wang, C.-R. Polymorphisms in CD1d affect antigen presentation and the activation of CD1d-restricted T cells. Proc. Natl. Acad. Sci. USA 2009, 106, 1909–1914. [Google Scholar] [CrossRef] [PubMed]
- Thomas, S.Y.; Scanlon, S.T.; Griewank, K.G.; Constantinides, M.G.; Savage, A.K.; Barr, K.A.; Meng, F.; Luster, A.D.; Bendelac, A. PLZF induces an intravascular surveillance program mediated by long-lived LFA-1–ICAM-1 interactions. J. Exp. Med. 2011, 208, 1179–1188. [Google Scholar] [CrossRef]
- Wingender, G.; Sag, D.; Kronenberg, M. NKT10 cells: A novel iNKT cell subset. Oncotarget 2015, 6, 26552–26553. [Google Scholar] [CrossRef]
- Liew, P.X.; Lee, W.-Y.; Kubes, P. iNKT Cells Orchestrate a Switch from Inflammation to Resolution of Sterile Liver Injury. Immunity 2017, 47, 752–765.e5. [Google Scholar] [CrossRef]
- Webster, K.E.; Kim, H.-O.; Kyparissoudis, K.; Corpuz, T.M.; Pinget, G.V.; Uldrich, A.P.; Brink, R.; Belz, G.T.; Cho, J.-H.; Godfrey, D.I.; et al. IL-17-producing NKT cells depend exclusively on IL-7 for homeostasis and survival. Mucosal Immunol. 2014, 7, 1058–1067. [Google Scholar] [CrossRef]
- Pillai, A.B.; George, T.I.; Dutt, S.; Teo, P.; Strober, S. Host NKT cells can prevent graft-versus-host disease and permit graft antitumor activity after bone marrow transplantation. J. Immunol. 2007, 178, 6242–6251. [Google Scholar] [CrossRef] [PubMed]
- Liman, N.; Park, J.-H. Markers and makers of NKT17 cells. Exp. Mol. Med. 2023, 55, 1090–1098. [Google Scholar] [CrossRef] [PubMed]
- Motomura, Y.; Kitamura, H.; Hijikata, A.; Matsunaga, Y.; Matsumoto, K.; Inoue, H.; Atarashi, K.; Hori, S.; Watarai, H.; Zhu, J.; et al. The transcription factor E4BP4 regulates the production of IL-10 and IL-13 in CD4+ T cells. Nat. Immunol. 2011, 12, 450–459. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, M.; Almeida, C.F.; Caridade, M.; Ribot, J.C.; Duarte, J.; Agua-Doce, A.; Wollenberg, I.; Silva-Santos, B.; Graca, L. Identification of Regulatory Foxp3+ Invariant NKT Cells Induced by TGF-β. J. Immunol. 2010, 185, 2157–2163. [Google Scholar] [CrossRef] [PubMed]
- LaMarche, N.M.; Kane, H.; Kohlgruber, A.C.; Dong, H.; Lynch, L.; Brenner, M.B. Distinct iNKT cell populations use IFNγ or ER stress-induced IL-10 to control adipose tissue homeostasis. Cell Metab. 2020, 32, 243–258.e6. [Google Scholar] [CrossRef] [PubMed]
- Burrello, C.; Strati, F.; Lattanzi, G.; Diaz-Basabe, A.; Mileti, E.; Giuffrè, M.R.; Lopez, G.; Cribiù, F.M.; Trombetta, E.; Kallikourdis, M.; et al. IL10 Secretion Endows Intestinal Human iNKT Cells with Regulatory Functions Towards Pathogenic T Lymphocytes. J. Crohns. Colitis. 2022, 16, 1461–1474. [Google Scholar] [CrossRef] [PubMed]
- Cameron, G.; Godfrey, D.I. Differential surface phenotype and context-dependent reactivity of functionally diverse NKT cells. Immunol. Cell Biol. 2018, 96, 759–771. [Google Scholar] [CrossRef]
- Saavedra-Avila, N.A.; Keshipeddy, S.; Guberman-Pfeffer, M.J.; Pérez-Gallegos, A.; Saini, N.K.; Schäfer, C.; Carreño, L.J.; Gascón, J.A.; Porcelli, S.A.; Howell, A.R. Amide-Linked C4″-Saccharide Modification of KRN7000 Provides Potent Stimulation of Human Invariant NKT Cells and Anti-Tumor Immunity in a Humanized Mouse Model. ACS Chem. Biol. 2020, 15, 3176–3186. [Google Scholar] [CrossRef]
- Lee, Y.J.; Holzapfel, K.L.; Zhu, J.; Jameson, S.C.; Hogquist, K.A. Steady state production of IL-4 modulates immunity in different strains and is determined by lineage diversity of iNKT cells. Nat. Immunol. 2013, 14, 1146–1154. [Google Scholar] [CrossRef]
- Natori, T.; Koezuka, Y.; Higa, T. Agelasphins, novel α-galactosylceramides from the marine sponge Agelas mauritianus. Tetrahedron Lett. 1993, 34, 5591–5592. [Google Scholar] [CrossRef]
- Watson, F.N.; Shears, M.J.; Matsubara, J.; Kalata, A.C.; Seilie, A.M.; Cruz Talavera, I.; Olsen, T.M.; Tsuji, M.; Chakravarty, S.; Kim Lee Sim, B.; et al. Cryopreserved Sporozoites with and without the Glycolipid Adjuvant 7DW8-5 Protect in Prime-and-Trap Malaria Vaccination. Am. J. Trop. Med. Hyg. 2022, 106, 1227–1236. [Google Scholar] [CrossRef] [PubMed]
- Pifferi, C.; Fuentes, R.; Fernández-Tejada, A. Natural and synthetic carbohydrate-based vaccine adjuvants and their mechanisms of action. Nat. Rev. Chem. 2021, 5, 197–216. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Bhave, M.S.; Yagita, H.; Cardell, S.L. Natural Killer T-Cell Agonist α-Galactosylceramide and PD-1 Blockade Synergize to Reduce Tumor Development in a Preclinical Model of Colon Cancer. Front. Immunol. 2020, 11, 581301. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.A.; Kang, M.H.; Lee, Y.S.; Kim, Y.J.; Kim, D.H.; Ko, H.J.; Kang, C.-Y. A distinct subset of natural killer T cells produces IL-17, contributing to airway infiltration of neutrophils but not to airway hyperreactivity. Cell Immunol. 2008, 251, 50–55. [Google Scholar] [CrossRef]
- Eberl, G.; MacDonald, H.R. Selective induction of NK cell proliferation and cytotoxicity by activated NKT cells. Eur. J. Immunol. 2000, 30, 985–992. [Google Scholar] [CrossRef]
- Nieuwenhuis, E.E.; Matsumoto, T.; Exley, M.; Schleipman, R.A.; Glickman, J.; Bailey, D.T.; Corazza, N.; Colgan, S.P.; Onderdonk, A.B.; Blumberg, R.S. CD1d-dependent macrophage-mediated clearance of Pseudomonas aeruginosa from lung. Nat. Med. 2002, 8, 588–593. [Google Scholar] [CrossRef]
- Gorbachev, A.V.; Fairchild, R.L. Activated NKT cells increase dendritic cell migration and enhance CD8+ T cell responses in the skin. Eur. J. Immunol. 2006, 36, 2494–2503. [Google Scholar] [CrossRef]
- Eberl, G.; Brawand, P.; MacDonald, H.R. Selective bystander proliferation of memory CD4+ and CD8+ T cells upon NK T or T cell activation. J. Immunol. 2000, 165, 4305–4311. [Google Scholar] [CrossRef]
- Leadbetter, E.A.; Brigl, M.; Illarionov, P.; Cohen, N.; Luteran, M.C.; Pillai, S.; Besra, G.S.; Brenner, M.B. NK T cells provide lipid antigen-specific cognate help for B cells. Proc. Natl. Acad. Sci. USA 2008, 105, 8339–8344. [Google Scholar] [CrossRef]
- Cerundolo, V.; Silk, J.D.; Masri, S.H.; Salio, M. Harnessing invariant NKT cells in vaccination strategies. Nat. Rev. Immunol. 2009, 9, 28–38. [Google Scholar] [CrossRef]
- Nishimura, T.; Kitamura, H.; Iwakabe, K.; Yahata, T.; Ohta, A.; Sato, M.; Takeda, K.; Okumura, K.; Van Kaer, L.; Kawano, T.; et al. The interface between innate and acquired immunity: Glycolipid antigen presentation by CD1d-expressing dendritic cells to NKT cells induces the differentiation of antigen-specific cytotoxic T lymphocytes. Int. Immunol. 2000, 12, 987–994. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.; Hong, S.; Scherer, D.C.; Serizawa, I.; Burdin, N.; Kronenberg, M.; Koezuka, Y.; Van Ker, L. Cutting edge: Activation of NK T cells by CD1d and alpha-galactosylceramide directs conventional T cells to the acquisition of a Th2 phenotype. J. Immunol. 1999, 163, 2373–2377. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.Q.; Yan, C.; Luo, R.; Liu, Z. iNKT cell agonists as vaccine adjuvants to combat infectious diseases. Carbohydr. Res. 2022, 513, 108527. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Aseguinolaza, G.; Van Kaer, L.; Bergmann, C.C.; Wilson, J.M.; Schmieg, J.; Kronenberg, M.; Nakayama, T.; Taniguchi, M.; Koezuka, T.; Tsuji, M. Natural killer T cell ligand alpha-galactosylceramide enhances protective immunity induced by malaria vaccines. J. Exp. Med. 2002, 195, 617–624. [Google Scholar] [CrossRef] [PubMed]
- Dondji, B.; Deak, E.; Goldsmith-Pestana, K.; Perez-Jimenez, E.; Esteban, M.; Miyake, S.; Yamamura, T.; McMahon-Pratt, D. Intradermal NKT cell activation during DNA priming in heterologous prime-boost vaccination enhances T cell responses and protection against Leishmania. Eur. J. Immunol. 2008, 38, 706–719. [Google Scholar] [CrossRef] [PubMed]
- Romagnani, S. The Th1/Th2 paradigm. Immunol. Today 1997, 18, 263–266. [Google Scholar] [CrossRef] [PubMed]
- Wilson, S.B.; Delovitch, T.L. Janus-like role of regulatory iNKT cells in autoimmune disease and tumour immunity. Nat. Rev. Immunol. 2003, 3, 211–222. [Google Scholar] [CrossRef]
- Parekh, V.V.; Wilson, M.T.; Olivares-Villagomez, D.; Singh, A.K.; Wu, L.; Wang, C.R.; Joyce, S.; Van Ker, L. Glycolipid antigen induces long-term natural killer T cell anergy in mice. J. Clin. Investig. 2005, 115, 2572–2583. [Google Scholar] [CrossRef]
- Osman, Y.; Kawamura, T.; Naito, T.; Takeda, K.; Van Kaer, L.; Okumura, K.; Abo, T. Activation of hepatic NKT cells and subsequent liver injury following administration of alpha-galactosylceramide. Eur. J. Immunol. 2000, 30, 1919–1928. [Google Scholar] [CrossRef]
- Laurent, X.; Bertin, B.; Renault, N.; Farce, A.; Speca, S.; Milhomme, O.; Mille, R.; Desreumau, P.; Hénon, E.; Chavatte, P. Switching invariant natural killer T (iNKT) cell response from anticancerous to anti-inflammatory effect: Molecular bases. J. Med. Chem. 2014, 57, 5489–5508. [Google Scholar] [CrossRef]
- Li, X.; Fujio, M.; Imamura, M.; Wu, D.; Vasan, S.; Wong, C.H.; Ho, D.D.; Tsuji, M. Design of a potent CD1d-binding NKT cell ligand as a vaccine adjuvant. Proc. Natl. Acad. Sci. USA 2010, 107, 13010–13015. [Google Scholar] [CrossRef]
- Venkataswamy, M.M.; Ng, T.W.; Kharkwal, S.S.; Carreño, L.J.; Johnson, A.J.; Kunnath-Velayudhan, S.; Liu, Z.; Bittman, R.; Jervis, P.J.; Cox, L.R.; et al. Improving Mycobacterium bovis bacillus Calmette-Guèrin as a vaccine delivery vector for viral antigens by incorporation of glycolipid activators of NKT cells. PLoS ONE 2014, 9, e108383. [Google Scholar] [CrossRef]
- Xu, X.; Hegazy, W.A.; Guo, L.; Gao, X.; Courtney, A.N.; Kurbanov, S.; Liu, D.; Tian, G.; Manuel, E.R.; Diamond, D.J.; et al. Effective cancer vaccine platform based on attenuated salmonella and a type III secretion system. Cancer Res. 2014, 74, 6260–6270. [Google Scholar] [CrossRef]
- Tsuji, M.; Nair, M.S.; Masuda, K.; Castagna, C.; Chong, Z.; Darling, T.L.; Seehra, K.; Hwang, Y.; Ribeiro, Á.L.; Ferreira, G.M.; et al. An immunostimulatory glycolipid that blocks SARS-CoV-2, RSV, and influenza infections in vivo. Nat. Commun. 2023, 14, 3959. [Google Scholar] [CrossRef]
- Velmourougane, G.; Raju, R.; Bricard, G.; Im, J.S.; Besra, G.S.; Porcelli, S.A.; Howell, A.R. Synthesis and evaluation of an acyl-chain unsaturated analog of the Th2 biasing, immunostimulatory glycolipid, OCH. Bioorg. Med. Chem. Lett. 2009, 19, 3386–3388. [Google Scholar] [CrossRef]
- Wei, J.; Hegde, V.L.; Yanamandra, A.V.; O’Hara, M.P.; Keegan, B.; Jones, K.M.; Strych, U.; Bottazzi, M.E.; Zhan, B.; Jagannadha Sastry, K.; et al. Mucosal Vaccination With Recombinant Tm-WAP49 Protein Induces Protective Humoral and Cellular Immunity Against Experimental Trichuriasis in AKR Mice. Front. Immunol. 2022, 13, 800295. [Google Scholar] [CrossRef]
- Jeong, D.; Woo, Y.D.; Chung, D.H. Invariant natural killer T cells in lung diseases. Exp. Mol. Med. 2023, 55, 1885–1894. [Google Scholar] [CrossRef]
- Rothchild, A.C.; Jayaraman, P.; Nunes-Alves, C.; Behar, S.M. iNKT cell production of GM-CSF controls Mycobacterium tuberculosis. PLoS Pathog. 2014, 10, 1003805. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.; Singh, S.; Galvan, G.; Jagannath, C.; Sastry, K.J. Prophylactic Sublingual Immunization with Mycobacterium tuberculosis Subunit Vaccine Incorporating the Natural Killer T Cell Agonist Alpha-Galactosylceramide Enhances Protective Immunity to Limit Pulmonary and Extra-Pulmonary Bacterial Burden in Mice. Vaccines 2017, 5, 47. [Google Scholar] [CrossRef] [PubMed]
- Venkataswamy, M.M.; Baena, A.; Goldberg, M.F.; Bricard, G.; Im, J.S.; Chan, J.; Reddington, F.; Besra, G.S.; Jacobs, W.R., Jr.; Porcelli, S.A. Incorporation of NKT Cell-Activating Glycolipids Enhances Immunogenicity and Vaccine Efficacy of Mycobacterium bovis Bacillus Calmette-Guérin1. J. Immunol. 2009, 183, 1644–1656. [Google Scholar] [CrossRef] [PubMed]
- Noda, K.; Kodama, S.; Umemoto, S.; Abe, N.; Hirano, T.; Suzuki, M. Nasal vaccination with P6 outer membrane protein and alpha-galactosylceramide induces nontypeable Haemophilus influenzae-specific protective immunity associated with NKT cell activation and dendritic cell expansion in nasopharynx. Vaccine 2010, 28, 5068–5074. [Google Scholar] [CrossRef] [PubMed]
- Noda, K.; Kodama, S.; Umemoto, S.; Nomi, N.; Hirano, T.; Suzuki, M. Th17 cells contribute to nontypeable Haemophilus influenzae-specific protective immunity induced by nasal vaccination with P6 outer membrane protein and alpha-galactosylceramide. Microbiol. Immunol. 2011, 55, 574–581. [Google Scholar] [CrossRef] [PubMed]
- Kakimi, K.; Guidotti, L.G.; Koezuka, Y.; Chisari, F.V. Natural killer T cell activation inhibits hepatitis B virus replication in vivo. J. Exp. Med. 2000, 192, 921–930. [Google Scholar] [CrossRef] [PubMed]
- Driver, J.P.; Carvalho Madrid, D.M.; Gu, W.; Artiaga, B.L.; Richt, J.A. Modulation of Immune Responses to Influenza A Virus Vaccines by Natural Killer T Cells. Front. Immunol. 2020, 11, 2172. [Google Scholar] [CrossRef] [PubMed]
- Guillonneau, C.; Mintern, J.D.; Hubert, F.-X.; Hurt, A.C.; Besra, G.S.; Porcelli, S.; Barr, I.G.; Doherty, P.C.; Godfrey, D.I.; Turner, S.J. Combined NKT cell activation and influenza virus vaccination boosts memory CTL generation and protective immunity. Proc. Natl. Acad. Sci. USA 2009, 106, 3330–3335. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Chen, A.; Li, X.; Chen, Z.; Zhang, W.; Song, Y.; Gurner, D.; Gardiner, D.; Basu, S.; Ho, D.D.; et al. Enhancement of HIV DNA vaccine immunogenicity by the NKT cell ligand, alpha-galactosylceramide. Vaccine 2008, 26, 1807–1816. [Google Scholar] [CrossRef]
- Singh, S.; Yang, G.; Byrareddy, S.N.; Barry, M.A.; Sastry, K.J. Natural killer T cell and TLR9 agonists as mucosal adjuvants for sublingual vaccination with clade C HIV-1 envelope protein. Vaccine 2014, 32, 6934–6940. [Google Scholar] [CrossRef]
- Singh, S.; Nehete, P.N.; Yang, G.; He, H.; Nehete, B.; Hanley, P.W.; Barry, M.A.; Sastry, K.J. Enhancement of Mucosal Immunogenicity of Viral Vectored Vaccines by the NKT Cell Agonist Alpha-Galactosylceramide as Adjuvant. Vaccines 2014, 2, 686–706. [Google Scholar] [CrossRef]
- Lu, H.; Liu, Z.; Deng, X.; Chen, S.; Zhou, R.; Zhao, R.; Parandaman, R.; Thind, A.; Henley, J.; Tian, L.; et al. Potent NKT cell ligands overcome SARS-CoV-2 immune evasion to mitigate viral pathogenesis in mouse models. PLoS Pathog. 2023, 19, 1011240. [Google Scholar] [CrossRef]
- Wang, J.; Wen, Y.; Zhou, S.H.; Zhang, H.W.; Peng, X.Q.; Zhang, R.Y.; Yin, X.-G.; Qiu, H.; Gong, R.; Yang, G.-F.; et al. Self-Adjuvanting Lipoprotein Conjugate alphaGalCer-RBD Induces Potent Immunity against SARS-CoV-2 and its Variants of Concern. J. Med. Chem. 2022, 65, 2558–2570. [Google Scholar] [CrossRef]
- Montoya, C.J.; Catano, J.C.; Ramirez, Z.; Rugeles, M.T.; Wilson, S.B.; Landay, A.L. Invariant NKT cells from HIV-1 or Mycobacterium tuberculosis-infected patients express an activated phenotype. Clin. Immunol. 2008, 127, 1–6. [Google Scholar] [CrossRef]
- Almeida, J.-S.; Casanova, J.M.; Santos-Rosa, M.; Tarazona, R.; Solana, R.; Rodrigues-Santos, P. Natural Killer T-like Cells: Immunobiology and Role in Disease. Int. J. Mol. Sci. 2023, 24, 2743. [Google Scholar] [CrossRef]
- Cho, S.; Knox, K.S.; Kohli, L.M.; He, J.J.; Exley, M.A.; Wilson, S.B.; Brutkiewicz, R.R. Impaired cell surface expression of human CD1d by the formation of an HIV-1 Nef/CD1d complex. Virology 2005, 337, 242–252. [Google Scholar] [CrossRef]
- Churiso, G.; Husen, G.; Bulbula, D.; Abebe, L. Immunity Cell Responses to RSV and the Role of Antiviral Inhibitors: A Systematic Review. Infect. Drug Resist. 2022, 15, 7413–7430. [Google Scholar] [CrossRef] [PubMed]
- Galvez, N.M.S.; Bohmwald, K.; Pacheco, G.A.; Andrade, C.A.; Carreno, L.J.; Kalergis, A.M. Type I Natural Killer T Cells as Key Regulators of the Immune Response to Infectious Diseases. Clin. Microbiol. Rev. 2021, 34. [Google Scholar] [CrossRef]
- Rey-Jurado, E.; Bohmwald, K.; Galvez, N.M.S.; Becerra, D.; Porcelli, S.A.; Carreno, L.J.; Kalergis, A.M. Contribution of NKT cells to the immune response and pathogenesis triggered by respiratory viruses. Virulence 2020, 11, 580–593. [Google Scholar] [CrossRef]
- Sanchez, D.J.; Gumperz, J.E.; Ganem, D. Regulation of CD1d expression and function by a herpesvirus infection. J. Clin. Investig. 2005, 115, 1369–1378. [Google Scholar] [CrossRef] [PubMed]
- Yuan, W.; Dasgupta, A.; Cresswell, P. Herpes simplex virus evades natural killer T cell recognition by suppressing CD1d recycling. Nat. Immunol. 2006, 7, 835–842. [Google Scholar] [CrossRef] [PubMed]
- Eberl, G.; Lees, R.; Smiley, S.T.; Taniguchi, M.; Grusby, M.J.; MacDonald, H.R. Tissue-specific segregation of CD1d-dependent and CD1d-independent NK T cells. J. Immunol. 1999, 162, 6410–6419. [Google Scholar] [CrossRef]
- Gao, B.; Jeong, W.I.; Tian, Z. Liver: An organ with predominant innate immunity. Hepatology 2008, 47, 729–736. [Google Scholar] [CrossRef]
- Shen, H.; Gu, C.; Liang, T.; Liu, H.; Guo, F.; Liu, X. Unveiling the heterogeneity of NKT cells in the liver through single cell RNA sequencing. Sci. Rep. 2020, 10, 19453. [Google Scholar] [CrossRef]
- Lei, Z.; Tang, R.; Qi, Q.; Gu, P.; Wang, J.; Xu, L.; Wei, C.; Pu, Y.; Qi, X.; Chen, Y.; et al. Hepatocyte CD1d protects against liver immunopathology in mice with schistosomiasis japonica. Immunology 2021, 162, 328–338. [Google Scholar] [CrossRef]
- Zeissig, S.; Peuker, K.; Iyer, S.; Gensollen, T.; Dougan, S.K.; Olszak, T.; Kaser, A.; Blumberg, R.S. CD1d-Restricted pathways in hepatocytes control local natural killer T cell homeostasis and hepatic inflammation. Proc. Natl. Acad. Sci. USA 2017, 114, 10449–10454. [Google Scholar] [CrossRef]
- Lee, Y.J.; Wang, H.; Starrett, G.J.; Phuong, V.; Jameson, S.C.; Hogquist, K.A. Tissue-Specific Distribution of iNKT Cells Impacts Their Cytokine Response. Immunity 2015, 43, 566–578. [Google Scholar] [CrossRef]
- Gu, X.; Chu, Q.; Ma, X.; Wang, J.; Chen, C.; Guan, J.; Ren, Y.; Wu, S.; Zhu, H. New insights into iNKT cells and their roles in liver diseases. Front. Immunol. 2022, 13, 1035950. [Google Scholar] [CrossRef]
- Shimaoka, T.; Seino, K.; Kume, N.; Minami, M.; Nishime, C.; Suematsu, M.; Kita, T.; Taniguchi, M.; Matsushima, K.; Yonehara, S. Critical role for CXC chemokine ligand 16 (SR-PSOX) in Th1 response mediated by NKT cells. J. Immunol. 2007, 179, 8172–8179. [Google Scholar] [CrossRef]
- Zhu, H.; Zhang, Q.; Chen, G. CXCR6 deficiency ameliorates ischemia-reperfusion injury by reducing the recruitment and cytokine production of hepatic NKT cells in a mouse model of non-alcoholic fatty liver disease. Int. Immunopharmacol. 2019, 72, 224–234. [Google Scholar] [CrossRef]
- Elbahrawy, A.; Atalla, H.; Alboraie, M.; Alwassief, A.; Madian, A.; El Fayoumie, M.; Tabll, A.A.; Aly, H.H. Recent Advances in Protective Vaccines against Hepatitis Viruses: A Narrative Review. Viruses 2023, 15, 214. [Google Scholar] [CrossRef]
- Torres, H.A.; Shigle, T.L.; Hammoudi, N.; Link, J.T.; Samaniego, F.; Kaseb, A.; Mallet, V. The oncologic burden of hepatitis C virus infection: A clinical perspective. CA Cancer J. Clin. 2017, 67, 411–431. [Google Scholar] [CrossRef]
- Min, Y.; Wei, X.; Xia, X.; Wei, Z.; Li, R.; Jin, J.; Liu, Z.; Hu, X.; Peng, X. Hepatitis B virus infection: An insight into the clinical connection and molecular interaction between hepatitis B virus and host extrahepatic cancer risk. Front. Immunol. 2023, 14, 1141956. [Google Scholar] [CrossRef]
- Zeissig, S.; Murata, K.; Sweet, L.; Publicover, J.; Hu, Z.; Kaser, A.; Bosse, E.; Iqbal, J.; Mahmood Hussain, M.; Balschun, K.; et al. Hepatitis B virus-induced lipid alterations contribute to natural killer T cell-dependent protective immunity. Nat. Med. 2012, 18, 1060–1068. [Google Scholar] [CrossRef] [PubMed]
- Baron, J.L.; Gardiner, L.; Nishimura, S.; Shinkai, K.; Locksley, R.; Ganem, D. Activation of a nonclassical NKT cell subset in a transgenic mouse model of hepatitis B virus infection. Immunity 2002, 16, 583–594. [Google Scholar] [CrossRef] [PubMed]
- Jin, Z.; Sun, R.; Wei, H.; Gao, X.; Chen, Y.; Tian, Z. Accelerated liver fibrosis in hepatitis B virus transgenic mice: Involvement of natural killer T cells. Hepatology 2011, 53, 219–229. [Google Scholar] [CrossRef]
- You, H.; Wang, X.; Ma, L.; Zhang, F.; Zhang, H.; Wang, Y.; Pan, X.; Zheng, K.; Kong, F.; Tang, R. Insights into the impact of hepatitis B virus on hepatic stellate cell activation. Cell Commun. Signal 2023, 21, 70. [Google Scholar] [CrossRef]
- Michel, M.L.; Keller, A.C.; Paget, C.; Fujio, M.; Trottein, F.; Savage, P.B.; Wong, C.-H.; Schneider, E.; Dy, M.; Leite-de-Moraes, M.C. Identification of an IL-17-producing NK1.1(neg) iNKT cell population involved in airway neutrophilia. J. Exp. Med. 2007, 204, 995–1001. [Google Scholar] [CrossRef] [PubMed]
- Tan, X.; Ding, Y.; Zhu, P.; Dou, R.; Liang, Z.; Yang, D.; Huang, Z.; Wang, W.; Wu, X.; Weng, X. Elevated Hepatic CD1d Levels Coincide with Invariant NKT Cell Defects in Chronic Hepatitis B Virus Infection. J. Immunol. 2018, 200, 3530–3538. [Google Scholar] [CrossRef] [PubMed]
- Deignan, T.; Curry, M.P.; Doherty, D.G.; Golden-Mason, L.; Volkov, Y.; Norris, S.; Nolan, N.; Traynor, O.; McEntee, G.; Hegarty, J.E.; et al. Decrease in hepatic CD56(+) T cells and V alpha 24(+) natural killer T cells in chronic hepatitis C viral infection. J. Hepatol. 2002, 37, 101–108. [Google Scholar] [CrossRef]
- Gonzalez, L.R.; Conesa, A.; Blanca, I.; Machado, I.; Fernandez, S.; Pujol, F.H.; Toro, F. Increased number of non-invariant NKT cells and low number of circulating CD1-expressing leukocytes in patients infected with hepatitis C virus. Ann. Biol. Clin. 2012, 70, 695–701. [Google Scholar] [CrossRef]
- Miyaki, E.; Hiraga, N.; Imamura, M.; Uchida, T.; Kan, H.; Tsuge, M.; Abe-Chayama, H.; Hayes, C.N.; Makokha, G.N.; Serikawa, M.; et al. Interferon alpha treatment stimulates interferon gamma expression in type I NKT cells and enhances their antiviral effect against hepatitis C virus. PLoS ONE 2017, 12, 0172412. [Google Scholar] [CrossRef]
- van Elsland, D.; Neefjes, J. Bacterial infections and cancer. EMBO Rep. 2018, 19, e46632. [Google Scholar] [CrossRef]
- Zembower, T.R. Epidemiology of Infections in Cancer Patients. In Infectious Complications in Cancer Patients; Stosor, V., Zembower, T.R., Eds.; Springer International Publishing: Cham, Switzerland, 2014; pp. 43–89. [Google Scholar] [CrossRef]
- Murata, M. Inflammation and cancer. Environ. Health Prev. Med. 2018, 23, 50. [Google Scholar] [CrossRef]
- El-Serag, H.B. Epidemiology of viral hepatitis and hepatocellular carcinoma. Gastroenterology 2012, 142, 1264–1273.e1. [Google Scholar] [CrossRef]
- Jindal, A.; Thadi, A.; Shailubhai, K. Hepatocellular Carcinoma: Etiology and Current and Future Drugs. J. Clin. Exp. Hepatol. 2019, 9, 221–232. [Google Scholar] [CrossRef]
- Cheng, X.; Tan, X.; Wang, W.; Zhang, Z.; Zhu, R.; Wu, M.; Li, M.; Chen, Y.; Liang, Z.; Zhu, P.; et al. Long-Chain Acylcarnitines Induce Senescence of Invariant Natural Killer T Cells in Hepatocellular Carcinoma. Cancer Res. 2023, 83, 582–594. [Google Scholar] [CrossRef]
- Fu, S.; He, K.; Tian, C.; Sun, H.; Zhu, C.; Bai, S.; Liu, J.; Wu, Q.; Xie, D.; Yue, T.; et al. Impaired lipid biosynthesis hinders anti-tumor efficacy of intratumoral iNKT cells. Nat. Commun. 2020, 11, 438. [Google Scholar] [CrossRef]
- Lee, I.-C.; Huang, Y.-H.; Chau, G.-Y.; Huo, T.-I.; Su, C.-W.; Wu, J.-C.; Lin, H.-C. Serum interferon gamma level predicts recurrence in hepatocellular carcinoma patients after curative treatments. Int. J. Cancer 2013, 133, 2895–2902. [Google Scholar] [CrossRef]
- Li, Y.-R.; Ochoa, C.J.; Zhu, Y.; Kramer, A.; Wilson, M.; Fang, Y.; Chen, Y.; Singh, T.; Di Bernardo, G.; Zhu, E.; et al. Profiling ovarian cancer tumor and microenvironment during disease progression for cell-based immunotherapy design. iScience 2023, 26, 107952. [Google Scholar] [CrossRef]
- Krijgsman, D.; Hokland, M.; Kuppen, P.J.K. The Role of Natural Killer T Cells in Cancer—A Phenotypical and Functional Approach. Front. Immunol. 2018, 9, 367. [Google Scholar] [CrossRef]
- Liu, X.; Li, L.; Si, F.; Huang, L.; Zhao, Y.; Zhang, C.; Hoft, D.F.; Peng, G. NK and NKT cells have distinct properties and functions in cancer. Oncogene 2021, 40, 4521–4537. [Google Scholar] [CrossRef]
- Stavrou, V.; Fultang, L.; Booth, S.; De Simone, D.; Bartnik, A.; Scarpa, U.; Gneo, L.; Panetti, S.; Potluri, S.; Almowaled, M.; et al. Invariant NKT cells metabolically adapt to the acute myeloid leukaemia environment. Cancer Immunol. Immunother. 2023, 72, 543–560. [Google Scholar] [CrossRef]
- Yi, Q.; Wang, J.; Liu, T.; Yao, Y.; Loveless, I.; Subedi, K.; Toor, J.; Adrianto, I.; Xiao, H.; Chen, B.; et al. scRNA-Seq and imaging mass cytometry analyses unveil iNKT cells-mediated anti-tumor immunity in pancreatic cancer liver metastasis. Cancer Lett. 2023, 561, 216149. [Google Scholar] [CrossRef]
- Ascui, G.; Gálvez-Jirón, F.; Kramm, K.; Schäfer, C.; Siña, J.; Pola, V.; Cristi, F.; Hernández, C.; Garrido-Tapia, M.; Pesce, B.; et al. Decreased invariant natural killer T-cell-mediated antitumor immune response in patients with gastric cancer. Immunol. Cell Biol. 2020, 98, 500–513. [Google Scholar] [CrossRef]
- Torre, E.; Pinton, G.; Lombardi, G.; Fallarini, S. Melanoma Cells Inhibit iNKT Cell Functions via PGE2 and IDO1. Cancers 2023, 15, 3498. [Google Scholar] [CrossRef]
- Singh, M.; Quispe-Tintaya, W.; Chandra, D.; Jahangir, A.; Venkataswamy, M.M.; Ng, T.W.; Sharma-Kharkwal, S.; Carreño, L.J.; Porcelli, S.A.; Gravekamp, C. Direct incorporation of the NKT-cell activator α-galactosylceramide into a recombinant Listeria monocytogenes improves breast cancer vaccine efficacy. Br. J. Cancer 2014, 111, 1945–1954. [Google Scholar] [CrossRef]
- Bae, E.-A.; Seo, H.; Kim, I.-K.; Jeon, I.; Kang, C.-Y. Roles of NKT cells in cancer immunotherapy. Arch. Pharm. Res. 2019, 42, 543–548. [Google Scholar] [CrossRef]
- Nelson, A.; Lukacs, J.D.; Johnston, B. The Current Landscape of NKT Cell Immunotherapy and the Hills Ahead. Cancers 2021, 13, 5174. [Google Scholar] [CrossRef]
- Yamashita, K.; Arimoto, A.; Nishi, M.; Tanaka, T.; Fujita, M.; Fukuoka, E.; Sugita, Y.; Nakagawa, A.; Hasegawa, H.; Suzuki, S.; et al. Application of iNKT Cell-targeted Active Immunotherapy in Cancer Treatment. Anticancer. Res. 2018, 38, 4233–4239. [Google Scholar] [CrossRef]
- Gebremeskel, S.; Clattenburg, D.R.; Slauenwhite, D.; Lobert, L.; Johnston, B. Natural killer T cell activation overcomes immunosuppression to enhance clearance of postsurgical breast cancer metastasis in mice. OncoImmunology 2015, 4, e995562. [Google Scholar] [CrossRef]
- Li, Y.-R.; Zhou, Y.; Kim, Y.J.; Zhu, Y.; Ma, F.; Yu, J.; Wang, Y.-C.; Chen, X.; Li, Z.; Zeng, S.; et al. Development of allogeneic HSC-engineered iNKT cells for off-the-shelf cancer immunotherapy. Cell Rep. Med. 2021, 2, 100449. [Google Scholar] [CrossRef]
- Toyoda, T.; Kamata, T.; Tanaka, K.; Ihara, F.; Takami, M.; Suzuki, H.; Nakajuma, T.; Ikeuchi, T.; Kawasaki, Y.; Hanaoka, H.; et al. Phase II study of α-galactosylceramide-pulsed antigen-presenting cells in patients with advanced or recurrent non-small cell lung cancer. J. Immunother. Cancer 2020, 8, e000316. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.-R.; Zhou, Y.; Wilson, M.; Kramer, A.; Hon, R.; Zhu, Y.; Fang, Y.; Yang, L. Tumor-Localized Administration of α-GalCer to Recruit Invariant Natural Killer T Cells and Enhance Their Antitumor Activity against Solid Tumors. Int. J. Mol. Sci. 2022, 23, 7547. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Guo, J.; Bao, X.; Xiong, F.; Ma, Y.; Tan, B.; Yu, L.; Zhao, Y.; Lu, J. Adoptive Transfer of Autologous Invariant Natural Killer T Cells as Immunotherapy for Advanced Hepatocellular Carcinoma: A Phase I Clinical Trial. Oncologist 2021, 26, e1919–e1930. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Bao, X.; Liu, F.; Guo, J.; Wu, Y.; Xiong, F.; Lu, J. Efficacy of Invariant Natural Killer T Cell Infusion Plus Transarterial Embolization vs Transarterial Embolization Alone for Hepatocellular Carcinoma Patients: A Phase 2 Randomized Clinical Trial. JHC 2023, 10, 1379–1388. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Cheng, X.; Jin, Y.; Xia, B.; Qin, R.; Zhang, W.; Hu, H.; Mao, X.; Zhou, L.; Yan, J.; et al. Safety and Clinical Response to Combined Immunotherapy with Autologous iNKT Cells and PD-1+CD8+ T Cells in Patients Failing First-line Chemotherapy in Stage IV Pancreatic Cancer. Cancer Res. Commun. 2023, 3, 991–1003. [Google Scholar] [CrossRef] [PubMed]
- Frontiers|α-GalCer and iNKT Cell-Based Cancer Immunotherapy: Realizing the Therapeutic Potentials n.d. Available online: https://www.frontiersin.org/articles/10.3389/fimmu.2019.01126/full (accessed on 2 October 2023).
- Ishibashi, F.; Sakairi, Y.; Iwata, T.; Moriya, Y.; Mizobuchi, T.; Hoshino, H.; Yoshida, S.; Hanaoka, H.; Yoshino, I.; Motohashi, S. A phase I study of loco-regional immunotherapy by transbronchial injection of α-galactosylceramide-pulsed antigen presenting cells in patients with lung cancer. Clin. Immunol. 2020, 215, 108457. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Springfield, R.; Chen, S.; Li, X.; Feng, X.; Moshirian, R.; Yang, R.; Yuan, W. α-GalCer and iNKT Cell-Based Cancer Immunotherapy: Realizing the Therapeutic Potentials. Front. Immunol. 2019, 10, 1126. [Google Scholar] [CrossRef] [PubMed]
- Nowak, M.; Schmidt-Wolf, I.G.H. Natural Killer T Cells Subsets in Cancer, Functional Defects in Prostate Cancer and Implications for Immunotherapy. Cancers 2011, 3, 3661–3675. [Google Scholar] [CrossRef]
- Tahir, S.M.; Cheng, O.; Shaulov, A.; Koezuka, Y.; Bubley, G.J.; Wilson, S.B.; Balk, S.P.; Exley, M.A. Loss of IFN-gamma production by invariant NK T cells in advanced cancer. J. Immunol. 2001, 167, 4046–4050. [Google Scholar] [CrossRef]
- Yanagisawa, K.; Seino, K.; Ishikawa, Y.; Nozue, M.; Todoroki, T.; Fukao, K. Impaired Proliferative Response of Vα24 NKT Cells from Cancer Patients Against α-Galactosylceramide1. J. Immunol. 2002, 168, 6494–6499. [Google Scholar] [CrossRef]
- Cortesi, F.; Delfanti, G.; Grilli, A.; Calcinotto, A.; Gorini, F.; Pucci, F.; Luciano, R.; Grioni, M.; Recchia, A.; Benigni, F.; et al. Bimodal CD40/Fas-Dependent Crosstalk between iNKT Cells and Tumor-Associated Macrophages Impairs Prostate Cancer Progression. Cell Rep. 2018, 22, 3006–3020. [Google Scholar] [CrossRef]
- Wang, Y.; Sedimbi, S.; Löfbom, L.; Singh, A.K.; Porcelli, S.A.; Cardell, S.L. Unique invariant natural killer T cells promote intestinal polyps by suppressing TH1 immunity and promoting regulatory T cells. Mucosal Immunol. 2018, 11, 131–143. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.-J.; Huang, J.-R.; Tsai, Y.-C.; Hung, J.-T.; Wu, D.; Fujio, M.; Wong, C.-H.; Yu, A.L. Potent immune-modulating and anticancer effects of NKT cell stimulatory glycolipids. Proc. Natl. Acad. Sci. USA 2007, 104, 10299–10304. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Basabe, A.; Burrello, C.; Lattanzi, G.; Botti, F.; Carrara, A.; Cassinotti, E.; Caprioli, F.; Facciotti, F. Human intestinal and circulating invariant natural killer T cells are cytotoxic against colorectal cancer cells via the perforin–granzyme pathway. Mol. Oncol. 2021, 15, 3385–3403. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, K.; Ueda, S.; Kawamura, M.; Aoshima, H.; Satoh, M.; Nakabayashi, J.; Fujii, S. Combination of cancer vaccine with CD122-biased IL-2/anti-IL-2 Ab complex shapes the stem-like effector NK and CD8+ T cells against tumor. J. Immunother. Cancer 2023, 11, e006409. [Google Scholar] [CrossRef] [PubMed]
- Carreño, L.J.; Kharkwal, S.S.; Porcelli, S.A. Optimizing NKT cell ligands as vaccine adjuvants. Immunotherapy 2014, 6, 309–320. [Google Scholar] [CrossRef]
- Zhang, H.; Bai, L. Challenges of iNKT cell-based antitumor immunotherapies. Cell Mol. Immunol. 2021, 18, 1077–1078. [Google Scholar] [CrossRef]
- Kunii, N.; Horiguchi, S.; Motohashi, S.; Yamamoto, H.; Ueno, N.; Yamamoto, S.; Sakurai, D.; Taniguchi, M.; Nakayama, T.; Okamoto, Y.; et al. Combination therapy of in vitro-expanded natural killer T cells and α-galactosylceramide-pulsed antigen-presenting cells in patients with recurrent head and neck carcinoma. Cancer Sci. 2009, 100, 1092–1098. [Google Scholar] [CrossRef]
- Shimizu, K.; Goto, A.; Fukui, M.; Taniguchi, M.; Fujii, S. Tumor Cells Loaded with α-Galactosylceramide Induce Innate NKT and NK Cell-Dependent Resistance to Tumor Implantation in Mice1. J. Immunol. 2007, 178, 2853–2861. [Google Scholar] [CrossRef]
- Fujii, S.-I.; Shimizu, K. Immune Networks and Therapeutic Targeting of iNKT Cells in Cancer. Trends Immunol. 2019, 40, 984–997. [Google Scholar] [CrossRef]
- Ashrafizadeh, M.; Delfi, M.; Zarrabi, A.; Bigham, A.; Sharifi, E.; Rabiee, N.; Paiva-Santos, A.C.; Prem Kumar, A.; Cheng Tan, S.; Hushmandi, K.; et al. Stimuli-responsive liposomal nanoformulations in cancer therapy: Pre-clinical & clinical approaches. J. Control. Release 2022, 351, 50–80. [Google Scholar] [CrossRef]
- Jia, J.-X.; Peng, S.-L.; Kalisa, N.Y.; Chao, Q.; Zhou, Z.; Gao, X.-D.; Wang, N. A liposomal carbohydrate vaccine, adjuvanted with an NKT cell agonist, induces rapid and enhanced immune responses and antibody class switching. J. Nanobiotechnol. 2023, 21, 175. [Google Scholar] [CrossRef]
- Khan, M.A.; Khan, A. Role of NKT Cells during Viral Infection and the Development of NKT Cell-Based Nanovaccines. Vaccines 2021, 9, 949. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, T.; Yamazaki, D.; Yamauchi, J.; Harashima, H. The nanoparticulation by octaarginine-modified liposome improves α-galactosylceramide-mediated antitumor therapy via systemic administration. J. Control. Release 2013, 171, 216–224. [Google Scholar] [CrossRef]
- Stolk, D.A.; de Haas, A.; Vree, J.; Duinkerken, S.; Lübbers, J.; van de Ven, R.; Ambrosini, M.; Kalay, H.; Brujins, S.; van der Vliet, H.J.; et al. Lipo-Based Vaccines as an Approach to Target Dendritic Cells for Induction of T- and iNKT Cell Responses. Front. Immunol. 2020, 11, 990. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Dong, C. DePICting T cell–APC crosstalk in cancer. Nat. Cancer 2022, 3, 265–267. [Google Scholar] [CrossRef] [PubMed]
- Kürten, C.H.L.; Kulkarni, A.; Cillo, A.R.; Santos, P.M.; Roble, A.K.; Onkar, S.; Reeder, C.; Lang, S.; Chen, X.; Duvvuri, U.; et al. Investigating immune and non-immune cell interactions in head and neck tumors by single-cell RNA sequencing. Nat. Commun. 2021, 12, 7338. [Google Scholar] [CrossRef] [PubMed]
- Sathe, A.; Grimes, S.M.; Lau, B.T.; Chen, J.; Suarez, C.; Huang, R.J.; Poultsides, G.; Ji, H.P. Single-Cell Genomic Characterization Reveals the Cellular Reprogramming of the Gastric Tumor Microenvironment. Clin. Cancer Res. 2020, 26, 2640–2653. [Google Scholar] [CrossRef]
- Chen, S.; Zhou, Z.; Li, Y.; Du, Y.; Chen, G. Application of single-cell sequencing to the research of tumor microenvironment. Front. Immunol. 2023, 14, 1285540. [Google Scholar] [CrossRef]
- Zhou, L.; Adrianto, I.; Wang, J.; Wu, X.; Datta, I.; Mi, Q.-S. Single-Cell RNA-Seq Analysis Uncovers Distinct Functional Human NKT Cell Sub-Populations in Peripheral Blood. Front. Cell Dev. Biol. 2020, 8, 384. [Google Scholar] [CrossRef]
- Xie, Z.; Huang, J.; Li, Y.; Zhu, Q.; Huang, X.; Chen, J.; Wei, C.; Luo, S.; Yang, S.; Gao, J. Single-cell RNA sequencing revealed potential targets for immunotherapy studies in hepatocellular carcinoma. Sci. Rep. 2023, 13, 18799. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
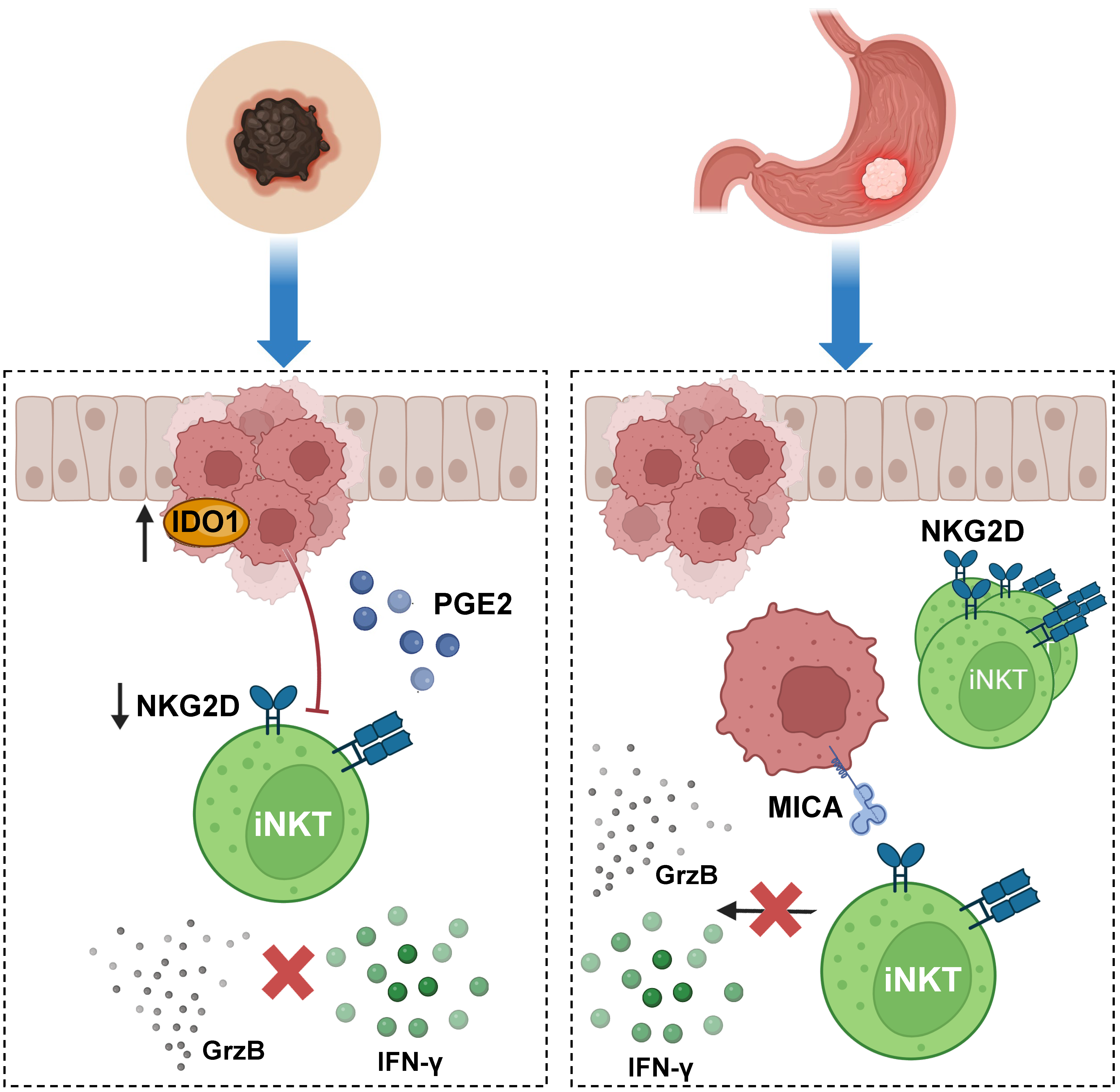{kind=link}
| Cells | Cancer Cell Type (In Vitro) | Effects In Vitro | Model In Vivo | Effects In Vivo | Reference |
|---|---|---|---|---|---|
| Allogeneic Human Stem cell- Engineered iNKT (AlloHSCiNKT cells) | A375 (Melanoma), K562 (Myelogenous leukemia, H292 (Lung cancer), PC3 (Prostate cancer), MM.1S (Multiple myeloma) | Reduction in metastasis, enhanced tumor killing efficacy | Human melanoma xenografth NSG mouse | Suppresed tumor growth | Ruide Li et al., 2021 [201] |
| α-GalCer-pulsed APCs to activate iNKT cells | - | - | NSCLC | Decreased iNKT and increase in NK cells, interferon-γ-producing cells and effector CD8+ T cells | Toyoda et al., 2020 [202] |
| PBMC-iNKT | A375-CD1d-FG and H292-CD1d-FG | Enhanced tumor cell killing and cytotoxic function | Human lung cancer xenograft NSG mouse | Enhanced antitumor ability | Ruide Li et al., 2022 [203] |
| Hepatocellular carcinoma | Greater production of Th1 cytokines and less IL-4. Safe and well tolerated treatment | Gao et al., 2021 [204] | |||
| Unresectable hepatocellular carcinoma after TACE failure | Showed improvements in the development of the disease in comparation to TAE treatment alone | Guo et al., 2023 [205] | |||
| Stage IV Pancreatic Cancer | Safe treatment, prolonged survival | Wang et al., 2023 [206] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tognarelli, E.I.; Gutiérrez-Vera, C.; Palacios, P.A.; Pasten-Ferrada, I.A.; Aguirre-Muñoz, F.; Cornejo, D.A.; González, P.A.; Carreño, L.J. Natural Killer T Cell Diversity and Immunotherapy. Cancers 2023, 15, 5737. https://doi.org/10.3390/cancers15245737
Tognarelli EI, Gutiérrez-Vera C, Palacios PA, Pasten-Ferrada IA, Aguirre-Muñoz F, Cornejo DA, González PA, Carreño LJ. Natural Killer T Cell Diversity and Immunotherapy. Cancers. 2023; 15(24):5737. https://doi.org/10.3390/cancers15245737
Chicago/Turabian StyleTognarelli, Eduardo I., Cristián Gutiérrez-Vera, Pablo A. Palacios, Ignacio A. Pasten-Ferrada, Fernanda Aguirre-Muñoz, Daniel A. Cornejo, Pablo A. González, and Leandro J. Carreño. 2023. "Natural Killer T Cell Diversity and Immunotherapy" Cancers 15, no. 24: 5737. https://doi.org/10.3390/cancers15245737