Understanding the Risks of Diffusion of Cyanobacteria Toxins in Rivers, Lakes, and Potable Water
by
 , , , and
, , , and
Amin Mahmood Thawabteh
1,2 ,
,
Hani A Naseef
1 ,
,
Donia Karaman
3, 
Sabino A. Bufo
4,5,
Laura Scrano
6 and
and
Rafik Karaman
3,4,*
1
Faculty of Pharmacy, Nursing and Health Professions, Birzeit University, Ramallah 00972, Palestine
2
General Safety Section, General Services Department, Birzeit University, Bir Zeit 71939, Palestine
3
Faculty of Pharmacy, Al-Quds University, Jerusalem 20002, Palestine
4
Department of Sciences, University of Basilicata, Via dell’Ateneo Lucano 10, 85100 Potenza, Italy
5
Department of Geography, Environmental Management and Energy Studies, University of Johannesburg, Auckland Park Kingsway Campus, Johannesburg 2092, South Africa
6
Department of European and Mediterranean Cultures, University of Basilicata, Via Lanera 20, 75100 Matera, Italy
*
Author to whom correspondence should be addressed.
Toxins 2023, 15(9), 582; https://doi.org/10.3390/toxins15090582
Submission received: 28 June 2023
/
Revised: 10 August 2023
/
Accepted: 18 September 2023
/
Published: 20 September 2023
Abstract
:Blue-green algae, or cyanobacteria, may be prevalent in our rivers and tap water. These minuscule bacteria can grow swiftly and form blooms in warm, nutrient-rich water. Toxins produced by cyanobacteria can pollute rivers and streams and harm the liver and nervous system in humans. This review highlights the properties of 25 toxin types produced by 12 different cyanobacteria genera. The review also covered strategies for reducing and controlling cyanobacteria issues. These include using physical or chemical treatments, cutting back on fertilizer input, algal lawn scrubbers, and antagonistic microorganisms for biocontrol. Micro-, nano- and ultrafiltration techniques could be used for the removal of internal and extracellular cyanotoxins, in addition to powdered or granular activated carbon, ozonation, sedimentation, ultraviolet radiation, potassium permanganate, free chlorine, and pre-treatment oxidation techniques. The efficiency of treatment techniques for removing intracellular and extracellular cyanotoxins is also demonstrated. These approaches aim to lessen the risks of cyanobacterial blooms and associated toxins. Effective management of cyanobacteria in water systems depends on early detection and quick action. Cyanobacteria cells and their toxins can be detected using microscopy, molecular methods, chromatography, and spectroscopy. Understanding the causes of blooms and the many ways for their detection and elimination will help the management of this crucial environmental issue.
Keywords:
cyanobacteria blooms; cyanotoxins; Microcystis; Anabaena; cylindrospermopsin; satellite imagery; biocontrolKey Contribution: This article focuses on cyanobacteria, looking into their origins, how they spread, and any issues connected to this type of toxicity diffusion in water. Many strategies are also discussed to reduce these risks and secure drinking water.
1. Introduction
On occasion, drinking tap water results in an earthy flavor that is detectable right away. In addition to colored scum that can develop on river surfaces, foam can occasionally form on lake or rive’ surfaces. Cyanobacteria are depicted in Figure 1 [1,2,3,4] as the colorful scum and the earthy taste of tap water. This unpleasant, old-growth organism contaminates many of our freshwater supplies, which threatens human health. While cyanobacteria are naturally present in our rivers and water supply, green growth and environmental modifications brought on by human activity might hasten their spread [2,3,4,5,6,7]. Our rivers and tap water may include cyanobacteria. These tiny bacteria can swiftly produce blooms in warm, nutrient-rich water, endangering humans and aquatic life. Toxins produced by cyanobacteria include Microcystins, which are well-known for their toxicity and capacity to harm plants. These poisons contaminate rivers and streams and threaten our drinking water supply since they can sneak through standard water treatment systems unnoticed [6,7,8].
Since these bacteria thrive in warmer climates, they pose a severe risk when river levels rise in the summer. To make matters worse, getting rid of cyanobacteria from nearby bodies of water can be challenging once a bloom has occurred. Because of this, we must comprehend the dangers of cyanobacteria and how to stop their spread in our drinking water [2,7,8,9].
When there is an abundance of warm, stagnant water and high amounts of phosphorus and nitrogen from sources like agricultural runoff or untreated sewage, cyanobacteria blooms commonly take place. Additionally, elements like global warming and the frequency of extreme weather events may make these blooms more likely [8,9,10].
The summer months experience mild temperatures, when these blossoms most frequently appear. They can also happen throughout extended periods of low precipitation and dryness. Due to long-term excess nutrient loading in a specific body of water, heavy rains may occasionally result in cyanobacterial blooms [11].
It is essential to remember that blooms can occur at any moment, so it is best to exercise caution when swimming in water bodies susceptible to blooming. Even if tests show that a river or lake’s cyanobacterial toxin levels are low, if a bloom happens when a person is swimming there, they may still be at risk [10,11,12]. In more detail, we will examine the topic in this review, learning where cyanobacteria originate, how they spread, and what dangers come with this form of toxicity diffusion in our waters. We will also discuss some measures implemented to reduce these hazards and maintain the safety of our tap water.
2. Common Genera of Cyanobacteria and Their Characteristics
In practically every freshwater ecosystem, cyanobacteria can be found alone or in combination with other organisms. In addition, there are numerous distinct cyanobacteria genera, each with unique properties. The common cyanobacteria genera discovered in freshwater systems are Synechococcus, Anabaena, Rivularia, Gloeotrichia, Oscillatoria, Cylindrospermopsis, Aphanizomenon, Planktothrix, Scytonema, Tolypothrix, Merismopedia, and Microcystis [13,14,15,16].
2.1. Microcystis: Colonial, Spherical Cells, Toxin Producer, Toxic, and Bloom-Forming
One of the most prevalent genera of cyanobacteria found in freshwater environments is Microcystis, a genus of colonial, spherical cells. Microcystis can produce toxins; they are poisonous and can cause blooms. Their colonies can be any shade of green, from deep olive to vivid blue-green. Microcystis colonies expand swiftly and have a milliliter cell density of over 1 million [14,15,16]. They prefer to form mats on the water’s surface along with other species like Anabaena, Oscillatoria, or Aphanizomenon and are photosynthetic organisms [17,18,19].
Moving water like rivers or streams and standing water like ponds, lakes, or reservoirs can harbor Microcystis. Because of the lower levels of oxygen and warmer temperatures throughout the summer, blooms are more likely to happen. Due to decreased oxygen levels and the development of chemicals that can harm humans and plant life, the blooms may result in drop-down water quality [20,21].
2.2. Anabaena: Filamentous, Heterocysts for Nitrogen Fixation and Diazotrophic
Anabaena inhabits environments in freshwater. These cyanobacteria are particularly important for their capacity to fix nitrogen since they include heterocyst cells capable of doing so. Anabaena also has akinetes cells, which act as protective spores and can withstand harsh environmental conditions [22,23].
The filaments that make up an Anabaena comprise individual trichome cells, which are one cell thick. Each trichome contains photosynthetic cells with a thick cell wall and an external peptidoglycan coating. The nucleoids, thylakoids, and carboxysomes of the cells are home to the enzymes involved in photosynthesis. Along the length of the trichome, heterocysts act as the central locations for nitrogen fixation [23,24,25]. Anabaena has undergone extensive research due to its ability to produce diazotrophic endospores that can be used to study other cyanobacterial species further. It is commonly used in aquatic systems to increase their nutritional content because of its ability to fix atmospheric nitrogen [25,26]. By doing so, it can supply other species with fixed nitrogen in settings where it is scarce or absent.
2.3. Oscillatoria: Filamentous, Motile, No Heterocysts
Oscillatoria is a widespread genus of freshwater cyanobacteria, the bacteria that cause bodies of water to get murkier and greener. It has a slimy, thin filamentous structure. Inhabiting shallow and deep regions of nutrient-rich water, such as seas, rivers, and lakes, Oscillatoria prefers to coil in an oscillatory motion [27,28].
It belongs to the cyanobacteria family and is not heterocytic, meaning it does not have heterocysts, a specific cell that fixes nitrogen in plants’ roots. Oscillatoria is frequently grouped with the Chroococcales family regarding their growth and physiological circumstances [29].
Under some circumstances, these mobile bacteria can create floating mats of colonies on the water’s surface. Oscillatoria can withstand dry seasons’ droughts thanks to the viscous slime that coats its cell walls. Furthermore, by utilizing nitrates to generate energy, it can survive in low light or even complete darkness because of its unique photosynthesis style [28,29,30].
2.4. Cylindrospermopsis: Filamentous, Motile, Toxin Producer
A filamentous and mobile genus of cyanobacteria called Cylindrospermopsis makes toxins. Freshwater systems like lakes and rivers and artificial reservoirs like ponds and canals frequently contain it. In temperate areas, this genus displays sporadic flowers in the summer [31].
Cylindrospermopsis spreads swiftly and takes over a system due to its motile habit, which makes it tough to eradicate. Despite being filamentous creatures, they can develop both deep blooms that can extend several meters below the surface of the water and enormous mats close to the water’s surface [32,33].
In addition to hepatotoxins (cyanotoxins), neurotoxins (ciguatoxins), and components of bacterial cell walls (lipopolysaccharides or LPS), Cylindrospermopsis species also produce toxins. Scientists must watch for these species in freshwater systems because these cyanotoxins are very hazardous to people and animals, even at low concentrations [32,33,34].
2.5. Aphanizomenon: Filamentous, Motile, Nitrogen Fixer
The genus Aphanizomenon of motile, free-floating filamentous cyanobacteria is widely found in freshwater bodies of water. They are known as nitrogen fixers because they can convert atmospheric nitrogen into a form that plants can utilize. As a result, they contribute nutrients to the food chain and maintain high water quality, making them vital to aquatic environments [35,36]. Certain Aphanizomenon species can harm humans and other animals when present in more significant proportions than usual. To further comprehend the potential risks these cyanobacteria pose, it is essential to frequently sample water and evaluate the levels of cyanobacterial metabolites [37]. Aphanizomenon is usually found in blossoms; therefore, that should also be considered. This indicates that an increase in their population may be caused by their quick growth, spurred by an abundance of nutrients or higher temperatures. It can help to prevent possible damage from increased toxin production or other negative impacts on aquatic plants and fauna by routinely checking for bloom indications [38,39].
2.6. Planktothrix: Filamentous, Motile, No Heterocysts
Numerous distinct species of cyanobacteria that have evolved to flourish in freshwater ecosystems comprise the Planktothrix genus. Planktothrix agardhii, P. rubescens, and P. limnetica are the most prevalent species. Due to their preference for nutrient-rich waters, these organisms are frequently found in eutrophic lakes with high levels of phosphate and nitrogen [40,41,42].
Phytotoxicity diffusion is the process of producing toxic exudates on the cell walls of Planktothrix species due to dense blooms that decompose. Due to the pigments (such as carotenoids) they produce, Planktothrix blooms can also cause water to become discolored [43]. In addition, Planktothrix is renowned for its capacity to multiply quickly and, under the right circumstances, to create dense aggregates or mats at the water’s surface. Optimal temperatures for Planktothrix development and survival in freshwater systems are between 15 and 25 °C and photosynthetically active radiation (PAR) [43,44].
2.7. Synechococcus: Unicellular and Colonial
The most prevalent genus of cyanobacteria is Synechococcus. It can be single-celled or colonial, consisting of one or more cells that can cluster and form filaments or colonies containing single or multiple cells. These colonies typically have a spherical form, a single gas exchange aperture, and a polysaccharide wall [45,46].
Synechococcus is renowned for its capacity to endure harsh conditions, including acidic and hypersaline waters. Due to its tolerance for high salt concentrations, it is also a significant species in coral reefs and soils, both aquatic environments. Its capacity for photosynthesis has also made it a vital component of the oxygen cycle in aquatic settings [47,48,49].
2.8. Rivularia: The Bubble-like Colonies
Rivularia create colonies that resemble bubbles, with each cell living inside a separate, protective “home”. Several characteristics set Rivularia apart from other cyanobacterial taxa. First, unlike most other cyanobacteria, Rivularia colonies are relatively solid in the water. The individual cells each have a distinct shape, with flat bottoms and rounded tops [50,51,52].
2.9. Gloeotrichia: Filamentous Cyanobacteria with Distinct Branching
The Gloeotrichia genus is distinguished by its distinctive branching, which gives its members the appearance of a starburst or a dandelion gone berserk. Due to their characteristic morphology, Gloeotrichia members are frequently observed in freshwater or brackish water habitats. They are anoxygenic phototrophs, producing sulfur rather than needing oxygen to use the light energy from photosynthesis [55,56].
These creatures’ internal sheaths have a spiral arrangement of cells that facilitates their movement through their environment, improving nutrition intake and mobility throughout your aquascape. The cell’s structure and surface area influence its interaction with its surroundings [56,57,58].
Most organisms have a variety of photosynthetic pigments in their photosynthetic system, allowing them to utilize light from locations where it is most abundant efficiently. Additionally, they have a variety of flagella to assist them in moving around in water deficient in nutrients or containing a lot of dissolved materials, such as salts and metals [58,59].
2.10. Scytonema: Irregularly Branched Filaments
Cyanobacteria belonging to the genus Scytonema have filaments that are erratically branched. When growing in colonies, cells can form cords, tufts, and even mats and are typically oriented spirally. The second name of this particular cyanobacterium species refers to the color of its distinctive brilliant reddish-brown spores. It can be found on mosses and lichens and often grows on damp surfaces [60,61].
Scytonema stands out from other cyanobacteria in several ways. It does not require light for photosynthesis, enabling it to flourish in dark places like caves or deep-sea cracks. A bud forms from the side of the bacteria cell and splits off to create a new organism, which is how they mainly reproduce [61,62].
2.11. Tolypothrix: Pseudoparenchymatous Filaments
Because of its pseudo-parenchymatous filaments, Tolypothrix can be identified. As a filamentous cyanobacterium that resembles a thread, Tolypothrix can join with other Tolypothrix cells to form lengthy chains of cells. This genus is extensively distributed worldwide and can be found in fresh and saltwater settings [64].
Typically comprised of four to six cells, Tolypothrix cells are protected from external challenges like UV radiation and drying out by a rigid coating consisting of glycoproteins and polysaccharides. The glycoprotein sheath keeps Their cell walls together, making it easier for them to build long chains [64,65,66].
Contrary to other cyanobacterial genera, such as rivularia or Gloeotrichia, Tolypothrix filaments rely on robust glycoprotein sheaths to make these connections rather than a visible sheath or stalk-like structure. This distinctive structure provides them a distinct edge when adapting to various settings, which explains why they are such a widespread genus globally [65,67].
2.12. Merismopedia: Cubicpacket-Shaped Cells
Merismopedia, a common genus of cyanobacteria, has a distinctive cubic packet-shaped cell and can split up to four times before finally dispersing. Merismopedia cells typically come in two shapes: flattened and cubed, ranging in size from 2 to 6 m [68,69]. When these cells cluster together in their surroundings, their morphology enables them to create a slimy, jelly like substance. This characteristic slimy mass typically comprises numerous layers with various types of Merismopedia cells [70].
Although it can also be found in wet soil and other moist habitats, Merismopedia is primarily found in oceans, seas, and other aquatic environments. Additionally, Merismopedia is frequently linked to cyanobacterial mats because of its ability to adapt to different light, temperature, and salt concentrations [71,72].
Merismopedia needs specific minerals like potassium and nitrogen to thrive appropriately and does it best in direct sunshine. It can grow rather quickly under ideal circumstances, such as temperatures between 25 and 35 °C and pH levels between 6 and 8.1. It is crucial to understand where Merismopedia is typically located and how to recognize it because this tiny but mighty genus of cyanobacteria can create toxins that can be dangerous to people if swallowed or even inhaled [71,72,73].
3. Cyanobacteria Diffusion and Spreads in Water Bodies
When cyanobacteria, single-celled organisms, disperse in water bodies, a process known as cyanobacteria diffusion and spread takes place. This may lead to higher concentrations of blue-green algae, or cyanobacteria, in certain conditions. It is critical to comprehend the elements that encourage bloom production and dissemination to understand the spread of this group of organisms [74,75,76].
3.1. Natural Factors Promoting Cyanobacteria Bloom Formation and Diffusion
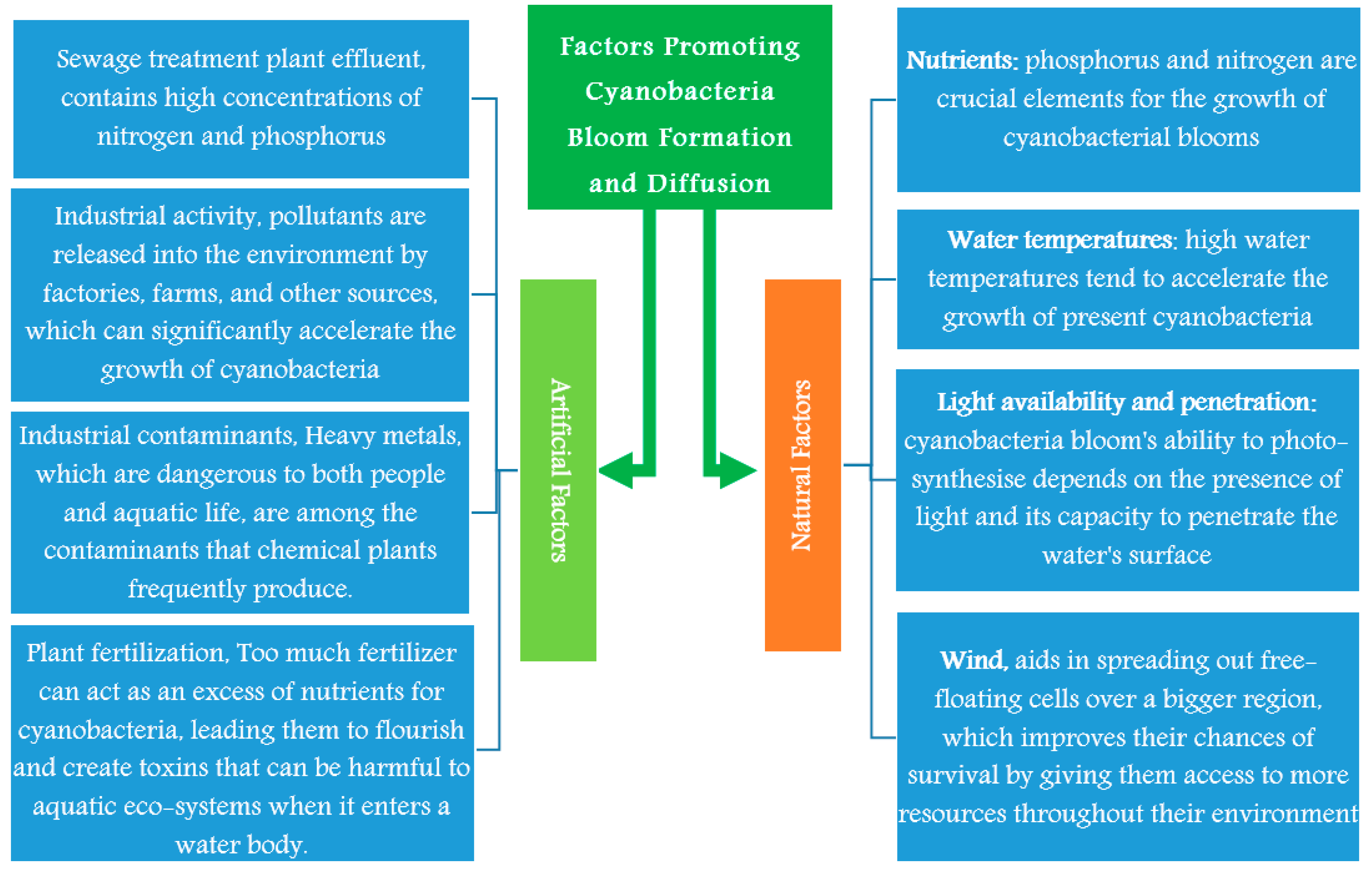
The availability of nutrients greatly influences blooms, and both phosphorus and nitrogen are crucial elements for the growth of cyanobacterial blooms. As a result, it is expected that some bloom may develop in water bodies where these nutrients are present in high concentrations [76,77,78]. Other factors as water temperatures, light availability and penetration will be discussed in this section and Figure 2.
First of all, phosphorus promotes cyanobacterial development and the production of toxic toxins. This is so it can function appropriately in cyanobacterial photosynthesis and cellular metabolism. Additionally, an overabundance of phosphorus can cause populations of cyanobacteria to increase quickly, resulting in a bloom [79]. On the other hand, because it is both a source of energy and a crucial element for the synthesis of cyanobacteria biomolecules, nitrogen also significantly impacts bloom development. As a result, too much nitrogen can alter the chemistry of the water, which can lead to a bloom [79,80].
Conversely, high water temperatures significantly aid cyanobacteria bloom development and dissemination. A body of water stratifies into two or more strata (thermoclines) according to its density as the temperature rises. This stratification prevents vertical mixing, which would otherwise separate nutrients near the surface from those in deeper waters, giving the cyanobacteria a surplus of food resources [81]. Additionally, a decline in vertical mixing may cause the near-surface waters to stagnate. Due to their tolerance to low oxygen levels, cyanobacteria can flourish in oxygen-depleted systems, creating the perfect setting for their growth. Additionally, high water temperatures tend to accelerate the growth of present cyanobacteria, which promotes the creation and diffusion of blooms [81,82,83].
Light availability and penetration are the third elements favoring the creation and diffusion of cyanobacterial blooms. The cyanobacteria bloom’s ability to photosynthesize depends on the presence of light and its capacity to penetrate the water’s surface. There are two types of light availability; depending on the atmosphere, it may be direct or indirect. While indirect light is sunlight reflected off an object or surface before entering the water, natural light enters the water directly but may be constrained by shadowing from other creatures. This could lead to uneven lighting, which would impact cyanobacteria blooms’ photosynthesis rate differently than direct light [83,84,85,86].
It is necessary to consider light penetration via water in addition to direct and indirect light. This depends on variables such as those that alter water clarity due to dissolved organic matter and the presence of other species suspended in the water column, which may limit the depth to which sunlight can penetrate. Knowing how light penetration and availability affect cyanobacteria blooms helps us understand where, when, and why these blooms occur as well as how to prevent them from occurring in undesirable locations [87,88].
The growth and diffusion of cyanobacteria blooms are significantly aided by water mixing and movement. One of the leading promoters or inhibitors of blooms is hydrodynamics, the study of the movement of fluids, which includes ocean currents, water flow, and wave action. Geographical location, seasonality, and a variety of environmental conditions, like as temperature gradients, all have a role in this. For example, a large-scale temperature gradient in the water might boost mixing, which is suitable for algae development—because of the blending, nutrients from the bottom rise to the surface, giving the algae more food [88,89,90].
Another significant aspect in the production and dispersion of cyanobacteria blooms is wind because it helps to stir up the water column, which again aids in bringing vital nutrients up from the bottom for algae growth. Additionally, it aids in spreading out free-floating cells over a more extensive region, which improves their chances of survival by giving them access to more resources throughout their environment [91,92].
These are all strongly connected to human activity, including climate change and eutrophication from excessive fertilizer runoff. It is well known that the presence of these elements alters the behavior of bacteria: they become less mobile, and their metabolic activity rises, assisting them in settling on substrates and remaining there more quickly. This causes cyanobacteria to spread throughout a river or even into drinkable water sources.
3.2. Artificial Factors Promoting Growth of Cyanobacteria Bloom Formation
The main artificial factors promoting the growth of cyanobacteria bloom formation are sewage treatment plant effluent, industrial activity, industrial contaminants, and plant fertilization; these sources will be discussed in this section and Figure 2.
One of the most critical sources of cyanobacterial toxicity in aquatic bodies is sewage treatment plant (STP) effluent. This is because STP effluent typically contains high concentrations of nitrogen and phosphorus released into water bodies, creating the perfect conditions for cyanobacteria to flourish. Additionally, these nutrients may build up in the water, causing eutrophication. As a result, cyanobacteria and other microbes proliferate quickly, causing blooms that may be hazardous to aquatic life. By disrupting oxygen levels and elevating pH levels, the chemicals emitted by these blooms can harm both plants and animals [93,94,95,96,97].
Wastewater treatment facilities must be appropriately maintained to ensure adequate nutrient removal from the effluent before it enters the receiving waterbody. This will help to limit the effects of STP effluents on water bodies. Additionally, to minimize excessive algal growth and lessen its impact on aquatic life, effective management measures should be implemented when dealing with excessive nutrients from agricultural runoff or urban areas [98,99,100,101,102].
Industrial activity is one of the main causes of cyanobacterial toxicity. Pollutants are released into the environment by factories, farms, and other sources, which can significantly accelerate the growth of cyanobacteria, or blue-green algae [103,104,105]. Nutrient runoff is the main factor driving the proliferation of cyanobacteria. Rainfall typically causes neighboring water bodies to rapidly expand in population by introducing nutrients like nitrogen and phosphorus [105,106,107,108,109].
Other industrial contaminants also aid the development of phytotoxic cyanobacteria. Heavy metals, dangerous to both people and aquatic life, are among the pollutants that chemical plants frequently produce. Additionally, agricultural practices can lead to a rise in contaminants like pesticides and fertilizers, which can lower the quality of water bodies by raising the likelihood of cyanobacterial growth [110,111,112,113,114,115,116].
Plant fertilization can result in cyanobacterial growth. Too much fertilizer can produce an excess of nutrients for cyanobacteria, leading them to flourish and create toxins that can harm aquatic ecosystems when they enter a water body. Therefore, it is crucial to take care while applying fertilizer close to their local water bodies [117,118,119]. In addition to causing cyanobacterial toxicity, excessive fertilizer use also causes eutrophication, in which algae density and biomass exceed normal levels due to an abundance of nutrients, depleting the water’s oxygen content and upsetting the local ecosystem [120,121,122,123].
4. The Common Toxins Produced from Cyanobacterial Blooms
Hazardous toxins, also called cyanotoxins, are present in cyanobacterial blooms. These cyanotoxins have been linked to several health issues, including cancer, liver damage, neurological problems, rashes, and skin irritation. Hepatotoxins, neurotoxins, dermatoxins, cytotoxins, and endotoxins are the most typical cyanotoxins produced by water blooms [124,125,126,127,128].
Hepatotoxins and neurotoxins are the two most prevalent cyanobacterial toxins. Hepatotoxins are harmful substances that harm the liver and can result in nausea, vomiting, and in severe cases, jaundice. When the neurological system is targeted by neurotoxins, symptoms such as headaches, nausea, muscle weakness, and even paralysis may result from exposure to excessive dosages. In addition, there are reproductive poisons that harm both men’s and women’s reproductive systems. If pregnant women are exposed to dangerous levels of cyanobacteria, this can lead to infertility or even miscarriage [126,127].
Hepatotoxic conditions are brought on by cyanotoxins, among which microcystin is one of the most often produced by cyanobacteria. Hepatotoxic chemicals build up in the water during dangerous cyanobacterial blooms, where they can be consumed through contaminated drinking water or inhaled. When a toxin enters the bloodstream and builds up in the liver, it damages the liver cells by rupturing their membranes [129,130,131].
Abdominal pain, nausea, and vomiting, as well as weakness, exhaustion, jaundice, dark urine, and abnormal liver enzymes, are indications of this disruption. Acute liver failure and possibly death can result from microcystin poisoning in severe situations [132,133,134].
When consuming food from polluted water, neurotoxic shellfish poisoning (NSP), which is brought on by the cyanotoxin anatoxin-A, is a serious cause for concern. Research suggests that it attaches to nicotinic acetylcholine receptors and interacts with our nervous system. This binding causes motor neurons to become paralyzed, which can cause problems with coordination, weakness, and in severe cases, even death [135,136].
However, it may take up to 12 h for symptoms to appear. They could include nausea, vomiting, headaches, and dizziness. When ingested in high doses, anatoxins can cause respiratory failure and muscle paralysis, which, if not treated right away, can soon result in death [136,137].
The common toxins created by cyanobacterial blooms is described in this section, and the analysis, methodologies, and mitigation techniques of cyanobacteria and cyanotoxins in water are discussed in Section 5.
4.1. Microcystin
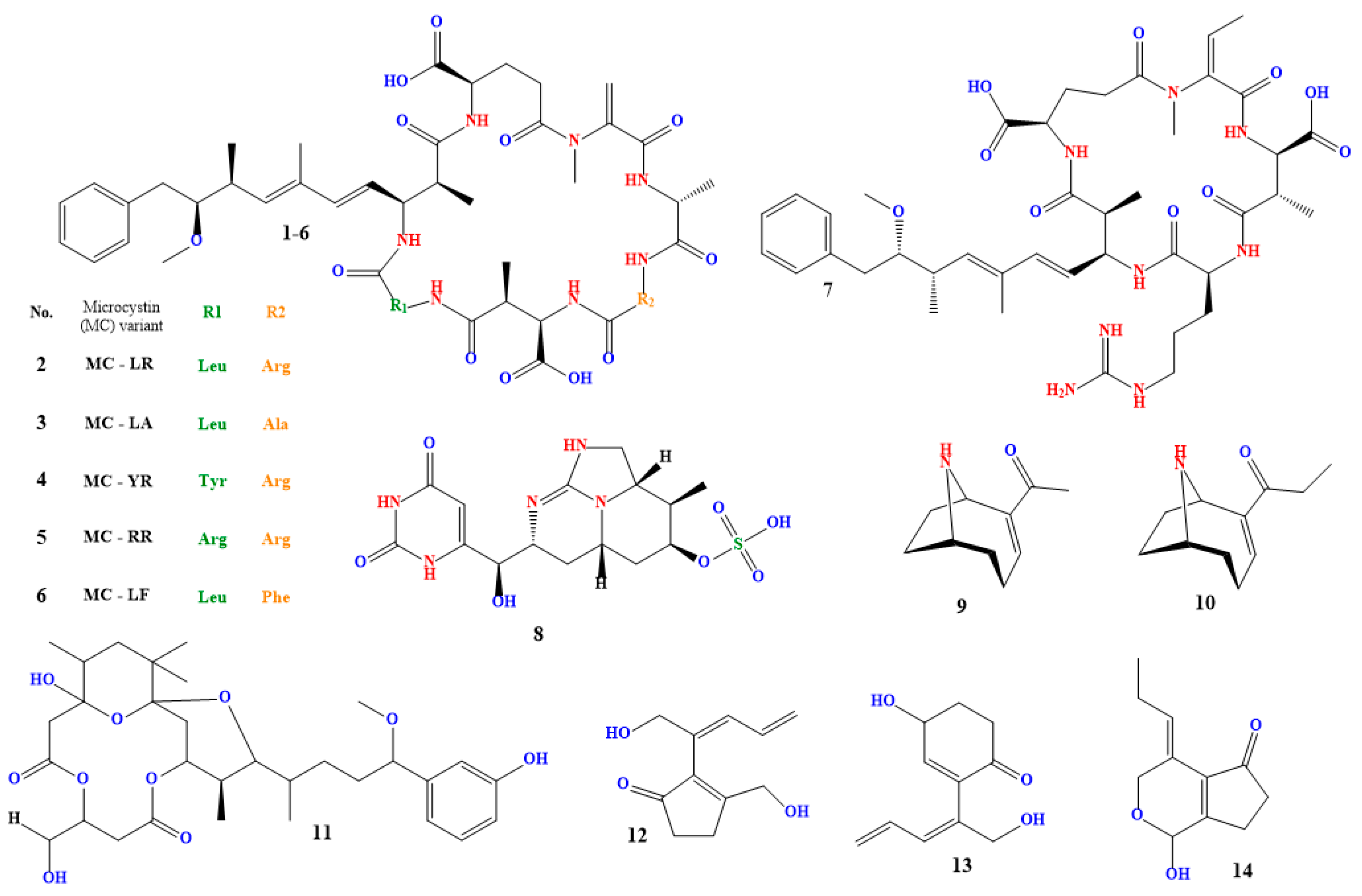
A class of cyclic hepatotoxins known as microcystins (compound 1 in Figure 3) are produced by many cyanobacterial species, including Microcystis aeruginosa, Anabaena spp., and Planktothrix spp. It is one of the cyanobacterial blooms’ most often made cyanotoxins [129,130,131].
Small peptides known as microcystins can be classified into various structural groups according to the amino acid make-up of their molecules [132,133]. Microcystin-LR (compound 2 in Figure 2), which has a leucine (L) and an arginine (R) residue in its structure, is the most extensively researched and toxic microcystin. Among others [134,135,136,137,138], other microcystin variants include microcystin-LA, microcystin-YR, microcystin-RR, and microcystin-LF (compounds 3–6 in Figure 3).
Microcystins are toxic to liver cells and can cause serious liver damage or even liver cancer in both humans and animals. They inhibit protein phosphatases, which are responsible for removing phosphate groups from proteins [139]. Because of the accumulation of phosphorylated proteins in liver cells, normal cellular function is disrupted, which is bad for the liver [140]. The kidney, reproductive system, and liver are all adversely affected by microcystins [141,142,143].
Ingestion of tainted water, skin contact, or inhalation of aerosolized water droplets containing the toxin are all ways people can be exposed to microcystins [141,142]. Although various nations may have their standards and laws regarding cyanotoxins in drinking water, the World Health Organization (WHO) has set a provisional recommendation value for microcystin-LR in drinking water of 1 g/L [143,144].
4.2. Nodularin
Nodularia spumigena, Nodularia harveyana, and Nodularia moravica are three species of cyanobacteria that generate nodularin (compound 7 in Figure 3), a cyclic hepatotoxin [145,146,147]. It exhibits the same toxicity mechanism as microcystin and is structurally identical to it. Nodularin inhibits protein phosphatases, which causes phosphorylated proteins to accumulate in liver cells and harm the liver. Nodularin is known to affect other organs, including the kidneys, and has been associated with the development of tumors in test animals [148,149,150].
Nodularin exposure can happen when a person consumes tainted water, comes into contact with it while touching their skin, or breathes in water droplets that have been aerosolized and contain the toxin [151,152,153,154]. Nodularin in drinking water has a provisional guideline value of 0.2 µg/L, according to the World Health Organization (WHO) [155,156,157,158].
4.3. Cylindrospermopsin
The cyanobacteria Cylindrospermopsis raciborskii, which is currently known as Raphidiopsis raciborskii and Aphanizomenon ovalisporum generate the toxin known as cylindrospermopsin (compound 8 in Figure 3). It is a tricyclic alkaloid that can be hazardous to humans and animals in acute and chronic phases [159,160,161,162]. Strong liver toxins, such as cylindrospermopsin, can cause cirrhosis and liver necrosis in people and experimental animals. In severe cases, it can also harm the kidneys, leading to renal failure. Cylindrospermopsin has also been shown to cause oxidative stress and DNA damage in cells [163,164].
When contaminated water is drunk, comes into contact with the skin, or is breathed as aerosolized water droplets, cyanospermopsin exposure can occur [165,166]. The World Health Organization (WHO) has set a preliminary recommended threshold for cylindrospermopsin in drinking water at 1 µg/L [167,168,169].
4.4. Anatoxin-a
Anabaena spp., Aphanizomenon spp., and Cylindrospermum spp. are cyanobacteria species that generate the powerful neurotoxin anatoxin-a (compound 9 in Figure 3). A secondary, bicyclic amine alkaloid with acute neurotoxicity, anatoxin-a is also referred to as Very Fast Death Factor. It was initially found in Canada in the early 1960s, and it was isolated in 1972. The potent nicotinic acetylcholine receptor agonist anatoxin-A irreversibly binds to these receptors in the central and peripheral nervous systems. As a result of the receptors being overstimulated, the release of the neurotransmitter acetylcholine, which causes muscle contractions, occurs. At high doses, anatoxin-A can cause respiratory collapse and death [170,171].
Ingestion of tainted water, skin contact, or inhalation of water droplets that have been aerosolized can all result in anatoxin-a exposure. Animal deaths and illnesses brought on by cyanobacterial blooms have frequently been attributed to anatoxin-a. The World Health Organization (WHO) has established a provisional guideline value for anatoxin-a in drinking water of 0.2 µg/L [172,173].
4.5. Homoanatoxin
Anabaena circinalis and Anabaena lemmermannii are cyanobacteria species that generate the toxin known as homoanatoxin (compound 10 in Figure 3). It is a cyclic alkaloid and a member of the anatoxins toxin subclass [174,175,176,177].
Homoanatoxin is a powerful neurotoxin that functions as a cholinergic agonist, much like anatoxin-a. In the central and peripheral neurological systems, it binds to nicotinic acetylcholine receptors, overstimulating the receptors and causing the release of acetylcholine, a neurotransmitter that causes muscle contractions. At high doses, homo-anatoxin can cause respiratory collapse and death [178].
4.6. Oscillatoxin A
Several cyanobacterial species, such as Oscillatoria spp. and Phormidium spp., produce the toxin known as oscillatoxin A (compound 11 in Figure 3). It has strong hepato- and neurotoxic properties [181,182]. Oscillatoxin A is a cyclic peptide toxin that shares structural similarities with microcystins. Protein phosphatases 1 and 2A, crucial enzymes involved in various cellular functions like cell division, metabolism, and death, are strongly inhibited by it. Cell damage, particularly in the liver and neurological system, can result from this inhibition [183].
4.7. Nakienones A–C
Toxins known as nikienones A–C (compounds 12–14 in Figure 3) are made by specific cyanobacteria, such as Nostoc and Anabaena. They are referred to as lipopeptides and are cytotoxic and antifungal. While nakienone C comprises three separate non-cyclic polypeptides, nakienones A and B are cyclic heptapeptides [185,186].
Toxins from nikienones A–C can cause the body harm, such as digestive problems, skin rashes, liver damage, infertility, and even death. Furthermore, it has been established that each of the three may have medical applications. Nakienone A is an anti-inflammatory and an antifungal, whilst nakienone B and C may potentially inhibit the proliferation of cancer cells. In some cell cultures, nikienone B has been found to have an anti-prostate cancer effect [185,186,187].
It is critical to remember that, like all cyanotoxins, these toxins can be deadly when consumed or inhaled in high doses, even though they may have qualities that make them useful for some medical therapies [188].
4.8. Aphantoxin
Aphantoxin made by Anabaena flosaquae and Aphanizomenon spp. It is a neurotoxin and a member of the PSPs (paralytic shellfish poisons) family. Aphantoxins work by blocking the neurotransmitters that regulate the relaxation and contraction of human muscle [189,190]. This results in severe muscle cramps or paralysis along with a variety of additional systemic symptoms like vomiting or altered mental states [191].
Consuming tainted water or shellfish that have collected the toxin are two ways people can be exposed to aphantoxin. Animal deaths and human illnesses related to cyanobacterial blooms have been linked to aphantoxin in multiple instances [192]. For aphantoxin in shellfish, the World Health Organization (WHO) has established a provisional guideline value of 20 µg/kg [192,193].
4.9. Debromoaplysiatoxin and Aplysiatoxin
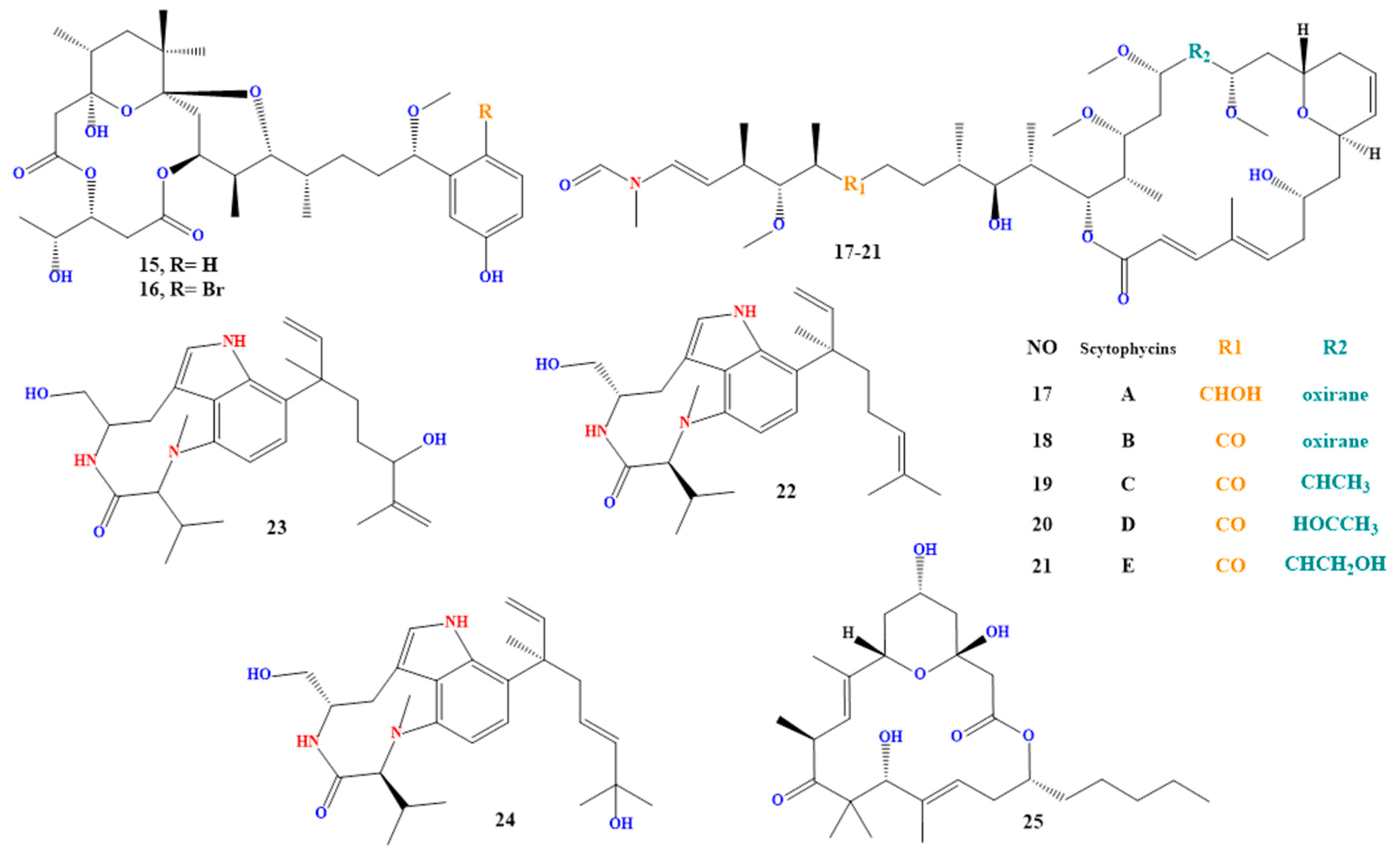
Aplysiatoxin and debromoaplysiatoxin (compounds 15 and 16 in Figure 4) are lipophilic toxins made by the fungi Lyngbya majuscula, Lyngbya sordida, and Schizothrix [194,195,196,197].
It is well known that these poisons are highly poisonous and can have various adverse health impacts on people and animals [190]. Exposure to these poisons can result in liver damage, neurotoxicity, and even death in extreme situations. Additionally, it has been discovered that they possess mutagenic and carcinogenic qualities, raising the risk of cancer. These poisons can be consumed through contact with contaminated water while engaging in recreational activities [196,197,198,199].
4.10. Scytophycins and Lyngbyatoxin
Both groups of toxins are lipophilic. Scytonema hofmanni produces scytophycins A to E (compounds 17 to 21 in Figure 4), which have been discovered in freshwater and marine settings and are harmful to aquatic and terrestrial species [200,201,202]. While Moorea producens and Lyngbya majuscula produce lyngbyatoxins A to C (compounds 22 to 24 in Figure 4). It has been demonstrated to have cytotoxic and apoptotic effects on cells [203,204] and is a potent protein synthesis inhibitor.
Exposure to scytophycin and lyngbyatoxin can have a variety of negative health effects on both humans and animals. It has been linked to skin irritation, respiratory problems, and liver damage [205]. Lyngbyatoxin has been demonstrated in lab studies to have a tumor-promoting effect, increasing the risk of cancer. Scytophycin has also been shown to have cytotoxic and apoptotic effects on cells, as well as neurotoxic effects on animals [205,206,207].
4.11. Acutiphycin
Acutiphycin is a class of indole alkaloid poisons produced by the genera Westiellopsis, Fischerella, and Hapalosiphon [208,209,210].
Acutiphycin exposure had a variety of adverse health impacts on both people and animals. This toxin’s ability to influence ribosomes and prevent the formation of peptide bonds makes it a potent protein synthesis inhibitor. Animals were also exposed to hepatotoxicity and neurotoxicity [211].
5. Managing and Mitigating Cyanobacterial Blooms and Toxins
Cyanobacteria can survive and grow in practically any environment. Increased human activity has caused climatic conditions to change, which favors the occurrence and severity of dangerous cyanobacterial blooms everywhere in the world. Even while some environmental variables, such as water temperature, pH, and nutrient levels, may be associated with higher cyanotoxin levels, the interaction of these variables with the particular cyanobacteria species present makes the issue complex. Effective management of a poisonous cyanobacterial bloom requires an understanding of how different environmental conditions affect the regulation of cyanotoxins [213,214,215].
As cyanobacterial blooms can occur in any body of fresh or salt water, it is crucial to know how to spot them. Early detection is essential because it enables quicker response times and more efficient management [216,217]. Additionally, it is critical to comprehend the hazards and probable health issues connected to cyanobacterial blooms [218,219].
Monitoring cyanobacteria is a crucial procedure that aids in assessing the condition of water bodies and identifying potential health risks to people and animals. Accurate sample collection is essential for producing results representative of the population. Choosing sampling sites that are indicative of the region of interest, such as those that are close to the coast, in the middle of the water body, or in areas where there is a high likelihood of finding cyanobacteria, are some factors that must be considered while collecting cyanobacteria samples. The site’s accessibility and security must also be considered [220,221,222,223]. Additionally, using sterile and non-toxic sampling tools like a plastic bucket or a sampler made specifically for cyanobacteria sampling. Avoid using metal containers, as they may change the material’s chemical. To obtain a representative sample of the water body, sample the water column vertically as another sampling technique. To capture any fluctuation, take repeated samples at various depths, particularly deep lakes or reservoirs. Each sample should include a minimum of 500 cc of water. Since samples are taken during the cyanobacteria’s active growing season, typically from late spring to early fall, it is crucial to pick the proper time to collect them. To prevent any time-of-day variations, collect samples simultaneously every day. To keep the samples from oxidizing, sodium thiosulfate at 0.1% is added. Within 24 h, transfer samples to the lab while keeping them on ice [223,224,225,226]. Techniques and methods for managing and mitigating cyanobacterial blooms and toxins are summarized in Figure 5.
5.1. Identifying and Measuring Cyanobacteria and Cyanotoxins
Cyanobacterial blooms may be identified with the naked eye due to their unusual appearance and color. If a blue-green algal bloom is observed, experts recommend sampling a nearby water source to learn more about the local flora and fauna. Furthermore, scientific examination of bloom samples might reveal the existence of specific bacterial strains and any possible toxins [227,228,229].
Microcystin, saxitoxin, cylindrospermopsin, anatoxin-A, sumoyltoxin A, and lyngbyatoxin A are the most prevalent cyanotoxins that must be watched. Toxin concentrations should be measured as part of a comprehensive risk assessment for aquatic or human health [230,231]. To track the growth of cyanobacteria in lakes and reservoirs, France established the Cyanobacteria Monitoring Network (CyMNet). More than 50 stations comprise this network, tracking variables like temperature, nutritional concentrations, and microcystin toxin levels [232,233,234].
These metrics offer crucial data for tracking and evaluating the dangers posed by cyanobacteria in various global ecosystems. To prevent and mitigate potentially deadly cyanobacterial blooms, one of the most important measures is accurately detecting and measuring cyanotoxins [235,236].
5.1.1. Microscopy Technology
Cyanobacteria cells are frequently found and recognized using a microscope. A standard method for locating and identifying cyanobacteria cells is light microscopy. The sample is viewed under a microscope when a light source and the cells illuminate. Cyanobacteria cells may be recognized by their distinctive blue-green hue, and the shape and size of the cells can determine different species. Another method for locating and identifying cyanobacteria cells is fluorescence microscopy. In this method, the cells are marked with a fluorescent dye and viewed using a fluorescence microscope. The fluorescent label can distinguish between various cyanobacteria cell types and find toxin concentrations [237,238].
A high-resolution method that can be utilized to see cyanobacteria cells and their structures at the nanoscale level is electron microscopy. In this method, the material is illuminated by an electron beam, and the cells are viewed via an electron microscope. Electron microscopy can be used to identify many structures, including thylakoids and gas vesicles, and can provide precise information on the internal structure of the cells. Confocal microscopy is a method that can be used to produce three-dimensional, high-resolution images of cyanobacteria cells. In this technique, a laser illuminated the sample, and several photographs are taken using various focus planes. A three-dimensional representation of the cells is then created by combining the photos [239,240].
There are not many case studies that show how to employ microscopy to find cyanobacterial cells. The blooms in Lake Taihu, a huge freshwater lake in China, were found during a survey to identify potentially harmful cyanobacterial blooms. The cyanobacterium Microcystis aeruginosa bloom caused a water crisis for millions of people in the vicinity. Cyanobacteria were found and identified using microscopy in the lake water sampled. The results of the microscope showed that the dominant cyanobacterium in the bloom was M. aeruginosa, which produces the potent hepatoxin microcystin [241]. Another study used microscopy to monitor the cyanobacterial populations during a 12-month period in a Brazilian eutrophic reservoir. The cyanobacteria population was dominated by Microcystis, Planktothrix, and Anabaena, according the microscope findings. The cyanobacteria communities fluctuated seasonally, with more diversity and abundance in the warmer months, according to another study finding [242].
The disadvantage of microscopic strategies is their lack of accuracy. The detection of microscopic cells is difficult due to the 200 nm resolution limit of optical microscopy. Erroneous cell estimations and non-homogeneous cell distribution may be brought on by the presence of dense colonies. An erroneous estimation of cell abundance results from the diverse dispersion of cells. However, the aggregation state of some Microcystis cyanobacterial colonies enables species identification based on morphology. Furthermore, lethal species identification and bloom lethality prediction are not possible with microscope examination. The inability to distinguish between toxic and non-toxic cyanobacteria, which prevents the risk assessment of a bloom, is increasingly the fundamental drawback of this technique [243,244].
5.1.2. Molecular Techniques
Cyanobacteria cells can also frequently be found using molecular methods. These methods entail searching for particular DNA identifiers or genes exclusive to cyanobacteria. Cyanobacteria can be found and identified using the molecular technique known as polymerase chain reaction (PCR). Specific DNA sequences of the cyanobacteria are amplified via PCR, making it simple to find and classify them. Other molecular methods include next-generation sequencing (NGS), which can sequence the entire genome of the cyanobacteria and compare it to known sequences in databases to identify the specifics, and fluorescent in situ hybridization (FISH), which uses fluorescently labeled DNA probes to bind to specific DNA sequences in the cyanobacteria cells [245,246,247].
In many studies, researchers used quantitative polymerase chain reaction PCR (qPCR) to locate and measure cyanobacteria in water samples from an Australian river. This is an illustration of how cyanobacterial cells can be found using molecular methods. The researchers extracted DNA from the water samples and used qPCR to amplify and quantify the mcyA gene, which is involved in producing microcystin, a characteristic cyanobacteria toxin. They found that cyanobacteria that produce microcystin were heavily concentrated in the river [248].
In a different investigation, quantitative polymerase chain reaction (qPCR) was used to count the cyanobacteria in a Japanese eutrophic lake. The qPCR method, which concentrated on the cyanobacteria’s 16S rRNA gene, facilitate the detection and quantification of a number of species, including Microcystis, Anabaena, and Aphanizomenon. The most common cyanobacterium in the lake, according to the study, was Microcystis, and its abundance was positively correlated with both the total nitrogen concentration and water temperature [249]. Additionally, qPCR and fluorescence in situ hybridization (FISH) were used in a study in a reservoir in Spain to identify and quantify harmful cyanobacterial species. FISH was used to determine the cyanobacteria’s physical features, and qPCR experiments concentrated on genes specific to Microcystis and Anabaena. Microcystis was found to be the dominant cyanobacterium in the reservoir and to be capable of producing the hepatotoxin microcystin [250].
qPCR has some limitations, one of which is the decrease in amplification efficiency with the length of the reaction product, despite the significant advantages in the detection and quantification of cyanobacteria and the genes that encode toxins, as well as the ability to detect toxic cyanobacteria in water long before the manifestation of cyanobacterial blooms. Furthermore, the transcription and translation of the toxin’s product by cyanobacteria are not always associated with the presence of an enlarged toxin-encoding gene. As a result, other techniques, such as chemical and biological processes, must be used to integrate and confirm qPCR results [251,252].
5.1.3. Chromatography and Spectroscopy
Chromatography and spectroscopy, two trusted techniques for examining in-water cyanobacterial cells, can be used to identify the presence of cyanobacterial pigments. While spectroscopy involves examining molecules via light absorption or reflection, chromatography is based on differences in the rate at which components migrate down a column or thin layer of adsorbent material. These two approaches work together to give researchers a thorough grasp of the toxins in a particular water system. In addition to identifying pigments, spectroscopy provides helpful information about the relative abundance or ratio of various pigments, which helps scientists more precisely assess potential risk levels. Numerous watery pigments, including beta-carotene, chlorophylls a and b, diadinoxanthin, and diatoxanthin, can be detected using chromatography. Microcystins and Cylindrospermopsin, compounds found in drinking water sources, can be seen by spectroscopy at very low quantities (1ppb) [253,254].
In a Lake Taihu, China investigation, HPLC was used to test and identify the microcystins produced by the common cyanobacterium Microcystis aeruginosa, which is responsible for harmful algal blooms. The investigation revealed that M. aeruginosa produced several microcystins, including the potent liver toxin microcystin-LR [255]. In another 2018 study—the authors used fluorescence spectroscopy to locate cyanobacteria cells and their toxic byproducts in water samples. Fluorescence spectra were utilized to differentiate between different cyanobacteria cell types and to detect the presence of microcystin in water samples. In order to swiftly and affordably identify cyanobacteria species and their toxins in water samples, fluorescence spectroscopy was used [256]. A portable, high-resolution fiber-optic Raman system and an ab-based Raman system, used for cell detection and identification, were used to study the cyanobacteria diffusion in the Adriatic Sea. In order to distinguish between distinct cyanobacterial species based on their spectral signatures, Raman spectroscopy was discovered to offer great sensitivity and specificity for detecting small amounts of cyanobacterial cells [257].
In a study conducted in a freshwater lake in France using GC/MS, volatile organic compounds (VOCs) produced by the common filamentous cyanobacterial species Anabaena flosaquae were investigated. Depending on the growth stage and environmental variables, A. flosaquae produces a variety of VOCs, including alkanes, alkenes, ketones, and alcohols [258].
Due to the existence of additional substances with comparable absorbance spectra, identification only based on UV absorbance is insufficient. Cyanotoxins are not specifically detected by the diode-array detector, which is also susceptible to interference from other analytes. Results are typically presented as microcystin-LR equivalents because many microcystin variants are difficult to identify and lack standards. Additionally, because some chemicals quickly degrade into anatoxin-A, chromatographic methods combined with UV spectroscopic detection are insufficiently sensitive, necessitating derivatization to increase their detection limits. Better sensitivity and selectivity are made possible by coupling with MS. On the other hand, the various LC-MS techniques make it rather simple and easy to analyze microcystins, nodularin, and cylindrospermopsin. However, utilizing LC-MS techniques to analyze highly polar chemicals with multiple isomers, such as saxitoxins, is extremely challenging [259,260,261].
Due to its outstanding sensitivity and selectivity, liquid chromatography with triple quadrupole mass spectrometry (LC-MS/MS) enables the precise identification and quantification of various and unidentified toxins in environmental materials. These MS/MS transitions are found when specific daugh-terons are produced from the precursor molecular ion in a collision cell [262].
m/z 135 [263] is a distinctive fragment ion shared by the majority of microcystins and nodularin. Despite having a different chemical structure from microcystins and nodularins, cylindrospermopsin is frequently analyzed alongside these cyanotoxins via LC-MS/MS using the particular MS-MS transitions m/z 416 > 336, 274, and 194 [264,265]. Anatoxin-a can be detected using the particular MS/MS transitions m/z 166 > 149, 131, 107, and 91 [266,267]. With the precise MS/MS transitions m/z 300 > 282, 204, saxitoxins can be found [268]. Table 1 lists various LC-MS and LC-MS/MS techniques for analyzing various cyanotoxins.
Chromatography techniques have some limits when it comes to detecting cyanotoxins, despite being effective and frequently used for the detection and analysis of many substances, including cyanotoxins. One type of sample complexity is as follows: the isolation and identification of cyanotoxins can be hampered by the presence of a complex matrix of organic and inorganic substances in water samples taken from natural bodies like lakes, rivers, and reservoirs. Reduced sensitivity and accuracy in quantification can be caused by matrix effects [269].
The use of chromatography techniques has drawbacks and limits due to sample complexity and sample preparation. A complex matrix of organic and inorganic chemicals, with varying chemical and physical properties, can obstruct the isolation and detection of cyanotoxins in water samples taken from lakes, rivers, and reservoirs. The sensitivity and accuracy of quantification can be decreased by matrix effects. Detection limits are also considered as one of the limitations of using chromatography techniques, as some cyanotoxins can be present in water at very low concentrations, which requires highly sensitive detection methods. Achieving lower detection limits using chromatography may be limited by factors such as sensitivity of the equipment and interference from the sample matrix. In addition to the development and validation of a chromatographic method for the analysis of cyanotoxins can be time consuming and resource intensive. The need to validate different methods for cyanotoxins and sample matrices can complicate the process [270].
In order to overcome these restrictions, chromatographic techniques are frequently combined with other techniques like immunoassays and biosensors.
5.1.4. Immunoassays
Immunoassays are frequently employed methods for identifying and measuring cyanotoxins. These methods, which rely on antibodies that are particular to the target cyanotoxin, can offer an efficient way to identify and measure these toxins in various environmental samples [271]. Cyanotoxins can be found using several different immunoassays, such as the following:
Enzyme-Linked Immunosorbent Assay (ELISA)
The Enzyme-Linked Immunosorbent Assay (ELISA) is an antibody-based assay which can be either monoclonal, specific for a certain molecule variant, or polyclonal, recognizing several types of molecules. ELISA are rapid and sensitive, and a low level of expertise is required. They are useful as primary quantitative screening tools with limits of detection around 0.1 μg/L, but they are susceptible to interferences that limit their use for quantitative analyses.
Anatoxin, microcystins, and cylindrospermopsin are a few examples of particular cyanotoxins that can be detected and measured using the widely used immunoassay known as ELISA. This method involves adding a sample to a microtiter plate coated with antibodies specific to the target cyanotoxin. A secondary antibody that is connected to an enzyme is introduced after the cyanotoxin in the sample binds to the antibodies. A reaction that the enzyme then catalyzes results in a measurable signal that may be measured using a spectrophotometer [272]. In a study published in 2019, ELISA was used to find anatoxin-a in surface water samples gathered from all around Victoria, Australia, where a cyanobacteria bloom occurred. They found anatoxin-a gene sequences in Cuspidothrix issatschenkoi, Aphanizomenon sp., D. circinale, Anabaena sp., and Oscillatoria sp., as well as their existence and distribution. The results indicate that the ELISA approach could be used to check water bodies for harmful algal blooms regularly and was successful in detecting anatoxin-a in water samples [272]. Table 1 summarizes the ELISA methods for the detection of microcystins, nodularins, and saxitoxins.
The use of widely accessible Enzyme-Linked Immunosorbent Assay (ELISA) test kits, which do not require pricey equipment or intensive training to use, is one of the most used cyanotoxin testing approaches. Semi-quantitative field screening ELISA kits for detecting the presence or absence of cyanotoxins are available. A repeat study using a quantitative ELISA test or one of the other analytical techniques is recommended if cyano-toxins are found by a field screening kit. More accurate, quantitative ELISA test kits are available for microcystins, nodularins, saxitoxin, anatoxin-a, and cylindrospermopsin. While ELISA kits offer quick results, their selectivity is limited, and they are not congener-specific. Additionally, the quantitative ability of ELISA to distinguish unique variants or congeners of cyanotoxins may vary due to varying cross-reactivities [272,273].
Abraxis Microcystins Kits, Microcystin-ADDA Test Strips, and the Microcystins-DM ELISA Kit are the top three microcystin test kits available today. Microcystin-LR and microcystin-RR-specific antibodies are used in the Abraxis kit’s ELISA to detect microcystins. The strips identify the ADDA moiety present in all types of microcystin. Although they are good for on-site screening, they are not as accurate as the Abraxis ELISA kit. Microcystins-DM from Abraxis employs the ELISA technique as well, but it uses an antibody to detect microcystins by binding to the D-glutamic acid (DM) component of the microcystin molecule. It gives reliable results for the most prevalent microcystin variants but may not pick up on other less prevalent forms [274,275].
Protein Phosphatase Inhibition Assay (PPIA)
Microcystins and nodularins, which are potent inhibitors of protein phosphatase, can be detected using a biochemical Protein Phosphatase Inhibition Assay (PPIA). This assay is a simple, rapid, sensitive, and reproducible colorimetric method [268]. The inhibition of eukaryotic protein phosphatases is a well-established indicator of toxin concentration. There are two methods for measuring this inhibition: radio-isotopic techniques using radiolabeled substrates and colorimetric assays using substrates like p-nitrophenyl phosphate [267,273]. The radio-isotopic method relies on radiolabeled proteins and is not suitable for routine monitoring. In the PPIA, the enzyme is exposed to a sample containing the toxin before incubation with the substrate [276]. By measuring the absorbance of the mixture at a specific wavelength, the substrate (or its transformation product) can be detected, and the enzyme activity can be assessed. The enzyme activity is inversely proportional to the concentration of the toxin. The PPIA method facilitates toxin detection within a few hours and can quantify microcystin-LR with a detection limit of 0.01 μg/L [268].
However, it is important to note that PPIA cannot differentiate between co-occurring variants of microcystins or distinguish microcystins from nodularins. Therefore, the results are often reported as equivalent microcystin-LR/L. Additionally, when analyzing water containing blooms, potential interferences with unknown compounds may lead to overestimation or underestimation of toxin concentration. Furthermore, since PPIA only detects microcystins and nodularins, further analysis is necessary to detect other cyanotoxins that may be present in the sample [277]. Depending on the class of microcystins, the PPIA method exhibits varying sensitivities to different classes of toxins. It provides a measure of relative toxicity, but it cannot identify the specific toxins [267,268,271,277,278]. A commercially available PPIA test in a 96-well microplate format is currently available [279]. Table 1 shows the analytical PPIA methods for the determination of microcystins and Nodularin. The analytical PPIA methods for determining microcystins and nodularin are displayed in Table 1.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
LC-MS, LC-MS/MS, ELISA, and PPIA methods for the analysis of cyanotoxins in environmental samples.
Table 1.
LC-MS, LC-MS/MS, ELISA, and PPIA methods for the analysis of cyanotoxins in environmental samples.
| ID Method | Cyanotoxin | Detection Limit | Sample Preparation | Water Type |
|---|---|---|---|---|
| LC-MS | Anatoxin-a [280] | 0.0021 μg/L | Solid phase extraction disk | Freshwaters |
| Microcystins [261] | 1 μg/g | Solid phase dispersion | Cyanobacteria strain-cultures water | |
| LCMS/MS | Microcystins [281] | 0.002 μg/L | Solid phase extraction | Spiked surfacewater |
| Nodularin [282] | 0.1 μg/L | Solid phase extraction | Lake water | |
| Anatoxin-a [283] | 0.5 μg/L | Freezing and thawing of cells | Surface waters | |
| Cylindrospermopsin [284] | 0.3 μg/L | Solid phase dispersion | Lake waters | |
| Microcystins [285] | <0.02 μg/L | Solid phase dispersion | Surface and Drinking waters | |
| ELISA | Saxitoxin [260] | 0.02 μg/L | Filtration and sonication | Freshwater ecosystems |
| Microcystins [286] | 0.05 μg/L | Solid phase dispersion | River | |
| Nodularin [287] | 0.1 μg/L | Filtration | Surface waters | |
| Microcystin LR in clams [288] | 0.1 ng/mL | Solid phase dispersion | Coastal ponds | |
| Microcystins in mussels [289] | 0.1 μg | Lyophilisation | Estuary | |
| PPIA | Microcystins [290] | 0.2 μg/L | Freeze drying | Lake water |
| Nodularin [291] | Not given | Solid phase dispersion | Lake water | |
| Microcystins [292] | 0.01 μg/L | Filtration | Water supply |
5.1.5. Biosensors and Satellite Imagery
Using a biosensor is the most reliable method for determining whether cyanobacteria and their toxins are in the environment. Specialized devices called biosensors use receptors that bind to specific toxins and send out signals when they are present. These sensors can detect low-concentration contaminants in water samples, enabling early detection and prompt action [293].
Whole-Cell Biosensors
Whole-cell biosensors use entire, living cells as sensing components to find the target analytes’ existence. These biosensors can react to particular cyanobacterial metabolic pathways or molecular markers. By detecting the metabolic activity of the cells and producing an electrical signal, for instance, a biosensor based on immobilized whole-cell cyanobacteria was utilized to determine the presence of cyanobacteria in water samples [293,294].
Antibody-Based Biosensors
Immobilized antibodies are used in antibody-based biosensors to selectively capture and detect particular chemicals or biomolecules linked to cyanobacteria. For instance, phycocyanin was found in water samples using a biosensor based on immobilized antibodies. The biosensor measured the binding of phycocyanin to the immobilized antibodies using a fluorescence-based method, which was detected through variations in the fluorescence signal [295].
Aptamer-Based Biosensors
To specifically bind and detect particular chemicals or biomolecules connected to cyanobacteria, aptamer-based biosensors use synthetic nucleic acid molecules known as aptamers. A biosensor based on immobilized aptamers was employed to find microcystin in water samples. The biosensor measured the binding of microcystin to the immobilized aptamers using a colorimetric assay, which was detected by variations in the color of the test solution [296].
Enzyme-Based Biosensors
Immobilized enzymes are used in enzyme-based biosensors to catalyze particular processes connected to the presence of cyanobacteria cells or their metabolic activity. It can be assumed that a phosphorus-containing substance-based biosensor based on immobilized alkaline phosphatase was employed to identify the molecule’s presence in water samples after cyanobacteria cells discharged it. The biosensor used a colorimetric test to quantify the immobilized enzyme’s activity, which was picked up via alterations in the assay solution’s color [297].
Using satellite imagery as a detection method, authorities can cheaply and effectively monitor broad regions of water thanks to the ability to detect cyanobacteria blooms from space based on changes in the color of the water. To monitor blooms in the future, it is beneficial to pinpoint regions where they have already occurred. The National Oceanic and Atmospheric Administration (NOAA) and the European Space Agency’s Sentinel-3 satellite use satellite photography for cyanobacteria detection and monitoring [298].
The Ocean and Land Color Instrument (OLCI) on the Sentinel-3 satellite can identify the spectral signature of cyanobacteria pigments in water bodies. Scientists may recognize and monitor cyanobacteria blooms’ spatial extent and temporal dynamics by examining the OLCI data. The Baltic Sea experienced a significant bloom of poisonous cyanobacteria in 2019 that was observed by the Sentinel-3 satellite and connected to multiple incidences of respiratory sickness in humans and animals [299].
The Cyanobacteria Monitoring Program of NOAA uses satellite imagery and other data sources to monitor cyanobacteria blooms throughout the United States. The initiative frequently informs resource managers and the general public about the location and severity of cyanobacterial blooms. Numerous beaches and fishing sites had to be closed as a result of a massive cyanobacteria bloom that occurred in the Florida lake Okeechobee in 2018 [300].
5.1.6. Enzyme Inhibition Methods
To find cyanotoxins in the environment, biochemical approaches are frequently used. Enzyme inhibition techniques are one such highly effective strategy for achieving this. By exposing the material to enzymes, this technique determines how much the enzymes are inhibited. Even in complicated water samples, modest quantities of cyanobacterial toxins can be found using enzyme inhibition techniques. This implies it is an excellent instrument for identifying tiny levels of poisons that other techniques might miss. Additionally, it can detect various poisons, including cylindrospermopsin, anatoxins, and microcystins [301,302].
Since cyanobacteria depend on photosynthesis to generate energy, their existence can be detected using photosynthesis inhibitors. Cyanobacteria are found in water samples using a substance known as 3-(3,4-dichlorophenyl)-1,1-dimethylurea (DCMU), a photosynthesis inhibitor. The DCMU stopped the cyanobacteria from producing oxygen, which reduced oxygen production—a cyanobacteria indicator—and was used to ascertain the presence of the cyanobacteria [303]. Others were successful in controlling the growth of bloom-forming M. aeruginosa by employing 2-Hydroxychalcone as a cyanobacterial photosynthesis inhibitor [304].
Since cyanobacteria are known to fix nitrogen, their presence can be determined using inhibitors of nitrogen fixation. The acetylene nitrogen fixation inhibitor can be used to find cyanobacteria in sediments from eutrophic lakes. Because the acetylene reduced the cyanobacteria’s ability to fix nitrogen, less ethylene, a cyanobacterium indicator, was produced [305]. Researchers also used L-phenylalanine, a phosphatase inhibitor, to find cyanobacteria in water samples. Because L-phenylalanine inhibits cyanobacterial phosphatase activity, orthophosphate, a cyanobacteria indicator, was secreted less frequently [306].
5.2. Control of Cyanobacteria and Cyanotoxins
5.2.1. Strategies for Managing the Spread and Control of Cyanobacteria
Fortunately, many tactics may be used to manage the cyanobacteria’s spread and containment.
Biocontrol
One typical biocontrol technique is the introduction of a naturally occurring bacterium that has antagonistic effects on cyanobacteria, either through competition for resources or via the creation of chemicals toxic to cyanobacteria, such as Bacillus spp. and Pseudomonas spp. [307,308,309]. On the other side, cyanophages, a natural virus that feeds on cyanobacteria cells and infects them, can kill cyanobacteria when introduced into infected waters [310]. Other biocontrol strategies involve adding plant extracts or certain aquatic organisms to waters that are already infested [311,312,313]. Several plant extracts, including Salvinia molesta [313,314,315] inhibit cyanobacteria growth. Phytoplanktivorous fishes, including silver carp, big-head carp (Hypophthalmichthys nobilis, previously Aristichys nobilis) and tilapia (Oreochromis niloticus) are direct consumers of phytoplankton (including MC-producing M. aeruginosa) and zooplankton, and thus are widely used in the non-traditional bio-manipulation of cyanobacterial blooms [142]. Aquatic creatures that consume cyanobacteria, like filter-feeding invertebrates and zooplankton Daphnia, can aid in lowering their population. Long-term monitoring of this approach can be challenging and it is not always effective [142,315].
Source Reduction
It is a management strategy that aims to reduce the nutrient inputs that support cyanobacteria growth and, as a result, aids in controlling the spread of cyanobacterial blooms. Reducing fertilizer use (primarily nitrogen and phosphorus) on surrounding agricultural fields, reducing wastewater inputs, and regulating storm water runoff are all examples of nutrient management strategies [316,317]. Additionally, vegetation is crucial in controlling cyanobacterial reproduction because it lowers nutrient inputs and offers shade, reducing the quantity of light available for cyanobacterial growth. The restoration or creation of wetlands, the planting of buffer strips, and the application of riparian zone management techniques are among the vegetation management options used there [318,319].
Septic systems are an essential source of nutrients added to water bodies, particularly in places where residences are built close to bodies of water. Nutrient inputs are reduced and the spread of cyanobacteria is controlled through proper septic system management and maintenance. Additionally, soil degradation increases the amount of nutrients added to water bodies, which might promote the growth of cyanobacteria. Soil erosion and nutrient inputs are reduced by implementing soil conservation practices, such as no-till farming and cover crops [315,318,319].
Algal Turf Scrubbers (ATS)
By eliminating extra nutrients from bodies of water, algal turf scrubbers (ATS) are a sort of device that can be used to control the growth of cyanobacteria blooms [320,321]. ATS is an algae-growing substrate that is a flow-through channel lined with mesh or another material. Pumping contaminated water through the channel causes the algae to absorb nutrients like nitrogen and phosphorus from the water as it passes over the substrate [322]. After being cleaned up, the water is released back into the environment, which can help control the growth of cyanobacterial blooms [322,323]. ATS can offer additional advantages besides nutrient removal, including carbon sequestration, habitat building, and erosion prevention [323,324,325].
5.2.2. Physical and Chemical Control and Removal of Cyanobacterial Blooms
Physical management and chemical removal techniques can be used to control and reduce cyanobacterial blooms. Chemical removal entails utilizing chemical agents to lower the population of the bacteria or the toxins they produce, whereas physical control involves adding mechanical mechanisms to disturb the blooms [326,327].
Physical Control
Cyanobacteria blooms can be physically removed or prevented from spreading using physical control methods. For example, adding oxygen to the water (aeration) can help to disrupt cyanobacteria blooms by encouraging the growth of beneficial bacteria that compete with cyanobacteria for nutrients [328,329]. The water column can be mixed up by aeration, which might lessen the amount of light accessible for cyanobacteria growth. A different approach is to stop their spread by harvesting cyanobacteria from water bodies using nets, screens, or other collecting tools [329,330]. Cyanobacterial blooms can be stopped from spreading to other parts of a body of water by using physical barriers like curtains or booms. In regions where cyanobacteria blooms are concentrated, such as close to a point source of nitrogen inputs, barriers can be beneficial [329,330,331].
Chemical Control
Algicides, copper sulfate, and hydrogen peroxide are a few chemical tools that can be employed to control the growth of cyanobacteria blooms. However, due to potential dangers to the environment and public health, chemical management measures are often only used as a last option [334,335,336,337]. Due to its toxic qualities, copper sulfate mainly kills smaller organisms like cyanobacteria. Still, hydrogen peroxide is frequently employed as a non-toxic substitute for larger organisms or in areas with sensitive species [338,339,340]. Additionally, sodium carbonate peroxyhydrate can be utilized to restrict cyanobacterial growth by removing their ability to photosynthesize by releasing oxygen into the water [339,341].
5.2.3. Cyanobacteria Removal Methods
Cyanobacterial cells and small amounts of toxins were successfully eliminated using conventional portable water treatment techniques such as coagulation, sedimentation, filtration, and chlorination [342,343]. The removal strategy, however, must be carefully chosen during a strong bloom when there are significant amounts of cyanobacterial cells and/or their cyanotoxins in the water because some cyanotoxins can be treated with this strategy, while others cannot [343].
To choose the best course of action for treatment, one must have a thorough understanding of the cyanobacteria species, growth patterns that make up the majority of the bloom, and the characteristics of the cyanotoxins. Cyanotoxins that are intracellular and extracellular (dissolved) can be removed using various techniques. Numerous techniques have been utilized to remove internal cyanotoxins, including coagulation/sedimentation, coagulation-dissolved air flotation (DAF), micro- and ultrafiltration, and pre-treatment oxidation [343,344,345,346,347,348,349]. While some techniques for eliminating extracellular (dissolved) cyanotoxins have been mentioned in the literature, these techniques include membranes (nano- and ultrafiltration), potassium permanganate, ozonation, the use of free chlorine, ultraviolet radiation, physical cyanotoxin adsorption using powdered activated carbon (PAC), granular activated carbon (GAC), and activated carbon adsorption [350,351,352,353,354]. Each method has benefits and is effective, and the efficacy depends on several variables, including chemical, physical, and biological variables [342,349].
6. Summary and Conclusions
The risks linked with cyanobacteria, sometimes referred to as blue-green algae, spreading in bodies of water are discussed in the manuscript. When conditions are ideal, cyanobacteria can spread swiftly. Their growth may be aided by surplus nitrogen and phosphorus from sewage and fertilizer runoff sources. The toxins they produce are toxic to both people and aquatic life.
This review also outlines the characteristics of many cyanobacterial taxa. Oscillatoria, Microcystis, Anabaena, and other organisms are listed. Numerous cyanobacterial species, such as microcystins, nodularin, anatoxin-a, and cylindrospermopsin, can produce toxins that harm the liver and nervous system.
Overabundances of nutrients, warm temperatures, lots of sunlight, and slow water flow all encourage cyanobacteria blooms. Sewage treatment facilities and industrial and agricultural endeavors provide the nutrients required for cyanobacteria to flourish.
Different techniques are discussed for finding and getting rid of cyanobacteria. Cyanobacteria cells and their toxins can be found using microscopy, molecular methods, chromatography, and spectroscopy. Biosensors and immunoassays are also reliable detection techniques. Large bodies of water can be monitored with satellite images.
The manuscript also discussed ways to control and lessen cyanobacteria problems. These techniques include physical or chemical treatments, fertilizer input reduction, the use of algal lawn scrubbers, and biocontrol utilizing antagonistic bacteria. These solutions strive to reduce the dangers that cyanobacterial blooms and their toxins offer. Cyanobacteria in water systems must be managed effectively, which requires early diagnosis and quick action.
Risks associated with abridgment and cyanobacteria dispersion call for thorough monitoring and mitigation measures. Management of this crucial environmental issue can be enhanced by better understanding the elements that encourage blooms and the many detection and eradication options available.
Author Contributions
A.M.T., H.A.N., D.K., S.A.B., L.S. and R.K. wrote and edited the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Ethical review and approval were waived for this review, which does not report new experimental results obtained by authors but contains a summary of already published data.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Bertani, I.; Steger, C.E.; Obenour, D.R.; Fahnenstiel, G.L.; Bridgeman, T.B.; Johengen, T.H.; Sayers, M.J.; Shuchman, R.A.; Scavia, D. Tracking cyanobacteria blooms: Do different monitoring approaches tell the same story? Sci. Total Environ. 2017, 575, 294–308. [Google Scholar] [CrossRef]
- Lürling, M.; Mello, M.M.E.; Van Oosterhout, F.; de Senerpont Domis, L.; Marinho, M.M. Response of natural cyanobacteria and algae assemblages to a nutrient pulse and elevated temperature. Front. Microbiol. 2018, 9, 1851. [Google Scholar] [CrossRef]
- Lyu, T.; Song, L.; Chen, Q.; Pan, G. Lake and river restoration: Method, evaluation and management. Water 2020, 12, 977. [Google Scholar] [CrossRef]
- Metcalf, J.S.; Codd, G.A. Co-occurrence of cyanobacteria and cyanotoxins with other environmental health hazards: Impacts and implications. Toxins 2020, 12, 629. [Google Scholar] [CrossRef]
- Zahra, Z.; Choo, D.H.; Lee, H.; Parveen, A. Cyanobacteria: Review of current potentials and applications. Environments 2020, 7, 13. [Google Scholar] [CrossRef]
- Chorus, I.; Welker, M. Toxic Cyanobacteria in Water: A Guide to Their Public Health Consequences, Monitoring and Management; Taylor & Francis: Abingdon, UK, 2021. [Google Scholar]
- Vidal, L.; Ballot, A.; Azevedo, S.; Padisák, J.; Welker, M. Introduction to cyanobacteria. In Toxic Cyanobacteria in Water, 2nd ed.; Chorus, I., Welker, M., Eds.; CRC Press: Boca Raton, FL, USA, 2021; pp. 163–211. [Google Scholar]
- Moreira, C.; Vasconcelos, V.; Antunes, A. Cyanobacterial blooms: Current knowledge and new perspectives. Earth 2022, 3, 127–135. [Google Scholar] [CrossRef]
- Burford, M.; Carey, C.; Hamilton, D.; Huisman, J.; Paerl, H.; Wood, S.; Wulff, A. Perspective: Advancing the research agenda for improving understanding of cyanobacteria in a future of global change. Harmful Algae 2020, 91, 101601. [Google Scholar] [CrossRef]
- Kultschar, B.; Llewellyn, C. Secondary metabolites in cyanobacteria. Second. Metab. Sources Appl. 2018, 64, 23–27. [Google Scholar] [CrossRef]
- Soule, T.; Garcia-Pichel, F. Cyanobacteria. In Encyclopedia of Microbiology; Elsevier: Amsterdam, The Netherlands, 2019; pp. 799–817. [Google Scholar]
- Molina-Grima, E.; García-Camacho, F.; Acién-Fernández, F.G.; Sánchez-Mirón, A.; Plouviez, M.; Shene, C.; Chisti, Y. Pathogens and predators impacting commercial production of microalgae and cyanobacteria. Biotechnol. Adv. 2022, 55, 107884. [Google Scholar] [CrossRef]
- Patel, V.K.; Sundaram, S.; Patel, A.K.; Kalra, A. Characterization of seven species of cyanobacteria for high-quality biomass production. Arab. J. Sci. Eng. 2018, 43, 109–121. [Google Scholar] [CrossRef]
- Wei, N.; Hu, L.; Song, L.; Gan, N. Microcystin-bound protein patterns in different cultures of Microcystis aeruginosa and field samples. Toxins 2016, 8, 293. [Google Scholar] [CrossRef]
- Huisman, J.; Codd, G.A.; Paerl, H.W.; Ibelings, B.W.; Verspagen, J.M.; Visser, P.M. Cyanobacterial blooms. Nat. Rev. Microbiol. 2018, 16, 471–483. [Google Scholar] [CrossRef]
- Kurmayer, R.; Christiansen, G.; Chorus, I. The abundance of microcystin-producing genotypes correlates positively with colony size in Microcystis sp. and determines its microcystin net production in Lake Wannsee. Appl. Environ. Microbiol. 2003, 69, 787–795. [Google Scholar] [CrossRef]
- Radkova, M.; Stefanova, K.; Uzunov, B.; Gärtner, G.; Stoyneva-Gärtner, M. Morphological and molecular identification of microcystin-producing cyanobacteria in nine shallow Bulgarian water bodies. Toxins 2020, 12, 39. [Google Scholar] [CrossRef]
- Xiao, M.; Li, M.; Reynolds, C.S. Colony formation in the cyanobacterium Microcystis. Biol. Rev. 2018, 93, 1399–1420. [Google Scholar] [CrossRef]
- Jacinavicius, F.R.; Pacheco, A.B.F.; Chow, F.; da Costa, G.C.V.; Kalume, D.E.; Rigonato, J.; Schmidt, E.C.; Sant’Anna, C.L. Different ecophysiological and structural strategies of toxic and non-toxic Microcystis aeruginosa (cyanobacteria) strains assessed under culture conditions. Algal Res. 2019, 41, 101548. [Google Scholar] [CrossRef]
- Dick, G.J.; Duhaime, M.B.; Evans, J.T.; Errera, R.M.; Godwin, C.M.; Kharbush, J.J.; Nitschky, H.S.; Powers, M.A.; Vanderploeg, H.A.; Schmidt, K.C. The genetic and ecophysiological diversity of Microcystis. Environ. Microbiol. 2021, 23, 7278–7313. [Google Scholar] [CrossRef]
- Park, B.S.; Li, Z.; Kang, Y.-H.; Shin, H.H.; Joo, J.-H.; Han, M.-S. Distinct bloom dynamics of toxic and non-toxic Microcystis (cyanobacteria) subpopulations in Hoedong Reservoir (Korea). Microb. Ecol. 2018, 75, 163–173. [Google Scholar] [CrossRef]
- Czarny, K.; Krawczyk, B.; Szczukocki, D. Toxic effects of bisphenol A and its analogues on cyanobacteria Anabaena variabilis and Microcystis aeruginosa. Chemosphere 2021, 263, 128299. [Google Scholar] [CrossRef]
- Österholm, J.; Popin, R.V.; Fewer, D.P.; Sivonen, K. Phylogenomic analysis of secondary metabolism in the toxic cyanobacterial genera Anabaena, Dolichospermum and Aphanizomenon. Toxins 2020, 12, 248. [Google Scholar] [CrossRef]
- Zervou, S.-K.; Moschandreou, K.; Paraskevopoulou, A.; Christophoridis, C.; Grigoriadou, E.; Kaloudis, T.; Triantis, T.M.; Tsiaoussi, V.; Hiskia, A. Cyanobacterial toxins and peptides in Lake Vegoritis, Greece. Toxins 2021, 13, 394. [Google Scholar] [CrossRef] [PubMed]
- Gugger, M.; Lyra, C.; Henriksen, P.; Coute, A.; Humbert, J.-F.; Sivonen, K. Phylogenetic comparison of the cyanobacterial genera Anabaena and Aphanizomenon. Int. J. Syst. Evol. Microbiol. 2002, 52, 1867–1880. [Google Scholar] [PubMed]
- Bramburger, A.J.; Filstrup, C.T.; Reavie, E.D.; Sheik, C.S.; Haffner, G.D.; Depew, D.C.; Downing, J.A. Paradox versus paradigm: A disconnect between understanding and management of freshwater cyanobacterial harmful algae blooms. Freshw. Biol. 2023, 68, 191–201. [Google Scholar] [CrossRef]
- Meriluoto, J.; Sandström, A.; Eriksson, J.; Remaud, G.; Graig, A.G.; Chattopadhyaya, J. Structure and toxicity of a peptide hepatotoxin from the cyanobacterium Oscillatoria agardhii. Toxicon 1989, 27, 1021–1034. [Google Scholar] [CrossRef] [PubMed]
- Eriksson, J.E.; Meriluoto, J.A.; Lindholm, T. Accumulation of a peptide toxin from the cyanobacterium Oscillatoria agardhii in the freshwater mussel Anadonta cygnea. Hydrobiologia 1989, 183, 211–216. [Google Scholar] [CrossRef]
- Bon, I.C.; Salvatierra, L.M.; Lario, L.D.; Morató, J.; Pérez, L.M. Prospects in cadmium-contaminated water management using free-living cyanobacteria (Oscillatoria sp.). Water 2021, 13, 542. [Google Scholar] [CrossRef]
- Ilieva, V.; Kondeva-Burdina, M.; Georgieva, T.; Pavlova, V. Toxicity of cyanobacteria. Organotropy of cyanotoxins and toxicodynamics of cyanotoxins by species. Pharmacia 2019, 66, 91–97. [Google Scholar] [CrossRef]
- Yang, Y.; Chen, Y.; Cai, F.; Liu, X.; Wang, Y.; Li, R. Toxicity-associated changes in the invasive cyanobacterium Cylindrospermopsis raciborskii in response to nitrogen fluctuations. Environ. Pollut. 2018, 237, 1041–1049. [Google Scholar] [CrossRef]
- Willis, A.; Woodhouse, J.N.; Ongley, S.E.; Jex, A.R.; Burford, M.A.; Neilan, B.A. Genome variation in nine co-occurring toxic Cylindrospermopsis raciborskii strains. Harmful Algae 2018, 73, 157–166. [Google Scholar] [CrossRef]
- Rzymski, P.; Horyn, O.; Budzyńska, A.; Jurczak, T.; Kokociński, M.; Niedzielski, P.; Klimaszyk, P.; Falfushynska, H. A report of Cylindrospermopsis raciborskii and other cyanobacteria in the water reservoirs of power plants in Ukraine. Environ. Sci. Pollut. Res. 2018, 25, 15245–15252. [Google Scholar] [CrossRef]
- Santos, G.D.; Vilar, M.C.P.; de Oliveira Azevedo, S.M.F. Acute toxicity of neurotoxin-producing Raphidiopsis (Cylindrospermopsis) raciborskii ITEP-A1 (Cyanobacteria) on the neotropical cladoceran Macrothrix spinosa. Ecotoxicol. Environ. Contam. 2020, 15, 1–8. [Google Scholar] [CrossRef]
- Falfushynska, H.; Horyn, O.; Osypenko, I.; Rzymski, P.; Wejnerowski, Ł.; Dziuba, M.K.; Sokolova, I.M. Multibiomarker-based assessment of toxicity of central European strains of filamentous cyanobacteria Aphanizomenon gracile and Raphidiopsis raciborskii to zebrafish Danio rerio. Water Res. 2021, 194, 116923. [Google Scholar] [CrossRef] [PubMed]
- Jin, H.; Ma, H.; Gan, N.; Wang, H.; Li, Y.; Wang, L.; Song, L. Non-targeted metabolomic profiling of filamentous cyanobacteria Aphanizomenon flos-aquae exposed to a concentrated culture filtrate of Microcystis aeruginosa. Harmful Algae 2022, 111, 102170. [Google Scholar] [CrossRef]
- Wejnerowski, Ł.; Falfushynska, H.; Horyn, O.; Osypenko, I.; Kokociński, M.; Meriluoto, J.; Jurczak, T.; Poniedziałek, B.; Pniewski, F.; Rzymski, P. In Vitro Toxicological screening of stable and senescing cultures of Aphanizomenon, Planktothrix, and Raphidiopsis. Toxins 2020, 12, 400. [Google Scholar] [CrossRef]
- Puschner, B. Cyanobacterial (blue-green algae) toxins. In Veterinary Toxicology; Elsevier: Amsterdam, The Netherlands, 2018; pp. 763–777. [Google Scholar]
- Yancey, C.E.; Mathiesen, O.; Dick, G.J. Transcriptionally active nitrogen fixation and biosynthesis of diverse secondary metabolites by Dolichospermum and Aphanizomenon-like Cyanobacteria in western Lake Erie Microcystis blooms. Harmful Algae 2023, 124, 102408. [Google Scholar] [CrossRef] [PubMed]
- Benayache, N.-Y.; Afri-Mehennaoui, F.-Z.; Kherief-Nacereddine, S.; Vo-Quoc, B.; Hushchyna, K.; Nguyen-Quang, T.; Bouaïcha, N. Massive fish death associated with the toxic cyanobacterial Planktothrix sp. bloom in the Béni-Haroun Reservoir (Algeria). Environ. Sci. Pollut. Res. 2022, 29, 80849–80859. [Google Scholar] [CrossRef]
- Pawlik-Skowrońska, B.; Toporowska, M.; Mazur-Marzec, H. Toxic oligopeptides in the cyanobacterium Planktothrix agardhii-dominated blooms and their effects on duckweed (Lemnaceae) development. Knowl. Manag. Aquat. Ecosyst. 2018, 419, 41. [Google Scholar] [CrossRef]
- Toporowska, M.; Mazur-Marzec, H.; Pawlik-Skowrońska, B. The effects of cyanobacterial bloom extracts on the biomass, Chl-a, MC and other oligopeptides contents in a natural Planktothrix agardhii population. Int. J. Environ. Res. Public Health 2020, 17, 2881. [Google Scholar] [CrossRef]
- Mallia, V.; Ivanova, L.; Eriksen, G.S.; Harper, E.; Connolly, L.; Uhlig, S. Investigation of in vitro endocrine activities of Microcystis and Planktothrix cyanobacterial strains. Toxins 2020, 12, 228. [Google Scholar] [CrossRef]
- Zastepa, A.; Miller, T.R.; Watson, L.C.; Kling, H.; Watson, S.B. Toxins and other bioactive metabolites in deep chlorophyll layers containing the cyanobacteria Planktothrix cf. isothrix in two Georgian Bay Embayments, Lake Huron. Toxins 2021, 13, 445. [Google Scholar] [CrossRef]
- Zhang, Y.; He, D.; Bu, Z.; Li, Y.; Guo, J.; Li, Q. The transcriptomic analysis revealed sulfamethoxazole stress at environmentally relevant concentration on the mechanisms of toxicity of cyanobacteria Synechococcus sp. J. Environ. Chem. Eng. 2022, 10, 107637. [Google Scholar] [CrossRef]
- Te, S.H.; Kok, J.W.K.; Luo, R.; You, L.; Sukarji, N.H.; Goh, K.C.; Sim, Z.Y.; Zhang, D.; He, Y.; Gin, K.Y.-H. Coexistence of Synechococcus and Microcystis Blooms in a Tropical Urban Reservoir and Their Links with Microbiomes. Environ. Sci. Technol. 2023, 57, 1613–1624. [Google Scholar] [CrossRef]
- Zeng, H.; Hu, X.; Ouyang, S.; Zhou, Q. Microplastics Weaken the Adaptability of Cyanobacterium Synechococcus sp. to Ocean Warming. Environ. Sci. Technol. 2023, 57, 9005–9017. [Google Scholar] [CrossRef] [PubMed]
- Sarker, I.; Moore, L.R.; Tetu, S.G. Investigating zinc toxicity responses in marine Prochlorococcus and Synechococcus. Microbiology 2021, 167, 001064. [Google Scholar] [CrossRef] [PubMed]
- Moradinejad, S.; Trigui, H.; Guerra Maldonado, J.F.; Shapiro, J.; Terrat, Y.; Zamyadi, A.; Dorner, S.; Prévost, M. Diversity assessment of toxic cyanobacterial blooms during oxidation. Toxins 2020, 12, 728. [Google Scholar] [CrossRef] [PubMed]
- Breinlinger, S.; Phillips, T.J.; Haram, B.N.; Mareš, J.; Martínez Yerena, J.A.; Hrouzek, P.; Sobotka, R.; Henderson, W.M.; Schmieder, P.; Williams, S.M. Hunting the eagle killer: A cyanobacterial neurotoxin causes vacuolar myelinopathy. Science 2021, 371, eaax9050. [Google Scholar] [CrossRef]
- Metcalf, J.; Souza, N. Cyanobacteria and their toxins. In Separation Science and Technology; Elsevier: Amsterdam, The Netherlands, 2019; Volume 11, pp. 125–148. [Google Scholar]
- Mlewski, E.C.; Pisapia, C.; Gomez, F.; Lecourt, L.; Soto Rueda, E.; Benzerara, K.; Ménez, B.; Borensztajn, S.; Jamme, F.; Réfrégiers, M. Characterization of pustular mats and related rivularia-rich laminations in oncoids from the Laguna Negra lake (Argentina). Front. Microbiol. 2018, 9, 996. [Google Scholar] [CrossRef]
- Nowruzi, B.; Bouaïcha, N.; Metcalf, J.S.; Porzani, S.J.; Konur, O. Plant-cyanobacteria interactions: Beneficial and harmful effects of cyanobacterial bioactive compounds on soil-plant systems and subsequent risk to animal and human health. Phytochemistry 2021, 192, 112959. [Google Scholar] [CrossRef]
- Lyimo, T.J. The Microalgae and Cyanobacteria of Chwaka Bay. In People, Nature and Research in Chwaka Bay, Zanzibar, Tanzania; de la Torre-Castro, M., Lyimo, T.J., Eds.; WIOMSA: Zanzibar Town, Tanzania, 2012; pp. 125–141. ISBN 978-9987-9559-1-6. [Google Scholar]
- Gabyshev, V.; Davydov, D.; Vilnet, A.; Sidelev, S.; Chernova, E.; Barinova, S.; Gabysheva, O.; Zhakovskaya, Z. Gloeotrichia cf. natans (Cyanobacteria) in the Continuous Permafrost Zone of Buotama River, Lena Pillars Nature Park, in Yakutia (Russia). Water 2023, 15, 2370. [Google Scholar] [CrossRef]
- Namsaraev, Z.; Melnikova, A.; Ivanov, V.; Komova, A.; Teslyuk, A. Cyanobacterial Bloom in the World Largest Freshwater Lake Baikal. In IOP Conference Series: Earth and Environmental Science; IOP Publishing: Bristol, UK, 2018; p. 032039. [Google Scholar]
- Kirpenko, N.; Krot, Y.G.; Usenko, O. Toxicological Aspects of the Surface Water” Blooms”(a Review). Hydrobiol. J. 2020, 56, 3–16. [Google Scholar] [CrossRef]
- Genuario, D.B.; Andreote, A.P.D.; Vaz, M.G.M.V.; Fiore, M.F. Heterocyte-forming cyanobacteria from Brazilian saline-alkaline lakes. Mol. Phylogenet. Evol. 2017, 109, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Zhang, J.; He, S.; Yan, X. A review study on macrolides isolated from cyanobacteria. Mar. Drugs 2017, 15, 126. [Google Scholar] [CrossRef] [PubMed]
- Klemm, L.C.; Czerwonka, E.; Hall, M.L.; Williams, P.G.; Mayer, A.M. Cyanobacteria scytonema javanicum and scytonema ocellatum lipopolysaccharides elicit release of superoxide anion, matrix-metalloproteinase-9, cytokines and chemokines by rat microglia in vitro. Toxins 2018, 10, 130. [Google Scholar] [CrossRef]
- Wood, S.A.; Kelly, L.; Bouma-Gregson, K.; Humbert, J.F.; Laughinghouse IV, H.D.; Lazorchak, J.; McAllister, T.; McQueen, A.; Pokrzywinski, K.; Puddick, J. Toxic benthic freshwater cyanobacterial proliferations: Challenges and solutions for enhancing knowledge and improving monitoring and mitigation. Freshw. Biol. 2020, 65, 1824. [Google Scholar] [CrossRef]
- Jaiswal, T.P.; Chakraborty, S.; Sharma, S.; Mishra, A.; Mishra, A.K.; Singh, S.S. Prospects of a hot spring–originated novel cyanobacterium, Scytonema ambikapurensis, for wastewater treatment and exopolysaccharide-enriched biomass production. Environ. Sci. Pollut. Res. 2023, 30, 53424–53444. [Google Scholar] [CrossRef]
- Barzegari Naeeni, R.; Amani, J.; Nobakht, M.; Mirhosseini, S.A.; Mahmoodzadeh Hosseini, H. Human Risk of Saxitoxins Poisoning: Incidence, Source, Toxicity and Diagnostic Methods. J. Mar. Med. 2022, 4, 16–25. [Google Scholar]
- Dulić, T.; Meriluoto, J.; Malešević, T.P.; Gajić, V.; Važić, T.; Tokodi, N.; Obreht, I.; Kostić, B.; Kosijer, P.; Khormali, F. Cyanobacterial diversity and toxicity of biocrusts from the Caspian Lowland loess deposits, North Iran. Quat. Int. 2017, 429, 74–85. [Google Scholar] [CrossRef]
- Velu, C.; Cirés, S.; Brinkman, D.L.; Heimann, K. Effect of CO2 and metal-rich waste water on bioproduct potential of the diazotrophic freshwater cyanobacterium, Tolypothrix sp. Heliyon 2019, 5, e01549. [Google Scholar] [CrossRef]
- Singh, A.K.; Singh, A.; Gaurav, N.; Srivastava, A.; Kumar, A. Morphology & Ecology of Selected BGA (Aulosira, Tolypothrix, Anabaena, Nostoc). Int. J. Gen. Med. Pharm. 2018, 5, 37–46. [Google Scholar]
- Yadav, P.; Gupta, R.K.; Singh, R.P.; Yadav, P.K.; Patel, A.K.; Pandey, K.D. Role of cyanobacteria in green remediation. In Sustainable Environmental Clean-Up; Elsevier: Amsterdam, The Netherlands, 2021; pp. 187–210. [Google Scholar]
- Krings, M.; Harper, C.J. A microfossil resembling Merismopedia (cyanobacteria) from the 410-million-yr-old rhynie and windyfield cherts-Rhyniococcus uniformis revisited. Nova Hedwig. 2019, 108, 17–35. [Google Scholar] [CrossRef]
- Curren, E.; Leong, S.C.Y. Natural and anthropogenic dispersal of cyanobacteria: A review. Hydrobiologia 2020, 847, 2801–2822. [Google Scholar] [CrossRef]
- Poot-Delgado, C.A.; Okolodkov, Y.B.; Aké-Castillo, J.A. Potentially harmful cyanobacteria in oyster banks of Términos lagoon, southeastern Gulf of Mexico. Acta Biol. Colomb. 2018, 23, 51–58. [Google Scholar] [CrossRef]
- Bakr, A. Occurrence of Merismopedia minima in a drinking water treatment plant in Sohag city and removal of their microcystins by sediments. Egypt. J. Bot. 2022, 62, 659–669. [Google Scholar] [CrossRef]
- Youn, S.J.; Kim, H.N.; Yu, S.J.; Byeon, M.S. Cyanobacterial occurrence and geosmin dynamics in Paldang Lake watershed, South Korea. Water Environ. J. 2020, 34, 634–643. [Google Scholar] [CrossRef]
- Häggqvist, K.; Akçaalan, R.; Echenique-Subiabre, I.; Fastner, J.; Horecká, M.; Humbert, J.F.; Izydorczyk, K.; Jurczak, T.; Kokociński, M.; Lindholm, T. Case Studies of Environmental Sampling, Detection, and Monitoring of Potentially Toxic Cyanobacteria. In Handbook of Cyanobacterial Monitoring and Cyanotoxin Analysis; Wiley: Hoboken, NJ, USA, 2016; pp. 70–83. [Google Scholar]
- Volk, A.; Lee, J. Cyanobacterial blooms: A player in the freshwater environmental resistome with public health relevance? Environ. Res. 2022, 216, 114612. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, T.; Umehara, A. Blooming of toxic cyanobacteria in the Isahaya Bay reclaimed land reservoir, South Japan: Present conditions and prospects to resolve the problem. In Environment and Fisheries in the Ariake Sea; Association of Researchers Calling for the Opening the Floodgates in Isahaya Bay; Ariake Sea Network of Fishermen and Citizens, 2017; Volume 4, pp. 19–28. Available online: http://www.ariake-gyomin.net/info/171107aef04.pdf#page=19 (accessed on 13 November 2020).
- Scholz, S.N.; Esterhuizen-Londt, M.; Pflugmacher, S. Rise of toxic cyanobacterial blooms in temperate freshwater lakes: Causes, correlations and possible countermeasures. Toxicol. Environ. Chem. 2017, 99, 543–577. [Google Scholar] [CrossRef]
- Gibbs, M.M.; Roygard, J.; Patterson, M.; Brown, L.; Brown, D. Factors influencing cyanobacteria blooms: Review of the historical monitoring data to assess management options for Lake Horowhenua. N. Z. J. Mar. Freshw. Res. 2022, 1–27. [Google Scholar] [CrossRef]
- Wang, Z.; Akbar, S.; Sun, Y.; Gu, L.; Zhang, L.; Lyu, K.; Huang, Y.; Yang, Z. Cyanobacterial dominance and succession: Factors, mechanisms, predictions, and managements. J. Environ. Manag. 2021, 297, 113281. [Google Scholar] [CrossRef]
- Song, H.; Xu, J.; Lavoie, M.; Fan, X.; Liu, G.; Sun, L.; Fu, Z.; Qian, H. Biological and chemical factors driving the temporal distribution of cyanobacteria and heterotrophic bacteria in a eutrophic lake (West Lake, China). Appl. Microbiol. Biotechnol. 2017, 101, 1685–1696. [Google Scholar] [CrossRef]
- Grasso, C.R.; Pokrzywinski, K.L.; Waechter, C.; Rycroft, T.; Zhang, Y.; Aligata, A.; Kramer, M.; Lamsal, A. A review of cyanophage–host relationships: Highlighting cyanophages as a potential cyanobacteria control strategy. Toxins 2022, 14, 385. [Google Scholar] [CrossRef]
- Fang, F.; Gao, Y.; Gan, L.; He, X.; Yang, L. Effects of different initial pH and irradiance levels on cyanobacterial colonies from Lake Taihu, China. J. Appl. Phycol. 2018, 30, 1777–1793. [Google Scholar] [CrossRef]
- Su, X.; Steinman, A.D.; Tang, X.; Xue, Q.; Zhao, Y.; Xie, L. Response of bacterial communities to cyanobacterial harmful algal blooms in Lake Taihu, China. Harmful Algae 2017, 68, 168–177. [Google Scholar] [CrossRef] [PubMed]
- Plaas, H.E.; Paerl, H.W. Toxic cyanobacteria: A growing threat to water and air quality. Environ. Sci. Technol. 2020, 55, 44–64. [Google Scholar] [CrossRef] [PubMed]
- Codd, G.A.; Testai, E.; Funari, E.; Svirčev, Z. Cyanobacteria, cyanotoxins, and human health. In Water Treatment for Purification from Cyanobacteria and Cyanotoxins; Wiley: Hoboken, NJ, USA, 2020; pp. 37–68. [Google Scholar]
- Ranjbar, M.H.; Hamilton, D.P.; Etemad-Shahidi, A.; Helfer, F. Individual-based modelling of cyanobacteria blooms: Physical and physiological processes. Sci. Total Environ. 2021, 792, 148418. [Google Scholar] [CrossRef] [PubMed]
- Drugă, B.; Buda, D.-M.; Szekeres, E.; Chiş, C.; Chiş, I.; Sicora, C. The impact of cation concentration on Microcystis (cyanobacteria) scum formation. Sci. Rep. 2019, 9, 3017. [Google Scholar] [CrossRef] [PubMed]
- Wood, S.A.; Atalah, J.; Wagenhoff, A.; Brown, L.; Doehring, K.; Young, R.G.; Hawes, I. Effect of river flow, temperature, and water chemistry on proliferations of the benthic anatoxin-producing cyanobacterium Phormidium. Freshw. Sci. 2017, 36, 63–76. [Google Scholar] [CrossRef]
- Seeger, H. Diffusion of Innovations and Public Communication Campaigns: An Examination of the 4R Nutrient Stewardship Program; Illinois State University: Normal, IL, USA, 2018. [Google Scholar]
- Verschoor, M.J. Relationship of Hypolimnetic Iron, Sediment Biogeochemistry, and Sulfate in the Promotion of Cyanobacterial Blooms. 2020. Available online: http://hdl.handle.net/10315/37863 (accessed on 13 November 2020).
- Tan, C.-Y.; Dodd, I.C.; Chen, J.E.; Phang, S.-M.; Chin, C.F.; Yow, Y.-Y.; Ratnayeke, S. Regulation of algal and cyanobacterial auxin production, physiology, and application in agriculture: An overview. J. Appl. Phycol. 2021, 33, 2995–3023. [Google Scholar] [CrossRef]
- DeMarco, J.R. Cyanobacterial Blooms in Chautauqua Lake, NY: Nutrient Sources and Toxin Analyses; Bowling Green State University: Bowling Green, OH, USA, 2021. [Google Scholar]
- Chorus, I.; Fastner, J.; Welker, M. Cyanobacteria and cyanotoxins in a changing environment: Concepts, controversies, challenges. Water 2021, 13, 2463. [Google Scholar] [CrossRef]
- Lee, J.; Lee, S.; Jiang, X. Cyanobacterial toxins in freshwater and food: Important sources of exposure to humans. Annu. Rev. Food Sci. Technol. 2017, 8, 281–304. [Google Scholar] [CrossRef]
- Luan, G.; Lu, X. Tailoring cyanobacterial cell factory for improved industrial properties. Biotechnol. Adv. 2018, 36, 430–442. [Google Scholar] [CrossRef]
- Chittora, D.; Meena, M.; Barupal, T.; Swapnil, P.; Sharma, K. Cyanobacteria as a source of biofertilizers for sustainable agriculture. Biochem. Biophys. Rep. 2020, 22, 100737. [Google Scholar] [CrossRef]
- Schuergers, N.; Mullineaux, C.W.; Wilde, A. Cyanobacteria in motion. Curr. Opin. Plant Biol. 2017, 37, 109–115. [Google Scholar] [CrossRef] [PubMed]
- Qin, B.; Paerl, H.W.; Brookes, J.D.; Liu, J.; Jeppesen, E.; Zhu, G.; Zhang, Y.; Xu, H.; Shi, K.; Deng, J. Why Lake Taihu continues to be plagued with cyanobacterial blooms through 10 years (2007–2017) efforts. Sci. Bull. 2019, 64, 354–356. [Google Scholar] [CrossRef] [PubMed]
- Khalifa, S.A.; Shedid, E.S.; Saied, E.M.; Jassbi, A.R.; Jamebozorgi, F.H.; Rateb, M.E.; Du, M.; Abdel-Daim, M.M.; Kai, G.-Y.; Al-Hammady, M.A. Cyanobacteria—From the oceans to the potential biotechnological and biomedical applications. Mar. Drugs 2021, 19, 241. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Pichel, F.; Zehr, J.P.; Bhattacharya, D.; Pakrasi, H.B. What’s in a name? The case of cyanobacteria. J. Phycol. 2020, 56, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Francés, E.; Escudero-Oñate, C. Cyanobacteria and microalgae in the production of valuable bioactive compounds. Microalgal Biotechnol. 2018, 6, 104–128. [Google Scholar]
- Magonono, M.; Oberholster, P.J.; Shonhai, A.; Makumire, S.; Gumbo, J.R. The presence of toxic and non-toxic cyanobacteria in the sediments of the Limpopo River Basin: Implications for human health. Toxins 2018, 10, 269. [Google Scholar] [CrossRef]
- Cirés, S.; Casero, M.C.; Quesada, A. Toxicity at the edge of life: A review on cyanobacterial toxins from extreme environments. Mar. Drugs 2017, 15, 233. [Google Scholar] [CrossRef]
- Coffer, M.M.; Schaeffer, B.A.; Foreman, K.; Porteous, A.; Loftin, K.A.; Stumpf, R.P.; Werdell, P.J.; Urquhart, E.; Albert, R.J.; Darling, J.A. Assessing cyanobacterial frequency and abundance at surface waters near drinking water intakes across the United States. Water Res. 2021, 201, 117377. [Google Scholar] [CrossRef]
- Kibuye, F.A.; Zamyadi, A.; Wert, E.C. A critical review on operation and performance of source water control strategies for cyanobacterial blooms: Part I-chemical control methods. Harmful Algae 2021, 109, 102099. [Google Scholar] [CrossRef]
- Singh, P.K.; Singh, V.P.; Passarini, M.R.Z.; Kumar, A. Cyanobacterial biology in twenty-first century. Front. Microbiol. 2023, 14, 1184669. [Google Scholar] [CrossRef] [PubMed]
- McKay, C.P. Water sources for cyanobacteria below desert rocks in the Negev Desert determined by conductivity. Glob. Ecol. Conserv. 2016, 6, 145–151. [Google Scholar] [CrossRef]
- Canizales, S.; Sliwszcinka, M.; Russo, A.; Bentvelzen, S.; Temmink, H.; Verschoor, A.; Wijffels, R.H.; Janssen, M. Cyanobacterial growth and cyanophycin production with urea and ammonium as nitrogen source. J. Appl. Phycol. 2021, 33, 3565–3577. [Google Scholar] [CrossRef]
- Ndlela, L.L.; Oberholster, P.J.; Van Wyk, J.; Cheng, P.-H. An overview of cyanobacterial bloom occurrences and research in Africa over the last decade. Harmful Algae 2016, 60, 11–26. [Google Scholar] [CrossRef]
- Erratt, K.J. Urea as an Effective Nitrogen Source for Cyanobacteria; The University of Western Ontario (Canada): London, ON, Canada, 2017. [Google Scholar]
- Leao, T.; Castelão, G.; Korobeynikov, A.; Monroe, E.A.; Podell, S.; Glukhov, E.; Allen, E.E.; Gerwick, W.H.; Gerwick, L. Comparative genomics uncovers the prolific and distinctive metabolic potential of the cyanobacterial genus Moorea. Proc. Natl. Acad. Sci. USA 2017, 114, 3198–3203. [Google Scholar] [CrossRef]
- Singh, A.K.; Singh, P.P.; Tripathi, V.; Verma, H.; Singh, S.K.; Srivastava, A.K.; Kumar, A. Distribution of cyanobacteria and their interactions with pesticides in paddy field: A comprehensive review. J. Environ. Manag. 2018, 224, 361–375. [Google Scholar] [CrossRef]
- Sidelev, S.; Babanazarova, O. Detection of cyanobacterial toxins in water supply sources and tap water in some Russian cities: Searching producers and testing removal methods. Water Resour. 2020, 47, 304–314. [Google Scholar] [CrossRef]
- de la Cruz, A.A.; Chernoff, N.; Sinclair, J.L.; Hill, D.; Diggs, D.L.; Lynch, A.T. Introduction to cyanobacteria and cyanotoxins. In Water Treatment for Purification from Cyanobacteria and Cyanotoxins; Wiley: Hoboken, NJ, USA, 2020; pp. 1–35. [Google Scholar]
- Facey, J.A.; Apte, S.C.; Mitrovic, S.M. A review of the effect of trace metals on freshwater cyanobacterial growth and toxin production. Toxins 2019, 11, 643. [Google Scholar] [CrossRef]
- Chen, M.-Y.; Teng, W.-K.; Zhao, L.; Hu, C.-X.; Zhou, Y.-K.; Han, B.-P.; Song, L.-R.; Shu, W.-S. Comparative genomics reveals insights into cyanobacterial evolution and habitat adaptation. ISME J. 2021, 15, 211–227. [Google Scholar] [CrossRef]
- Graham, J.L.; Dubrovsky, N.M.; Eberts, S.M. Cyanobacterial Harmful Algal Blooms and US Geological Survey Science Capabilities; US Department of the Interior, US Geological Survey: Reston, VA, USA, 2016. [Google Scholar]
- Zhang, W.; Song, X. Synthetic Biology of Cyanobacteria; Springer: Berlin/Heidelberg, Germany, 2018; Volume 1080. [Google Scholar]
- Rodgers, K.J.; Main, B.J.; Samardzic, K. Cyanobacterial neurotoxins: Their occurrence and mechanisms of toxicity. Neurotox. Res. 2018, 33, 168–177. [Google Scholar] [CrossRef]
- Pazouki, P.; Prévost, M.; McQuaid, N.; Barbeau, B.; de Boutray, M.-L.; Zamyadi, A.; Dorner, S. Breakthrough of cyanobacteria in bank filtration. Water Res. 2016, 102, 170–179. [Google Scholar] [CrossRef] [PubMed]
- Trung, B.; Dao, T.-S.; Faassen, E.; Lürling, M. Cyanobacterial blooms and microcystins in Southern Vietnam. Toxins 2018, 10, 471. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Liu, Y.-L.; Conklin, A.; Westrick, J.; Weavers, L.K.; Dionysiou, D.D.; Lenhart, J.J.; Mouser, P.J.; Szlag, D.; Walker, H.W. Toxic cyanobacteria and drinking water: Impacts, detection, and treatment. Harmful Algae 2016, 54, 174–193. [Google Scholar] [CrossRef]
- Kidron, G.J.; Kronenfeld, R.; Tal, S.Y.; Temina, M.; Starinsky, A.; McKay, C.P. The effect of the water source on niche partioning of chlorolichens and cyanobacteria—Implications for resilience? Planta 2023, 258, 8. [Google Scholar] [CrossRef] [PubMed]
- Svirčev, Z.; Dulić, T.; Obreht, I.; Codd, G.A.; Lehmkuhl, F.; Marković, S.B.; Hambach, U.; Meriluoto, J. Cyanobacteria and loess—An underestimated interaction. Plant Soil 2019, 439, 293–308. [Google Scholar] [CrossRef]
- Grattan, L.M.; Holobaugh, S.; Morris, J.G., Jr. Harmful algal blooms and public health. Harmful Algae 2016, 57, 2–8. [Google Scholar] [CrossRef]
- Gobler, C.J.; Burkholder, J.M.; Davis, T.W.; Harke, M.J.; Johengen, T.; Stow, C.A.; Van de Waal, D.B. The dual role of nitrogen supply in controlling the growth and toxicity of cyanobacterial blooms. Harmful Algae 2016, 54, 87–97. [Google Scholar] [CrossRef]
- Massey, I.Y.; Al Osman, M.; Yang, F. An overview on cyanobacterial blooms and toxins production: Their occurrence and influencing factors. Toxin Rev. 2022, 41, 326–346. [Google Scholar] [CrossRef]
- Karlson, B.; Andersen, P.; Arneborg, L.; Cembella, A.; Eikrem, W.; John, U.; West, J.J.; Klemm, K.; Kobos, J.; Lehtinen, S. Harmful algal blooms and their effects in coastal seas of Northern Europe. Harmful Algae 2021, 102, 101989. [Google Scholar] [CrossRef]
- Metcalf, J.; Banack, S.; Wessel, R.; Lester, M.; Pim, J.; Cassani, J.; Cox, P. Toxin analysis of freshwater cyanobacterial and marine harmful algal blooms on the west coast of Florida and implications for estuarine environments. Neurotox. Res. 2021, 39, 27–35. [Google Scholar] [CrossRef]
- Mckay, R.M.L.; Tuttle, T.; Reitz, L.A.; Bullerjahn, G.S.; Cody, W.R.; McDowell, A.J.; Davis, T.W. Early onset of a microcystin-producing cyanobacterial bloom in an agriculturally-influenced Great Lakes tributary. J. Oceanol. Limnol. 2018, 36, 1112–1125. [Google Scholar] [CrossRef]
- Lad, A.; Breidenbach, J.D.; Su, R.C.; Murray, J.; Kuang, R.; Mascarenhas, A.; Najjar, J.; Patel, S.; Hegde, P.; Youssef, M. As we drink and breathe: Adverse health effects of microcystins and other harmful algal bloom toxins in the liver, gut, lungs and beyond. Life 2022, 12, 418. [Google Scholar] [CrossRef] [PubMed]
- Skafi, M.; Duy, S.V.; Munoz, G.; Dinh, Q.T.; Simon, D.F.; Juneau, P.; Sauvé, S. Occurrence of microcystins, anabaenopeptins and other cyanotoxins in fish from a freshwater wildlife reserve impacted by harmful cyanobacterial blooms. Toxicon 2021, 194, 44–52. [Google Scholar] [CrossRef]
- Schaefer, A.M.; Yrastorza, L.; Stockley, N.; Harvey, K.; Harris, N.; Grady, R.; Sullivan, J.; McFarland, M.; Reif, J.S. Exposure to microcystin among coastal residents during a cyanobacteria bloom in Florida. Harmful Algae 2020, 92, 101769. [Google Scholar] [CrossRef] [PubMed]
- Pamplona-Silva, M.T.; Gonçalves, L.C.; Marin-Morales, M.A. Genetic toxicity of water contaminated by microcystins collected during a cyanobacteria bloom. Ecotoxicol. Environ. Saf. 2018, 166, 223–230. [Google Scholar] [CrossRef]
- Kim, M.; Lee, J.; Yang, D.; Park, H.Y.; Park, W. Seasonal dynamics of the bacterial communities associated with cyanobacterial blooms in the Han River. Environ. Pollut. 2020, 266, 115198. [Google Scholar] [CrossRef]
- Nwaji, A.R.; Arieri, O.; Anyang, A.S.; Nguedia, K.; Abiade, E.B.; Forcados, G.E.; Oladipo, O.O.; Makama, S.; Elisha, I.L.; Ozele, N. Natural toxins and one health: A review. Sci. One Health 2022, 1, 100013. [Google Scholar] [CrossRef]
- Chrapusta, E.; Węgrzyn, M.; Zabaglo, K.; Kaminski, A.; Adamski, M.; Wietrzyk, P.; Bialczyk, J. Microcystins and anatoxin-a in Arctic biocrust cyanobacterial communities. Toxicon 2015, 101, 35–40. [Google Scholar] [CrossRef]
- Corbel, S.; Mougin, C.; Bouaïcha, N. Cyanobacterial toxins: Modes of actions, fate in aquatic and soil ecosystems, phytotoxicity and bioaccumulation in agricultural crops. Chemosphere 2014, 96, 1–15. [Google Scholar] [CrossRef]
- Janssen, E.M.-L. Cyanobacterial peptides beyond microcystins–A review on co-occurrence, toxicity, and challenges for risk assessment. Water Res. 2019, 151, 488–499. [Google Scholar] [CrossRef]
- Kosiba, J.; Krztoń, W.; Wilk-Woźniak, E. Effect of microcystins on proto-and metazooplankton is more evident in artificial than in natural waterbodies. Microb. Ecol. 2018, 75, 293–302. [Google Scholar] [CrossRef] [PubMed]
- Palagama, D.S.; Baliu-Rodriguez, D.; Snyder, B.K.; Thornburg, J.A.; Bridgeman, T.B.; Isailovic, D. Identification and quantification of microcystins in western Lake Erie during 2016 and 2017 harmful algal blooms. J. Great Lakes Res. 2020, 46, 289–301. [Google Scholar] [CrossRef]
- Chen, L.; Chen, J.; Zhang, X.; Xie, P. A review of reproductive toxicity of microcystins. J. Hazard. Mater. 2016, 301, 381–399. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Giesy, J.P.; Adamovsky, O.; Svirčev, Z.; Meriluoto, J.; Codd, G.A.; Mijovic, B.; Shi, T.; Tuo, X.; Li, S.C.; et al. Challenges of using blooms of Microcystis spp. in animal feeds: A comprehensive review of nutritional, toxicological and microbial health evaluation. Sci. Total Environ. 2021, 764, 142319. [Google Scholar] [CrossRef] [PubMed]
- Botha, N.; van de Venter, M.; Downing, T.G.; Shephard, E.G.; Gehringer, M.M. The effect of intraperitoneally administered microcystin-LR on the gastrointestinal tract of Balb/c mice. Toxicon 2004, 43, 251–254. [Google Scholar] [CrossRef]
- Su, X.; Steinman, A.D.; Oudsema, M.; Hassett, M.; Xie, L. The influence of nutrients limitation on phytoplankton growth and microcystins production in Spring Lake, USA. Chemosphere 2019, 234, 34–42. [Google Scholar] [CrossRef]
- Carlsson, P.; Rita, D. Sedimentation of Nodularia spumigena and distribution of nodularin in the food web during transport of a cyanobacterial bloom from the Baltic Sea to the Kattegat. Harmful Algae 2019, 86, 74–83. [Google Scholar] [CrossRef]
- Konkel, R.; Toruńska-Sitarz, A.; Cegłowska, M.; Ežerinskis, Ž.; Šapolaitė, J.; Mažeika, J.; Mazur-Marzec, H. Blooms of toxic cyanobacterium Nodularia spumigena in norwegian fjords during Holocene warm periods. Toxins 2020, 12, 257. [Google Scholar] [CrossRef]
- Omara, T.; Nagawa, C.B.; Kyarimpa, C.; Böhmdorfer, S.; Rosenau, T.; Lugasi, S.O.; Matovu, H.; Odongo, S.; Ssebugere, P. Lacustrine Cyanobacteria, Algal Blooms and Cyanotoxins in East Africa: Implications for Human and Ecological Health Protection. Phycology 2023, 3, 147–167. [Google Scholar] [CrossRef]
- Wharton, R.E.; Cunningham, B.R.; Schaefer, A.M.; Guldberg, S.M.; Hamelin, E.I.; Johnson, R.C. Measurement of microcystin and nodularin activity in human urine by immunocapture-protein phosphatase 2A assay. Toxins 2019, 11, 729. [Google Scholar] [CrossRef]
- Mchau, G.J.; Machunda, R.; Kimanya, M.; Makule, E.; Gong, Y.Y.; Mpolya, E.; Meneely, J.P.; Elliott, C.T.; Greer, B. First Report of the co-occurrence of cylindrospermopsin, nodularin and microcystins in the Freshwaters of Lake Victoria, Tanzania. Expo. Health 2021, 13, 185–194. [Google Scholar] [CrossRef]
- Kubickova, B.; Babica, P.; Hilscherová, K.; Šindlerová, L. Effects of cyanobacterial toxins on the human gastrointestinal tract and the mucosal innate immune system. Environ. Sci. Eur. 2019, 31, 1–27. [Google Scholar] [CrossRef]
- Turner, A.D.; Beach, D.G.; Foss, A.; Samdal, I.A.; Løvberg, K.L.; Waack, J.; Edwards, C.; Lawton, L.A.; Dean, K.J.; Maskrey, B.H. A Feasibility Study into the Production of a Mussel Matrix Reference Material for the Cyanobacterial Toxins Microcystins and Nodularins. Toxins 2023, 15, 27. [Google Scholar] [CrossRef] [PubMed]
- Nowruzi, B.; Blanco, S.; Nejadsattari, T. Chemical and molecular evidences for the poisoning of a duck by anatoxin-a, nodularin and cryptophycin at the coast of lake Shoormast (Mazandaran province, Iran). Int. J. Algae 2018, 20, 359–376. [Google Scholar] [CrossRef]
- Chorus, I.; Welker, M. Cyanobacterial toxins. In Toxic Cyanobacteria in Water; CRC Press: Boca Raton, FL, USA, 2021; pp. 13–162. [Google Scholar]
- Chen, G.; Wang, L.; Wang, M.; Hu, T. Comprehensive insights into the occurrence and toxicological issues of nodularins. Mar. Pollut. Bull. 2021, 162, 111884. [Google Scholar] [CrossRef]
- Silveira, S.B.; Odebrecht, C. Effects of salinity and temperature on the growth, toxin production, and akinete germination of the cyanobacterium Nodularia spumigena. Front. Mar. Sci. 2019, 6, 339. [Google Scholar] [CrossRef]
- Schmitz, M.J.; Colombo, G.M.; dos Santos Simião, C.; Ortiz, C.R.; Costa, L.D.F.; da Silva, T.V.N.; Ramos, P.B.; Yunes, J.S.; Wasielesky, W., Jr.; Tesser, M.B. Modulation of nodularin toxicity in shrimp Litopenaeus vannamei (BOONE, 1931) fed with dietary açai (Euterpe oleracea) inclusion. Fish Shellfish Immunol. 2020, 103, 464–471. [Google Scholar] [CrossRef]
- Kaur, S.; Srivastava, A.; Ahluwalia, A.S.; Mishra, Y. Cyanobacterial blooms and Cyanotoxins: Occurrence and Detection. In Algae Multifarious Applications for a Sustainable World; Springer: Singapore, 2021; pp. 339–352. [Google Scholar]
- Mutoti, M.; Gumbo, J.; Jideani, A.I.O. Occurrence of cyanobacteria in water used for food production: A review. Phys. Chem. Earth Parts A/B/C 2022, 125, 103101. [Google Scholar] [CrossRef]
- Hercog, K.; Štern, A.; Maisanaba, S.; Filipič, M.; Žegura, B. Plastics in cyanobacterial blooms—Genotoxic effects of binary mixtures of cylindrospermopsin and bisphenols in HepG2 cells. Toxins 2020, 12, 219. [Google Scholar] [CrossRef]
- Burford, M.A.; Willis, A.; Chuang, A.; Man, X.; Orr, P.T. Recent insights into physiological responses to nutrients by the cylindrospermopsin producing cyanobacterium, Cylindrospermopsis raciborskii. J. Oceanol. Limnol. 2018, 36, 1032–1039. [Google Scholar] [CrossRef]
- Magana-Arachchi, D.N.; Wanigatunge, R.P. Cyanotoxin in Hydrosphere and Human Interface. In One Health: Human, Animal, and Environment Triad; Wiley: Hoboken, NJ, USA, 2023; pp. 75–85. [Google Scholar]
- Yang, Y.; Yu, G.; Chen, Y.; Jia, N.; Li, R. Four decades of progress in cylindrospermopsin research: The ins and outs of a potent cyanotoxin. J. Hazard. Mater. 2021, 406, 124653. [Google Scholar] [CrossRef] [PubMed]
- Kubickova, B.; Laboha, P.; Hildebrandt, J.-P.; Hilscherová, K.; Babica, P. Effects of cylindrospermopsin on cultured immortalized human airway epithelial cells. Chemosphere 2019, 220, 620–628. [Google Scholar] [CrossRef] [PubMed]
- Adamski, M.; Wołowski, K.; Kaminski, A.; Hindáková, A. Cyanotoxin cylindrospermopsin producers and the catalytic decomposition process: A review. Harmful Algae 2020, 98, 101894. [Google Scholar] [CrossRef] [PubMed]
- da Silva, R.d.C.; Grötzner, S.R.; Moura Costa, D.D.; Garcia, J.R.E.; Muelbert, J.; de Magalhães, V.F.; Filipak Neto, F.; de Oliveira Ribeiro, C.A. Comparative bioaccumulation and effects of purified and cellular extract of cylindrospermopsin to freshwater fish Hoplias malabaricus. J. Toxicol. Environ. Health Part A 2018, 81, 620–632. [Google Scholar] [CrossRef]
- Zhang, Y.; Duy, S.V.; Munoz, G.; Sauvé, S. Phytotoxic effects of microcystins, anatoxin-a and cylindrospermopsin to aquatic plants: A meta-analysis. Sci. Total Environ. 2022, 810, 152104. [Google Scholar] [CrossRef]
- Molnár, C.M.; Pînzaru, S.C.; Chis, V.; Feher, I.; Glamuzina, B. SERS of cylindrospermopsin cyanotoxin: Prospects for quantitative analysis in solution and in fish tissue. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2023, 286, 121984. [Google Scholar] [CrossRef]
- Mohamed, Z.A.; Bakr, A. Concentrations of cylindrospermopsin toxin in water and tilapia fish of tropical fishponds in Egypt, and assessing their potential risk to human health. Environ. Sci. Pollut. Res. 2018, 25, 36287–36297. [Google Scholar] [CrossRef]
- Blahova, L.; Sehnal, L.; Lepsova-Skacelova, O.; Szmucova, V.; Babica, P.; Hilscherova, K.; Teikari, J.; Sivonen, K.; Blaha, L. Occurrence of cylindrospermopsin, anatoxin-a and their homologs in the southern Czech Republic–Taxonomical, analytical, and molecular approaches. Harmful Algae 2021, 108, 102101. [Google Scholar] [CrossRef]
- Christensen, V.G.; Khan, E. Freshwater neurotoxins and concerns for human, animal, and ecosystem health: A review of anatoxin-a and saxitoxin. Sci. Total Environ. 2020, 736, 139515. [Google Scholar] [CrossRef]
- Li, Q.; Gu, P.; Zhang, C.; Luo, X.; Zhang, H.; Zhang, J.; Zheng, Z. Combined toxic effects of anatoxin-a and microcystin-LR on submerged macrophytes and biofilms. J. Hazard. Mater. 2020, 389, 122053. [Google Scholar] [CrossRef]
- World Health Organization. Cyanobacterial Toxins: Anatoxin-A and Analogues; World Health Organization: Geneva, Switzerland, 2020. [Google Scholar]
- Falfushynska, H.; Kasianchuk, N.; Siemens, E.; Henao, E.; Rzymski, P. A review of common cyanotoxins and their effects on fish. Toxics 2023, 11, 118. [Google Scholar] [CrossRef] [PubMed]
- Maliaka, V.; Lürling, M.; Fritz, C.; Verstijnen, Y.J.; Faassen, E.J.; Van Oosterhout, F.; Smolders, A.J. Interannual and spatial variability of cyanotoxins in the Prespa lake area, Greece. Water 2021, 13, 357. [Google Scholar] [CrossRef]
- Svirčev, Z.; Lalić, D.; Bojadžija Savić, G.; Tokodi, N.; Drobac Backović, D.; Chen, L.; Meriluoto, J.; Codd, G.A. Global geographical and historical overview of cyanotoxin distribution and cyanobacterial poisonings. Arch. Toxicol. 2019, 93, 2429–2481. [Google Scholar] [CrossRef] [PubMed]
- Lovin, L.M.; Brooks, B.W. Global scanning of anatoxins in aquatic systems: Environment and health hazards, and research needs. Mar. Freshw. Res. 2019, 71, 689–700. [Google Scholar] [CrossRef]
- Foss, A.J.; Butt, J.; Aubel, M.T. Benthic periphyton from Pennsylvania, USA is a source for both hepatotoxins (microcystins/nodularin) and neurotoxins (anatoxin-a/homoanatoxin-a). Toxicon 2018, 150, 13–16. [Google Scholar] [CrossRef] [PubMed]
- Toporowska, M.; Ferencz, B.; Dawidek, J. Effects of Extracts Containing Metabolites of Different Cyanobacteria from an Ambient Spring (Central Europe) on Zooplankters Daphnia magna and Duckweed Spirodela polyrhiza. Water 2022, 14, 4107. [Google Scholar] [CrossRef]
- Painter, K.J.; Venkiteswaran, J.J.; Simon, D.F.; Duy, S.V.; Sauvé, S.; Baulch, H.M. Early and late cyanobacterial bloomers in a shallow, eutrophic lake. Environ. Sci. Process. Impacts 2022, 24, 1212–1227. [Google Scholar] [CrossRef]
- Huang, I.-S.; Zimba, P.V. Cyanobacterial bioactive metabolites—A review of their chemistry and biology. Harmful Algae 2019, 86, 139–209. [Google Scholar] [CrossRef]
- Pradhan, B.; Kim, H.; Abassi, S.; Ki, J.-S. Toxic Effects and Tumor Promotion Activity of Marine Phytoplankton Toxins: A Review. Toxins 2022, 14, 397. [Google Scholar] [CrossRef]
- Shoptaugh, N.H.; Schantz, E.J. Algal Toxins. In CRC Handbook of Foodborne Diseases of Biological Origin; CRC Press: Boca Raton, FL, USA, 2020; pp. 271–275. [Google Scholar]
- Zhang, H.-H.; Zhang, X.-K.; Si, R.-R.; Shen, S.-C.; Liang, T.-T.; Fan, T.-T.; Chen, W.; Xu, L.-H.; Han, B.-N. Chemical and biological study of novel aplysiatoxin derivatives from the marine cyanobacterium Lyngbya sp. Toxins 2020, 12, 733. [Google Scholar] [CrossRef]
- Wani, A.K.; Akhtar, N.; Datta, B.; Pandey, J.; Mannan, M.A.-u. Cyanobacteria-derived small molecules: A new class of drugs. In Volatiles and Metabolites of Microbes; Elsevier: Amsterdam, The Netherlands, 2021; pp. 283–303. [Google Scholar]
- Zahran, E.M.; Albohy, A.; Khalil, A.; Ibrahim, A.H.; Ahmed, H.A.; El-Hossary, E.M.; Bringmann, G.; Abdelmohsen, U.R. Bioactivity potential of marine natural products from Scleractinia-associated microbes and in silico anti-SARS-CoV-2 evaluation. Mar. Drugs 2020, 18, 645. [Google Scholar] [CrossRef] [PubMed]
- Thawabteh, A.M.; Swaileh, Z.; Ammar, M.; Jaghama, W.; Yousef, M.; Karaman, R.; Bufo, S.A.; Scrano, L. Antifungal and antibacterial activities of isolated marine compounds. Toxins 2023, 15, 93. [Google Scholar] [CrossRef] [PubMed]
- Dabas, P.; Tripathi, S.; Chauhan, S.; Malik, I.; Kumar, N.; Kumari, S.; Kumari, R.; Bhatnagar, S. Cyanobacteria—An Unexplored Source for Biomolecules: A Review. Adv. Sci. Eng. Med. 2014, 6, 921–930. [Google Scholar] [CrossRef]
- Singh, R.; Parihar, P.; Singh, M.; Bajguz, A.; Kumar, J.; Singh, S.; Singh, V.P.; Prasad, S.M. Uncovering potential applications of cyanobacteria and algal metabolites in biology, agriculture and medicine: Current status and future prospects. Front. Microbiol. 2017, 8, 515. [Google Scholar] [CrossRef] [PubMed]
- Chong, R.S.-M. Harmful algal blooms. In Aquaculture Pathophysiology; Elsevier: Amsterdam, The Netherlands, 2022; pp. 745–757. [Google Scholar]
- .Spoerke, D.G.; Rumack, B.H. Blue-green algae poisoning. J. Emerg. Med. 1985, 2, 353–355. [Google Scholar] [CrossRef] [PubMed]
- Diehl, F.; Ramos, P.B.; Santos, J.M.d.; Barros, D.M.; Yunes, J.S. Behavioral alterations induced by repeated saxitoxin exposure in drinking water. J. Venom. Anim. Toxins Incl. Trop. Dis. 2016, 22, 18. [Google Scholar] [CrossRef] [PubMed]
- Shaikh, R.; Rizvi, A.; Pandit, S.; Desai, N.; Patil, R. Microalgae: Classification, bioactives, medicinal properties, industrial applications, and future prospectives. In An Integration of Phycoremediation Processes in Wastewater Treatment; Elsevier: Amsterdam, The Netherlands, 2022; pp. 451–486. [Google Scholar]
- Cires, S.; Ballot, A. A review of the phylogeny, ecology and toxin production of bloom-forming Aphanizomenon spp. and related species within the Nostocales (cyanobacteria). Harmful Algae 2016, 54, 21–43. [Google Scholar] [CrossRef] [PubMed]
- Osborne, N.J. Marine Dermatotoxine. In Toxic Cyanobacteria in Water, 2nd ed.; Chorus, I., Welker, M., Eds.; CRC Press: Boca Raton, FL, USA; World Health Organization: Geneva, Switzerland, 2021. [Google Scholar]
- Han, B.-N.; Liang, T.-T.; Keen, L.J.; Fan, T.-T.; Zhang, X.-D.; Xu, L.; Zhao, Q.; Wang, S.-P.; Lin, H.-W. Two marine cyanobacterial aplysiatoxin polyketides, neo-debromoaplysiatoxin A and B, with K+ channel inhibition activity. Org. Lett. 2018, 20, 578–581. [Google Scholar] [CrossRef]
- Tang, Y.-H.; Wu, J.; Fan, T.-T.; Zhang, H.-H.; Gong, X.-X.; Cao, Z.-Y.; Zhang, J.; Lin, H.-W.; Han, B.-N. Chemical and biological study of aplysiatoxin derivatives showing inhibition of potassium channel Kv1.5. RSC Adv. 2019, 9, 7594–7600. [Google Scholar] [CrossRef]
- Puschner, B.; Bautista, A.C.; Wong, C. Debromoaplysiatoxin as the Causative Agent of Dermatitis in a Dog after Exposure to Freshwater in California. Front. Vet. Sci. 2017, 4, 50. [Google Scholar] [CrossRef]
- Ducrot, Y.M.; Thomas, O.P.; Nicolas, M.; Kakue, G.; Desnues, A.; Payri, C.; Bertolotti, A. Toxic seaweed dermatitis in New Caledonia: An epidemiological and clinical study of 83 cases. J. Eur. Acad. Dermatol. Venereol. 2022, 37, e66–e69. [Google Scholar] [CrossRef]
- Aranda, Y.N.; Bhatt, P.; Ates, N.; Engel, B.A.; Simsek, H. Cyanophage-cyanobacterial interactions for sustainable aquatic environment. Environ. Res. 2023, 229, 115728. [Google Scholar] [CrossRef] [PubMed]
- Nowruzi, B.; Porzani, S.J. Toxic compounds produced by cyanobacteria belonging to several species of the order Nostocales: A review. J. Appl. Toxicol. 2021, 41, 510–548. [Google Scholar] [CrossRef] [PubMed]
- Shishido Joutsen, T.K. Cyanobacterial Bioactive Compounds: Biosynthesis, Evolution, Structure and Bioactivity. 2015. Available online: http://hdl.handle.net/10138/154414 (accessed on 13 November 2020).
- Singh, N.K.; Dhar, D.W.; Tabassum, R. Role of cyanobacteria in crop protection. Proc. Natl. Acad. Sci. India Sect. B Biol. Sci. 2016, 86, 1–8. [Google Scholar] [CrossRef]
- Aithal Sadanand, V. The Potential Applications of Cyanobacteria (Blue-Green Algae): A Brief Review. Int. J. Adv. Sci. Res. Manag. 2019, 4, 181–187. [Google Scholar]
- Parwani, L.; Shrivastava, M.; Singh, J. Potential of Cyanobacteria in Wound Healing. In Cyanobacteria-Recent Advances in Taxonomy and Applications; IntechOpen: London, UK, 2021. [Google Scholar]
- Liu, L. New Bioactive Secondary Metabolites from Cyanobacteria. 2014. Available online: http://hdl.handle.net/10138/136091 (accessed on 13 November 2020).
- Soriagalvarro, J.C., Jr. The Identification and Dereplication of Natural Products from Cyanobacteria; University of Illinois at Chicago: Chicago, IL, USA, 2019. [Google Scholar]
- Ananya, A.K.; Ahmad, I.Z. Cyanobacteria “the blue green algae” and its novel applications: A brief review. Int. J. Innov. Appl. Stud. 2014, 7, 251. [Google Scholar]
- Chauhan, A.; Ranjan, A.; Basniwal, R.K.; Jindal, T. Cytotoxic and Antibiotic Properties of Cyanobacterial Extracts. In New Frontiers in Environmental Toxicology; Springer: Cham, Switzerland, 2022; pp. 23–34. [Google Scholar] [CrossRef]
- Singh, S.P.; Pathak, J.; Sinha, R.P. Cyanobacterial factories for the production of green energy and value-added products: An integrated approach for economic viability. Renew. Sustain. Energy Rev. 2017, 69, 578–595. [Google Scholar]
- Vasudevan, S.; Arulmoorthy, M.; Suresh, R. Isolation, purification and structural elucidation of secondary metabolites from Microcystis aeruginosa bloom from Muttukadu estuary and its in vitro antibacterial, antioxidant and anticancer potency. S. Afr. J. Bot. 2020, 132, 59–67. [Google Scholar] [CrossRef]
- Rizwan, M.; Mujtaba, G.; Memon, S.A.; Lee, K.; Rashid, N. Exploring the potential of microalgae for new biotechnology applications and beyond: A review. Renew. Sustain. Energy Rev. 2018, 92, 394–404. [Google Scholar] [CrossRef]
- Sivaramakrishnan, R.; Suresh, S.; Kanwal, S.; Ramadoss, G.; Ramprakash, B.; Incharoensakdi, A. Microalgal biorefinery concepts’ developments for biofuel and bioproducts: Current perspective and bottlenecks. Int. J. Mol. Sci. 2022, 23, 2623. [Google Scholar] [CrossRef]
- Rastogi, R.P.; Madamwar, D.; Incharoensakdi, A. Bloom dynamics of cyanobacteria and their toxins: Environmental health impacts and mitigation strategies. Front. Microbiol. 2015, 6, 1254. [Google Scholar] [CrossRef]
- Bashir, F.; Bashir, A.; Bouaïcha, N.; Chen, L.; Codd, G.A.; Neilan, B.; Xu, W.-L.; Ziko, L.; Rajput, V.D.; Minkina, T. Cyanotoxins, biosynthetic gene clusters, and factors modulating cyanotoxin biosynthesis. World J. Microbiol. Biotechnol. 2023, 39, 241. [Google Scholar] [CrossRef]
- Pearson, L.A.; Dittmann, E.; Mazmouz, R.; Ongley, S.E.; D’Agostino, P.M.; Neilan, B.A. The genetics, biosynthesis and regulation of toxic specialized metabolites of cyanobacteria. Harmful Algae 2016, 54, 98–111. [Google Scholar] [CrossRef]
- Paerl, H.W. Mitigating harmful cyanobacterial blooms in a human-and climatically-impacted world. Life 2014, 4, 988–1012. [Google Scholar] [CrossRef] [PubMed]
- Paerl, H.W.; Barnard, M.A. Mitigating the global expansion of harmful cyanobacterial blooms: Moving targets in a human-and climatically-altered world. Harmful Algae 2020, 96, 101845. [Google Scholar] [CrossRef] [PubMed]
- Burford, M.; Gobler, C.; Hamilton, D.; Visser, P.; Lurling, M.; Codd, G. Solutions for Managing Cyanobacterial Blooms: A Scientific Summary for Policy Makers; IOC/UNESCO: Paris, France, 2019; 16p, IOC/INF-1382. [Google Scholar] [CrossRef]
- Weber, S.J.; Mishra, D.R.; Wilde, S.B.; Kramer, E. Risks for cyanobacterial harmful algal blooms due to land management and climate interactions. Sci. Total Environ. 2020, 703, 134608. [Google Scholar] [CrossRef]
- Moreira, C.; Ramos, V.; Azevedo, J.; Vasconcelos, V. Methods to detect cyanobacteria and their toxins in the environment. Appl. Microbiol. Biotechnol. 2014, 98, 8073–8082. [Google Scholar] [CrossRef] [PubMed]
- Roy-Lachapelle, A.; Solliec, M.; Bouchard, M.F.; Sauvé, S. Detection of cyanotoxins in algae dietary supplements. Toxins 2017, 9, 76. [Google Scholar] [CrossRef]
- Makhalanyane, T.P.; Valverde, A.; Velázquez, D.; Gunnigle, E.; Van Goethem, M.W.; Quesada, A.; Cowan, D.A. Ecology and biogeochemistry of cyanobacteria in soils, permafrost, aquatic and cryptic polar habitats. Biodivers. Conserv. 2015, 24, 819–840. [Google Scholar] [CrossRef]
- Sinha, A.K.; Eggleton, M.A.; Lochmann, R.T. An environmentally friendly approach for mitigating cyanobacterial bloom and their toxins in hypereutrophic ponds: Potentiality of a newly developed granular hydrogen peroxide-based compound. Sci. Total Environ. 2018, 637, 524–537. [Google Scholar] [CrossRef]
- Drikas, M.; Chow, C.W.; House, J.; Burch, M.D. Using coagulation, flocculation, and settling to remove toxic cyanobacteria. J. Am. Water Work. Assoc. 2001, 93, 100–111. [Google Scholar] [CrossRef]
- Kaushik, R.; Balasubramanian, R. Methods and approaches used for detection of cyanotoxins in environmental samples: A review. Crit. Rev. Environ. Sci. Technol. 2013, 43, 1349–1383. [Google Scholar] [CrossRef]
- Zamyadi, A.; MacLeod, S.L.; Fan, Y.; McQuaid, N.; Dorner, S.; Sauvé, S.; Prévost, M. Toxic cyanobacterial breakthrough and accumulation in a drinking water plant: A monitoring and treatment challenge. Water Res. 2012, 46, 1511–1523. [Google Scholar] [CrossRef] [PubMed]
- Sundaravadivelu, D.; Sanan, T.T.; Venkatapathy, R.; Mash, H.; Tettenhorst, D.; DAnglada, L.; Frey, S.; Tatters, A.O.; Lazorchak, J. Determination of Cyanotoxins and Prymnesins in Water, Fish Tissue, and Other Matrices: A Review. Toxins 2022, 14, 213. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Zhang, S.; Yu, T.; Dai, Z.; Wei, Q. Aptamer-based fluorescent sensor array for multiplexed detection of cyanotoxins on a smartphone. Anal. Chem. 2019, 91, 10448–10457. [Google Scholar] [CrossRef]
- Greer, B.; Maul, R.; Campbell, K.; Elliott, C.T. Detection of freshwater cyanotoxins and measurement of masked microcystins in tilapia from Southeast Asian aquaculture farms. Anal. Bioanal. Chem. 2017, 409, 4057–4069. [Google Scholar] [CrossRef]
- Du, X.; Liu, H.; Yuan, L.; Wang, Y.; Ma, Y.; Wang, R.; Chen, X.; Losiewicz, M.D.; Guo, H.; Zhang, H. The diversity of cyanobacterial toxins on structural characterization, distribution and identification: A systematic review. Toxins 2019, 11, 530. [Google Scholar] [CrossRef]
- Gaget, V.; Lau, M.; Sendall, B.; Froscio, S.; Humpage, A.R. Cyanotoxins: Which detection technique for an optimum risk assessment? Water Res. 2017, 118, 227–238. [Google Scholar] [CrossRef]
- Graham, J.L.; Dubrovsky, N.M.; Foster, G.M.; King, L.R.; Loftin, K.A.; Rosen, B.H.; Stelzer, E.A. Cyanotoxin occurrence in large rivers of the United States. Inland Waters 2020, 10, 109–117. [Google Scholar] [CrossRef]
- Moreira, C.; Gomes, C.; Vasconcelos, V.; Antunes, A. Cyanotoxins occurrence in Portugal: A new report on their recent multiplication. Toxins 2020, 12, 154. [Google Scholar] [CrossRef]
- Westrick, J.A.; Szlag, D. A cyanotoxin primer for drinking water professionals. J. Am. Water Work. Assoc. 2018, 110, E1–E16. [Google Scholar] [CrossRef]
- Facciponte, D.N.; Bough, M.W.; Seidler, D.; Carroll, J.L.; Ashare, A.; Andrew, A.S.; Tsongalis, G.J.; Vaickus, L.J.; Henegan, P.L.; Butt, T.H. Identifying aerosolized cyanobacteria in the human respiratory tract: A proposed mechanism for cyanotoxin-associated diseases. Sci. Total Environ. 2018, 645, 1003–1013. [Google Scholar] [CrossRef] [PubMed]
- Almuhtaram, H.; Cui, Y.; Zamyadi, A.; Hofmann, R. Cyanotoxins and cyanobacteria cell accumulations in drinking water treatment plants with a low risk of bloom formation at the source. Toxins 2018, 10, 430. [Google Scholar] [CrossRef]
- Gregor, J.; Maršálek, B.; Šípková, H. Detection and estimation of potentially toxic cyanobacteria in raw water at the drinking water treatment plant by in vivo fluorescence method. Water Res. 2007, 41, 228–234. [Google Scholar] [CrossRef] [PubMed]
- Aguilera, A.; Klemenčič, M.; Sueldo, D.J.; Rzymski, P.; Giannuzzi, L.; Martin, M.V. Cell death in cyanobacteria: Current understanding and recommendations for a consensus on its nomenclature. Front. Microbiol. 2021, 12, 631654. [Google Scholar] [CrossRef] [PubMed]
- Nagayama, K.; Danev, R. Phase contrast electron microscopy: Development of thin-film phase plates and biological applications. Philos. Trans. R. Soc. B Biol. Sci. 2008, 363, 2153–2162. [Google Scholar] [CrossRef]
- Karthikeyan, N.; Prasanna, R.; Sood, A.; Jaiswal, P.; Nayak, S.; Kaushik, B. Physiological characterization and electron microscopic investigation of cyanobacteria associated with wheat rhizosphere. Folia Microbiol. 2009, 54, 43–51. [Google Scholar] [CrossRef]
- Donghao, W.; Fangfei, C.; Jiaxin, H.; Guanning, J.; YaDong, S.; Aichun, S. The declining cyanobacterial blooms in Lake Taihu (China) in 2021: The interplay of nutrients and meteorological determinants. Ecol. Indic. 2022, 145, 109590. [Google Scholar] [CrossRef]
- Barros, M.U.; Wilson, A.E.; Leitão, J.I.; Pereira, S.P.; Buley, R.P.; Fernandez-Figueroa, E.G.; Capelo-Neto, J. Environmental factors associated with toxic cyanobacterial blooms across 20 drinking water reservoirs in a semi-arid region of Brazil. Harmful Algae 2019, 86, 128–137. [Google Scholar] [CrossRef]
- Teta, R.; Esposito, G.; De Sterlich, C.; Lega, M.; Costantino, V. Early detection of cyanobacterial blooms and associated cyanotoxins using fast detection strategy. JoVE 2021, 168, e61889. [Google Scholar]
- Voráčová, K.; Paichlová, J.; Vicková, K.; Hrouzek, P. Screening of cyanobacterial extracts for apoptotic inducers: A combined approach of caspase-3/7 homogeneous assay and time-lapse microscopy. J. Appl. Phycol. 2017, 29, 1933–1943. [Google Scholar] [CrossRef]
- Pierella Karlusich, J.J.; Pelletier, E.; Lombard, F.; Carsique, M.; Dvorak, E.; Colin, S.; Picheral, M.; Cornejo-Castillo, F.M.; Acinas, S.G.; Pepperkok, R. Global distribution patterns of marine nitrogen-fixers by imaging and molecular methods. Nat. Commun. 2021, 12, 4160. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Lou, I.; Ung, W.K.; Kong, Y.; Mok, K.M. Application of PCR and real-time PCR for monitoring cyanobacteria, Microcystis spp. and Cylindrospermopsis raciborskii in Macau freshwater reservoir. Front. Earth Sci. 2014, 8, 291–301. [Google Scholar] [CrossRef]
- Martinez De La Escalera, G.; Antoniades, D.; Bonilla, S.; Piccini, C. Application of ancient DNA to the reconstruction of past microbial assemblages and for the detection of toxic cyanobacteria in subtropical freshwater ecosystems. Mol. Ecol. 2014, 23, 5791–5802. [Google Scholar] [CrossRef] [PubMed]
- Orr, P.T.; Rasmussen, J.P.; Burford, M.A.; Eaglesham, G.K.; Lennox, S.M. Evaluation of quantitative real-time PCR to characterise spatial and temporal variations in cyanobacteria, Cylindrospermopsis raciborskii (Woloszynska) Seenaya et Subba Raju and cylindrospermopsin concentrations in three subtropical Australian reservoirs. Harmful Algae 2010, 9, 243–254. [Google Scholar] [CrossRef]
- Tomioka, N.; Nagai, T.; Kawasaki, T.; Imai, A.; Matsushige, K.; Kohata, K. Quantification of Microcystis in a eutrophic lake by simple DNA extraction and SYBR Green real-time PCR. Microbes Environ. 2008, 23, 306–312. [Google Scholar] [CrossRef]
- Vieira-Lanero, R.; Barca, S.; Cobo, M.C.; Cobo, F. Occurrence of freshwater cyanobacteria and bloom Records in Spanish Reservoirs (1981–2017). Hydrobiology 2022, 1, 122–136. [Google Scholar] [CrossRef]
- Lu, K.-Y.; Chiu, Y.-T.; Burch, M.; Senoro, D.; Lin, T.-F. A molecular-based method to estimate the risk associated with cyanotoxins and odor compounds in drinking water sources. Water Res. 2019, 164, 114938. [Google Scholar] [CrossRef]
- Devi, A.; Chiu, Y.-T.; Hsueh, H.-T.; Lin, T.-F. Quantitative PCR based detection system for cyanobacterial geosmin/2-methylisoborneol (2-MIB) events in drinking water sources: Current status and challenges. Water Res. 2021, 188, 116478. [Google Scholar] [CrossRef]
- Jehlička, J.; Edwards, H.G.; Oren, A. Raman spectroscopy of microbial pigments. Appl. Environ. Microbiol. 2014, 80, 3286–3295. [Google Scholar] [CrossRef]
- Kleinteich, J.; Puddick, J.; Wood, S.A.; Hildebrand, F.; Laughinghouse IV, H.D.; Pearce, D.A.; Dietrich, D.R.; Wilmotte, A. Toxic cyanobacteria in Svalbard: Chemical diversity of microcystins detected using a liquid chromatography mass spectrometry precursor ion screening method. Toxins 2018, 10, 147. [Google Scholar] [CrossRef] [PubMed]
- Parker, C.H.; Stutts, W.L.; DeGrasse, S.L. Development and validation of a liquid chromatography-tandem mass spectrometry method for the quantitation of microcystins in blue-green algal dietary supplements. J. Agric. Food Chem. 2015, 63, 10303–10312. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.; Shan, K.; Lin, L.; Shen, W.; Huang, L.; Gan, N.; Song, L. Multi-year assessment of toxic genotypes and microcystin concentration in northern Lake Taihu, China. Toxins 2016, 8, 23. [Google Scholar] [CrossRef]
- Grigoryeva, N.; Chistyakova, L. Fluorescence microscopic spectroscopy for investigation and monitoring of biological diversity and physiological state of cyanobacterial cultures. In Cyanobacteria; IntechOpen: Rijeka, Croatia, 2018; pp. 11–44. [Google Scholar]
- Pinzaru, S.C.; Müller, C.; Brezestean, I.; Barchewits, D.; Glamuzina, B. Cyanobacteria detection and Raman spectroscopy characterization with a highly sensitive, high resolution fiber optic portable Raman system. Stud. Univ. Babes Bolyai Phys. 2016, 61, 99–108. [Google Scholar]
- Oehrle, S.A.; Southwell, B.; Westrick, J. Detection of various freshwater cyanobacterial toxins using ultra-performance liquid chromatography tandem mass spectrometry. Toxicon 2010, 55, 965–972. [Google Scholar] [CrossRef] [PubMed]
- Kleinteich, J.; Wood, S.A.; Puddick, J.; Schleheck, D.; Küpper, F.C.; Dietrich, D. Potent toxins in Arctic environments–presence of saxitoxins and an unusual microcystin variant in Arctic freshwater ecosystems. Chem. Biol. Interact. 2013, 206, 423–431. [Google Scholar] [CrossRef]
- Camean, A.; Moreno, I.M.; Ruiz, M.J.; Pico, Y. Determination of microcystins in natural blooms and cyanobacterial strain cultures by matrix solid-phase dispersion and liquid chromatography–mass spectrometry. Anal. Bioanal. Chem. 2004, 380, 537–544. [Google Scholar] [CrossRef]
- Qin, Y.; Zhong, Q.; Zhang, Y.; Lin, X.; Fu, P.; Lin, H. Micro-flow hydrophilic interaction liquid chromatography coupled with triple quadrupole mass spectrometry detects modified nucleosides in the transfer RNA pool of cyanobacteria. J. Sep. Sci. 2021, 44, 3208–3218. [Google Scholar] [CrossRef]
- Fayad, P.B.; Roy-Lachapelle, A.; Duy, S.V.; Prévost, M.; Sauvé, S. On-line solid-phase extraction coupled to liquid chromatography tandem mass spectrometry for the analysis of cyanotoxins in algal blooms. Toxicon 2015, 108, 167–175. [Google Scholar] [CrossRef]
- Zastepa, A.; Pick, F.R.; Blais, J.M.; Saleem, A. Analysis of intracellular and extracellular microcystin variants in sediments and pore waters by accelerated solvent extraction and high performance liquid chromatography-tandem mass spectrometry. Anal. Chim. Acta 2015, 872, 26–34. [Google Scholar] [CrossRef]
- Rodriguez, I.; Alfonso, C.; Alfonso, A.; Otero, P.; Meyer, T.; Breitenbach, U.; Botana, L. Toxin profile in samples collected in fresh and brackish water in Germany. Toxicon 2014, 91, 35–44. [Google Scholar] [CrossRef]
- Bogialli, S.; Bruno, M.; Curini, R.; Di Corcia, A.; Laganà, A. Simple and rapid determination of anatoxin-a in lake water and fish muscle tissue by liquid-chromatography–tandem mass spectrometry. J. Chromatogr. A 2006, 1122, 180–185. [Google Scholar] [CrossRef]
- Furey, A.; Crowley, J.; Lehane, M.; James, K.J. Liquid chromatography with electrospray ion-trap mass spectrometry for the determination of anatoxins in cyanobacteria and drinking water. Rapid Commun. Mass Spectrom. 2003, 17, 583–588. [Google Scholar] [CrossRef]
- Dell’Aversano, C.; Eaglesham, G.K.; Quilliam, M.A. Analysis of cyanobacterial toxins by hydrophilic interaction liquid chromatography–mass spectrometry. J. Chromatogr. A 2004, 1028, 155–164. [Google Scholar] [CrossRef] [PubMed]
- Koreivienė, J.; Belous, O. Methods for cyanotoxins detection. Bot. Lith. 2012, 18, 58–65. [Google Scholar] [CrossRef]
- Lawton, L.A.; Edwards, C. Conventional laboratory methods for cyanotoxins. In Cyanobacterial Harmful Algal Blooms: State of the Science and Research Needs; Springer: Berlin/Heidelberg, Germany, 2008; pp. 513–537. [Google Scholar]
- Vallod, D.; Cravedi, J.; Hillenweck, A.; Robin, J. Analysis of the off-flavor risk in carp production in ponds in Dombes and Forez (France). Aquac. Int. 2007, 15, 287–298. [Google Scholar] [CrossRef]
- Metcalf, J.S.; Codd, G.A. Immunoassays and Other Antibody Applications. In Handbook of Cyanobacterial Monitoring and Cyanotoxin Analysis; Wiley: Hoboken, NJ, USA, 2016; pp. 263–266. [Google Scholar]
- Sheng, J.-W.; He, M.; Shi, H.-C.; Qian, Y. A comprehensive immunoassay for the detection of microcystins in waters based on polyclonal antibodies. Anal. Chim. Acta 2006, 572, 309–315. [Google Scholar] [CrossRef] [PubMed]
- Kitsiou, D.; Karydis, M. Coastal marine eutrophication assessment: A review on data analysis. Environ. Int. 2011, 37, 778–801. [Google Scholar] [CrossRef] [PubMed]
- Wagner, C.; Adrian, R. Cyanobacteria dominance: Quantifying the effects of climate change. Limnol. Oceanogr. 2009, 54, 2460–2468. [Google Scholar] [CrossRef]
- Srivastava, A.; Singh, S.; Ahn, C.-Y.; Oh, H.-M.; Asthana, R.K. Monitoring approaches for a toxic cyanobacterial bloom. Environ. Sci. Technol. 2013, 47, 8999–9013. [Google Scholar] [CrossRef]
- Merel, S.; Walker, D.; Chicana, R.; Snyder, S.; Baurès, E.; Thomas, O. State of knowledge and concerns on cyanobacterial blooms and cyanotoxins. Environ. Int. 2013, 59, 303–327. [Google Scholar] [CrossRef] [PubMed]
- Mountfort, D.O.; Holland, P.; Sprosen, J. Method for detecting classes of microcystins by combination of protein phosphatase inhibition assay and ELISA: Comparison with LC-MS. Toxicon 2005, 45, 199–206. [Google Scholar] [CrossRef] [PubMed]
- Kaloudis, T.; Zervou, S.-K.; Tsimeli, K.; Triantis, T.M.; Fotiou, T.; Hiskia, A. Determination of microcystins and nodularin (cyanobacterial toxins) in water by LC–MS/MS. Monitoring of Lake Marathonas, a water reservoir of Athens, Greece. J. Hazard. Mater. 2013, 263, 105–115. [Google Scholar] [CrossRef]
- Takino, M.; Daishima, S.; Yamaguchi, K. Analysis of anatoxin-a in freshwaters by automated on-line derivatization–liquid chromatography–electrospray mass spectrometry. J. Chromatogr. A 1999, 862, 191–197. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Ping, X.; Yang, Z. Determination of microcystin-LR in surface water using high-performance liquid chromatography/tandem electrospray ionization mass detector. Talanta 2004, 62, 191–198. [Google Scholar] [CrossRef]
- Ortea, P.M.; Allis, O.; Healy, B.M.; Lehane, M.; Shuilleabháin, A.N.; Furey, A.; James, K.J. Determination of toxic cyclic heptapeptides by liquid chromatography with detection using ultra-violet, protein phosphatase assay and tandem mass spectrometry. Chemosphere 2004, 55, 1395–1402. [Google Scholar] [CrossRef]
- Hedman, C.J.; Krick, W.R.; Karner Perkins, D.A.; Harrahy, E.A.; Sonzogni, W.C. New measurements of cyanobacterial toxins in natural waters using high performance liquid chromatography coupled to tandem mass spectrometry. J. Environ. Qual. 2008, 37, 1817–1824. [Google Scholar] [CrossRef]
- Messineo, V.; Bogialli, S.; Melchiorre, S.; Sechi, N.; Lugliè, A.; Casiddu, P.; Mariani, M.A.; Padedda, B.M.; Di Corcia, A.; Mazza, R. Cyanobacterial toxins in Italian freshwaters. Limnologica 2009, 39, 95–106. [Google Scholar] [CrossRef]
- Triantis, T.; Tsimeli, K.; Kaloudis, T.; Thanassoulias, N.; Lytras, E.; Hiskia, A. Development of an integrated laboratory system for the monitoring of cyanotoxins in surface and drinking waters. Toxicon 2010, 55, 979–989. [Google Scholar] [CrossRef]
- Dixit, R.B.; Patel, A.K.; Toppo, K.; Nayaka, S. Emergence of toxic cyanobacterial species in the Ganga River, India, due to excessive nutrient loading. Ecol. Indic. 2017, 72, 420–427. [Google Scholar] [CrossRef]
- Sedda, T.; Baralla, E.; Varoni, M.V.; Pasciu, V.; Lorenzoni, G.; Demontis, M.P. Determination of microcystin-LR in clams (Tapes decussatus) of two Sardinian coastal ponds (Italy). Mar. Pollut. Bull. 2016, 108, 317–320. [Google Scholar] [CrossRef]
- Preece, E.; Moore, B.C.; Hardy, F.J. Transfer of microcystin from freshwater lakes to Puget Sound, WA and toxin accumulation in marine mussels (Mytilus trossulus). Ecotoxicol. Environ. Saf. 2015, 122, 98–105. [Google Scholar] [CrossRef] [PubMed]
- Heresztyn, T.; Nicholson, B.C. Determination of cyanobacterial hepatotoxins directly in water using a protein phosphatase inhibition assay. Water Res. 2001, 35, 3049–3056. [Google Scholar] [CrossRef] [PubMed]
- Almeida, V.P.; Cogo, K.; Tsai, S.M.; Moon, D.H. Colorimetric test for the monitoring of microcystins in cyanobacterial culture and environmental samples from southeast-Brazil. Braz. J. Microbiol. 2006, 37, 192–198. [Google Scholar] [CrossRef]
- John, N.; Baker, L.; Ansell, B.R.; Newham, S.; Crosbie, N.D.; Jex, A.R. First report of anatoxin-a producing cyanobacteria in Australia illustrates need to regularly up-date monitoring strategies in a shifting global distribution. Sci. Rep. 2019, 9, 10894. [Google Scholar] [CrossRef]
- Pescheck, M.; Schweizer, A.; Bláha, L. Innovative electrochemical biosensor for toxicological investigations on algae and cyanobacteria. Bioelectrochemistry 2022, 143, 107926. [Google Scholar] [CrossRef]
- Salouti, M.; Khadivi Derakhshan, F. Biosensors and nanobiosensors in environmental applications. In Biogenic Nano-Particles and Their Use in Agro-Ecosystems; Springer: Singapore, 2020; pp. 515–591. [Google Scholar]
- Hosseini, S.; Espinosa-Hernandez, M.A.; Garcia-Ramirez, R.; Cerda-Kipper, A.S.; Reveles-Huizar, S.; Acosta-Soto, L.; Acosta-Soto, L.; Hosseini, S. Bio-microelectromechanical Systems (BioMEMS) in Bio-sensing Applications-Fluorescence Detection Strategies. In BioMEMS: Biosensing Applications; Springer: Singapore, 2021; pp. 69–97. [Google Scholar]
- Kumar, P.; Rautela, A.; Kesari, V.; Szlag, D.; Westrick, J.; Kumar, S. Recent developments in the methods of quantitative analysis of microcystins. J. Biochem. Mol. Toxicol. 2020, 34, e22582. [Google Scholar] [CrossRef] [PubMed]
- Devic, E.; Li, D.; Dauta, A.; Henriksen, P.; Codd, G.A.; Marty, J.-L.; Fournier, D. Detection of anatoxin-a (s) in environmental samples of cyanobacteria by using a biosensor with engineered acetylcholinesterases. Appl. Environ. Microbiol. 2002, 68, 4102–4106. [Google Scholar] [CrossRef] [PubMed]
- Vander Woude, A.; Ruberg, S.; Johengen, T.; Miller, R.; Stuart, D. Spatial and temporal scales of variability of cyanobacteria harmful algal blooms from NOAA GLERL airborne hyperspectral imagery. J. Great Lakes Res. 2019, 45, 536–546. [Google Scholar] [CrossRef]
- Mishra, S.; Stumpf, R.P.; Schaeffer, B.A.; Werdell, P.J.; Loftin, K.A.; Meredith, A. Measurement of cyanobacterial bloom magnitude using satellite remote sensing. Sci. Rep. 2019, 9, 18310. [Google Scholar] [CrossRef]
- Kramer, B.J.; Davis, T.W.; Meyer, K.A.; Rosen, B.H.; Goleski, J.A.; Dick, G.J.; Oh, G.; Gobler, C.J. Nitrogen limitation, toxin synthesis potential, and toxicity of cyanobacterial populations in Lake Okeechobee and the St. Lucie River Estuary, Florida, during the 2016 state of emergency event. PLoS ONE 2018, 13, e0196278. [Google Scholar] [CrossRef]
- Bucur, B.; Munteanu, F.-D.; Marty, J.-L.; Vasilescu, A. Advances in enzyme-based biosensors for pesticide detection. Biosensors 2018, 8, 27. [Google Scholar] [CrossRef]
- Chen, Y.; Huang, X.; Wang, J.; Li, C. Effect of pure microcystin-LR on activity and transcript level of immune-related enzymes in the white shrimp (Litopenaeus vannamei). Ecotoxicology 2017, 26, 702–710. [Google Scholar] [CrossRef]
- Pansook, S.; Incharoensakdi, A.; Phunpruch, S. Effects of the photosystem II inhibitors CCCP and DCMU on hydrogen production by the unicellular halotolerant cyanobacterium Aphanothece halophytica. Sci. World J. 2019, 2019, 1030236. [Google Scholar] [CrossRef]
- Zhang, X.; Yang, X.; Huang, Y.; Hu, J.; Wu, D.; Yang, N.; Wang, H. 2-Hydroxychalcone as a Novel Natural Photosynthesis Inhibitor against Bloom-Forming Cyanobacteria. J. Agric. Food Chem. 2022, 70, 15069–15079. [Google Scholar] [CrossRef] [PubMed]
- Yi, X.; Gao, K. Impact of ultraviolet radiation nearly overrides the effects of elevated p CO2 on a prominent nitrogen-fixing cyanobacterium. Limnol. Oceanogr. 2023, 68, 557–568. [Google Scholar] [CrossRef]
- Rodriguez, I.; Fraga, M.; Alfonso, A.; Guillebault, D.; Medlin, L.; Baudart, J.; Jacob, P.; Helmi, K.; Meyer, T.; Breitenbach, U. Monitoring of freshwater toxins in European environmental waters by using novel multi-detection methods. Environ. Toxicol. Chem. 2017, 36, 645–654. [Google Scholar] [CrossRef]
- Singh, D.; Kaur, G.; Singh, J.; Satija, S. Microbial Strategies for Controlling Harmful Cyanobacterial Blooms. In Microbial Biotechnology: Basic Research and Applications; Springer: Singapore, 2020; pp. 189–204. [Google Scholar]
- Pandit, M.A.; Kumar, J.; Gulati, S.; Bhandari, N.; Mehta, P.; Katyal, R.; Rawat, C.D.; Mishra, V.; Kaur, J. Major biological control strategies for plant pathogens. Pathogens 2022, 11, 273. [Google Scholar] [CrossRef] [PubMed]
- Shah, S.T.; Basit, A.; Ullah, I.; Mohamed, H.I. Cyanobacteria and algae as biocontrol agents against fungal and bacterial plant pathogens. In Plant Growth-Promoting Microbes for Sustainable Biotic and Abiotic Stress Management; Springer: Cham, Switzerland, 2021; pp. 1–23. [Google Scholar]
- Jawed, A.; Kar, P.; Verma, R.; Shukla, K.; Hemanth, P.; Thakur, V.K.; Pandey, L.M.; Gupta, R.K. Integration of biological control with engineered heterojunction nano-photocatalysts for sustainable and effective management of water hyacinth weed. J. Environ. Chem. Eng. 2022, 10, 106976. [Google Scholar] [CrossRef]
- Kibuye, F.A.; Zamyadi, A.; Wert, E.C. A critical review on operation and performance of source water control strategies for cyanobacterial blooms: Part II-mechanical and biological control methods. Harmful Algae 2021, 109, 102119. [Google Scholar] [CrossRef]
- Yadav, P.; Singh, R.P.; Patel, A.K.; Pandey, K.; Gupta, R.K. Cyanobacteria as a Biocontrol Agent. In Microbial Biocontrol: Food Security and Post Harvest Management: Volume 2; Springer: Berlin/Heidelberg, Germany, 2022; pp. 167–185. [Google Scholar]
- El Semary, N.; Al Naim, H.; Aldayel, M.F. A novel application of laser in biocontrol of plant pathogenic bacteria. Appl. Sci. 2022, 12, 4933. [Google Scholar] [CrossRef]
- Banerji, A.; Benesh, K. Incorporating Microbial Species Interaction in Management of Freshwater Toxic Cyanobacteria: A Systems Science Challenge. Ecologies 2022, 3, 570–587. [Google Scholar] [CrossRef]
- Rawat, S.; Shahi, N.; Mallik, S.K.; Pathak, R.; Singh, B.; Pande, V. Biocontrol of Toxin Producing Cyanobacterium Microcystis aeruginosa by Algicidal Bacterium Exiguobacterium acetylicum Strain TM2 Isolated from Mid-Altitudinal Himalayan Lake of Northern India. Int. J. Curr. Microbiol. App. Sci. 2021, 10, 170–187. [Google Scholar]
- Ying, A.; Bi, Y.; Hu, Z. Response of predominant phytoplankton species to anthropogenic impacts in Lake Taihu. J. Freshw. Ecol. 2015, 30, 99–112. [Google Scholar] [CrossRef]
- Wurtsbaugh, W.A.; Paerl, H.W.; Dodds, W.K. Nutrients, eutrophication and harmful algal blooms along the freshwater to marine continuum. Wiley Interdiscip. Rev. Water 2019, 6, e1373. [Google Scholar] [CrossRef]
- Wu, W.-M.; Yang, J.; Criddle, C.S. Microplastics pollution and reduction strategies. Front. Environ. Sci. Eng. 2017, 11, 1–4. [Google Scholar] [CrossRef]
- Zaragüeta, M.; Acebes, P. Controlling eutrophication in a Mediterranean shallow reservoir by phosphorus loading reduction: The need for an integrated management approach. Environ. Manag. 2017, 59, 635–651. [Google Scholar] [CrossRef]
- Leong, Y.K.; Huang, C.-Y.; Chang, J.-S. Pollution prevention and waste phycoremediation by algal-based wastewater treatment technologies: The applications of high-rate algal ponds (HRAPs) and algal turf scrubber (ATS). J. Environ. Manag. 2021, 296, 113193. [Google Scholar] [CrossRef]
- Velu, C.; Cirés, S.; Brinkman, D.L.; Heimann, K. Bioproduct potential of outdoor cultures of Tolypothrix sp.: Effect of carbon dioxide and metal-rich wastewater. Front. Bioeng. Biotechnol. 2020, 8, 51. [Google Scholar] [CrossRef]
- Mitsch, W.J.; Mander, Ü. Ecological Engineering of Sustainable Landscapes; Elsevier: Amsterdam, The Netherlands, 2017; Volume 108, pp. 351–357. [Google Scholar]
- Overman, C.; Wells, S. Modeling Cyanobacteria Vertical Migration. Water 2022, 14, 953. [Google Scholar] [CrossRef]
- Rajendran, A.; Fox, T.; Reis, C.R.; Wilson, B.; Hu, B. Deposition of manure nutrients in a novel mycoalgae biofilm for nutrient management. Biocatal. Agric. Biotechnol. 2018, 14, 120–128. [Google Scholar] [CrossRef]
- Chong, J.W.R.; Yew, G.Y.; Khoo, K.S.; Ho, S.-H.; Show, P.L. Recent advances on food waste pretreatment technology via microalgae for source of polyhydroxyalkanoates. J. Environ. Manag. 2021, 293, 112782. [Google Scholar] [CrossRef] [PubMed]
- Paerl, H.W.; Otten, T.G.; Kudela, R. Mitigating the Expansion of Harmful Algal Blooms across the Freshwater-to-Marine Continuum; ACS Publications: Washington, DC, USA, 2018. [Google Scholar]
- Qin, B.; Li, W.; Zhu, G.; Zhang, Y.; Wu, T.; Gao, G. Cyanobacterial bloom management through integrated monitoring and forecasting in large shallow eutrophic Lake Taihu (China). J. Hazard. Mater. 2015, 287, 356–363. [Google Scholar] [CrossRef]
- Bormans, M.; Maršálek, B.; Jančula, D. Controlling internal phosphorus loading in lakes by physical methods to reduce cyanobacterial blooms: A review. Aquat. Ecol. 2016, 50, 407–422. [Google Scholar] [CrossRef]
- Nwankwegu, A.S.; Li, Y.; Huang, Y.; Wei, J.; Norgbey, E.; Sarpong, L.; Lai, Q.; Wang, K. Harmful algal blooms under changing climate and constantly increasing anthropogenic actions: The review of management implications. 3 Biotech 2019, 9, 1–19. [Google Scholar]
- Pang, C.; Radomyski, A.; Subramanian, V.; Nadimi-Goki, M.; Marcomini, A.; Linkov, I. Multi-criteria decision analysis applied to harmful algal bloom management: A case study. Integr. Environ. Assess. Manag. 2017, 13, 631–639. [Google Scholar] [CrossRef]
- Hamilton, D.P.; Salmaso, N.; Paerl, H.W. Mitigating harmful cyanobacterial blooms: Strategies for control of nitrogen and phosphorus loads. Aquat. Ecol. 2016, 50, 351–366. [Google Scholar] [CrossRef]
- Hamilton, D.P.; Wood, S.A.; Dietrich, D.R.; Puddick, J. Costs of harmful blooms of freshwater cyanobacteria. In Cyanobacteria: An Economic Perspective; Wiley: Hoboken, NJ, USA, 2014; pp. 245–256. [Google Scholar]
- Zeng, G.; Wang, P.; Wang, Y. Algicidal efficiency and mechanism of Phanerochaete chrysosporium against harmful algal bloom species. Algal Res. 2015, 12, 182–190. [Google Scholar] [CrossRef]
- Huang, H.; Xiao, X.; Lin, F.; Grossart, H.-P.; Nie, Z.; Sun, L.; Xu, C.; Shi, J. Continuous-release beads of natural allelochemicals for the long-term control of cyanobacterial growth: Preparation, release dynamics and inhibitory effects. Water Res. 2016, 95, 113–123. [Google Scholar] [CrossRef]
- Kumari, N.; Mishra, S.; Häder, D.-P.; Sinha, R.P. Cyanobacterial blooms and their implications in the changing environment. Adv. Environ. Eng. Res. 2022, 3, 1–41. [Google Scholar]
- Matthijs, H.C.; Jančula, D.; Visser, P.M.; Maršálek, B. Existing and emerging cyanocidal compounds: New perspectives for cyanobacterial bloom mitigation. Aquat. Ecol. 2016, 50, 443–460. [Google Scholar] [CrossRef]
- Park, Y.; Pyo, J.; Kwon, Y.S.; Cha, Y.; Lee, H.; Kang, T.; Cho, K.H. Evaluating physico-chemical influences on cyanobacterial blooms using hyperspectral images in inland water, Korea. Water Res. 2017, 126, 319–328. [Google Scholar] [CrossRef] [PubMed]
- Greenfield, D.I.; Duquette, A.; Goodson, A.; Keppler, C.J.; Williams, S.H.; Brock, L.M.; Stackley, K.D.; White, D.; Wilde, S.B. The effects of three chemical algaecides on cell numbers and toxin content of the cyanobacteria Microcystis aeruginosa and Anabaenopsis sp. Environ. Manag. 2014, 54, 1110–1120. [Google Scholar] [CrossRef] [PubMed]
- Amorim, C.A.; Dantas, Ê.W.; do Nascimento Moura, A. Modeling cyanobacterial blooms in tropical reservoirs: The role of physicochemical variables and trophic interactions. Sci. Total Environ. 2020, 744, 140659. [Google Scholar] [CrossRef] [PubMed]
- Huh, J.-H.; Ahn, J.-W. A perspective of chemical treatment for cyanobacteria control toward sustainable freshwater development. Environ. Eng. Res. 2017, 22, 1–11. [Google Scholar] [CrossRef]
- Sneed, J.M.; Meickle, T.; Engene, N.; Reed, S.; Gunasekera, S.; Paul, V.J. Bloom dynamics and chemical defenses of benthic cyanobacteria in the Indian River Lagoon, Florida. Harmful Algae 2017, 69, 75–82. [Google Scholar] [CrossRef]
- Rorar, J.; Garcia, L.D.; Cutright, T. Removal of saxitoxin and anatoxin-a by PAC in the presence and absence of microcystin-LR and/or cyanobacterial cells. J. Environ. Sci. 2023, 128, 161–170. [Google Scholar] [CrossRef]
- Qi, J.; Ma, B.; Miao, S.; Liu, R.; Hu, C.; Qu, J. Pre-oxidation enhanced cyanobacteria removal in drinking water treatment: A review. J. Environ. Sci. 2021, 110, 160–168. [Google Scholar] [CrossRef]
- Serrà, A.; Philippe, L.; Gómez, E. Removal of cyanobacteria and cyanotoxins in waters. Toxins 2021, 13, 636. [Google Scholar] [CrossRef]
- Pinkanjananavee, K.; Teh, S.J.; Kurobe, T.; Lam, C.H.; Tran, F.; Young, T.M. Potential Impacts on Treated Water Quality of Recycling Dewatered Sludge Supernatant during Harmful Cyanobacterial Blooms. Toxins 2021, 13, 99. [Google Scholar] [CrossRef]
- Castro-Jiménez, C.C.; Grueso-Domínguez, M.C.; Correa-Ochoa, M.A.; Saldarriaga-Molina, J.C.; García, E.F. A Coagulation Process Combined with a Multi-Stage Filtration System for Drinking Water Treatment: An Alternative for Small Communities. Water 2022, 14, 3256. [Google Scholar] [CrossRef]
- El Bouaidi, W.; Libralato, G.; Douma, M.; Ounas, A.; Yaacoubi, A.; Lofrano, G.; Albarano, L.; Guida, M.; Loudiki, M. A review of plant-based coagulants for turbidity and cyanobacteria blooms removal. Environ. Sci. Pollut. Res. 2022, 29, 42601–42615. [Google Scholar] [CrossRef] [PubMed]
- Akyol, Ç.; Ozbayram, E.G.; Accoroni, S.; Radini, S.; Eusebi, A.L.; Gorbi, S.; Vignaroli, C.; Bacchiocchi, S.; Campacci, D.; Gigli, F. Monitoring of cyanobacterial blooms and assessing polymer-enhanced microfiltration and ultrafiltration for microcystin removal in an Italian drinking water treatment plant. Environ. Pollut. 2021, 286, 117535. [Google Scholar] [CrossRef] [PubMed]
- Chae, S.; Noeiaghaei, T.; Oh, Y.; Kim, I.S.; Park, J.-S. Effective removal of emerging dissolved cyanotoxins from water using hybrid photocatalytic composites. Water Res. 2019, 149, 421–431. [Google Scholar] [CrossRef] [PubMed]
- Jalili, F.; Moradinejad, S.; Zamyadi, A.; Dorner, S.; Sauvé, S.; Prévost, M. Evidence-Based Framework to Manage Cyanobacteria and Cyanotoxins in Water and Sludge from Drinking Water Treatment Plants. Toxins 2022, 14, 410. [Google Scholar] [CrossRef]
- Munoz, M.; Cirés, S.; de Pedro, Z.M.; Colina, J.Á.; Velásquez-Figueroa, Y.; Carmona-Jiménez, J.; Caro-Borrero, A.; Salazar, A.; Fuster, M.-C.S.M.; Contreras, D. Overview of toxic cyanobacteria and cyanotoxins in Ibero-American freshwaters: Challenges for risk management and opportunities for removal by advanced technologies. Sci. Total Environ. 2021, 761, 143197. [Google Scholar] [CrossRef] [PubMed]
- Vlad, S.; Anderson, W.B.; Peldszus, S.; Huck, P.M. Removal of the cyanotoxin anatoxin-a by drinking water treatment processes: A review. J. Water Health 2014, 12, 601–617. [Google Scholar] [CrossRef] [PubMed]
- Rastogi, R.P.; Sinha, R.P.; Moh, S.H.; Lee, T.K.; Kottuparambil, S.; Kim, Y.-J.; Rhee, J.-S.; Choi, E.-M.; Brown, M.T.; Häder, D.-P. Ultraviolet radiation and cyanobacteria. J. Photochem. Photobiol. B Biol. 2014, 141, 154–169. [Google Scholar] [CrossRef] [PubMed]
- Abbas, T.; Kajjumba, G.W.; Ejjada, M.; Masrura, S.U.; Marti, E.J.; Khan, E.; Jones-Lepp, T.L. Recent advancements in the removal of cyanotoxins from water using conventional and modified adsorbents—A contemporary review. Water 2020, 12, 2756. [Google Scholar] [CrossRef]
- Weir, M.H.; Wood, T.A.; Zimmer-Faust, A. Development of methods to estimate microcystins removal and water treatment resiliency using mechanistic risk modelling. Water Res. 2021, 190, 116763. [Google Scholar] [CrossRef]
Figure 1.
Cyanobacterial growth and dissemination on lake (a–c) and river (d) surfaces [3,8]. Reproduced with permission from Tao Lyu, Lirong Song, Qiuwen Chen, and Gang Pan, Water Journal; published by MDPI, 2020. Reproduced with permission from Moreira, C.; Vasconcelos, V.; Antunes, A., Earth Journal; published by MDPI, 2022.
Figure 1.
Cyanobacterial growth and dissemination on lake (a–c) and river (d) surfaces [3,8]. Reproduced with permission from Tao Lyu, Lirong Song, Qiuwen Chen, and Gang Pan, Water Journal; published by MDPI, 2020. Reproduced with permission from Moreira, C.; Vasconcelos, V.; Antunes, A., Earth Journal; published by MDPI, 2022.

Figure 2.
Factors promoting cyanobacteria bloom formation and diffusion.

Figure 3.
Chemical structures of microcystin (1), microcystin-LR (2), microcystin-LA (3), microcystin-YR (4), microcystin-RR (5), microcystin-LF (6), Nodularin (7), Cylindrospermopsin (8), Anatoxin-a (9), Homoanatoxin (10), Oscillatoxin A (11), and Nakienones A–C (12–14).
Figure 3.
Chemical structures of microcystin (1), microcystin-LR (2), microcystin-LA (3), microcystin-YR (4), microcystin-RR (5), microcystin-LF (6), Nodularin (7), Cylindrospermopsin (8), Anatoxin-a (9), Homoanatoxin (10), Oscillatoxin A (11), and Nakienones A–C (12–14).

Figure 4.
Chemical structures of aphantoxin (15), debromoaplysiatoxin (16), scytophycins A-E (17–21), lyngbyatoxins A-C (22–24), and acutiphycin (25).
Figure 4.
Chemical structures of aphantoxin (15), debromoaplysiatoxin (16), scytophycins A-E (17–21), lyngbyatoxins A-C (22–24), and acutiphycin (25).

Figure 5.
Techniques and methods for managing and mitigating cyanobacterial blooms and toxins.

Table 2.
A summary of the effectiveness of removal methods for intracellular cyanotoxins.
| Treatment Technique | Effectiveness of the Technique |
|---|---|
| Oxidation (pre-treatment) [343] | Oxidation stresses or kills cyanobacteria cells that release cyanotoxin into the water. |
| Coagulation/Sedimentation/Filtration [346,347] | After sludge separation, it must be guaranteed that the supernatant sludge does not re-enter the supply. |
| Filter membranes [349] | Intracellular cyanotoxins (cyanobacteria cells) can be effectively removed. Microfiltration and ultrafiltration work well when cells are not allowed to accumulate on membranes for extended periods of time. More frequent cleaning may be required during a bloom episode. |
| Flotation [350] | Since many cyanobacteria that produce toxins are buoyant, flotation techniques like Dissolved Air Flotation (DAF) are effective at removing cyanotoxins from the body. |
Table 3.
A summary of the effectiveness of removal methods for extracellular cyanotoxins.
| Treatment Technique | Effectiveness of the Technique |
|---|---|
| Filter membranes [350] |
|
| KMnO4 [351] | Microcystins and anatoxins are effectively oxidized, whereas saxitoxin is not. |
| Ozone [352] | Microcystins, anatoxin-a, and cylindrospermopsin are all quite effective at being oxidized. Saxitoxin oxidation is ineffective. |
| Free Chlorine [351] | As long as the pH is less than 8, it is effective for oxidizing microcystins. Cylindrospermopsin and saxitoxin are effectively oxidized. Anatoxin-a oxidation is ineffective. |
| Ultraviolet radiation [354] | Microcystins and cylindrospermopsin cannot be oxidized by UV radiation alone at dosages typically used in drinking water treatment. UV light is effective at oxidizing anatoxin-a, cylindrospermopsin, and, at high UV dosages, microcystins when mixed with ozone or hydrogen peroxide. |
| Activated Carbon Adsorption [354] |
|
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Thawabteh, A.M.; Naseef, H.A.; Karaman, D.; Bufo, S.A.; Scrano, L.; Karaman, R. Understanding the Risks of Diffusion of Cyanobacteria Toxins in Rivers, Lakes, and Potable Water. Toxins 2023, 15, 582. https://doi.org/10.3390/toxins15090582
AMA Style
Thawabteh AM, Naseef HA, Karaman D, Bufo SA, Scrano L, Karaman R. Understanding the Risks of Diffusion of Cyanobacteria Toxins in Rivers, Lakes, and Potable Water. Toxins. 2023; 15(9):582. https://doi.org/10.3390/toxins15090582
Chicago/Turabian StyleThawabteh, Amin Mahmood, Hani A Naseef, Donia Karaman, Sabino A. Bufo, Laura Scrano, and Rafik Karaman. 2023. "Understanding the Risks of Diffusion of Cyanobacteria Toxins in Rivers, Lakes, and Potable Water" Toxins 15, no. 9: 582. https://doi.org/10.3390/toxins15090582
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.