
Seed-Encapsulation of Desiccation-Tolerant Microorganisms for the Protection of Maize from Drought: Phenotyping Effects of a New Dry Bioformulation

 , , and
, , and 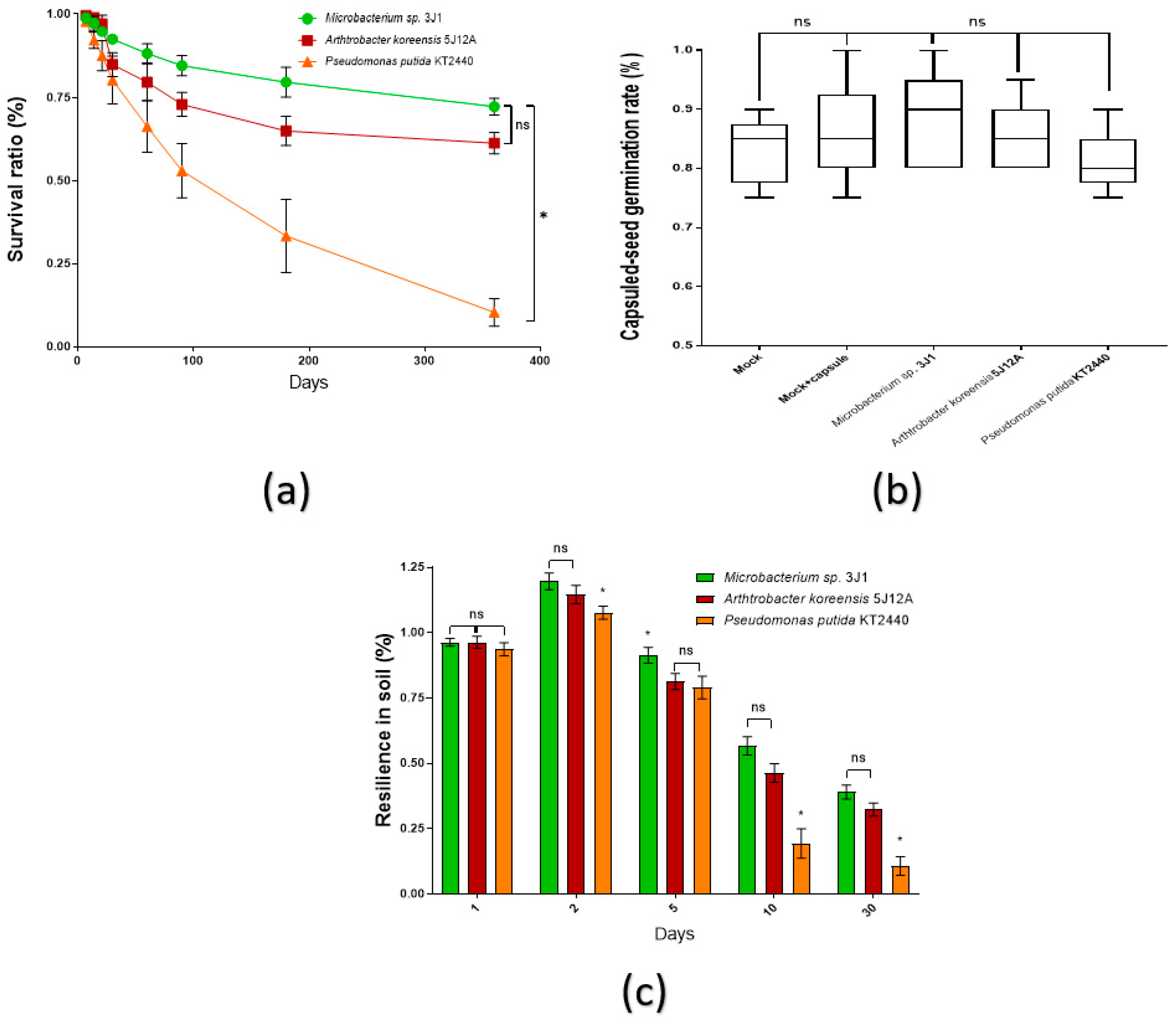{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
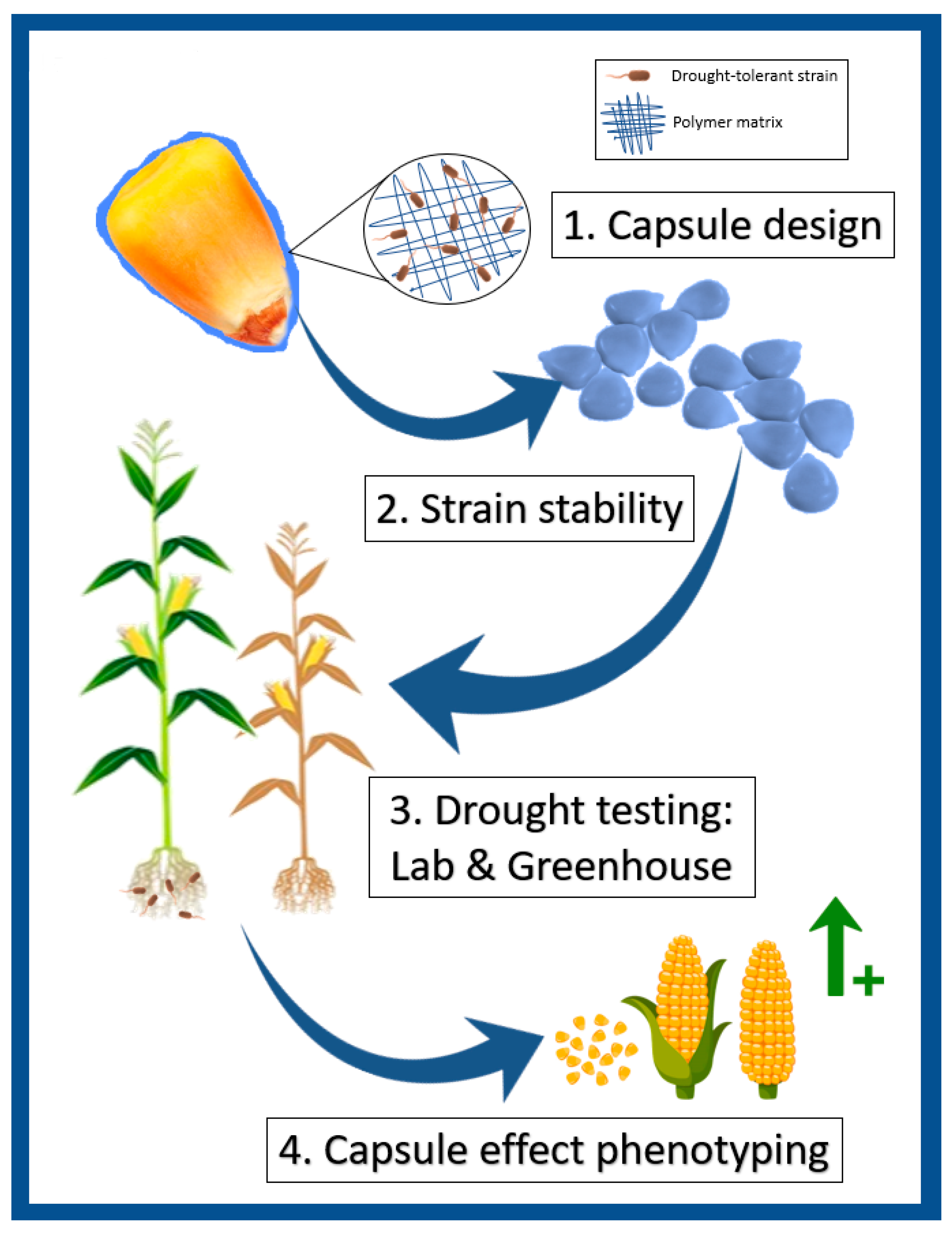
2.1. Strains Survival in Coating Formulation, Capsuled Seeds Germination and Resilience Evaluation in Soil
2.2. Laboratory Tests
2.3. Greenhouse Tests
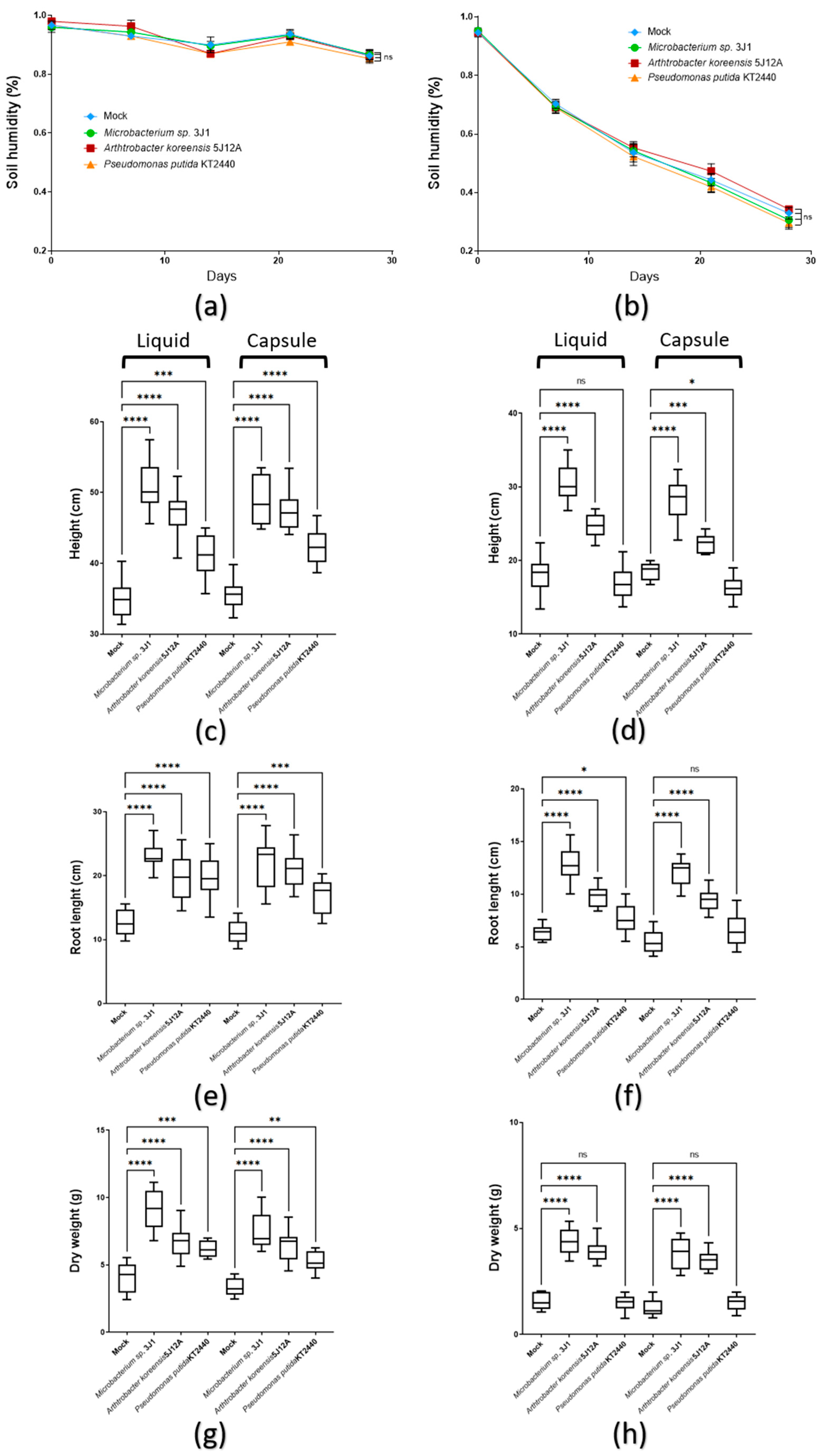
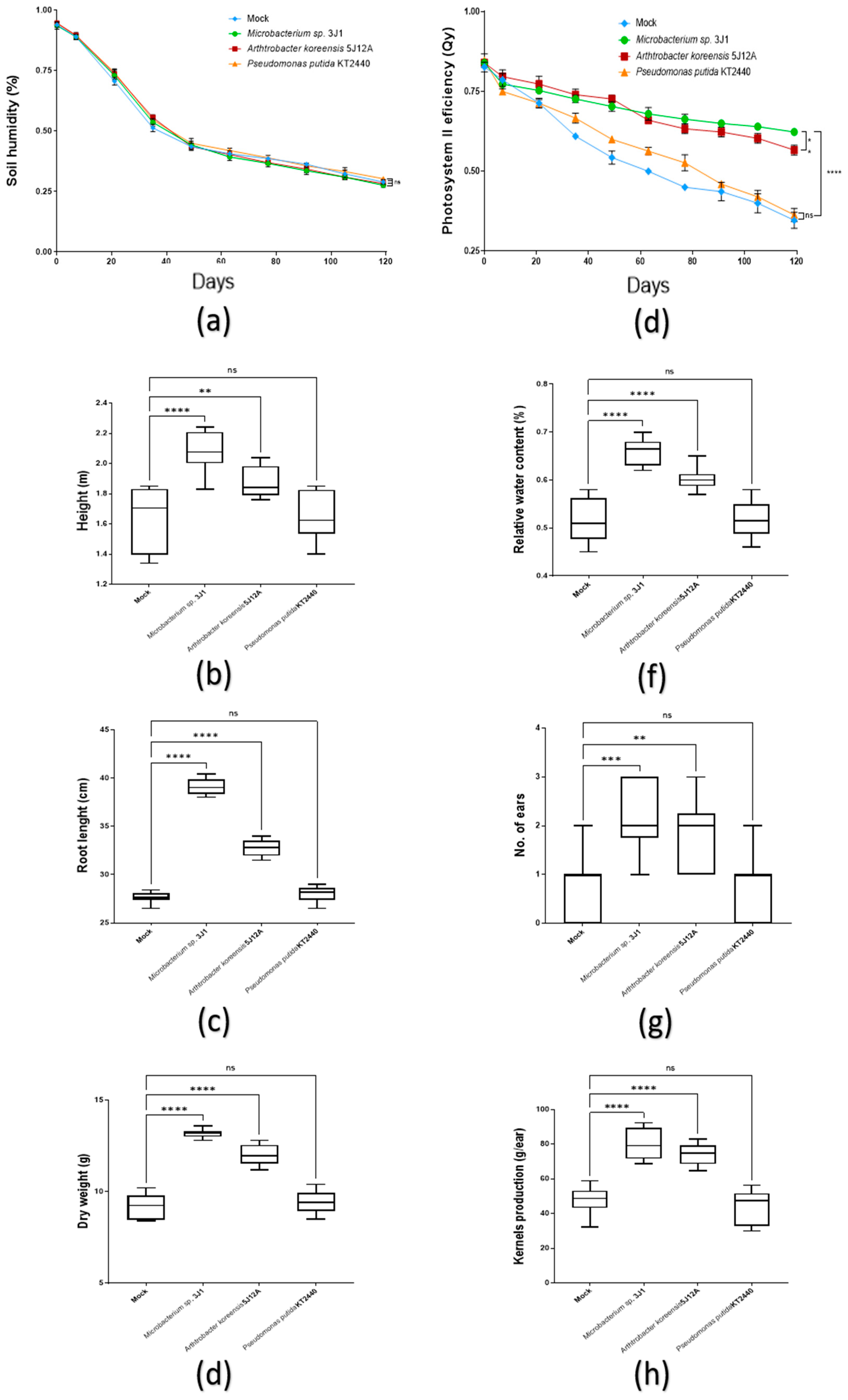
2.3.1. Plant Growth Promotion
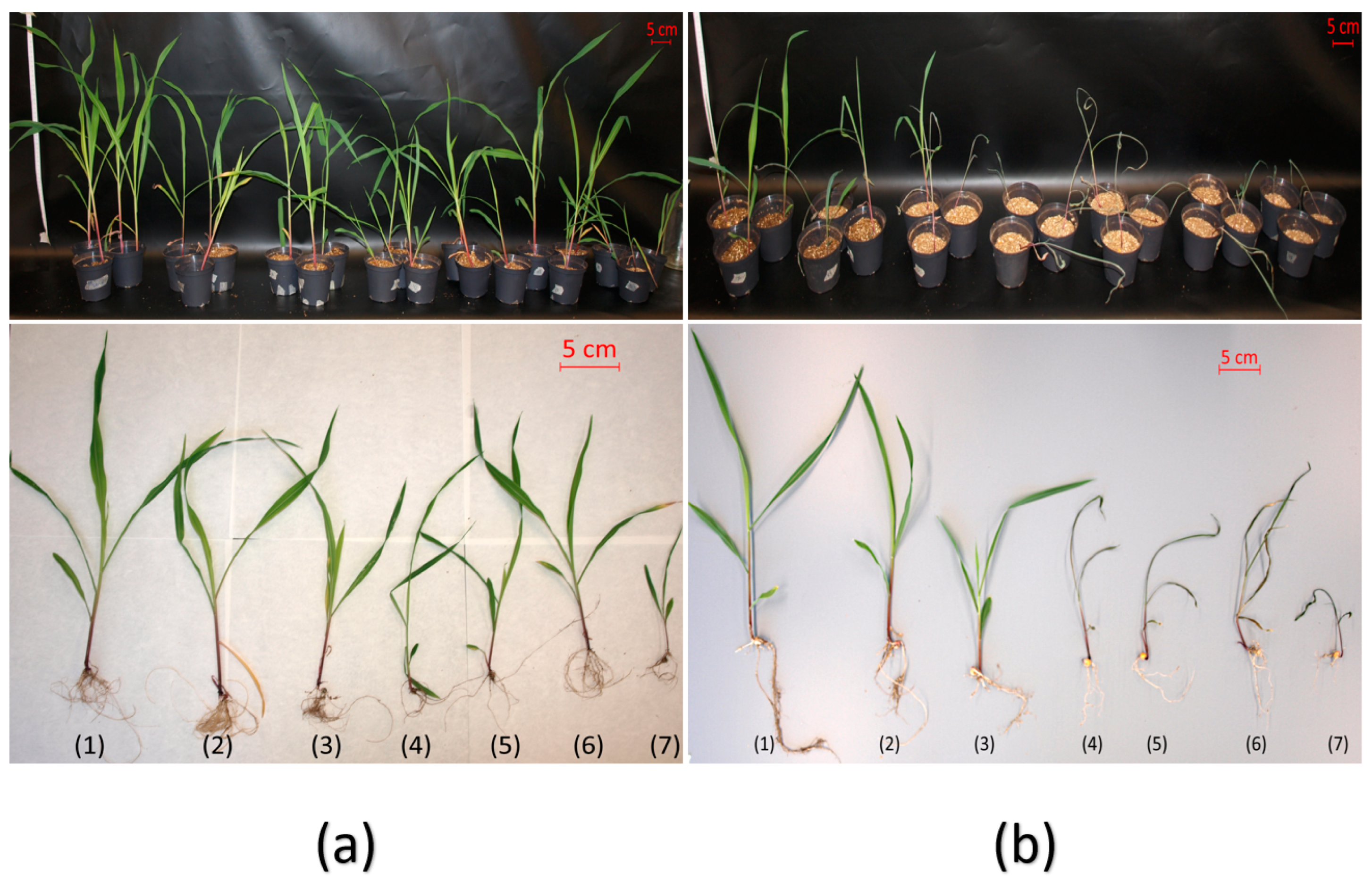
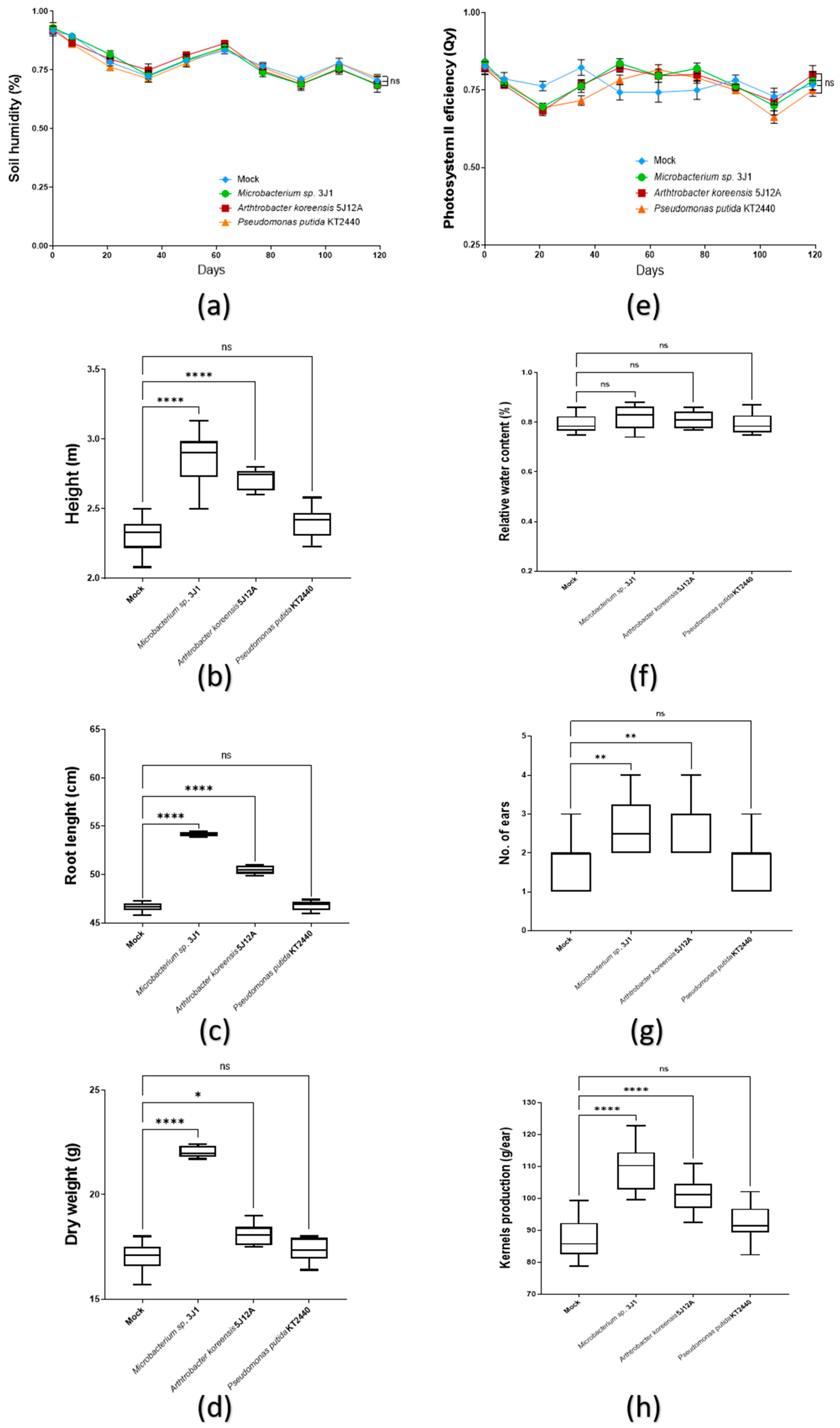
2.3.2. Drought Tolerance Test
3. Discussion
4. Materials and Methods
4.1. Culture and Preparation of the Strains
4.2. Preparation of Liquid Inoculum and Capsules. Assessment of the Viability of the Cells in Coating and Germination of Seeds Evaluation of Resilience in Soil Evaluation
4.3. Laboratory Tests
4.4. Greenhouse Tests
4.5. Phenotype Evaluation
4.6. Statistical Study
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mohanty, P.; Singh, P.K.; Chakraborty, D.; Mishra, S.; Pattnaik, R. Insight into the Role of PGPR in Sustainable Agriculture and Environment. Front. Sustain. Food Syst. 2021, 5, 183. [Google Scholar] [CrossRef]
- Markets & Markets. Biofertilizers Market by form (Liquid, Carrier-Based), Mode of Application (Soil Treatment, Seed Treatment), Crop Type, Type (Nitrogen-Fixing, Phosphate Solubilizing & Mobilizing, Potash Solubilizing & Mobilizing), Region—Global Forecast to 2025; Markets & Markets: Pune, India, 2020. [Google Scholar]
- Goldstein Market Intelligence. Global Biofertilizer Industry Analysis by Bio-Fertilizer Type, by Crop Type, by Application and, by Geography with COVID-19 Impact | Market Outlook 2017–2030; Goldstein Market Intelligence: New York, NY, USA, 2020. [Google Scholar]
- Research and Markets. Biofertilizers Market Research and Analysis, 2020—Trends, Growth Opportunities and Forecasts to 2030; Research and Markets: The Liberties, Ireland, 2020. [Google Scholar]
- Barros-Rodríguez, A.; Rangseekaew, P.; Lasudee, K.; Pathom-aree, W.; Manzanera, M. Regulatory risks associated with bacteria as biostimulants and biofertilizers in the frame of the European Regulation (EU) 2019/1009. Sci. Total Environ. 2020, 740, 140239. [Google Scholar] [CrossRef] [PubMed]
- Schütz, L.; Gattinger, A.; Meier, M.; Müller, A.; Boller, T.; Mäder, P.; Mathimaran, N. Improving Crop Yield and Nutrient Use Efficiency via Biofertilization—A Global Meta-analysis. Front. Plant Sci. 2018, 8, 2204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basu, A.; Prasad, P.; Das, S.N.; Kalam, S.; Sayyed, R.Z.; Reddy, M.S.; El Enshasy, H. Plant Growth Promoting Rhizobacteria (PGPR) as Green Bioinoculants: Recent Developments, Constraints, and Prospects. Sustainability 2021, 13, 1140. [Google Scholar] [CrossRef]
- Herrmann, L.; Lesueur, D. Challenges of formulation and quality of biofertilizers for successful inoculation. Appl. Microbiol. Biotechnol. 2013, 97, 8859–8873. [Google Scholar] [CrossRef] [PubMed]
- Mitter, E.K.; Tosi, M.; Obregón, D.; Dunfield, K.E.; Germida, J.J. Rethinking Crop Nutrition in Times of Modern Microbiology: Innovative Biofertilizer Technologies. Front. Sustain. Food Syst. 2021, 5, 29. [Google Scholar] [CrossRef]
- Rocha, I.; Souza-Alonso, P.; Pereira, G.; Ma, Y.; Vosátka, M.; Freitas, H.; Oliveira, R.S. Using microbial seed coating for improving cowpea productivity under a low-input agricultural system. J. Sci. Food Agric. 2020, 100, 1092–1098. [Google Scholar] [CrossRef]
- Agake, S.; Ramirez, M.D.A.; Kojima, K.; Ookawa, T.; Ohkama-Ohtsu, N.; Yokoyama, T. Seed coating by biofertilizer containing spores of Bacillus pumilus TUAT1 strain enhanced initial growth of Oryza sativa L. Agron. J. 2021, 113, 3708–3717. [Google Scholar] [CrossRef]
- Huang, X.-F.; Zhou, D.; Lapsansky, E.R.; Reardon, K.F.; Andales, M.J.; Vivanco, J.M.; Manter, D.K.; Guo, J. Mitsuaria sp. and Burkholderia sp. from Arabidopsis rhizosphere enhance drought tolerance in Arabidopsis thaliana and maize (Zea mays L.). Plant Soil 2017, 419, 523–539. [Google Scholar] [CrossRef]
- Danish, S.; Zafar-Ul-Hye, M.; Fahad, S.; Saud, S.; Brtnicky, M.; Hammerschmiedt, T.; Datta, R. Drought Stress Alleviation by ACC Deaminase Producing Achromobacter xylosoxidans and Enterobacter cloacae, with and without Timber Waste Biochar in Maize. Sustainability 2020, 12, 6286. [Google Scholar] [CrossRef]
- Alori, E.T.; Dare, M.O.; Babalola, O.O. Microbial Inoculants for Soil Quality and Plant Health. In Sustainable Agriculture Reviews; Lichtfouse, E., Ed.; Springer International Publishing: Cham, Switzerland, 2017; pp. 281–307. [Google Scholar]
- Fontana, C.G.; Narváez-Reinaldo, J.J.; Castillo, F.; González-López, J.; Luque, I.; Manzanera, M. A New Physiological Role for the DNA Molecule as a Protector against Drying Stress in Desiccation-Tolerant Microorganisms. Front. Microbiol. 2016, 7, 2066. [Google Scholar] [CrossRef] [Green Version]
- Vílchez, J.I.; Niehaus, K.; Dowling, D.N.; González-López, J.; Manzanera, M. Protection of pepper plants from drought by Microbacterium sp. 3J1 by modulation of the plant’s glutamine and α-ketoglutarate Content: A Comparative Metabolomics Approach. Front. Microbiol. 2018, 9, 284. [Google Scholar] [CrossRef] [PubMed]
- García-Fontana, C.; Vilchez, J.I.; Manzanera, M. Proteome Comparison Between Natural Desiccation-Tolerant Plants and Drought-Protected Caspicum annuum Plants by Microbacterium sp. 3J1. Front. Microbiol. 2020, 11, 1537. [Google Scholar] [CrossRef]
- Narváez-Reinaldo, J.J.; Barba, I.; González-López, J.; Tunnacliffe, A.; Manzanera, M. Rapid Method for Isolation of Desiccation-Tolerant Strains and Xeroprotectants. Appl. Environ. Microbiol. 2010, 76, 5254–5262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, N.; Nesme, J.; Røder, H.L.; Li, X.; Zuo, Z.; Petersen, M.; Burmølle, M.; Sørensen, S.J. Emergent bacterial community properties induce enhanced drought tolerance in Arabidopsis. NPJ Biofilms Microbiomes 2021, 7, 1574. [Google Scholar] [CrossRef] [PubMed]
- Vurukonda, S.S.K.P.; Vardharajula, S.; Shrivastava, M.; SkZ, A. Enhancement of drought stress tolerance in crops by plant growth promoting rhizobacteria. Microbiol. Res. 2016, 184, 13–24. [Google Scholar] [CrossRef] [PubMed]
- Niu, X.; Song, L.; Xiao, Y.; Ge, W. Drought-tolerant plant growth-promoting rhizobacteria associated with foxtail millet in a semi-arid agroecosystem and their potential in alleviating drought stress. Front. Microbiol. 2018, 8, 2580. [Google Scholar] [CrossRef]
- Vilchez, S.; Manzanera, M. Biotechnological uses of desiccation-tolerant microorganisms for the rhizoremediation of soils subjected to seasonal drought. Appl. Microbiol. Biotechnol. 2011, 91, 1297–1304. [Google Scholar] [CrossRef]
- Manzanera, M. Dealing with water stress and microbial preservation. Environ. Microbiol. 2021, 23, 3351–3359. [Google Scholar] [CrossRef]
- Guo, J.-J.; Huang, X.-P.; Xiang, L.; Wang, Y.-Z.; Li, Y.-W.; Li, H.; Cai, Q.-Y.; Mo, C.-H.; Wong, M.-H. Source, migration and toxicology of microplastics in soil. Environ. Int. 2020, 137, 105263. [Google Scholar] [CrossRef]
- Yang, L.; Zhang, Y.; Kang, S.; Wang, Z.; Wu, C. Microplastics in soil: A review on methods, occurrence, sources, and potential risk. Sci. Total Environ. 2021, 780, 146546. [Google Scholar] [CrossRef] [PubMed]
- Nelson, L.M. Plant Growth Promoting Rhizobacteria (PGPR): Prospects for New Inoculants. Crop Manag. 2004, 3, 1–7. [Google Scholar] [CrossRef]
- Baez, A.; Morales-García, Y.E.; Quintero-Hernández, V.; Muñoz-Rojas, J. Next generation of microbial inoculants for agriculture and bioremediation. Microb. Biotechnol. 2017, 10, 19–21. [Google Scholar] [CrossRef] [PubMed]
- Kumari, R.; Singh, D.P. Nano-biofertilizer: An Emerging Eco-friendly Approach for Sustainable Agriculture. Natl. Acad. Sci. India Sect. B Biol. Sci. 2020, 90, 733–741. [Google Scholar] [CrossRef]
- Kloepper, J.W. Development of a Powder Formulation of Rhizobacteria for Inoculation of Potato Seed Pieces. Phytopathology 1981, 71, 590–592. [Google Scholar] [CrossRef] [Green Version]
- Manzanera, M.; Vilchez, S.; Tunnacliffe, A. Plastic Encapsulation of Stabilized Escherichia coli and Pseudomonas putida. Appl. Environ. Microbiol. 2004, 70, 3143–3145. [Google Scholar] [CrossRef] [Green Version]
- Malusa, E.; Sas-Paszt, L.; Ciesielska, J. Technologies for Beneficial Microorganisms Inocula Used as Biofertilizers. Sci. World J. 2012, 2012, 491206. [Google Scholar] [CrossRef]
- Adjanohoun, A.; Allagbe, M.; Noumavo, A.P.; Gotoechan-Hodonou, H.; Sikirou, R.; Dossa, K.K.; Glele Kakaï, R.L.; Kotchoni, S.; Baba-Moussa, L. Effects of plant growth promoting rhizobacteria on field grown maize. J. Anim. Plant Sci. 2011, 11, 1457–1465. [Google Scholar]
- Mondal, S.; Dutta, S.; Banerjee, A.; Banerjee, S.; Datta, R.; Roy, P.; Podder, A.; Roy, R.; Basu, P. Production and application of phosphate solubilizing bacteria as biofertilizer: Field trial at maize field, Uchalan, Burdwan District, West Bengal. Int. J. Environ. Agric. Res. 2017, 3, 1–9. [Google Scholar]
- Cortivo, C.D.; Ferrari, M.; Visioli, G.; Lauro, M.; Fornasier, F.; Barion, G.; Panozzo, A.; Vamerali, T. Effects of Seed-Applied Biofertilizers on Rhizosphere Biodiversity and Growth of Common Wheat (Triticum aestivum L.) in the Field. Front. Plant Sci. 2020, 11, 72. [Google Scholar] [CrossRef] [Green Version]
- Benizri, E.; Baudoin, E.; Guckert, A. Root Colonization by Inoculated Plant Growth-Promoting Rhizobacteria. Biocontrol Sci. Technol. 2001, 11, 557–574. [Google Scholar] [CrossRef]
- Segura, A.; Rodríguez-Conde, S.; Ramos, C.; Ramos, J.L. Bacterial responses and interactions with plants during rhizoremediation. Microb. Biotechnol. 2009, 2, 452–464. [Google Scholar] [CrossRef] [PubMed]
- Albareda, M.; Dardanelli, M.S.; Sousa, C.; Megãas, M.; Temprano, F.; Rodrãguez-Navarro, D.N. Factors affecting the attachment of rhizospheric bacteria to bean and soybean roots. FEMS Microbiol. Lett. 2006, 259, 67–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bloemberg, G.; Lugtenberg, B.; Bloemberg, G.V.; Lugtenberg, B.J.J. Molecular basis of plant growth promotion and biocontrol by rhizobacteria. Curr. Opin. Plant Biol. 2001, 4, 343–350. [Google Scholar] [CrossRef]
- Trabelsi, D.; Mhamdi, R. Microbial Inoculants and Their Impact on Soil Microbial Communities: A Review. BioMed Res. Int. 2013, 2013, 863240. [Google Scholar] [CrossRef] [PubMed]
- Bergsma-Vlami, M.; Prins, M.E.; Raaijmakers, J.M. Influence of plant species on population dynamics, genotypic diversity and antibiotic production in the rhizosphere by indigenous Pseudomonas spp. FEMS Microbiol. Ecol. 2005, 52, 59–69. [Google Scholar] [CrossRef] [PubMed]
- Vacheron, J.; Desbrosses, G.; Bouffaud, M.-L.; Touraine, B.; Moënne-Loccoz, Y.; Muller, D.; Legendre, L.; Wisniewski-Dyé, F.; Prigent-Combaret, C. Plant growth-promoting rhizobacteria and root system functioning. Front. Plant Sci. 2013, 4, 356. [Google Scholar] [CrossRef] [Green Version]
- Vílchez, J.I. Mejora en la Tolerancia a la Sequía de Plantas con Interés Biotecnológico en Zonas de Secano; University of Granada: Granada, Spain, 2016. [Google Scholar]
- Manzanera, M.; Narváez-Reinaldo, J.J.; García-Fontana, C.; Vílchez, J.I.; González-López, J. Genome Sequence of Arthrobacter koreensis 5J12A, a Plant Growth-Promoting and Desiccation-Tolerant Strain. Genome Announc. 2015, 3, e00648-15. [Google Scholar] [CrossRef] [Green Version]
- Manzanera, M.; García-Fontana, C.; Vílchez, J.I.; Narváez-Reinaldo, J.J.; González-López, J. Genome Sequence of Microbacterium sp. Strain 3J1, a Highly Desiccation-Tolerant Bacterium That Promotes Plant Growth. Genome Announc. 2015, 3, e00713-15. [Google Scholar] [CrossRef] [Green Version]
- Manzanera, M.; Santa-Cruz-Calvo, L.; Vílchez, J.I.; García-Fontana, C.; Silva-Castro, G.A.; Calvo, C.; González-López, J. Genome Sequence of Arthrobacter siccitolerans 4J27, a Xeroprotectant-Producing Desiccation-Tolerant Microorganism. Genome Announc. 2014, 2, e00526-14. [Google Scholar] [CrossRef] [Green Version]
- Manzanera, M.; Vílchez, J.I.; García-Fontana, C.; Calvo, C.; González-López, J. Genome Sequence of Leucobacter sp. 4J7B1, a Plant-Osmoprotectant Soil Microorganism. Genome Announc. 2015, 3, e00398-15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manzanera, M.; García-Fontana, C.; Vílchez, J.I.; González-López, J. Genome Sequence of Rhodococcus sp. 4J2A2, a Desiccation-Tolerant Bacterium Involved in Biodegradation of Aromatic Hydrocarbons. Genome Announc. 2015, 3, e00592-15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manzanera, M.; García de Castro, A.; Tøndervik, A.; Rayner-Brandes, M.; Strøm, A.R.; Tunnacliffe, A. Hydroxyectoine is superior to trehalose for anhydrobiotic engineering of Pseudomonas putida KT2440. Appl. Environ. Microbiol. 2002, 68, 4328–4333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pieczynski, M.; Marczewski, W.; Hennig, J.; Dolata, J.; Bielewicz, D.; Piontek, P.; Wyrzykowska, A.; Krusiewicz, D.; Strzelczyk-Zyta, D.; Konopka-Postupolska, D.; et al. Down-regulation ofCBP80gene expression as a strategy to engineer a drought-tolerant potato. Plant Biotechnol. J. 2013, 11, 459–469. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rebelo Romão, I.; Rodrigues dos Santos, A.S.; Velasco, L.; Martínez-Ferri, E.; Vilchez, J.I.; Manzanera, M. Seed-Encapsulation of Desiccation-Tolerant Microorganisms for the Protection of Maize from Drought: Phenotyping Effects of a New Dry Bioformulation. Plants 2022, 11, 1024. https://doi.org/10.3390/plants11081024
Rebelo Romão I, Rodrigues dos Santos AS, Velasco L, Martínez-Ferri E, Vilchez JI, Manzanera M. Seed-Encapsulation of Desiccation-Tolerant Microorganisms for the Protection of Maize from Drought: Phenotyping Effects of a New Dry Bioformulation. Plants. 2022; 11(8):1024. https://doi.org/10.3390/plants11081024
Chicago/Turabian StyleRebelo Romão, Inês, Ana Sofia Rodrigues dos Santos, Leonardo Velasco, Elsa Martínez-Ferri, Juan Ignacio Vilchez, and Maximino Manzanera. 2022. "Seed-Encapsulation of Desiccation-Tolerant Microorganisms for the Protection of Maize from Drought: Phenotyping Effects of a New Dry Bioformulation" Plants 11, no. 8: 1024. https://doi.org/10.3390/plants11081024