
Screening and Research on Skin Barrier Damage Protective Efficacy of Different Mannosylerythritol Lipids


Abstract
:1. Introduction
2. Results
2.1. Isolation and TLC Verification of MELs
2.2. Quantitative Detection of MELs
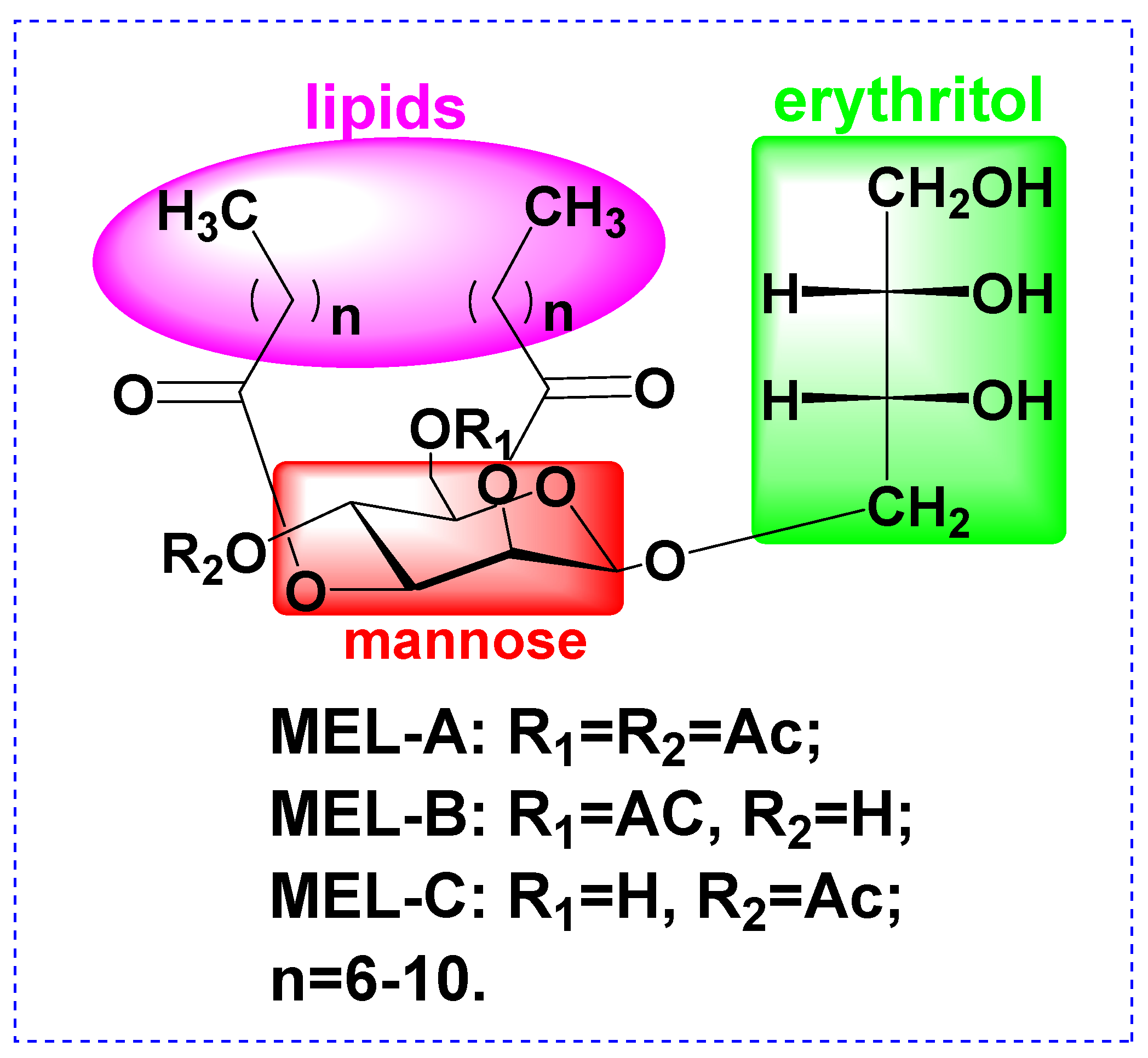
2.3. Physicochemical Characterisation of MELs
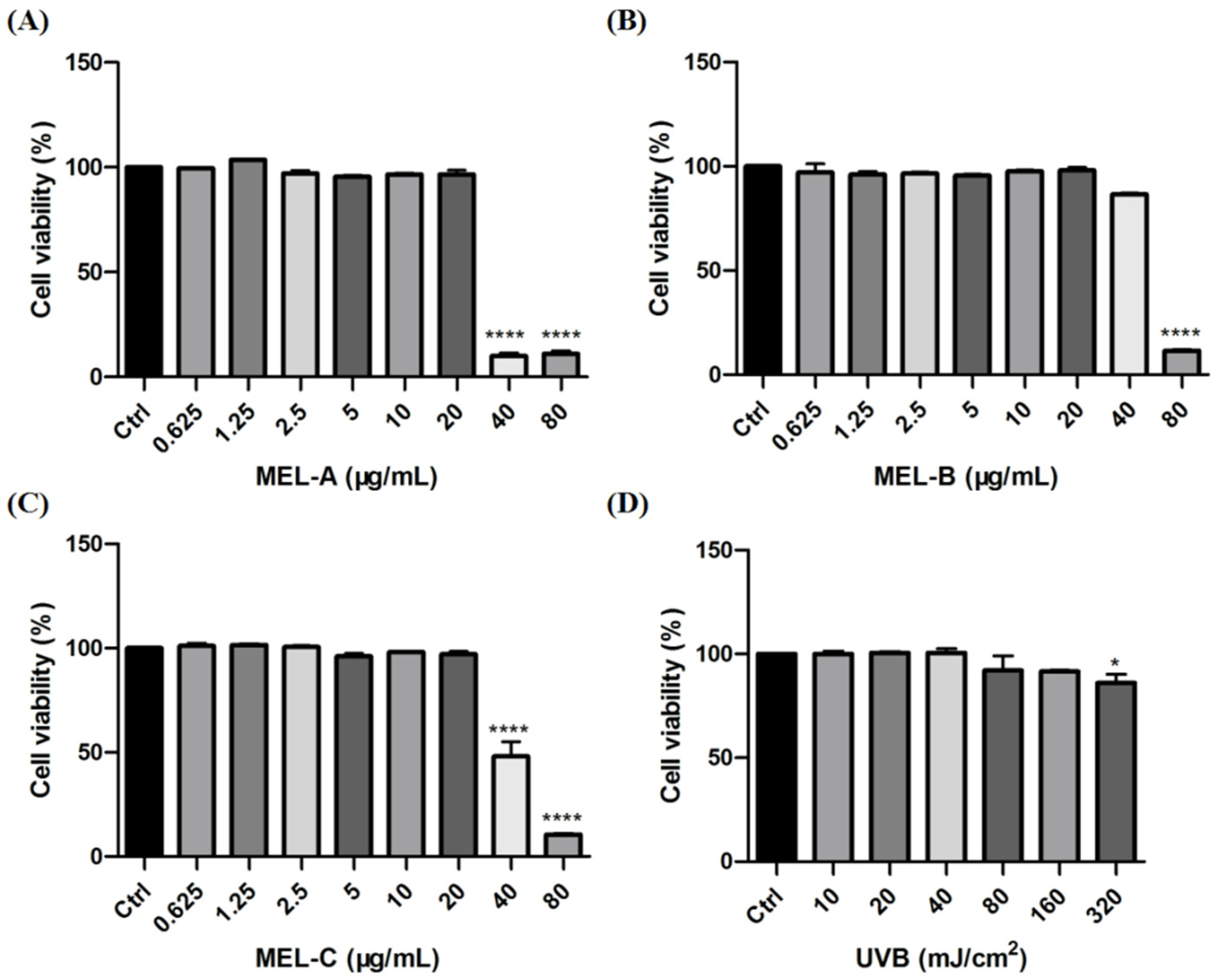
2.4. Effect of MEL and UVB on Cell Viability in HaCaT Cells
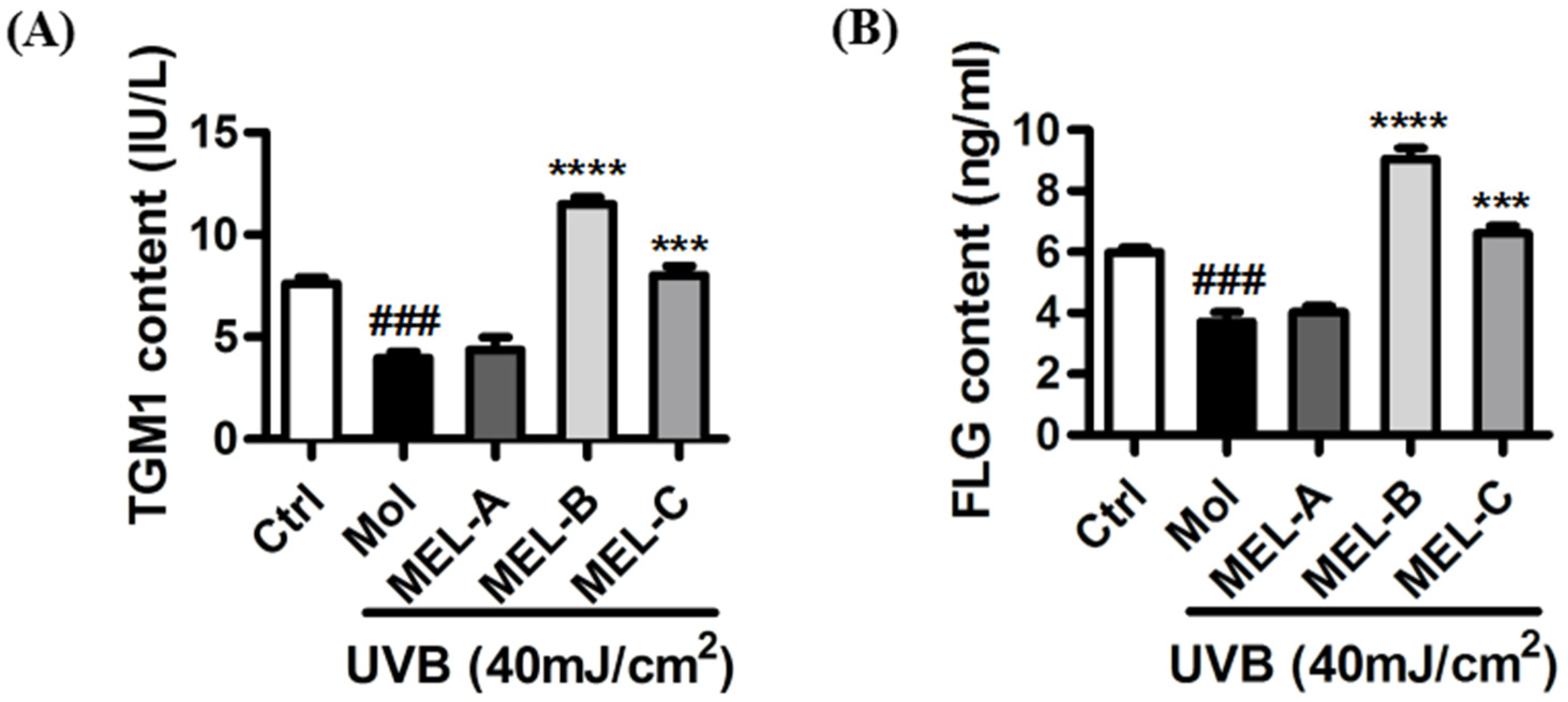
2.5. Effect of MELs on Contents of FLG and TGM1 in UVB-Induced HaCaT Cells
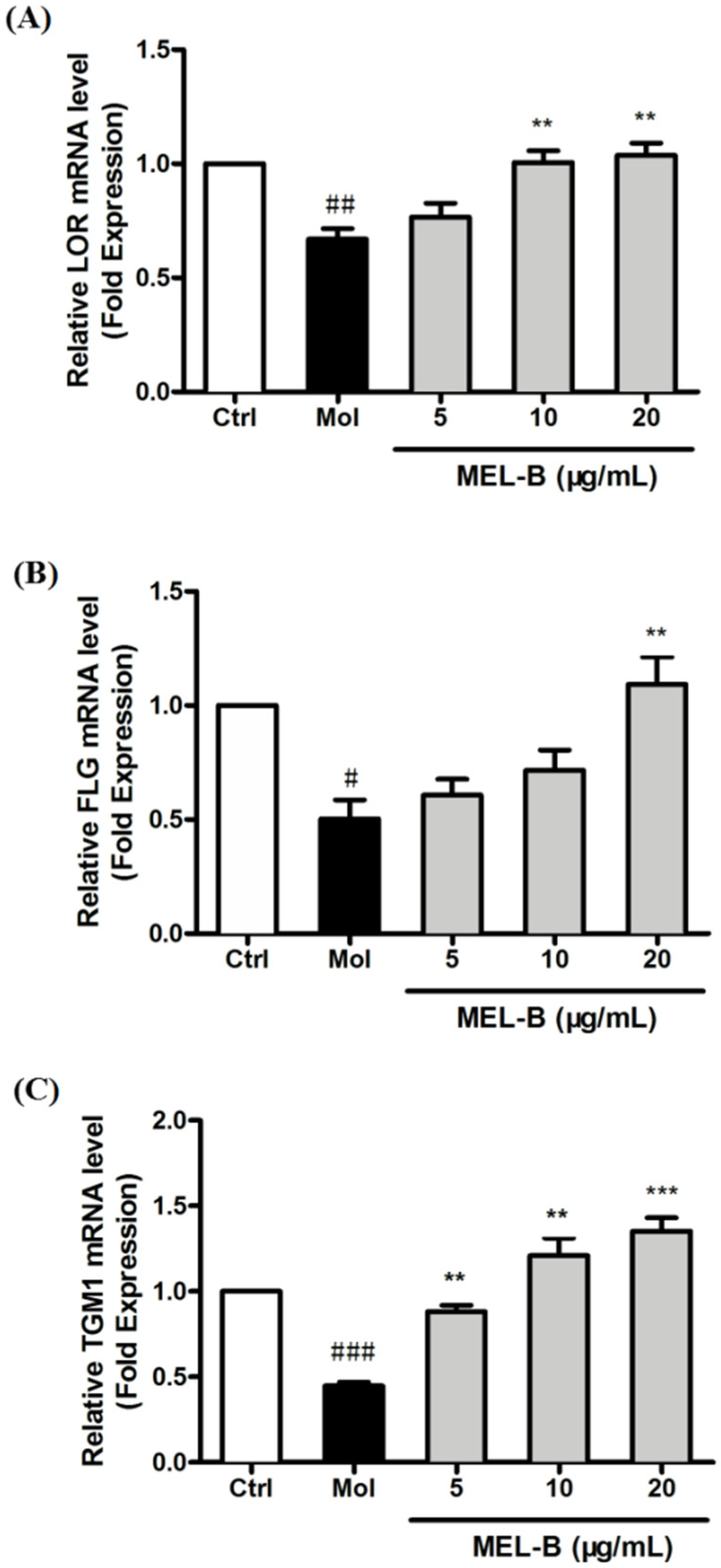
2.6. Increased mRNA Expression of Genes Associated with Skin Barrier in UVB-Induced HaCaT Cells after MEL-B Treatment
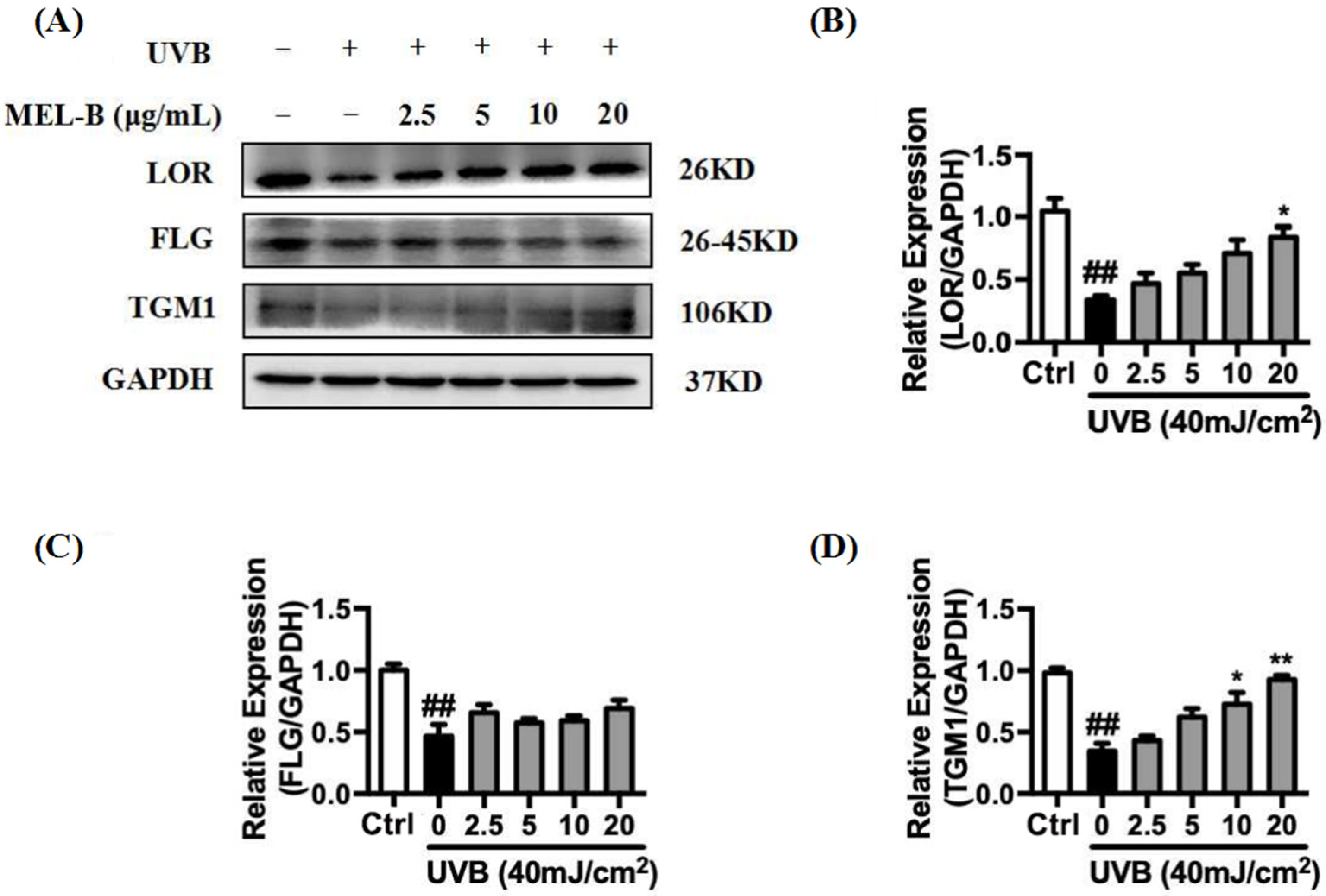
2.7. Upregulation of MEL-B Treatment on Expression Levels of LOR, FLG, TGM1 Protein in UVB-Induced HaCaT Cells
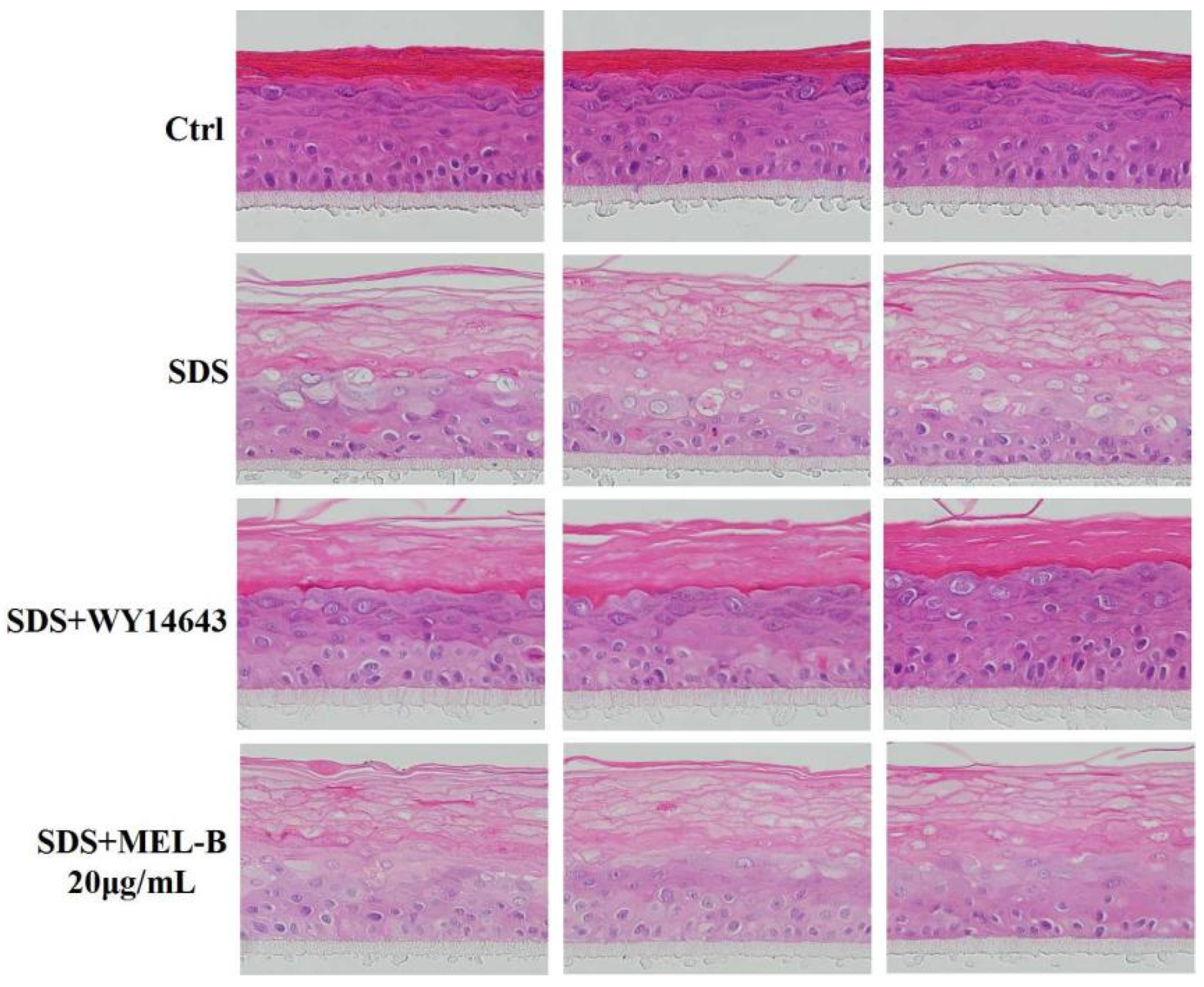
2.8. Protective Effects of MEL-B on Skin Barrier Damage as Assessed in SDS-Induced EpiKutis®3D Human Skin Cells
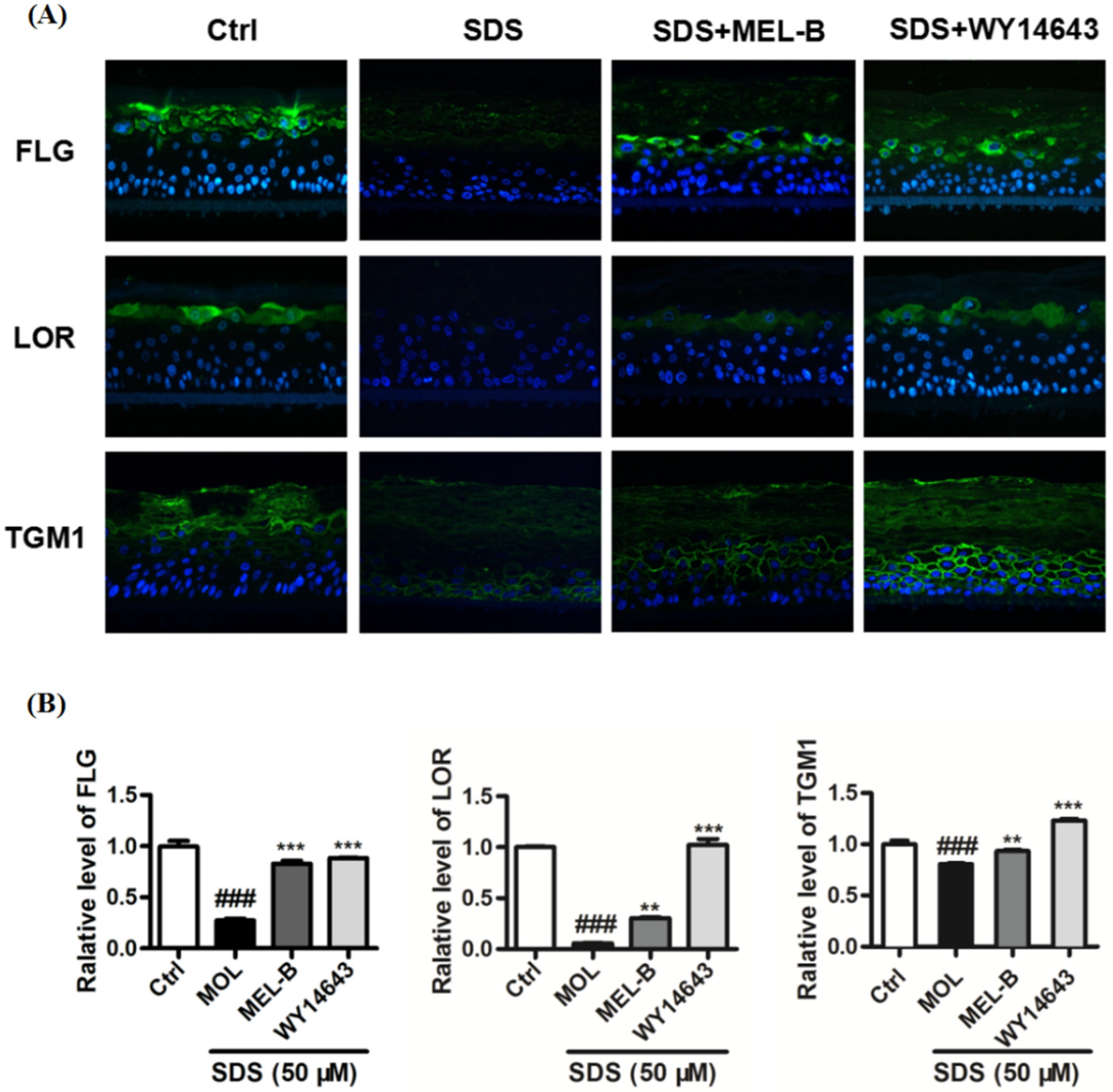
2.9. Effects of MEL-B Treatment on Expression Levels of LOR, FLG, and TGM1 in Cells of the SDS-Induced EpiKutis®3D Human Skin Cell Damage Model
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. General Experimental Procedures
4.2.1. Production of MELs
4.2.2. Preliminary Separation and Purification of Fermentation Broth
4.2.3. Separation and Purification of MELs by Silica Gel Column Chromatography
4.2.4. Quantification of MELs by High-Performance Liquid Chromatography (HPLC)
4.2.5. Structure Analysis of MELs by HRESIMS and NMR
4.3. Bioactivity Assay
4.3.1. Cell Culture
4.3.2. UVB Irradiation and Treatments
4.3.3. Cell Viability Assay
4.3.4. ELISA
4.3.5. Quantitative Real-Time PCR (RT-PCR) Analysis
4.3.6. Western Blotting
4.3.7. Immunohistochemistry
4.3.8. Immunofluorescence
4.3.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Hesse, M.; Zimek, A.; Weber, K.; Magin, T.M. Comprehensive analysis of keratin gene clusters in humans and rodents. Eur. J. Cell Biol. 2004, 83, 19–26. [Google Scholar] [CrossRef] [Green Version]
- Jung, Y.O.; Jeong, H.; Cho, Y.; Lee, E.O.; Jang, H.W.; Kim, J.; Nam, K.T.; Lim, K.M. Lysates of a probiotic, lactobacillus rhamnosus, can improve skin barrier function in a reconstructed human epidermis model. Int. J. Mol. Sci. 2019, 20, 4289. [Google Scholar] [CrossRef] [Green Version]
- Vávrová, K.; Henkes, D.; Strüver, K.; Sochorová, M.; Školová, B.; Witting, M.Y.; Friess, W.; Schreml, S.; Meier, R.J.; Schäfer-Korting, M.; et al. Filaggrin deficiency leads to impaired lipid profile and altered acidification pathways in a 3D skin construct. J. Investig. Dermatol. 2014, 134, 746–753. [Google Scholar] [CrossRef] [Green Version]
- Dai, X.; Utsunomiya, R.; Shiraishi, K.; Mori, H.; Muto, J.; Murakami, M.; Sayama, K. Nuclear IL-33 plays an important role in the suppression of filaggrin, loricrin, keratin 1, and keratin 10 by IL-4 and IL-13 in human keratinocytes. J. Investig. Dermatol. 2021, 141, 2646–2655. [Google Scholar] [CrossRef]
- Gao, P.S.; Rafaels, N.M.; Hand, T.; Murray, T.; Boguniewicz, M.; Hata, T.; Schneider, L.; Hanifin, J.M.; Gallo, R.L.; Gao, L.; et al. Filaggrin mutations that confer risk of atopic dermatitis confer greater risk for eczema herpeticum. J. Allergy Clin. Immunol. 2009, 124, 507–513. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.Y.; Lin, K.; Chen, Y.J.; Chiang, Y.J.; Ho, K.C.; Shen, L.F.; Song, I.W.; Liu, K.M.; Yang-Yen, H.F.; Chen, Y.J.; et al. Palmitoyl Acyltransferase Activity of ZDHHC13 Regulates Skin Barrier Development Partly by Controlling PADi3 and TGM1 Protein Stability. J. Investig. Dermatol. 2020, 140, 959–970. [Google Scholar] [CrossRef]
- D’ Orazio, J.; Jarrett, S.; Amaro-Ortiz, A.; Scott, T. UV radiation and the skin. Int. J. Mol. Sci. 2013, 14, 12222–12248. [Google Scholar] [CrossRef] [Green Version]
- Martinez, R.M.; Fattori, V.; Saito, P.; Melo, C.B.P.; Borghi, S.M.; Pinto, I.C.; Bussmann, A.J.C.; Baracat, M.M.; Georgetti, S.R.; Verri, W.A., Jr.; et al. Lipoxin A4 inhibits UV radiation-induced skin inflammation and oxidative stress in mice. J. Dermatol. Sci. 2018, 1811, 30201–30209. [Google Scholar] [CrossRef] [Green Version]
- Arutchelvi, J.I.; Bhaduri, S.; Uppara, P.V.; Doble, M. Mannosylerythritol lipids: A review. J. Ind. Microbiol. Biot. 2008, 35, 1559–1570. [Google Scholar] [CrossRef]
- Dubey, K.V.; Charde, P.N.; Meshram, S.U.; Shendre, L.P.; Dubey, V.S.; Juwarkar, A.A. Surface-active potential of biosurfactants produced in curd whey by Pseudomonas aeruginosa strain-PP2 and Kocuria turfanesis strain-J at extreme environmental conditions. Bioresour. Technol. 2012, 126, 368–374. [Google Scholar] [CrossRef]
- Shu, Q.; Niu, Y.W.; Zhao, W.J.; Chen, Q. Antibacterial activity and mannosylerythritol lipids against vegetative cells and spores of Bacillus cereus. Food Control 2019, 106, 106711. [Google Scholar] [CrossRef]
- Díaz De Rienzo, M.A.; Stevenson, P.; Marchant, R.; Banat, I.M. Antibacterial properties of biosurfactants against selected Gram-positive and -negative bacteria. FEMS Microbiol. Lett. 2016, 363, fnv224. [Google Scholar] [CrossRef] [Green Version]
- Nashida, J.; Nishi, N.; Takahashi, Y.; Hayashi, C.; Igarashi, M.; Takahashi, D.; Toshima, K. Systematic and stereoselective total synthesis of mannosylerythritol lipids and evaluation of their antibacterial activity. J. Org. Chem. 2018, 83, 7281–7289. [Google Scholar] [CrossRef]
- Coelho, A.L.S.; Feuser, P.E.; Carciofi, B.A.M.; de Andrade, C.J.; de Oliveira, D. Mannosylerythritol lipids: Antimicrobial and biomedical properties. Appl. Microbiol. Biotechnol. 2020, 104, 2297–2318. [Google Scholar] [CrossRef]
- Isakov, N. Protein kinase C (PKC) isoforms in cancer, tumor promotion and tumor suppression. Semin. Cancer Biol. 2018, 48, 36–52. [Google Scholar] [CrossRef]
- Kömüves, L.G.; Hanley, K.; Lefebvre, A.M.; Man, M.Q.; Ng, D.C.; Bikle, D.D.; Williams, M.L.; Elias, P.M.; Auwerx, J.; Feingold, K.R. Stimulation of PPARα promotes epidermal keratinocyte differentiation in vivo. J. Investig. Dermatol. 2000, 115, 353–360. [Google Scholar] [CrossRef] [Green Version]
- Wallmeyer, L.; Lehnen, D.; Eger, N.; Sochorová, M.; Opálka, L.; Kováčik, A.; Vávrová, K.; Hedtrich, S. Stimulation of PPARα normalizes the skin lipid ratio and improves the skin barrier of normal and filaggrin deficient reconstructed skin. J. Dermatol. Sci. 2015, 80, 102–110. [Google Scholar] [CrossRef]
- Tanaka, Y.; Uchi, H.; Furue, M. Antioxidant cinnamaldehyde attenuates UVB-induced photoaging. J. Dermatol. Sci. 2019, 96, 151–158. [Google Scholar] [CrossRef]
- Kang, M.C.; Yumnam, S.; Kim, S.Y. Oral intake of collagen peptide attenuates ultraviolet B irradiation-induced skin dehydration in vivo by regulating hyaluronic acid synthesis. Int. J. Mol. Sci. 2018, 19, 3551. [Google Scholar] [CrossRef] [Green Version]
- Hung, C.F.; Chen, W.Y.; Aljuffali, I.A.; Lin, Y.K.; Shih, H.C.; Fang, J.Y. Skin aging modulates percutaneous drug absorption: The impact of ultraviolet irradiation and ovariectomy. Age 2015, 37, 21. [Google Scholar] [CrossRef] [Green Version]
- Yang, G.; Seok, J.K.; Kang, H.C.; Cho, Y.Y.; Lee, H.S.; Lee, J.Y. Skin barrier abnormalities and immune dysfunction in atopic dermatitis. Int. J. Mol. Sci. 2020, 21, 2867. [Google Scholar] [CrossRef] [Green Version]
- Karim, N.; Phinney, B.S.; Salemi, M.; Wu, P.W.; Naeem, M.; Rice, R.H. Human stratum corneum proteomics reveals cross-linking of a broad spectrum of proteins in cornified envelopes. Exp. Dermatol. 2019, 28, 618–622. [Google Scholar] [CrossRef] [Green Version]
- Sandilands, A.; Sutherland, C.; Irvine, A.D.; McLean, W.I. Filaggrin in the frontline: Role in skin barrier function and disease. J. Cell Sci. 2009, 122, 1285–1294. [Google Scholar] [CrossRef] [Green Version]
- Debinska, A. New treatments for atopic dermatitis targeting skin barrier repair via the regulation of FLG expression. J. Clin. Med. 2021, 10, 2506. [Google Scholar] [CrossRef]
- Furue, M. Regulation of filaggrin, loricrin, and involucrin by IL-4, IL-13, IL-17A, IL-22, AHR, and NRF2: Pathogenic implications in atopic dermatitis. Int. J. Mol. Sci. 2020, 21, 5382. [Google Scholar] [CrossRef]
- Kitamoto, D.; Yanagishita, H.; Shinbo, T.; Nakane, T.; Kamisawa, C.; Nakahara, T. Surface active properties and antimicrobial activities of mannosylerythritol lipids as biosurfactants produced by Candida antarctica. J. Biotechnol. 1993, 29, 91–96. [Google Scholar] [CrossRef]
- Fukuoka, T.; Morita, T.; Konishi, M.; Imura, T.; Kitamoto, D. Characterization of new glycolipid biosurfactants, tri-acylated mannosylerythritol lipids, produced by Pseudozyma yeasts. Biotechnol. Lett. 2007, 29, 1111–1118. [Google Scholar] [CrossRef]
- Morita, T.; Fukuoka, T.; Imura, T.; Kitamoto, D. Formation of the two novel glycolipid biosurfactants, mannosylribitol lipid and mannosylarabitol lipid, by Pseudozyma parantarctica JCM 11752T. Appl. Microbiol. Biotechmol. 2012, 96, 931–938. [Google Scholar] [CrossRef]
- Spoeckner, S.; Wray, V.; Nimtz, M.; Lang, S. Glycolipids of the smut fungus Ustilago maydis from cultivation on renewable resources. Appl. Microbiol. Biotechnol. 1999, 51, 33–39. [Google Scholar] [CrossRef]
- Yamamoto, S.; Fukuoka, T.; Imura, T.; Yanagidani, S.; Kitamoto, D.; Kitagawa, M. Production of a novel mannosylerythritol lipid containing a hydroxy fatty acid from castor oil by Pseudozyma tsukubaensis. J. Oleo Sci. 2013, 62, 381–389. [Google Scholar] [CrossRef]
- Fukuoka, T.; Morita, T.; Konishi, M.; Imura, T.; Sakai, H.; Kitamoto, D. Structural characterization and surface-active properties of a new glycolipid biosurfactant, mono-acylated mannosylerythritol lipid, produced from glucose by Pseudozyma antarctica. Appl. Microbiol. Biotechnol. 2007, 76, 801–810. [Google Scholar] [CrossRef]
- Morita, T.; Konishi, M.; Fukuoka, T.; Imura, T.; Kitamoto, H.K.; Kitamoto, D. Characterization of the genus Pseudozyma by the formation of glycolipid biosurfactants, mannosylerythritol lipids. FEMS Yeast Res. 2007, 7, 286–292. [Google Scholar] [CrossRef] [Green Version]
- Kitamoto, D.; Morita, T.; Fukuoka, T.; Konishi, M.A.; Imura, T. Self-assembling properties of glycolipid biosurfactants and their potential applications. Curr. Opin. Colloid Interface Sci. 2009, 14, 315–328. [Google Scholar] [CrossRef]
- Worakitkanchanakul, W.; Imura, T.; Fukuoka, T.; Morita, T.; Sakai, H.; Abe, M.; Rujiravanit, R.; Chavadej, S.; Minamikawa, H.; Kitamoto, D. Phase behavior of ternary mannosylerythritol lipid/water/oil systems. Colloids Surf. B Biointerfaces 2009, 68, 207–212. [Google Scholar] [CrossRef]
- Yamamoto, S.; Morita, T.; Fukuoka, T.; Imura, T.; Yanagidani, S.; Sogabe, A.; Kitamoto, D.; Kitagawa, M. The moisturizing effects of glycolipid biosurfactants, mannosylerythritol lipids, on human skin. J. Oleo Sci. 2012, 61, 407–412. [Google Scholar] [CrossRef] [Green Version]
- Morita, T.; Kitagawa, M.; Yamamoto, S.; Sogabe, A.; Imura, T.; Fukuoka, T.; Kitamoto, D. Glycolipid biosurfactants, mannosylerythritol lipids, repair the damaged hair. J. Oleo Sci. 2010, 59, 267–272. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| MEL-A a | MEL-B a | MEL-C b | ||||
|---|---|---|---|---|---|---|
| Functional Groups | 1H NMR δ (ppm) | 13C NMR δ (ppm) | 1H NMR δ (ppm) | 13C NMR δ (ppm) | 1H NMR δ (ppm) | 13C NMR δ (ppm) |
| D-Mannose | ||||||
| H-1′ (C-1′) | 5.50, d | 99.35 | 5.50, dd | 99.39 | 5.50, dd | 99.14 |
| H-2′ (C-2′) | 5.06, dd | 68.56 | 4.91, dd | 68.74 | 5.10, dd | 68.70 |
| H-3′ (C-3′) | 5.24, t | 70.60 | 4.77, m | 71.23 | 5.18, t | 70.64 |
| H-4′ (C-4′) | 4.72, d | 65.90 | 3.98, m | 65.64 | 4.77, d | 66.19 |
| H-5′ (C-5′) | 3.69, m | 72.55 | 3.85, m | 74.58 | 4.04, m | 74.88 |
| H-6′ (C-6′) a | 4.19, dd | 62.40 | 3.81, m | 63.15 | 3.77, m | 61.42 |
| H-6′ (C-6′) b | 4.24, dd | 62.40 | 4.43, m | 63.15 | 3.79, m | 61.42 |
| meso-Erythritol | ||||||
| H-1 (C-1) | 3.76, m | 63.60 | 3.72, m | 63.63 | 3.73, m | 63.48 |
| H-2 (C-2) | 3.66, m | 71.86 | 3.66, m | 73.12 | 3.65, m | 73.10 |
| H-3 (C-3) | 3.72, m | 71.19 | 3.77, m | 71.93 | 3.75, m | 71.10 |
| H-4a (C-4a) | 3.84, dd | 72.38 | 3.56, m | 72.23 | 3.68, m | 71.99 |
| H-4b (C-4b) | 3.99, dd | 72.38 | 3.75, m | 72.23 | 3.54, m | 71.99 |
| Acetyl group(s) | ||||||
| -CH3 | 2.09, 2.03, s | (20.75, 20.68) | 2.13, s | 20.86 | 2.06, s | 20.73 |
| -C=O | (170.76, 169.46) | 171.67 | 170.21 | |||
| Acyl groups | ||||||
| -CH3 | 0.86, m | (14.12, 14.11) | 0.87, m | (14.12, 14.07) | 0.88, m | (14.13, 14.07) |
| -(CH2)n- | 1.23–1.31, brs | (22.67–31.88) | 1.24–1.26, brs | (22.61–31.94) | 1.25–1.29, brs | (22.68–31.90) |
| -CO-CH2- (C-2′) position | 2.42, m | 34.16 | 2.40, m | 34.11 | 2.43, m | 34.11 |
| -CO-CH2- (C-3′) position | 2.21, m | 34.03 | 2.29, m | 34.20 | 2.22, m | 34.05 |
| -C=O | (173.43, 172.73) | (173.54, 172.41) | (173.59, 172.79) | |||
| Gene | Sequence |
|---|---|
| LOR-F | 5′-GAGCTACGGAGGCGTCTCTA-3′ |
| LOR-R | 5′-AGAGTAGCCGCAGACAGA-3′ |
| FLG-F | 5′-GGTCTGGACGTTCAGGGTCT-3′ |
| FLG-R | 5′-GGATGTGGTGTGGCTGTGAT-3′ |
| TGM1-F | 5′-CCCCAAGAGACTAGCAGTGG-3′ |
| TGM1-R | 5′-AGACCAGGCCATTCTTGATG-3′ |
| GAPDH-F | 5′-GTGAAGGTCGGAGTCAACG-3′ |
| GAPDH-R | 5′-TGAGGTCAATGAAGGGGTC-3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jing, C.; Guo, J.; Li, Z.; Xu, X.; Wang, J.; Zhai, L.; Liu, J.; Sun, G.; Wang, F.; Xu, Y.; et al. Screening and Research on Skin Barrier Damage Protective Efficacy of Different Mannosylerythritol Lipids. Molecules 2022, 27, 4648. https://doi.org/10.3390/molecules27144648
Jing C, Guo J, Li Z, Xu X, Wang J, Zhai L, Liu J, Sun G, Wang F, Xu Y, et al. Screening and Research on Skin Barrier Damage Protective Efficacy of Different Mannosylerythritol Lipids. Molecules. 2022; 27(14):4648. https://doi.org/10.3390/molecules27144648
Chicago/Turabian StyleJing, Chenxu, Jiling Guo, Zhenzhuo Li, Xiaohao Xu, Jing Wang, Lu Zhai, Jianzeng Liu, Guang Sun, Fei Wang, Yangfen Xu, and et al. 2022. "Screening and Research on Skin Barrier Damage Protective Efficacy of Different Mannosylerythritol Lipids" Molecules 27, no. 14: 4648. https://doi.org/10.3390/molecules27144648