
Potential Beneficial Effects of Naringin and Naringenin on Long COVID—A Review of the Literature
1
Guangdong Engineering and Technology Research Center for Quality and Efficacy Re-Evaluation of Post-Market Traditional Chinese Medicine, State Key Laboratory of Biocontrol, Guangdong Provincial Key Laboratory of Plant Resources, School of Life Sciences, Sun Yat-sen University, Guangzhou 510275, China
2
Maoming Branch, Guangdong Laboratory for Lingnan Modern Agriculture, Maoming 525000, China
*
Author to whom correspondence should be addressed.
Microorganisms 2024, 12(2), 332; https://doi.org/10.3390/microorganisms12020332
Submission received: 9 January 2024
/
Revised: 29 January 2024
/
Accepted: 31 January 2024
/
Published: 4 February 2024
(This article belongs to the Special Issue Coronaviruses: Past, Present, and Future)
Abstract
:Coronavirus disease 2019 (COVID-19) caused a severe epidemic due to severe acute respiratory syndrome coronavirus-2 (SARS-CoV-2). Recent studies have found that patients do not completely recover from acute infections, but instead, suffer from a variety of post-acute sequelae of SARS-CoV-2 infection, known as long COVID. The effects of long COVID can be far-reaching, with a duration of up to six months and a range of symptoms such as cognitive dysfunction, immune dysregulation, microbiota dysbiosis, myalgic encephalomyelitis/chronic fatigue syndrome, myocarditis, pulmonary fibrosis, cough, diabetes, pain, reproductive dysfunction, and thrombus formation. However, recent studies have shown that naringenin and naringin have palliative effects on various COVID-19 sequelae. Flavonoids such as naringin and naringenin, commonly found in fruits and vegetables, have various positive effects, including reducing inflammation, preventing viral infections, and providing antioxidants. This article discusses the molecular mechanisms and clinical effects of naringin and naringenin on treating the above diseases. It proposes them as potential drugs for the treatment of long COVID, and it can be inferred that naringin and naringenin exhibit potential as extended long COVID medications, in the future likely serving as nutraceuticals or clinical supplements for the comprehensive alleviation of the various manifestations of COVID-19 complications.
1. Introduction
COVID-19 encompasses a range of illnesses resulting from the contraction of severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2). It is spread in the population mainly through droplets, but also via aerosol transmission [1], contamination transmission [2], and the fecal–oral route of transmission [3]. SARS-CoV-2 binds to the angiotensin-converting enzyme II (ACE2) receptor in the body via the S protein, which then invades human tissues and releases RNA via cytotoxicity [4]. The ACE2 receptor is mainly found in type II alveolar cells and is also found in many tissues, such as the heart, esophagus, kidneys, and bladder [5]. Therefore, SARS-CoV-2 invades the human body and causes a multi-tissue disease, including respiratory symptoms. SARS-CoV-2 has a high rate of transmission. The majority of patients experience acute symptoms within a span of 2–14 days following viral exposure. The symptoms are primarily fever, cough, sore throat, myalgia, and an altered sense of smell/taste [6,7]. In addition to cardiac disease [8], neurologic disease [9] and gastrointestinal disease are also seen in critical patients [10]. Most acute symptoms subside within 2–3 weeks of being infected with the virus, although subsequent research has revealed that certain individuals do not completely recuperate from their ailment, but instead, experience post-acute consequences of SARS-CoV-2 infection (PASC), commonly referred to as long COVID [11].
Long COVID is characterized by the presence of persistent, relapsing, new symptoms, or other health effects that manifest following acute SARS-CoV-2 infection [12]. Approximately half of the infected individuals worldwide will develop long COVID, especially in Asia (51%), Europe (44%), and the United States of America (31%), and this figure could be higher due to insufficient statistics and a lack of standardized scoring [13,14]. The symptoms of long COVID are more widespread and have a greater impact on human health and working ability [15]. Statistical analysis of the symptoms of COVID-19 patients one year after infection found that the main symptoms of long COVID include dizziness, fatigue, sleep difficulties, memory loss, anxiety and depression, hair loss, smell and taste disorder, decreased appetite, thirst, chronic cough, pulmonary fibrosis, palpitations, chest pain, diarrhea or vomiting, gastrointestinal disorders, diabetes, changes in sexual desire or capacity, joint and muscle pain, post-exertional malaise, abnormal movements, and thrombus formation [12,16] (Figure 1). Among them, fatigue, memory loss, multiorgan abnormalities, pulmonary fibrosis, heart damage, sleep difficulties, and anxiety and depression will continue unimproved for four months up to two years [17,18]. Furthermore, women and individuals with chronic underlying conditions are more susceptible to the detrimental effects of long COVID, and the intensity of symptoms can be affected by vaccination and the mutated strains of the virus [19]. Current research suggests that the main pathogenic mechanisms of long COVID are the persistence of the SARS-CoV-2 virus [20], microthrombosis [21], immune dysregulation [22], the disruption of mitochondrial function [23], the activation of pathogens such as Epstein–Barr virus (EBV) and human herpesvirus 6 (HHV6) [24], changes in the body microbiota [25], persistent inflammation [26], the activation of autoimmunity [27], brainstem signal transduction, and vagal signaling dysfunction [28,29].
At present, the primary treatment for long COVID is based on distinct symptoms, and there are no definitive medications and therapies for long COVID treatment [30]. However, a large number of studies have identified several potential therapeutic agents against long COVID in the natural constituents of medicinal plants, including the organic sulfur compound from garlic (Allium sativum L.) [31], curcumin from Curcuma longa [32], nigellone and α-hederin from Nigella sativ [33,34], phytocompound 6-gingerol from Zingiber officinale [35], and flavonoids from garlic, propolis, and honey [31,36]. In particular, natural and semi-synthetic flavonoids and neutrophil elastase inhibitors isolated from natural sources show great advantages in fighting SARS-CoV-2 or treating long COVID [37,38,39].
Naringin and naringenin are two of the most common flavanones found in citrus plants (Figure 2). They are known to cause bitterness in citrus fruits and have a wide range of biological functions. Naringin is the glycoside form of naringenin. After the oral ingestion of naringin or naringenin by the human body, naringin is converted to naringenin in the presence of intestinal microorganisms and absorbed in the intestinal epithelium [40]. The efficacy of naringin and naringenin is due to anti-inflammatory [41], anticancer [42], antioxidant properties [43,44], treatment of cardiovascular disease [45], antiviral [36] and immune-modulation effects [41], which may play an effective role in treating long COVID. The anti-inflammatory and antioxidant effects of naringenin can effectively counteract the cytokine storm induced by SARS-CoV-2, especially targeting and inhibiting interleukin(IL)-6, the main pro-inflammatory factor of COVID-19 [46]. Moreover, numerous molecular docking studies reveal that naringenin binds to viral spiking proteins, viral major proteases, host receptors, and host viral transport channels and has multiple antiviral effects against SARS-CoV-2 [47]. For instance, naringenin can influence the replication of the viral genome by attaching to SARS-CoV-2 macrodomain RNA polymerase (NSP3), SARS-CoV-2 RNA-dependent RNA polymerase (NSP12), and 3-chymotrypsin-like protease, as well as affect viral invasion by attaching to the ACE2 receptor to achieve an antiviral effect [48,49]. However, despite the abundance of evidence suggesting that naringin and naringenin can be more effective in treating various illnesses, there is yet to be a review of their combined potential in treating multiple coexisting conditions of long COVID. The purpose of this research was to assess the potential beneficial effects of naringin and naringenin in treating long COVID.
2. Therapeutic Potential of Naringin and Naringenin in Long COVID
In the following, we will discuss the pathogenesis of 11 common symptoms in patients with long COVID and make reasonable assumptions about the potential therapeutic role of naringin and naringenin in treating these symptoms (Figure 3).
2.1. Cognitive Dysfunction
Cognitive dysfunction is an extremely common persistent psychiatric manifestation after COVID-19 referred to as “brain fog”. It is characterized by attention/processing speed deficits, mainly in memory and executive functioning [50]. Fatigue, sleep disturbances, language disorders, and loss of smell/taste accompany it [51,52]. According to meta-statistics, cognitive dysfunction was found to be present in 22% of COVID-19 patients 19 weeks after diagnosis, and the symptom was continued in 19.7% at 9 months after infection [53,54]. The condition is seen in both mildly and severely ill patients and persists for long periods without relief, and the longer the duration of symptoms, the greater the cognitive impairment [55]. The main way that COVID-19 survivors experience cognitive decline is through the extended inflammation caused by SARS-CoV-2, which can increase blood–brain barrier permeability and activate microglia and astrocyte subtypes, leading to cellular stress and neuronal damage [56,57]. The invasion of SARS-CoV-2 also causes midbrain dopamine neuron senescence, which has been implicated in Parkinson’s disease [58]. The downregulation of new neuron formation caused by long-term inflammation leads to reduced synaptic plasticity in the hippocampus, resulting in memory loss in COVID-19 survivors [59]. Moreover, some of the pro-inflammatory factors are involved in mood regulation, leading to depression and other mood abnormalities in survivors [60]. In addition, SARS-CoV-2 directly infects the nervous system and causes neurological destruction mediated by neurological inflammation [61,62]. The virus directly damages the olfactory epithelium via neuropilin-1 and ACE2, thereby affecting the olfactory neural network and causing olfactory malfunction [63]. Patients have exhibited notable enhancements in IL-6, CD70, C-reactive protein (CRP), C-C Motif Chemokine Ligand 11 (CCL11), serotonin, and serum biomarkers of neuronal and gliotic degeneration in the blood, as well as alterations in the brain’s microstructure and functional brain integrity, as indicated by neuropsychological tests, blood tests, and diagnostic brain imaging [64,65,66].
Naringin and naringenin have been shown to provide protection and potentially offer therapeutic benefits against cognitive decline in neurological disorders like Alzheimer’s disease [67,68]. Therefore, they can be reasonably hypothesized to have an ameliorative effect on cognitive dysfunction after COVID-19. Naringin shows a protective effect on the nervous system, mainly through the modulation of glial cell activation and protection against nervous system stresses. Microglia and astrocytes are both essential for safeguarding neurons. Microglia have two activation phenotypes: the M1 type is pro-inflammatory and damages neurons, while the M2 phenotype exerts anti-inflammatory and neuronal-repair functions. Naringin can promote microglia activation toward the M2 type and inhibit its activation toward the M1 type by modulating the janus kinase/signal transd ucer and activator of transcription 3 (JAK/STAT3) signaling pathway [69]. Astrocytes are protective of neuronal cells [70]. Naringin regulates the expression of nuclear factor erythroid 2-related factor 2 (Nrf2), thereby enhancing neuronal protection by astrocytes [71]. Naringenin combination therapy attenuates the pro-inflammatory activation of astrocytes and significantly attenuates 3-nitro propionic acid-induced neuronal cell death [72]. Naringin and naringenin have been shown to protect the nervous system from oxidative stress and inflammation in various studies. Naringenin reduces the production of reactive oxygen species, modulates pro-inflammatory cytokines (IL-1β, IL-6, and (tumor necrosis factor)TNF-α) and anti-inflammatory cytokines (IL-10 and IL-4) to reduce inflammation, and enhances synaptic plasticity by increasing the expression of N-methyl-D-aspartate receptors associated with learning and memory [73]. Naringin significantly attenuates D-galactose, doxorubicin, and Bisphenol A-induced oxidative stress in the nervous system and improves cognitive performance after treatment [74,75,76]. Naringenin exerts a protective effect against lead damage to the nervous system by maintaining the antioxidant enzyme system (superoxide dismutase (SOD), catalase (CAT), and glutathione (GSH)) and inhibiting the elevation of inflammatory factors (NF-κB) and proapoptotic-related protein (Bcl-2 and caspase-3) [77].
Moreover, naringenin has depression-relieving effects. Depression disorders are linked to decreased levels of brain-derived neurotrophic factor (BDNF), which is controlled by the cyclic adenosine monophosphate (cAMP)-cAMP response element binding protein (CREB)-BDNF signaling pathway [78,79]. Naringenin reverses the reduction in BDNF expression induced by high cortisol levels and modulates the mitochondrial apoptotic pathway to inhibit hippocampal apoptosis [80]. Naringenin can potentially enhance the endocrine nervous system by controlling the levels of glucocorticoid receptors and monoamines in the hippocampus, resulting in an antidepressant impact [81]. Furthermore, naringenin and SARS-CoV-2 can cross the blood–brain barrier and enter brain tissue [73]. The combination of naringenin and naringin, which block primary viral proteases, decrease receptor activity, and attach to viral spiny proteins, could potentially lessen neurological harm [48,82].
2.2. Immune Dysregulation
SARS-CoV-2 invasion activates the body’s innate and adaptive immunity to clear the invading pathogen [83]. Patients with long COVID have exhibited changes in immune cells, increased levels of autoantibodies, and the reactivation of dormant viruses [27,84,85]. The long-term effects of COVID-19 on the patient’s immune system can be seen through alterations in the epigenetic and transcriptional makeup of monocytes, which can result in long-term inflammation, excessive autoimmune activity, and long-term consequences in the body [86]. Intermediate monocyte abnormalities and T-cell activation were found in recovering COVID-19 patients, which were associated with persistent endothelial cell activation and coagulation dysfunction [87,88]. Patients with long COVID exhibited a decline in cluster of differentiation (CD)4+ and CD8+ effector memory cells [22], and the expression of the pro-inflammatory cytokines interferon (IFN)-β and IFN-λ1 remained high during the 8 months after infection [89]. The persistence of SARS-CoV-2 also leads to a lack of dendritic cells in the host [90], along with a reduction in non-classical monocyte and lymphocyte subsets [91]. The persistent aberrant activation of immunity produces a broad spectrum of self-targeting antibodies and is hypothesized to be associated with different long COVID symptoms [92]. Long COVID patients exhibit abnormally high levels of functional autoantibodies targeting different G protein-coupled receptors, which are associated with persistent neurological and cardiovascular symptoms in patients [93]. SARS-CoV-2-induced autoimmunity causes immune blood disorders, antiphospholipid syndrome, systemic lupus erythematosus, vasculitis, acute arthritis, and Kawasaki-like syndromes [94].
Naringenin has a palliative effect on T cell-mediated autoimmune diseases [95,96]. Following antigen induction, CD4+ T cells differentiate into distinct subpopulations, encompassing pro-inflammatory T cells and anti-inflammatory regulatory T cells, which maintain equilibrium within healthy organisms [97]. Naringenin modulates the immune system’s response to autoantigens by influencing the growth and specialization of T cells, as well as cytokine signaling [98,99]. Naringenin effectively alleviates symptoms of rheumatoid arthritis in rats by modulating lymphocyte polarization, primarily through the reduction in T helper (Th)1 and Th17 cell differentiation and the reduction in IL-6 and TNF-α levels [99,100]. Similarly, naringenin alleviates symptoms of multiple sclerosis and systemic lupus erythematosus by inhibiting the growth and specialization of harmful pro-inflammatory T cells while maintaining the differentiation of anti-inflammatory subgroups [101,102]. Naringenin also modulates cellular inflammatory responses by regulating the transcription of inflammatory factors and enhancing the lysosomal degradation of cellular inflammatory factors [103,104].
2.3. Microbiota Dysbiosis
SARS-CoV-2 leads to long-term changes in the microbiota, with decreased gut microbial diversity, fewer beneficial commensal bacteria, and increased opportunistic pathogens in patients [105]. Viral invasion dysregulates gut ecology by activating immunity, deregulating ACE2 expression, disrupting the intestinal barrier, and directly infecting bacteria [106,107]. Moreover, the antibiotics administered during treatment can cause more harm to the patient’s intestinal flora [108]. Furthermore, COVID-19 patients exhibited noteworthy alterations in the lung [109], oral [110], and nasopharyngeal microbiomes [111]. Changes in the microbiome persist for an extended period, and patients do not return to normal microbial abundance after 6 months of rehabilitation therapy [112]. Gut microbes remained significantly different from healthy controls after 3 months of recovery and showed a correlation with persistent symptoms of long COVID [113]. Changes in the composition and abundance of a patient’s gut microbes also affect their susceptibility to long COVID and complication severity [25]. These changes can have a serious impact on the health of the survivor, causing an inflammatory immune response in the body, changes in the levels of basal metabolite levels (e.g., amino acids, carbohydrates, and neurotransmitters), and clinical gastrointestinal symptoms [114]. Gut microflora dysbiosis can have neurological and pulmonary effects through cytokines and metabolites [115], which are mediated through the gut–lung and brain–gut axes [116,117].
Naringin and naringenin have been found to have beneficial regulatory effects on the composition and metabolism of gut flora [118,119]. The dietary intake of naringin and naringenin helps to control the intestinal microenvironment [120]. Naringin and naringenin can directly regulate the intestinal microbiota and its metabolism. Naringin significantly reduces the abundance of gastrointestinal disease-related bacteria (e.g., Lachnoclostridium and Bilophila) and enhances probiotic content in experiments [121]. Naringenin can regulate the growth and gene expression pattern of intestinal commensal microorganisms through interaction and activate genes related to cellular metabolism [122]. In addition, gut barrier dysfunction caused by microbiota dysbiosis may increase bacterial ectopia and contribute to host immune destabilization [123]. Naringin has been shown to reduce inflammation-associated protein expression and colonic damage caused by dextran sulfate sodium, as well as improve colonic barrier dysfunction [124].
2.4. Myalgic Encephalomyelitis/Chronic Fatigue Syndrome (ME/CFS)
ME/CFS is a chronic multisystem disease that often follows infectious diseases and may be associated with chronic viral infections such as human herpesviruses, human parvovirus B19V, and enteroviruses [125]. Patients with long COVID often experience persistent fatigue, myalgia, post-exertional malaise, insomnia, and exercise intolerance, with symptoms resembling the diagnostic criteria for ME/CFS [126]. Although there are no studies directly proving that acute COVID-19 triggers ME/CFS, several studies have identified multiple overlapping or similar symptoms between long COVID and ME/CFS [127]. COVID-19 also greatly increases the number of patients with ME/CFS [128], and bioinformatics analysis has identified a common network of genetic interactions between the two [129]. According to the diagnostic criteria for ME/CFS, the overall prevalence of ME/CFS was as high as 43% among patients with long COVID, with fatigue, malaise on exertion, and insomnia representing the main symptoms [130]. The invasion of SARS-CoV-2 significantly impacts both mitochondrial function and reserves, and the SARS-CoV-2 membrane protein causes mitochondrial apoptosis in epithelial cells [131]. Virus-induced decreases in mitochondrial membrane potential [132] and the downregulation of nuclear-encoded mitochondrial genes [133] have been observed in patients. In addition, the infiltration of amyloid-containing deposits and skeletal muscle injuries have also been observed in patients with long COVID [134]. The pathogenesis of ME/CFS involves neuroinflammation, redox imbalance, mitochondrial dysfunction, autoantibodies, and autonomic dysfunction [135,136,137].
The treatment of ME/CFS is still unclear and there are no effective treatments. Most treatments are based on the clinical condition and include central nervous system drugs, antiviral drugs, immunomodulators, analgesics, and nutritional supplements [138]. Fatigue and post-exercise discomfort may be due to reduced energy sources, mitochondrial dysfunction, and redox imbalance. Serum matrix metalloproteinase 9 (MMP-9) and muscle damage-associated lactate dehydrogenase (LDH) are associated with muscle fatigue. By stabilizing redox and elevating blood glucose levels, naringin effectively diminishes MMP-9 and LDH concentrations, thereby augmenting energy sources. This prolongs the duration of fatigue-inducing exercise and relieves post-exercise discomfort [139,140]. Moreover, naringenin can exert anti-fatigue effects by participating in the promotion of testosterone secretion [141]. Additionally, naringin can help to stabilize mitochondrial membrane potential, preserve mitochondrial integrity, and sustain mitochondrial complex activity [142,143], all of which can help to reduce mitochondrial dysfunction. Naringenin also protects neuronal mitochondrial function by activating the transcription factor Nrf2 [144]. In addition, naringin and naringenin are important redox regulators. Naringin significantly attenuated antigen-induced oxidative stress and reduced TNF-α levels in fatigued mice [145] while reinstating oxidative stress markers in rodent brains [146].
2.5. Myocarditis
Certain individuals who have survived the initial stages of COVID-19 experience enduring cardiac impairment following their recovery, ultimately resulting in myocarditis [147]. The clinical signs of myocarditis are heterogeneous and non-specific, typically consisting of chest pain, arrhythmias, generalized fatigue, dyspnea, and tachycardia [148]. The analysis of cardiac symptoms among COVID-19 survivors revealed that 9.79 percent displayed indications of chest pain, whereas 8.22 percent presented with arrhythmias [149]. Within a span of 2–3 months following SARS-CoV-2 infection, 19% of patients developed persistent symptoms of myocarditis [150]. One year after rehabilitation, patients had a significant 4.16-fold increased risk of myocarditis [151] and poorer clinical outcomes, with a mortality rate of 1.36% to 5% [152]. The direct pathogenic mechanism of myocarditis in long COVID is the direct interaction of SARS-CoV-2 with ACE2 receptors in cardiomyocytes and pericytes, causing immune dysregulation. Subsequently, immune dysregulation leads to cardiomyocyte injury and the release of inflammatory factors, such as IL-2, IL-6, IL-7, TNF-a, IFN-α/β, C-X-C motif cytokine 10 (CXCL10), and C-C motif ligand 2 (CCL2), which, in turn, induces persistent low-level inflammation [153]. Indirect pathogenic mechanisms include hypoxemia caused by the cytokine storm [154], high levels of antiheart autoantibodies caused by CD4+ T-lymphocytes [155], and cardiac mitochondrial dysfunction [156]. Myocardial injury can be identified by the presence of markers such as myoglobin, troponin, creatine kinase-MB, IL-6, LDH, and N-terminal pro-b-type natriuretic peptide [157].
Naringin and naringenin have been shown to have cardioprotective properties [45]. First, the anti-inflammatory properties of naringin can drastically reduce the inflammatory factors associated with cardiovascular injury, such as NF-κB, IL-6, IL-1β, and TNF-α, and suppress the inflammatory response [158]. Furthermore, the antioxidant effects of naringin can scavenge free radicals and increase the activity of antioxidant enzymes (superoxide dismutase and catalase) and GSH levels, effectively protecting the heart mitochondria from damage, thereby reducing DOX-induced apoptosis and vacuolization in cardiomyocytes [67]. In addition, naringin has the potential to alter the cellular channel currents in mouse ventricular myocytes, consequently exerting antiarrhythmic effects [159].
2.6. Pulmonary Fibrosis
Pulmonary fibrosis is one of the serious sequelae of COVID-19. It is estimated that 19% of patients recovering from COVID-19 have residual lung abnormalities [160]. The lungs are the primary target organ for viral invasion. SARS-CoV-2 causes severe lung damage, ultimately leading to post-COVID-19 pulmonary fibrosis (PCPF). Radiological manifestations of pulmonary fibrosis usually include bilateral lung infiltrates, “ground glass” opacity, and “honeycomb” lungs [161]. Its cellular and molecular characteristics include a decrease in lymphocytes and an increase in CRP and IL-6 [162]. Multiple studies have shown that the prevalence of PCPF among COVID-19 survivors is more than 9.3% [163]. Alveolar injury caused by SARS-CoV-2 leads to the secretion of pro-fibrotic and pro-inflammatory cytokines by alveolar macrophages and type 2 alveolar epithelial cells (AEC), including IFN-γ, transforming growth factor (TGF)-β, and IL-6 and IL-17. Cytokines induce myofibroblast differentiation by activating the WNT/β-catenin and YAP/TAZ pathways, ultimately leading to the combined effects of pulmonary fibrosis [164,165].
Naringin and naringenin are effective inhibitors of pulmonary fibrosis. The main target of current anti-pulmonary fibrosis therapy is the TGF-β/Smad pathway [166]. It has been shown that naringin inhibits cellular fibrosis by suppressing TGF-β overexpression and reducing downstream regulatory factor phosphorylation [167]. Moreover, the anti-inflammatory and antioxidant effects of naringin and naringenin can be used against pulmonary fibrosis [168]. Naringin has been shown to significantly reduce the infiltration of inflammatory cells induced by lipopolysaccharide and decrease the production of macrophage nitrogen monoxide (NO) and IL-6 [169]. In critically ill COVID-19 patients, naringenin also showed excellent IL-6 inhibition compared to synthetic monoclonal antibodies [46]. In a paraquat-induced acute lung injury model, naringin not only decreased the production of the inflammatory cytokines TNF-α and TNF-β1 but also inhibited oxidative stress by activating the expression of antioxidant enzymes (superoxide dismutase, glutathione peroxidase, and heme oxygenase 1) and regulated collagen formation by modulating the ratio of tissue inhibitors of metalloproteinases-1 (TIMP-1) to MMP-9, thereby preventing lung fibrosis [170]. Furthermore, naringin modulates the activating transcription factor 3/PTEN-induced kinase 1 (ATF3/PINK1) pathway and enhances mitophagy in lung tissues, resulting in the alleviation of bleomycin-induced idiopathic pulmonary fibrosis [171]. Naringenin also protects against cigarette-induced lung injury by regulating miRNAs in extracellular vesicles [172].
2.7. Cough
Coughing is the most common symptom of COVID-19 sequelae and is present in approximately 23% to 57% of COVID-19 survivors in all countries, and is highly prevalent in both mild and severe cases [173,174]. Cough can persist for months, with a prevalence of up to 2.5% even after one year of recovery [175], making it a major source of distress for patients. SARS-CoV-2 may induce chronic coughing through neuroinflammatory and neuroimmune mechanisms. Viral invasion causes the release of neuroinflammatory mediators (IL-1β, TNF, IFN, adenosine triphosphate (ATP)) and, in turn, activates vagal sensory neurons via transient receptor potential (TRP) channels. Sensory neurons release a variety of neuropeptides (calcitonin gene-related peptide, substance P, and neurokinin A), eventually leading to hypersensitivity of the cough pathway [176]. In addition, the underlying mechanisms of chronic cough may also be related to post-infectious lung abnormalities, cough underlying disease, upper airway cough syndrome, or cough-variant asthma (CVA) [177,178,179].
The current treatment of chronic coughing focuses on suppressing the cough reflex, reducing airway inflammation, drying and relieving coughing, and resolving phlegm to relieve coughing [180,181]. Naringin and naringenin have long been shown to have positive antitussive effects [182]. Naringenin can reduce inflammation caused by cigarette smoke in mice with chronic obstructive pulmonary disease by inhibiting the production of the pro-inflammatory factors IL-8, TNF-α, and MMP9, as well as the NF-κB pathway [183]. In an experimental CVA model, naringin has been shown to reduce irritation-induced cough and suppress the growth of airway-inflammatory factors (IL-4, IL-5, and IL-13) and leukocytes [184]. Moreover, naringin has been shown to significantly attenuate cigarette smoke-induced airway neurogenic inflammation by reducing substance P and NK-1 receptors [185]. Furthermore, naringin can be used in asthma treatment by promoting the proliferation of AECs through the bitter taste receptor (TAS2R)-related signal pathway, thereby inducing the relaxation of airway smooth muscle cells [186].
2.8. Diabetes
The occurrence of new-onset diabetes mellitus can be attributed to acute infection with SARS-CoV-2, and complications such as diabetic ketoacidosis (DKA) and hyperglycemic hyperosmolar syndrome are highly prevalent [187,188]. COVID-19 survivors have a 64–66% higher risk of developing diabetes than uninfected individuals [189]. The risk of developing diabetes in the first three months after COVID-19 infection was up to 95%, with a higher risk of developing type 2 diabetes (T2D) than type 1 diabetes (T1D) (70% and 48%, respectively) [190]. The data indicated that during the initial year of the COVID-19 outbreak, there was a notable surge of 9.5% and 25% in the occurrence of new pediatric T1Ds and DKAs, correspondingly, alongside a substantial rise in blood glucose levels [191]. A follow-up study found that most patients with new-onset T2D were cured 3 months after discharge from the hospital. In contrast, the remaining 37% of patients with T2D were diagnosed with persistent diabetes mellitus [192]. COVID-19-related abnormalities in glucose metabolism also recovered after one year [193]. On this bases, diabetes was also associated with serious illness, hospitalization, and death in COVID-19, so a bidirectional causal relationship between COVID-19 and diabetes was hypothesized [194]. Both COVID-19 patients and survivors exhibit insulin resistance and beta-cell dysfunction, which endure even after recuperation from the illness [195].
The pathogenesis of new-onset diabetes is mainly related to pancreatic autoimmunity, pancreatic injury, pro-inflammatory cytokine storms, the triggering of steroid drugs, and underlying diabetic activation of the body. T1D is an autoimmune disease, and the invasion of SARS-CoV-2 leads to autoimmune hyperactivation in the body [196]. Additionally, researchers have found pancreatic damage in some COVID-19 patients [197]. Thus there has been a hypothesis suggesting that ACE2 exhibits elevated expression levels in exocrine glands and pancreas islets [198] when SARS-CoV-2 enters them to induce pancreatic β-cell dysfunction or apoptosis, hindering insulin signaling [199]. T2D is mainly caused by short-term treatment and inflammation. Steroids, a widely prescribed medication for COVID-19, can potentially impact insulin sensitivity; nevertheless, the discontinuation of steroid therapy may lead to the recurrence of new-onset diabetes [200]. The inflammatory factor TNF has also been recognized as a common pathogenic target of COVID-19 and T2D [201].
Naringin and naringenin may exert antidiabetic potential by attenuating pancreatic β-cell damage or activating their proliferation. Naringenin protects pancreatic β-cells from streptozotocin (STZ)-induced immune stress by activating Nrf2, leading to a substantial decrease in blood glucose levels and restoring normal insulin levels [202]. Naringin inhibits mitochondria-mediated and death receptor-mediated apoptosis in pancreatic β cells [203]. Forkhead box M1 (FoxM1) transcription factor affects pancreatic adult beta cell proliferation, and naringin increases beta cell mass and treats diabetes by upregulating FoxM1 [204]. Naringin and naringenin also showed significant ameliorative effects on insulin resistance. In the an STZ- and a nicotinamide-induced T2D model, naringin and naringenin enhanced insulin secretory responses and the expression of insulin receptors and their sensitizers [205]. In T2D patients, naringin was found to improve glucose metabolism, increase residual insulin secretion, and stimulate glycogen synthesis; however, absolute insulin deficiency prevented it from regulating glucose levels in T1D patients, yet it could reverse T1D-induced DKA [206]. Furthermore, naringin and naringenin regulate carbohydrate and lipid metabolism. Naringin modulates the activity of glycolysis-related enzymes and regulates intestinal carbohydrate absorption [207]. Naringenin can reduce metabolic disorders by modulating immune-related inflammatory factors and inhibiting the infiltration of inflammatory cells into adipose tissue [208]. In addition, naringenin has a mitigating effect on the complications of diabetes. Naringenin, when used in conjunction with insulin, modulates matrix metalloproteinases to reduce neuropathic pain caused by diabetes [209]. Naringenin inhibits high-glucose-induced vasculopathy by downregulating the hyperproliferation and migration of vascular smooth muscle cells [210].
2.9. Pain
COVID-19 patients often have accompanying pain, including headache, musculoskeletal pain, and testicular pain, during the acute infection period and after rehabilitation [211,212]. Although the majority of pain symptoms subside two months after recovery, 10% of COVID-19 survivors still experience persistent musculoskeletal muscle pain, and the length of the pain does not correlate with the intensity of COVID-19 [213]. The post-infection follow-up of COVID-19 survivors unveiled a range of symptoms, encompassing general pain (13.40%), joint pain (28.25%), muscle pain (13.30%), headache (9.50%), and chest pain (12.12%) [214]. Pain development is mainly associated with direct action through ACE2 receptors [215], the inflammatory cytokine storm [216], and the driving effect of prostaglandins [217]. ACE2 receptors are abundant in skeletal muscle and other organs, resulting in viral harm to the muscles, and their neurophilicity can also lead to neuronal damage, resulting in pain [218]. The virus triggers an extended inflammatory reaction, resulting in heightened hyperexcitability of the central nervous system, thereby exacerbating pain [219]. COVID-19 patients experiencing headache symptoms exhibited elevated concentrations of inflammatory molecules and harmful molecules (high-mobility group protein B1 (HMGB1), NOD-like receptor thermal protein domain-associated protein 3 (NLRP3), and IL-6), potentially contributing to the initiation of trigeminal activation [220,221].
Naringin and naringenin have effective analgesic effects. Naringin and naringenin can attenuate neuronal stimulation directly by inhibiting the inflammatory response. Naringin attenuates iodoacetate-induced osteoarthritis pain by inhibiting the secretion of adrenaline and pro-inflammatory factors (IL-6, NO, and TNF-α) [222]. The administration of naringenin in a rat model of neuropathic pain resulted in the inhibition of glial cell activation caused by spinal nerve ligation injury, as well as a reduction in elevated levels of inflammatory factors (TNF-α, IL-1β, and monocyte chemotactic protein 1) [223]. Moreover, naringin and naringenin can directly modulate the damage perception pathway to achieve analgesic effects. Transient receptor potential vanilloid member 1 (TRPV1) is associated with noxious temperature sensation and inflammatory pain, and its antagonists have been considered potential analgesics [224,225,226]. By interacting with TRPV1, naringin suppresses nervous system hyperexcitability and potentially mitigates nerve pain by inhibiting oxidative stress [227]. Transient receptor potential melastatin-3 (TRPM3) ion channels are involved in organismal injury perception and are expressed on somatosensory neurons. Naringenin can effectively block TRPM3 channels in vivo and in vitro, thus supporting its analgesic effect [228]. Moreover, naringenin inhibits superoxide anion-induced inflammatory pain by activating the NO-cGMP-PKG-KAP signaling pathway and reducing nociceptive cytokine expression [229].
2.10. Reproductive Dysfunction
The acute infection phase of COVID-19 leads to reproductive dysfunction in both genders, including erectile dysfunction, orchitis, reduced testosterone levels, ovarian dysfunction, and menstrual changes [230,231]. After 3.8 months of recovery, 35.9% of men and 27.7% of women were found to have sexual dysfunction, with different mechanisms in both cases [232]. A continued high prevalence of erectile dysfunction in men was observed 3 months after COVID-19 recovery [233]. The impact of long COVID on women’s reproductive health is mainly characterized by menstrual irregularities, gonadal dysfunction, and fertility problems [234]. Both ACE2 and transmembrane serine protease 2 (TMPRSS2) are highly expressed in the gonads, with higher expression in male gonads and detectable SARS-CoV-2 infection in the testis [235]. Therefore, it is hypothesized that the gonads, especially the testes, are susceptible to SARS-CoV-2 infection and damage. Male long COVID patients with coagulation abnormalities and endothelial damage also experience testicular inflammation [236]. Testicular injury, oxidative stress by reactive oxygen species [237], and decreased testosterone due to dysfunction of the hypothalamic–pituitary–gonadal axis [238] can lead to erectile dysfunction [239]. Moreover, the autoimmune inflammatory response of the body can also cause a decrease in sperm quality [240].
Naringenin also has an attenuating effect on testicular and sperm damage. Naringenin can reduce the harmful effects of bisphenol A on the testes by suppressing oxidative stress and mitochondrial apoptosis [241]. Naringenin has been shown to reduce the harmful effects of antiretroviral medication on the male reproductive system of rats while preserving the normal physical structure of the testes and sperm viability [242]. Furthermore, naringenin has a protective effect on the ovary and can alleviate polycystic ovary syndrome. In vivo naringin treatment in rats inhibits steroidogenic enzymes and consequently controls ovulation, thereby restoring ovarian morphology and cystic follicle levels in patients with polycystic ovary syndrome [243].
2.11. Thrombus Formation
Endothelial damage caused by SARS-CoV-2 can lead to coagulation dysfunction in certain patients, ultimately resulting in the development of long-term concomitant thrombosis [244]. Individuals afflicted with long COVID face an elevated likelihood of experiencing arterial thromboembolic events and venous thromboembolic events. They are susceptible to acute pulmonary embolism (PE), deep vein thrombosis (DVT), myocardial infarction, and acute respiratory distress syndrome (ARDS) [245,246]. COVID-19 survivors had a significantly higher incidence of thrombophilia, PE (2.5–6.3%), and DVT (1.2–6.4%), which was 2 to 3 times higher than in uninfected individuals [247]. Long COVID-induced thrombosis can be caused by various factors, such as damage to the endothelium, the abnormal production of fibrin during platelet aggregation, and dysregulation of the immune system [248]. SARS-CoV-2 invasion can promote endothelial damage and dysfunction through direct interaction with ACE2 receptors [244]. Its spiny proteins can also interact with platelets and fibrin, inducing the formation of fibrin-like microclots, which are difficult to hydrolyze [249]. Moreover, viral invasion inducing a storm of inflammatory factors (IL-1β, IL-6, and TNF) activates exogenous coagulation pathways and promotes the inhibition of anticoagulant routes [250]. In addition, the persistence of the virus in the outer vesicles, the formation of autoantibodies, and chronic hypoxia can lead to long-term coagulation disorders [251]. Patients with long COVID who displayed coagulation abnormalities exhibited increased levels of hepatocyte growth factor (HGF), IL-6, and D-dimer [252].
Naringin can exert its antithrombotic effect by protecting endothelial cells, inhibiting platelet activation, and inhibiting thrombin activity. Naringin has been shown to reduce inflammation and alter the permeability of endothelial cells, thus mitigating endothelial dysfunction [169]. Naringin protects endothelial cell function by upregulating NO bioavailability [253], decreasing levels of inflammatory factors (IL-1β, IL-6, and IL-18), and reversing YAP downregulation [254]. Naringin inhibits excessive autophagy in endothelial cells by activating the PI3K-Akt-mTOR pathway [255]. Moreover, naringenin can impede platelet aggregation. Naringenin has a structure-dependent inhibitory effect on platelet function in both whole blood and plasma [256], and can also inhibit platelet activation by targeting the PI3K/Akt pathway, preventing FeCl3-induced carotid artery thrombosis and vascular occlusion and displaying effective antithrombotic properties [257]. In addition, research on molecular docking demonstrated that naringenin is strongly attracted to thrombin and can attach itself to the active core of thrombin, potentially impeding its activity [258]. Protein disulfide isomerase (PDI) in plasma is involved in the conformational formation of coagulation-associated proteins, and naringin can also affect thrombus formation and stabilization by binding to PDI and inducing conformational changes [259].
3. Conclusions and Prospects
This review addresses the pathogenesis and statistical status of eleven complications of long COVID and specifically addresses the therapeutic potential of naringin and naringenin in long COVID (Table 1). The clinical symptoms and biomarkers of long COVID have been widely reported, and long COVID can indeed have serious long-term effects on human health [260,261]. There are no definitive medications that directly treat multiple long COVID complications [262], and the treatment approach for long COVID is still based solely on clinical symptoms and the use of nutraceuticals and probiotics to improve symptoms [263]. Therefore, treating patients with multiple syndromes requires a combination of drugs. Naringin and naringenin, as important constituents of medicinal plants, are characterized by strong safety, multiple targets, and few side effects, with excellent performance in treating various diseases. They demonstrate protection against biotic stress by inducing a hormonal dose response in various cell models [264]. Moreover, it is worth noting that nano-preparations of natural products such as naringenin can improve their bioavailability and drug targeting after oral administration [265]. We enumerate the therapeutic and palliative effects of naringin and naringenin on conditions with symptoms or pathogenic mechanisms similar to those of long COVID, which are based on their anti-inflammatory, antimicrobial, antiviral, anti-free radical, cardiovascular-protection, microbiota-modulation, and neuron-protection effects. The purpose of this review is to provide a theoretical foundation for the use of naringin and naringenin as potential treatments for long COVID. Further research is necessary to ascertain the true efficacy and appropriate dosage of naringin and naringenin in treating long COVID, as well as conduct thorough follow-up clinical trials.
Author Contributions
Conceptualization, S.L. and P.L.; methodology, M.Z.; validation, Y.W., P.L., and H.W.; investigation, S.L.; resources, P.L.; writing—original draft preparation, S.L.; writing—review and editing, M.Z.; visualization, H.W.; supervision, W.S.; project administration, P.L.; funding acquisition, W.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Natural Science Foundation of Guangdong Province, grant number 2023A1515011953, the Open Competition Program of Ten Major Directions of Agricultural Science and Technology Innovation for the 14th Five-Year Plan of Guangdong Province, grant number 2022SDZG07, and the Research Fund of Maoming Branch, Guangdong Laboratory for Lingnan Modern Agriculture, grant number 2022ZD006.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Yu, I.T.S.; Li, Y.; Wong, T.W.; Tam, W.; Chan, A.T.; Lee, J.H.W.; Leung, D.Y.C.; Ho, T. Evidence of Airborne Transmission of the Severe Acute Respiratory Syndrome Virus. N. Engl. J. Med. 2004, 350, 1731–1739. [Google Scholar] [CrossRef]
- Cai, J.; Sun, W.; Huang, J.; Gamber, M.; Wu, J.; He, G. Indirect Virus Transmission in Cluster of COVID-19 Cases, Wenzhou, China, 2020. Emerg. Infect. Dis. 2020, 26, 1343–1345. [Google Scholar] [CrossRef] [PubMed]
- da Silva, F.A.F.; de Brito, B.B.; Santos, M.L.C.; Marques, H.S.; da Silva Junior, R.T.; de Carvalho, L.S.; de Sousa Cruz, S.; Rocha, G.R.; Santos, G.L.C.; de Souza, K.C.; et al. Transmission of Severe Acute Respiratory Syndrome Coronavirus 2 via Fecal-Oral: Current Knowledge. World J. Clin. Cases 2021, 9, 8280–8294. [Google Scholar] [CrossRef]
- Zhou, P.; Yang, X.-L.; Wang, X.-G.; Hu, B.; Zhang, L.; Zhang, W.; Si, H.-R.; Zhu, Y.; Li, B.; Huang, C.-L.; et al. A Pneumonia Outbreak Associated with a New Coronavirus of Probable Bat Origin. Nature 2020, 579, 270–273. [Google Scholar] [CrossRef]
- Beyerstedt, S.; Casaro, E.B.; Rangel, É.B. COVID-19: Angiotensin-Converting Enzyme 2 (ACE2) Expression and Tissue Susceptibility to SARS-CoV-2 Infection. Eur. J. Clin. Microbiol. Infect. Dis. 2021, 40, 905–919. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Ahmad Farouk, I.; Lal, S.K. COVID-19: A Review on the Novel Coronavirus Disease Evolution, Transmission, Detection, Control and Prevention. Viruses 2021, 13, 202. [Google Scholar] [CrossRef]
- World Health Organization. Operational Planning Guidance to Support Country Preparedness and Response. Available online: https://www.who.int/publications-detail-redirect/draft-operational-planning-guidance-for-un-country-teams (accessed on 20 December 2023).
- Wang, D.; Hu, B.; Hu, C.; Zhu, F.; Liu, X.; Zhang, J.; Wang, B.; Xiang, H.; Cheng, Z.; Xiong, Y.; et al. Clinical Characteristics of 138 Hospitalized Patients with 2019 Novel Coronavirus–Infected Pneumonia in Wuhan, China. JAMA 2020, 323, 1061–1069. [Google Scholar] [CrossRef] [PubMed]
- Liotta, E.M.; Batra, A.; Clark, J.R.; Shlobin, N.A.; Hoffman, S.C.; Orban, Z.S.; Koralnik, I.J. Frequent Neurologic Manifestations and Encephalopathy-associated Morbidity in COVID-19 Patients. Ann. Clin. Transl. Neurol. 2020, 7, 2221–2230. [Google Scholar] [CrossRef]
- Tian, Y.; Rong, L.; Nian, W.; He, Y. Review Article: Gastrointestinal Features in COVID-19 and the Possibility of Faecal Transmission. Aliment. Pharmacol. Ther. 2020, 51, 843–851. [Google Scholar] [CrossRef]
- Chen, B.; Julg, B.; Mohandas, S.; Bradfute, S.B. Viral Persistence, Reactivation, and Mechanisms of Long COVID. eLife 2023, 12, e86015. [Google Scholar] [CrossRef]
- Thaweethai, T.; Jolley, S.E.; Karlson, E.W.; Levitan, E.B.; Levy, B.; McComsey, G.A.; McCorkell, L.; Nadkarni, G.N.; Parthasarathy, S.; Singh, U.; et al. Development of a Definition of Postacute Sequelae of SARS-CoV-2 Infection. JAMA 2023, 329, 1934–1946. [Google Scholar] [CrossRef]
- Chen, C.; Haupert, S.R.; Zimmermann, L.; Shi, X.; Fritsche, L.G.; Mukherjee, B. Global Prevalence of Post-Coronavirus Disease 2019 (COVID-19) Condition or Long COVID: A Meta-Analysis and Systematic Review. J. Infect. Dis. 2022, 226, 1593–1607. [Google Scholar] [CrossRef]
- Kingery, J.R.; Safford, M.M.; Martin, P.; Lau, J.D.; Rajan, M.; Wehmeyer, G.T.; Li, H.A.; Alshak, M.N.; Jabri, A.; Kofman, A.; et al. Health Status, Persistent Symptoms, and Effort Intolerance One Year after Acute COVID-19 Infection. J. Gen. Intern. Med. 2022, 37, 1218–1225. [Google Scholar] [CrossRef] [PubMed]
- Kerksieck, P.; Ballouz, T.; Haile, S.R.; Schumacher, C.; Lacy, J.; Domenghino, A.; Fehr, J.S.; Bauer, G.F.; Dressel, H.; Puhan, M.A.; et al. Post COVID-19 Condition, Work Ability and Occupational Changes in a Population-Based Cohort. Lancet Reg. Health-Eur. 2023, 31, 100671. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Huang, L.; Wang, Y.; Li, X.; Ren, L.; Gu, X.; Kang, L.; Guo, L.; Liu, M.; Zhou, X.; et al. 6-Month Consequences of COVID-19 in Patients Discharged from Hospital: A Cohort Study. Lancet 2023, 401, e21–e33. [Google Scholar] [CrossRef]
- Yong, S.J. Long COVID or Post-COVID-19 Syndrome: Putative Pathophysiology, Risk Factors, and Treatments. Infect. Dis. 2021, 53, 737–754. [Google Scholar] [CrossRef] [PubMed]
- Ballouz, T.; Menges, D.; Anagnostopoulos, A.; Domenghino, A.; Aschmann, H.E.; Frei, A.; Fehr, J.S.; Puhan, M.A. Recovery and Symptom Trajectories up to Two Years after SARS-CoV-2 Infection: Population Based, Longitudinal Cohort Study. BMJ 2023, 381, e074425. [Google Scholar] [CrossRef]
- Bostanci, A.; Gazi, U.; Tosun, O.; Suer, K.; Unal Evren, E.; Evren, H.; Sanlidag, T. Long-COVID-19 in Asymptomatic, Non-Hospitalized, and Hospitalized Populations: A Cross-Sectional Study. J. Clin. Med. 2023, 12, 2613. [Google Scholar] [CrossRef] [PubMed]
- Vibholm, L.K.; Nielsen, S.S.F.; Pahus, M.H.; Frattari, G.S.; Olesen, R.; Andersen, R.; Monrad, I.; Andersen, A.H.F.; Thomsen, M.M.; Konrad, C.V.; et al. SARS-CoV-2 Persistence Is Associated with Antigen-Specific CD8 T-Cell Responses. EBioMedicine 2021, 64, 103230. [Google Scholar] [CrossRef] [PubMed]
- Ahamed, J.; Laurence, J. Long COVID Endotheliopathy: Hypothesized Mechanisms and Potential Therapeutic Approaches. J. Clin. Investig. 2022, 132, e161167. [Google Scholar] [CrossRef]
- Glynne, P.; Tahmasebi, N.; Gant, V.; Gupta, R. Long COVID Following Mild SARS-CoV-2 Infection: Characteristic T Cell Alterations and Response to Antihistamines. J. Investig. Med. Off. Publ. Am. Fed. Clin. Res. 2022, 70, 61–67. [Google Scholar] [CrossRef]
- Noonong, K.; Chatatikun, M.; Surinkaew, S.; Kotepui, M.; Hossain, R.; Bunluepuech, K.; Noothong, C.; Tedasen, A.; Klangbud, W.K.; Imai, M.; et al. Mitochondrial Oxidative Stress, Mitochondrial ROS Storms in Long COVID Pathogenesis. Front. Immunol. 2023, 14, 1275001. [Google Scholar] [CrossRef]
- Zubchenko, S.; Kril, I.; Nadizhko, O.; Matsyura, O.; Chopyak, V. Herpesvirus Infections and Post-COVID-19 Manifestations: A Pilot Observational Study. Rheumatol. Int. 2022, 42, 1523–1530. [Google Scholar] [CrossRef] [PubMed]
- Álvarez-Santacruz, C.; Tyrkalska, S.D.; Candel, S. The Microbiota in Long COVID. Int. J. Mol. Sci. 2024, 25, 1330. [Google Scholar] [CrossRef]
- Talla, A.; Vasaikar, S.V.; Szeto, G.L.; Lemos, M.P.; Czartoski, J.L.; MacMillan, H.; Moodie, Z.; Cohen, K.W.; Fleming, L.B.; Thomson, Z.; et al. Persistent Serum Protein Signatures Define an Inflammatory Subcategory of Long COVID. Nat. Commun. 2023, 14, 3417. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.; Yuan, D.; Chen, D.G.; Ng, R.H.; Wang, K.; Choi, J.; Li, S.; Hong, S.; Zhang, R.; Xie, J.; et al. Multiple Early Factors Anticipate Post-Acute COVID-19 Sequelae. Cell 2022, 185, 881–895. [Google Scholar] [CrossRef]
- Matschke, J.; Lütgehetmann, M.; Hagel, C.; Sperhake, J.P.; Schröder, A.S.; Edler, C.; Mushumba, H.; Fitzek, A.; Allweiss, L.; Dandri, M.; et al. Neuropathology of Patients with COVID-19 in Germany: A Post-Mortem Case Series. Lancet Neurol. 2020, 19, 919–929. [Google Scholar] [CrossRef] [PubMed]
- Spudich, S.; Nath, A. Nervous System Consequences of COVID-19. Science 2022, 375, 267–269. [Google Scholar] [CrossRef] [PubMed]
- Yong, S.J.; Halim, A.; Halim, M.; Ming, L.C.; Goh, K.W.; Alfaresi, M.; AlShehail, B.M.; Al Fares, M.A.; Alissa, M.; Sulaiman, T.; et al. Experimental Drugs in Randomized Controlled Trials for Long-COVID: What’s in the Pipeline? A Systematic and Critical Review. Expert Opin. Investig. Drugs 2023, 32, 655–667. [Google Scholar] [CrossRef] [PubMed]
- Khubber, S.; Hashemifesharaki, R.; Mohammadi, M.; Gharibzahedi, S.M.T. Garlic (Allium Sativum L.): A Potential Unique Therapeutic Food Rich in Organosulfur and Flavonoid Compounds to Fight with COVID-19. Nutr. J. 2020, 19, 124. [Google Scholar] [CrossRef]
- Suresh, M.V.; Francis, S.; Aktay, S.; Kralovich, G.; Raghavendran, K. Therapeutic Potential of Curcumin in ARDS and COVID-19. Clin. Exp. Pharmacol. Physiol. 2023, 50, 267–276. [Google Scholar] [CrossRef]
- Imran, M.; Khan, S.A.; Abida; Alshammari, M.K.; Alkhaldi, S.M.; Alshammari, F.N.; Kamal, M.; Alam, O.; Asdaq, S.M.B.; Alzahrani, A.K.; et al. Nigella sativa L. and COVID-19: A Glance at The Anti-COVID-19 Chemical Constituents, Clinical Trials, Inventions, and Patent Literature. Molecules 2022, 27, 2750. [Google Scholar] [CrossRef]
- Pakkir Maideen, N.M.; Hassan Jumale, A.; Ramadan Barakat, I.; Khalifa Albasti, A. Potential of Black Seeds (Nigella Sativa) in the Management of Long COVID or Post-Acute Sequelae of COVID-19 (PASC) and Persistent COVID-19 Symptoms—An Insight. Infect. Disord. Drug Targets 2023, 23, e230223213955. [Google Scholar] [CrossRef]
- Rathinavel, T.; Palanisamy, M.; Srinivasan, P.; Subramanian, A.; Thangaswamy, S. Phytochemical 6-Gingerol-A Promising Drug of Choice for COVID-19. Int. J. Adv. Sci. Eng. 2020, 6, 1482–1489. [Google Scholar] [CrossRef]
- Ali, A.M.; Kunugi, H. Propolis, Bee Honey, and Their Components Protect against Coronavirus Disease 2019 (COVID-19): A Review of in Silico, in Vitro, and Clinical Studies. Molecules 2021, 26, 1232. [Google Scholar] [CrossRef] [PubMed]
- Melrose, J.; Smith, M.M. Natural and Semi-Synthetic Flavonoid Anti-SARS-CoV-2 Agents for the Treatment of Long COVID-19 Disease and Neurodegenerative Disorders of Cognitive Decline. Front. Biosci. 2022, 14, 27. [Google Scholar] [CrossRef] [PubMed]
- Rizzuti, B.; Ceballos-Laita, L.; Ortega-Alarcon, D.; Jimenez-Alesanco, A.; Vega, S.; Grande, F.; Conforti, F.; Abian, O.; Velazquez-Campoy, A. Sub-Micromolar Inhibition of SARS-CoV-2 3CLpro by Natural Compounds. Pharmaceuticals 2021, 14, 892. [Google Scholar] [CrossRef] [PubMed]
- Marinaccio, L.; Stefanucci, A.; Scioli, G.; Della Valle, A.; Zengin, G.; Cichelli, A.; Mollica, A. Peptide Human Neutrophil Elastase Inhibitors from Natural Sources: An Overview. Int. J. Mol. Sci. 2022, 23, 2924. [Google Scholar] [CrossRef]
- Bai, Y.; Peng, W.; Yang, C.; Zou, W.; Liu, M.; Wu, H.; Fan, L.; Li, P.; Zeng, X.; Su, W. Pharmacokinetics and Metabolism of Naringin and Active Metabolite Naringenin in Rats, Dogs, Humans, and the Differences between Species. Front. Pharmacol. 2020, 11, 364. [Google Scholar] [CrossRef] [PubMed]
- Miles, E.A.; Calder, P.C. Effects of Citrus Fruit Juices and Their Bioactive Components on Inflammation and Immunity: A Narrative Review. Front. Immunol. 2021, 12, 712608. [Google Scholar] [CrossRef]
- Stabrauskiene, J.; Kopustinskiene, D.M.; Lazauskas, R.; Bernatoniene, J. Naringin and Naringenin: Their Mechanisms of Action and the Potential Anticancer Activities. Biomedicines 2022, 10, 1686. [Google Scholar] [CrossRef]
- Chen, X.; Xue, Y.; Jia, G.; Zhao, H.; Liu, G.; Huang, Z. Antifatigue Effect of Naringin on Improving Antioxidant Capacity and Mitochondrial Function and Preventing Muscle Damage. Exp. Biol. Med. 2022, 247, 1776–1784. [Google Scholar] [CrossRef] [PubMed]
- Casacchia, T.; Occhiuzzi, M.A.; Grande, F.; Rizzuti, B.; Granieri, M.C.; Rocca, C.; Gattuso, A.; Garofalo, A.; Angelone, T.; Statti, G. A Pilot Study on the Nutraceutical Properties of the Citrus Hybrid Tacle® as a Dietary Source of Polyphenols for Supplementation in Metabolic Disorders. J. Funct. Foods 2019, 52, 370–381. [Google Scholar] [CrossRef]
- Heidary Moghaddam, R.; Samimi, Z.; Moradi, S.Z.; Little, P.J.; Xu, S.; Farzaei, M.H. Naringenin and Naringin in Cardiovascular Disease Prevention: A Preclinical Review. Eur. J. Pharmacol. 2020, 887, 173535. [Google Scholar] [CrossRef] [PubMed]
- AmeliMojarad, M.; AmeliMojarad, M. Interleukin-6 Inhibitory Effect of Natural Product Naringenin Compared to a Synthesised Monoclonal Antibody against Life-Threatening COVID-19. Rev. Med. Virol. 2023, 33, e2445. [Google Scholar] [CrossRef]
- Agrawal, P.K.; Agrawal, C.; Blunden, G. Naringenin as a Possible Candidate against SARS-CoV-2 Infection and in the Pathogenesis of COVID-19. Nat. Prod. Commun. 2021, 16, 1934578X211066723. [Google Scholar] [CrossRef]
- Tutunchi, H.; Naeini, F.; Ostadrahimi, A.; Hosseinzadeh-Attar, M.J. Naringenin, a Flavanone with Antiviral and Anti-inflammatory Effects: A Promising Treatment Strategy against COVID-19. Phytother. Res. 2020, 34, 3137–3147. [Google Scholar] [CrossRef] [PubMed]
- Aleebrahim-Dehkordi, E.; Ghoshouni, H.; Koochaki, P.; Esmaili-Dehkordi, M.; Aleebrahim, E.; Chichagi, F.; Jafari, A.; Hanaei, S.; Heidari-Soureshjani, E.; Rezaei, N. Targeting the Vital Non-Structural Proteins (NSP12, NSP7, NSP8 and NSP3) from SARS-CoV-2 and Inhibition of RNA Polymerase by Natural Bioactive Compound Naringenin as a Promising Drug Candidate against COVID-19. J. Mol. Struct. 2023, 1287, 135642. [Google Scholar] [CrossRef]
- Matias-Guiu, J.A.; Herrera, E.; González-Nosti, M.; Krishnan, K.; Delgado-Alonso, C.; Díez-Cirarda, M.; Yus, M.; Martínez-Petit, Á.; Pagán, J.; Matías-Guiu, J.; et al. Development of Criteria for Cognitive Dysfunction in Post-COVID Syndrome: The IC-CoDi-COVID Approach. Psychiatry Res. 2023, 319, 115006. [Google Scholar] [CrossRef]
- Premraj, L.; Kannapadi, N.V.; Briggs, J.; Seal, S.M.; Battaglini, D.; Fanning, J.; Suen, J.; Robba, C.; Fraser, J.; Cho, S.-M. Mid and Long-Term Neurological and Neuropsychiatric Manifestations of Post-COVID-19 Syndrome: A Meta-Analysis. J. Neurol. Sci. 2022, 434, 120162. [Google Scholar] [CrossRef]
- Reiss, A.B.; Greene, C.; Dayaramani, C.; Rauchman, S.H.; Stecker, M.M.; De Leon, J.; Pinkhasov, A. Long COVID, the Brain, Nerves, and Cognitive Function. Neurol. Int. 2023, 15, 821–841. [Google Scholar] [CrossRef]
- Ceban, F.; Ling, S.; Lui, L.M.W.; Lee, Y.; Gill, H.; Teopiz, K.M.; Rodrigues, N.B.; Subramaniapillai, M.; Di Vincenzo, J.D.; Cao, B.; et al. Fatigue and Cognitive Impairment in Post-COVID-19 Syndrome: A Systematic Review and Meta-Analysis. Brain Behav. Immun. 2022, 101, 93–135. [Google Scholar] [CrossRef] [PubMed]
- Zeng, N.; Zhao, Y.-M.; Yan, W.; Li, C.; Lu, Q.-D.; Liu, L.; Ni, S.-Y.; Mei, H.; Yuan, K.; Shi, L.; et al. A Systematic Review and Meta-Analysis of Long Term Physical and Mental Sequelae of COVID-19 Pandemic: Call for Research Priority and Action. Mol. Psychiatry 2023, 28, 423–433. [Google Scholar] [CrossRef] [PubMed]
- Cheetham, N.J.; Penfold, R.; Giunchiglia, V.; Bowyer, V.; Sudre, C.H.; Canas, L.S.; Deng, J.; Murray, B.; Kerfoot, E.; Antonelli, M.; et al. The Effects of COVID-19 on Cognitive Performance in a Community-Based Cohort: A COVID Symptom Study Biobank Prospective Cohort Study. EClinicalMedicine 2023, 62, 102086. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Jin, L.; Zhang, M.; Wu, Y.; Duan, Z.; Guo, Y.; Wang, C.; Guo, Y.; Chen, W.; Liao, Z.; et al. Blood–Brain Barrier Injury and Neuroinflammation Induced by SARS-CoV-2 in a Lung–Brain Microphysiological System. Nat. Biomed. Eng. 2023, 1–16. [Google Scholar] [CrossRef]
- Soung, A.L.; Vanderheiden, A.; Nordvig, A.S.; Sissoko, C.A.; Canoll, P.; Mariani, M.B.; Jiang, X.; Bricker, T.; Rosoklija, G.B.; Arango, V.; et al. COVID-19 Induces CNS Cytokine Expression and Loss of Hippocampal Neurogenesis. Brain 2022, 145, 4193–4201. [Google Scholar] [CrossRef]
- Yang, L.; Kim, T.W.; Han, Y.; Nair, M.S.; Harschnitz, O.; Zhu, J.; Wang, P.; Koo, S.Y.; Lacko, L.A.; Chandar, V.; et al. SARS-CoV-2 Infection Causes Dopaminergic Neuron Senescence. Cell Stem Cell 2024, S1934590923004423. [Google Scholar] [CrossRef]
- Kavanagh, E. Long COVID Brain Fog: A Neuroinflammation Phenomenon? Oxf. Open Immunol. 2022, 3, iqac007. [Google Scholar] [CrossRef]
- Beurel, E.; Toups, M.; Nemeroff, C.B. The Bidirectional Relationship of Depression and Inflammation: Double Trouble. Neuron 2020, 107, 234–256. [Google Scholar] [CrossRef]
- Leng, A.; Shah, M.; Ahmad, S.A.; Premraj, L.; Wildi, K.; Li Bassi, G.; Pardo, C.A.; Choi, A.; Cho, S.-M. Pathogenesis Underlying Neurological Manifestations of Long COVID Syndrome and Potential Therapeutics. Cells 2023, 12, 816. [Google Scholar] [CrossRef]
- Pröbstel, A.-K.; Schirmer, L. SARS-CoV-2-Specific Neuropathology: Fact or Fiction? Trends Neurosci. 2021, 44, 933–935. [Google Scholar] [CrossRef]
- Cantuti-Castelvetri, L.; Ojha, R.; Pedro, L.D.; Djannatian, M.; Franz, J.; Kuivanen, S.; van der Meer, F.; Kallio, K.; Kaya, T.; Anastasina, M.; et al. Neuropilin-1 Facilitates SARS-CoV-2 Cell Entry and Infectivity. Science 2020, 370, 856–860. [Google Scholar] [CrossRef] [PubMed]
- Quan, M.; Wang, X.; Gong, M.; Wang, Q.; Li, Y.; Jia, J. Post-COVID Cognitive Dysfunction: Current Status and Research Recommendations for High Risk Population. Lancet Reg. Health West. Pac. 2023, 38, 100836. [Google Scholar] [CrossRef] [PubMed]
- Hugon, J.; Msika, E.-F.; Queneau, M.; Farid, K.; Paquet, C. Long COVID: Cognitive Complaints (Brain Fog) and Dysfunction of the Cingulate Cortex. J. Neurol. 2022, 269, 44–46. [Google Scholar] [CrossRef] [PubMed]
- Wong, A.C.; Devason, A.S.; Umana, I.C.; Cox, T.O.; Dohnalová, L.; Litichevskiy, L.; Perla, J.; Lundgren, P.; Etwebi, Z.; Izzo, L.T.; et al. Serotonin Reduction in Post-Acute Sequelae of Viral Infection. Cell 2023, 186, 4851–4867. [Google Scholar] [CrossRef] [PubMed]
- Kwatra, M.; Kumar, V.; Jangra, A.; Mishra, M.; Ahmed, S.; Ghosh, P.; Vohora, D.; Khanam, R. Ameliorative Effect of Naringin against Doxorubicin-Induced Acute Cardiac Toxicity in Rats. Pharm. Biol. 2016, 54, 637–647. [Google Scholar] [CrossRef] [PubMed]
- Nouri, Z.; Fakhri, S.; El-Senduny, F.F.; Sanadgol, N.; Abd-ElGhani, G.E.; Farzaei, M.H.; Chen, J.-T. On the Neuroprotective Effects of Naringenin: Pharmacological Targets, Signaling Pathways, Molecular Mechanisms, and Clinical Perspective. Biomolecules 2019, 9, 690. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Liu, R.; He, J.; Li, J.; Guo, J.; Chen, Y.; Ji, K. Naringin Regulates Microglia BV-2 Activation and Inflammation via the JAK/STAT3 Pathway. Evid.-Based Complement. Altern. Med. 2022, 2022, 3492058. [Google Scholar] [CrossRef] [PubMed]
- Liddell, J.R. Are Astrocytes the Predominant Cell Type for Activation of Nrf2 in Aging and Neurodegeneration? Antioxidants 2017, 6, 65. [Google Scholar] [CrossRef]
- Wang, G.-Q.; Zhang, B.; He, X.-M.; Li, D.-D.; Shi, J.-S.; Zhang, F. Naringenin Targets on Astroglial Nrf2 to Support Dopaminergic Neurons. Pharmacol. Res. 2019, 139, 452–459. [Google Scholar] [CrossRef]
- Salman, M.; Sharma, P.; Alam, M.d.I.; Tabassum, H.; Parvez, S. Naringenin Mitigates Behavioral Alterations and Provides Neuroprotection against 3-Nitropropinoic Acid-Induced Huntington’s Disease like Symptoms in Rats. Nutr. Neurosci. 2022, 25, 1898–1908. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, Y.; Liu, Y.; Niu, X. Naringenin Attenuates Cognitive Impairment in a Rat Model of Vascular Dementia by Inhibiting Hippocampal Oxidative Stress and Inflammatory Response and Promoting N-Methyl-D-Aspartate Receptor Signaling Pathway. Neurochem. Res. 2022, 47, 3402–3413. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Prakash, A.; Dogra, S. Naringin Alleviates Cognitive Impairment, Mitochondrial Dysfunction and Oxidative Stress Induced by D-Galactose in Mice. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2010, 48, 626–632. [Google Scholar] [CrossRef] [PubMed]
- Kwatra, M.; Jangra, A.; Mishra, M.; Sharma, Y.; Ahmed, S.; Ghosh, P.; Kumar, V.; Vohora, D.; Khanam, R. Naringin and Sertraline Ameliorate Doxorubicin-Induced Behavioral Deficits through Modulation of Serotonin Level and Mitochondrial Complexes Protection Pathway in Rat Hippocampus. Neurochem. Res. 2016, 41, 2352–2366. [Google Scholar] [CrossRef] [PubMed]
- Mahdavinia, M.; Ahangarpour, A.; Zeidooni, L.; Samimi, A.; Alizadeh, S.; Dehghani, M.A.; Alboghobeish, S. Protective Effect of Naringin on Bisphenol A-Induced Cognitive Dysfunction and Oxidative Damage in Rats. Int. J. Mol. Cell. Med. 2019, 8, 141–153. [Google Scholar] [CrossRef] [PubMed]
- Mansour, L.A.H.; Elshopakey, G.E.; Abdelhamid, F.M.; Albukhari, T.A.; Almehmadi, S.J.; Refaat, B.; El-Boshy, M.; Risha, E.F. Hepatoprotective and Neuroprotective Effects of Naringenin against Lead-Induced Oxidative Stress, Inflammation, and Apoptosis in Rats. Biomedicines 2023, 11, 1080. [Google Scholar] [CrossRef] [PubMed]
- Phillips, C. Brain-Derived Neurotrophic Factor, Depression, and Physical Activity: Making the Neuroplastic Connection. Neural. Plast. 2017, 2017, 7260130. [Google Scholar] [CrossRef]
- Esvald, E.-E.; Tuvikene, J.; Sirp, A.; Patil, S.; Bramham, C.R.; Timmusk, T. CREB Family Transcription Factors Are Major Mediators of BDNF Transcriptional Autoregulation in Cortical Neurons. J. Neurosci. 2020, 40, 1405–1426. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Lu, R.-R.; Xu, R.-H.; Wang, H.-H.; Feng, W.-S.; Zheng, X.-K. Naringenin and Apigenin Ameliorates Corticosterone-Induced Depressive Behaviors. Heliyon 2023, 9, e15618. [Google Scholar] [CrossRef]
- Yi, L.-T.; Li, J.; Li, H.-C.; Su, D.-X.; Quan, X.-B.; He, X.-C.; Wang, X.-H. Antidepressant-like Behavioral, Neurochemical and Neuroendocrine Effects of Naringenin in the Mouse Repeated Tail Suspension Test. Prog. Neuropsychopharmacol. Biol. Psychiatry 2012, 39, 175–181. [Google Scholar] [CrossRef]
- Jain, A.S.; Sushma, P.; Dharmashekar, C.; Beelagi, M.S.; Prasad, S.K.; Shivamallu, C.; Prasad, A.; Syed, A.; Marraiki, N.; Prasad, K.S. In Silico Evaluation of Flavonoids as Effective Antiviral Agents on the Spike Glycoprotein of SARS-CoV-2. Saudi J. Biol. Sci. 2021, 28, 1040–1051. [Google Scholar] [CrossRef]
- Sette, A.; Crotty, S. Adaptive Immunity to SARS-CoV-2 and COVID-19. Cell 2021, 184, 861–880. [Google Scholar] [CrossRef]
- Davis, H.E.; McCorkell, L.; Vogel, J.M.; Topol, E.J. Long COVID: Major Findings, Mechanisms and Recommendations. Nat. Rev. Microbiol. 2023, 21, 133–146. [Google Scholar] [CrossRef] [PubMed]
- Klein, J.; Wood, J.; Jaycox, J.R.; Dhodapkar, R.M.; Lu, P.; Gehlhausen, J.R.; Tabachnikova, A.; Greene, K.; Tabacof, L.; Malik, A.A.; et al. Distinguishing Features of Long COVID Identified through Immune Profiling. Nature 2023, 623, 139–148. [Google Scholar] [CrossRef] [PubMed]
- Cheong, J.-G.; Ravishankar, A.; Sharma, S.; Parkhurst, C.N.; Grassmann, S.A.; Wingert, C.K.; Laurent, P.; Ma, S.; Paddock, L.; Miranda, I.C.; et al. Epigenetic Memory of Coronavirus Infection in Innate Immune Cells and Their Progenitors. Cell 2023, 186, 3882–3902.e24. [Google Scholar] [CrossRef] [PubMed]
- Fogarty, H.; Ward, S.E.; Townsend, L.; Karampini, E.; Elliott, S.; Conlon, N.; Dunne, J.; Kiersey, R.; Naughton, A.; Gardiner, M.; et al. Sustained VWF-ADAMTS-13 Axis Imbalance and Endotheliopathy in Long COVID Syndrome Is Related to Immune Dysfunction. J. Thromb. Haemost. 2022, 20, 2429–2438. [Google Scholar] [CrossRef] [PubMed]
- Townsend, L.; Dyer, A.H.; Naughton, A.; Kiersey, R.; Holden, D.; Gardiner, M.; Dowds, J.; O’Brien, K.; Bannan, C.; Nadarajan, P.; et al. Longitudinal Analysis of COVID-19 Patients Shows Age-Associated T Cell Changes Independent of Ongoing Ill-Health. Front. Immunol. 2021, 12, 676932. [Google Scholar] [CrossRef] [PubMed]
- Phetsouphanh, C.; Darley, D.R.; Wilson, D.B.; Howe, A.; Munier, C.M.L.; Patel, S.K.; Juno, J.A.; Burrell, L.M.; Kent, S.J.; Dore, G.J.; et al. Immunological Dysfunction Persists for 8 Months Following Initial Mild-to-Moderate SARS-CoV-2 Infection. Nat. Immunol. 2022, 23, 210–216. [Google Scholar] [CrossRef] [PubMed]
- Sumi, T.; Harada, K. Immune Response to SARS-CoV-2 in Severe Disease and Long COVID-19. iScience 2022, 25, 104723. [Google Scholar] [CrossRef] [PubMed]
- Gruber, C.N.; Patel, R.S.; Trachtman, R.; Lepow, L.; Amanat, F.; Krammer, F.; Wilson, K.M.; Onel, K.; Geanon, D.; Tuballes, K.; et al. Mapping Systemic Inflammation and Antibody Responses in Multisystem Inflammatory Syndrome in Children (Mis-c). Cell 2020, 183, 982–995. [Google Scholar] [CrossRef]
- Wang, E.Y.; Mao, T.; Klein, J.; Dai, Y.; Huck, J.D.; Jaycox, J.R.; Liu, F.; Zhou, T.; Israelow, B.; Wong, P.; et al. Diverse Functional Autoantibodies in Patients with COVID-19. Nature 2021, 595, 283–288. [Google Scholar] [CrossRef] [PubMed]
- Wallukat, G.; Hohberger, B.; Wenzel, K.; Fürst, J.; Schulze-Rothe, S.; Wallukat, A.; Hönicke, A.-S.; Müller, J. Functional Autoantibodies against G-Protein Coupled Receptors in Patients with Persistent Long-COVID-19 Symptoms. J. Transl. Autoimmun. 2021, 4, 100100. [Google Scholar] [CrossRef]
- Novelli, L.; Motta, F.; De Santis, M.; Ansari, A.A.; Gershwin, M.E.; Selmi, C. The JANUS of Chronic Inflammatory and Autoimmune Diseases Onset during COVID-19—A Systematic Review of the Literature. J. Autoimmun. 2021, 117, 102592. [Google Scholar] [CrossRef]
- Liu, Z.; Niu, X.; Wang, J. Naringenin as a Natural Immunomodulator against T Cell-Mediated Autoimmune Diseases: Literature Review and Network-Based Pharmacology Study. Crit. Rev. Food Sci. Nutr. 2022, 63, 11026–11043. [Google Scholar] [CrossRef]
- Wang, H.-K.; Yeh, C.-H.; Iwamoto, T.; Satsu, H.; Shimizu, M.; Totsuka, M. Dietary Flavonoid Naringenin Induces Regulatory T Cells via an Aryl Hydrocarbon Receptor Mediated Pathway. J. Agric. Food Chem. 2012, 60, 2171–2178. [Google Scholar] [CrossRef]
- Zhu, J.; Yamane, H.; Paul, W.E. Differentiation of Effector CD4 T Cell Populations. Annu. Rev. Immunol. 2010, 28, 445–489. [Google Scholar] [CrossRef] [PubMed]
- Niu, X.; Wu, C.; Li, M.; Zhao, Q.; Meydani, S.N.; Wang, J.; Wu, D. Naringenin Is an Inhibitor of T Cell Effector Functions. J. Nutr. Biochem. 2018, 58, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Niu, X.; Wu, C.; Wu, D. Naringenin Modifies the Development of Lineage-Specific Effector CD4+ t Cells. Front. Immunol. 2018, 9, 2267. [Google Scholar] [CrossRef]
- Jiang, Y.-P.; Wen, J.-J.; Zhao, X.-X.; Gao, Y.-C.; Ma, X.; Song, S.-Y.; Jin, Y.; Shao, T.-J.; Yu, J.; Wen, C.-P. The Flavonoid Naringenin Alleviates Collagen-Induced Arthritis through Curbing the Migration and Polarization of Cd4+ t Lymphocyte Driven by Regulating Mitochondrial Fission. Int. J. Mol. Sci. 2022, 24, 279. [Google Scholar] [CrossRef]
- Abrego-Peredo, A.; Romero-Ramírez, H.; Espinosa, E.; López-Herrera, G.; García-García, F.; Flores-Muñoz, M.; Sandoval-Montes, C.; Rodríguez-Alba, J.C. Naringenin Mitigates Autoimmune Features in Lupus-Prone Mice by Modulation of T-Cell Subsets and Cytokines Profile. PLoS ONE 2020, 15, e0233138. [Google Scholar] [CrossRef]
- Wang, J.; Qi, Y.; Niu, X.; Tang, H.; Meydani, S.N.; Wu, D. Dietary Naringenin Supplementation Attenuates Experimental Autoimmune Encephalomyelitis by Modulating Autoimmune Inflammatory Responses in Mice. J. Nutr. Biochem. 2018, 54, 130–139. [Google Scholar] [CrossRef] [PubMed]
- Zeng, W.; Jin, L.; Zhang, F.; Zhang, C.; Liang, W. Naringenin as a Potential Immunomodulator in Therapeutics. Pharmacol. Res. 2018, 135, 122–126. [Google Scholar] [CrossRef] [PubMed]
- Jin, L.; Zeng, W.; Zhang, F.; Zhang, C.; Liang, W. Naringenin Ameliorates Acute Inflammation by Regulating Intracellular Cytokine Degradation. J. Immunol. 2017, 199, 3466–3477. [Google Scholar] [CrossRef] [PubMed]
- Yeoh, Y.K.; Zuo, T.; Lui, G.C.-Y.; Zhang, F.; Liu, Q.; Li, A.Y.; Chung, A.C.; Cheung, C.P.; Tso, E.Y.; Fung, K.S.; et al. Gut Microbiota Composition Reflects Disease Severity and Dysfunctional Immune Responses in Patients with COVID-19. Gut 2021, 70, 698–706. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Lau, R.I.; Liu, Q.; Su, Q.; Chan, F.K.L.; Ng, S.C. Gut Microbiota in COVID-19: Key Microbial Changes, Potential Mechanisms and Clinical Applications. Nat. Rev. Gastroenterol. Hepatol. 2023, 20, 323–337. [Google Scholar] [CrossRef] [PubMed]
- de Ana Patrícia, O.; Lopes, A.L.F.; Pacheco, G.; de Sá Guimarães Nolêto, I.R.; Nicolau, L.A.D.; Medeiros, J.V.R. Premises among SARS-CoV-2, Dysbiosis and Diarrhea: Walking through the ACE2/mTOR/Autophagy Route. Med. Hypotheses 2020, 144, 110243. [Google Scholar] [CrossRef]
- Gu, S.; Chen, Y.; Wu, Z.; Chen, Y.; Gao, H.; Lv, L.; Guo, F.; Zhang, X.; Luo, R.; Huang, C.; et al. Alterations of the Gut Microbiota in Patients with COVID-19 or H1n1 Influenza. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2020, 71, ciaa709. [Google Scholar] [CrossRef]
- Zhong, H.; Wang, Y.; Shi, Z.; Zhang, L.; Ren, H.; He, W.; Zhang, Z.; Zhu, A.; Zhao, J.; Xiao, F.; et al. Characterization of Respiratory Microbial Dysbiosis in Hospitalized COVID-19 Patients. Cell Discov. 2021, 7, 23. [Google Scholar] [CrossRef]
- Soffritti, I.; D’Accolti, M.; Fabbri, C.; Passaro, A.; Manfredini, R.; Zuliani, G.; Libanore, M.; Franchi, M.; Contini, C.; Caselli, E. Oral Microbiome Dysbiosis Is Associated with Symptoms Severity and Local Immune/Inflammatory Response in COVID-19 Patients: A Cross-Sectional Study. Front. Microbiol. 2021, 12, 687513. [Google Scholar] [CrossRef]
- Nardelli, C.; Gentile, I.; Setaro, M.; Di Domenico, C.; Pinchera, B.; Buonomo, A.R.; Zappulo, E.; Scotto, R.; Scaglione, G.L.; Castaldo, G.; et al. Nasopharyngeal Microbiome Signature in COVID-19 Positive Patients: Can We Definitively Get a Role to Fusobacterium Periodonticum? Front. Cell. Infect. Microbiol. 2021, 11, 625581. [Google Scholar] [CrossRef]
- Chen, Y.; Gu, S.; Chen, Y.; Lu, H.; Shi, D.; Guo, J.; Wu, W.-R.; Yang, Y.; Li, Y.; Xu, K.-J.; et al. Six-Month Follow-up of Gut Microbiota Richness in Patients with COVID-19. Gut 2022, 71, 222–225. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Zhang, J.; Zhang, D.; Ma, W.-L.; Wang, X. Linking the Gut Microbiota to Persistent Symptoms in Survivors of COVID-19 after Discharge. J. Microbiol. 2021, 59, 941–948. [Google Scholar] [CrossRef] [PubMed]
- Nagata, N.; Takeuchi, T.; Masuoka, H.; Aoki, R.; Ishikane, M.; Iwamoto, N.; Sugiyama, M.; Suda, W.; Nakanishi, Y.; Terada-Hirashima, J.; et al. Human Gut Microbiota and Its Metabolites Impact Immune Responses in COVID-19 and Its Complications. Gastroenterology 2023, 164, 272–288. [Google Scholar] [CrossRef] [PubMed]
- de Gislane Lelis Vilela, O.; Oliveira, C.N.S.; Pinzan, C.F.; de Salis, L.V.V.; Cardoso, C.R.d.B. Microbiota Modulation of the Gut-Lung Axis in COVID-19. Front. Immunol. 2021, 12, 635471. [Google Scholar] [CrossRef]
- Chen, J.; Vitetta, L. The Role of the Gut-Lung Axis in COVID-19 Infections and Its Modulation to Improve Clinical Outcomes. Front. Biosci. Sch. Ed. 2022, 14, 23. [Google Scholar] [CrossRef] [PubMed]
- Carloni, S.; Bertocchi, A.; Mancinelli, S.; Bellini, M.; Erreni, M.; Borreca, A.; Braga, D.; Giugliano, S.; Mozzarelli, A.M.; Manganaro, D.; et al. Identification of a Choroid Plexus Vascular Barrier Closing during Intestinal Inflammation. Science 2021, 374, 439–448. [Google Scholar] [CrossRef] [PubMed]
- Feng, W.; Ao, H.; Peng, C.; Yan, D. Gut Microbiota, a New Frontier to Understand Traditional Chinese Medicines. Pharmacol. Res. 2019, 142, 176–191. [Google Scholar] [CrossRef] [PubMed]
- Cheng, H.; Zhang, D.; Wu, J.; Liu, J.; Zhou, Y.; Tan, Y.; Feng, W.; Peng, C. Interactions between Gut Microbiota and Polyphenols: A Mechanistic and Metabolomic Review. Phytomedicine Int. J. Phytother. Phytopharm. 2023, 119, 154979. [Google Scholar] [CrossRef]
- Li, P.; Yao, X.; Zhou, Q.; Meng, X.; Zhou, T.; Gu, Q. Citrus Peel Flavonoid Extracts: Health-Beneficial Bioactivities and Regulation of Intestinal Microecology in Vitro. Front. Nutr. 2022, 9, 888745. [Google Scholar] [CrossRef]
- Pan, L.; Ye, H.; Pi, X.; Liu, W.; Wang, Z.; Zhang, Y.; Zheng, J. Effects of Several Flavonoids on Human Gut Microbiota and Its Metabolism by in Vitro Simulated Fermentation. Front. Microbiol. 2023, 14, 1092729. [Google Scholar] [CrossRef]
- Firrman, J.; Liu, L.; Argoty, G.A.; Zhang, L.; Tomasula, P.; Wang, M.; Pontious, S.; Kobori, M.; Xiao, W. Analysis of Temporal Changes in Growth and Gene Expression for Commensal Gut Microbes in Response to the Polyphenol Naringenin. Microbiol. Insights 2018, 11, 1178636118775100. [Google Scholar] [CrossRef]
- Sun, Z.; Song, Z.-G.; Liu, C.; Tan, S.; Lin, S.; Zhu, J.; Dai, F.-H.; Gao, J.; She, J.-L.; Mei, Z.; et al. Gut Microbiome Alterations and Gut Barrier Dysfunction Are Associated with Host Immune Homeostasis in COVID-19 Patients. BMC Med. 2022, 20, 24. [Google Scholar] [CrossRef]
- Wu, X.; Guo, H.; Pan, X.; Huang, R.; Wang, G.; Liu, J. Naringin Exhibited Therapeutic Effects against DSS-Induced Mice Ulcerative Colitis in Intestinal Barrier–Dependent Manner. Molecules 2021, 26, 6604. [Google Scholar] [CrossRef]
- Rasa, S.; Nora-Krukle, Z.; Henning, N.; Eliassen, E.; Shikova, E.; Harrer, T.; Scheibenbogen, C.; Murovska, M.; Prusty, B.K. Chronic Viral Infections in Myalgic Encephalomyelitis/Chronic Fatigue Syndrome (ME/CFS). J. Transl. Med. 2018, 16, 268. [Google Scholar] [CrossRef]
- Renz-Polster, H.; Scheibenbogen, C. Post-COVID-Syndrom Mit Fatigue Und Belastungsintoleranz: Myalgische Enzephalomyelitis Bzw. Chronisches Fatigue-Syndrom. Inn. Med. 2022, 63, 830–839. [Google Scholar] [CrossRef]
- Komaroff, A.L.; Lipkin, W.I. ME/CFS and Long COVID Share Similar Symptoms and Biological Abnormalities: Road Map to the Literature. Front. Med. 2023, 10, 1187163. [Google Scholar] [CrossRef] [PubMed]
- Komaroff, A.L.; Bateman, L. Will COVID-19 Lead to Myalgic Encephalomyelitis/Chronic Fatigue Syndrome? Front. Med. 2021, 7, 606824. [Google Scholar] [CrossRef] [PubMed]
- Lv, Y.; Zhang, T.; Cai, J.; Huang, C.; Zhan, S.; Liu, J. Bioinformatics and Systems Biology Approach to Identify the Pathogenetic Link of Long COVID and Myalgic Encephalomyelitis/Chronic Fatigue Syndrome. Front. Immunol. 2022, 13, 952987. [Google Scholar] [CrossRef]
- Bonilla, H.; Quach, T.C.; Tiwari, A.; Bonilla, A.E.; Miglis, M.; Yang, P.C.; Eggert, L.E.; Sharifi, H.; Horomanski, A.; Subramanian, A.; et al. Myalgic Encephalomyelitis/Chronic Fatigue Syndrome Is Common in Post-Acute Sequelae of SARS-CoV-2 Infection (PASC): Results from a Post-COVID-19 Multidisciplinary Clinic. Front. Neurol. 2023, 14, 1090747. [Google Scholar] [CrossRef]
- Yang, Y.; Wu, Y.; Meng, X.; Wang, Z.; Younis, M.; Liu, Y.; Wang, P.; Huang, X. SARS-CoV-2 Membrane Protein Causes the Mitochondrial Apoptosis and Pulmonary Edema via Targeting BOK. Cell Death Differ. 2022, 29, 1395–1408. [Google Scholar] [CrossRef]
- Díaz-Resendiz, K.J.G.; Benitez-Trinidad, A.B.; Covantes-Rosales, C.E.; Toledo-Ibarra, G.A.; Ortiz-Lazareno, P.C.; Girón-Pérez, D.A.; Bueno-Durán, A.Y.; Pérez-Díaz, D.A.; Barcelos-García, R.G.; Girón-Pérez, M.I. Loss of Mitochondrial Membrane Potential (ΔΨ m) in Leucocytes as post-COVID-19 Sequelae. J. Leukoc. Biol. 2022, 112, 23–29. [Google Scholar] [CrossRef]
- Miller, B.; Silverstein, A.; Flores, M.; Cao, K.; Kumagai, H.; Mehta, H.H.; Yen, K.; Kim, S.-J.; Cohen, P. Host Mitochondrial Transcriptome Response to SARS-CoV-2 in Multiple Cell Models and Clinical Samples. Sci. Rep. 2021, 11, 3. [Google Scholar] [CrossRef]
- Appelman, B.; Charlton, B.T.; Goulding, R.P.; Kerkhoff, T.J.; Breedveld, E.A.; Noort, W.; Offringa, C.; Bloemers, F.W.; Van Weeghel, M.; Schomakers, B.V.; et al. Muscle Abnormalities Worsen after Post-Exertional Malaise in Long COVID. Nat. Commun. 2024, 15, 17. [Google Scholar] [CrossRef]
- Chaves-Filho, A.M.; Braniff, O.; Angelova, A.; Deng, Y.; Tremblay, M.-È. Chronic Inflammation, Neuroglial Dysfunction, and Plasmalogen Deficiency as a New Pathobiological Hypothesis Addressing the Overlap between Post-COVID-19 Symptoms and Myalgic Encephalomyelitis/Chronic Fatigue Syndrome. Brain Res. Bull. 2023, 201, 110702. [Google Scholar] [CrossRef]
- Komaroff, A.L.; Lipkin, W.I. Insights from Myalgic Encephalomyelitis/Chronic Fatigue Syndrome May Help Unravel the Pathogenesis of Postacute COVID-19 Syndrome. Trends Mol. Med. 2021, 27, 895–906. [Google Scholar] [CrossRef] [PubMed]
- Paul, B.D.; Lemle, M.D.; Komaroff, A.L.; Snyder, S.H. Redox Imbalance Links COVID-19 and Myalgic Encephalomyelitis/Chronic Fatigue Syndrome. Proc. Natl. Acad. Sci. USA 2021, 118, e2024358118. [Google Scholar] [CrossRef] [PubMed]
- Toogood, P.L.; Clauw, D.J.; Phadke, S.; Hoffman, D. Myalgic Encephalomyelitis/Chronic Fatigue Syndrome (ME/CFS): Where Will the Drugs Come From? Pharmacol. Res. 2021, 165, 105465. [Google Scholar] [CrossRef] [PubMed]
- Zamanian, M.; Hajizadeh, M.; Shamsizadeh, A.; Moemenzadeh, M.; Amirteimouri, M.; Elshiekh, M.; Allahtavakoli, M. Effects of Naringin on Physical Fatigue and Serum MMP-9 Concentration in Female Rats. Pharm. Biol. 2016, 55, 423–427. [Google Scholar] [CrossRef] [PubMed]
- Balachandran, A.; Choi, S.B.; Beata, M.-M.; Małgorzata, J.; Froemming, G.R.A.; Lavilla, C.A.; Billacura, M.P.; Siyumbwa, S.N.; Okechukwu, P.N. Antioxidant, Wound Healing Potential and in Silico Assessment of Naringin, Eicosane and Octacosane. Molecules 2023, 28, 1043. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Deng, S.; Li, J.; Gong, S.; Song, T.; Ge, J.; Zhao, Y.; Zhang, J.; Ma, L.; Zheng, Y.; et al. UPLC-MS Analysis and Network Pharmacology-Based Investigation into the Active Ingredients and Molecular Mechanisms of Anti-Fatigue of Male Flowers with Eucommia Ulmoides Oliv. Fundam. Clin. Pharmacol. 2022, 36, 1083–1098. [Google Scholar] [CrossRef] [PubMed]
- Kulasekaran, G.; Ganapasam, S. Neuroprotective Efficacy of Naringin on 3-Nitropropionic Acid-Induced Mitochondrial Dysfunction through the Modulation of Nrf2 Signaling Pathway in PC12 Cells. Mol. Cell. Biochem. 2015, 409, 199–211. [Google Scholar] [CrossRef] [PubMed]
- Varshney, V.; Garabadu, D. Naringin Exhibits Mas Receptor–Mediated Neuroprotection against Amyloid Beta–Induced Cognitive Deficits and Mitochondrial Toxicity in Rat Brain. Neurotox. Res. 2021, 39, 1023–1043. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Chen, Z.; Huang, L.; Meng, B.; Zhou, X.; Wen, X.; Ren, D. Naringenin Reduces Oxidative Stress and Improves Mitochondrial Dysfunction via Activation of the Nrf2/ARE Signaling Pathway in Neurons. Int. J. Mol. Med. 2017, 40, 1582–1590. [Google Scholar] [CrossRef] [PubMed]
- Vij, G.; Gupta, A.; Chopra, K. Modulation of Antigen-Induced Chronic Fatigue in Mouse Model of Water Immersion Stress by Naringin, a Polyphenolic Antioxidant. Fundam. Clin. Pharmacol. 2009, 23, 331–337. [Google Scholar] [CrossRef] [PubMed]
- Viswanatha, G.L.; Shylaja, H.; Moolemath, Y. The Beneficial Role of Naringin- a Citrus Bioflavonoid, against Oxidative Stress-Induced Neurobehavioral Disorders and Cognitive Dysfunction in Rodents: A Systematic Review and Meta-Analysis. Biomed. Pharmacother. 2017, 94, 909–929. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Xu, E.; Bowe, B.; Al-Aly, Z. Long-Term Cardiovascular Outcomes of COVID-19. Nat. Med. 2022, 28, 583–590. [Google Scholar] [CrossRef]
- Huseynov, A.; Akin, I.; Duerschmied, D.; Scharf, R.E. Cardiac Arrhythmias in Post-COVID Syndrome: Prevalence, Pathology, Diagnosis, and Treatment. Viruses 2023, 15, 389. [Google Scholar] [CrossRef]
- Guo, B.; Zhao, C.; He, M.Z.; Senter, C.; Zhou, Z.; Peng, J.; Li, S.; Fitzpatrick, A.L.; Lindström, S.; Stebbins, R.C.; et al. Identifying Patterns of Reported Findings on Long-Term Cardiac Complications of COVID-19: A Systematic Review and Meta-Analysis. BMC Med. 2023, 21, 468. [Google Scholar] [CrossRef]
- Szarpak, L.; Pruc, M.; Filipiak, K.J.; Popieluch, J.; Bielski, A.; Jaguszewski, M.J.; Gilis-Malinowska, N.; Chirico, F.; Afique, Z.R.; Peacock, F.W. Myocarditis: A Complication of COVID-19 and Long-COVID-19 Syndrome as a Serious Threat in Modern Cardiology. Cardiol. J. 2022, 29, 178–179. [Google Scholar] [CrossRef]
- Zuin, M.; Rigatelli, G.; Bilato, C.; Porcari, A.; Merlo, M.; Roncon, L.; Sinagra, G. One-Year Risk of Myocarditis after COVID-19 Infection: A Systematic Review and Meta-Analysis. Can. J. Cardiol. 2023, 39, 839–844. [Google Scholar] [CrossRef] [PubMed]
- Sattar, Y.; Sandhyavenu, H.; Patel, N.; Victor, V.; Patel, D.; Hussain, B.; Titus, A.; Thyagaturu, H.; Alraiyes, M.; Atti, L.; et al. In-Hospital Outcomes of COVID-19 Associated Myocarditis (from a Nationwide Inpatient Sample Database Study). Am. J. Cardiol. 2023, 192, 39–44. [Google Scholar] [CrossRef]
- Okor, I.; Bob-Manuel, T.; Price, J.; Sleem, A.; Amoran, O.; Kelly, J.; Ekerete, M.F.; Bamgbose, M.O.; Bolaji, O.A.; Krim, S.R. COVID-19 Myocarditis: An Emerging Clinical Conundrum. Curr. Probl. Cardiol. 2022, 47, 101268. [Google Scholar] [CrossRef]
- Elseidy, S.A.; Awad, A.K.; Vorla, M.; Fatima, A.; Elbadawy, M.A.; Mandal, D.; Mohamad, T. Cardiovascular Complications in the Post-Acute COVID-19 Syndrome (Pacs). Int. J. Cardiol. Heart Vasc. 2022, 40, 101012. [Google Scholar] [CrossRef] [PubMed]
- Silva, L.S.; Boldrini, V.O.; Marques, A.M.; Yasuda, C.L. Post-COVID Myocarditis with Antiheart Antibodies Persistence: Clues for Autoimmunity? Clin. Cardiol. 2023, 46, 350–351. [Google Scholar] [CrossRef] [PubMed]
- Chang, X.; Ismail, N.I.; Rahman, A.; Xu, D.; Chan, R.W.Y.; Ong, S.-G.; Ong, S.-B. Long COVID-19 and the Heart: Is Cardiac Mitochondria the Missing Link? Antioxid. Redox Signal. 2023, 38, 599–618. [Google Scholar] [CrossRef]
- Naeem, A.; Tabassum, S.; Gill, S.; Khan, M.Z.; Mumtaz, N.; Qaiser, Q.; Karamat, M.; Arif, M.; Naeem, F.; Afifi, A.; et al. COVID-19 and Cardiovascular Diseases: A Literature Review from Pathogenesis to Diagnosis. Cureus 2023, 15, e35658. [Google Scholar] [CrossRef]
- Sun, L.-J.; Qiao, W.; Xiao, Y.-J.; Cui, L.; Wang, X.; Ren, W.-D. Naringin Mitigates Myocardial Strain and the Inflammatory Response in Sepsis-Induced Myocardial Dysfunction through Regulation of PI3K/AKT/NF-κB Pathway. Int. Immunopharmacol. 2019, 75, 105782. [Google Scholar] [CrossRef]
- Li, S.-H.; Ma, G.-L.; Zhang, S.-L.; Yang, Y.-Y.; Liu, H.-F.; Luo, A.; Wen, J.; Cao, Z.-Z.; Jia, Y.-Z. Naringin Exerts Antiarrhythmic Effects by Inhibiting Channel Currents in Mouse Cardiomyocytes. J. Electrocardiol. 2023, 80, 69–80. [Google Scholar] [CrossRef]
- Stewart, I.; Jacob, J.; George, P.M.; Molyneaux, P.L.; Porter, J.C.; Allen, R.J.; Aslani, S.; Baillie, J.K.; Barratt, S.L.; Beirne, P.; et al. Residual Lung Abnormalities after COVID-19 Hospitalization: Interim Analysis of the UKILD Post–COVID-19 Study. Am. J. Respir. Crit. Care Med. 2023, 207, 693–703. [Google Scholar] [CrossRef]
- Farghaly, S.; Badedi, M.; Ibrahim, R.; Sadhan, M.H.; Alamoudi, A.; Alnami, A.; Muhajir, A. Clinical Characteristics and Outcomes of Post-COVID-19 Pulmonary Fibrosis. Medicine 2022, 101, e28639. [Google Scholar] [CrossRef]
- Espín, E.; Yang, C.; Shannon, C.P.; Assadian, S.; He, D.; Tebbutt, S.J. Cellular and Molecular Biomarkers of Long COVID: A Scoping Review. EBioMedicine 2023, 91, 104552. [Google Scholar] [CrossRef]
- Hama Amin, B.J.; Kakamad, F.H.; Ahmed, G.S.; Ahmed, S.F.; Abdulla, B.A.; Mohammed, S.H.; Mikael, T.M.; Salih, R.Q.; Ali, R.k.; Salh, A.M.; et al. Post COVID-19 Pulmonary Fibrosis; a Meta-Analysis Study. Ann. Med. Surg. 2022, 77, 103590. [Google Scholar] [CrossRef]
- Hirawat, R.; Jain, N.; Aslam Saifi, M.; Rachamalla, M.; Godugu, C. Lung Fibrosis: Post-COVID-19 Complications and Evidences. Int. Immunopharmacol. 2023, 116, 109418. [Google Scholar] [CrossRef]
- Patrucco, F.; Solidoro, P.; Gavelli, F.; Apostolo, D.; Bellan, M. Idiopathic Pulmonary Fibrosis and Post-COVID-19 Lung Fibrosis: Links and Risks. Microorganisms 2023, 11, 895. [Google Scholar] [CrossRef]
- George, P.M.; Wells, A.U.; Jenkins, R.G. Pulmonary Fibrosis and COVID-19: The Potential Role for Antifibrotic Therapy. Lancet Respir. Med. 2020, 8, 807–815. [Google Scholar] [CrossRef]
- Wang, R.; Wu, G.; Dai, T.; Lang, Y.; Chi, Z.; Yang, S.; Dong, D. Naringin Attenuates Renal Interstitial Fibrosis by Regulating the TGF-β/Smad Signaling Pathway and Inflammation. Exp. Ther. Med. 2021, 21, 66. [Google Scholar] [CrossRef]
- Wang, Q.; Li, W.; Hu, H.; Lu, X.; Qin, S. Monomeric Compounds from Traditional Chinese Medicine: New Hopes for Drug Discovery in Pulmonary Fibrosis. Biomed. Pharmacother. 2023, 159, 114226. [Google Scholar] [CrossRef]
- Zhang, H.; Zhou, X.; Zhong, Y.; Ji, L.; Yu, W.; Fang, J.; Ying, H.; Li, C. Naringin Suppressed Airway Inflammation and Ameliorated Pulmonary Endothelial Hyperpermeability by Upregulating Aquaporin1 in Lipopolysaccharide/Cigarette Smoke-Induced Mice. Biomed. Pharmacother. 2022, 150, 113035. [Google Scholar] [CrossRef]
- Chen, Y.; Nie, Y.; Luo, Y.; Lin, F.; Zheng, Y.; Cheng, G.; Wu, H.; Zhang, K.; Su, W.; Shen, J.; et al. Protective Effects of Naringin against Paraquat-Induced Acute Lung Injury and Pulmonary Fibrosis in Mice. Food Chem. Toxicol. 2013, 58, 133–140. [Google Scholar] [CrossRef]
- Wei, Y.; Sun, L.; Liu, C.; Li, L. Naringin Regulates Endoplasmic Reticulum Stress and Mitophagy through the ATF3/PINK1 Signaling Axis to Alleviate Pulmonary Fibrosis. Naunyn. Schmiedebergs Arch. Pharmacol. 2023, 396, 1155–1169. [Google Scholar] [CrossRef]
- Fan, W.; Xu, Z.; Zhang, J.; Guan, M.; Zheng, Y.; Wang, Y.; Wu, H.; Su, W.; Li, P. Naringenin Regulates Cigarette Smoke Extract-Induced Extracellular Vesicles from Alveolar Macrophage to Attenuate the Mouse Lung Epithelial Ferroptosis through Activating EV miR-23a-3p/ACSL4 Axis. Phytomedicine 2024, 124, 155256. [Google Scholar] [CrossRef] [PubMed]
- Mahmoud, N.; Radwan, N.; Alkattan, A.; Hassanien, M.; Elkajam, E.; Alqahtani, S.; Haji, A.; Alfaifi, A.; Alfaleh, A.; Alabdulkareem, K. Post-COVID-19 Syndrome: Nature of Symptoms and Associated Factors. Z. Cesundh. Wiss. 2023, 32, 207–212. [Google Scholar] [CrossRef] [PubMed]
- Chudzik, M.; Babicki, M.; Kapusta, J.; Kałuzińska-Kołat, Ż.; Kołat, D.; Jankowski, P.; Mastalerz-Migas, A. Long-COVID Clinical Features and Risk Factors: A Retrospective Analysis of Patients from the STOP-COVID Registry of the poLoCOV Study. Viruses 2022, 14, 1755. [Google Scholar] [CrossRef] [PubMed]
- Fernández-de-las-Peñas, C.; Guijarro, C.; Plaza-Canteli, S.; Hernández-Barrera, V.; Torres-Macho, J. Prevalence of Post-COVID-19 Cough One Year after SARS-CoV-2 Infection: A Multicenter Study. Lung 2021, 199, 249–253. [Google Scholar] [CrossRef] [PubMed]
- Song, W.-J.; Hui, C.K.M.; Hull, J.H.; Birring, S.S.; McGarvey, L.; Mazzone, S.B.; Chung, K.F. Confronting COVID-19-Associated Cough and the Post-COVID Syndrome: Role of Viral Neurotropism, Neuroinflammation, and Neuroimmune Responses. Lancet Respir. Med. 2021, 9, 533–544. [Google Scholar] [CrossRef] [PubMed]
- Nguyen-Ho, L.; Nguyen-Nhu, V.; Tran-Thi, T.-T.; Solomon, J.J. Severe Chronic Cough Relating to Post-COVID-19 Interstitial Lung Disease: A Case Report. Asia Pac. Allergy 2022, 12, e42. [Google Scholar] [CrossRef] [PubMed]
- Rouadi, P.W.; Idriss, S.A.; Bousquet, J.; Laidlaw, T.M.; Azar, C.R.; Al-Ahmad, M.S.; Yañez, A.; Al-Nesf, M.A.Y.; Nsouli, T.M.; Bahna, S.L.; et al. WAO-ARIA Consensus on Chronic Cough—Part II: Phenotypes and Mechanisms of Abnormal Cough Presentation—Updates in COVID-19. World Allergy Organ. J. 2021, 14, 100618. [Google Scholar] [CrossRef] [PubMed]
- Gencer, A.; Caliskaner Ozturk, B.; Borekci, S.; Gemicioglu, B. Bronchodilator Reversibility Testing in Long-Term Cough and Dyspnea after COVID-19 Viral Infection: A Trigger for Asthma? J. Asthma Off. J. Assoc. Care Asthma 2023, 60, 1221–1226. [Google Scholar] [CrossRef]
- Rai, D.K.; Sharma, P.; Karmakar, S.; Thakur, S.; Ameet, H.; Yadav, R.; Gupta, V.B. Approach to Post COVID-19 Persistent Cough: A Narrative Review. Lung India 2023, 40, 149–154. [Google Scholar] [CrossRef]
- Zanasi, A.; Fontana, G.A.; Mutolo, D. Cough: Pathophysiology, Diagnosis and Treatment, 1st ed.; Springer International Publishing: Cham, Switzerland, 2020; ISBN 10:3030485714. [Google Scholar] [CrossRef]
- Gupta, G.; Almalki, W.H.; Kazmi, I.; Fuloria, N.K.; Fuloria, S.; Subramaniyan, V.; Sekar, M.; Singh, S.K.; Chellappan, D.K.; Dua, K. Current Update on the Protective Effect of Naringin in Inflammatory Lung Diseases. EXCLI J. 2022, 21, 573–579. [Google Scholar] [CrossRef]
- Liu, J.; Yao, J.; Zhang, J. Naringenin Attenuates Inflammation in Chronic Obstructive Pulmonary Disease in Cigarette Smoke Induced Mouse Model and Involves Suppression of NF-κB. J. Microbiol. Biotechnol. 2018. [Google Scholar] [CrossRef]
- Jiao, H.; Su, W.; Li, P.; Liao, Y.; Zhou, Q.; Zhu, N.; He, L. Therapeutic Effects of Naringin in a Guinea Pig Model of Ovalbumin-Induced Cough-Variant Asthma. Pulm. Pharmacol. Ther. 2015, 33, 59–65. [Google Scholar] [CrossRef]
- Luo, Y.; Li, P.; Zhang, C.; Zheng, Y.; Wang, S.; Nie, Y.; Zhang, K.; Su, W. Effects of Four Antitussives on Airway Neurogenic Inflammation in a Guinea Pig Model of Chronic Cough Induced by Cigarette Smoke Exposure. Inflamm. Res. 2013, 62, 1053–1061. [Google Scholar] [CrossRef]
- Ni, K.; Guo, J.; Bu, B.; Pan, Y.; Li, J.; Liu, L.; Luo, M.; Deng, L. Naringin as a Plant-Derived Bitter Tastant Promotes Proliferation of Cultured Human Airway Epithelial Cells via Activation of TAS2R Signaling. Phytomedicine 2021, 84, 153491. [Google Scholar] [CrossRef]
- Khunti, K.; Valabhji, J.; Misra, S. Diabetes and the COVID-19 Pandemic. Diabetologia 2023, 66, 255–266. [Google Scholar] [CrossRef]
- Suwanwongse, K.; Shabarek, N. Newly Diagnosed Diabetes Mellitus, DKA, and COVID-19: Causality or Coincidence? A Report of Three Cases. J. Med. Virol. 2021, 93, 1150–1153. [Google Scholar] [CrossRef] [PubMed]
- Ssentongo, P.; Zhang, Y.; Witmer, L.; Chinchilli, V.M.; Ba, D.M. Association of COVID-19 with Diabetes: A Systematic Review and Meta-Analysis. Sci. Rep. 2022, 12, 20191. [Google Scholar] [CrossRef]
- Zhang, T.; Mei, Q.; Zhang, Z.; Walline, J.H.; Liu, Y.; Zhu, H.; Zhang, S. Risk for Newly Diagnosed Diabetes after COVID-19: A Systematic Review and Meta-Analysis. BMC Med. 2022, 20, 444. [Google Scholar] [CrossRef]
- Rahmati, M.; Keshvari, M.; Mirnasuri, S.; Yon, D.K.; Lee, S.W.; Il Shin, J.; Smith, L. The Global Impact of COVID-19 Pandemic on the Incidence of Pediatric New-onset Type 1 Diabetes and Ketoacidosis: A Systematic Review and Meta-analysis. J. Med. Virol. 2022, 94, 5112–5127. [Google Scholar] [CrossRef]
- Lu, J.Y.; Wilson, J.; Hou, W.; Fleysher, R.; Herold, B.C.; Herold, K.C.; Duong, T.Q. Incidence of New-Onset in-Hospital and Persistent Diabetes in COVID-19 Patients: Comparison with Influenza. EBioMedicine 2023, 90, 104487. [Google Scholar] [CrossRef]
- Laurenzi, A.; Caretto, A.; Molinari, C.; Mercalli, A.; Melzi, R.; Nano, R.; Tresoldi, C.; Rovere Querini, P.; Ciceri, F.; Lampasona, V.; et al. No Evidence of Long-Term Disruption of Glycometabolic Control after SARS-CoV-2 Infection. J. Clin. Endocrinol. Metab. 2021, 107, e1009–e1019. [Google Scholar] [CrossRef]
- Cao, H.; Baranova, A.; Wei, X.; Wang, C.; Zhang, F. Bidirectional Causal Associations between Type 2 Diabetes and COVID-19. J. Med. Virol. 2023, 95, e28100. [Google Scholar] [CrossRef] [PubMed]
- Montefusco, L.; Ben Nasr, M.; D’Addio, F.; Loretelli, C.; Rossi, A.; Pastore, I.; Daniele, G.; Abdelsalam, A.; Maestroni, A.; Dell’Acqua, M.; et al. Acute and Long-Term Disruption of Glycometabolic Control after SARS-CoV-2 Infection. Nat. Metab. 2021, 3, 774–785. [Google Scholar] [CrossRef]
- Fotea, S.; Ghiciuc, C.M.; Stefanescu, G.; Cianga, A.L.; Mihai, C.M.; Lupu, A.; Butnariu, L.I.; Starcea, I.M.; Salaru, D.L.; Mocanu, A.; et al. Pediatric COVID-19 and Diabetes: An Investigation into the Intersection of Two Pandemics. Diagnostics 2023, 13, 2436. [Google Scholar] [CrossRef]
- Liu, F.; Long, X.; Zhang, B.; Zhang, W.; Chen, X.; Zhang, Z. ACE2 Expression in Pancreas May Cause Pancreatic Damage after SARS-CoV-2 Infection. Clin. Gastroenterol. Hepatol. 2020, 18, 2128–2130. [Google Scholar] [CrossRef]
- Kusmartseva, I.; Wu, W.; Syed, F.; Van Der Heide, V.; Jorgensen, M.; Joseph, P.; Tang, X.; Candelario-Jalil, E.; Yang, C.; Nick, H.; et al. Expression of SARS-CoV-2 Entry Factors in the Pancreas of Normal Organ Donors and Individuals with COVID-19. Cell Metab. 2020, 32, 1041–1051. [Google Scholar] [CrossRef] [PubMed]
- Hayden, M.R. An Immediate and Long-Term Complication of COVID-19 May Be Type 2 Diabetes Mellitus: The Central Role of β-Cell Dysfunction, Apoptosis and Exploration of Possible Mechanisms. Cells 2020, 9, 2475. [Google Scholar] [CrossRef]
- Birabaharan, M.; Kaelber, D.C.; Pettus, J.H.; Smith, D.M. Risk of New-Onset Type 2 Diabetes Mellitus in 600,055 Persons after COVID-19: A Cohort Study. Diabetes Obes. Metab. 2022, 24, 1176–1179. [Google Scholar] [CrossRef]
- Zhao, T.; Ding, J.; Liang, Z.; Cui, X.; He, K.; Chen, L.; Li, J. When Type II Diabetes Mellitus Meets COVID-19—Identification of the Shared Gene Signatures and Biological Mechanism between the Two Diseases. Eur. J. Clin. Investig. 2023, 53, e13955. [Google Scholar] [CrossRef]
- Rajappa, R.; Sireesh, D.; Salai, M.B.; Ramkumar, K.M.; Sarvajayakesavulu, S.; Madhunapantula, S.V. Treatment with Naringenin Elevates the Activity of Transcription Factor Nrf2 to Protect Pancreatic β-Cells from Streptozotocin-Induced Diabetes in Vitro and in Vivo. Front. Pharmacol. 2018, 9, 1562. [Google Scholar] [CrossRef]
- Lim, Y.J.; Kim, J.H.; Pan, J.H.; Kim, J.K.; Park, T.-S.; Kim, Y.J.; Lee, J.H.; Kim, J.H. Naringin Protects Pancreatic β-Cells against Oxidative Stress-Induced Apoptosis by Inhibiting Both Intrinsic and Extrinsic Pathways in Insulin-Deficient Diabetic Mice. Mol. Nutr. Food Res. 2018, 62, 1700810. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, M.; Thotakura, B.; Chandra Sekaran, S.P.; Jyothi, A.K.; Sundaramurthi, I. Naringin Ameliorates Streptozotocin-Induced Diabetes through Forkhead Box M1-Mediated Beta Cell Proliferation. Cells Tissues Organs 2019, 206, 242–253. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, O.M.; Hassan, M.A.; Abdel-Twab, S.M.; Abdel Azeem, M.N. Navel Orange Peel Hydroethanolic Extract, Naringin and Naringenin Have Anti-Diabetic Potentials in Type 2 Diabetic Rats. Biomed. Pharmacother. 2017, 94, 197–205. [Google Scholar] [CrossRef] [PubMed]
- Murunga, A.N.; Miruka, D.O.; Driver, C.; Nkomo, F.S.; Cobongela, S.Z.Z.; Owira, P.M.O. Grapefruit Derived Flavonoid Naringin Improves Ketoacidosis and Lipid Peroxidation in Type 1 Diabetes Rat Model. PLoS ONE 2016, 11, e0153241. [Google Scholar] [CrossRef] [PubMed]
- Mahmoud, A.M.; Ahmed, O.M.; Ashour, M.B.; Abdel-Moneim, A. In Vivo and in Vitro Antidiabetic Effects of Citrus Flavonoids; a Study on the Mechanism of Action. Int. J. Diabetes Dev. Ctries. 2015, 35, 250–263. [Google Scholar] [CrossRef]
- Yoshida, H.; Watanabe, H.; Ishida, A.; Watanabe, W.; Narumi, K.; Atsumi, T.; Sugita, C.; Kurokawa, M. Naringenin Suppresses Macrophage Infiltration into Adipose Tissue in an Early Phase of High-Fat Diet-Induced Obesity. Biochem. Biophys. Res. Commun. 2014, 454, 95–101. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.; Bansal, S.; Kuhad, A.; Kumar, A.; Chopra, K. Naringenin Ameliorates Diabetic Neuropathic Pain by Modulation of Oxidative-Nitrosative Stress, Cytokines and MMP-9 Levels. Food Funct. 2020, 11, 4548–4560. [Google Scholar] [CrossRef] [PubMed]
- He, W.; Wang, Y.; Yang, R.; Ma, H.; Qin, X.; Yan, M.; Rong, Y.; Xie, Y.; Li, L.; Si, J.; et al. Molecular Mechanism of Naringenin against High-Glucose-Induced Vascular Smooth Muscle Cells Proliferation and Migration Based on Network Pharmacology and Transcriptomic Analyses. Front. Pharmacol. 2022, 13, 862709. [Google Scholar] [CrossRef]
- Fiala, K.; Martens, J.; Abd-Elsayed, A. Post-COVID Pain Syndromes. Curr. Pain Headache Rep. 2022, 26, 379–383. [Google Scholar] [CrossRef]
- Malik, P.; Patel, K.; Pinto, C.; Jaiswal, R.; Tirupathi, R.; Pillai, S.; Patel, U. Post-acute COVID-19 Syndrome (PCS) and Health-related Quality of Life (HRQoL)—A Systematic Review and Meta-analysis. J. Med. Virol. 2022, 94, 253–262. [Google Scholar] [CrossRef]
- Fernández-de-las-Peñas, C.; Navarro-Santana, M.; Plaza-Manzano, G.; Palacios-Ceña, D.; Arendt-Nielsen, L. Time Course Prevalence of Post-COVID Pain Symptoms of Musculoskeletal Origin in Patients Who Had Survived Severe Acute Respiratory Syndrome Coronavirus 2 Infection: A Systematic Review and Meta-Analysis. Pain 2022, 163, 1220. [Google Scholar] [CrossRef]
- Natarajan, A.; Shetty, A.; Delanerolle, G.; Zeng, Y.; Zhang, Y.; Raymont, V.; Rathod, S.; Halabi, S.; Elliot, K.; Shi, J.Q.; et al. A Systematic Review and Meta-Analysis of Long COVID Symptoms. Syst. Rev. 2023, 12, 88. [Google Scholar] [CrossRef]
- Wu, Y.; Xu, X.; Chen, Z.; Duan, J.; Hashimoto, K.; Yang, L.; Liu, C.; Yang, C. Nervous System Involvement after Infection with COVID-19 and Other Coronaviruses. Brain Behav. Immun. 2020, 87, 18–22. [Google Scholar] [CrossRef] [PubMed]
- Bussmann, A.J.C.; Ferraz, C.R.; Lima, A.V.A.; Castro, J.G.S.; Ritter, P.D.; Zaninelli, T.H.; Saraiva-Santos, T.; Verri, W.A.; Borghi, S.M. Association between IL-10 Systemic Low Level and Highest Pain Score in Patients during Symptomatic SARS-CoV-2 Infection. Pain Pract. Pain 2022, 22, 453–462. [Google Scholar] [CrossRef] [PubMed]
- Rani, M.; Uniyal, A.; Akhilesh; Tiwari, V. Decrypting the Cellular and Molecular Intricacies Associated with COVID-19-Induced Chronic Pain. Metab. Brain Dis. 2022, 37, 2629–2642. [Google Scholar] [CrossRef] [PubMed]
- Takeshita, H.; Yamamoto, K. Tryptophan Metabolism and COVID-19-Induced Skeletal Muscle Damage: Is ACE2 a Key Regulator? Front. Nutr. 2022, 9, 868845. [Google Scholar] [CrossRef] [PubMed]
- Tana, C.; Bentivegna, E.; Cho, S.-J.; Harriott, A.M.; García-Azorín, D.; Labastida-Ramirez, A.; Ornello, R.; Raffaelli, B.; Beltrán, E.R.; Ruscheweyh, R.; et al. Long COVID Headache. J. Headache Pain 2022, 23, 93. [Google Scholar] [CrossRef] [PubMed]
- Bolay, H.; Karadas, Ö.; Oztürk, B.; Sonkaya, R.; Tasdelen, B.; Bulut, T.D.S.; Gülbahar, Ö.; Özge, A.; Baykan, B. HMGB1, NLRP3, IL-6 and ACE2 Levels Are Elevated in COVID-19 with Headache: A Window to the Infection-Related Headache Mechanism. J. Headache Pain 2021, 22, 94. [Google Scholar] [CrossRef] [PubMed]
- Caronna, E.; Ballvé, A.; Llauradó, A.; Gallardo, V.J.; Ariton, D.M.; Lallana, S.; López Maza, S.; Olivé Gadea, M.; Quibus, L.; Restrepo, J.L.; et al. Headache: A Striking Prodromal and Persistent Symptom, Predictive of COVID-19 Clinical Evolution. Cephalalgia 2020, 40, 1410–1421. [Google Scholar] [CrossRef]
- Xu, Q.; Zhang, Z.; Sun, W. Effect of Naringin on Monosodium Iodoacetate-Induced Osteoarthritis Pain in Rats. Med. Sci. Monit. 2017, 23, 3746–3751. [Google Scholar] [CrossRef]
- Hu, C.; Zhao, Y.-T. Analgesic Effects of Naringenin in Rats with Spinal Nerve Ligation-Induced Neuropathic Pain. Biomed. Rep. 2014, 2, 569–573. [Google Scholar] [CrossRef]
- Aghazadeh Tabrizi, M.; Baraldi, P.G.; Baraldi, S.; Gessi, S.; Merighi, S.; Borea, P.A. Medicinal Chemistry, Pharmacology, and Clinical Implications of TRPV1 Receptor Antagonists. Med. Res. Rev. 2017, 37, 936–983. [Google Scholar] [CrossRef]
- Katz, B.; Zaguri, R.; Edvardson, S.; Maayan, C.; Elpeleg, O.; Lev, S.; Davidson, E.; Peters, M.; Kfir-Erenfeld, S.; Berger, E.; et al. Nociception and Pain in Humans Lacking a Functional TRPV1 Channel. J. Clin. Investig. 2023, 133, e153558. [Google Scholar] [CrossRef]
- Gouin, O.; L’Herondelle, K.; Lebonvallet, N.; Le Gall-Ianotto, C.; Sakka, M.; Buhé, V.; Plée-Gautier, E.; Carré, J.-L.; Lefeuvre, L.; Misery, L.; et al. TRPV1 and TRPA1 in Cutaneous Neurogenic and Chronic Inflammation: Pro-Inflammatory Response Induced by Their Activation and Their Sensitization. Protein Cell 2017, 8, 644–661. [Google Scholar] [CrossRef]
- Eom, S.; Lee, B.-B.; Lee, S.; Park, Y.; Yeom, H.D.; Kim, T.-H.; Nam, S.-H.; Lee, J.H. Antioxidative and Analgesic Effects of Naringin through Selective Inhibition of Transient Receptor Potential Vanilloid Member 1. Antioxidants 2021, 11, 64. [Google Scholar] [CrossRef] [PubMed]
- Straub, I.; Mohr, F.; Stab, J.; Konrad, M.; Philipp, S.; Oberwinkler, J.; Schaefer, M. Citrus Fruit and Fabacea Secondary Metabolites Potently and Selectively Block TRPM3. Br. J. Pharmacol. 2013, 168, 1835–1850. [Google Scholar] [CrossRef] [PubMed]
- Manchope, M.F.; Calixto-Campos, C.; Coelho-Silva, L.; Zarpelon, A.C.; Pinho-Ribeiro, F.A.; Georgetti, S.R.; Baracat, M.M.; Casagrande, R.; Verri, W.A. Naringenin Inhibits Superoxide Anion-Induced Inflammatory Pain: Role of Oxidative Stress, Cytokines, Nrf-2 and the NO−cGMP−PKG−KATP Channel Signaling Pathway. PLoS ONE 2016, 11, e0153015. [Google Scholar] [CrossRef] [PubMed]
- Pourmasumi, S.; Kounis, N.G.; Naderi, M.; Hosseinisadat, R.; Khoradmehr, A.; Fagheirelahee, N.; Kouni, S.N.; de Gregorio, C.; Dousdampanis, P.; Mplani, V.; et al. Effects of COVID-19 Infection and Vaccination on the Female Reproductive System: A Narrative Review. Balk. Med. J. 2023, 40, 153–164. [Google Scholar] [CrossRef] [PubMed]
- Nassau, D.E.; Best, J.C.; Kresch, E.; Gonzalez, D.C.; Khodamoradi, K.; Ramasamy, R. Impact of the SARS-CoV-2 Virus on Male Reproductive Health. Bju Int. 2022, 129, 143–150. [Google Scholar] [CrossRef] [PubMed]
- Yelin, D.; Daitch, V.; Kalfon, T.; Mor, M.; Buchrits, S.; Shafir, Y.; Awwad, M.; Ghantous, N.; Shapira-Lichter, I.; Leibovici, L.; et al. Long COVID Sexual Dysfunction among Both Genders: Evaluation of a Cohort of COVID-19 Recoverees. Infect. Dis. Now 2023, 53, 104750. [Google Scholar] [CrossRef] [PubMed]
- Harirugsakul, K.; Wainipitapong, S.; Phannajit, J.; Paitoonpong, L.; Tantiwongse, K. Erectile Dysfunction after COVID-19 Recovery: A Follow-up Study. PLoS ONE 2022, 17, e0276429. [Google Scholar] [CrossRef] [PubMed]
- Pollack, B.; Von Saltza, E.; McCorkell, L.; Santos, L.; Hultman, A.; Cohen, A.K.; Soares, L. Female Reproductive Health Impacts of Long COVID and Associated Illnesses Including ME/CFS, POTS, and Connective Tissue Disorders: A Literature Review. Front. Rehabil. Sci. 2023, 4, 1122673. [Google Scholar] [CrossRef] [PubMed]
- Bechmann, N.; Maccio, U.; Kotb, R.; Dweik, R.A.; Cherfane, M.; Moch, H.; Bornstein, S.R.; Varga, Z. COVID-19 Infections in Gonads: Consequences on Fertility? Horm. Metab. Res. 2022, 54, 549–555. [Google Scholar] [CrossRef] [PubMed]
- Corona, G.; Baldi, E.; Isidori, A.M.; Paoli, D.; Pallotti, F.; De Santis, L.; Francavilla, F.; La Vignera, S.; Selice, R.; Caponecchia, L.; et al. SARS-CoV-2 Infection, Male Fertility and Sperm Cryopreservation: A Position Statement of the Italian Society of Andrology and Sexual Medicine (SIAMS) (Società Italiana Di Andrologia e Medicina Della Sessualità). J. Endocrinol. Investig. 2020, 43, 1153–1157. [Google Scholar] [CrossRef] [PubMed]
- Sengupta, P.; Dutta, S.; Roychoudhury, S.; D’Souza, U.J.A.; Govindasamy, K.; Kolesarova, A. COVID-19, Oxidative Stress and Male Reproduction: Possible Role of Antioxidants. Antioxidants 2022, 11, 548. [Google Scholar] [CrossRef] [PubMed]
- Sauve, F.; Nampoothiri, S.; Clarke, S.A.; Fernandois, D.; Ferreira Coêlho, C.F.; Dewisme, J.; Mills, E.G.; Ternier, G.; Cotellessa, L.; Iglesias-Garcia, C.; et al. Long-COVID Cognitive Impairments and Reproductive Hormone Deficits in Men May Stem from GnRH Neuronal Death. EBioMedicine 2023, 96, 104784. [Google Scholar] [CrossRef]
- Al-kuraishy, H.M.; Al-Gareeb, A.I.; Alarfaj, S.J.; Al-Akeel, R.K.; Faidah, H.; El-Bouseary, M.M.; Sabatier, J.-M.; De Waard, M.; El-Masry, T.A.; Batiha, G.E.-S. Long COVID and Risk of Erectile Dysfunction in Recovered Patients from Mild to Moderate COVID-19. Sci. Rep. 2023, 13, 5977. [Google Scholar] [CrossRef]
- Li, H.; Xiao, X.; Zhang, J.; Zafar, M.I.; Wu, C.; Long, Y.; Lu, W.; Pan, F.; Meng, T.; Zhao, K.; et al. Impaired Spermatogenesis in COVID-19 Patients. EClinicalMedicine 2020, 28, 100604. [Google Scholar] [CrossRef]
- Chen, H.; Chen, J.; Shi, X.; Li, L.; Xu, S. Naringenin Protects Swine Testis Cells from Bisphenol A-Induced Apoptosis via Keap1/Nrf2 Signaling Pathway. BioFactors 2022, 48, 190–203. [Google Scholar] [CrossRef]
- Adana, M.Y.; Akang, E.N.; Peter, A.I.; Jegede, A.I.; Naidu, E.C.S.; Tiloke, C.; Chuturgoon, A.A.; Azu, O.O. Naringenin Attenuates Highly Active Antiretroviral Therapy-Induced Sperm DNA Fragmentations and Testicular Toxicity in Sprague-Dawley Rats. Andrology 2018, 6, 166–175. [Google Scholar] [CrossRef]
- Rashid, R.; Tripathi, R.; Singh, A.; Sarkar, S.; Kawale, A.; Bader, G.N.; Gupta, S.; Gupta, R.K.; Jha, R.K. Naringenin Improves Ovarian Health by Reducing the Serum Androgen and Eliminating Follicular Cysts in Letrozole-Induced Polycystic Ovary Syndrome in the Sprague Dawley Rats. Phytother. Res. 2023, 37, 4018–4041. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Xiang, M.; Jing, H.; Wang, C.; Novakovic, V.A.; Shi, J. Damage to Endothelial Barriers and Its Contribution to Long COVID. Angiogenesis 2023, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Candeloro, M.; Schulman, S. Arterial Thrombotic Events in Hospitalized COVID-19 Patients: A Short Review and Meta-Analysis. Semin. Thromb. Hemost. 2023, 49, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Di Minno, A.; Ambrosino, P.; Calcaterra, I.; Di Minno, M.N.D. COVID-19 and Venous Thromboembolism: A Meta-Analysis of Literature Studies. Semin. Thromb. Hemost. 2020, 46, 763–771. [Google Scholar] [CrossRef] [PubMed]
- Zuin, M.; Engelen, M.M.; Barco, S.; Spyropoulos, A.C.; Vanassche, T.; Hunt, B.J.; Vandenbriele, C.; Verhamme, P.; Kucher, N.; Rashidi, F.; et al. Incidence of Venous Thromboembolic Events in COVID-19 Patients after Hospital Discharge: A Systematic Review and Meta-Analysis. Thromb. Res. 2022, 209, 94–98. [Google Scholar] [CrossRef]
- Martins-Gonçalves, R.; Hottz, E.D.; Bozza, P.T. Acute to Post-Acute COVID-19 Thromboinflammation Persistence: Mechanisms and Potential Consequences. Curr. Res. Immunol. 2023, 4, 100058. [Google Scholar] [CrossRef]
- Grobbelaar, L.M.; Venter, C.; Vlok, M.; Ngoepe, M.; Laubscher, G.J.; Lourens, P.J.; Steenkamp, J.; Kell, D.B.; Pretorius, E. SARS-CoV-2 Spike Protein S1 Induces Fibrin(Ogen) Resistant to Fibrinolysis: Implications for Microclot Formation in COVID-19. Biosci. Rep. 2021, 41, BSR20210611. [Google Scholar] [CrossRef]
- Schultheiß, C.; Willscher, E.; Paschold, L.; Gottschick, C.; Klee, B.; Henkes, S.-S.; Bosurgi, L.; Dutzmann, J.; Sedding, D.; Frese, T.; et al. The IL-1β, IL-6, and TNF Cytokine Triad Is Associated with Post-Acute Sequelae of COVID-19. Cell Rep. Med. 2022, 3, 100663. [Google Scholar] [CrossRef]
- Turner, S.; Khan, M.A.; Putrino, D.; Woodcock, A.; Kell, D.B.; Pretorius, E. Long COVID: Pathophysiological Factors and Abnormalities of Coagulation. Trends Endocrinol. Metab. 2023, 34, 321. [Google Scholar] [CrossRef]
- Zaira, B.; Yulianti, T.; Levita, J. Correlation between Hepatocyte Growth Factor (HGF) with D-Dimer and Interleukin-6 as Prognostic Markers of Coagulation and Inflammation in Long COVID-19 Survivors. Curr. Issues Mol. Biol. 2023, 45, 5725–5740. [Google Scholar] [CrossRef]
- Malakul, W.; Pengnet, S.; Kumchoom, C.; Tunsophon, S. Naringin Ameliorates Endothelial Dysfunction in Fructose-Fed Rats. Exp. Ther. Med. 2018, 15, 3140–3146. [Google Scholar] [CrossRef]
- Zhao, H.; Liu, M.; Liu, H.; Suo, R.; Lu, C. Naringin Protects Endothelial Cells from Apoptosis and Inflammation by Regulating the Hippo-YAP Pathway. Biosci. Rep. 2020, 40, BSR20193431. [Google Scholar] [CrossRef]
- Wang, K.; Peng, S.; Xiong, S.; Niu, A.; Xia, M.; Xiong, X.; Zeng, G.; Huang, Q. Naringin Inhibits Autophagy Mediated by PI3K-Akt-mTOR Pathway to Ameliorate Endothelial Cell Dysfunction Induced by High Glucose/High Fat Stress. Eur. J. Pharmacol. 2020, 874, 173003. [Google Scholar] [CrossRef]
- Wright, B.; Spencer, J.P.E.; Lovegrove, J.A.; Gibbins, J.M. Flavonoid Inhibitory Pharmacodynamics on Platelet Function in Physiological Environments. Food Funct. 2013, 4, 1803–1810. [Google Scholar] [CrossRef]
- Huang, M.; Deng, M.; Nie, W.; Zou, D.; Wu, H.; Xu, D. Naringenin Inhibits Platelet Activation and Arterial Thrombosis through Inhibition of Phosphoinositide 3-Kinase and Cyclic Nucleotide Signaling. Front. Pharmacol. 2021, 12, 722257. [Google Scholar] [CrossRef]
- Li, Q.-Q.; Yang, Y.-X.; Qv, J.-W.; Hu, G.; Hu, Y.-J.; Xia, Z.-N.; Yang, F.-Q. Investigation of Interactions between Thrombin and Ten Phenolic Compounds by Affinity Capillary Electrophoresis and Molecular Docking. J. Anal. Methods Chem. 2018, 2018, 4707609. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.B.; Siddiqui, U.; Fatima, S.; Rehman, A.A.; Jairajpuri, M.A. Naringin Binds to Protein Disulfide Isomerase to Inhibit Its Activity and Modulate the Blood Coagulation Rates: Implications in Controlling Thrombosis. Int. J. Biol. Macromol. 2023, 252, 126241. [Google Scholar] [CrossRef] [PubMed]
- Tsilingiris, D.; Vallianou, N.G.; Karampela, I.; Christodoulatos, G.S.; Papavasileiou, G.; Petropoulou, D.; Magkos, F.; Dalamaga, M. Laboratory Findings and Biomarkers in Long COVID: What Do We Know so Far? Insights into Epidemiology, Pathogenesis, Therapeutic Perspectives and Challenges. Int. J. Mol. Sci. 2023, 24, 10458. [Google Scholar] [CrossRef] [PubMed]
- Kenny, G.; Townsend, L.; Savinelli, S.; Mallon, P.W.G. Long COVID: Clinical Characteristics, Proposed Pathogenesis and Potential Therapeutic Targets. Front. Mol. Biosci. 2023, 10, 1157651. [Google Scholar] [CrossRef] [PubMed]
- Bonilla, H.; Peluso, M.J.; Rodgers, K.; Aberg, J.A.; Patterson, T.F.; Tamburro, R.; Baizer, L.; Goldman, J.D.; Rouphael, N.; Deitchman, A.; et al. Therapeutic Trials for Long COVID-19: A Call to Action from the Interventions Taskforce of the RECOVER Initiative. Front. Immunol. 2023, 14, 1129459. [Google Scholar] [CrossRef] [PubMed]
- Chee, Y.J.; Fan, B.E.; Young, B.E.; Dalan, R.; Lye, D.C. Clinical Trials on the Pharmacological Treatment of Long COVID: A Systematic Review. J. Med. Virol. 2023, 95, e28289. [Google Scholar] [CrossRef] [PubMed]
- Calabrese, E.J.; Pressman, P.; Hayes, A.W.; Dhawan, G.; Kapoor, R.; Agathokleous, E.; Manes, P.; Calabrese, V. Naringin Commonly Acts via Hormesis. Sci. Total Environ. 2023, 896, 164728. [Google Scholar] [CrossRef] [PubMed]
- Davatgaran Taghipour, Y.; Hajialyani, M.; Naseri, R.; Hesari, M.; Mohammadi, P.; Stefanucci, A.; Mollica, A.; Farzaei, M.H.; Abdollahi, M. Nanoformulations of Natural Products for Management of Metabolic Syndrome. Int. J. Nanomed. 2019, 14, 5303–5321. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Long COVID symptoms.

Figure 2.
Chemical structures of flavanone, naringenin, and its derivatives. The 2D structure images were obtained from PubChem (https://pubchem.ncbi.nlm.nih.gov).
Figure 2.
Chemical structures of flavanone, naringenin, and its derivatives. The 2D structure images were obtained from PubChem (https://pubchem.ncbi.nlm.nih.gov).

Figure 3.
Summary of long COVID symptom pathogenesis (left) and potential therapeutic mechanisms (right). Upward arrows (↑) in the colored panels of the table indicate increases and downward arrows (↓) indicate decreases. Leftward arrows (←) between the panels indicate the palliative and therapeutic effects of naringin on long COVID symptoms.
Figure 3.
Summary of long COVID symptom pathogenesis (left) and potential therapeutic mechanisms (right). Upward arrows (↑) in the colored panels of the table indicate increases and downward arrows (↓) indicate decreases. Leftward arrows (←) between the panels indicate the palliative and therapeutic effects of naringin on long COVID symptoms.


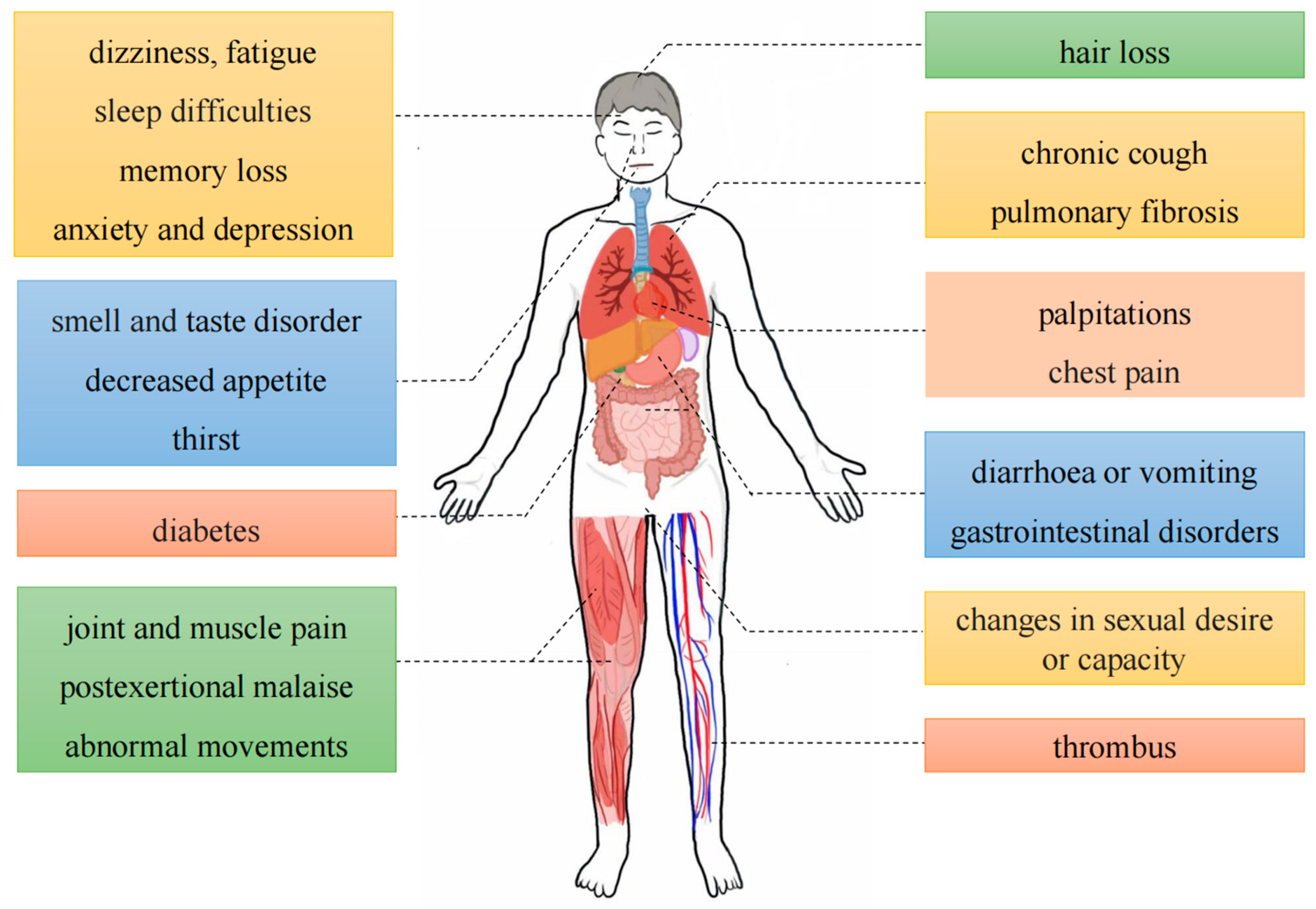{kind=link}
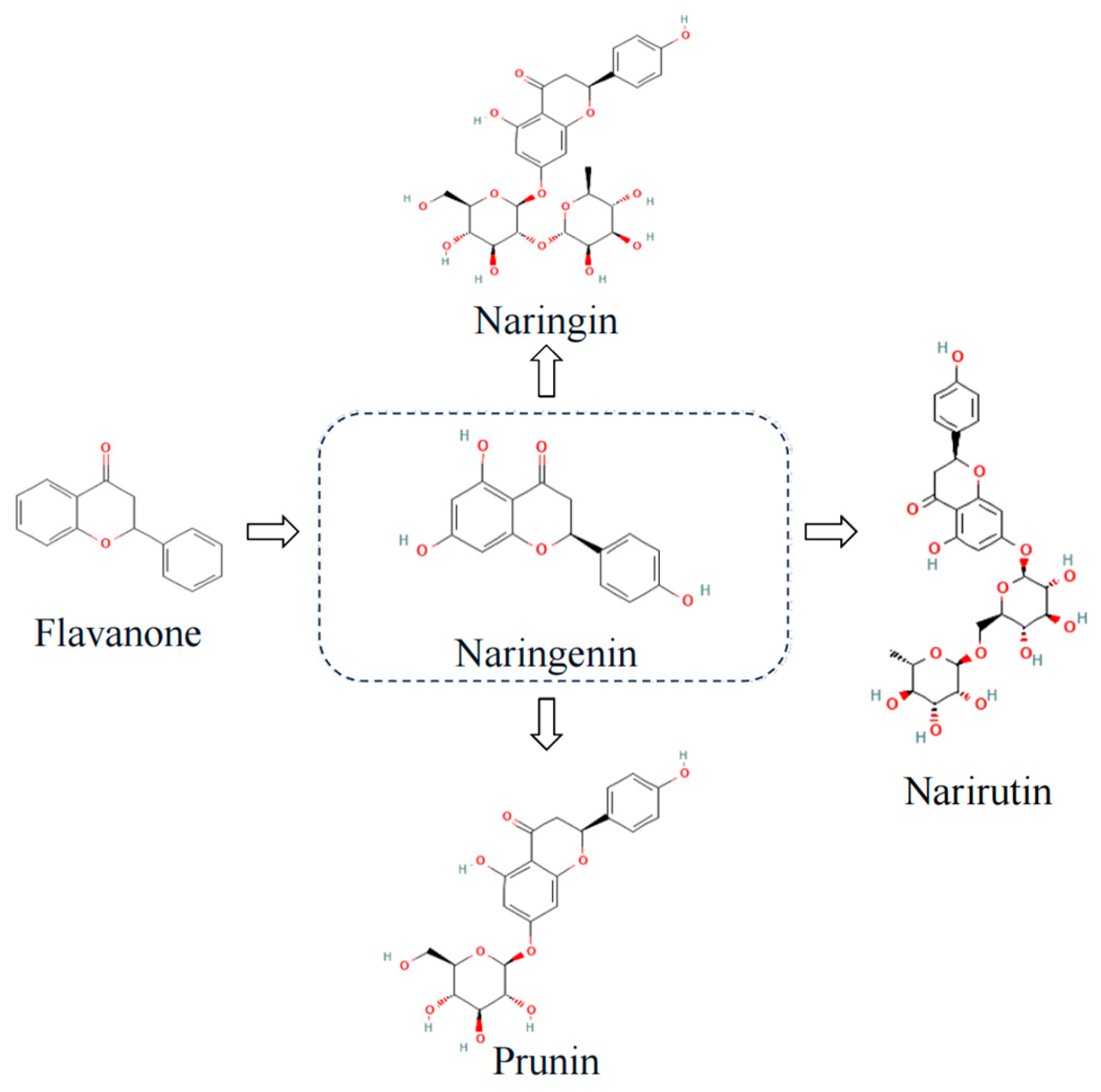{kind=link}
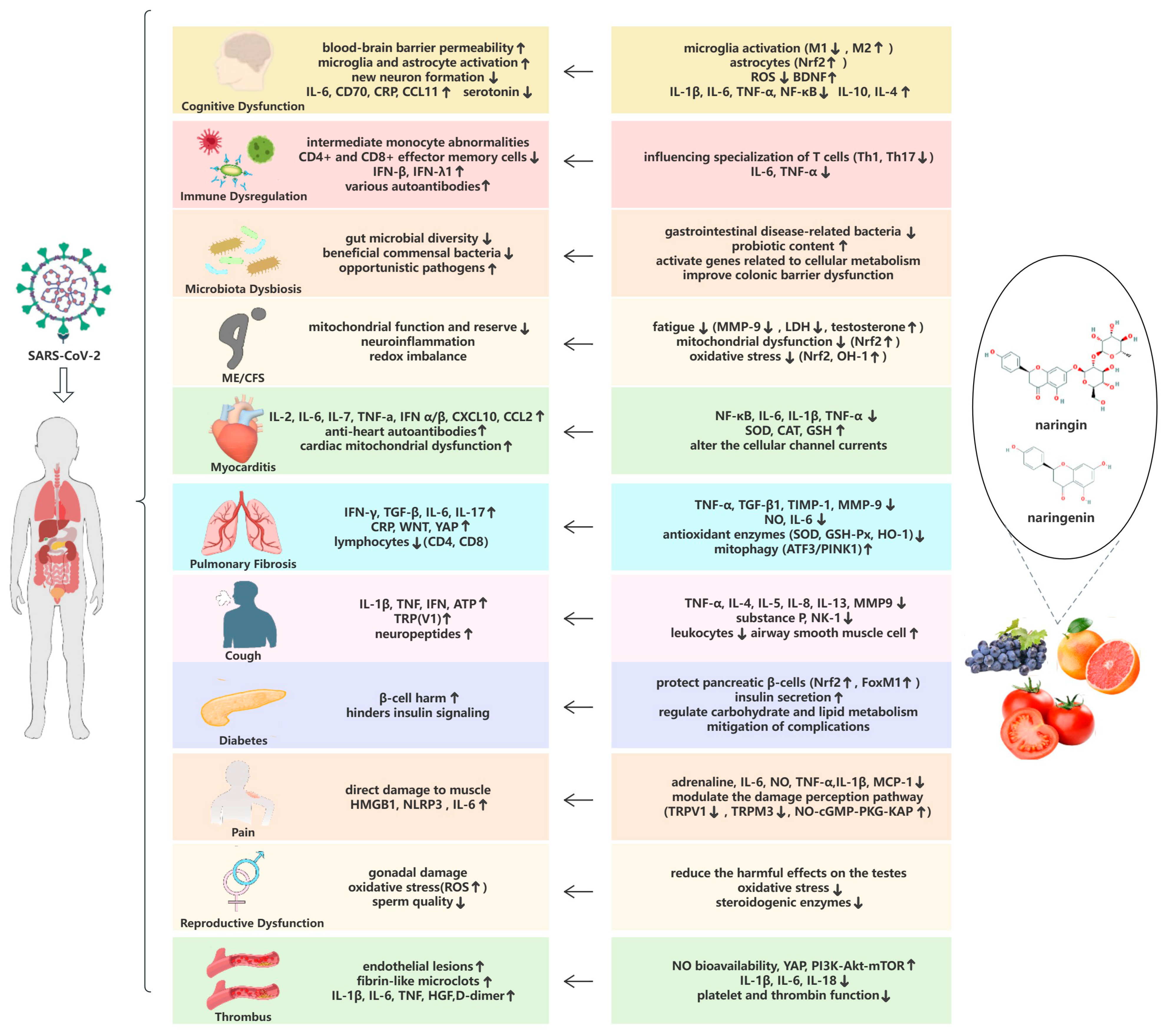{kind=link}
Table 1.
Potential pathophysiological mechanisms of long COVID symptoms and the functions of naringin and/or naringenin.
Table 1.
Potential pathophysiological mechanisms of long COVID symptoms and the functions of naringin and/or naringenin.
| Long COVID Symptoms | Potential Pathophysiological Mechanisms of Long COVID | Functions of Naringin and/or Naringenin |
|---|---|---|
| Cognitive Dysfunction | Interference with blood–brain barrier led to neuronal damage [56,57] Downregulation of new neuron formation [59] Pro-inflammatory factors led to depression [60] Direct damage to the olfactory epithelium [63] | Modulation of glial cell activation [69,71,72] Enhanced synaptic plasticity [73] Protected the nervous system from oxidative stress and inflammation [77] Inhibited hippocampal apoptosis [80] Enhanced the endocrine nervous system [81] |
| Immune Dysregulation | Intermediated monocyte abnormalities and T-cell activation [87,88] Decline in CD4+ and CD8+ effector memory cells [22] Lack of dendritic cells [90] Reduction in non-classical monocyte and lymphocyte subsets [91] Increased levels of autoantibodies [93] | Influenced growth and specialization of T cells [98,99,100,101,102] Regulated the transcription and lysosomal degradation of inflammatory factors [103,104] |
| Microbiota Dysbiosis | Changes in the composition and abundance of the gut [105], lung [109], oral [110], and nasopharyngeal microbiomes [111] Altered basal metabolite levels [114] | Controlled the intestinal microenvironment [120] and colonic barrier [124]. Regulated the intestinal microbiota [121] Regulated gene expression pattern of intestinal commensal microorganisms [122] |
| ME/CFS | Impacted both mitochondrial function and reserves [131,132,133] Infiltration of amyloid-containing deposits [134] | Stabilization of energy sources [139,140,141] Reduced mitochondrial dysfunction [142,143,144] Attenuated oxidative stress [145,146] |
| Myocarditis | Inflammation of cardiomyocytes through direct action of SARS-CoV-2 [153] Hypoxemia [154] High levels of antiheart autoantibodies [155] Cardiac mitochondrial dysfunction [156] | Reduced the inflammatory factors associated with cardiovascular injury [158] Protected cardiac mitochondria from oxidative damage [67] Altered the cellular channel currents [159] |
| Pulmonary Fibrosis | Alveolar injury led to secretion of pro-fibrotic and pro-inflammatory cytokines [164,165] | Inhibition of TGF-β overexpression [167] Reduced the infiltration of inflammatory cells [169] Regulated collagen formation [170] Enhanced mitophagy in lung tissues [171] Regulated miRNAs in extracellular vesicles [172] |
| Cough | Neuroinflammatory mediators led to hypersensitivity of the cough pathway [176] | Inhibited the production of pro-inflammatory factors [183,184] Attenuated airway neurogenic inflammation [185] Induced relaxation of airway smooth muscle cells [186] |
| Diabetes | Autoimmune hyperactivation in the body [196] β-cell harm and pancreatic damage [197,199] Steroids impacted insulin sensitivity [200] | Attenuated pancreatic β-cell damage or activated their proliferation [202,203,204] Ameliorative effect on insulin resistance [205,206] Regulated carbohydrate and lipid metabolism [207,208] Mitigated effect on the complications of diabetes [209,210] |
| Pain | Direct damage to muscle by the virus [218] Inflammation led to overstimulation of the nervous system [219,220,221] | Attenuated neuronal stimulation by inhibiting the inflammatory response [222,223] Modulated the damage perception pathway [227,228,229] |
| Reproductive Dysfunction | Virus-induced gonadal damage [235] Coagulation abnormalities and endothelial damage [236] Decreased testosterone [238] Decrease in sperm quality caused by inflammatory response [240] | Inhibition of oxidative stress and mitochondrial apoptosis in testes [241] Preserved the structure of the testes and sperm viability [242] Inhibited steroidogenic enzymes and controlled ovulation [243] |
| Thrombus Formation | Endothelial damage and dysfunction [244] Formation of fibrin-like microclots that are difficult to hydrolyze [249] Inflammatory factor storm activated exogenous coagulation pathways [250] Formation of autoantibodies and chronic hypoxia [251] | Upregulated NO bioavailability [253] Decreasing levels of inflammatory factors [254] Inhibited excessive autophagy in endothelial cells [255] Inhibited platelet activation [256,257] Inhibited thrombin activity [258,259] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Liu, S.; Zhong, M.; Wu, H.; Su, W.; Wang, Y.; Li, P. Potential Beneficial Effects of Naringin and Naringenin on Long COVID—A Review of the Literature. Microorganisms 2024, 12, 332. https://doi.org/10.3390/microorganisms12020332
AMA Style
Liu S, Zhong M, Wu H, Su W, Wang Y, Li P. Potential Beneficial Effects of Naringin and Naringenin on Long COVID—A Review of the Literature. Microorganisms. 2024; 12(2):332. https://doi.org/10.3390/microorganisms12020332
Chicago/Turabian StyleLiu, Siqi, Mengli Zhong, Hao Wu, Weiwei Su, Yonggang Wang, and Peibo Li. 2024. "Potential Beneficial Effects of Naringin and Naringenin on Long COVID—A Review of the Literature" Microorganisms 12, no. 2: 332. https://doi.org/10.3390/microorganisms12020332
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.