
Molecular Epidemiology, Antimicrobial Susceptibility, and Clinical Features of Methicillin-Resistant Staphylococcus aureus Bloodstream Infections over 30 Years in Barcelona, Spain (1990–2019)
 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Setting, Definitions, and Clinical Data
2.2. Bacterial Isolates and Antibiotic Susceptibility Testing
2.3. Molecular Typing, Whole-Genome Sequencing, and Bioinformatic Analysis
2.4. Statistical Analysis
2.5. Ethics
3. Results
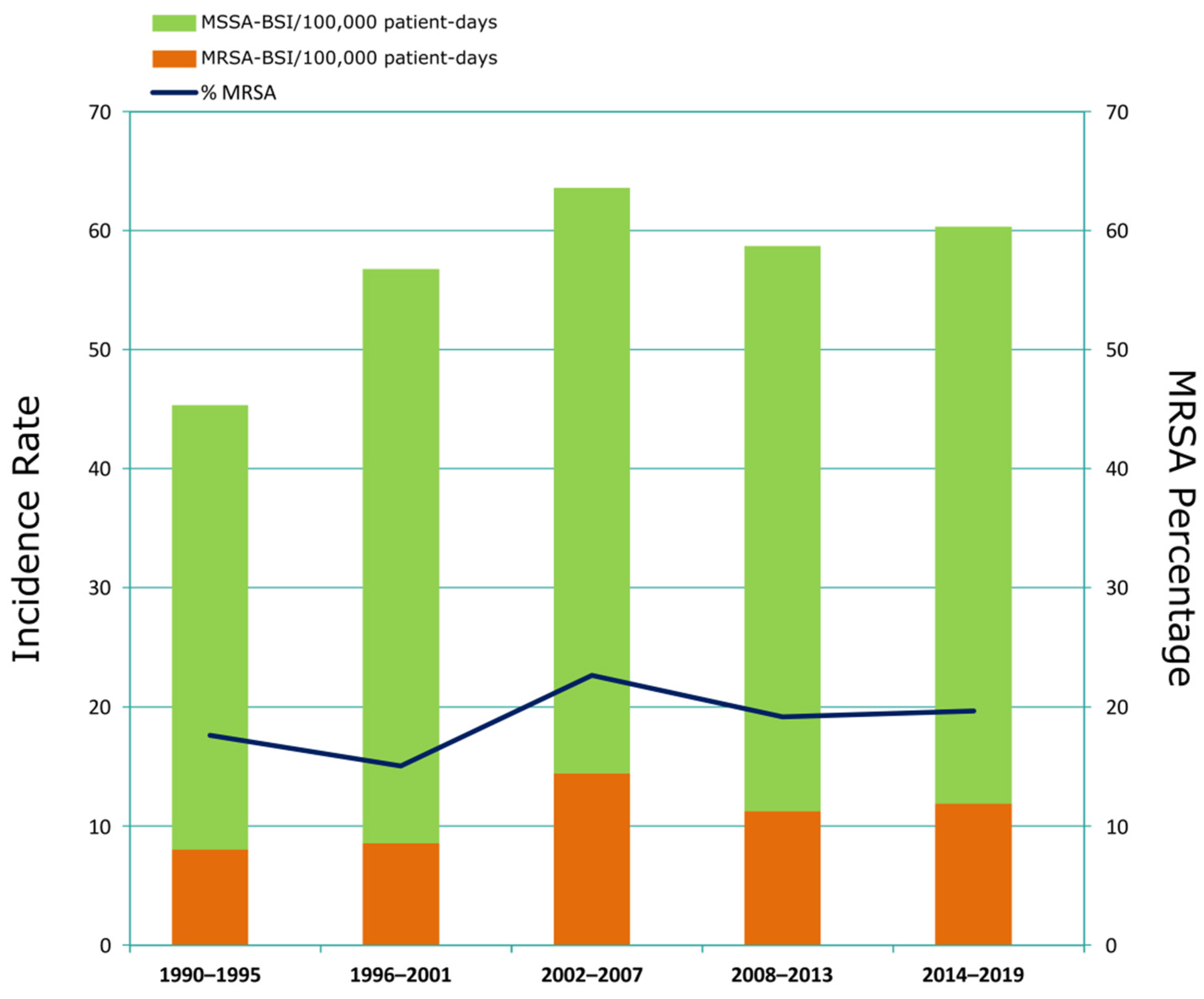
3.1. Incidence and Clinical Characteristics of MRSA-BSI Episodes
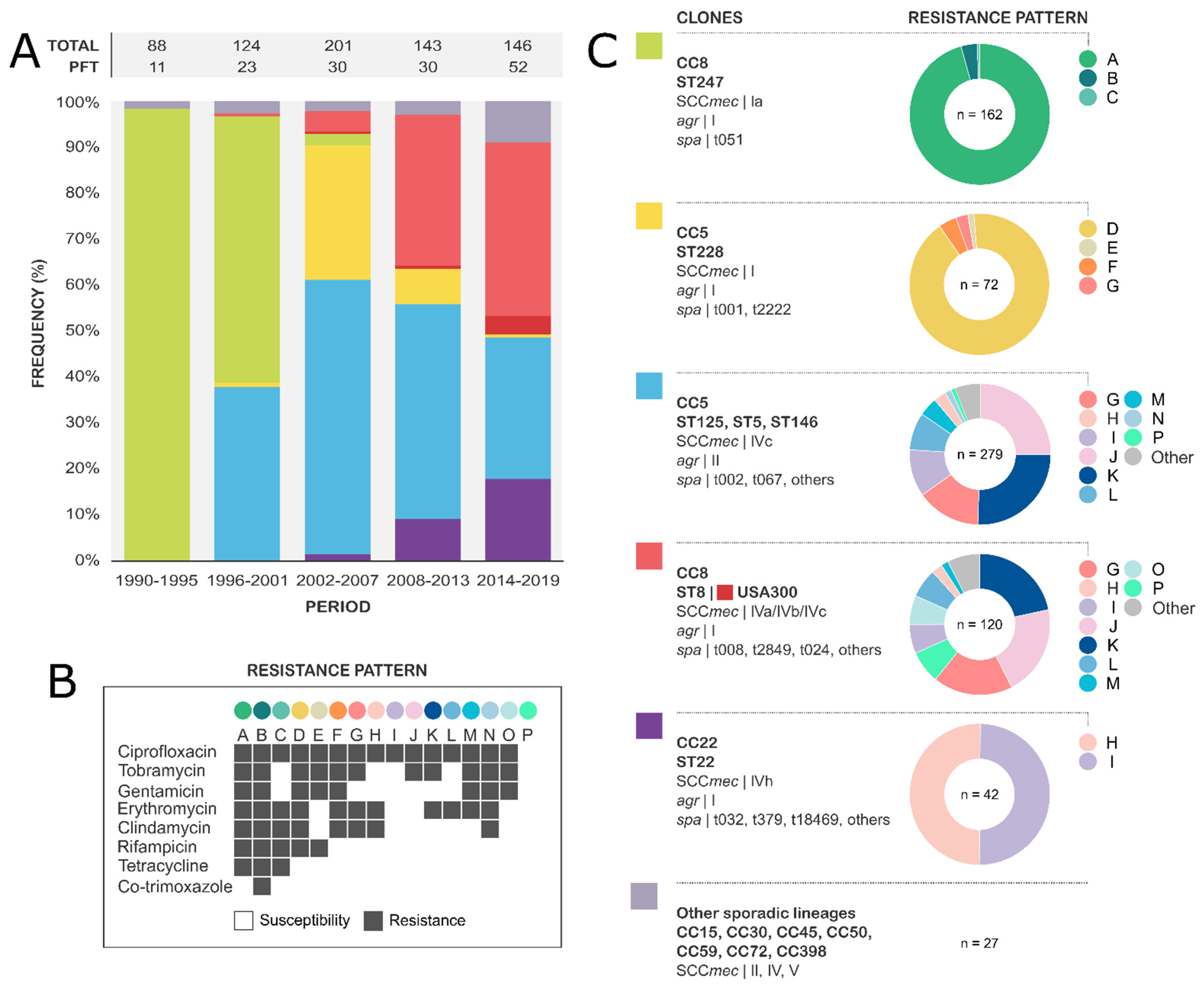
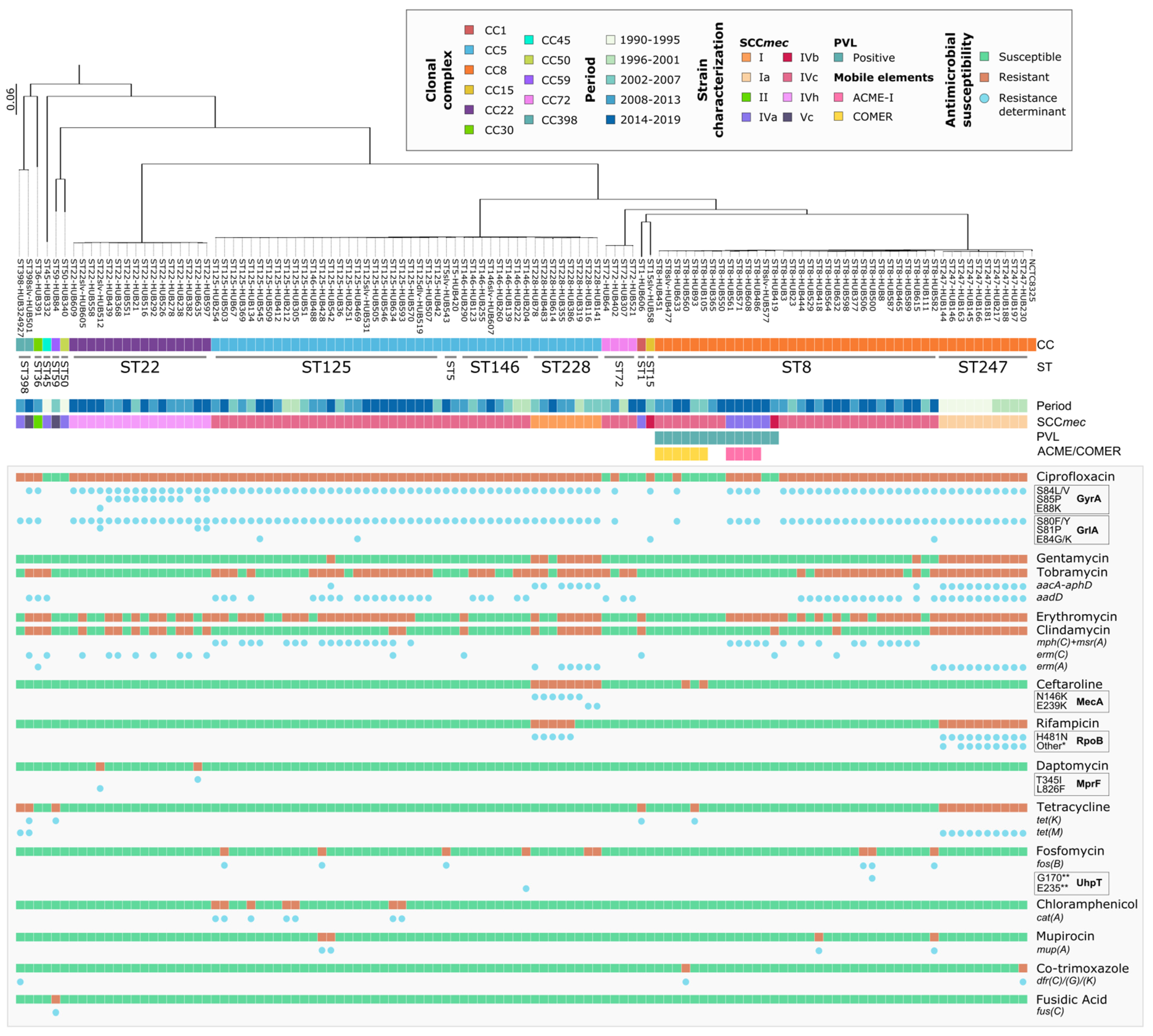
3.2. Molecular Epidemiology of MRSA-BSI
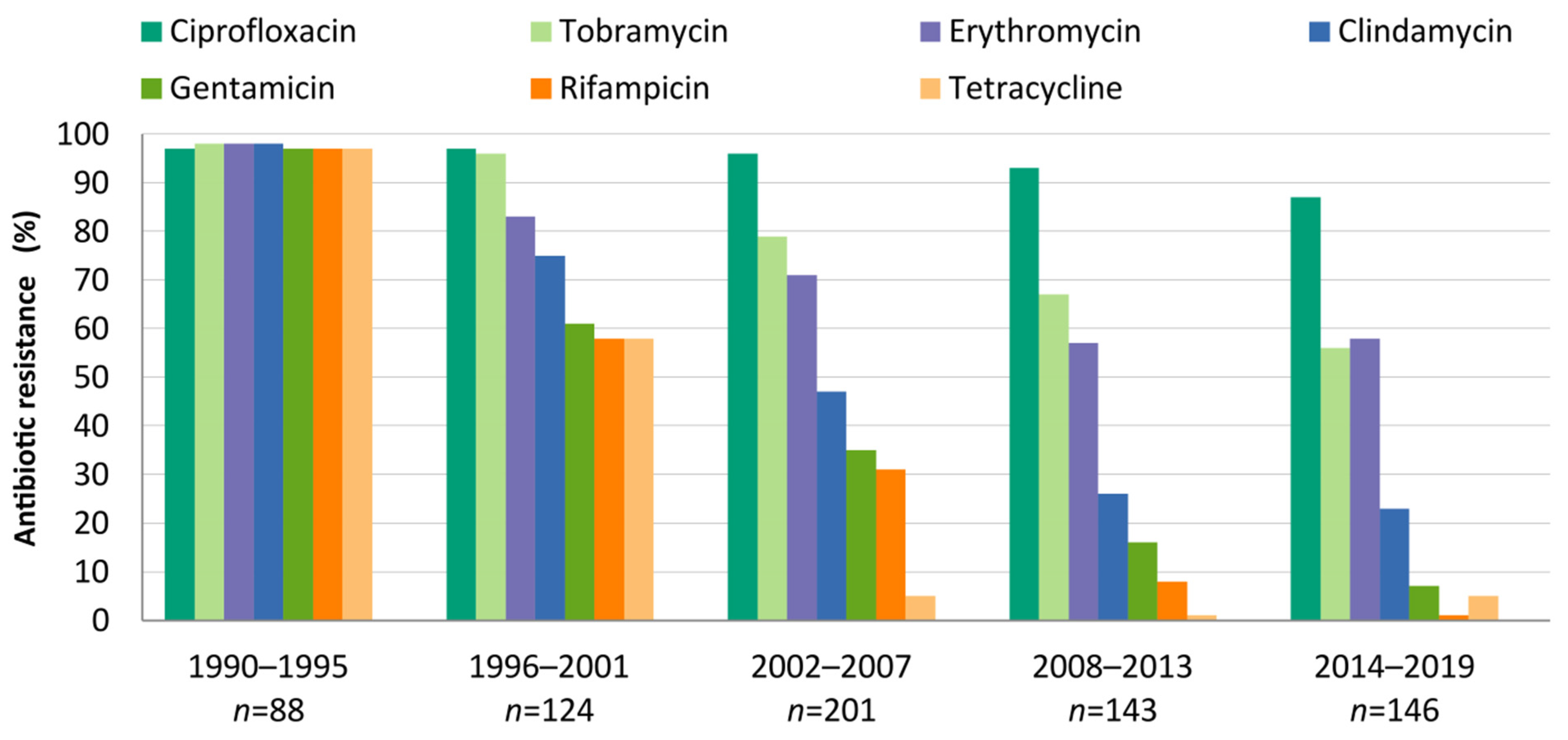
3.3. Evolution and Molecular Basis of Antimicrobial Resistance
3.4. Major Lineages and Clinical Characteristics of Recent MRSA-BSI Episodes (2008–2019)
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lee, A.S.; de Lencastre, H.H.; Garau, J.; Kluytmans, J.; Malhotra-Kumar, S.; Peschel, A.; Harbarth, S. Methicillin-Resistant Staphylococcus aureus. Nat. Rev. Dis. Prim. 2018, 4, 18033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization. Antimicrobial Resistance: Global Report on Surveillance 2014. In WHO 2014 AMR Report; WHO: Geneva, Switzerland, 2014; pp. 1–8. [Google Scholar]
- Munckhof, W.J.; Nimmo, G.R.; Carney, J.; Schooneveldt, J.M.; Huygens, F.; Inman-Bamber, J.; Tong, E.; Morton, A.; Giffard, P. Methicillin-Susceptible, Non-Multiresistant Methicillin-Resistant and Multiresistant Methicillin-Resistant Staphylococcus aureus Infections: A Clinical, Epidemiological and Microbiological Comparative Study. Eur. J. Clin. Microbiol. Infect. Dis. 2008, 27, 355–364. [Google Scholar] [CrossRef] [PubMed]
- Bai, A.D.; Lo, C.K.L.; Komorowski, A.S.; Suresh, M.; Guo, K.; Garg, A.; Tandon, P.; Senecal, J.; del Corpo, O.; Stefanova, I.; et al. Staphylococcus aureus Bacteraemia Mortality: A Systematic Review and Meta-Analysis. Clin. Microbiol. Infect. 2022, 28, 1076–1084. [Google Scholar] [CrossRef]
- Gasch, O.; Ayats, J.; Ángeles Dominguez, M.; Tubau, F.; Liñares, J.; Peña, C.; Grau, I.; Pallarés, R.; Gudiol, F.; Ariza, J.; et al. Epidemiology of Methicillin-Resistant Staphylococcus aureus (MRSA) Bloodstream Infection: Secular Trends over 19 Years at a University Hospital. Medicine 2011, 90, 319–327. [Google Scholar] [CrossRef] [PubMed]
- Jensen, S.O.; Lyon, B.R. Genetics of Antimicrobial Resistance in Staphylococcus Aureus. Future Microbiol. 2009, 4, 565–582. [Google Scholar] [CrossRef] [PubMed]
- Ito, T.; Katayama, Y.; Hiramatsu, K. Cloning and Nucleotide Sequence Determination of the Entire Mec DNA of Pre-Methicillin-Resistant Staphylococcus aureus N315. Antimicrob. Agents Chemother. 1999, 43, 1449–1458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robinson, D.A.; Enright, M.C. Evolutionary Models of the Emergence of Methicillin-Resistant Staphylococcus aureus. Antimicrob. Agents Chemother. 2003, 47, 3926–3934. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nübel, U.; Roumagnac, P.; Feldkamp, M.; Song, J.H.; Ko, K.S.; Huang, Y.C.; Coombs, G.; Ip, M.; Westh, H.; Skov, R.; et al. Frequent Emergence and Limited Geographic Dispersal of Methicillin- Resistant Staphylococcus aureus. Proc. Natl. Acad. Sci. USA 2008, 105, 14130–14135. [Google Scholar] [CrossRef] [Green Version]
- Kashyap, R.; Shah, A.; Dutt, T.; Wieruszewski, P.M.; Ahdal, J.; Jain, R. Treatments and Limitations for Methicillin-Resistant Staphylococcus aureus: A Review of Current Literature. World J. Clin. Infect Dis. 2019, 9, 1–10. [Google Scholar] [CrossRef]
- Watkins, R.R.; Holubar, M.; David, M.Z. Antimicrobial Resistance in Methicillin-Resistant Staphylococcus aureus to Newer Antimicrobial Agents. Antimicrob. Agents Chemother. 2019, 63, e01216-19. [Google Scholar] [CrossRef]
- Asokan, G.V.; Ramadhan, T.; Ahmed, E.; Sanad, H. WHO Global Priority Pathogens List: A Bibliometric Analysis of Medline-PubMed for Knowledge Mobilization to Infection Prevention and Control Practices in Bahrain. Oman Med. J. 2019, 34, 184. [Google Scholar] [CrossRef]
- WHO. Antibiotic Resistance: Multi-Country Public Awareness Survey; WHO: Geneva, Switzerland, 2015; ISBN 978 92 4 150981 7. [Google Scholar]
- Planet, P.J. Life after USA300: The Rise and Fall of a Superbug. J. Infect. Dis. 2017, 215, S71–S77. [Google Scholar] [CrossRef] [PubMed]
- SJ, P.; GK, P. Mechanisms of Methicillin Resistance in Staphylococcus aureus. Ann. Rev. Biochem. 2015, 84, 577–601. [Google Scholar] [CrossRef]
- Vestergaard, M.; Frees, D.; Ingmer, H. Antibiotic Resistance and the MRSA Problem. Microbiol. Spectr. 2019, 7, 2–7. [Google Scholar] [CrossRef] [PubMed]
- Robinson, J.O.; Pearson, J.C.; Christiansen, K.J.; Coombs, G.W.; Murray, R.J. Community-Associated versus Healthcare-Associated Methicillin-Resistant Staphylococcus aureus Bacteraemia: A 10-Year Retrospective Review. Eur. J. Clin. Microbiol. Infect. Dis. 2009, 28, 353–361. [Google Scholar] [CrossRef]
- Nimmo, G.R.; Coombs, G.W. Community-Associated Methicillin-Resistant Staphylococcus aureus (MRSA) in Australia. Int. J. Antimicrob. Agents 2008, 31, 401–410. [Google Scholar] [CrossRef] [PubMed]
- Nimmo, G.R.; Bergh, H.; Nakos, J.; Whiley, D.; Marquess, J.; Huygens, F.; Paterson, D.L. Replacement of Healthcare-Associated MRSA by Community-Associated MRSA in Queensland: Confirmation by Genotyping. J. Infect. 2013, 67, 439–447. [Google Scholar] [CrossRef]
- Turner, N.A.; Sharma-Kuinkel, B.K.; Maskarinec, S.A.; Eichenberger, E.M.; Shah, P.P.; Carugati, M.; Holland, T.L.; Fowler, V.G. Methicillin-Resistant Staphylococcus aureus: An Overview of Basic and Clinical Research. Nat. Rev. Microbiol. 2019, 17, 203–218. [Google Scholar] [CrossRef]
- Lakhundi, S.; Zhang, K. Methicillin-Resistant Staphylococcus aureus: Molecular Characterization, Evolution, and Epidemiology. Clin. Microbiol. Rev. 2018, 31, 1–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sullivan, M.J.; Altman, D.R.; Chacko, K.I.; Ciferri, B.; Webster, E.; Pak, T.R.; Deikus, G.; Lewis-Sandari, M.; Khan, Z.; Beckford, C.; et al. A Complete Genome Screening Program of Clinical Methicillin-Resistant Staphylococcus aureus Isolates Identifies the Origin and Progression of a Neonatal Intensive Care Unit Outbreak. J. Clin. Microbiol. 2019, 57, e01261-19. [Google Scholar] [CrossRef]
- Friedman, N.D.; Kaye, K.S.; Stout, J.E.; McGarry, S.A.; Trivette, S.L.; Briggs, J.P.; Lamm, W.; Clark, C.; MacFarquhar, J.; Walton, A.L.; et al. Health Care-Associated Bloodstream Infections in Adults: A Reason to Change the Accepted Definition of Community-Acquired Infections. Ann. Intern. Med. 2002, 137, 791–797. [Google Scholar] [CrossRef] [PubMed]
- Klevens, R.M.; Morrison, M.A.; Nadle, J.; Petit, S.; Gershman, K.; Ray, S.; Harrison, L.H.; Lynfield, R.; Dumyati, G.; Townes, J.M.; et al. Invasive Methicillin-Resistant Staphylococcus aureus Infections in the United States. JAMA 2007, 298, 1763–1771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clinical and Laboratory Standards Institute (CLSI). Performance Standards for Antimicrobial Susceptibility Testing—30th Edition: A CLSI supplement for global application. M100. J. Serv. Mark. 2020, 40, 1–296. [Google Scholar]
- Magiorakos, A.P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-Resistant, Extensively Drug-Resistant and Pandrug-Resistant Bacteria: An International Expert Proposal for Interim Standard Definitions for Acquired Resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tenover, F.C.; Arbeit, R.D.; Goering, R.V.; Mickelsen, P.a.; Murray, B.E.; Persing, D.H.; Swaminathan, B. Interpreting Chromosomal DNA Restriction Patterns Produced by Pulsed-Field Gel Electrophoresis: Criteria for Bacterial Strain Typing. J. Clin. Microbiol. 1995, 33, 2233–2239. [Google Scholar] [CrossRef] [Green Version]
- Gasch, O.; Camoez, M.; Dominguez, M.A.; Padilla, B.; Pintado, V.; Almirante, B.; Molina, J.; Lopez-Medrano, F.; Ruiz, E.; Martinez, J.A.; et al. Predictive Factors for Mortality in Patients with Methicillin-Resistant Staphylococcus aureus Bloodstream Infection: Impact on Outcome of Host, Microorganism and Therapy. Clin. Microbiol. Infect. 2013, 19, 1049–1057. [Google Scholar] [CrossRef] [Green Version]
- Seemann, T. Prokka: Rapid Prokaryotic Genome Annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef] [Green Version]
- Larsen, M.V.; Cosentino, S.; Rasmussen, S.; Friis, C.; Hasman, H.; Marvig, R.L.; Jelsbak, L.; Sicheritz-Pontén, T.; Ussery, D.W.; Aarestrup, F.M.; et al. Multilocus Sequence Typing of Total-Genome-Sequenced Bacteria. J. Clin. Microbiol. 2012, 50, 1355–1361. [Google Scholar] [CrossRef] [Green Version]
- Bartels, M.D.; Petersen, A.; Worning, P.; Nielsen, J.B.; Larner-Svensson, H.; Johansen, H.K.; Andersen, L.P.; Jarløv, J.O.; Boye, K.; Larsen, A.R.; et al. Comparing Whole-Genome Sequencing with Sanger Sequencing for Spa Typing of Methicillin-Resistant Staphylococcus aureus. J. Clin. Microbiol. 2014, 52, 4305. [Google Scholar] [CrossRef] [Green Version]
- Kaya, H.; Hasman, H.; Larsen, J.; Stegger, M.; Johannesen, T.B.; Allesøe, R.L.; Lemvigh, C.K.; Aarestrup, F.M.; Lund, O.; Larsen, A.R. SCCmec Finder, a Web-Based Tool for Typing of Staphylococcal Cassette Chromosome mec in Staphylococcus aureus Using Whole-Genome Sequence Data. mSphere 2018, 3, e00612-17. [Google Scholar] [CrossRef] [Green Version]
- Croucher, N.J.; Page, A.J.; Connor, T.R.; Delaney, A.J.; Keane, J.A.; Bentley, S.D.; Parkhill, J.; Harris, S.R. Rapid Phylogenetic Analysis of Large Samples of Recombinant Bacterial Whole Genome Sequences Using Gubbins. Nucleic Acids Res. 2015, 43, e15. [Google Scholar] [CrossRef] [PubMed]
- Argimón, S.; Abudahab, K.; Goater, R.J.E.; Fedosejev, A.; Bhai, J.; Glasner, C.; Feil, E.J.; Holden, M.T.G.; Yeats, C.A.; Grundmann, H.; et al. Microreact: Visualizing and Sharing Data for Genomic Epidemiology and Phylogeography. Microb. Genom. 2016, 2, e000093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Page, A.J.; Cummins, C.A.; Hunt, M.; Wong, V.K.; Reuter, S.; Holden, M.T.G.; Fookes, M.; Falush, D.; Keane, J.A.; Parkhill, J. Roary: Rapid Large-Scale Prokaryote Pan Genome Analysis. Bioinformatics 2015, 31, 3691–3693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bortolaia, V.; Kaas, R.S.; Ruppe, E.; Roberts, M.C.; Schwarz, S.; Cattoir, V.; Philippon, A.; Allesoe, R.L.; Rebelo, A.R.; Florensa, A.F.; et al. ResFinder 4.0 for Predictions of Phenotypes from Genotypes. J. Antimicrob. Chemother. 2020, 75, 3491–3500. [Google Scholar] [CrossRef] [PubMed]
- Dominguez, M.A.; de Lencastre, H.; Linares, J.; Tomasz, A. Spread and Maintenance of a Dominant Methicillin-Resistant Staphylococcus aureus (MRSA) Clone during an Outbreak of MRSA Disease in a Spanish Hospital. J. Clin. Microbiol. 1994, 32, 2081–2087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kadlec, K.; Schwarz, S. Identification of a Novel Trimethoprim Resistance Gene, DfrK, in a Methicillin-Resistant Staphylococcus aureus ST398 Strain and Its Physical Linkage to the Tetracycline Resistance Gene Tet(L). Antimicrob. Agents Chemother. 2009, 53, 776–778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- European Centre for Disease Prevention and Control (ECDC). Antimicrobial Resistance in the EU/EEA (EARS-Net)—AER for 2019. In Annual Epidemiological Report 2019; European Centre for Disease Prevention and Control (ECDC): Solna, Sweden, 2020. [Google Scholar]
- Souli, M.; Ruffin, F.; Choi, S.H.; Park, L.P.; Gao, S.; Lent, N.C.; Sharma-Kuinkel, B.K.; Thaden, J.T.; Maskarinec, S.A.; Wanda, L.; et al. Changing Characteristics of Staphylococcus aureus Bacteremia: Results from a 21-Year, Prospective, Longitudinal Study. Clin. Infect. Dis. 2019, 69, 1868–1877. [Google Scholar] [CrossRef]
- Dupper, A.C.; Sullivan, M.J.; Chacko, K.I.; Mishkin, A.; Ciferri, B.; Kumaresh, A.; Berbel Caban, A.; Oussenko, I.; Beckford, C.; Zeitouni, N.E.; et al. Blurred Molecular Epidemiological Lines between the Two Dominant Methicillin-Resistant Staphylococcus aureus Clones. Open Forum. Infect. Dis. 2019, 6, ofz302. [Google Scholar] [CrossRef] [Green Version]
- Saliba, P.; Hornero, A.; Cuervo, G.; Grau, I.; Jimenez, E.; Berbel, D.; Martos, P.; Verge, J.M.; Tebe, C.; Martínez-Sánchez, J.M.; et al. Interventions to Decrease Short-Term Peripheral Venous Catheter-Related Bloodstream Infections: Impact on Incidence and Mortality. J. Hosp. Infect. 2018, 100, e178–e186. [Google Scholar] [CrossRef]
- van Hal, S.J.; Jensen, S.O.; Vaska, V.L.; Espedido, B.A.; Paterson, D.L.; Gosbell, I.B. Predictors of Mortality in Staphylococcus aureus Bacteremia. Clin. Microbiol. Rev. 2012, 25, 362–386. [Google Scholar] [CrossRef] [Green Version]
- Mick, V.; Domínguez, M.A.; Tubau, F.; Liñares, J.; Pujol, M.; Martín, R. Molecular Characterization of Resistance to Rifampicin in an Emerging Hospital-Associated Methicillin-Resistant Staphylococcus aureus Clone ST228, Spain. BMC Microbiol. 2010, 10, 68. [Google Scholar] [CrossRef] [PubMed]
- Monecke, S.; Coombs, G.; Shore, A.C.; Coleman, D.C.; Akpaka, P.; Borg, M.; Chow, H.; Ip, M.; Jatzwauk, L.; Jonas, D.; et al. A Field Guide to Pandemic, Epidemic and Sporadic Clones of Methicillin-Resistant Staphylococcus aureus. PLoS ONE 2011, 6, e17936. [Google Scholar] [CrossRef] [PubMed]
- Holden, M.T.G.; Hsu, L.Y.; Kurt, K.; Weinert, L.A.; Mather, A.E.; Harris, S.R.; Strommenger, B.; Layer, F.; Witte, W.; de Lencastre, H.; et al. A Genomic Portrait of the Emergence, Evolution, and Global Spread of a Methicillin-Resistant Staphylococcus aureus Pandemic. Genome Res. 2013, 23, 653–664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Challagundla, L.; Reyes, J.; Rafiqullah, I.; Sordelli, D.O.; Echaniz-Aviles, G.; Velazquez-Meza, M.E.; Castillo-Ramírez, S.; Fittipaldi, N.; Feldgarden, M.; Chapman, S.B.; et al. Phylogenomic Classification and the Evolution of Clonal Complex 5 Methicillin-Resistant Staphylococcus aureus in the Western Hemisphere. Front. Microbiol. 2018, 9, 1901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuzi, M. Dissimilar Fitness Associated with Resistance to Fluoroquinolones Influences Clonal Dynamics of Various Multiresistant Bacteria. Front. Microbiol. 2016, 7, 1017. [Google Scholar] [CrossRef] [Green Version]
- Haaber, J.; Penadés, J.R.; Ingmer, H. Transfer of Antibiotic Resistance in Staphylococcus aureus. Trends. Microbiol. 2017, 25, 893–905. [Google Scholar] [CrossRef]
- Mlynarczyk-Bonikowska, B.; Kowalewski, C.; Krolak-Ulinska, A.; Marusza, W. Molecular Mechanisms of Drug Resistance in Staphylococcus aureus. Int. J. Mol. Sci 2022, 23, 8088. [Google Scholar] [CrossRef]
- Adriaenssens, N.; Bruyndonckx, R.; Versporten, A.; Hens, N.; Monnet, D.L.; Molenberghs, G.; Goossens, H.; Weist, K.; Coenen, S. Consumption of Macrolides, Lincosamides and Streptogramins in the Community, European Union/European Economic Area, 1997–2017. J. Antimicrob. Chemother. 2021, 76, 30–36. [Google Scholar] [CrossRef]
- Doan, T.; Worden, L.; Hinterwirth, A.; Arzika, A.M.; Maliki, R.; Abdou, A.; Zhong, L.; Chen, C.; Cook, C.; Lebas, E.; et al. Macrolide and Nonmacrolide Resistance with Mass Azithromycin Distribution. N. Engl. J. Med. 2020, 383, 1941–1950. [Google Scholar] [CrossRef]
- Sabat, A.J.; Tinelli, M.; Grundmann, H.; Akkerboom, V.; Monaco, M.; del Grosso, M.; Errico, G.; Pantosti, A.; Friedrich, A.W. Daptomycin Resistant Staphylococcus aureus Clinical Strain with Novel Non-Synonymous Mutations in the MprF and VraS Genes: A New Insight into Daptomycin Resistance. Front. Microbiol. 2018, 9, 2705. [Google Scholar] [CrossRef] [Green Version]
- Weber, R.E.; Fuchs, S.; Layer, F.; Sommer, A.; Bender, J.K.; Thürmer, A.; Werner, G.; Strommenger, B. Genome-Wide Association Studies for the Detection of Genetic Variants Associated with Daptomycin and Ceftaroline Resistance in Staphylococcus aureus. Front. Microbiol. 2021, 12, 639660. [Google Scholar] [CrossRef] [PubMed]
- Deresinski, S. Methicillin-Resistant Staphylococcus aureus: An Evolutionary, Epidemiologic, and Therapeutic Odyssey. Clin. Infect. Dis. 2005, 40, 562–573. [Google Scholar] [CrossRef] [PubMed]
- Armand-Lefevre, L.; Buke, C.; Ruppe, E.; Barbier, F.; Lolom, I.; Andremont, A.; Ruimy, R.; Lucet, J.C. Secular Trends and Dynamics of Hospital Associated Methicillin-Resistant Staphylococcus aureus. Clin. Microbiol. Infect. 2010, 16, 1435–1441. [Google Scholar] [CrossRef] [PubMed]
- Carrera-Salinas, A.; González-Díaz, A.; Vázquez-Sánchez, D.A.; Camoez, M.; Niubó, J.; Càmara, J.; Ardanuy, C.; Martí, S.; Domínguez, M.Á.; Garcia, M.; et al. Staphylococcus aureus Surface Protein G (SasG) Allelic Variants: Correlation between Biofilm Formation and Their Prevalence in Methicillin-Resistant S. aureus (MRSA) Clones. Res. Microbiol. 2022, 173, 103921. [Google Scholar] [CrossRef] [PubMed]
- Monecke, S.; Ruppelt-Lorz, A.; Müller, E.; Reissig, A.; Thürmer, A.; Shore, A.C.; Coleman, D.C.; Ehricht, R.; Jatzwauk, L. Dissemination of High-Level Mupirocin-Resistant CC22-MRSA-IV in Saxony. GMS Hyg. Infect Control 2017, 12, Doc19. [Google Scholar] [CrossRef]
- Toleman, M.S.; Reuter, S.; Jamrozy, D.; Wilson, H.J.; Blane, B.; Harrison, E.M.; Coll, F.; Hope, R.J.; Kearns, A.; Parkhill, J.; et al. Prospective Genomic Surveillance of Methicillin Resistant Staphylococcus aureus (MRSA) Associated with Bloodstream Infection, England, 1 October 2012 to 30 September 2013. Eurosurveillance 2019, 24, 1800215. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Period | ||||||
|---|---|---|---|---|---|---|
| 1990–1995 | 1996–2001 | 2002–2007 | 2008–2013 | 2014–2019 | ||
| Number of episodes | 137 | 142 | 207 | 149 | 149 | |
| Sex, female | 34 (25) | 38 (27) | 76 (37) | 62 (42) | 68 (46) * | |
| Age, mean ± SD (range) | 60.2 ± 18.9 (22–94) | 68.3 ± 16.3 (23–99) | 70.2 ± 14.9 (24–98) | 70.1 ± 14.1 (27–94) | 71.2 ± 14.5 (19–85) * | |
| Acquisition | HO-HCA | 124 (91) | 113 (80) | 98 (47) | 70 (47) | 39 (26) * |
| CO-HCA | 13 (9) | 29 (20) | 107 (52) | 71 (48) | 89 (60) * | |
| CA | 0 (0) | 0 (0) | 2 (1) | 8 (5) | 21 (14) * | |
| Source of infection | ||||||
| Catheter | 78 (57) | 71 (50) | 62 (30) | 35 (24) | 30 (20) * | |
| Skin and soft tissue | 8 (6) | 12 (8) | 34 (16) | 20 (13) | 29 (20) * | |
| Respiratory tract | 1 (1) | 14 (10) | 17 (8) | 17 (11) | 24 (16) * | |
| Urinary | 2 (1) | 5 (4) | 22 (11) | 21 (14) | 12 (8) * | |
| Osteoarticular | 4 (3) | 5 (4) | 12 (6) | 11 (7) | 13 (9) * | |
| Endocarditis | 5 (3) | 3 (2) | 4 (2) | 4 (3) | 11 (7) | |
| Other source ‡ | 16 (12) | 22 (15) | 35 (17) | 13 (9) | 10 (7) | |
| Unknown source | 23 (17) | 10 (7) | 21 (10) | 28 (19) | 20 (13) | |
| CCI | >2 | 13 (10) | 20 (25) | 92 (49) | 117 (82) | 140 (94) * |
| ≥5 | 7 (6) | 3 (4) | 33 (18) | 94 (66) | 112 (75) * | |
| Not available | 13 | 61 | 21 | 7 | 0 | |
| 30-day mortality | 39 (28) | 41 (29) | 57 (28) | 57 (38) | 54 (36) | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vázquez-Sánchez, D.A.; Grillo, S.; Carrera-Salinas, A.; González-Díaz, A.; Cuervo, G.; Grau, I.; Camoez, M.; Martí, S.; Berbel, D.; Tubau, F.; et al. Molecular Epidemiology, Antimicrobial Susceptibility, and Clinical Features of Methicillin-Resistant Staphylococcus aureus Bloodstream Infections over 30 Years in Barcelona, Spain (1990–2019). Microorganisms 2022, 10, 2401. https://doi.org/10.3390/microorganisms10122401
Vázquez-Sánchez DA, Grillo S, Carrera-Salinas A, González-Díaz A, Cuervo G, Grau I, Camoez M, Martí S, Berbel D, Tubau F, et al. Molecular Epidemiology, Antimicrobial Susceptibility, and Clinical Features of Methicillin-Resistant Staphylococcus aureus Bloodstream Infections over 30 Years in Barcelona, Spain (1990–2019). Microorganisms. 2022; 10(12):2401. https://doi.org/10.3390/microorganisms10122401
Chicago/Turabian StyleVázquez-Sánchez, Daniel Antonio, Sara Grillo, Anna Carrera-Salinas, Aida González-Díaz, Guillermo Cuervo, Inmaculada Grau, Mariana Camoez, Sara Martí, Dàmaris Berbel, Fe Tubau, and et al. 2022. "Molecular Epidemiology, Antimicrobial Susceptibility, and Clinical Features of Methicillin-Resistant Staphylococcus aureus Bloodstream Infections over 30 Years in Barcelona, Spain (1990–2019)" Microorganisms 10, no. 12: 2401. https://doi.org/10.3390/microorganisms10122401