

Innovative Bioactive Products with Medicinal Value from Microalgae and Their Overall Process Optimization through the Implementation of Life Cycle Analysis—An Overview



Abstract
:1. Introduction
2. Bioactive Compounds of Marine Origin
2.1. Carotenoids
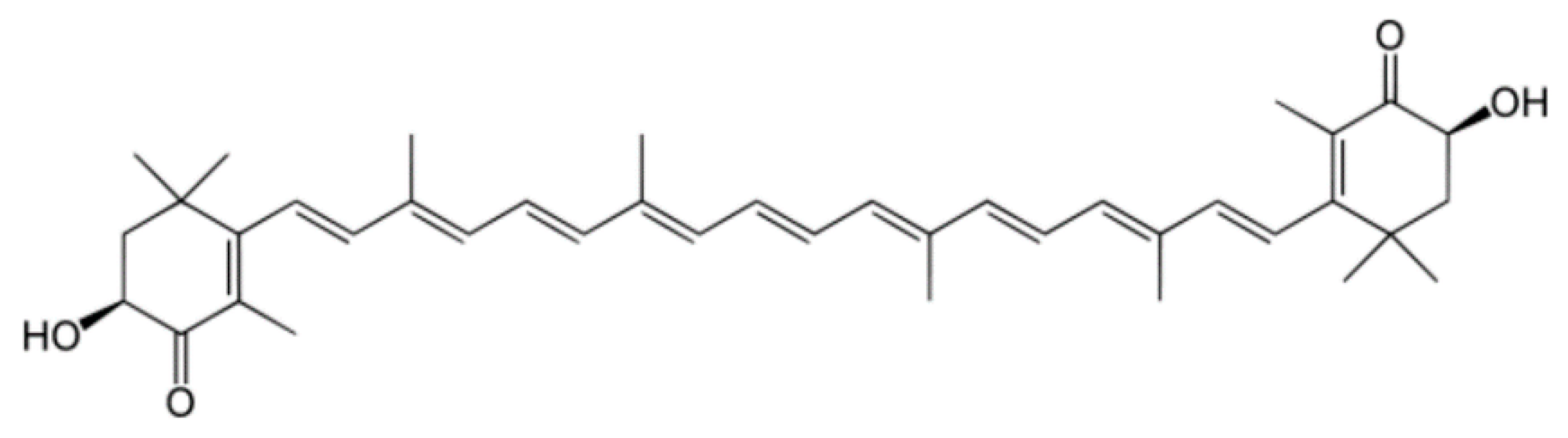
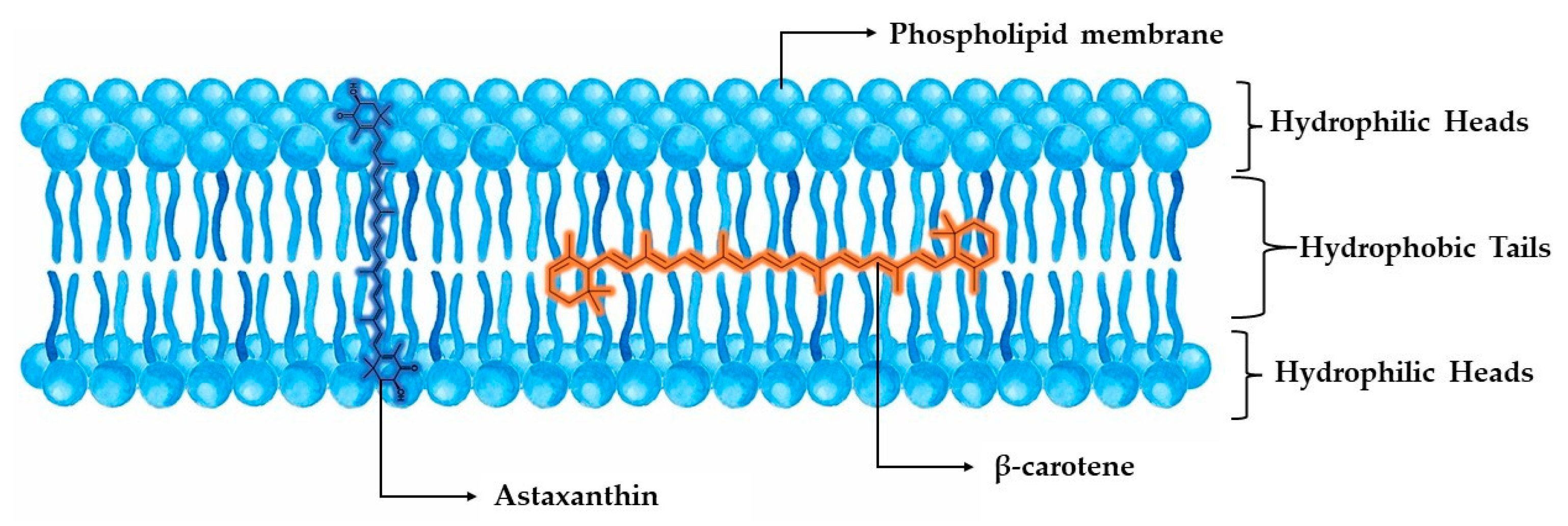
2.1.1. Astaxanthin
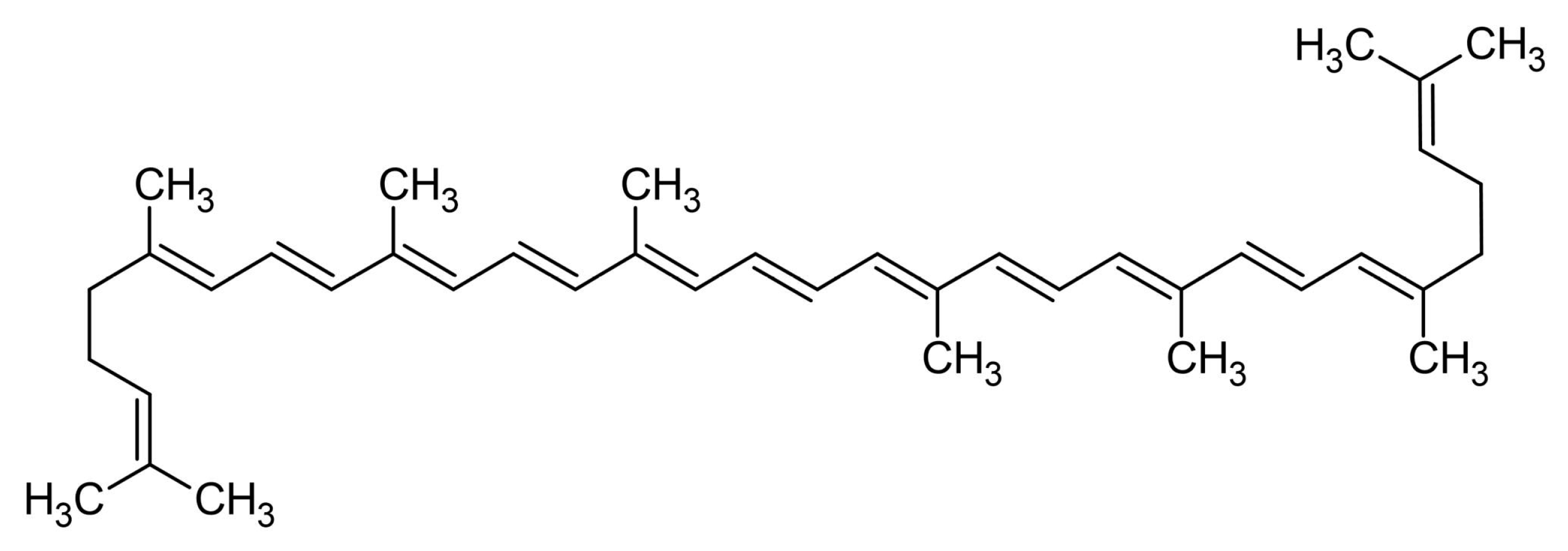
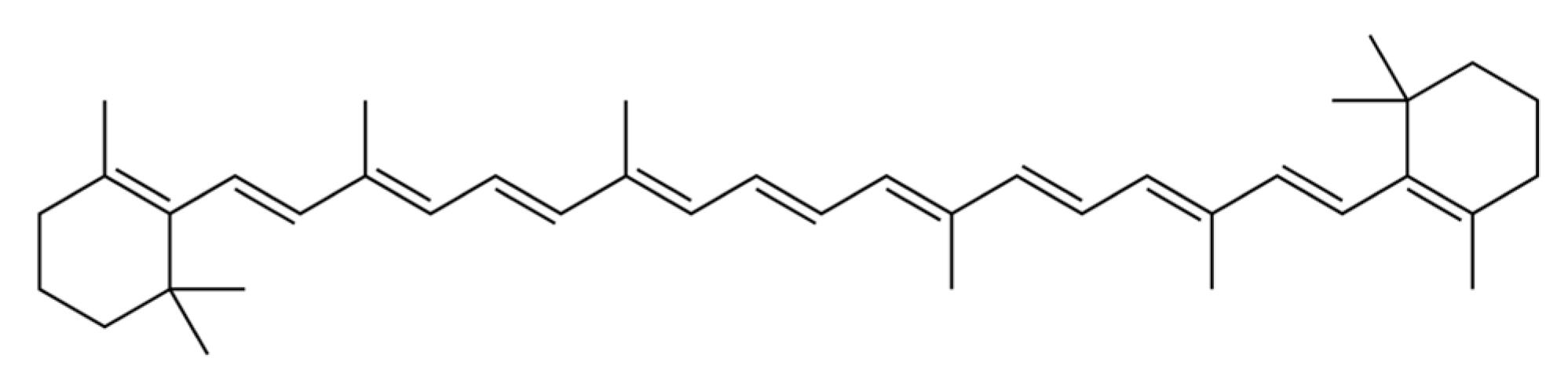
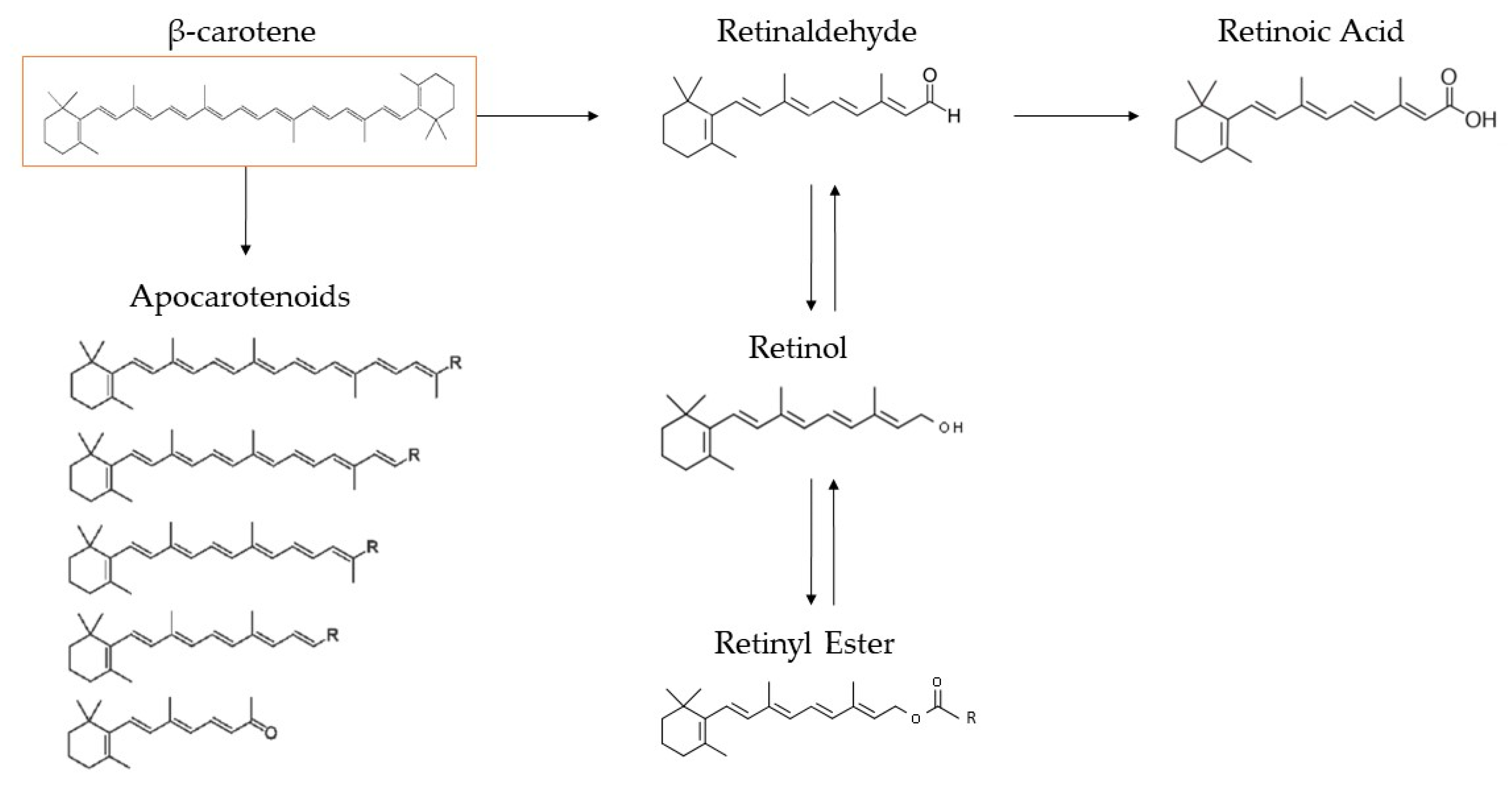
2.1.2. β-Carotene
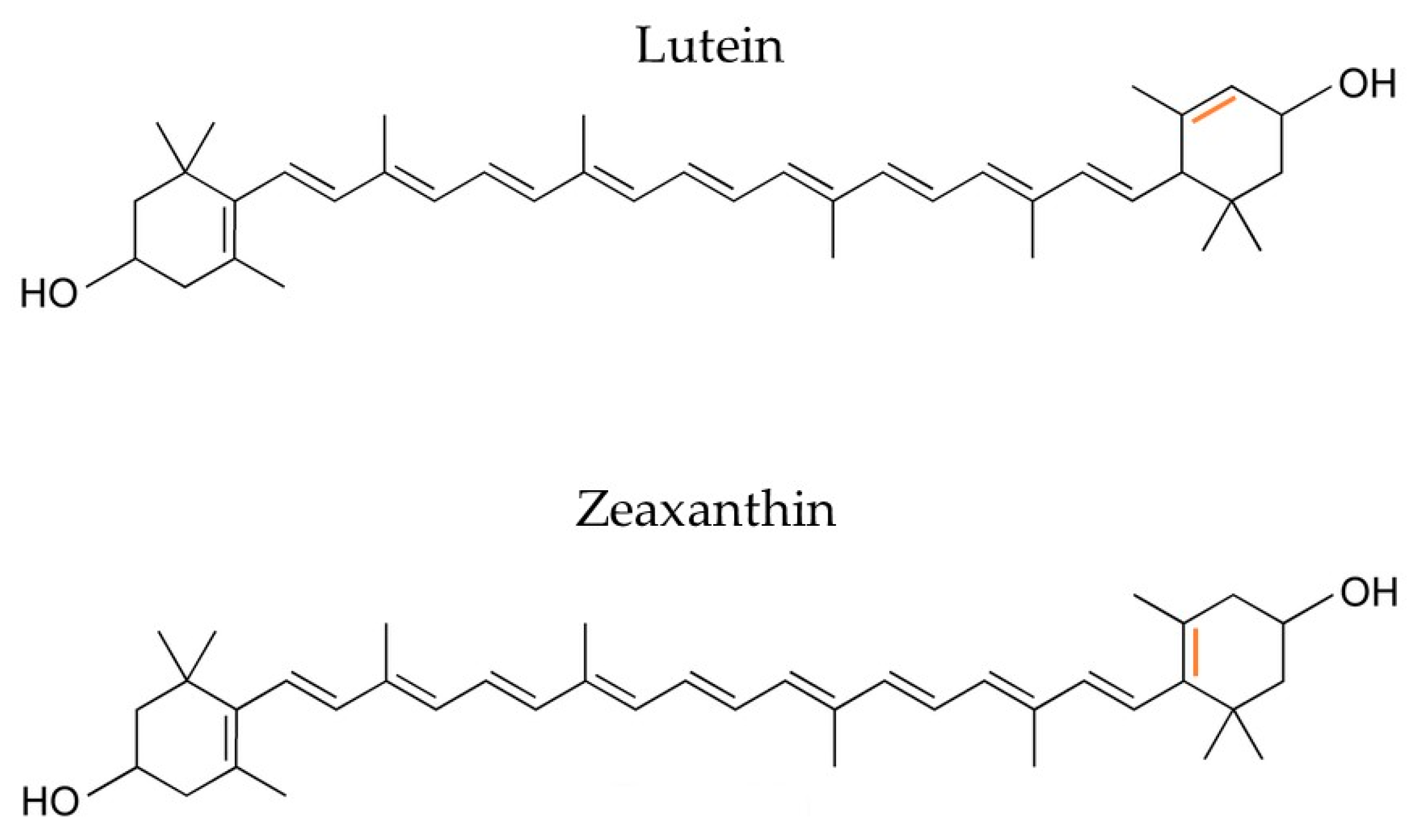
2.1.3. Lutein and Zeaxanthin
2.2. Proteins and Bioactive Hydrolysates
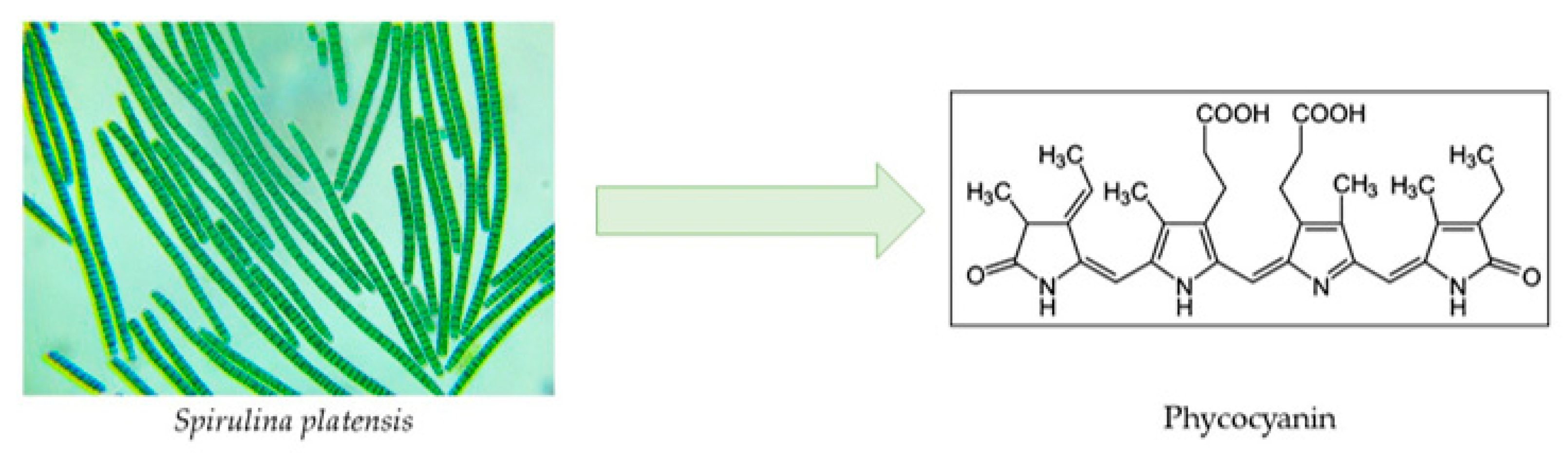
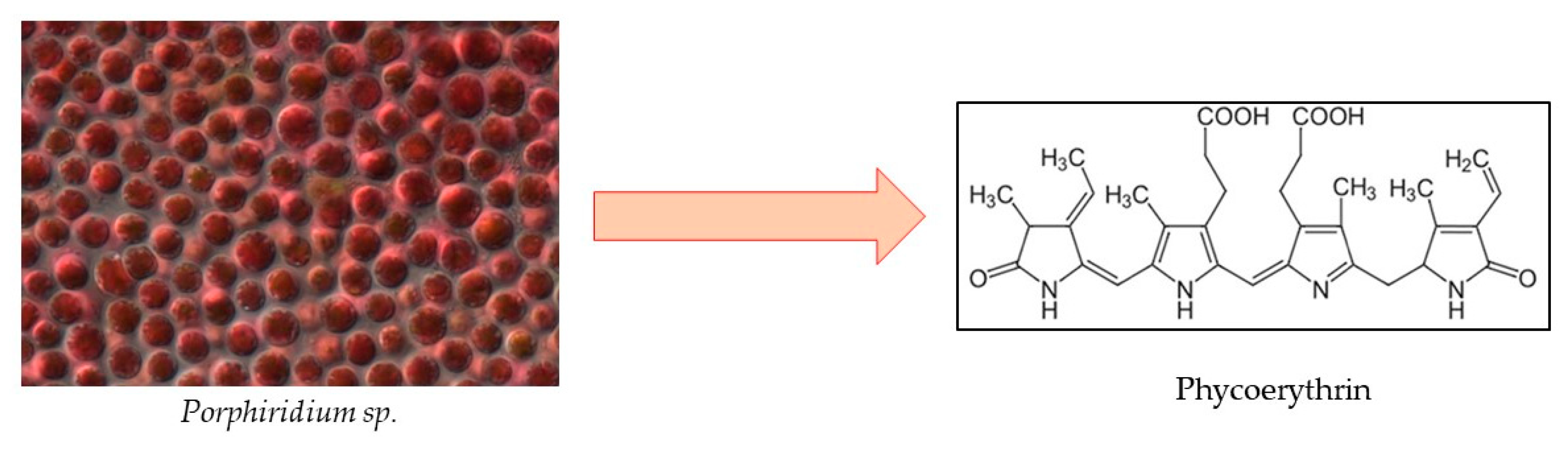
2.2.1. Phycobiliproteins
Phycocyanin
Phycoerythrin
2.2.2. Bioactive Hydrolysates
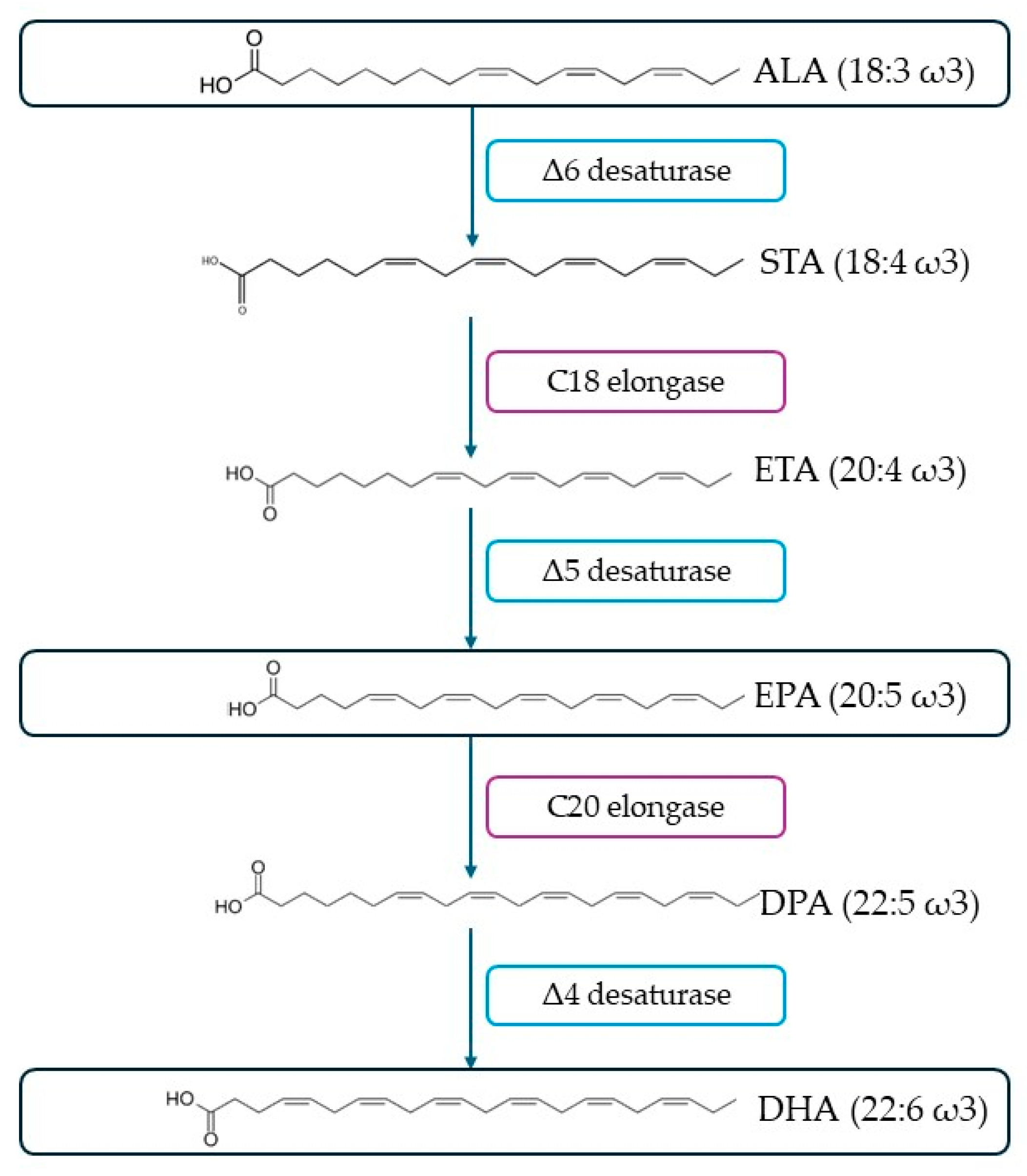
2.3. Polyunsaturated Fatty Acids
3. Extraction of Bioactive Compounds from Microalgae
3.1. Ultrasound-Assisted Extraction
3.2. Microwave-Assisted Extraction
3.3. Pressurized Liquid Extraction
4. Encapsulation of Bioactive Compounds
4.1. Electrohydrodynamic Process (Electrospinning)
- It is a simpler and more flexible method with a higher containment efficiency.
- It produces very fine fibers of a few nanometers in size with a large surface area.
- There is the potential to produce nanofibers on a large scale.
- Compared to the extrusion method, the solution is not heated during the creation of the fibers, as a result of which any effects of degradation on the active ingredients from heat are limited.
- A stainless steel needle with a blunt tip or a capillary tube.
- A pump, whose role is to move the feeding syringe, which contains the solution to be fiberized.
- A power supply, a source of a high-voltage supply.
- A grounded collector for the fibers produced.
- The parameters related to the properties of the solution, such as concentration, viscosity, molecular weight, surface tension, nature of the solvent, and conductivity.
- The parameters related to the device, e.g., flow rate, applied electric field strength, solution volumetric flow rate, needle–collector distance, temperature, humidity, and air flow inside the chamber, whose shape regulates and affects the fibers’ properties, such as diameter, uniformity, porosity, or even their various defects.
- The composition of the solution (by changing the type of polymer, its concentration, or the type of solvent).
- The properties of the solution (by changing pH, temperature, and ionic strength).
- The device parameters (by varying the applied voltage, the distance between the source and collection surface, and the relative humidity of the chamber).
4.2. Spray–Freeze Drying
- Spray–freezing into a vapor, where the feed solution is sprayed into a chamber with cold gas causing the particles to freeze.
- Spray–freezing into a liquid, where the nozzle of the atomizer is placed into a cryogenic liquid and the spray occurs directly in it, producing frozen microparticles.
- Spray–freezing into a vapor over a liquid, where the atomization occurs in contact with a cold gas. The particles begin to freeze during their dispersion into the vapor and fall into the cryogenic liquid, where they become completely frozen.
5. Life Cycle Analysis (LCA) as a Sustainability Tool for Process Optimization
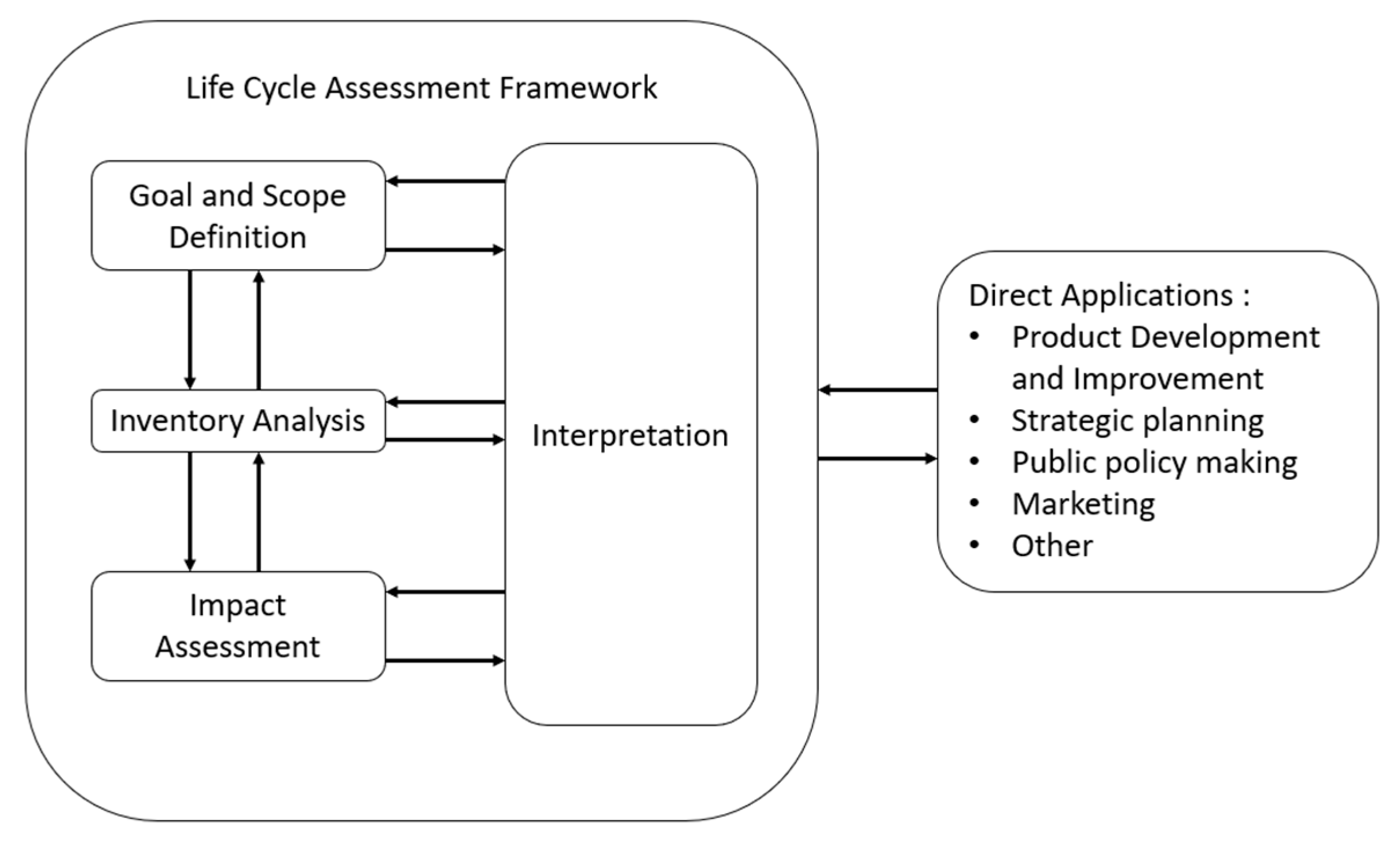
5.1. The ISO Framework
- Goal and scope definition.
- Inventory analysis.
- Life cycle impact assessment.
- Life cycle interpretation.
5.1.1. Goal and Scope Definition
5.1.2. Inventory Analysis
5.1.3. Impact Assessment
- Determining impact categories that are indicative of the evaluation criteria established throughout the scope definition process. A relevant indicator and an environmental model for each impact category are used to estimate the effect of primary streams on the indicator.
- The elementary flows from the inventory are grouped into impact categories based on how they could potentially influence the selected indicator.
- The characterization of the impact category utilizing environmental frameworks to assess how each allocated primary flow affects the category indicator. A single measure for the impact category is used to express the characterized impact scores that are obtained. This consolidates all of the contributions into a single score that reflects the overall influence that the product system has on that category.
- By presenting each specified score for each effect category with respect to a single reference impact for each impact category, the normalization serves to provide insight into the relative severity of each impact. The product system’s standardized impact profile, which expresses each category indicator score using the same metric, is the final product of the normalization process.
- By grouping and possibly ranking the impact categories based on their perceived intensity, or by weighting them using weighting factors that provide a quantitative expression of each impact category’s severity concerning the other impact categories, grouping, and weighting, stimulates comparisons across the impact categories.
5.1.4. Interpretation
5.2. Applications for Bioactive Compounds Isolation of Marine Origin
6. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Totelin, L. When Foods Become Remedies in Ancient Greece: The Curious Case of Garlic and Other Substances. J. Ethnopharmacol. 2015, 167, 30–37. [Google Scholar] [CrossRef] [PubMed]
- Tapsell, L.C.; Hemphill, I.; Cobiac, L.; Sullivan, D.R.; Fenech, M.; Patch, C.S.; Roodenrys, S.; Keogh, J.B.; Clifton, P.M.; Williams, P.G.; et al. Health Benefits of Herbs and Spices: The Past, the Present, the Future. Med. J. Aust. 2006, 185, S1–S24. [Google Scholar] [CrossRef] [PubMed]
- Rao, A.; Rao, L. Carotenoids and Human Health. Pharmacol. Res. 2007, 55, 207–216. [Google Scholar] [CrossRef] [PubMed]
- Ames, B.N.; Shigenaga, M.K.; Hagen, T.M. Oxidants, Antioxidants, and the Degenerative Diseases of Aging. Proc. Natl. Acad. Sci. USA 1993, 90, 7915–7922. [Google Scholar] [CrossRef] [PubMed]
- Shahidi, F.; Ambigaipalan, P. Omega-3 Polyunsaturated Fatty Acids and Their Health Benefits. Annu. Rev. Food Sci. Technol. 2018, 9, 345–381. [Google Scholar] [CrossRef] [PubMed]
- Friedman, M. Nutritional Value of Proteins from Different Food Sources. A Review. J. Agric. Food Chem. 1996, 44, 6–29. [Google Scholar] [CrossRef]
- DataM Intelligence Bioactive Market Trends in Health & Nutrition Research. Available online: https://www.datamintelligence.com/research-report/bioactive-ingredients-market (accessed on 15 January 2024).
- Uma, V.S.; Usmani, Z.; Sharma, M.; Diwan, D.; Sharma, M.; Guo, M.; Tuohy, M.G.; Makatsoris, C.; Zhao, X.; Thakur, V.K.; et al. Valorisation of Algal Biomass to Value-Added Metabolites: Emerging Trends and Opportunities. Phytochem. Rev. 2023, 22, 1015–1040. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.; Sarkar, T.; Pati, S.; Kari, Z.A.; Edinur, H.A.; Chakraborty, R. Novel Bioactive Compounds from Marine Sources as a Tool for Functional Food Development. Front. Mar. Sci. 2022, 9, 832957. [Google Scholar] [CrossRef]
- Martínez-Francés, E.; Escudero-Oñate, C. Cyanobacteria and Microalgae in the Production of Valuable Bioactive Compounds. In Microalgal Biotechnology; InTech: Orlando, FL, USA, 2018. [Google Scholar]
- Corrêa, P.S.; Morais Júnior, W.G.; Martins, A.A.; Caetano, N.S.; Mata, T.M. Microalgae Biomolecules: Extraction, Separation and Purification Methods. Processes 2020, 9, 10. [Google Scholar] [CrossRef]
- Quitério, E.; Grosso, C.; Ferraz, R.; Delerue-Matos, C.; Soares, C. A Critical Comparison of the Advanced Extraction Techniques Applied to Obtain Health-Promoting Compounds from Seaweeds. Mar. Drugs 2022, 20, 677. [Google Scholar] [CrossRef]
- de Santos, P.D.F.; Rubio, F.T.V.; da Silva, M.P.; Pinho, L.S.; Favaro-Trindade, C.S. Microencapsulation of Carotenoid-Rich Materials: A Review. Food Res. Int. 2021, 147, 110571. [Google Scholar] [CrossRef] [PubMed]
- Khoshnoudi-Nia, S.; Forghani, Z.; Jafari, S.M. A Systematic Review and Meta-Analysis of Fish Oil Encapsulation within Different Micro/Nanocarriers. Crit. Rev. Food Sci. Nutr. 2022, 62, 2061–2082. [Google Scholar] [CrossRef] [PubMed]
- Čuček, L.; Klemeš, J.J.; Kravanja, Z. Overview of Environmental Footprints. In Assessing and Measuring Environmental Impact and Sustainability; Elsevier: Amsterdam, The Netherlands, 2015; pp. 131–193. [Google Scholar]
- Hellweg, S.; Milà i Canals, L. Emerging Approaches, Challenges and Opportunities in Life Cycle Assessment. Science 2014, 344, 1109–1113. [Google Scholar] [CrossRef] [PubMed]
- Haoujar, I.; Cacciola, F.; Abrini, J.; Mangraviti, D.; Giuffrida, D.; Oulad El Majdoub, Y.; Kounnoun, A.; Miceli, N.; Fernanda Taviano, M.; Mondello, L.; et al. The Contribution of Carotenoids, Phenolic Compounds, and Flavonoids to the Antioxidative Properties of Marine Microalgae Isolated from Mediterranean Morocco. Molecules 2019, 24, 4037. [Google Scholar] [CrossRef] [PubMed]
- Jacob-Lopes, E.; Maroneze, M.M.; Deprá, M.C.; Sartori, R.B.; Dias, R.R.; Zepka, L.Q. Bioactive Food Compounds from Microalgae: An Innovative Framework on Industrial Biorefineries. Curr. Opin. Food Sci. 2019, 25, 1–7. [Google Scholar] [CrossRef]
- Balasubramaniam, V.; Gunasegavan, R.D.-N.; Mustar, S.; Lee, J.C.; Mohd Noh, M.F. Isolation of Industrial Important Bioactive Compounds from Microalgae. Molecules 2021, 26, 943. [Google Scholar] [CrossRef] [PubMed]
- Borowitzka, M.A. High-Value Products from Microalgae-Their Development and Commercialisation. J. Appl. Phycol. 2013, 25, 743–756. [Google Scholar] [CrossRef]
- Spolaore, P.; Joannis-Cassan, C.; Duran, E.; Isambert, A. Commercial Applications of Microalgae. J. Biosci. Bioeng. 2006, 101, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Britton, G. Getting to Know Carotenoids. In Methods in Enzymology; Academic Press Inc.: Cambridge, MA, USA, 2022; Volume 670, pp. 1–56. ISBN 9780323999755. [Google Scholar]
- Ellison, S.L. Carotenoids: Physiology. In Encyclopedia of Food and Health; Elsevier Inc.: Amsterdam, The Netherlands, 2015; pp. 670–675. ISBN 9780123849533. [Google Scholar]
- Ladygin, V.G. Biosynthesis of Carotenoids in the Chloroplasts of Algae and Higher Plants. Russ. J. Plant Physiol. 2000, 47, 796–814. [Google Scholar] [CrossRef]
- Bohn, T. Bioavailability of Non-Provitamin A Carotenoids. Curr. Nutr. Food Sci. 2008, 4, 240–258. [Google Scholar] [CrossRef]
- Stahl, W.; Sies, H. Antioxidant Activity of Carotenoids. Mol. Asp. Med. 2003, 24, 345–351. [Google Scholar] [CrossRef] [PubMed]
- Miyashita, K. Marine Antioxidants. In Antioxidants and Functional Components in Aquatic Foods; Wiley: Hoboken, NJ, USA, 2014; pp. 219–235. [Google Scholar]
- Liu, Z.; van den Berg, C.; Weusthuis, R.A.; Dragone, G.; Mussatto, S.I. Strategies for an Improved Extraction and Separation of Lipids and Carotenoids from Oleaginous Yeast. Sep. Purif. Technol. 2021, 257, 117946. [Google Scholar] [CrossRef]
- Jain, A.; Sirisha, V.L. Algal Carotenoids. In Encyclopedia of Marine Biotechnology; Wiley: Hoboken, NJ, USA, 2020; pp. 33–64. [Google Scholar]
- Mehariya, S.; Goswami, R.K.; Karthikeysan, O.P.; Verma, P. Microalgae for High-Value Products: A Way towards Green Nutraceutical and Pharmaceutical Compounds. Chemosphere 2021, 280, 130553. [Google Scholar] [CrossRef] [PubMed]
- Ranga Rao, A.; Raghunath Reddy, R.L.; Baskaran, V.; Sarada, R.; Ravishankar, G.A. Characterization of Microalgal Carotenoids by Mass Spectrometry and Their Bioavailability and Antioxidant Properties Elucidated in Rat Model. J. Agric. Food Chem. 2010, 58, 8553–8559. [Google Scholar] [CrossRef] [PubMed]
- Del Campo, J.A.; Rodriguez, H.; Moreno, J.; Vargas, M.A.; Rivas, J.; Guerrero, M.G. Accumulation of Astaxanthin and Lutein in Chlorella zofingiensis (Chlorophyta). Appl. Microbiol. Biotechnol. 2004, 64, 848–854. [Google Scholar] [CrossRef] [PubMed]
- Ranga Rao, A.; Baskaran, V.; Sarada, R.; Ravishankar, G.A. In Vivo Bioavailability and Antioxidant Activity of Carotenoids from Microalgal Biomass—A Repeated Dose Study. Food Res. Int. 2013, 54, 711–717. [Google Scholar] [CrossRef]
- Inbaraj, B.S.; Chien, J.T.; Chen, B.H. Improved High Performance Liquid Chromatographic Method for Determination of Carotenoids in the Microalga chlorella Pyrenoidosa. J. Chromatogr. A 2006, 1102, 193–199. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Lin, Y.; He, M.; Gong, Y.; Huang, J. Induced High-Yield Production of Zeaxanthin, Lutein, and β-Carotene by a Mutant of Chlorella zofingiensis. J. Agric. Food Chem. 2018, 66, 891–897. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.-C.; Lin, J.-T.; Lu, F.-J.; Chou, F.-P.; Yang, D.-J. Determination of Carotenoids in Dunaliella salina Cultivated in Taiwan and Antioxidant Capacity of the Algal Carotenoid Extract. Food Chem. 2008, 109, 439–446. [Google Scholar] [CrossRef] [PubMed]
- Murthy, K.N.C.; Rajesha, J.; Swamy, M.M.; Ravishankar, G.A. Comparative Evaluation of Hepatoprotective Activity of Carotenoids of Microalgae. J. Med. Food 2005, 8, 523–528. [Google Scholar] [CrossRef] [PubMed]
- Villaró, S.; Ciardi, M.; Morillas-España, A.; Sánchez-Zurano, A.; Acién-Fernández, G.; Lafarga, T. Microalgae Derived Astaxanthin: Research and Consumer Trends and Industrial Use as Food. Foods 2021, 10, 2303. [Google Scholar] [CrossRef] [PubMed]
- Debnath, T.; Bandyopadhyay, T.K.; Vanitha, K.; Bobby, M.N.; Nath Tiwari, O.; Bhunia, B.; Muthuraj, M. Astaxanthin from Microalgae: A Review on Structure, Biosynthesis, Production Strategies and Application. Food Res. Int. 2024, 176, 113841. [Google Scholar] [CrossRef] [PubMed]
- Renstrøm, B.; Liaaen-Jensen, S. Fatty Acid Composition of Some Esterified Carotenols. Comp. Biochem. Physiol. Part B Comp. Biochem. 1981, 69, 625–627. [Google Scholar] [CrossRef]
- Zhekisheva, M.; Zarka, A.; Khozin-Goldberg, I.; Cohen, Z.; Boussiba, S. Inhibition of Astaxanthin Synthesis Under High Irradiance Does Not Abolish Triacylglycerol Accumulation In The Green Alga Haematococcus pluvialis (Chlorophyceae). J. Phycol. 2005, 41, 819–826. [Google Scholar] [CrossRef]
- Panis, G.; Carreon, J.R. Commercial Astaxanthin Production Derived by Green Alga Haematococcus pluvialis: A Microalgae Process Model and a Techno-Economic Assessment All through Production Line. Algal Res. 2016, 18, 175–190. [Google Scholar] [CrossRef]
- Ashokkumar, V.; Flora, G.; Sevanan, M.; Sripriya, R.; Chen, W.H.; Park, J.-H.; Rajesh, B.J.; Kumar, G. Technological Advances in the Production of Carotenoids and Their Applications– A Critical Review. Bioresour. Technol. 2023, 367, 128215. [Google Scholar] [CrossRef] [PubMed]
- Jannel, S.; Caro, Y.; Bermudes, M.; Petit, T. Novel Insights into the Biotechnological Production of Haematococcus pluvialis-Derived Astaxanthin: Advances and Key Challenges to Allow Its Industrial Use as Novel Food Ingredient. J. Mar. Sci. Eng. 2020, 8, 789. [Google Scholar] [CrossRef]
- Aly, H.K.; El-Baz, F.; Ali, S.I.; Salama, A. Safety of Astaxanthin-Rich Fraction of Haematococcus pluvialis Microalgae. Egypt. J. Chem. 2021, 65, 479–489. [Google Scholar] [CrossRef]
- Niu, T.; Zhou, J.; Wang, F.; Xuan, R.; Chen, J.; Wu, W.; Chen, H. Safety Assessment of Astaxanthin from Haematococcus pluvialis: Acute Toxicity, Genotoxicity, Distribution and Repeat-Dose Toxicity Studies in Gestation Mice. Regul. Toxicol. Pharmacol. 2020, 115, 104695. [Google Scholar] [CrossRef]
- Naguib, Y.M.A. Antioxidant Activities of Astaxanthin and Related Carotenoids. J. Agric. Food Chem. 2000, 48, 1150–1154. [Google Scholar] [CrossRef]
- Goto, S.; Kogure, K.; Abe, K.; Kimata, Y.; Kitahama, K.; Yamashita, E.; Terada, H. Efficient Radical Trapping at the Surface and inside the Phospholipid Membrane Is Responsible for Highly Potent Antiperoxidative Activity of the Carotenoid Astaxanthin. Biochim. Biophys. Acta (BBA) Biomembr. 2001, 1512, 251–258. [Google Scholar] [CrossRef]
- Nagai, H.; Kim, Y.H. Cancer Prevention from the Perspective of Global Cancer Burden Patterns. J. Thorac. Dis. 2017, 9, 448–451. [Google Scholar] [CrossRef] [PubMed]
- Pahwa, R.; Jialal, I. Atherosclerosis. In StatPearls [Internet]; StatPearls Publishing LLC: Treasure Island, FL, USA, 2024. [Google Scholar]
- Yang, Y.; Seo, J.M.; Nguyen, A.; Pham, T.X.; Park, H.J.; Park, Y.; Kim, B.; Bruno, R.S.; Lee, J. Astaxanthin-Rich Extract from the Green Alga Haematococcus pluvialis Lowers Plasma Lipid Concentrations and Enhances Antioxidant Defense in Apolipoprotein E Knockout Mice. J. Nutr. 2011, 141, 1611–1617. [Google Scholar] [CrossRef] [PubMed]
- Chan, K.; Chen, S.; Chen, P. Astaxanthin Attenuated Thrombotic Risk Factors in Type 2 Diabetic Patients. J. Funct. Foods 2019, 53, 22–27. [Google Scholar] [CrossRef]
- Mashhadi, N.S.; Zakerkish, M.; Mohammadiasl, J.; Zarei, M.; Mohammadshahi, M.; Haghighizadeh, M.H. Astaxanthin Improves Glucose Metabolism and Reduces Blood Pressure in Patients with Type 2 Diabetes Mellitus. Asia Pac. J. Clin. Nutr. 2018, 27, 341–346. [Google Scholar] [CrossRef] [PubMed]
- Urakaze, M.; Kobashi, C.; Satou, Y.; Shigeta, K.; Toshima, M.; Takagi, M.; Takahashi, J.; Nishida, H. The Beneficial Effects of Astaxanthin on Glucose Metabolism and Modified Low-Density Lipoprotein in Healthy Volunteers and Subjects with Prediabetes. Nutrients 2021, 13, 4381. [Google Scholar] [CrossRef] [PubMed]
- Kurihara, H.; Koda, H.; Asami, S.; Kiso, Y.; Tanaka, T. Contribution of the Antioxidative Property of Astaxanthin to Its Protective Effect on the Promotion of Cancer Metastasis in Mice Treated with Restraint Stress. Life Sci. 2002, 70, 2509–2520. [Google Scholar] [CrossRef]
- Palozza, P.; Torelli, C.; Boninsegna, A.; Simone, R.; Catalano, A.; Mele, M.C.; Picci, N. Growth-Inhibitory Effects of the Astaxanthin-Rich Alga Haematococcus pluvialis in Human Colon Cancer Cells. Cancer Lett. 2009, 283, 108–117. [Google Scholar] [CrossRef] [PubMed]
- Park, J.S.; Chyun, J.H.; Kim, Y.K.; Line, L.L.; Chew, B.P. Astaxanthin Decreased Oxidative Stress and Inflammation and Enhanced Immune Response in Humans. Nutr. Metab. 2010, 7, 18. [Google Scholar] [CrossRef]
- Jiang, X.; Chen, L.; Shen, L.; Chen, Z.; Xu, L.; Zhang, J.; Yu, X. Trans-Astaxanthin Attenuates Lipopolysaccharide-Induced Neuroinflammation and Depressive-like Behavior in Mice. Brain Res. 2016, 1649, 30–37. [Google Scholar] [CrossRef]
- Nakajima, Y.; Inokuchi, Y.; Shimazawa, M.; Otsubo, K.; Ishibashi, T.; Hara, H. Astaxanthin, a Dietary Carotenoid, Protects Retinal Cells against Oxidative Stress in-Vitro and in Mice in-Vivo. J. Pharm. Pharmacol. 2008, 60, 1365–1374. [Google Scholar] [CrossRef] [PubMed]
- Ito, N.; Seki, S.; Ueda, F. The Protective Role of Astaxanthin for UV-Induced Skin Deterioration in Healthy People—A Randomized, Double-Blind, Placebo-Controlled Trial. Nutrients 2018, 10, 817. [Google Scholar] [CrossRef] [PubMed]
- Shete, V.; Quadro, L. Mammalian Metabolism of β-Carotene: Gaps in Knowledge. Nutrients 2013, 5, 4849–4868. [Google Scholar] [CrossRef] [PubMed]
- Mordor Intelligence Beta Carotene Market Size & Share Analysis—Growth Trends & Forecasts (2024–2029). Available online: https://www.mordorintelligence.com/industry-reports/beta-carotene-market (accessed on 25 January 2024).
- El Baz, F.K.; Aboul-Enein, A.M.; El-Baroty, G.S.; Youssef, A.M.; Abdel-Baky, H.H. Accumulation of Antioxidant Vitamins in Dunaliella salina. J. Biol. Sci. 2002, 2, 220–223. [Google Scholar] [CrossRef]
- Rigo Roso, G. The Bioeconomy of Microalgal Carotenoid-Rich Oleoresins Produced in Agroindustrial Biorefineries. J. Chem. Eng. Process Technol. 2015, 6, 1000218. [Google Scholar] [CrossRef]
- Ren, Y.; Sun, H.; Deng, J.; Huang, J.; Chen, F. Carotenoid Production from Microalgae: Biosynthesis, Salinity Responses and Novel Biotechnologies. Mar. Drugs 2021, 19, 713. [Google Scholar] [CrossRef] [PubMed]
- Von Lintig, J. Provitamin A Metabolism and Functions in Mammalian Biology. Am. J. Clin. Nutr. 2012, 96, 1234S–1244S. [Google Scholar] [CrossRef] [PubMed]
- Karppi, J.; Laukkanen, J.A.; Mäkikallio, T.H.; Ronkainen, K.; Kurl, S. Low β-Carotene Concentrations Increase the Risk of Cardiovascular Disease Mortality among Finnish Men with Risk Factors. Nutr. Metab. Cardiovasc. Dis. 2012, 22, 921–928. [Google Scholar] [CrossRef] [PubMed]
- Ito, Y.; Kurata, M.; Suzuki, K.; Hamajima, N.; Hishida, H.; Aoki, K. Cardiovascular Disease Mortality and Serum Carotenoid Levels: A Japanese Population-Based Follow-up Study. J. Epidemiol. 2006, 16, 154–160. [Google Scholar] [CrossRef]
- Karppi, J.; Laukkanen, J.A.; Mäkikallio, T.H.; Ronkainen, K.; Kurl, S. Serum β-Carotene and the Risk of Sudden Cardiac Death in Men: A Population-Based Follow-up Study. Atherosclerosis 2013, 226, 172–177. [Google Scholar] [CrossRef]
- Aly, H.K.; El-Baz, F.; Salama, A.; Ali, S.I.; El Askary, H.M.; Khalil, A. Assessment of β-Carotenoids Extracted from Dunaliella salina Microalgae: Toxicological Aspects. Egypt. J. Chem. 2021, 65, 727–739. [Google Scholar] [CrossRef]
- Lin, H.-W.; Liu, C.-W.; Yang, D.-J.; Chen, C.-C.; Chen, S.-Y.; Tseng, J.-K.; Chang, T.-J.; Chang, Y.-Y. Dunaliella salina Alga Extract Inhibits the Production of Interleukin-6, Nitric Oxide, and Reactive Oxygen Species by Regulating Nuclear Factor-ΚB/Janus Kinase/Signal Transducer and Activator of Transcription in Virus-Infected RAW264.7 Cells. J. Food Drug Anal. 2017, 25, 908–918. [Google Scholar] [CrossRef] [PubMed]
- Murthy, K.N.C.; Vanitha, A.; Rajesha, J.; Swamy, M.M.; Sowmya, P.R.; Ravishankar, G.A. In Vivo Antioxidant Activity of Carotenoids from Dunaliella salina—A Green Microalga. Life Sci. 2005, 76, 1381–1390. [Google Scholar] [CrossRef] [PubMed]
- Hsu, Y.-W.; Tsai, C.-F.; Chang, W.-H.; Ho, Y.-C.; Chen, W.-K.; Lu, F.-J. Protective Effects of Dunaliella salina—A Carotenoids-Rich Alga, against Carbon Tetrachloride-Induced Hepatotoxicity in Mice. Food Chem. Toxicol. 2008, 46, 3311–3317. [Google Scholar] [CrossRef] [PubMed]
- Heinrich, U.; Wiebusch, M.; Tronnier, H.; Gärtner, C.; Eichler, O.; Sies, H.; Stahl, W. Supplementation with β-Carotene or a Similar Amount of Mixed Carotenoids Protects Humans from UV-Induced Erythema. J. Nutr. 2003, 133, 98–101. [Google Scholar] [CrossRef] [PubMed]
- Tsai, C.-F.; Lu, F.-J.; Hsu, Y.-W. Protective Effects of Dunaliella salina—A Carotenoids-Rich Alga—Against Ultraviolet B-Induced Corneal Oxidative Damage in Mice. Mol. Vis. 2012, 18, 1540–1547. [Google Scholar] [PubMed]
- Zafar, J.; Aqeel, A.; Shah, F.I.; Ehsan, N.; Gohar, U.F.; Moga, M.A.; Festila, D.; Ciurea, C.; Irimie, M.; Chicea, R. Biochemical and Immunological Implications of Lutein and Zeaxanthin. Int. J. Mol. Sci. 2021, 22, 10910. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; George, E.W.; Rognon, G.T.; Gorusupudi, A.; Ranganathan, A.; Chang, F.-Y.; Shi, L.; Frederick, J.M.; Bernstein, P.S. Imaging Lutein and Zeaxanthin in the Human Retina with Confocal Resonance Raman Microscopy. Proc. Natl. Acad. Sci. USA 2020, 117, 12352–12358. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.E.; Leite, J.O.; deOgburn, R.; Smyth, J.A.; Clark, R.M.; Fernandez, M.L. A Lutein-Enriched Diet Prevents Cholesterol Accumulation and Decreases Oxidized LDL and Inflammatory Cytokines in the Aorta of Guinea Pigs. J. Nutr. 2011, 141, 1458–1463. [Google Scholar] [CrossRef]
- Qiao, Y.-Q.; Jiang, P.-F.; Gao, Y.-Z. Lutein Prevents Osteoarthritis through Nrf2 Activation and Downregulation of Inflammation. Arch. Med. Sci. 2018, 14, 617–624. [Google Scholar] [CrossRef]
- Murray, I.J.; Makridaki, M.; van der Veen, R.L.P.; Carden, D.; Parry, N.R.A.; Berendschot, T.T.J.M. Lutein Supplementation over a One-Year Period in Early AMD Might Have a Mild Beneficial Effect on Visual Acuity: The CLEAR Study. Investig. Ophthalmol. Vis. Sci. 2013, 54, 1781. [Google Scholar] [CrossRef] [PubMed]
- El-Baz, F.K.; Hussein, R.A.; Saleh, D.O.; Abdel Jaleel, G.A.R. Zeaxanthin Isolated from Dunaliella salina Microalgae Ameliorates Age Associated Cardiac Dysfunction in Rats through Stimulation of Retinoid Receptors. Mar. Drugs 2019, 17, 290. [Google Scholar] [CrossRef] [PubMed]
- Qi, J.; Kim, S.M. α-Glucosidase Inhibitory Activities of Lutein and Zeaxanthin Purified from Green Alga Chlorella ellipsoidea. J. Ocean. Univ. China 2018, 17, 983–989. [Google Scholar] [CrossRef]
- Chung, R.W.S.; Leanderson, P.; Lundberg, A.K.; Jonasson, L. Lutein Exerts Anti-Inflammatory Effects in Patients with Coronary Artery Disease. Atherosclerosis 2017, 262, 87–93. [Google Scholar] [CrossRef] [PubMed]
- United Nations Department of Economic and Social Affairs World Population Prospects—Population Division—United Nations 2022. Available online: http://population.un.org/wpp/ (accessed on 29 January 2024).
- Aiking, H.; de Boer, J. The next Protein Transition. Trends Food Sci. Technol. 2020, 105, 515–522. [Google Scholar] [CrossRef]
- Henchion, M.; Hayes, M.; Mullen, A.; Fenelon, M.; Tiwari, B. Future Protein Supply and Demand: Strategies and Factors Influencing a Sustainable Equilibrium. Foods 2017, 6, 53. [Google Scholar] [CrossRef] [PubMed]
- Becker, E.W. Micro-Algae as a Source of Protein. Biotechnol. Adv. 2007, 25, 207–210. [Google Scholar] [CrossRef] [PubMed]
- Assaye, H.; Belay, A.; Desse, G.; Gray, D. Seasonal Variation in the Nutrient Profile of Arthrospira Fusiformis Biomass Harvested from an Ethiopian Soda Lake, Lake Chitu. J. Appl. Phycol. 2018, 30, 1597–1606. [Google Scholar] [CrossRef]
- Brown, M.R. The Amino-Acid and Sugar Composition of 16 Species of Microalgae Used in Mariculture. J. Exp. Mar. Biol. Ecol. 1991, 145, 79–99. [Google Scholar] [CrossRef]
- Tokuşoglu, Ö.; üUnal, M.K. Biomass Nutrient Profiles of Three Microalgae: Spirulina platensis, Chlorella vulgaris, and Isochrisis galbana. J. Food Sci. 2003, 68, 1144–1148. [Google Scholar] [CrossRef]
- Waghmare, A.G.; Salve, M.K.; LeBlanc, J.G.; Arya, S.S. Concentration and Characterization of Microalgae Proteins from Chlorella Pyrenoidosa. Bioresour. Bioprocess. 2016, 3, 16. [Google Scholar] [CrossRef]
- Safi, C.; Charton, M.; Pignolet, O.; Silvestre, F.; Vaca-Garcia, C.; Pontalier, P.-Y. Influence of Microalgae Cell Wall Characteristics on Protein Extractability and Determination of Nitrogen-to-Protein Conversion Factors. J. Appl. Phycol. 2013, 25, 523–529. [Google Scholar] [CrossRef]
- Andreeva, A.; Budenkova, E.; Babich, O.; Sukhikh, S.; Ulrikh, E.; Ivanova, S.; Prosekov, A.; Dolganyuk, V. Production, Purification, and Study of the Amino Acid Composition of Microalgae Proteins. Molecules 2021, 26, 2767. [Google Scholar] [CrossRef] [PubMed]
- Millward, D.J. Amino Acid Scoring Patterns for Protein Quality Assessment. Br. J. Nutr. 2012, 108, S31–S43. [Google Scholar] [CrossRef] [PubMed]
- Masojídek, J.; Torzillo, G.; Koblížek, M. Photosynthesis in Microalgae. In Handbook of Microalgal Culture; Wiley: Hoboken, NJ, USA, 2013; pp. 21–36. [Google Scholar]
- Tounsi, L.; Ben Hlima, H.; Hentati, F.; Hentati, O.; Derbel, H.; Michaud, P.; Abdelkafi, S. Microalgae: A Promising Source of Bioactive Phycobiliproteins. Mar. Drugs 2023, 21, 440. [Google Scholar] [CrossRef] [PubMed]
- Jha, S.; Singh, V.K.; Singh, A.P.; Gupta, A.; Rana, P.; Sinha, R.P. The Radiant World of Cyanobacterial Phycobiliproteins: Examining Their Structure, Functions, and Biomedical Potentials. Targets 2024, 2, 32–51. [Google Scholar] [CrossRef]
- Chen, H.; Qi, H.; Xiong, P. Phycobiliproteins—A Family of Algae-Derived Biliproteins: Productions, Characterization and Pharmaceutical Potentials. Mar. Drugs 2022, 20, 450. [Google Scholar] [CrossRef] [PubMed]
- Sekar, S.; Chandramohan, M. Phycobiliproteins as a Commodity: Trends in Applied Research, Patents and Commercialization. J. Appl. Phycol. 2008, 20, 113–136. [Google Scholar] [CrossRef]
- Chakdar, H.; Pabbi, S. Cyanobacterial Phycobilins: Production, Purification, and Regulation. In Frontier Discoveries and Innovations in Interdisciplinary Microbiology; Springer: New Delhi, India, 2016; pp. 45–69. [Google Scholar]
- Li, Y.; Li, X.; Liang, Z.-P.; Chang, X.-Y.; Li, F.-T.; Wang, X.-Q.; Lian, X.-J. Progress of Microencapsulated Phycocyanin in Food and Pharma Industries: A Review. Molecules 2022, 27, 5854. [Google Scholar] [CrossRef]
- Khandual, S.; Sanchez, E.O.L.; Andrews, H.E.; de la Rosa, J.D.P. Phycocyanin Content and Nutritional Profile of Arthrospira Platensis from Mexico: Efficient Extraction Process and Stability Evaluation of Phycocyanin. BMC Chem. 2021, 15, 24. [Google Scholar] [CrossRef]
- Grover, P.; Bhatnagar, A.; Kumari, N.; Narayan Bhatt, A.; Kumar Nishad, D.; Purkayastha, J. C-Phycocyanin-a Novel Protein from Spirulina Platensis- In Vivo Toxicity, Antioxidant and Immunomodulatory Studies. Saudi J. Biol. Sci. 2021, 28, 1853–1859. [Google Scholar] [CrossRef] [PubMed]
- Shih, C.-M.; Cheng, S.-N.; Wong, C.-S.; Kuo, Y.-L.; Chou, T.-C. Antiinflammatory and Antihyperalgesic Activity of C-Phycocyanin. Anesth. Analg. 2009, 108, 1303–1310. [Google Scholar] [CrossRef] [PubMed]
- Environment Climate Change and Health Team of World Health Organization Radiation: The Known Health Effects of Ultraviolet Radiation. Available online: https://www.who.int/news-room/questions-and-answers/item/radiation-the-known-health-effects-of-ultraviolet-radiation (accessed on 1 February 2024).
- Jang, Y.A.; Kim, B.A. Protective Effect of Spirulina-Derived C-Phycocyanin against Ultraviolet B-Induced Damage in HaCaT Cells. Medicina 2021, 57, 273. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.M.; Lee, J.Y.; Im, A.-R.; Chae, S. Phycocyanin Protects Against UVB-Induced Apoptosis Through the PKC α/ΒII-Nrf-2/HO-1 Dependent Pathway in Human Primary Skin Cells. Molecules 2018, 23, 478. [Google Scholar] [CrossRef] [PubMed]
- Nagaoka, S.; Shimizu, K.; Kaneko, H.; Shibayama, F.; Morikawa, K.; Kanamaru, Y.; Otsuka, A.; Hirahashi, T.; Kato, T. A Novel Protein C-Phycocyanin Plays a Crucial Role in the Hypocholesterolemic Action of Spirulina Platensis Concentrate in Rats. J. Nutr. 2005, 135, 2425–2430. [Google Scholar] [CrossRef] [PubMed]
- Riss, J.; Décordé, K.; Sutra, T.; Delage, M.; Baccou, J.-C.; Jouy, N.; Brune, J.-P.; Oréal, H.; Cristol, J.-P.; Rouanet, J.-M. Phycobiliprotein C-Phycocyanin from Spirulina platensis Is Powerfully Responsible for Reducing Oxidative Stress and NADPH Oxidase Expression Induced by an Atherogenic Diet in Hamsters. J. Agric. Food Chem. 2007, 55, 7962–7967. [Google Scholar] [CrossRef] [PubMed]
- Hsiao, G.; Chou, P.-H.; Shen, M.-Y.; Chou, D.-S.; Lin, C.-H.; Sheu, J.-R. C-Phycocyanin, a Very Potent and Novel Platelet Aggregation Inhibitor from Spirulina platensis. J. Agric. Food Chem. 2005, 53, 7734–7740. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, M.; Perumal, Y.; Bansal, S.; Arora, S.; Chopra, K. Phycocyanin Alleviates ICV-STZ Induced Cognitive and Molecular Deficits via PI3-Kinase Dependent Pathway. Food Chem. Toxicol. 2020, 145, 111684. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Gan, L.; Yan, S.; Yan, Y.; Huang, W. Effect of C-Phycocyanin on HDAC3 and MiRNA-335 in Alzheimer’s Disease. Transl. Neurosci. 2020, 11, 161–172. [Google Scholar] [CrossRef]
- Cervantes-Llanos, M.; Lagumersindez-Denis, N.; Marín-Prida, J.; Pavón-Fuentes, N.; Falcon-Cama, V.; Piniella-Matamoros, B.; Camacho-Rodríguez, H.; Fernández-Massó, J.R.; Valenzuela-Silva, C.; Raíces-Cruz, I.; et al. Beneficial Effects of Oral Administration of C-Phycocyanin and Phycocyanobilin in Rodent Models of Experimental Autoimmune Encephalomyelitis. Life Sci. 2018, 194, 130–138. [Google Scholar] [CrossRef]
- Hao, S.; Yan, Y.; Li, S.; Zhao, L.; Zhang, C.; Liu, L.; Wang, C. The In Vitro Anti-Tumor Activity of Phycocyanin against Non-Small Cell Lung Cancer Cells. Mar. Drugs 2018, 16, 178. [Google Scholar] [CrossRef] [PubMed]
- Pan, D.; Huang, B.; Gan, Y.; Gao, C.; Liu, Y.; Tang, Z. Phycocyanin Ameliorates Colitis-Associated Colorectal Cancer by Regulating the Gut Microbiota and the IL-17 Signaling Pathway. Mar. Drugs 2022, 20, 260. [Google Scholar] [CrossRef] [PubMed]
- Liao, G.; Gao, B.; Gao, Y.; Yang, X.; Cheng, X.; Ou, Y. Phycocyanin Inhibits Tumorigenic Potential of Pancreatic Cancer Cells: Role of Apoptosis and Autophagy. Sci. Rep. 2016, 6, 34564. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Wang, Y.; Liu, G.; Liu, H.; Zhu, F.; Ji, H.; Li, B. C-Phycocyanin Exerts Anti-Cancer Effects via the MAPK Signaling Pathway in MDA-MB-231 Cells. Cancer Cell Int. 2018, 18, 12. [Google Scholar] [CrossRef] [PubMed]
- Siti Halimatul Munawaroh, H.; Gumilar, G.G.; Nurjanah, F.; Yuliani, G.; Aisyah, S.; Kurnia, D.; Wulandari, A.P.; Kurniawan, I.; Ningrum, A.; Koyande, A.K.; et al. In-Vitro Molecular Docking Analysis of Microalgae Extracted Phycocyanin as an Anti-Diabetic Candidate. Biochem. Eng. J. 2020, 161, 107666. [Google Scholar] [CrossRef]
- Ou, Y.; Ren, Z.; Wang, J.; Yang, X. Phycocyanin Ameliorates Alloxan-Induced Diabetes Mellitus in Mice: Involved in Insulin Signaling Pathway and GK Expression. Chem. Biol. Interact. 2016, 247, 49–54. [Google Scholar] [CrossRef] [PubMed]
- Ou, Y.; Lin, L.; Yang, X.; Pan, Q.; Cheng, X. Antidiabetic Potential of Phycocyanin: Effects on KKAy Mice. Pharm. Biol. 2013, 51, 539–544. [Google Scholar] [CrossRef] [PubMed]
- Jadaun, P.; Seniya, C.; Pal, S.K.; Kumar, S.; Kumar, P.; Nema, V.; Kulkarni, S.S.; Mukherjee, A. Elucidation of Antiviral and Antioxidant Potential of C-Phycocyanin against HIV-1 Infection through In Silico and In Vitro Approaches. Antioxidants 2022, 11, 1942. [Google Scholar] [CrossRef] [PubMed]
- Setthamongkol, P.; Kulert, W.; Wanmanee, S.; Swami, R.; Kutako, M.; Chanthathamrongsiri, N.; Semangoen, T.; Hiransuchalert, R. In Vitro Characterization and Assessment of a Potential Cosmetic Cream Containing Phycocyanin Extracted from Arthrospira Platensis BUUC1503 Blue-Green Algae. J. Appl. Phycol. 2023, 35, 1685–1697. [Google Scholar] [CrossRef]
- Nihal, B.; Gupta, N.V.; Gowda, D.V.M. Formulation and development of topical anti acne formulation of spirulina extract. Int. J. Appl. Pharm. 2018, 10, 229. [Google Scholar] [CrossRef]
- Sarada, D.V.L.; Sreenath Kumar, C.; Rengasamy, R. Purified C-Phycocyanin from Spirulina platensis (Nordstedt) Geitler: A Novel and Potent Agent against Drug Resistant Bacteria. World J. Microbiol. Biotechnol. 2011, 27, 779–783. [Google Scholar] [CrossRef]
- Tan, H.T.; Yusoff, F.M.; Khaw, Y.S.; Noor Mazli, N.A.I.; Nazarudin, M.F.; Shaharuddin, N.A.; Katayama, T.; Ahmad, S.A. A Review on a Hidden Gem: Phycoerythrin from Blue-Green Algae. Mar. Drugs 2022, 21, 28. [Google Scholar] [CrossRef] [PubMed]
- Future Market Insights Phycoerythrin Market. Available online: https://www.futuremarketinsights.com/reports/phycoerythrin-market (accessed on 2 February 2024).
- Patel, S.N.; Sonani, R.R.; Jakharia, K.; Bhastana, B.; Patel, H.M.; Chaubey, M.G.; Singh, N.K.; Madamwar, D. Antioxidant Activity and Associated Structural Attributes of Halomicronema Phycoerythrin. Int. J. Biol. Macromol. 2018, 111, 359–369. [Google Scholar] [CrossRef] [PubMed]
- Soni, B.; Visavadiya, N.P.; Madamwar, D. Attenuation of Diabetic Complications by C-Phycoerythrin in Rats: Antioxidant Activity of C-Phycoerythrin Including Copper-Induced Lipoprotein and Serum Oxidation. Br. J. Nutr. 2009, 102, 102–109. [Google Scholar] [CrossRef] [PubMed]
- Chaubey, M.G.; Patel, S.N.; Rastogi, R.P.; Srivastava, P.L.; Singh, A.K.; Madamwar, D.; Singh, N.K. Therapeutic Potential of Cyanobacterial Pigment Protein Phycoerythrin: In Silico and in Vitro Study of BACE1 Interaction and in Vivo Aβ Reduction. Int. J. Biol. Macromol. 2019, 134, 368–378. [Google Scholar] [CrossRef] [PubMed]
- Sonani, R.R.; Singh, N.K.; Kumar, J.; Thakar, D.; Madamwar, D. Concurrent Purification and Antioxidant Activity of Phycobiliproteins from Lyngbya sp. A09DM: An Antioxidant and Anti-Aging Potential of Phycoerythrin in Caenorhabditis elegans. Process Biochem. 2014, 49, 1757–1766. [Google Scholar] [CrossRef]
- Sonani, R.R.; Singh, N.K.; Awasthi, A.; Prasad, B.; Kumar, J.; Madamwar, D. Phycoerythrin Extends Life Span and Health Span of Caenorhabditis elegans. Age 2014, 36, 9717. [Google Scholar] [CrossRef] [PubMed]
- O’Connor, J.; Garcia-Vaquero, M.; Meaney, S.; Tiwari, B.K. Bioactive Peptides from Algae: Traditional and Novel Generation Strategies, Structure-Function Relationships, and Bioinformatics as Predictive Tools for Bioactivity. Mar. Drugs 2022, 20, 317. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.-K.; Kang, K.-H. Medicinal Effects of Peptides from Marine Microalgae. In Advances in Food and Nutrition Research; Academic Press: Cambridge, MA, USA, 2011; pp. 313–323. [Google Scholar]
- Bai, R.; Nguyen, T.T.; Zhou, Y.; Diao, Y.; Zhang, W. Identification of Antioxidative Peptides Derived from Arthrospira Maxima in the Biorefinery Process after Extraction of C-Phycocyanin and Lipids. Mar. Drugs 2023, 21, 146. [Google Scholar] [CrossRef] [PubMed]
- Sheih, I.-C.; Fang, T.J.; Wu, T.-K.; Lin, P.-H. Anticancer and Antioxidant Activities of the Peptide Fraction from Algae Protein Waste. J. Agric. Food Chem. 2010, 58, 1202–1207. [Google Scholar] [CrossRef] [PubMed]
- Ko, S.-C.; Kim, D.; Jeon, Y.-J. Protective Effect of a Novel Antioxidative Peptide Purified from a Marine Chlorella ellipsoidea Protein against Free Radical-Induced Oxidative Stress. Food Chem. Toxicol. 2012, 50, 2294–2302. [Google Scholar] [CrossRef] [PubMed]
- Sheih, I.-C.; Wu, T.-K.; Fang, T.J. Antioxidant Properties of a New Antioxidative Peptide from Algae Protein Waste Hydrolysate in Different Oxidation Systems. Bioresour. Technol. 2009, 100, 3419–3425. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.-F.; Zhang, Y.Y.; Di He, M.; Li, C.Y.; Zhou, C.X.; Hong, P.Z.; Qian, Z.-J. Antioxidant Peptide Purified from Enzymatic Hydrolysates of Isochrysis zhanjiangensis and Its Protective Effect against Ethanol Induced Oxidative Stress of HepG2 Cells. Biotechnol. Bioprocess. Eng. 2019, 24, 308–317. [Google Scholar] [CrossRef]
- Safitri, N.M.; Herawati, E.Y.; Hsu, J.L. Antioxidant Activity of Purified Active Peptide Derived from Spirulina Platensis Enzymatic Hydrolysates. Res. J. Life Sci. 2017, 4, 119–128. [Google Scholar] [CrossRef]
- Wong, M.K.S. Angiotensin Converting Enzymes. In Handbook of Hormones; Elsevier: Amsterdam, The Netherlands, 2016; pp. 505–508. [Google Scholar]
- Heo, S.-Y.; Ko, S.-C.; Kim, C.S.; Oh, G.-W.; Ryu, B.; Qian, Z.-J.; Kim, G.; Park, W.S.; Choi, I.-W.; Phan, T.T.V.; et al. A Heptameric Peptide Purified from Spirulina sp. Gastrointestinal Hydrolysate Inhibits Angiotensin I-Converting Enzyme- and Angiotensin II-Induced Vascular Dysfunction in Human Endothelial Cells. Int. J. Mol. Med. 2017, 39, 1072–1082. [Google Scholar] [CrossRef]
- Suetsuna, K.; Chen, J.-R. Identification of Antihypertensive Peptides from Peptic Digest of Two Microalgae, Chlorella Vulgaris and Spirulina platensis. Mar. Biotechnol. 2001, 3, 305–309. [Google Scholar] [CrossRef]
- Ko, S.-C.; Kang, N.; Kim, E.-A.; Kang, M.C.; Lee, S.-H.; Kang, S.-M.; Lee, J.-B.; Jeon, B.-T.; Kim, S.-K.; Park, S.-J.; et al. A Novel Angiotensin I-Converting Enzyme (ACE) Inhibitory Peptide from a Marine Chlorella ellipsoidea and Its Antihypertensive Effect in Spontaneously Hypertensive Rats. Process Biochem. 2012, 47, 2005–2011. [Google Scholar] [CrossRef]
- Samarakoon, K.W.; O-Nam, K.; Ko, J.-Y.; Lee, J.-H.; Kang, M.-C.; Kim, D.; Lee, J.B.; Lee, J.-S.; Jeon, Y.-J. Purification and Identification of Novel Angiotensin-I Converting Enzyme (ACE) Inhibitory Peptides from Cultured Marine Microalgae (Nannochloropsis oculata) Protein Hydrolysate. J. Appl. Phycol. 2013, 25, 1595–1606. [Google Scholar] [CrossRef]
- Vo, T.-S.; Kim, S.-K. Down-Regulation of Histamine-Induced Endothelial Cell Activation as Potential Anti-Atherosclerotic Activity of Peptides from Spirulina maxima. Eur. J. Pharm. Sci. 2013, 50, 198–207. [Google Scholar] [CrossRef]
- Shih, M.; Chen, L.; Cherng, J. Chlorella 11-Peptide Inhibits the Production of Macrophage-Induced Adhesion Molecules and Reduces Endothelin-1 Expression and Endothelial Permeability. Mar. Drugs 2013, 11, 3861–3874. [Google Scholar] [CrossRef]
- Wang, Z.; Zhang, X. Isolation and Identification of Anti-proliferative Peptides from Spirulina platensis Using Three-step Hydrolysis. J. Sci. Food Agric. 2017, 97, 918–922. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Zhang, X. Inhibitory Effects of Small Molecular Peptides from Spirulina (Arthrospira) Platensis on Cancer Cell Growth. Food Funct. 2016, 7, 781–788. [Google Scholar] [CrossRef]
- Morris, H.J.; Carrillo, O.; Almarales, A.; Bermúdez, R.C.; Lebeque, Y.; Fontaine, R.; Llauradó, G.; Beltrán, Y. Immunostimulant Activity of an Enzymatic Protein Hydrolysate from Green Microalga Chlorella Vulgaris on Undernourished Mice. Enzym. Microb. Technol. 2007, 40, 456–460. [Google Scholar] [CrossRef]
- Vo, T.-S.; Ryu, B.; Kim, S.-K. Purification of Novel Anti-Inflammatory Peptides from Enzymatic Hydrolysate of the Edible Microalgal Spirulina maxima. J. Funct. Foods 2013, 5, 1336–1346. [Google Scholar] [CrossRef]
- Vo, T.; Ngo, D.; Kang, K.; Park, S.; Kim, S. The Role of Peptides Derived from Spirulina maxima in Downregulation of FcεRI-mediated Allergic Responses. Mol. Nutr. Food Res. 2014, 58, 2226–2234. [Google Scholar] [CrossRef] [PubMed]
- Cherng, J.Y.; Liu, C.C.; Shen, C.R.; Lin, H.H.; Shih, M.F. Beneficial Effects of Chlorella -11 Peptide on Blocking LPS-Induced Macrophage Activation and Alleviating Thermal Injury-Induced Inflammation in Rats. Int. J. Immunopathol. Pharmacol. 2010, 23, 811–820. [Google Scholar] [CrossRef] [PubMed]
- Kumari, P.; Kumar, M.; Reddy, C.R.K.; Jha, B. Algal Lipids, Fatty Acids and Sterols. In Functional Ingredients from Algae for Foods and Nutraceuticals; Elsevier: Amsterdam, The Netherlands, 2013; pp. 87–134. [Google Scholar]
- Okazaki, Y.; Saito, K. Roles of Lipids as Signaling Molecules and Mitigators during Stress Response in Plants. Plant J. 2014, 79, 584–596. [Google Scholar] [CrossRef] [PubMed]
- Kaur, N.; Chugh, V.; Gupta, A.K. Essential Fatty Acids as Functional Components of Foods—A Review. J. Food Sci. Technol. 2014, 51, 2289–2303. [Google Scholar] [CrossRef] [PubMed]
- EFSA Panel on Dietetic Products, N. and A. Scientific Opinion on Dietary Reference Values for Fats, Including Saturated Fatty Acids, Polyunsaturated Fatty Acids, Monounsaturated Fatty Acids, Trans Fatty Acids, and Cholesterol. EFSA J. 2010, 8, 1461. [Google Scholar] [CrossRef]
- Wilson, B.A.; Pollard, R.D.; Ferguson, D.S. Nutriential Hazards: Macronutrients: Essential Fatty Acids. In Encyclopedia of Food Safety; Elsevier: Amsterdam, The Netherlands, 2014; pp. 95–102. [Google Scholar]
- Scott, D.D.; Krupanidhi, S.; Carani, B. Sanjeevi Omega-3 Fatty Acids for Nutrition and Medicine: Considering Microalgae Oil as a Vegetarian Source of EPA and DHA. Curr. Diabetes Rev. 2007, 3, 198–203. [Google Scholar] [CrossRef]
- Oliver, L.; Dietrich, T.; Marañón, I.; Villarán, M.C.; Barrio, R.J. Producing Omega-3 Polyunsaturated Fatty Acids: A Review of Sustainable Sources and Future Trends for the EPA and DHA Market. Resources 2020, 9, 148. [Google Scholar] [CrossRef]
- Tocher, D.; Betancor, M.; Sprague, M.; Olsen, R.; Napier, J. Omega-3 Long-Chain Polyunsaturated Fatty Acids, EPA and DHA: Bridging the Gap between Supply and Demand. Nutrients 2019, 11, 89. [Google Scholar] [CrossRef] [PubMed]
- Kris-Etherton, P.M.; Harris, W.S.; Appel, L.J. Fish Consumption, Fish Oil, Omega-3 Fatty Acids, and Cardiovascular Disease. Circulation 2002, 106, 2747–2757. [Google Scholar] [CrossRef] [PubMed]
- Dwyer, J. Vegeterian Diets. In Encyclopedia of Human Nutrition; Elsevier: Amsterdam, The Netherlands, 2013; pp. 316–322. [Google Scholar]
- Colombo, S.M.; Rodgers, T.F.M.; Diamond, M.L.; Bazinet, R.P.; Arts, M.T. Projected Declines in Global DHA Availability for Human Consumption as a Result of Global Warming. Ambio 2020, 49, 865–880. [Google Scholar] [CrossRef] [PubMed]
- Chi, G.; Xu, Y.; Cao, X.; Li, Z.; Cao, M.; Chisti, Y.; He, N. Production of Polyunsaturated Fatty Acids by Schizochytrium (Aurantiochytrium) spp. Biotechnol. Adv. 2022, 55, 107897. [Google Scholar] [CrossRef] [PubMed]
- Park, J.H.; Upadhaya, S.D.; Kim, I.H. Effect of Dietary Marine Microalgae (Schizochytrium) Powder on Egg Production, Blood Lipid Profiles, Egg Quality, and Fatty Acid Composition of Egg Yolk in Layers. Asian-Australas. J. Anim. Sci. 2015, 28, 391–397. [Google Scholar] [CrossRef] [PubMed]
- Patil, V.; Källqvist, T.; Olsen, E.; Vogt, G.; Gislerød, H.R. Fatty Acid Composition of 12 Microalgae for Possible Use in Aquaculture Feed. Aquac. Int. 2007, 15, 1–9. [Google Scholar] [CrossRef]
- Matos, Â.P.; Feller, R.; Moecke, E.H.S.; de Oliveira, J.V.; Junior, A.F.; Derner, R.B.; Sant’Anna, E.S. Chemical Characterization of Six Microalgae with Potential Utility for Food Application. J. Am. Oil Chem. Soc. 2016, 93, 963–972. [Google Scholar] [CrossRef]
- Yang, Y.; Du, L.; Hosokawa, M.; Miyashita, K. Total Lipids Content, Lipid Class and Fatty Acid Composition of Ten Species of Microalgae. J. Oleo Sci. 2020, 69, 1181–1189. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Wang, Z.; Yu, C.; Yin, Y.; Zhou, G. Evaluation of the Potential of 9 Nannochloropsis Strains for Biodiesel Production. Bioresour. Technol. 2014, 167, 503–509. [Google Scholar] [CrossRef]
- Otleş, S.; Pire, R. Fatty Acid Composition of Chlorella and Spirulina microalgae Species. J. AOAC Int. 2001, 84, 1708–1714. [Google Scholar] [CrossRef] [PubMed]
- Cakmak, Y.S.; Kaya, M.; Asan-Ozusaglam, M. Biochemical Composition and Bioactivity Screening of Various Extracts from Dunaliella Salina, a Green Microalga. EXCLI J. 2014, 13, 679–690. [Google Scholar] [PubMed]
- Swanson, D.; Block, R.; Mousa, S.A. Omega-3 Fatty Acids EPA and DHA: Health Benefits Throughout Life. Adv. Nutr. 2012, 3, 1–7. [Google Scholar] [CrossRef] [PubMed]
- van Beelen, V.A.; Spenkelink, B.; Mooibroek, H.; Sijtsma, L.; Bosch, D.; Rietjens, I.M.C.M.; Alink, G.M. An N-3 PUFA-Rich Microalgal Oil Diet Protects to a Similar Extent as a Fish Oil-Rich Diet against AOM-Induced Colonic Aberrant Crypt Foci in F344 Rats. Food Chem. Toxicol. 2009, 47, 316–320. [Google Scholar] [CrossRef] [PubMed]
- Ryckebosch, E.; Bruneel, C.; Termote-Verhalle, R.; Goiris, K.; Muylaert, K.; Foubert, I. Nutritional Evaluation of Microalgae Oils Rich in Omega-3 Long Chain Polyunsaturated Fatty Acids as an Alternative for Fish Oil. Food Chem. 2014, 160, 393–400. [Google Scholar] [CrossRef] [PubMed]
- Geppert, J.; Kraft, V.; Demmelmair, H.; Koletzko, B. Docosahexaenoic Acid Supplementation in Vegetarians Effectively Increases Omega-3 Index: A Randomized Trial. Lipids 2005, 40, 807–814. [Google Scholar] [CrossRef] [PubMed]
- Harris, W.S. The Omega-3 Index: Clinical Utility for Therapeutic Intervention. Curr. Cardiol. Rep. 2010, 12, 503–508. [Google Scholar] [CrossRef] [PubMed]
- García-Maldonado, E.; Alcorta, A.; Zapatera, B.; Vaquero, M.P. Changes in Fatty Acid Levels after Consumption of a Novel Docosahexaenoic Supplement from Algae: A Crossover Randomized Controlled Trial in Omnivorous, Lacto-Ovo Vegetarians and Vegans. Eur. J. Nutr. 2023, 62, 1691–1705. [Google Scholar] [CrossRef] [PubMed]
- Morgese, M.; Mhillaj, E.; Francavilla, M.; Bove, M.; Morgano, L.; Tucci, P.; Trabace, L.; Schiavone, S. Chlorella sorokiniana Extract Improves Short-Term Memory in Rats. Molecules 2016, 21, 1311. [Google Scholar] [CrossRef]
- Lai, Y.-J. Omega-3 Fatty Acid Obtained from Nannochloropsis oceanica Cultures Grown under Low Urea Protect against Abeta-Induced Neural Damage. J. Food Sci. Technol. 2015, 52, 2982–2989. [Google Scholar] [CrossRef]
- Lauritzen, L.; Brambilla, P.; Mazzocchi, A.; Harsløf, L.; Ciappolino, V.; Agostoni, C. DHA Effects in Brain Development and Function. Nutrients 2016, 8, 6. [Google Scholar] [CrossRef] [PubMed]
- Balakrishnan, J.; Krishnan, B.; Sekar, T.; Ayothi, P.; Muthu, R.; Sampathrajan, V.; Muthu, A.; Dhavamani, S.; Shanmugam, K. Maternal Supplementation of High-Value PUFA-Rich Isochrysis sp. Biomass Prevents Monosodium Glutamate-Induced Neurotoxicity in First-Generation Wistar Rats. Neurochem. Int. 2022, 154, 105292. [Google Scholar] [CrossRef] [PubMed]
- Conde, T.A.; Neves, B.F.; Couto, D.; Melo, T.; Neves, B.; Costa, M.; Silva, J.; Domingues, P.; Domingues, M.R. Microalgae as Sustainable Bio-Factories of Healthy Lipids: Evaluating Fatty Acid Content and Antioxidant Activity. Mar. Drugs 2021, 19, 357. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Z.; Wang, B.; Ibrar, M.; Ying, M.; Li, S.; Yang, X. Schizochytrium sp. Extracted Lipids Prevent Alopecia by Enhancing Antioxidation and Inhibiting Ferroptosis of Dermal Papilla Cells. Antioxidants 2023, 12, 1332. [Google Scholar] [CrossRef]
- Robertson, R.; Guihéneuf, F.; Bahar, B.; Schmid, M.; Stengel, D.; Fitzgerald, G.; Ross, R.; Stanton, C. The Anti-Inflammatory Effect of Algae-Derived Lipid Extracts on Lipopolysaccharide (LPS)-Stimulated Human THP-1 Macrophages. Mar. Drugs 2015, 13, 5402–5424. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez-Pliego, L.E.; Martínez-Carrillo, B.E.; Reséndiz-Albor, A.A.; Arciniega-Martínez, I.M.; Escoto-Herrera, J.A.; Rosales-Gómez, C.A.; Valdés-Ramos, R. Effect of Supplementation with n-3 Fatty Acids Extracted from Microalgae on Inflammation Biomarkers from Two Different Strains of Mice. J. Lipids 2018, 2018, 4765358. [Google Scholar] [CrossRef] [PubMed]
- Dawczynski, C.; Dittrich, M.; Neumann, T.; Goetze, K.; Welzel, A.; Oelzner, P.; Völker, S.; Schaible, A.M.; Troisi, F.; Thomas, L.; et al. Docosahexaenoic Acid in the Treatment of Rheumatoid Arthritis: A Double-Blind, Placebo-Controlled, Randomized Cross-over Study with Microalgae vs. Sunflower Oil. Clin. Nutr. 2018, 37, 494–504. [Google Scholar] [CrossRef] [PubMed]
- Vilakazi, H.; Olasehinde, T.A.; Olaniran, A.O. Chemical Characterization, Antiproliferative and Antioxidant Activities of Polyunsaturated Fatty Acid-Rich Extracts from Chlorella sp. S14. Molecules 2021, 26, 4109. [Google Scholar] [CrossRef] [PubMed]
- Castejón, N.; Marko, D. Fatty Acid Composition and Cytotoxic Activity of Lipid Extracts from Nannochloropsis Gaditana Produced by Green Technologies. Molecules 2022, 27, 3710. [Google Scholar] [CrossRef] [PubMed]
- Natesan, V.; Kim, S.-J. Lipid Metabolism, Disorders and Therapeutic Drugs—Review. Biomol. Ther. 2021, 29, 596–604. [Google Scholar] [CrossRef]
- Bernstein, A.M.; Ding, E.L.; Willett, W.C.; Rimm, E.B. A Meta-Analysis Shows That Docosahexaenoic Acid from Algal Oil Reduces Serum Triglycerides and Increases HDL-Cholesterol and LDL-Cholesterol in Persons without Coronary Heart Disease3. J. Nutr. 2012, 142, 99–104. [Google Scholar] [CrossRef] [PubMed]
- de Mello-Sampayo, C.; Paterna, A.; Polizzi, A.; Duarte, D.; Batista, I.; Pinto, R.; Gonçalves, P.; Raymundo, A.; Batista, A.; Gouveia, L.; et al. Evaluation of Marine Microalga Diacronema vlkianum Biomass Fatty Acid Assimilation in Wistar Rats. Molecules 2017, 22, 1097. [Google Scholar] [CrossRef] [PubMed]
- Rao, A.; Briskey, D.; Nalley, J.O.; Ganuza, E. Omega-3 Eicosapentaenoic Acid (EPA) Rich Extract from the Microalga Nannochloropsis Decreases Cholesterol in Healthy Individuals: A Double-Blind, Randomized, Placebo-Controlled, Three-Month Supplementation Study. Nutrients 2020, 12, 1869. [Google Scholar] [CrossRef]
- Li, T.-T.; Liu, Y.-Y.; Wan, X.-Z.; Huang, Z.-R.; Liu, B.; Zhao, C. Regulatory Efficacy of the Polyunsaturated Fatty Acids from Microalgae spirulina Platensis on Lipid Metabolism and Gut Microbiota in High-Fat Diet Rats. Int. J. Mol. Sci. 2018, 19, 3075. [Google Scholar] [CrossRef] [PubMed]
- Wan, X.; Li, T.; Zhong, R.; Chen, H.; Xia, X.; Gao, L.; Gao, X.; Liu, B.; Zhang, H.; Zhao, C. Anti-Diabetic Activity of PUFAs-Rich Extracts of Chlorella pyrenoidosa and Spirulina platensis in Rats. Food Chem. Toxicol. 2019, 128, 233–239. [Google Scholar] [CrossRef] [PubMed]
- Macías-Sánchez, M.D.; Mantell, C.; Rodríguez, M.; Martínez de la Ossa, E.; Lubián, L.M.; Montero, O. Comparison of Supercritical Fluid and Ultrasound-Assisted Extraction of Carotenoids and Chlorophyll a from Dunaliella salina. Talanta 2009, 77, 948–952. [Google Scholar] [CrossRef] [PubMed]
- Rostagno, M.A.; Palma, M.; Barroso, C.G. Ultrasound-Assisted Extraction of Soy Isoflavones. J. Chromatogr. A 2003, 1012, 119–128. [Google Scholar] [CrossRef]
- Tavakoli, S.; Hong, H.; Wang, K.; Yang, Q.; Gahruie, H.H.; Zhuang, S.; Li, Y.; Liang, Y.; Tan, Y.; Luo, Y. Ultrasonic-Assisted Food-Grade Solvent Extraction of High-Value Added Compounds from Microalgae Spirulina platensis and Evaluation of Their Antioxidant and Antibacterial Properties. Algal Res. 2021, 60, 102493. [Google Scholar] [CrossRef]
- Kumar, Y.; Singhal, S.; Tarafdar, A.; Pharande, A.; Ganesan, M.; Badgujar, P.C. Ultrasound Assisted Extraction of Selected Edible Macroalgae: Effect on Antioxidant Activity and Quantitative Assessment of Polyphenols by Liquid Chromatography with Tandem Mass Spectrometry (LC-MS/MS). Algal Res. 2020, 52, 102114. [Google Scholar] [CrossRef]
- McCabe, W.L.; Smith, J.C.; Harriott, P. Unit Operations of Chemical Engineering; McGraw-Hill Chemical Engineering Series; McGraw Hill: New York, NY, USA, 2001; ISBN 9780070393622. [Google Scholar]
- Zhang, Q.-W.; Lin, L.-G.; Ye, W.-C. Techniques for Extraction and Isolation of Natural Products: A Comprehensive Review. Chin. Med. 2018, 13, 20. [Google Scholar] [CrossRef]
- Pirzada, M.; Altintas, Z. Template Removal in Molecular Imprinting: Principles, Strategies, and Challenges. In Molecular Imprinting for Nanosensors and Other Sensing Applications; Elsevier: Amsterdam, The Netherlands, 2021; pp. 367–406. [Google Scholar]
- Carreira-Casais, A.; Otero, P.; Garcia-Perez, P.; Garcia-Oliveira, P.; Pereira, A.G.; Carpena, M.; Soria-Lopez, A.; Simal-Gandara, J.; Prieto, M.A. Benefits and Drawbacks of Ultrasound-Assisted Extraction for the Recovery of Bioactive Compounds from Marine Algae. Int. J. Env. Res. Public. Health 2021, 18, 9153. [Google Scholar] [CrossRef] [PubMed]
- Mehta, N.; Jeyapriya, S.; Kumar, P.; Verma, A.K.; Umaraw, P.; Khatkar, S.K.; Khatkar, A.B.; Pathak, D.; Kaka, U.; Sazili, A.Q. Ultrasound-Assisted Extraction and the Encapsulation of Bioactive Components for Food Applications. Foods 2022, 11, 2973. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.-F.; Yang, X.-H.; Wang, Y. Microwave Assisted Extraction of Secondary Metabolites from Plants: Current Status and Future Directions. Trends Food Sci. Technol. 2011, 22, 672–688. [Google Scholar] [CrossRef]
- Perez-Vazquez, A.; Carpena, M.; Barciela, P.; Cassani, L.; Simal-Gandara, J.; Prieto, M.A. Pressurized Liquid Extraction for the Recovery of Bioactive Compounds from Seaweeds for Food Industry Application: A Review. Antioxidants 2023, 12, 612. [Google Scholar] [CrossRef] [PubMed]
- Cvetanović, A. Extractions Without Organic Solvents: Advantages and Disadvantages. Chem. Afr. 2019, 2, 343–349. [Google Scholar] [CrossRef]
- Picot-Allain, C.; Mahomoodally, M.F.; Ak, G.; Zengin, G. Conventional versus Green Extraction Techniques—A Comparative Perspective. Curr. Opin. Food Sci. 2021, 40, 144–156. [Google Scholar] [CrossRef]
- Alzahrani, M.; Perera, C. Ultrasound-Assisted Extraction of Bioactive Compounds from Microalgae. In Handbook of Algal Technologies and Phytochemicals: Volume I: Food, Health and Nutraceutical Applications; Ravishankar, G., Rao Ambati, R., Eds.; CRC Press: Boca Raton, FL, USA, 2019; ISBN 9780429054242. [Google Scholar]
- Albarri, R.; Şahin, S. Kinetics, Thermodynamics, and Mass Transfer Mechanism of the Ultrasound-Assisted Extraction of Bioactive Molecules from Moringa Oleifera Leaves. Biomass Convers. Biorefinery 2023, 13, 7919–7926. [Google Scholar] [CrossRef]
- Vinatoru, M. An Overview of the Ultrasonically Assisted Extraction of Bioactive Principles from Herbs. Ultrason. Sonochem. 2001, 8, 303–313. [Google Scholar] [CrossRef]
- Vinatoru, M.; Toma, M.; Radu, O.; Filip, P.I.; Lazurca, D.; Mason, T.J. The Use of Ultrasound for the Extraction of Bioactive Principles from Plant Materials. Ultrason. Sonochem. 1997, 4, 135–139. [Google Scholar] [CrossRef]
- Yusoff, I.M.; Mat Taher, Z.; Rahmat, Z.; Chua, L.S. A Review of Ultrasound-Assisted Extraction for Plant Bioactive Compounds: Phenolics, Flavonoids, Thymols, Saponins and Proteins. Food Res. Int. 2022, 157, 111268. [Google Scholar] [CrossRef]
- Delgado-Povedano, M.M.; Luque de Castro, M.D. Ultrasound-Assisted Extraction and in Situ Derivatization. J. Chromatogr. A 2013, 1296, 226–234. [Google Scholar] [CrossRef] [PubMed]
- Briars, R.; Paniwnyk, L. Effect of Ultrasound on the Extraction of Artemisinin from Artemisia Annua. Ind. Crops Prod. 2013, 42, 595–600. [Google Scholar] [CrossRef]
- Leal-Ramos, M.Y.; Alarcon-Rojo, A.D.; Mason, T.J.; Paniwnyk, L.; Alarjah, M. Ultrasound-Enhanced Mass Transfer in Halal Compared with Non-Halal Chicken. J. Sci. Food Agric. 2011, 91, 130–133. [Google Scholar] [CrossRef] [PubMed]
- Chemat, F.; Vian, M.A.; Cravotto, G. Green Extraction of Natural Products: Concept and Principles. Int. J. Mol. Sci. 2012, 13, 8615–8627. [Google Scholar] [CrossRef] [PubMed]
- Osman, W.; Badri, N.; Chong, A.; Mohamad, M.; Sulaiman, A. Comparison of Ultrasound Assisted Extraction and Conventional Extraction Technique on Recovery of Phenolic and Flavonoid Compounds from Aloe barbadensis Miller. IOP Conf. Ser. Earth Env. Sci. 2021, 765, 012073. [Google Scholar] [CrossRef]
- Saifullah, M.; McCullum, R.; McCluskey, A.; Vuong, Q. Comparison of Conventional Extraction Technique with Ultrasound Assisted Extraction on Recovery of Phenolic Compounds from Lemon Scented Tea Tree (Leptospermum petersonii) Leaves. Heliyon 2020, 6, e03666. [Google Scholar] [CrossRef] [PubMed]
- Kumar, K.; Srivastav, S.; Sharanagat, V.S. Ultrasound Assisted Extraction (UAE) of Bioactive Compounds from Fruit and Vegetable Processing by-Products: A Review. Ultrason. Sonochem. 2021, 70, 105325. [Google Scholar] [CrossRef] [PubMed]
- Panagiotopoulou, M.; Papadaki, S.; Missirli, T.; Thanassoulia, I.; Krokida, M. Exploring the Valorisation Potential of Tomato Cultivation By-Products in the Frame of Circular Economy. Waste Biomass Valorization 2022, 13, 3957–3972. [Google Scholar] [CrossRef]
- Lavilla, I.; Bendicho, C. Fundamentals of Ultrasound-Assisted Extraction. In Water Extraction of Bioactive Compounds; Elsevier: Amsterdam, The Netherlands, 2017; pp. 291–316. [Google Scholar]
- Hamid Nour, A.; Ruth Oluwaseun, A.; Hamid Nour, A.; Suliman Omer, M.; Ahmed, N. Microwave-Assisted Extraction of Bioactive Compounds (Review). In Microwave Heating—Electromagnetic Fields Causing Thermal and Non-Thermal Effects; IntechOpen: Orlando, FL, USA, 2021. [Google Scholar]
- Bagade, S.B.; Patil, M. Recent Advances in Microwave Assisted Extraction of Bioactive Compounds from Complex Herbal Samples: A Review. Crit. Rev. Anal. Chem. 2021, 51, 138–149. [Google Scholar] [CrossRef]
- Torabi, P.; Hamdami, N.; Keramat, J. Microwave-Assisted Extraction of Sodium Alginate from Brown Macroalgae Nizimuddinia zanardini, Optimization and Physicochemical Properties. Sep. Sci. Technol. 2022, 57, 872–885. [Google Scholar] [CrossRef]
- Routray, W.; Orsat, V. Microwave-Assisted Extraction of Flavonoids: A Review. Food Bioproc. Tech. 2012, 5, 409–424. [Google Scholar] [CrossRef]
- Jain, T.; Jain, V.; Pandey, R.; Vyas, A.; Gandhi, S.; Shukla, S. Microwave Assisted Extraction for Phytoconstituents—An Overview. Asian J. Res. Chem. 2009, 2, 19–25. [Google Scholar]
- Li, Y.; Fabiano-Tixier, A.-S.; Abert-Vian, M.; Chemat, F. Microwave-Assisted Extraction of Antioxidants and Food Colors. In Microwave-Assisted Extraction for Bioactive Compounds: Theory and Practice; Springer: Berlin/Heidelberg, Germany, 2012; pp. 103–125. [Google Scholar]
- Alvarez-Rivera, G.; Bueno, M.; Ballesteros-Vivas, D.; Mendiola, J.A.; Ibañez, E. Pressurized Liquid Extraction. In Liquid-Phase Extraction; Elsevier: Amsterdam, The Netherlands, 2020; pp. 375–398. [Google Scholar]
- Picó, Y. Recent Advances in Sample Preparation for Pesticide Analysis. In Comprehensive Sampling and Sample Preparation; Elsevier: Amsterdam, The Netherlands, 2012; pp. 569–590. [Google Scholar]
- Papadaki, S.; Kyriakopoulou, K.; Krokida, M. Recovery and Encapsualtion of Bioactive Extracts from Haematococcus pluvialis and Phaedodactylum tricornutum for Food Applications. IOSR J. Environ. Sci. Toxicol. Food Technol. (IOSR-JESTFT) 2016, 10, 2319–2399. [Google Scholar] [CrossRef]
- Nootem, J.; Chalorak, P.; Meemon, K.; Mingvanish, W.; Pratumyot, K.; Ruckthong, L.; Srisuwannaket, C.; Niamnont, N. Electrospun Cellulose Acetate Doped with Astaxanthin Derivatives from Haematococcus pluvialis for In Vivo Anti-Aging Activity. RSC Adv. 2018, 8, 37151–37158. [Google Scholar] [CrossRef] [PubMed]
- Schmatz, D.A.; da Silveira Mastrantonio, D.J.; Vieira Costa, J.A.; de Morais, M.G. Encapsulation of Phycocyanin by Electrospraying: A Promising Approach for the Protection of Sensitive Compounds. Food Bioprod. Process. 2020, 119, 206–215. [Google Scholar] [CrossRef]
- Singh, H.; Kumar, C.; Singh, N.; Paul, S.; Jain, S.K. Nanoencapsulation of Docosahexaenoic Acid (DHA) Using a Combination of Food Grade Polymeric Wall Materials and Its Application for Improvement in Bioavailability and Oxidative Stability. Food Funct. 2018, 9, 2213–2227. [Google Scholar] [CrossRef] [PubMed]
- Chu, W.-L. Biotechnological Applications of Microalgae. Int. e-J. Sci. Med. Educ. 2012, 6, S24–S37. [Google Scholar] [CrossRef]
- Ataç Mogol, B.; Gökmen, V.; Shimoni, E. Nano-Encapsulation Improves Thermal Stability of Bioactive Compounds Omega Fatty Acids and Silymarin in Bread. Agro Food Ind. Hi Tech. 2013, 24, 62–65. [Google Scholar]
- Timilsena, Y.P.; Wang, B.; Adhikari, R.; Adhikari, B. Advances in Microencapsulation of Polyunsaturated Fatty Acids (PUFAs)-Rich Plant Oils Using Complex Coacervation: A Review. Food Hydrocoll. 2017, 69, 369–381. [Google Scholar] [CrossRef]
- Amin, A.A.; Salama, M.F.; Seliem, E.I.; Mahmoud, K.F. Encapsulation of Nano Carotenoids; Evaluation of Stability and Safety. Int. J. Curr. Microbiol. Appl. Sci. 2015, 4, 1017–1029. [Google Scholar]
- Marlina, D.; Novita, M.; Karwur, F.F.; Rodonuwu, F.S. Recent Development of Carotenoids Encapsulation Technology. In Proceedings of the International Conference on Research: Implementation and Education of Mathematics and Sciences, Yogyakarta, Indonesia, 18–20 May 2014; pp. 18–20. [Google Scholar]
- OROSA, M. Analysis and Enhancement of Astaxanthin Accumulation in Haematococcus pluvialis. Bioresour. Technol. 2005, 96, 373–378. [Google Scholar] [CrossRef]
- Kovač, D.J.; Simeunović, J.B.; Babić, O.B.; Mišan, A.Č.; Milovanović, I.L. Algae in Food and Feed. Food Feed. Res. 2013, 40, 21–32. [Google Scholar]
- Kerby, N.W.; Raven, J.A. Transport and Fixation of Inorganic Carbon by Marine Algae. In Advances in Botanical Research; Callow, J.A., Woolhouse, H.W., Eds.; Academic Press: Cambridge, MA, USA, 1985; Volume 11, pp. 71–123. [Google Scholar]
- Panagiotopoulou, M.; Papadaki, S.; Krokida, M. Formation and Characterization of Zein Electrosprayed Nanoparticles Containing Bioactive Compounds. S. Afr. J. Chem. Eng. 2022, 40, 32–47. [Google Scholar] [CrossRef]
- Suganya, V.; Anuradha, V. Microencapsulation and Nanoencapsulation: A Review. Int. J. Pharm. Clin. Res. 2017, 9, 233–239. [Google Scholar] [CrossRef]
- Onwulata, C.I. Microencapsulation and Functional Bioactive Foods. J. Food Process Preserv. 2013, 37, 510–532. [Google Scholar] [CrossRef]
- Nakilcioglu-Taş, E.; Otleş, S. Degradation Kinetics of Bioactive Compounds and Antioxidant Capacity of Brussels Sprouts during Microwave Processing. Int. J. Food Prop. 2017, 20, S2798–S2809. [Google Scholar] [CrossRef]
- Pérez-Masiá, R.; López-Nicolás, R.; Periago, M.J.; Ros, G.; Lagaron, J.M.; López-Rubio, A. Encapsulation of Folic Acid in Food Hydrocolloids through Nanospray Drying and Electrospraying for Nutraceutical Applications. Food Chem. 2015, 168, 124–133. [Google Scholar] [CrossRef] [PubMed]
- Soukoulis, C.; Bohn, T. A Comprehensive Overview on the Micro- and Nano-Technological Encapsulation Advances for Enhancing the Chemical Stability and Bioavailability of Carotenoids. Crit. Rev. Food Sci. Nutr. 2018, 58, 971353. [Google Scholar] [CrossRef] [PubMed]
- Jaworek, A.; Sobczyk, A.T. Electrospraying Route to Nanotechnology: An Overview. J. Electrostat. 2008, 66, 197–219. [Google Scholar] [CrossRef]
- Ayala-Fuentes, J.C.; Chavez-Santoscoy, R.A. Nanotechnology as a Key to Enhance the Benefits and Improve the Bioavailability of Flavonoids in the Food Industry. Foods 2021, 10, 2701. [Google Scholar] [CrossRef]
- Fathi, M.; Martín, Á.; McClements, D.J. Nanoencapsulation of Food Ingredients Using Carbohydrate Based Delivery Systems. Trends Food Sci. Technol. 2014, 39, 18–39. [Google Scholar] [CrossRef]
- Ezhilarasi, P.N.; Karthik, P.; Chhanwal, N.; Anandharamakrishnan, C. Nanoencapsulation Techniques for Food Bioactive Components: A Review. Food Bioprocess Technol. 2013, 6, 628–647. [Google Scholar] [CrossRef]
- Sozer, N.; Kokini, J.L. Nanotechnology and Its Applications in the Food Sector. Trends Biotechnol. 2009, 27, 82–89. [Google Scholar] [CrossRef] [PubMed]
- Pisoschi, A.M.; Pop, A.; Cimpeanu, C.; Turcuş, V.; Predoi, G.; Iordache, F. Nanoencapsulation Techniques for Compounds and Products with Antioxidant and Antimicrobial Activity—A Critical View. Eur. J. Med. Chem. 2018, 157, 1326–1345. [Google Scholar] [CrossRef] [PubMed]
- Casanova, F.; Santos, L. Encapsulation of Cosmetic Active Ingredients for Topical Application—A Review. J. Microencapsul. 2016, 33, 1115900. [Google Scholar] [CrossRef] [PubMed]
- Kikionis, S.; Ioannou, E.; Toskas, G.; Roussis, V. Electrospun Biocomposite Nanofibers of Ulvan/PCL and Ulvan/PEO. J. Appl. Polym. Sci. 2015, 132, 42153. [Google Scholar] [CrossRef]
- El-Aassar, M.R.; Ibrahim, O.M.; Al-Oanzi, Z.H. Biotechnological Applications of Polymeric Nanofiber Platforms Loaded with Diverse Bioactive Materials. Polymers 2021, 13, 3734. [Google Scholar] [CrossRef] [PubMed]
- Bhardwaj, N.; Kundu, S.C. Electrospinning: A Fascinating Fiber Fabrication Technique. Biotechnol. Adv. 2010, 28, 325–347. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, S.; Wendorff, J.H.; Greiner, A. Use of Electrospinning Technique for Biomedical Applications. Polymers 2008, 49, 5603–5621. [Google Scholar] [CrossRef]
- Drosou, C.G.; Krokida, M.K.; Biliaderis, C.G. Encapsulation of Bioactive Compounds through Electrospinning/Electrospraying and Spray Drying: A Comparative Assessment of Food-Related Applications. Dry. Technol. 2017, 35, 139–162. [Google Scholar] [CrossRef]
- Ahmadian, A.; Shafiee, A.; Aliahmad, N.; Agarwal, M. Overview of Nano-Fiber Mats Fabrication via Electrospinning and Morphology Analysis. Textiles 2021, 1, 206–226. [Google Scholar] [CrossRef]
- Drosou, C.; Krokida, M.; Biliaderis, C.G. Composite Pullulan-Whey Protein Nanofibers Made by Electrospinning: Impact of Process Parameters on Fiber Morphology and Physical Properties. Food Hydrocoll. 2018, 77, 726–735. [Google Scholar] [CrossRef]
- Xue, J.; Wu, T.; Dai, Y.; Xia, Y. Electrospinning and Electrospun Nanofibers: Methods, Materials, and Applications. Chem. Rev. 2019, 119, 5298–5415. [Google Scholar] [CrossRef] [PubMed]
- Wen, X.; Xiong, J.; Lei, S.; Wang, L.; Qin, X. Diameter Refinement of Electrospun Nanofibers: From Mechanism, Strategies to Applications. Adv. Fiber Mater. 2022, 4, 145–161. [Google Scholar] [CrossRef]
- Li, Z.; Wang, C. Effects of Working Parameters on Electrospinning. In One-Dimensional Nanostructures: Electrospinning Technique and Unique Nanofibers; Li, Z., Wang, C., Eds.; Springer: Berlin/Heidelberg, Germany, 2013; pp. 15–28. ISBN 978-3-642-36427-3. [Google Scholar]
- Rajam, R.; Anandharamakrishnan, C. Spray Freeze Drying Method for Microencapsulation of Lactobacillus plantarum. J. Food Eng. 2015, 166, 95–103. [Google Scholar] [CrossRef]
- Ishwarya, S.P.; Anandharamakrishnan, C.; Stapley, A.G.F. Spray-Freeze-Drying: A Novel Process for the Drying of Foods and Bioproducts. Trends Food Sci. Technol. 2015, 41, 161–181. [Google Scholar] [CrossRef]
- Adali, M.B.; Barresi, A.A.; Boccardo, G.; Pisano, R. Spray Freeze-Drying as a Solution to Continuous Manufacturing of Pharmaceutical Products in Bulk. Processes 2020, 8, 709. [Google Scholar] [CrossRef]
- Zhang, F.; Ma, X.; Wu, X.; Xu, Q.; Tian, W.; Li, Z. Inert Particles as Process Aid in Spray-Freeze Drying. Dry. Technol. 2020, 38, 71–79. [Google Scholar] [CrossRef]
- Laurin, L. Overview of LCA-History, Concept, and Methodology. In Encyclopedia of Sustainable Technologies; Elsevier: Amsterdam, The Netherlands, 2017; pp. 217–222. ISBN 9780128046777. [Google Scholar]
- Finnveden, G.; Potting, J. Life Cycle Assessment. In Encyclopedia of Toxicology, 3rd ed.; Elsevier: Amsterdam, The Netherlands, 2014; pp. 74–77. ISBN 9780123864543. [Google Scholar]
- Guinée, J.; Heijungs, R. Introduction to Life Cycle Assessment. In Sustainable Supply Chains: A Research-Based Textbook on Operations and Strategy; Springer: Berlin/Heidelberg, Germany, 2017; pp. 15–41. [Google Scholar]
- Clift, R.; Druckman, A. (Eds.) Taking Stock of Industrial Ecology; Springer International Publishing: Cham, Switzerland, 2016; ISBN 978-3-319-20570-0. [Google Scholar]
- Hauschild, M.Z.; Rosenbaum, R.K.; Olsen, S.I. (Eds.) Life Cycle Assessment: Theory and Practice; Springer International Publishing: Cham, Switzerland, 2018; ISBN 978-3-319-56474-6. [Google Scholar]
- Heijungs, R.; Guineév, J.B. An Overview of the Life Cycle Assessment Method-Past, Present, and Future. In Life Cycle Assessment Handbook: A Guide for Environmentally Sustainable Products; John Wiley and Sons: Hoboken, NJ, USA, 2012; pp. 15–41. ISBN 9781118099728. [Google Scholar]
- Yang, S.; Ma, K.; Liu, Z.; Ren, J.; Man, Y. Development and Applicability of Life Cycle Impact Assessment Methodologies. In Life Cycle Sustainability Assessment for Decision-Making: Methodologies and Case Studies; Elsevier: Amsterdam, The Netherlands, 2019; pp. 95–124. ISBN 9780128183557. [Google Scholar]
- ISO 14040:2006; Environmental Management—Life Cycle Assessment—Principles and Framework. International Organization for Standardization: Geneva, Switzerland, July 2006; pp. 1–20.
- Kyriakopoulou, K.; Papadaki, S.; Krokida, M. Life Cycle Analysis of β-Carotene Extraction Techniques. J. Food Eng. 2015, 167, 51–58. [Google Scholar] [CrossRef]
- Espada, J.J.; Pérez-Antolín, D.; Vicente, G.; Bautista, L.F.; Morales, V.; Rodríguez, R. Environmental and Techno-Economic Evaluation of β-Carotene Production from Dunaliella salina. A Biorefinery Approach. Biofuels Bioprod. Biorefining 2020, 14, 43–54. [Google Scholar] [CrossRef]
- Wang, T.H.; Hsu, C.L.; Huang, C.H.; Hsieh, Y.K.; Tan, C.S.; Wang, C.F. Environmental Impact of CO2-Expanded Fluid Extraction Technique in Microalgae Oil Acquisition. J. Clean. Prod. 2016, 137, 813–820. [Google Scholar] [CrossRef]
- Davis, D.; Morão, A.; Johnson, J.K.; Shen, L. Life Cycle Assessment of Heterotrophic Algae Omega-3. Algal Res. 2021, 60, 102494. [Google Scholar] [CrossRef]
- Qin, Z.H.; Hu, X.; Mou, J.H.; He, G.H.; Ye, G.B.; Li, H.Y.; Chopra, S.S.; Dong, L.; Lin, C.S.K.; Wang, X. Environmental Profiling Microalgae-Based Eicosapentaenoic Acid Production along the Technical Advancement via Life Cycle Assessment. J. Clean. Prod. 2023, 397, 136477. [Google Scholar] [CrossRef]
- Pérez-López, P.; González-García, S.; Allewaert, C.; Verween, A.; Murray, P.; Feijoo, G.; Moreira, M.T. Environmental Evaluation of Eicosapentaenoic Acid Production by Phaeodactylum tricornutum. Sci. Total Environ. 2014, 466–467, 991–1002. [Google Scholar] [CrossRef] [PubMed]
- Papadaki, S.G.; Kyriakopoulou, K.E.; Krokida, M.K. Life Cycle Analysis of Microalgae Extraction Techniques. Chem. Eng. Trans. 2016, 52, 1039–1044. [Google Scholar] [CrossRef]
- Aldaghi, S.A.; Ubais, R.; Schmitt, I.; Wendisch, V.F.; Costamagna, M.; Perucca, M. Life Cycle Assessment of Bacterial, Algal, and Synthetic Approaches for Astaxanthin Production at a Laboratory Scale: Comparative Environmental Analysis and Sensitivity of Energy Sources. Processes 2023, 11, 2911. [Google Scholar] [CrossRef]
- Zlaugotne, B.; Diaz Sanchez, F.A.; Pubule, J.; Blumberga, D. Life Cycle Impact Assessment of Microalgae and Synthetic Astaxanthin Pigments. Environ. Clim. Technol. 2023, 27, 233–242. [Google Scholar] [CrossRef]
- Papadaki, S.; Kyriakopoulou, K.; Tzovenis, I.; Krokida, M. Environmental Impact of Phycocyanin Recovery from Spirulina platensis Cyanobacterium. Innov. Food Sci. Emerg. Technol. 2017, 44, 217–223. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bioactive Pigment | Microalgae Strain | Carotenoid Content | Reference |
|---|---|---|---|
| Astaxanthin | Heamatococcus pluvialis | 7% dw * | [30] |
| 81.38% TC ** | [31] | ||
| Chromochloris zofingiensis | 1,5 mg/g dw | [32] | |
| Lutein | Spirulina platensis | 2556 μg/g dw | [33] |
| Chlorella pyrenoidosa | 125,034.4 μg/g (trans) 27,975.3 μg/g (cis) | [34] | |
| Botryococcus braunii | 74.6% TC | [31] | |
| Chlorella zofingiensis |
4000 µg/g
13.81 ± 1.23 mg/g | [32] [35] | |
| β-carotene | Spirulina platensis | 12,510 μg/g dw | [33] |
| 69.5% TC | [31] | ||
| Spirulina maxima | 80% TC | [30] | |
| Dunaliella salina | 138.25 mg/g (all-trans) 124.65 mg/g (9-cis) 10–13% dw | [30,36] | |
| Chlorella zofingiensis | 0.9–3.7% dw | [30] | |
| Zeaxanthin | Spirulina platensis | 558 μg/g dw | [33] |
| Porphyridium cruentum | 97.4% TC | [30] | |
| Chlorella pyrenoidosa | 2170.3 µg/g | [34] | |
| Chlorella zofingiensis | 7000 ± 820 µg/g | [35] |
| Microalgae | Protein Content (% Dry Matter) | Reference |
|---|---|---|
| Spirulina (Arthrospira) platensis | 46–63 | [87] |
| Spirulina (Arthrospira) maxima | 60–71 | [87] |
| Arthrospira fusiformis | 39–56 | [88] |
| Chlorella vulgaris | 51–58 | [87] |
| Chlorella pyrenoidosa | 57 | [87] |
| Nannochloropsis oculata | 35 | [89] |
| Isochrysis galbana | 27–29 | [89,90] |
| Phaeodactylum tricornutum | 30 | [89] |
| Porphyridium cruentum | 28–39 | [87] |
| Dunaliella salina | 57 | [87] |
| Amino Acid | SP | SM | CV | CP | NO | PC | DS | WHO/FAO Requirement for Adults (mg/kg/day) |
|---|---|---|---|---|---|---|---|---|
| Alanine | 9.5 | 6.8 | 7.9 | 5.08 ± 0.19 | 10.92 ± 0.01 | 6.67 ± 3.67 | 10.99 ± 0.32 | - |
| Arginine | 7.3 | 6.5 | 6.4 | 5.91 ± 0.07 | 5.93 ± 0.02 | 7.78 ± 0.29 | 8.16 ± 0.24 | - |
| Aspartic acid | 11.8 | 8.6 | 9.0 | 8.12 ± 0.16 | 9.14 ± 0.05 | 11.21 ± 0.45 | 9.56 ± 0.28 | - |
| Cysteine | 0.9 | 0.4 | 1.4 | 2.82 ± 0.06 | 0.19 ± 0.01 | 0.33 ± 0.01 | 1.63 ± 0.04 | - |
| Glutamic acid | 10.3 | 12.6 | 11.6 | 7.87 ± 0.23 | 10.30 ± 0.02 | 8.17 ± 0.29 | 12.41 ± 0.37 | - |
| Glycine | 5.7 | 4.8 | 5.8 | 9.73 ± 0.42 | 9.00 ± 0.01 | 6.86 ± 0.28 | 8.71 ± 0.26 | - |
| Histidine * | 2.2 | 1.8 | 2.0 | 1.64 ± 0.01 | 0.94 ± 0.01 | 1.11 ± 0.04 | 1.73 ± 0.05 | 10 |
| Isoleucine * | 6.7 | 6.0 | 3.8 | 6.20 ± 0.14 | 0.11 ± 0.01 | 5.25 ± 0.24 | 4.09 ± 0.12 | 20 |
| Leucine * | 9.8 | 8.0 | 8.8 | 3.44 ± 0.06 | 8.11 ± 0.05 | 5.83 ± 0.21 | 9.58 ± 0.28 | 39 |
| Lysine * | 4.8 | 4.6 | 8.4 | 8.14 ± 0.37 | 5.70 ± 0.01 | 5.50 ± 0.21 | 5.99 ± 0.17 | 30 |
| Methionine * | 2.5 | 1.4 | 2.2 | 3.30 ± 0.02 | 1.50 ± 0.01 | 2.78 ± 0.11 | 2.79 ± 0.08 | - |
| Phenylalanine * | 5.3 | 4.9 | 5.0 | 3.83 ± 0.11 | 5.05 ± 0.01 | 5.00 ± 0.20 | 6.98 ± 0.20 | - |
| Proline | 4.2 | 3.9 | 4.8 | - | 4.20 ± 0.07 | 2.53 ± 0.17 | 5.23 ± 0.15 | - |
| Serine | 5.1 | 4.2 | 4.1 | 2.79 ± 0.03 | 6.52 ± 0.01 | 8.11 ± 0.29 | 4.81 ± 0.14 | - |
| Threonine * | 6.2 | 4.6 | 4.8 | 3.45 ± 0.04 | 5.91 ± 0.03 | 6.25 ± 0.25 | 5.16 ± 0.15 | 15 |
| Tryptophan * | 0.3 | 1.4 | 2.1 | - | 1.24 ± 0.01 | 1.39 ± 0.05 | 0.18 ± 0.01 | - |
| Tyrosine | 5.3 | 3.9 | 3.4 | 1.22 ± 0.01 | 3.40 ± 0.02 | 4.43 ± 0.18 | 4.86 ± 0.14 | - |
| Valine * | 7.1 | 6.5 | 5.5 | 5.17 ± 0.05 | 3.29 ± 0.02 | 2.50 ± 0.10 | 7.23 ± 0.21 | - |
| Strain | PC Concentration | Inhibition Zone (mm) (Mean ± STD) | MIC * | Formulation |
|---|---|---|---|---|
| S. vitulinus | 100 μg/mL | 0.33 ± 0.11 | - | Cosmetic cream |
| 300 μg/mL | 0.28 ± 0.10 | - | ||
| 500 μg/mL | 0.39 ± 0.13 | - | ||
| 1000 μg/mL | 0.57 ± 0.05 | - | ||
| S. aureus | 100 μg/mL | 0.48 ± 0.18 | - | Cosmetic cream |
| 300 μg/mL | 0.85 ± 0.07 | - | ||
| 500 μg/mL | 0.90 ± 0.08 | - | ||
| 1000 μg/mL | 1.21 ± 0.01 | - | ||
| 100 μg/ disc | 9.33 ± 0.33 | 125 μg/mL | C-PC discs | |
| E. coli | 100 μg/mL | 0.53 ± 0.11 | - | Cosmetic cream |
| 300 μg/mL | 0.60 ± 0.07 | - | ||
| 500 μg/mL | 0.63 ± 0.04 | - | ||
| 1000 μg/mL | 1.13 ± 0.32 | - | ||
| 100 μg/ disc | 13.33 ± 0.67 | 100 μg/mL | C-PC discs | |
| P. acne | 10 g C-PC/100 g oleaginous base | 23.4 ± 1.0 | 1.6 ± 0.4 mg/mL | Anti-acne ointment |
| 10 g C-PC/100 g water-soluble base | 26.1 ± 1.2 | 1.5 ± 0.1 mg/mL | ||
| S. epidermidis | 10 g C-PC/100 g oleaginous base | 21.3 ± 1.4 | 2.1 ± 0.6 mg/mL | Anti-acne ointment |
| 10 g C-PC/100 g water-soluble base | 24.6 ± 1.6 | 1.8 ± 0.2 mg/mL | ||
| P. aeruginosa | 100 μg/disc | 18.00 ± 0.58 | 50 μg/mL | C-PC discs |
| K. pneumoniae | 100 μg/disc | 16.00 ± 0.58 | 75 μg/mL | C-PC discs |
| AA Sequence | Microalgal Source | IC50 | Reference |
|---|---|---|---|
| Leu-Asn-Gly-Asp-Val-Trp | Chlorella ellipsoidea | 0.2 mM (PR) | |
| 0.92 mM (DPPH) | [136] | ||
| 1.42 mM (HR) | |||
| Val-Glu-Cys-Tyr-Gly-Pro-Asn-Arg-Pro-Gln-Phe | Chlorella vulgaris algal waste | 9.8 ± 0.5 μΜ (ABTS) | |
| 8.3 ± 0.15 μΜ (HR) | [137] | ||
| 7.5 ± 0.12 μM (SR) | |||
| Asn-Asp-Ala-Glu-Tyr-Gly-Ile-Cys-Gly-Phe | Isochrysis zhanjiangensis | Antioxidant activity against DPPH, HR, SR, AR | [138] |
| Phe-Ser-Glu-Ser-Ser-Ala-Pro-Glu-Gln-His-Tyr | Arthrospira platensis | 171.47 μg/mL (DPPH) | [139] |
| Microalgae | Lipid Content (% dw) | PUFAs Content (% Total FAs) | EPA Content (% Total FAs) | DHA Content (% Total FAs) | References |
|---|---|---|---|---|---|
| Schizochytrium sp. | 46–78 | 46.96 | 0.72 | 37.63 | [164,165] |
| Phaeodactylum tricornutum | 14.9 | 40.04 | 30.57 | 0.22 | [166,167] |
| Porphyridium cruentum | 5.3 | 70.05 | 29.47 | - | [166,167] |
| Isochrysis galbana | 28.68 | 51.62 | 1.03 | 20.44 | [166,168] |
| Nannochloropsis oceanica | 51.67 | 38.69 | 23.95 | - | [166,169] |
| Nannochloropsis gaditana | 8.1 | 4.69 | - | - | [167] |
| Nannochloropsis oculata | 15.6 | 36.48 | 31.24 | 0.45 | [167] |
| Pavlova lutheri | 31.36 | 29.38 | 17.76 | 7.61 | [168] |
| Spirulina platensis | 6.2 | 30.17 | 0.19 | - | [170] |
| Chlorella pyrenoidosa | 11.4 | 43.67 | 0.31 | 0.22 | [170] |
| Chlorella vulgaris | 13.3 | 38.30 | - | 0.30 | [170] |
| Dunaliella salina | 24.85 | 31.06 | - | - | [171] |
| Microalgae | Lipid Content (% Dry Mass) | PUFAs Content (% of Total FAs) | ABTS Assay IC50 (μg/mL) | DPPH Assay IC20 (μg/mL) |
|---|---|---|---|---|
| Tetraselmis chui | 6.5 ± 0.4 | 51.8 ± 6.6 | 40.9 ± 4.7 | 225.7 ± 6.9 |
| Chlorella vulgaris | 8.8 ± 0.7 | 64.6 ± 3.5 | 51.1 ± 3.7 | 50.5 ± 12.3 |
| Spirulina sp. | 10.7 ± 1.0 | 44.8 ± 1.5 | 38.7 ± 1.6 | 96.9 ± 9.7 |
| Scendesmus obliquus | 11.1 ± 1.1 | 60.1 ± 4.8 | 29.4 ± 1.2 | 89.1 ± 6.6 |
| Phaeodactylum tricornutum | 12.6 ± 1.2 | 54.3 ± 2.0 | 57.3 ± 4.5 | 75.4 ± 4.6 |
| Chlorococcum amblystomatis | 16.6 ± 0.9 | 56.7 ± 2.6 | 52.6 ± 4.6 | 58.4 ± 10.7 |
| Nannochloropsis oceanica | 20.9 ± 3.4 | 39.7 ± 3.4 | 101.9 ± 1.7 | 175.6 ± 8.7 |
| Extraction Methods | Advantages | Disadvantages |
|---|---|---|
| Soxhlet extraction | Simple and inexpensive. | High solvent consumption; Labor and time intensive; Difficulty with automation and scaling-up. |
| Ultrasound-assisted extraction | Energy, solvent, and time savings; Extract quality preservation; Suitable for thermosensitive compounds. | Need for optimization to avoid overstimulation of treated materials; Filtration or centrifuge step needed. |
| Microwave-assisted extraction | Simple and low cost; Higher selectivity; Reduced extraction time. | Possible chemical structure modification of bioactive target compounds due to intense sudden heating; Filtration or centrifuge step needed. |
| Pressurized liquid extraction | Extract fractionation by pressure alterations; Solvent reduction; Lower extraction time. | High equipment cost. |
| Supercritical fluid extraction (CO2) | Exploitation of gas and liquid physicochemical properties as solvents; Time effective. | High equipment cost and capital investment; Poor selectivity towards polar compounds due to low polarity of supercritical CO2. |
| Enzyme-assisted extraction | High selectivity; Improved yield. | Expensive enzymes. Rigorous control of medium pH and temperature for optimal enzyme action. |
| Benefits from Encapsulation | Encapsulation Methods |
|---|---|
| Solubility increase | Phase separation |
| Storage time extension Covering unwanted odor and taste | Adsorption |
| Preservation of the aroma during storage | Formation of inclusion complexes |
| Protection against microbiological infections | Crystallization |
| Creation of stable and standardized components | Lyophilization/freeze drying |
| Protection of active ingredients from heat and light | Extrusion |
| Controlled release of substances | Electrohydrodynamic process (Electrospinning) |
| Avoiding degradation of thermosensitive compounds | Spray–freeze drying |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Papadaki, S.; Tricha, N.; Panagiotopoulou, M.; Krokida, M. Innovative Bioactive Products with Medicinal Value from Microalgae and Their Overall Process Optimization through the Implementation of Life Cycle Analysis—An Overview. Mar. Drugs 2024, 22, 152. https://doi.org/10.3390/md22040152
Papadaki S, Tricha N, Panagiotopoulou M, Krokida M. Innovative Bioactive Products with Medicinal Value from Microalgae and Their Overall Process Optimization through the Implementation of Life Cycle Analysis—An Overview. Marine Drugs. 2024; 22(4):152. https://doi.org/10.3390/md22040152
Chicago/Turabian StylePapadaki, Sofia, Nikoletta Tricha, Margarita Panagiotopoulou, and Magdalini Krokida. 2024. "Innovative Bioactive Products with Medicinal Value from Microalgae and Their Overall Process Optimization through the Implementation of Life Cycle Analysis—An Overview" Marine Drugs 22, no. 4: 152. https://doi.org/10.3390/md22040152