
Relationship between Carbon Sequestration and Soil Physicochemical Parameters in Northern Campeche, Mexico
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
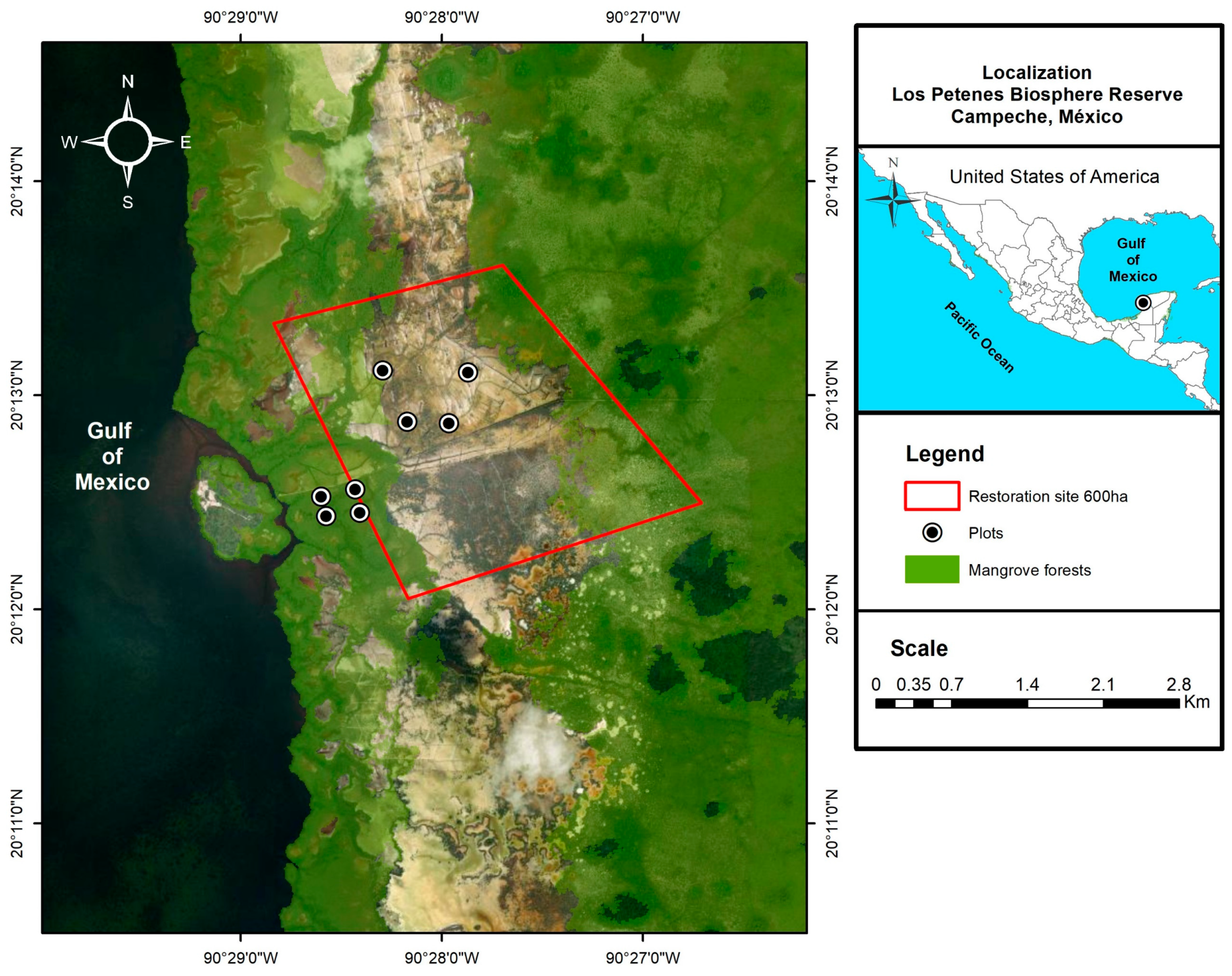
2.1. Study Area
2.2. Determination of pH, Redox Potential, Temperature, and Salinity
2.3. Determination of Bulk Density
2.4. Determination of Soil Carbon
2.5. Quantification of Organic Carbon Sequestration in Soil
2.6. Statistical Analysis
3. Results
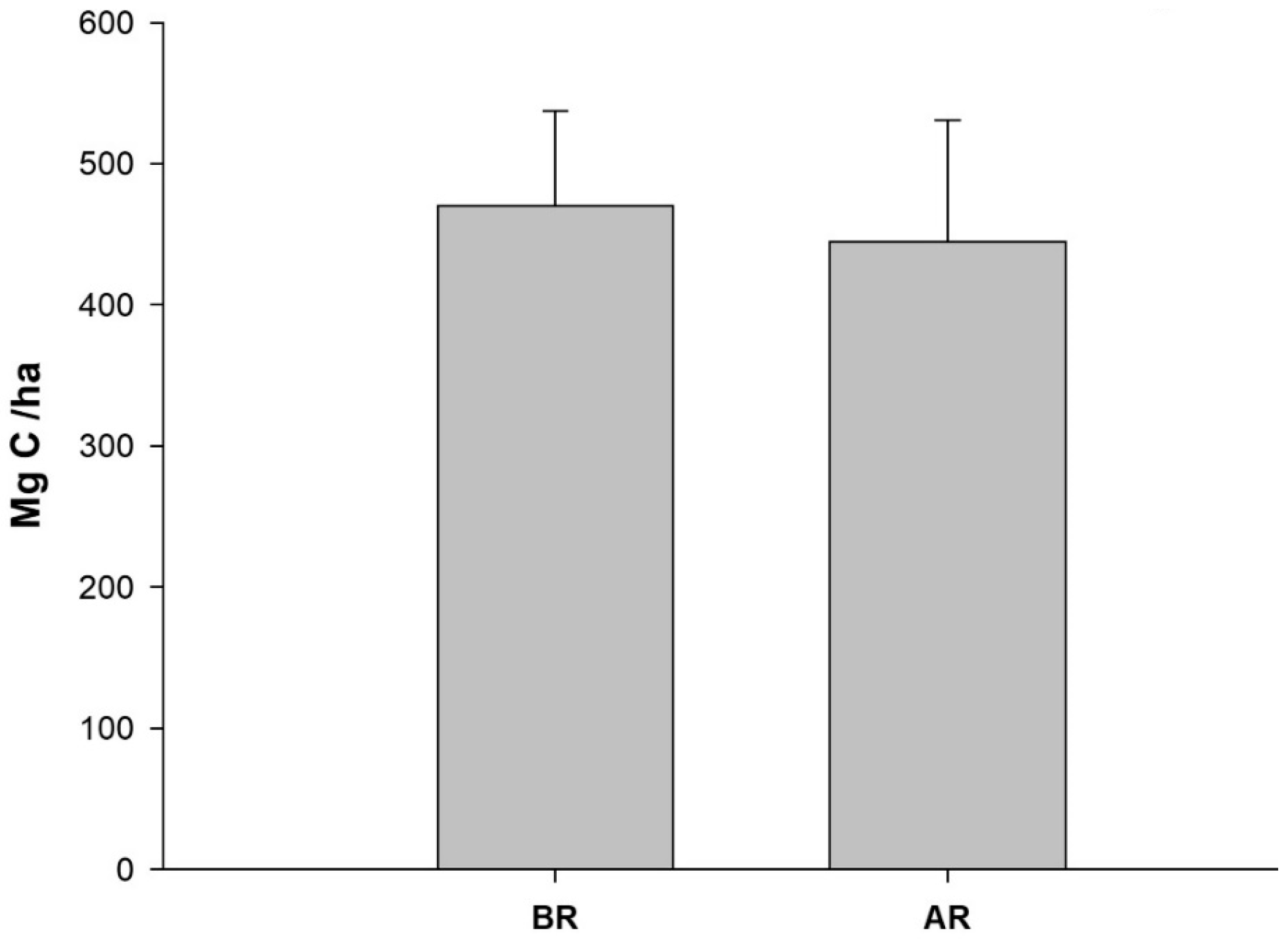
3.1. Organic Carbon Sequestration
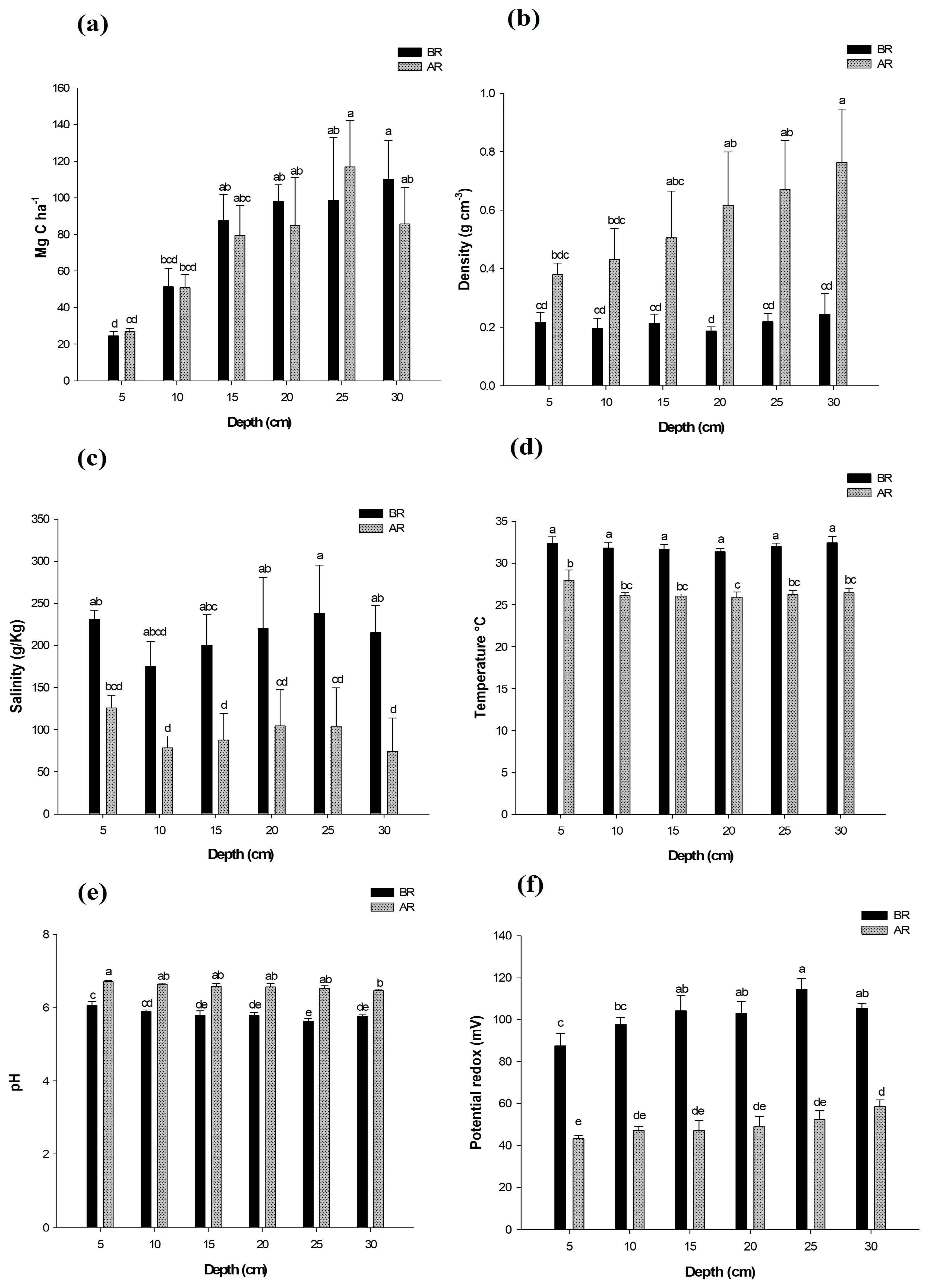
3.2. General Behavior of Soil Physicochemical Parameters
3.3. Relationship between Carbon Sequestration and Soil Physicochemical Parameters
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Agraz-Hernández, C.M.; Chan-Keb, C.A.; Muñiz-Salazar, R.; Pérez-Balan, R.A.; Vanegas, G.P.; Manzanilla, H.G.; Osti-Sáenz, J.; del Río Rodríguez, R. Pore Water Chemical Variability and Its Effect on Phenological Production in Three Mangrove Species under Drought Conditions in Southeastern Mexico. Diversity 2022, 14, 668. [Google Scholar] [CrossRef]
- Collins, M.E.; Kuehl, R. Organic Matter Accumulation and Organic Soils. In Wetlands Soils: Genesis, Hydrology, Landscapes and Classification; Wetlands Soils: Boca Raton, IL, USA, 2000; p. 432. ISBN 978-0-429-13332-9. [Google Scholar]
- Donato, D.C.; Kauffman, J.B.; Murdiyarso, D.; Kurnianto, S.; Stidham, M.; Kanninen, M. Mangroves among the Most Carbon-Rich Forests in the Tropics. Nat. Geosci. 2011, 4, 293–297. [Google Scholar] [CrossRef]
- Troxler, T.G.; Barr, J.G.; Fuentes, J.D.; Engel, V.; Anderson, G.; Sanchez, C.; Lagomasino, D.; Price, R.; Davis, S.E. Component-Specific Dynamics of Riverine Mangrove CO2 Efflux in the Florida Coastal Everglades. Agric. For. Meteorol. 2015, 213, 273–282. [Google Scholar] [CrossRef]
- Alongi, D.M. Present State and Future of the World’s Mangrove Forests. Environ. Conserv. 2002, 29, 331–349. [Google Scholar] [CrossRef]
- Thomas, N.; Lucas, R.; Bunting, P.; Hardy, A.; Rosenqvist, A.; Simard, M. Distribution and Drivers of Global Mangrove Forest Change, 1996–2010. PLoS ONE 2017, 12, e0179302. [Google Scholar] [CrossRef]
- Valderrama-Landeros, L.H.; Rodríguez-Zúñiga, M.T.; Troche-Souza, C.; Velázquez-Salazar, S.; Villeda-Chávez, E.; Alcántara-Maya, J.A. (Eds.) Manglares de México: Actualización y Exploración de los Datos del Sistema de Monitoreo 1970/1980–2015; CONABIO: México City, Mexico, 2017; ISBN 978-607-8328-77-2. [Google Scholar]
- IPCC; Eggleston, H.S.; Miwa, S.; Srivastava, N.; Tanabe, K. 2006 IPCC Guidelines for National Greenhouse Gas Inventories—A Primer, Prepared by the National Greenhouse Gas Inventories Programme; Institute for Global Environmental Strategies (IGES): Hayama, Japan, 2008. [Google Scholar]
- Twilley, R.R.; Castañeda-Moya, E.; Rivera-Monroy, V.H.; Rovai, A. Productivity and Carbon Dynamics in Mangrove Wetlands. In Mangrove Ecosystems: A Global Biogeographic Perspective; Rivera-Monroy, V.H., Lee, S.Y., Kristensen, E., Twilley, R.R., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 113–162. ISBN 978-3-319-62204-0. [Google Scholar]
- Fatoyinbo, T.E.; Simard, M.; Washington-Allen, R.A.; Shugart, H.H. Landscape-scale Extent, Height, Biomass, and Carbon Estimation of Mozambique’s Mangrove Forests with Landsat ETM+ and Shuttle Radar Topography Mission Elevation Data. J. Geophys. Res. 2008, 113, 2007JG000551. [Google Scholar] [CrossRef]
- Kairo, J.G.; Lang’at, J.K.S.; Dahdouh-Guebas, F.; Bosire, J.; Karachi, M. Structural Development and Productivity of Replanted Mangrove Plantations in Kenya. For. Ecol. Manag. 2008, 255, 2670–2677. [Google Scholar] [CrossRef]
- Adame, M.F.; Kauffman, J.B.; Medina, I.; Gamboa, J.N.; Torres, O.; Caamal, J.P.; Reza, M.; Herrera-Silveira, J.A. Carbon Stocks of Tropical Coastal Wetlands within the Karstic Landscape of the Mexican Caribbean. PLoS ONE 2013, 8, e56569. [Google Scholar] [CrossRef]
- Piedra, L.; Sierra, L. Metodología para la Cuantificación de Carbono en Bosques de Manglares. In Métodos para el Estudio de la Biodiversidad en Ecosistemas Marinos Tropicales de Iberoamérica para la Adaptación al Cambio Climático; Hernández-Zanuy, A.C., Alcolado, P.M., Eds.; La Habana: Red CYTED; Instituto de Oceanología: La Habana, Cuba, 2014; ISBN 978-959-298-031-0. [Google Scholar]
- Sitoe, A.; Mandlate, L.; Guedes, B. Biomass and Carbon Stocks of Sofala Bay Mangrove Forests. Forests 2014, 5, 1967–1981. [Google Scholar] [CrossRef]
- Li, T.; Huang, Y.; Zhang, W.; Yu, Y.-Q. Methane Emissions Associated with the Conversion of Marshland to Cropland and Climate Change on the Sanjiang Plain of Northeast China from 1950 to 2100. Biogeosciences 2012, 9, 5199–5215. [Google Scholar] [CrossRef]
- Zinn, Y.L.; Lal, R.; Resck, D.V.S. Changes in Soil Organic Carbon Stocks under Agriculture in Brazil. Soil Tillage Res. 2005, 84, 28–40. [Google Scholar] [CrossRef]
- Lai, R. Soil Carbon Sequestration in Natural and Managed Tropical Forest Ecosystems. J. Sustain. For. 2004, 21, 1–30. [Google Scholar] [CrossRef]
- Masera, O.R.; Cerón, A.D.; Ordóñez, A. Forestry Mitigation Options for Mexico: Finding Synergies between National Sustainable Development Priorities and Global Concerns. Mitig. Adapt. Strateg. Glob. Change 2001, 6, 291–312. [Google Scholar] [CrossRef]
- Or Ordóñez, J.A.B. Cómo entender el manejo forestal, la captura de carbono y el pago por servicios ambientales. Ciencias 2008, 90, 37–42. [Google Scholar]
- Moreno May, G.; Cerón Bretón, J.G.; Cerón Bretón, R.M.; Guerra Santos, J.J.; Amador del Angel, L.E.; Endañú Huerta, E. Estimación Del Potencial de Captura de Carbono en Suelos de Manglar de Isla Del Carmen. UNACAR Tecnociencia 2010, 1, 23–39. [Google Scholar]
- Gutiérrez-Ayala, L.V.; Torrescano-Valle, N.; Islebe, G.A. Reconstrucción paleoambiental del Holoceno tardío de la reserva los Petenes, Península de Yucatán, México. Rev. Mex. Cienc. Geol. 2012, 29, 749–763. [Google Scholar]
- Agraz-Hernández, C.M.; Osti-Sáenz, J.; Expósito-Díaz, G.; Martínez-Reyes, A.A.; Chan-Keb, C.; Martínez-Muñiz, G. Study of the Possible Environmental Impact on the Mangrove Ecosystem Due to the Operation of the Las Cruces Hydroelectric Project; Universidad Autónoma de Campeche/Comisión Federal de Electricidad: Campeche, Mexico, 2015. [Google Scholar]
- Valdés Velarde, E.; Valdez Hernández, J.I.; Ordaz Chaparro, V.M.; Gallardo Lancho, J.F.; Pérez Nieto, J.; Ayala Sánchez, C. Evaluación del carbono orgánico en suelos de los manglares de Nayarit. Rev. Mex. Cienc. For. 2011, 2, 47–58. [Google Scholar]
- Agraz Hernández, C.M.; García Zaragoza, C.; Iriarte-Vivar, S.; Flores-Verdugo, F.J.; Moreno Casasola, P. Forest Structure, Productivity and Species Phenology of Mangroves in the la Mancha Lagoon in the Atlantic Coast of Mexico. Wetl. Ecol. Manag. 2011, 19, 273–293. [Google Scholar] [CrossRef]
- Rodríguez Fuentes, H.R.; Rodríguez Absi, J.R. Métodos de Análisis de Suelos y Plantas; Trillas: Mexico City, Mexico, 2002; ISBN 978-968-24-6596-3. [Google Scholar]
- Bremner, J.M. Nitrogen-Total. In Methods of Soil Analysis; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 1996; pp. 1085–1121. ISBN 978-0-89118-866-7. [Google Scholar]
- Zar, J.H. Biostatistical Analysis; Prentice Hall: Englewood Cliffs, NJ, USA, 2010; ISBN 978-0-13-100846-5. [Google Scholar]
- Middleton, B.A.; McKee, K.L. Degradation of Mangrove Tissues and Implications for Peat Formation in Belizean Island Forests. J. Ecol. 2001, 89, 818–828. [Google Scholar] [CrossRef]
- Hilmi, E.; Parengrengi; Vikaliana, R.; Kusmana, C.; Iskandar; Sari, L.K. Setijanto the Carbon Conservation of Mangrove Ecosystem Applied REDD Program. Reg. Stud. Mar. Sci. 2017, 16, 152–161. [Google Scholar] [CrossRef]
- Ochoa-Gómez, J.G.; Lluch-Cota, S.E.; Rivera-Monroy, V.H.; Lluch-Cota, D.B.; Troyo-Diéguez, E.; Oechel, W.; Serviere-Zaragoza, E. Mangrove Wetland Productivity and Carbon Stocks in an Arid Zone of the Gulf of California (La Paz Bay, Mexico). For. Ecol. Manag. 2019, 442, 135–147. [Google Scholar] [CrossRef]
- Chávez-Barrera, J.C. Potential of Ecological Restoration to Recover Carbon Storage in Mangroves in the State of Campeche. Master’s Thesis, Universidad Autónoma de Campeche, Campeche, Mexico, 2020. [Google Scholar]
- May-Colli, L.O. Comportamiento del Carbono, Nitrógeno y Metano en Suelo de Manglar y su Relación con las Condiciones Ambientales; Universidad Autonoma de Campeche—Facultad de Ciencias Quimico Biológicas: Campeche, Mexico, 2018. [Google Scholar]
- Acosta-Velázquez, J. Análisis de la Condición del Bosque de Manglar en el Sistema Lagunar Teacapán-Agua Brava-Marismas Nacionales y Sus Relaciones a Nivel de Paisaje. Master’s Thesis, Centro de Investigación en Alimentación y Desarrollo, Hermosillo, Mexico, 2003. [Google Scholar]
- Maricusa Agraz Hernández, C.; Armando, C.; Keb, C.; Iriarte-Vivar, S.; Venegas, G.P.; Serratos, B.V.; Osti Sáenz, J.; Hernández, A.; Keb, C.A.C.; Iriarte-Vivar, S.I.; et al. Phenological Variation of Rhizophora Mangle and Ground Water Chemistry Associated to Changes of the Precipitation. Hidrobiológica 2015, 25, 49–61. [Google Scholar]
- Ruiz-Fernández, A.C.; Agraz-Hernández, C.M.; Sanchez-Cabeza, J.A.; Díaz-Asencio, M.; Pérez-Bernal, L.H.; Chan Keb, C.A.; López-Mendoza, P.G.; Blanco Y Correa, J.M.; Ontiveros-Cuadras, J.F.; Osti Saenz, J.; et al. Sediment Geochemistry, Accumulation Rates and Forest Structure in a Large Tropical Mangrove Ecosystem. Wetlands 2018, 38, 307–325. [Google Scholar] [CrossRef]
- Marín-Muñiz, J.L.; Hernández, M.E.; Moreno-Casasola, P. Comparing Soil Carbon Sequestration in Coastal Freshwater Wetlands with Various Geomorphic Features and Plant Communities in Veracruz, Mexico. Plant Soil 2014, 378, 189–203. [Google Scholar] [CrossRef]
- Agraz-Hernández, C.M.; Chan-Keb, C.A.; Chávez-Barrera, J.; Osti-Sáenz, J.; Expósito-Díaz, G.; Alonso-Campos, V.A.; Muñiz-Salazar, R.; Ruiz-Fernández, A.C.; Pérez-Bernal, L.H.; Sánchez-Cabeza, J.A.; et al. Reserva de carbón en un ecosistema de manglar al norte de México: Cambios ambientales durante 35 años. Rev. Mex. Biodivers. 2020, 91, e912910. [Google Scholar] [CrossRef]
- Bernal, B.; Mitsch, W.J. A Comparison of Soil Carbon Pools and Profiles in Wetlands in Costa Rica and Ohio. Ecol. Eng. 2008, 34, 311–323. [Google Scholar] [CrossRef]
- Kauffman, J.B.; Donato, D.C.; Adame, M.F. Protocolo para la Medición, Monitoreo y Reporte de la Estructura, Biomasa y Reservas de Carbono de los Manglares; CIFOR: Bogor, Indonesia, 2013. [Google Scholar]
- Mitsch, W.J.; Gosselink, J.G.; Zhang, L.; Anderson, C.J. Wetland Ecosystems; Wiley: Hoboken, NJ, USA, 2009; ISBN 978-0-470-28630-2. [Google Scholar]
- Marín Muñiz, J.L.; Hernández Alarcón, M.E.; Moreno-Casasola Barceló, P. Secuestro de Carbono en Suelos Humedales Costeros de Agua Dulce en Veracruz. Trop. Subtrop. Agroecosyst. 2011, 13, 365–372. [Google Scholar]
- Agraz-Hernandez, C.M.; Chan-Keb, C.A.; Posada-Vanegas, G.; Osti Saenz, J.; Reyes Castellanos, J.E.; Martínez Kumul, J.G.; Conde-Medina, K.P. Respuesta Ecofisiológica de los Manglares del Litoral de Campeche en Condiciones de Extrema Sequía. In Vulnerabilidad de las Zonas Costeras de Latinoamérica al Cambio Climático; UJAT, UNAM, UAC, Botello, A.V., Villanueva, S., Gutiérrez, J., Rojas Galaviz, J.L., Eds.; Departamento de Difusión y Publicaciones del Instituto Epomex-Universidad Autónoma de Campeche: Campeche, Mexico, 2017; p. 476. ISBN 978-607-606-416-0. [Google Scholar]
- Tomlinson, P.B. The Botany of Mangroves; Cambridge University Press: Cambridge, UK, 1986; ISBN 0-521-25567-8. [Google Scholar]
- Yáñez-Arancibia, A.; Lara-Domínguez, A.L. Ecosistemas de Manglar en América Tropical; Unión Mundial para la Naturaleza: Xalapa Veracruz, México, 1999; ISBN 978-968-7863-57-3. [Google Scholar]
- Yáñez-Arancibia, A.; Twilley, R.R.; Lara-Domínguez, A.L. Los Ecosistemas de Manglar Frente al Cambio Climático Global. Madera Bosques 1998, 4, 3–19. [Google Scholar] [CrossRef]
- Moreno Cáliz, E.; Guerrero Peña, A.; Gutiérrez Castorena, M.d.C.; Ortiz Solorio, C.A.; Palma López, D.J. Los manglares de Tabasco, una reserva natural de carbono. Madera Bosques 2002, 8, 115–128. [Google Scholar] [CrossRef]
- Agraz-Hernández, C.M.; Chan-Keb, C.A.; Muñiz-Salazar, R.; Pérez-Balan, R.A.; Osti-Sáenz, J.; Gutiérrez-Alcántara, E.J.; Reyes-Castellano, J.E.; May-Colli, L.O.; Conde-Medina, K.P.; Ruiz-Hernández, J. Relationship between Blue Carbon and Methane and the Hydrochemistry of Mangroves in Southeast Mexico. Appl. Ecol. Environ. Res. 2020, 18, 1091–1106. [Google Scholar] [CrossRef]
- Morales-Ojeda, S.M.; Herrera-Silveira, J.A.; Orellana, R. Almacenes de Carbono en Un Paisaje de Humedal Cárstico a Lo Largo de Un Corredor Transversal Costero de la Península de Yucatán. Madera Bosques 2021, 27. [Google Scholar] [CrossRef]
- Chan-Keb, C.A.; Agraz-Hernández, C.M.; Muñiz-Salazar, R.; Posada-Vanegas, G.; Osti-Sáenz, J.; Reyes Castellano, J.E.; Conde-Medina, K.P.; Vega-Serratos, B.E. Ecophysiological Response of Rhizophora Mangle to the Variation in Hydrochemistry during Five Years along the Coast of Campeche, México. Diversity 2018, 10, 9. [Google Scholar] [CrossRef]
- Agraz-Hernández, C.M.; Osti-Sáenz, J.; Chan-Keb, C.; Expósito-Díaz, E.; Márquez-García, J.; Cach-Ruiz, M.Y. Mangrove Ecosystems in the State of Campeche: Diagnosis of Conservation and Identification of Potential Areas for Restoration; Final Report; Mixed Funds of the Government of the State of Campeche and Science and Technology (CONACYT-FOMIX); Universidad Autónoma de Campeche: Campeche, Mexico, 2012. [Google Scholar]
- Alongi, D.M. Carbon Cycling and Storage in Mangrove Forests. Annu. Rev. Mar. Sci. 2014, 6, 195–219. [Google Scholar] [CrossRef] [PubMed]
- Tue, N.T.; Dung, L.V.; Nhuan, M.T.; Omori, K. Carbon Storage of a Tropical Mangrove Forest in Mui Ca Mau National Park, Vietnam. CATENA 2014, 121, 119–126. [Google Scholar] [CrossRef]
- Dung, L.V.; Tue, N.T.; Nhuan, M.T.; Omori, K. Carbon Storage in a Restored Mangrove Forest in Can Gio Mangrove Forest Park, Mekong Delta, Vietnam. For. Ecol. Manag. 2016, 380, 31–40. [Google Scholar] [CrossRef]
- Tue, N.T.; Hamaoka, H.; Sogabe, A.; Quy, T.D.; Nhuan, M.T.; Omori, K. The Application of δ13C and C/N Ratios as Indicators of Organic Carbon Sources and Paleoenvironmental Change of the Mangrove Ecosystem from Ba Lat Estuary, Red River, Vietnam. Environ. Earth Sci. 2011, 64, 1475–1486. [Google Scholar] [CrossRef]
- Murdiyarso, D.; Purbopuspito, J.; Kauffman, J.B.; Warren, M.W.; Sasmito, S.D.; Donato, D.C.; Manuri, S.; Krisnawati, H.; Taberima, S.; Kurnianto, S. The Potential of Indonesian Mangrove Forests for Global Climate Change Mitigation. Nat. Clim. Change 2015, 5, 1089–1092. [Google Scholar] [CrossRef]
- Krauss, K.W.; Lovelock, C.E.; McKee, K.L.; López-Hoffman, L.; Ewe, S.M.L.; Sousa, W.P. Environmental Drivers in Mangrove Establishment and Early Development: A Review. Aquat. Bot. 2008, 89, 105–127. [Google Scholar] [CrossRef]
- Setia, R.; Smith, P.; Marschner, P.; Baldock, J.; Chittleborough, D.; Smith, J. Introducing a Decomposition Rate Modifier in the Rothamsted Carbon Model to Predict Soil Organic Carbon Stocks in Saline Soils. Environ. Sci. Technol. 2011, 45, 6396–6403. [Google Scholar] [CrossRef]
- Naidoo, G. The Mangroves of South Africa: An Ecophysiological Review. S. Afr. J. Bot. 2016, 107, 101–113. [Google Scholar] [CrossRef]
- Kreuzwieser, J.; Buchholz, J.; Rennenberg, H. Emission of Methane and Nitrous Oxide by Australian Mangrove Ecosystems. Plant Biol. 2003, 5, 423–431. [Google Scholar] [CrossRef]
- Chan-Keb, C.A.; Agraz-Hernández, C.M.; Pérez-Balan, R.A.; Gutiérrez-Alcántara, E.J.; Muñiz-Salazar, R.; Reyes-Castellano, J.E.; Osti-Sáenz, J. Phytotoxicity in Seedlings of Rhizophora Mangle (L.) Exposed to 2,4-Dichlorophenoxyacetic Acid under Experimental Conditions. J. Mar. Sci. Eng. 2021, 9, 1417. [Google Scholar] [CrossRef]
- Adame, M.F.; Brown, C.J.; Bejarano, M.; Herrera-Silveira, J.A.; Ezcurra, P.; Kauffman, J.B.; Birdsey, R. The Undervalued Contribution of Mangrove Protection in Mexico to Carbon Emission Targets. Conserv. Lett. 2018, 11, e12445. [Google Scholar] [CrossRef]
- Kauffman, J.B.; Hernandez Trejo, H.; Del Carmen Jesus Garcia, M.; Heider, C.; Contreras, W.M. Carbon Stocks of Mangroves and Losses Arising from Their Conversion to Cattle Pastures in the Pantanos de Centla, Mexico. Wetl. Ecol. Manag. 2016, 24, 203–216. [Google Scholar] [CrossRef]
- Mata-Fernández, I.; Rodríguez-Gamiño, M.L.; López-Blanco, J.; Vela-Correa, G. Dinámica de la Salinidad en los Suelos. Rev. Digit. Dep. Hombre Ambiente 2014, 1, 26–35. [Google Scholar]
- Zedler, J.B.; Callaway, J.C. Tidal Wetland Functioning. J. Coast. Res. 2001, 27, 38–64. [Google Scholar]
- Barendregt, A.; Swarth, C.W. Tidal Freshwater Wetlands: Variation and Changes. Estuaries Coasts 2013, 36, 445–456. [Google Scholar] [CrossRef]
- Rincón Pérez, M.; Infante-Mata, D.; Moreno-Casasola, P.; Hernández Alarcón, M.E.; Barbas Macías, E.; García-Alfaro, J.R. Patrones de Distribución y Estructura de la Vegetación en el Gradiente de Humedales Costeros el Castaño, Chiapas, México. Rev. Biol. Trop. 2020, 68, 242–259. [Google Scholar] [CrossRef]
- Gleason, S.M.; Ewel, K.C.; Hue, N. Soil Redox Conditions and Plant–Soil Relationships in a Micronesian Mangrove Forest. Estuar. Coast. Shelf Sci. 2003, 56, 1065–1074. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Place | Depth (cm) | Organic Carbon (Mg C/ha) | Reference |
|---|---|---|---|
| Ohio, USA | 0–24 | 90.3 | [38] |
| Costa Rica | 0–24 | 68.2 | [38] |
| Indonesia | 0–10 | 28 | [39] |
| Mozambique | 0–30 | 53.3 | [14] |
| Bangladesh | 0–30 | 50.8 | [39] |
| Mexican Caribbean | 0–30 | 83.5 | [12] |
| Nayarit, Mexico | 0–20 | 89.7 | [23] |
| Nayarit, Mexico | 0–20 | 36.2 | [37] |
| Veracruz, Mexico | 0–20 | 969 | [36] |
| Campeche, Mexico | 0–50 | 332 | [31] |
| Campeche, Mexico | 0–10 | 174.6 | [32] |
| This study | 0–30 | ||
| Reference Forest (BR) | 470.17 | ||
| Restoration Area (AR) | 440.53 |
| Site/Variable Dependent (Y) | Equation | R2 | p |
|---|---|---|---|
| Reference Forest | |||
| SC (Mg C/ha) | Y = 20.39 + 3.313 X1 | 0.41 | 0.001 |
| SC (Mg C/ha) | Y = 571.3 − 85.20 X2 | 0.23 | 0.024 |
| CO (%) | Y = 33.97 − 49.51 X3 | 0.21 | 0.031 |
| Restoration Area | |||
| SC (Mg C/ha) | Y = 34.93 + 0.4175 X4 | 0.48 | 0.0001 |
| SC (Mg C/ha) | Y = −11.9 + 2.441 X1 + 0.4690 X4 | 0.79 | 0.0001 |
| CO (%) | Y = 28.50 − 28.49 X3 | 0.71 | 0.001 |
| PS (cm) | Y = 9.161 + 14.14 X3 | 0.23 | 0.018 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chan-Keb, C.A.; Agraz-Hernández, C.M.; Pérez-Balan, R.A.; Mas-Qui, O.O.; Osti-Sáenz, J.; Reyes-Castellanos, J.E. Relationship between Carbon Sequestration and Soil Physicochemical Parameters in Northern Campeche, Mexico. Land 2024, 13, 139. https://doi.org/10.3390/land13020139
Chan-Keb CA, Agraz-Hernández CM, Pérez-Balan RA, Mas-Qui OO, Osti-Sáenz J, Reyes-Castellanos JE. Relationship between Carbon Sequestration and Soil Physicochemical Parameters in Northern Campeche, Mexico. Land. 2024; 13(2):139. https://doi.org/10.3390/land13020139
Chicago/Turabian StyleChan-Keb, Carlos A., Claudia M. Agraz-Hernández, Román A. Pérez-Balan, Oscar O. Mas-Qui, Juan Osti-Sáenz, and Jordán E. Reyes-Castellanos. 2024. "Relationship between Carbon Sequestration and Soil Physicochemical Parameters in Northern Campeche, Mexico" Land 13, no. 2: 139. https://doi.org/10.3390/land13020139