1. Introduction
Soil salinization is recognized as the main threat to environmental resources and affects between 7 and 10% of the continental extension [
1]. A soil is considered saline when the ion concentration interferes with the growth of the species of agricultural interest and reaches an electrical conductivity >4 dS m
−1 (approximately 36 mM NaCl), determined in the soil paste at 25° [
2]. The presence of salts in the soil is of pedogenic (weathering) or anthropic (irrigation or fertilization water) origin and is exacerbated when rainfall is insufficient to leach ions from the soil profile, or when evaporation rates are high [
2,
3,
4].
Salinity decreases agricultural production in most crops by affecting the physical and chemical properties of the soil and the ecological balance of the cultivated area [
1,
4]. High levels of salinity (>4 dS m
−1) induce physiological stress because its ions produce toxicity, osmotic, and oxidative stress, and nutrient deficiency (N, Ca, K, P, Fe, Zn) which limit the uptake of nutrients from the soil. Salinity also affects photosynthesis, due to the reduction in leaf area, chlorophyll content, stomatal conductance and, to a lesser extent, the efficiency of photosystem II. This is reflected in a decrease in germination, vegetative growth, and reproductive development in different crops [
2,
3,
5].
In this context, the avocado has been identified as one of the fruit trees most sensitive to salts [
6,
7], since the trees of this species irrigated with water whose chloride and sodium content is at tolerable levels for most of the crops show severe damage to the leaves, restriction of vegetative growth, and decreased production volume [
6]. This situation occurs worldwide in those areas with an arid and semi-arid climate where avocados are grown [
8], which has led to the search for different measures to deal with salinity, among which the selection of salt-tolerant rootstocks represents a feasible but not sufficient alternative [
9].
On the desert coast of Peru, a pole of agro-industrial development and avocado cultivation has been established since 2018 as the main agro-export sector [
10]; there are approximately 33,000 hectares of “Hass” avocado [
11], where the degradation of these lands is mainly due to soil salinity, which affects approximately 40% of the arable surface in the Piura, Tumbes, and Lambayeque regions, which are the most important areas for agricultural production for export [
12].
In addition, it has been determined that in the coastal areas of Peru, there are approximately 300,000 hectares that have salinization problems, which not only affect avocado cultivation, but also, and especially, areas where rice is grown [
13] among others. These saline soils have been formed in part by natural conditions such as rainfall deficit, arid and semi-arid environments; although, in most cases, it has increased due to artificial conditions such as irrigation with saline waters, inadequate irrigation, and poor drainage [
14]. So, saline soils in Peru are widely distributed and are generally treated by amendments with calcium sulfate, application of organic amendments, and reseeding or change of crop, whose action is local and temporary and even expensive. Therefore, methods are required that are long-lasting and sustainable to the environment, while maintaining agricultural activities [
15].
Soil salinity then generates a depressing effect on plant growth, and some plant growth promoting bacteria (PGPR) or other microorganisms that can attenuate the stress that this produces [
16]. In the case of the genus
Bacillus, various authors indicate that
B. subtilis reduces the level of oxidative and osmotic stress induced in seedlings that manifests itself in the improvement of plant growth [
17,
18,
19]. In the same way, the genus
Pseudomonas has been shown to reduce the level of salinity stress in cultivated plants by decreasing peroxidase activity in the plant and increasing nutrient uptake and improving plant growth under salinity conditions at high concentrations [
20,
21].
On the other hand, arbuscular mycorrhizae (AM) associate with most plants of agricultural interest and improve growth under stress conditions due to adverse soil conditions; specifically, they modulate salinity stress [
22,
23] by root colonization, modifying antioxidant enzyme systems that reduce oxidative damage and stimulate growth [
24,
25,
26]. AM symbiosis increases proline synthesis and superoxide dismutase, catalase, and ascorbate peroxidase activities that decrease the synthesis of phenolic compounds related to salinity stress [
27].
Regarding the joint use of mycorrhizae and plant growth-promoting bacteria (PGPR), it is known that they can influence the superior response of plants to abiotic stresses such as drought and salinity through different mechanisms such as increased potassium uptake and decreased sodium by the plant [
28,
29]. In addition, synergistic effects potentially contribute to expanding crop production to soils that would otherwise be unproductive through interactions that may have a potential role in relieving stress [
29], or improving nutrient uptake and crop productivity cultures compared to a single inoculation [
30,
31].
Therefore, microorganisms that promote plant growth constitute potential alternatives for sustainable agriculture due to their low cost of production and high effectiveness. Hence, the objective of this research was to evaluate PGPR and arbuscular mycorrhizae in improving resistance to salinity stress in avocado seedlings of the Zutano variety.
2. Materials and Methods
2.1. Selection of PGPR Resistant to High Osmotic Concentrations
From 26 isolates of bacteria belonging to the genera
Bacillus and
Pseudomonas from the rhizosphere of
P. americana, 7 strains were selected according to the growth-promoting activity associated with phosphate solubilization [
32], synthesis of siderophores [
33], and antagonistic activity against
P. cinnamomi and/or
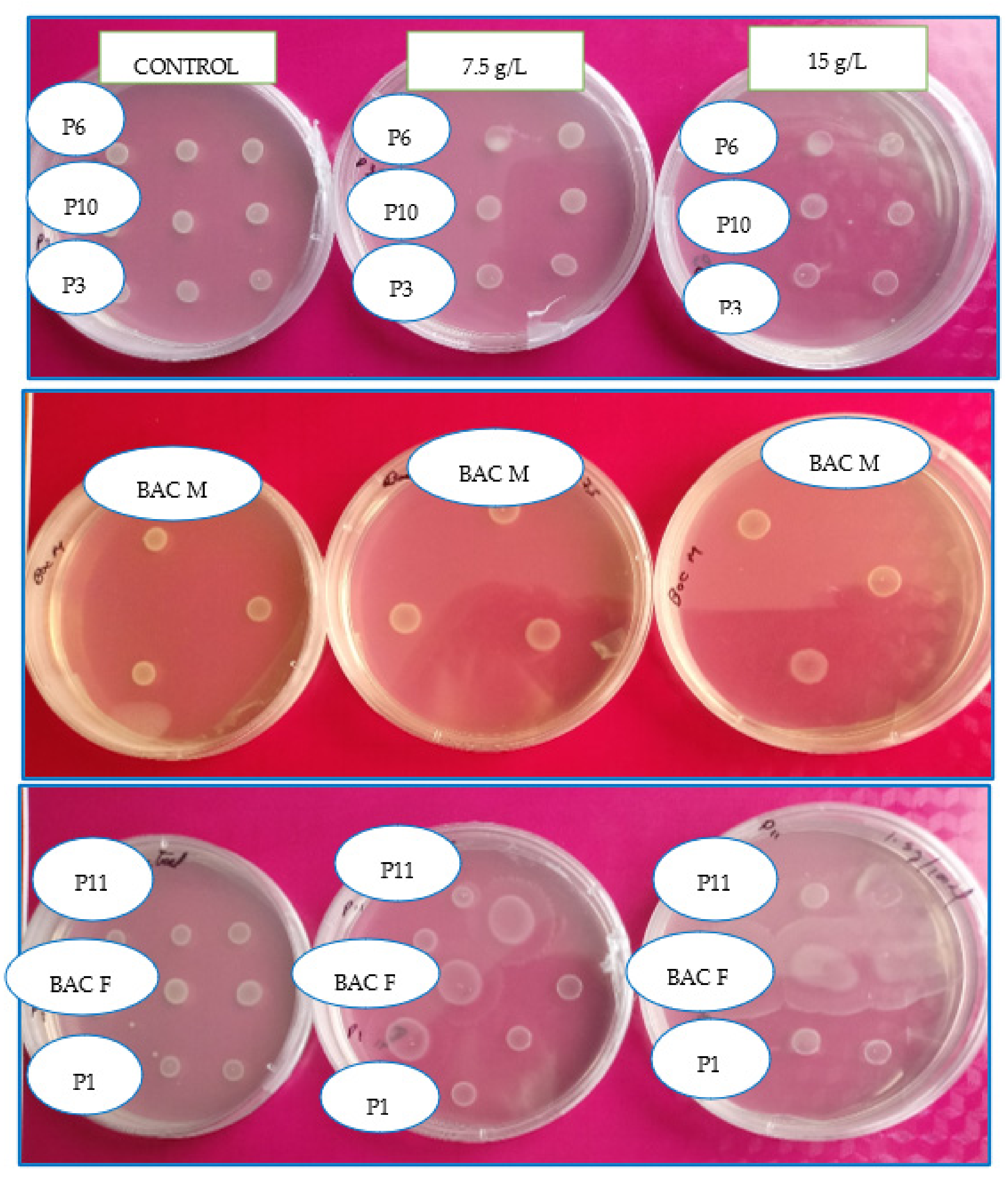
L. theobromae for representing the most important pathogens for avocado cultivation in Peru and the world. Then, those showing greater growth under salinity conditions in increasing concentrations of salinity in nutrient agar with 7.5 and 15 g·L
−1 of sodium chloride, respectively, were selected.
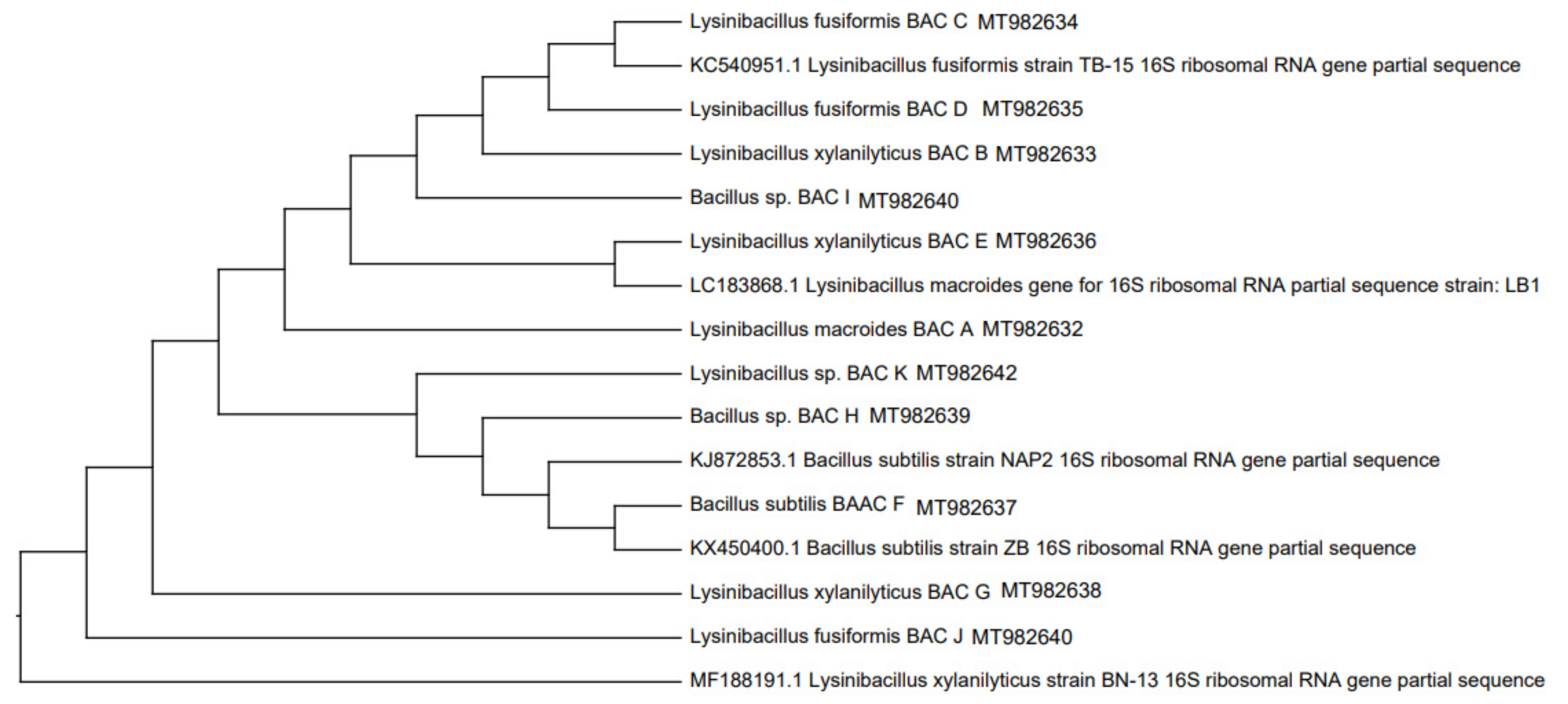
Previously, the strains isolated from the rhizosphere were identified by amplifying the 16S rDNA gene, for which they were activated in nutrient broth at 37 °C for 18 h, and the genomic DNA of each strain was extracted according to the instructions of the commercial kit (Thermo SCIENTIFIC, USA) and later sequenced by MacroGen Inc. (Seoul, Korea). To determine their taxonomic identity, BLAST analysis of the similarity between the sequences of the nucleotides obtained and those deposited in the Gen Bank database was used. Phylogenetic trees of 16S rRNA were deduced using the neighbor-joining analysis [
34]. Evolutionary analyses were performed in MEGA X (2018).
2.2. Description of the Substrate Used for the Tests
Premix ® N°8 substrate (pH: 5.5, EC: 0.75 dS m−1, P: 25 ppm and K: 100 ppm) was added to 2-L plastic pots; one avocado seed per pot was sown of the var. Zutano weighing approximately 50 g; the seeds were donated by the company Avo Hass Peru, previously sterilized in 10% sodium hypochlorite for 10 min and once rinsed, each one of them was allowed to germinate for ten days.
2.3. Preparation of Avocado Seeds for the Installation of the Experiment
Once the seeds germinated after ten days in the 2 L pots, they were inoculated with the bacteria or Glomeromycota fungi according to the corresponding test and they were allowed to grow for three weeks in a greenhouse with an average temperature of 20ºC and humidity of 60% until the seedlings were obtained, prior to treatment with irrigation with saline water.
2.4. Selection, Propagation, and Inoculation of Arbuscular Mycorrhizae
Two inocula of
Glomeromycota fungi isolated from Peru were used. One of them was obtained from rhizospheric soil from a fallow land in Pucallpa, Ucayali (Glomeromycota Fallow Inoculum-GFI) and the other one from rhizospheric soil of
Sporobolus sp. located in Pisco, Ica (Glomeromycota Wetland Inoculum-GWI), according to Castañeda et al. [
35]. Both were reproduced in trap pots with
Brachiaria decumbens in sterile sand, irrigated with Long Ashton’s solution, and a quarter of the dose of P every 15 days [
36]. Rhizospheric samples were taken to quantify spores and AM colonization of the rootlets [
37,
38] in each inoculum. A similar number of spores was obtained in GFI and GWI to inoculate the avocado plants in the experiment. The GFI inoculum consisted of the native species, mainly the fungus
R. intraradices and the genera
Acaulospora,
Gigaspora, and
Archaeospora, of which 3726 spores/mL per seedling were added from colonized rootlets (70%). In the GWI inoculum, the fungus
R. intraradices predominated, of which 3400 spores/mL per seedling were applied from colonized rootlets (90%). In both cases, the inocula were applied at the base of the pre-germinated seeds, spreading the inoculum around, slightly uncovering the substrate with a sterile spatula to expose the rootlets without damaging them.
2.5. Inoculation of the Seedlings with the Bacterial Strains
The bacteria selected for their resistance to growing in saline conditions in vitro were cultured for 48 h in nutrient broth at 28 °C; then, 30 mL of bacterial broth from each strain with a concentration of 108 cfu mL−1 was applied to the neck of the plant, 15 days after germination of the seed, and the area of influence of the rootlets. This procedure was repeated a second time 3 days after the first inoculation to guarantee its presence and adequacy in the rhizosphere of the plant before starting the saline irrigation treatments.
2.6. Greenhouse Environmental Conditions
Throughout the experiment, the maximum temperatures recorded varied between 24.51 and 27.43 °C, and the minimum between 18.91 and 20.37 °C. The relative humidity varied between 59.9 and 61.92%. These parameters were recorded by the meteorological station located inside the greenhouse of the Microbial Ecology and Biotechnology Laboratory of the La Molina National Agrarian University.
2.7. Effect of Plant Growth Promoting Rhizobacteria on Salt Stress Tolerance in Persea americana var. Zutano
Nine treatments were established based on two factors: (a) inoculation of plant growth promoting rhizobacteria (Bacillus subtilis, Pseudomonas plecoglossicida and without bacteria) and (b) sodium chloride concentrations (irrigation water without sodium chloride, irrigation water with 0.75 g/L of sodium chloride, and irrigation water with 1.5 g/L of sodium chloride). The test was maintained for 60 days with an irrigation frequency of three times a week and a dose of 500 mL of water per plant.
2.8. Effect of Inoculation with Glomeromycota fungi on Tolerance to Salt Stress in Persea americana var. Zutano
Seven treatments were established based on two factors: (a) mycorrhizae (isolated from wetland soils, GWI, and isolated from fallow soils, GFI) and (b) sodium chloride concentrations (irrigation water without sodium chloride, irrigation water with 0.75 g/L of sodium chloride and irrigation water with 1.5 g/L of sodium chloride). The test was maintained for 60 days with an irrigation frequency of three times a week and a dose of 500 mL per plant.
2.9. Evaluated Parameters
Growth parameters were evaluated in the seedlings associated with plant height, stem diameter, and the accumulation of fresh and dry matter both in the root and in the aerial part, for which the samples were dried at 40 °C for 5 days until no variation in the weight of the sample was observed. Additionally, the content of ions related to saline stress, such as K, Na, and Cl, was measured both in the aerial biomass (leaves and stems) and in the root biomass.
2.10. Statistical Analysis
For the statistical analysis, the Statistic Package for Social Sciences (SPSS) program from IBM company version 26 was used. The data obtained in each experiment were submitted to the analysis of variance for a complete randomized design (DCA) with factorial arrangement using the F test and by calculating the main effects of the tested factors; then, the treatments were compared in case of significance using Duncan’s test to determine differences between strains and salinity levels tested. The probability of alpha error of less than 5% was considered significant.
5. Conclusions
The inoculation of seedlings with both PGPR, P. plecoglossicida, and B. subtilis, contributes to the reduction in the accumulation of chlorine and sodium ions, while they increase the accumulation of potassium in avocado leaves. Chlorine and sodium ions also decrease while potassium accumulation in leaves and roots increases.
The inoculation of seedlings with GFI and GWI Glomeromycota fungi reduced the impact of salinity, improving plant growth, and decreasing the accumulation of chlorine and sodium ions in leaves and roots below control where mycorrhizae were not applied. The beneficial microorganisms tested are promising in the mitigation of salt stress in avocado, highlighting the inoculum native from saline soil (GWI).
In the future, it is necessary to study the feasibility of using the co-inoculation of Glomeromycota fungi and plant growth promoting rhizobacteria from saline soils in avocado seedlings, particularly with both bacteria used in this work and GWI Glomeromycota inoculum. In addition, efficacy tests should be conducted in the field to allow the development of commercial inoculants with these microorganisms for the treatment of saline soils in avocado cultivation.










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}