
AGER-1 Long Non-Coding RNA Levels Correlate with the Expression of the Advanced Glycosylation End-Product Receptor, a Regulator of the Inflammatory Response in Visceral Adipose Tissue of Women with Obesity and Type 2 Diabetes Mellitus

 , , ,
, , , 
Abstract
:1. Introduction
2. Results
2.1. Clinical and Biochemical Characteristics of Study Participants
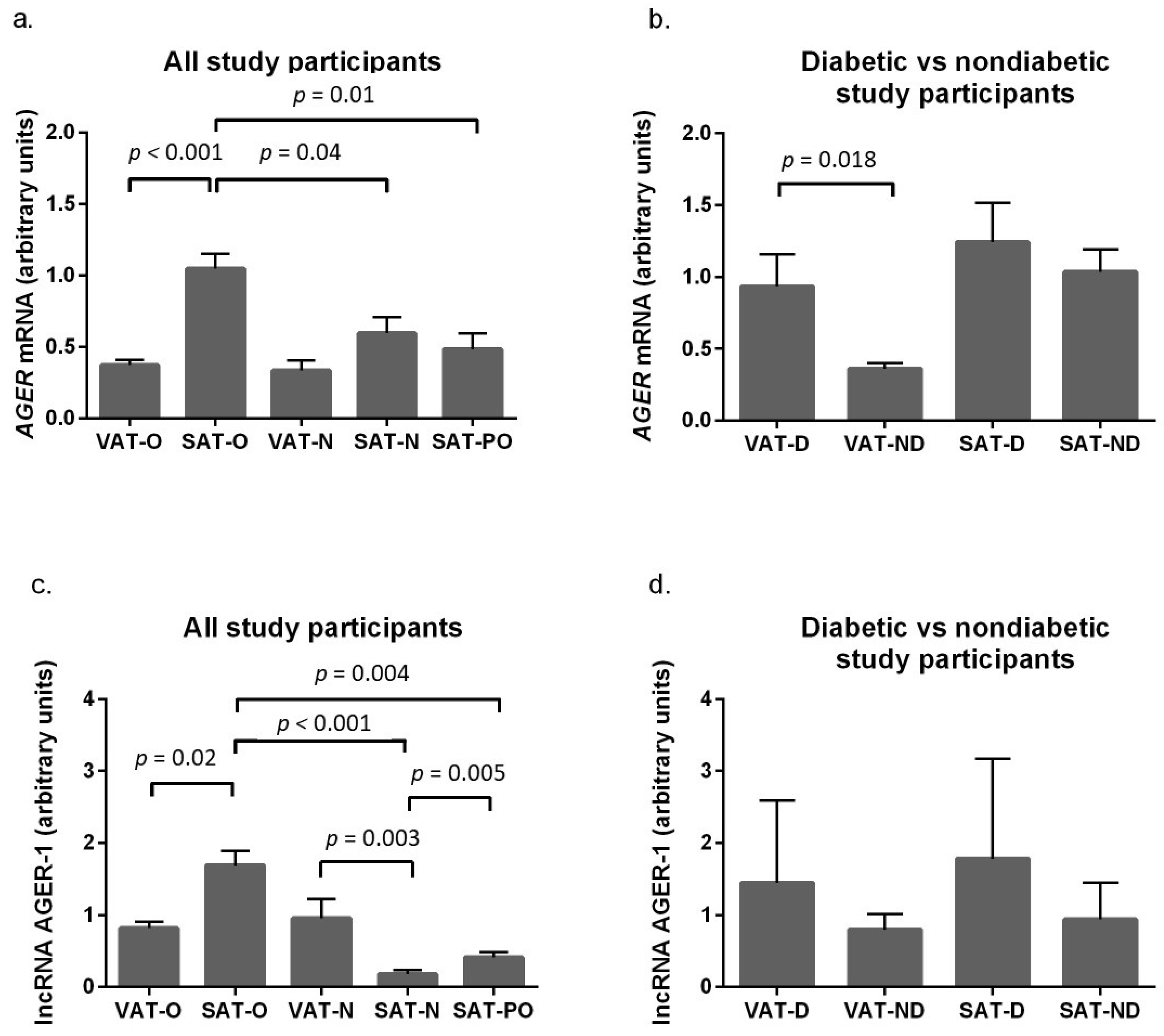
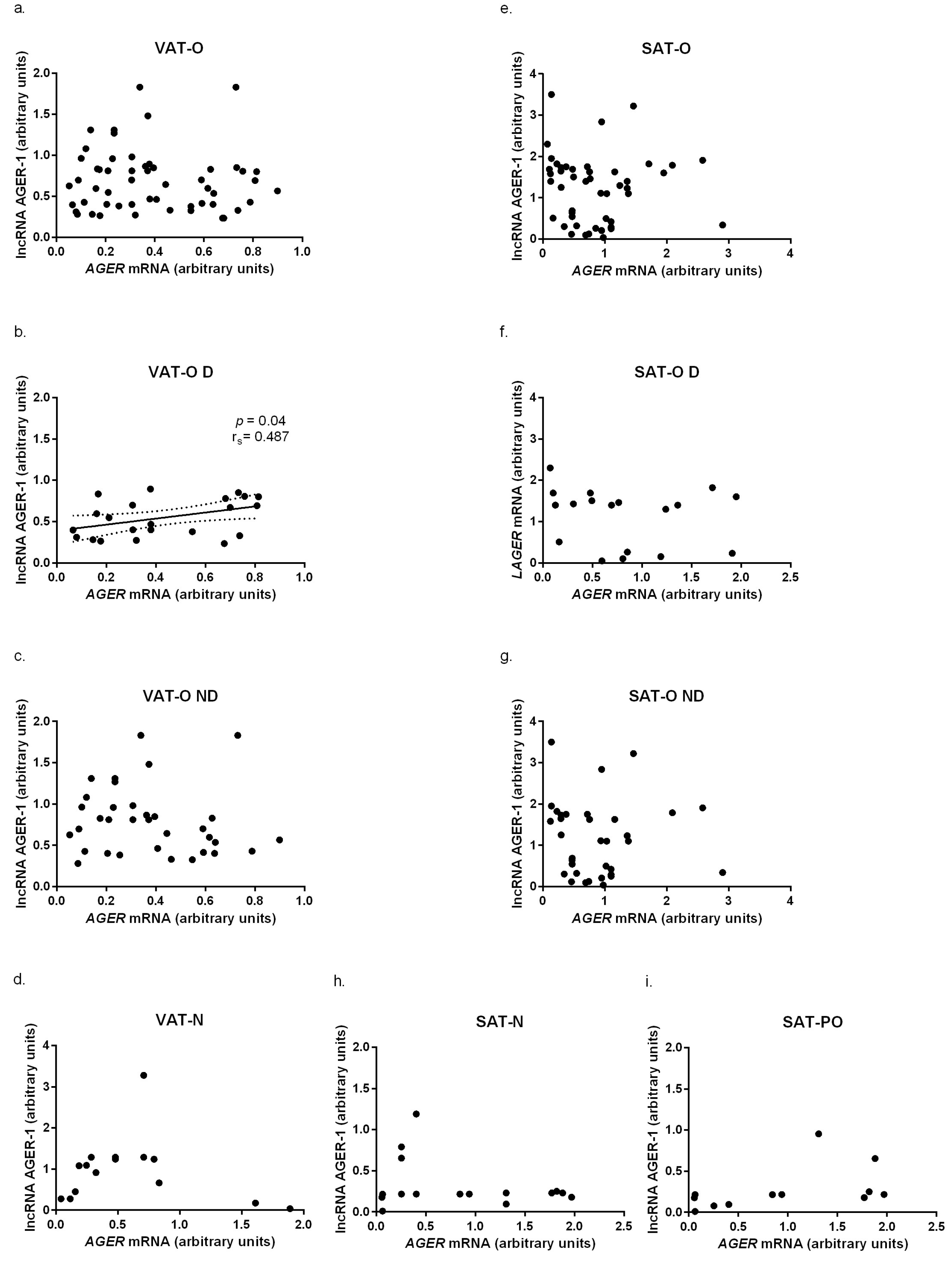
2.2. The Advanced Glycosylation End-Product Receptor (AGER) Gene and Long Non-Coding RNA AGER-1 Expression in Adipose Tissue of Obese Individuals before and after Bariatric Surgery and Normal Weight Subjects
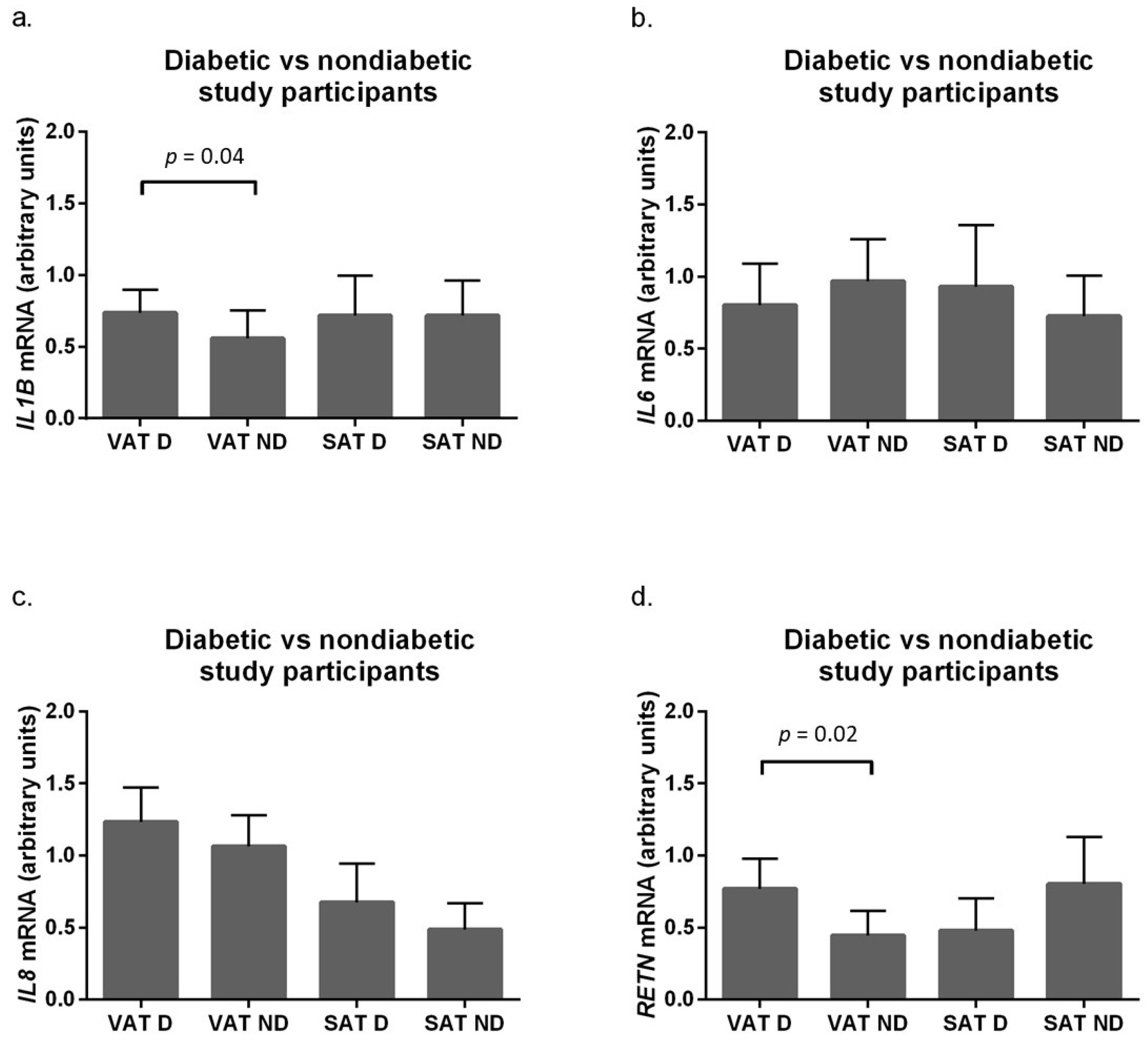
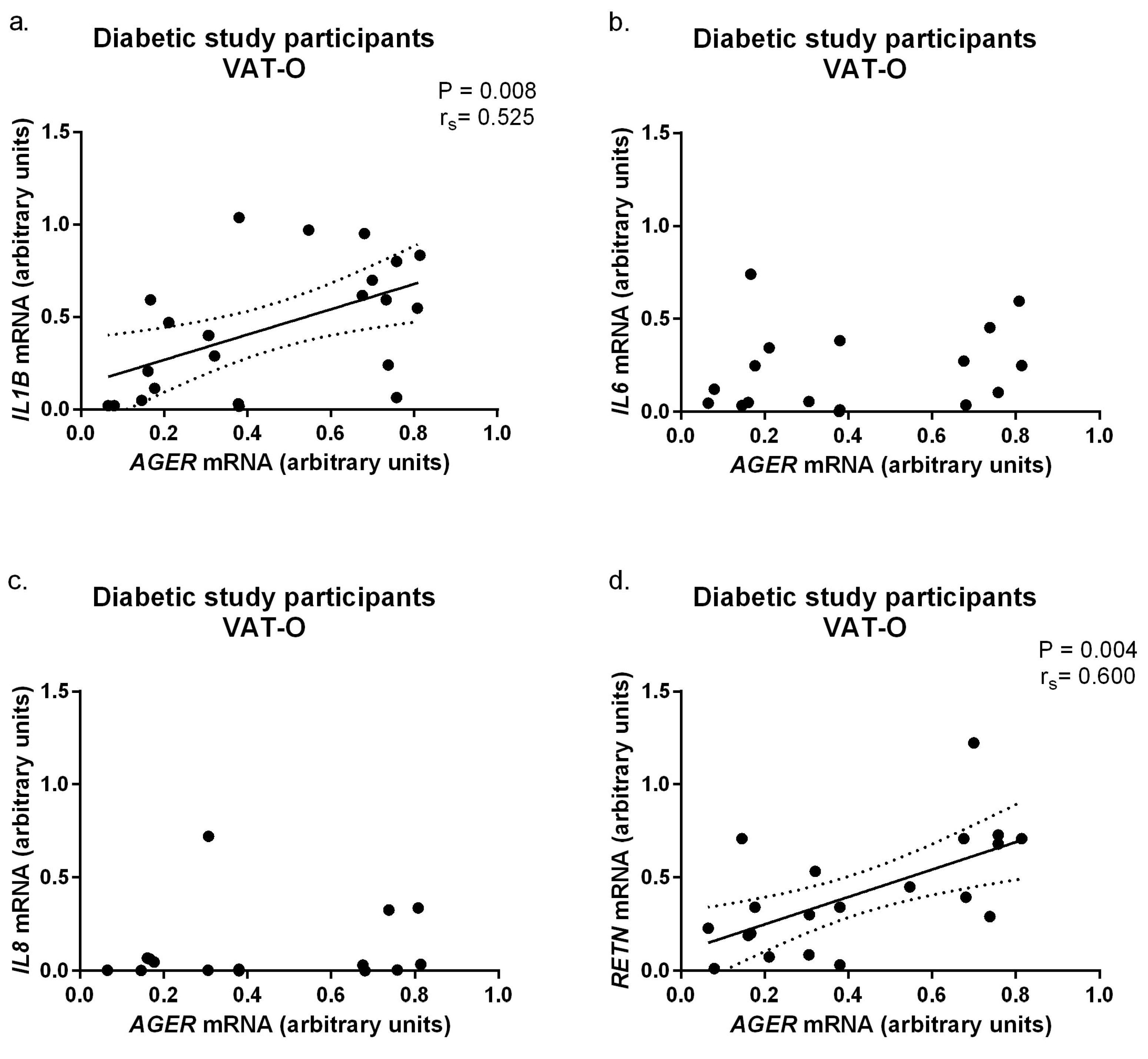
2.3. Expression of Pro-Inflammatory Adipokines in Adipose Tissue of Obese Subjects, Stratified by Diabetic Status
3. Discussion
4. Materials and Methods
4.1. Tissue and Blood Sample Collection
4.2. Isolation of Total RNA, Reverse Transcription, and Real-Time PCR
4.3. Isolation of a Protein Fraction from Adipose Tissue and Measurement of Cytokine Concentrations
4.4. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kwok, K.; Lam, K.; Xu, A. Heterogeneity of white adipose tissue: Molecular basis and clinical implications. Exp. Mol. Med. 2016, 48, e215. [Google Scholar] [CrossRef] [PubMed]
- Cypess, A.M. Reassessing Human Adipose Tissue. N. Engl. J. Med. 2022, 386, 768–779. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Sun, Q.; Wu, X.; Zhang, Y.; Xing, X.; Lin, K.; Feng, Y.; Wang, M.; Wang, Y.; Wang, R. Hypoxia as a Double-Edged Sword to Combat Obesity and Comorbidities. Cells 2022, 11, 3735. [Google Scholar] [CrossRef] [PubMed]
- Woo, C.Y.; Jang, J.E.; Lee, S.E.; Koh, E.H.; Lee, K.U. Mitochondrial Dysfunction in Adipocytes as a Primary Cause of Adipose Tissue Inflammation. Diabetes Metab. J. 2016, 43, 247–256. [Google Scholar] [CrossRef] [PubMed]
- Reilly, S.M.; Saltiel, A.R. Adapting to obesity with adipose tissue inflammation. Nat. Rev. Endocrinol. 2017, 13, 633–643. [Google Scholar] [CrossRef]
- Wondmkun, Y.T. Obesity, Insulin Resistance, and Type 2 Diabetes: Associations and Therapeutic Implications. Diabetes. Metab. Syndr. Obes. 2020, 13, 3611–3616. [Google Scholar] [CrossRef]
- Yan, S.F.; Ramasamy, R.; Schmidt, A.M. Mechanisms of disease: Advanced glycation end-products and their receptor in inflammation and diabetes complications. Nat. Clin. Pract. Endocrinol. Metab. 2008, 4, 285–293. [Google Scholar] [CrossRef]
- Ribeiro, P.V.M.; Tavares, J.F.; Costa, M.A.C.; Mattar, J.B.; Alfenas, R.C.G. Effect of reducing dietary advanced glycation end products on obesity-associated complications: A systematic review. Nutr. Rev. 2019, 77, 725–734. [Google Scholar] [CrossRef]
- Senatus, L.M.; Schmidt, A.M. The AGE-RAGE Axis: Implications for Age-Associated Arterial Diseases. Front. Genet. 2017, 8, 187. [Google Scholar] [CrossRef]
- Sessa, L.; Gatti, E.; Zeni, F.; Antonelli, A.; Catucci, A.; Koch, M.; Pompilio, G.; Fritz, G.; Raucci, A.; Bianchi, M.E. The receptor for advanced glycation end-products (RAGE) is only present in mammals and belongs to a family of cell adhesion molecules (CAMs). PLoS ONE 2014, 9, e86903. [Google Scholar] [CrossRef]
- Rouhiainen, A.; Kuja-Panula, J.; Tumova, S.; Rauvala, H. RAGE-mediated cell signaling. Methods Mol. Biol. 2013, 963, 239–263. [Google Scholar] [CrossRef] [PubMed]
- Chini, A.; Guha, P.; Malladi, V.S.; Guo, Z.; Mandal, S.S. Novel long non-coding RNAs associated with inflammation and macrophage activation in humans. Sci. Rep. 2023, 13, 4036. [Google Scholar] [CrossRef] [PubMed]
- Mattick, J.S.; Makunin, I.V. Non-coding RNA. Hum. Mol. Genet. Spec. 2006, 1, R17–R29. [Google Scholar] [CrossRef] [PubMed]
- Wijesinghe, S.N.; Nicholson, T.; Tsintzas, K.; Jones, S.W. Involvements of long noncoding RNAs in obesity-associated inflammatory diseases. Obes. Rev. 2021, 22, e13156. [Google Scholar] [CrossRef]
- Yang, L.; Wang, X.; Guo, H.; Zhang, W.; Wang, W.; Ma, H. Whole Transcriptome Analysis of Obese Adipose Tissue Suggests u001kfc.1 as a Potential Regulator to Glucose Homeostasis. Front. Genet. 2019, 10, 1133. [Google Scholar] [CrossRef]
- Sun, J.; Ruan, Y.; Wang, M.; Chen, R.; Yu, N.; Sun, L.; Liu, T.; Chen, H. Differentially expressed circulating LncRNAs and mRNA identified by microarray analysis in obese patients. Sci. Rep. 2016, 6, 35421. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.; Kerr, A.; Jiao, H.; Hon, C.C.; Rydén, M.; Dahlman, I.; Arner, P. Long Non-Coding RNAs Associated with Metabolic Traits in Human White Adipose Tissue. EBioMedicine 2018, 30, 248–260. [Google Scholar] [CrossRef] [PubMed]
- American Diabetes Association Professional Practice Committee. Classification and Diagnosis of Diabetes: Standards of Medical Care in Diabetes—2022. Diabetes Care 2022, 45, S17–S38. [Google Scholar] [CrossRef] [PubMed]
- Jonas, M.I.; Kurylowicz, A.; Bartoszewicz, Z.; Lisik, W.; Jonas, M.; Wierzbicki, Z.; Chmura, A.; Pruszczyk, P.; Puzianowska-Kuznicka, M. Interleukins 6 and 15 Levels Are Higher in Subcutaneous Adipose Tissue, but Obesity Is Associated with Their Increased Content in Visceral Fat Depots. Int. J. Mol. Sci. 2015, 16, 25817–25830. [Google Scholar] [CrossRef]
- Jonas, M.I.; Kurylowicz, A.; Bartoszewicz, Z.; Lisik, W.; Jonas, M.; Domienik-Karlowicz, J.; Puzianowska-Kuznicka, M. Adiponectin/resistin interplay in serum and adipose tissue of obese and normal-weight individuals. Diabetol. Metab. Syndr. 2017, 9, 95. [Google Scholar] [CrossRef]
- Gaens, K.H.; Goossens, G.H.; Niessen, P.M.; van Greevenbroek, M.M.; van der Kallen, C.J.; Niessen, H.W. Nε-(carboxymethyl)lysine-receptor for advanced glycation end product axis is a key modulator of obesity-induced dysregulation of adipokine expression and insulin resistance. Arter. Thromb. Vasc. Biol. 2014, 34, 1199–1208. [Google Scholar] [CrossRef] [PubMed]
- Baptista, L.S.; Silva, K.R.; Jobeili, L.; Guillot, L.; Sigaudo-Roussel, D. Unraveling White Adipose Tissue Heterogeneity and Obesity by Adipose Stem/Stromal Cell Biology and 3D Culture Models. Cells 2023, 12, 1583. [Google Scholar] [CrossRef] [PubMed]
- Kurylowicz, A. microRNAs in Human Adipose Tissue Physiology and Dysfunction. Cells 2021, 10, 3342. [Google Scholar] [CrossRef] [PubMed]
- Haberka, M.; Machnik, G.; Kowalówka, A.; Biedroń, M.; Skudrzyk, E.; Regulska-Ilow, B.; Gajos, G.; Manka, R.; Deja, M.; Okopień, B.; et al. Epicardial, paracardial, and perivascular fat quantity, gene expressions, and serum cytokines in patients with coronary artery disease and diabetes. Pol. Arch. Intern. Med. 2019, 129, 738–746. [Google Scholar] [CrossRef] [PubMed]
- Kawai, T.; Autieri, M.V.; Scalia, R. Adipose tissue inflammation and metabolic dysfunction in obesity. Am. J. Physiol. Cell. Physiol. 2021, 320, C375–C391. [Google Scholar] [CrossRef] [PubMed]
- Samaras, K.; Botelho, N.K.; Chisholm, D.J.; Lord, R.V. Subcutaneous and visceral adipose tissue gene expression of serum adipokines that predict type 2 diabetes. Obesity 2010, 18, 884–889. [Google Scholar] [CrossRef]
- Donath, M.Y.; Böni-Schnetzler, M. IL-1β activation as a response to metabolic disturbances. Cell Metab. 2010, 12, 427–428. [Google Scholar] [CrossRef] [PubMed]
- Eder, K.; Baffy, N.; Falus, A.; Fulop, A.K. The major inflammatory mediator interleukin-6 and obesity. Inflamm. Res. 2009, 58, 727–736. [Google Scholar] [CrossRef]
- Saito, T.; Murata, M.; Otani, T.; Tamemoto, H.; Kawakami, M.; Ishikawa, S.E. Association of subcutaneous and visceral fat mass with serum concentrations of adipokines in subjects with type 2 diabetes mellitus. Endocr. J. 2012, 59, 39–45. [Google Scholar] [CrossRef]
- Beasley, L.E.; Koster, A.; Newman, A.B.; Javaid, M.K.; Ferrucci, L.; Kritchevsky, S.B.; Kuller, L.H.; Pahor, M.; Schaap, L.A.; Visser, M.; et al. Health ABC study. Health ABC study. Inflammation and race and gender differences in computerized tomography-measured adipose depots. Obesity 2009, 17, 1062–1069. [Google Scholar] [CrossRef]
- Spoto, B.; Di Betta, E.; Mattace-Raso, F.; Sijbrands, E.; Vilardi, A.; Parlongo, R.M.; Pizzini, P.; Pisano, A.; Vermi, W.; Testa, A.; et al. Pro- and anti-inflammatory cytokine gene expression in subcutaneous and visceral fat in severe obesity. Nutr. Metab. Cardiovasc. Dis. 2014, 24, 1137–1143. [Google Scholar] [CrossRef] [PubMed]
- Fox, C.S.; Massaro, J.M.; Hoffmann, U.; Pou, K.M.; Maurovich-Horvat, P.; Liu, C.Y.; Vasan, R.S.; Murabito, J.M.; Meigs, J.B.; Cupples, L.A.; et al. Abdominal visceral and subcutaneous adipose tissue compartments: Association with metabolic risk factors in the Framingham Heart Study. Circulation 2007, 116, 39–48. [Google Scholar] [CrossRef]
- Kim, S.K.; Kim, H.J.; Hur, K.Y.; Choi, S.H.; Ahn, C.W.; Lim, S.K.; Kim, K.R.; Lee, H.C.; Huh, K.B.; Cha, B.S. Visceral fat thickness measured by ultrasonography can estimate not only visceral obesity but also risks of cardiovascular and metabolic diseases. Am. J. Clin. Nutr. 2004, 79, 593–599. [Google Scholar] [CrossRef] [PubMed]
- Alvehus, M.; Burén, J.; Sjöström, M.; Goedecke, J.; Olsson, T. The human visceral fat depot has a unique inflammatory profile. Obesity 2010, 18, 879–883. [Google Scholar] [CrossRef] [PubMed]
- Nagareddy, P.R.; Kraakman, M.; Masters, S.L.; Stirzaker, R.A.; Gorman, D.J.; Grant, R.W.; Dragoljevic, D.; Hong, E.S.; Abdel-Latif, A.; Smyth, S.S.; et al. Adipose tissue macrophages promote myelopoiesis and monocytosis in obesity. Cell Metab. 2014, 19, 821–835. [Google Scholar] [CrossRef] [PubMed]
- Thrum, S.; Sommer, M.; Raulien, N.; Gericke, M.; Massier, L.; Kovacs, P.; Krasselt, M.; Landgraf, K.; Körner, A.; Dietrich, A.; et al. Macrophages in obesity are characterized by increased IL-1β response to calcium-sensing receptor signals. Int. J. Obes. 2022, 46, 1883–1891. [Google Scholar] [CrossRef]
- Curti, M.L.; Jacob, P.; Borges, M.C.; Rogero, M.M.; Ferreira, S.R. Studies of gene variants related to inflammation, oxidative stress, dyslipidemia, and obesity: Implications for a nutrigenetic approach. J. Obes. 2011, 2011, 497401. [Google Scholar] [CrossRef]
- Luo, Z.; Zhang, Y.; Li, F.; He, J.; Ding, H.; Yan, L.; Cheng, H. Resistin induces insulin resistance by both AMPK-dependent and AMPK-independent mechanisms in HepG2 cells. Endocrine 2009, 36, 60–69. [Google Scholar] [CrossRef]
- Kaser, S.; Kaser, A.; Sandhofer, A.; Ebenbichler, C.F.; Tilg, H.; Patsch, J.R. Resistin messenger-RNA expression is increased by proinflammatory cytokines in vitro. Biochem. Biophys. Res. Commun. 2003, 309, 286–290. [Google Scholar] [CrossRef]
- Silswal, N.; Singh, A.K.; Aruna, B.; Mukhopadhyay, S.; Ghosh, S.; Ehtesham, N.Z. Human resistin stimulates the pro-inflammatory cytokines TNF-alpha and IL-12 in macrophages by NF-kappaB-dependent pathway. Biochem. Biophys. Res. Commun. 2005, 334, 1092–1101. [Google Scholar] [CrossRef]
- Siddiqui, K.; Scaria Joy, S.; George, T.P. Circulating resistin levels in relation with insulin resistance, inflammatory and endothelial dysfunction markers in patients with type 2 diabetes and impaired fasting glucose. End. Metab. Sci. 2020, 3–4, 100059. [Google Scholar] [CrossRef]
- Ruiz, H.H.; Nguyen, A.; Wang, C.; He, L.; Li, H.; Hallowell, P.; McNamara, C.; Schmidt, A.M. The AGE/RAGE/DIAPH1 axis is associated with immunometabolic markers and risk of insulin resistance in subcutaneous but not omental adipose tissue in human obesity. Int. J. Obes. 2021, 45, 2083–2094. [Google Scholar] [CrossRef] [PubMed]
- Monden, M.; Koyama, H.; Otsuka, Y.; Morioka, T.; Mori, K.; Shoji, T.; Mima, Y.; Motoyama, K.; Fukumoto, S.; Shioi, A.; et al. Receptor for advanced glycation end products regulates adipocyte hypertrophy and insulin sensitivity in mice: Involvement of Toll-like receptor 2. Diabetes 2013, 62, 478–489. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.W.; Chae, H.Y.; Kwon, S.J.; Park, S.Y.; Ihm, J.; Ihm, S.H. RAGE ligands induce apoptotic cell death of pancreatic β-cells via oxidative stress. Int. J. Mol. Med. 2010, 26, 813–818. [Google Scholar] [PubMed]
- Sanchez, C.A.; Kawamura, Y.; Yamamoto, Y.; Takeshita, F.; Ochiya, T. Emerging Roles of Long Non-Coding RNA in Cancer. Cancer Sci. 2018, 109, 2093–2100. [Google Scholar] [CrossRef]
- Chrysanthou, M.; Estruch, I.M.; Rietjens, I.M.C.M.; Wichers, H.J.; Hoppenbrouwers, T. In Vitro Methodologies to Study the Role of Advanced Glycation End Products (AGEs) in Neurodegeneration. Nutrients 2022, 14, 363. [Google Scholar] [CrossRef]
- Wu, P.; Cai, J.; Chen, Q.; Han, B.; Meng, X.; Li, Y.; Li, Z.; Wang, R.; Lin, L.; Duan, C.; et al. Lnc-TALC promotes O6-methylguanine-DNA methyltransferase expression via regulating the c-Met pathway by competitively binding with miR-20b-3p. Nat. Commun. 2019, 10, 2045. [Google Scholar] [CrossRef]
- Lin, M.; Li, Y.; Xian, J.; Chen, J.; Feng, Y.; Mao, C.; Pan, Y.; Li, Z.; Zeng, Y.; Yang, L.; et al. Long non-coding RNA AGER-1 inhibits colorectal cancer progression through sponging miR-182. Int. J. Biol. Markers 2020, 35, 10–18. [Google Scholar] [CrossRef]
- Pan, Z.; Liu, L.; Nie, W.; Miggin, S.; Qiu, F.; Cao, Y.; Chen, J.; Yang, B.; Zhou, Y.; Lu, J.; et al. Long non-coding RNA AGER-1 functionally upregulates the innate immunity gene AGER and approximates its anti-tumor effect in lung cancer. Mol. Carcinog. 2018, 57, 305–318. [Google Scholar] [CrossRef]
- Koźniewski, K.; Wąsowski, M.; Jonas, M.I.; Lisik, W.; Jonas, M.; Binda, A.; Jaworski, P.; Tarnowski, W.; Noszczyk, B.; Puzianowska-Kuźnicka, M.; et al. Epigenetic Regulation of Estrogen Receptor Genes’ Expressions in Adipose Tissue in the Course of Obesity. Int. J. Mol. Sci. 2022, 23, 5989. [Google Scholar] [CrossRef]
- Navel, V.; Malecaze, J.; Belville, C.; Choltus, H.; Henrioux, F.; Dutheil, F.; Malecaze, F.; Chiambaretta, F.; Blanchon, L.; Sapin, V. Dysregulation of Receptor for Advanced Glycation End Products (RAGE) Expression as a Biomarker of Keratoconus. Dis. Markers 2022, 2022, 1543742. [Google Scholar] [CrossRef] [PubMed]
- Perneger, T.V. What’s wrong with Bonferroni adjustments. BMJ 1998, 316, 1236–1238. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, S. A farewell to Bonferroni: The problems of low statistical power and publication bias. Behav. Ecol. 2004, 15, 1044–1045. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Obese Individuals before Weight Loss (N = 62) | Obese Individuals after Weight Loss (N = 15) | Normal Weight Controls (N = 20) | ||||
|---|---|---|---|---|---|---|
| Mean ± SD | Min–Max | Mean ± SD | Min–Max | Mean ± SD | Min–Max | |
| Age (years) | 40.48 ± 10.19 | 20–59 | 40.76 ± 8.13 | 29–59 | 44.08 ± 13.5 | 26–63 |
| Weight (kg) | 125.6 ± 17.11 | 87.8–170.0 | 73.11 ± 6.25 | 66.0–82.0 | 67.92 ± 10.48 | 54.0–74.0 |
| BMI (kg/m2) | 45.97 ± 5.70 | 35.43–63.42 | 28.2 ± 2.45 | 24.0–31.2 | 23.06 ± 1.33 | 20.7–24.93 |
| Adipose tissue (% body mass) | 48.54 ± 3.65 | 40.43–57.23 | 32.5 ± 3.4 | 26.0–37.0 | - | - |
| Weight loss (kg) | - | - | 32.4 ± 8.4 | 23.0–42.0 | - | - |
| Glucose (mmol/l) | 5.91 ± 1.37 | 3.67–9.85 | 4.82 ± 0.55 | 4.12–5.66 | 5.2 ± 0.3 | 4.8–5.4 |
| HbA1c (%) | 5.79 ± 0.51 | 5.06–7.20 | 5.1 ± 0.47 | 4.59–5.65 | 4.9 ± 0.28 | 4.6–5.3 |
| Total cholesterol (mmol/l) | 4.96 ± 0.91 | 3.13–7.87 | 4.76 ± 0.66 | 3.92–5.9 | 4.31 ± 0.20 | 4.24–4.78 |
| LDL cholesterol (mmol/l) | 3.04 ± 0.97 | 1.04–5.65 | 2.99 ± 0.35 | 2.54–3.58 | 2.82 ± 0.11 | 2.74–2.90 |
| HDL cholesterol (mmol/l) | 1.21 ± 0.25 | 0.59–1.78 | 1.38 ± 0.27 | 1.08–1.68 | 1.47 ± 0.26 | 1.24–1.76 |
| Triglycerides (mmol/l) | 1.43 ± 0.72 | 0.52–3.23 | 1.40 ± 0.38 | 1.02–1.85 | 1.32 ± 0.2 | 1.1–1.46 |
| CRP (mg/l) | 10.65 ± 5.29 | 1.21–21.61 | 4.51 ± 2.45 | 1.81–7.21 | 3.24 ± 1.72 | 0.7–5.20 |
| Comorbidities | N | (%) | N | (%) | N | (%) |
| Hypertension | 31 | 50.0% | 5 | 33.3% | none | none |
| Type 2 diabetes mellitus/prediabetes * | 23 | 37.0% | 3 | 20.0% | none | none |
| Hyperlipidemia | 34 | 55.0% | 6 | 40.0% | none | none |
| Gene/lncRNA | Description | Primers | Ref. | |
|---|---|---|---|---|
| AGER | advanced glycosylation end-product receptor | F | 5′ TGTGCTGATCCTCCCTGAGA 3′ | [51] |
| R | 5′ CGAGGAGGGGCCAACTGCA 3′ | |||
| lncAGER-1 | long non-coding RNA AGER-1 | F | 5′ AACCAGGAGGAAGAGGAGGA 3′ | [48] |
| R | 5′ TTGGCAAGG TGGGGTTATAC 3′ | |||
| IL1B | interleukin 1β | F | 5′ CACCAAGCTTTTTTGCTGTGAGT3′ | [19] |
| R | 5′ GCACGATGCACCTGTACGAT 3′ | |||
| IL6 | interleukin 6 | F | 5′ CCTTCGGTCCAGTTGCCTTC 3′ | [19] |
| R | 5′ GTGGGGCGGCTACATCTTTG 3′ | |||
| IL8 | interleukin 8 | F | 5′ CACCGGAAGAACCATCTCACT 3′ | [19] |
| R | 5′ TCAGCCCTCTTCAAAAACTTCTCC 3′ | |||
| RETN | resistin | F | 5′ GCTGTTGGTGTCTAGCAAGAC 3′ | [20] |
| R | 5′ CATCATCATCATCATCTCCAG 3′ | |||
| ACTB | β-actin | F | 5′ CAGCCTGGATAGCAACGTAC 3′ | [19,20,51] |
| R | 5′ TTCTACAATGAGCTGCGTGTG 3′ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gutowska, K.; Koźniewski, K.; Wąsowski, M.; Jonas, M.I.; Bartoszewicz, Z.; Lisik, W.; Jonas, M.; Binda, A.; Jaworski, P.; Tarnowski, W.; et al. AGER-1 Long Non-Coding RNA Levels Correlate with the Expression of the Advanced Glycosylation End-Product Receptor, a Regulator of the Inflammatory Response in Visceral Adipose Tissue of Women with Obesity and Type 2 Diabetes Mellitus. Int. J. Mol. Sci. 2023, 24, 17447. https://doi.org/10.3390/ijms242417447
Gutowska K, Koźniewski K, Wąsowski M, Jonas MI, Bartoszewicz Z, Lisik W, Jonas M, Binda A, Jaworski P, Tarnowski W, et al. AGER-1 Long Non-Coding RNA Levels Correlate with the Expression of the Advanced Glycosylation End-Product Receptor, a Regulator of the Inflammatory Response in Visceral Adipose Tissue of Women with Obesity and Type 2 Diabetes Mellitus. International Journal of Molecular Sciences. 2023; 24(24):17447. https://doi.org/10.3390/ijms242417447
Chicago/Turabian StyleGutowska, Klaudia, Krzysztof Koźniewski, Michał Wąsowski, Marta Izabela Jonas, Zbigniew Bartoszewicz, Wojciech Lisik, Maurycy Jonas, Artur Binda, Paweł Jaworski, Wiesław Tarnowski, and et al. 2023. "AGER-1 Long Non-Coding RNA Levels Correlate with the Expression of the Advanced Glycosylation End-Product Receptor, a Regulator of the Inflammatory Response in Visceral Adipose Tissue of Women with Obesity and Type 2 Diabetes Mellitus" International Journal of Molecular Sciences 24, no. 24: 17447. https://doi.org/10.3390/ijms242417447