
Studying the Human Microbiota: Advances in Understanding the Fundamentals, Origin, and Evolution of Biological Timekeeping

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
- How many other human-associated microorganisms possess biological clocks?
- Is there a single, conserved primordial timekeeping mechanism, or has evolution given rise to diverse proto-circadian systems?
- What constitutes the core of cellular timing systems? Is traditional self-sustainability necessary, or can adaptive advantages be achieved with simpler mechanisms?
- How do microbes track the time inside the human intestine in the absence of light cues?
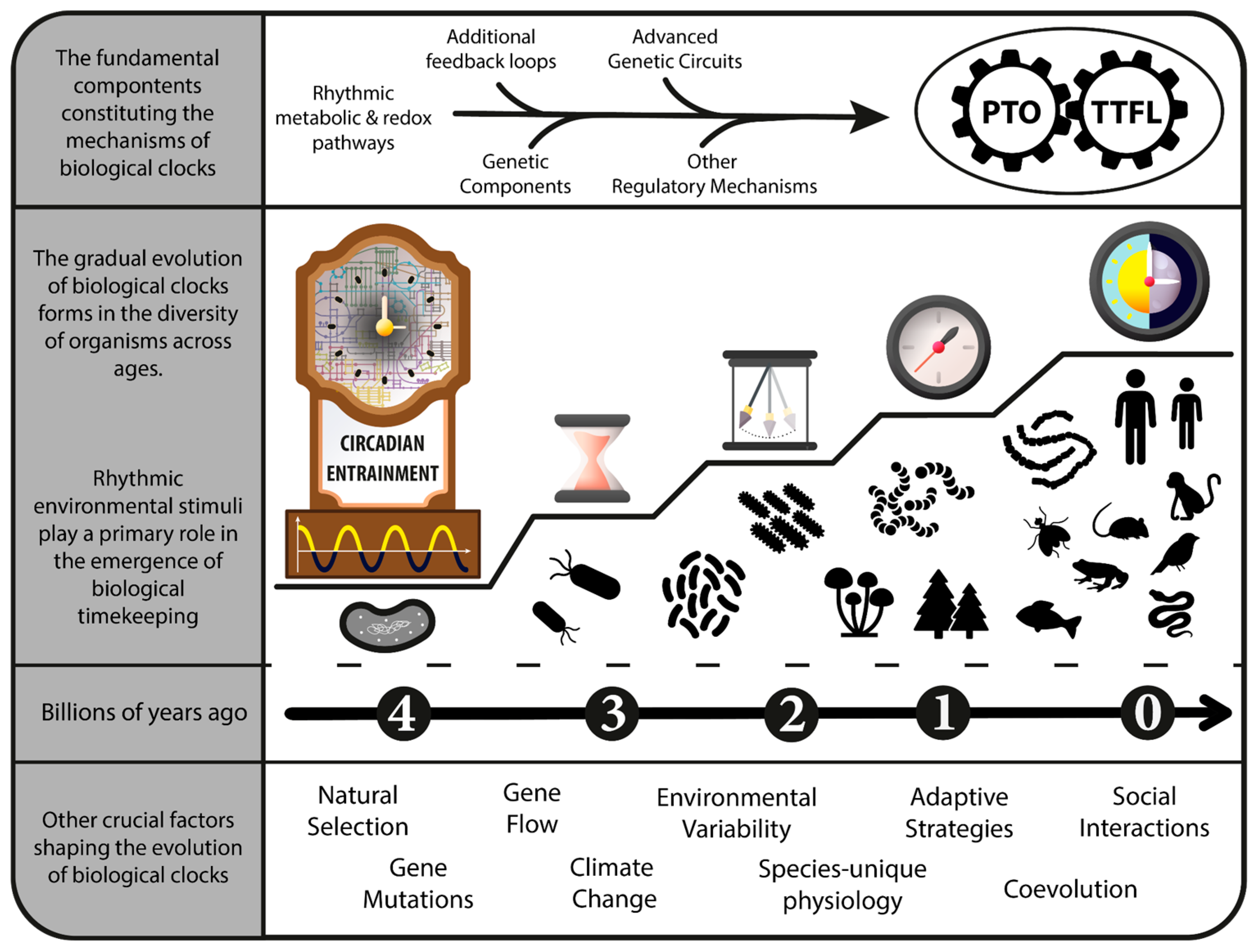
2. The Clock That Ticks the Life Rhythm
3. The Expanding World of Bacterial Biological Clocks
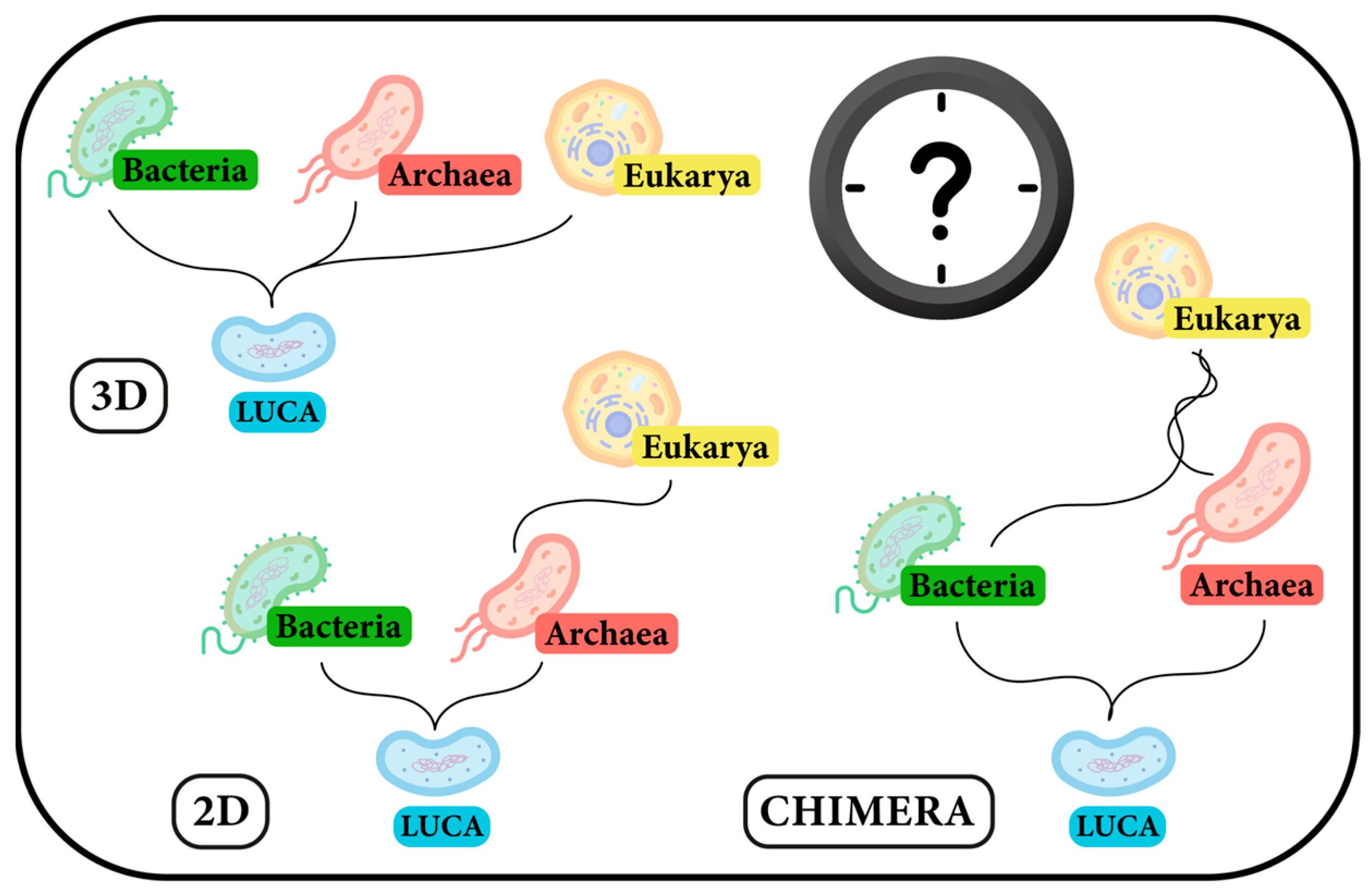
4. A Missing Line on the Evolutionary Map: Eukarya–Archaea
5. Subtle Traces of Archaeal Timekeeping Capabilities
6. Delving Deeper into Prokaryotic Oscillators Unveils the Landscape of Timing Systems
7. A Fork in the Road of Circadian Rhythms—TTFL and PTO Mechanisms
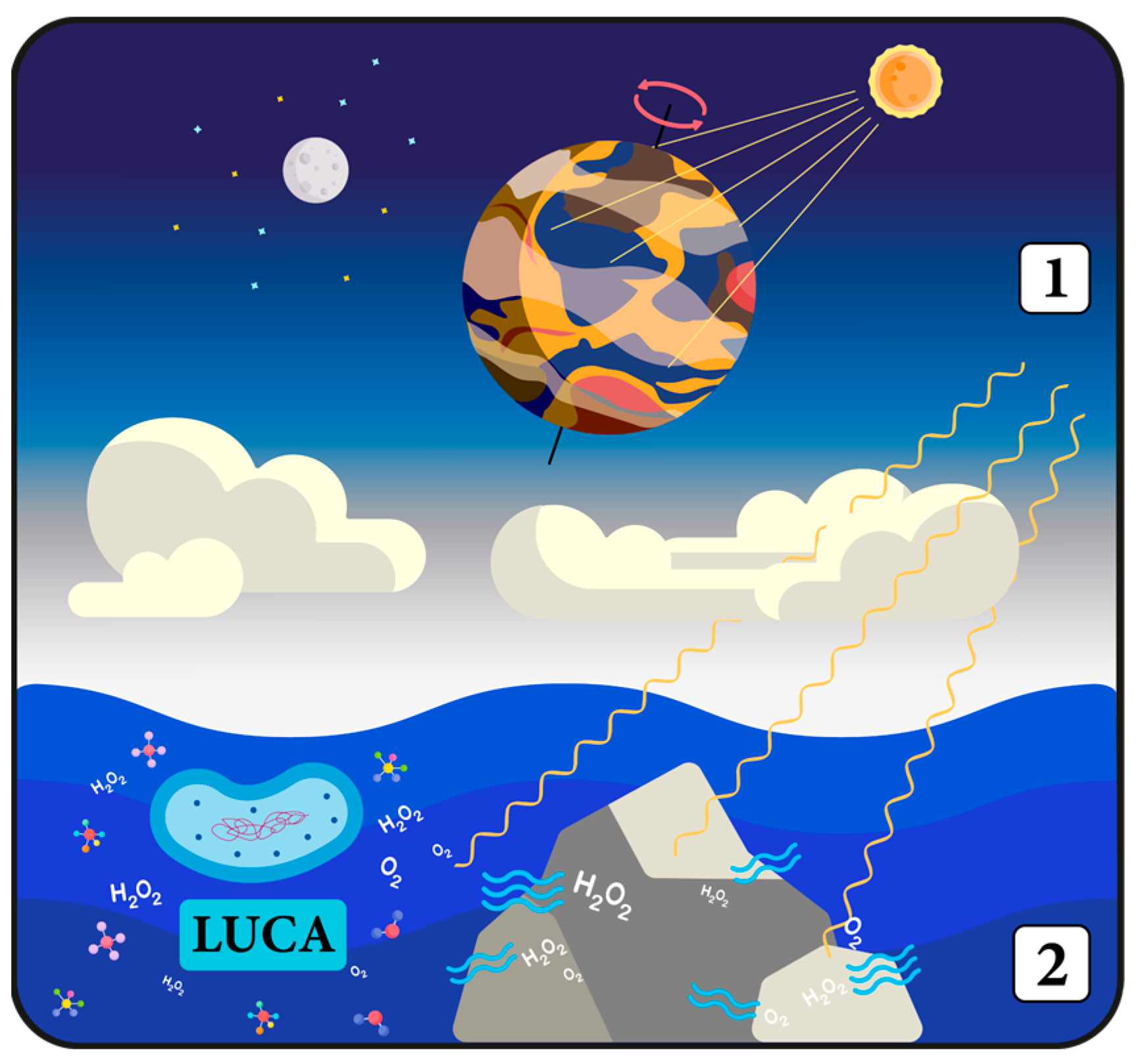
8. Why Has Evolution Decided to Start Measuring Time?
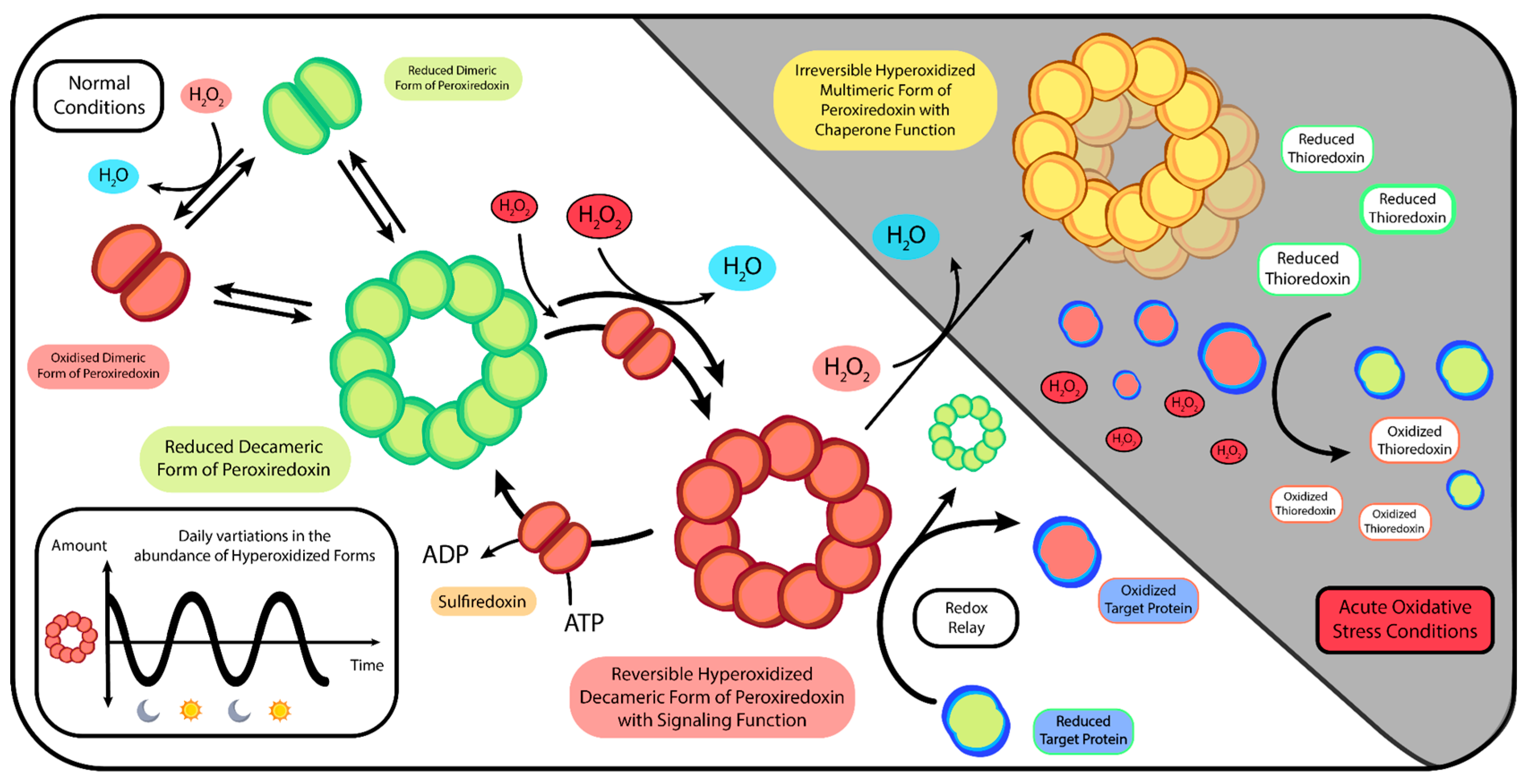
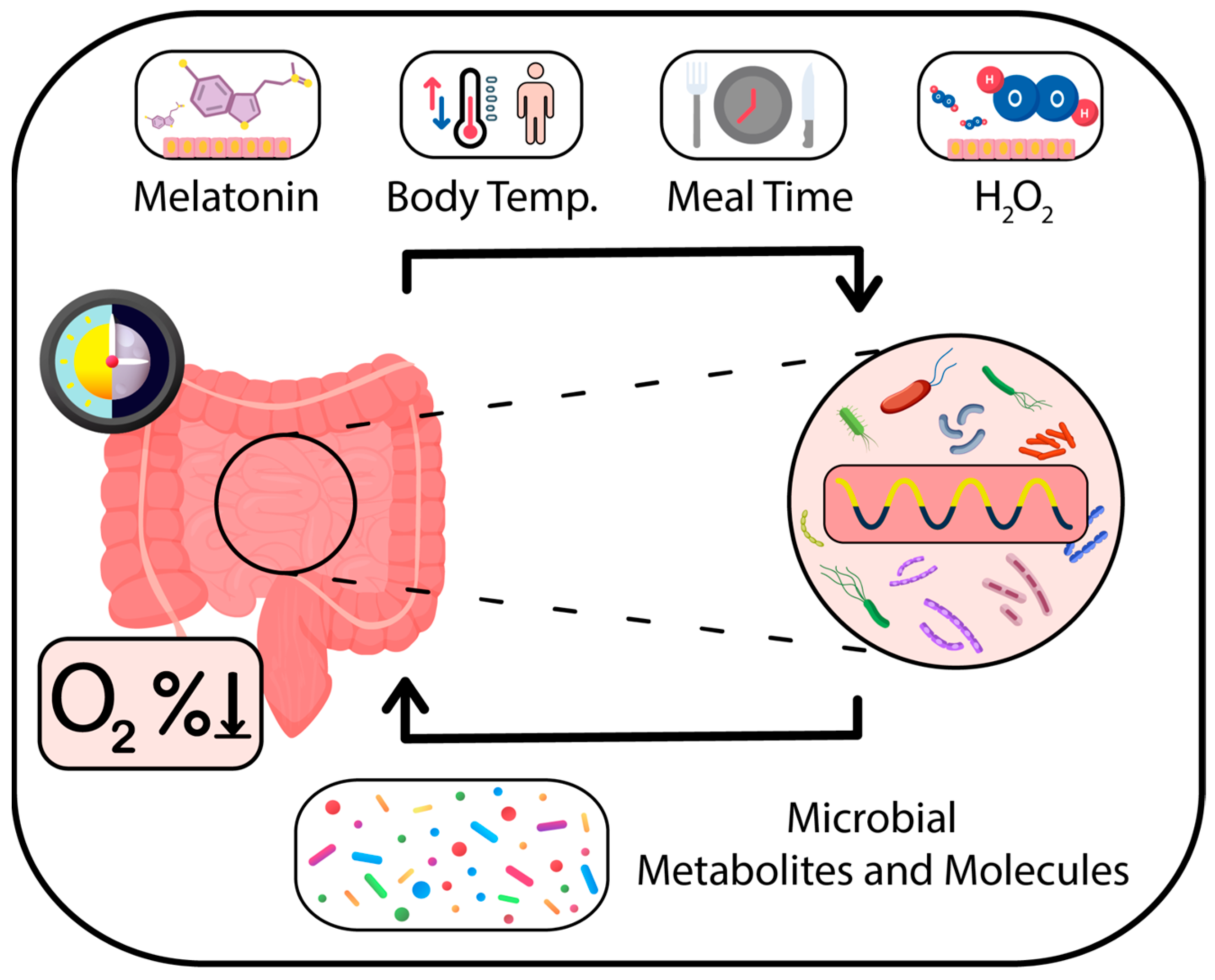
9. Bridging Redox Metabolism, Cellular Communication, and Antioxidant Function
10. Mycobiome May Be a Window into the Intricacies of Biological Timekeeping
11. The Crossroad of Metabolic and Antioxidant Functions within the Timekeeping Framework
12. Metabolic Rhythms: The Heartbeat of Circadian Timing
13. Paradigms Change around the Clock
14. Time Cues That Facilitate Rhythmic Cooperation
15. Concluding Remarks
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Simon, A.K.; Hollander, G.A.; McMichael, A. Evolution of the Immune System in Humans from Infancy to Old Age. Proc. R. Soc. B Biol. Sci. 2015, 282, 20143085. [Google Scholar] [CrossRef]
- Mueller, N.T.; Bakacs, E.; Combellick, J.; Grigoryan, Z.; Dominguez-Bello, M.G. The Infant Microbiome Development: Mom Matters. Trends Mol. Med. 2015, 21, 109–117. [Google Scholar] [CrossRef] [PubMed]
- Schroeder, B.O.; Bäckhed, F. Signals from the Gut Microbiota to Distant Organs in Physiology and Disease. Nat. Med. 2016, 22, 1079–1089. [Google Scholar] [CrossRef]
- Bienenstock, J.; Kunze, W.; Forsythe, P. Microbiota and the Gut-Brain Axis. Nutr. Rev. 2015, 73 (Suppl. 1), 28–31. [Google Scholar] [CrossRef]
- Qin, J.; Li, R.; Raes, J.; Arumugam, M.; Burgdorf, K.S.; Manichanh, C.; Nielsen, T.; Pons, N.; Levenez, F.; Yamada, T.; et al. A Human Gut Microbial Gene Catalog Established by Metagenomic Sequencing. Nature 2010, 464, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, J.A.; Blaser, M.J.; Caporaso, J.G.; Jansson, J.K.; Lynch, S.V.; Knight, R. Current Understanding of the Human Microbiome. Nat. Med. 2018, 24, 392–400. [Google Scholar] [CrossRef]
- Berg, G.; Rybakova, D.; Fischer, D.; Cernava, T.; Vergès, M.-C.C.; Charles, T.; Chen, X.; Cocolin, L.; Eversole, K.; Corral, G.H.; et al. Microbiome Definition Re-Visited: Old Concepts and New Challenges. Microbiome 2020, 8, 103. [Google Scholar] [CrossRef]
- Marchesi, J.R.; Ravel, J. The Vocabulary of Microbiome Research: A Proposal. Microbiome 2015, 3, 31. [Google Scholar] [CrossRef]
- Chen, Y.; Zhou, J.; Wang, L. Role and Mechanism of Gut Microbiota in Human Disease. Front. Cell. Infect. Microbiol. 2021, 11, 625913. [Google Scholar] [CrossRef]
- Liang, X.; FitzGerald, G.A. Timing the Microbes: The Circadian Rhythm of the Gut Microbiome. J. Biol. Rhythm. 2017, 32, 505–515. [Google Scholar] [CrossRef]
- Liang, X.; Bushman, F.D.; FitzGerald, G.A. Rhythmicity of the Intestinal Microbiota Is Regulated by Gender and the Host Circadian Clock. Proc. Natl. Acad. Sci. USA 2015, 112, 10479–10484. [Google Scholar] [CrossRef]
- Thaiss, C.A.; Zeevi, D.; Levy, M.; Zilberman-Schapira, G.; Suez, J.; Tengeler, A.C.; Abramson, L.; Katz, M.N.; Korem, T.; Zmora, N.; et al. Transkingdom Control of Microbiota Diurnal Oscillations Promotes Metabolic Homeostasis. Cell 2014, 159, 514–529. [Google Scholar] [CrossRef] [PubMed]
- Tognini, P.; Murakami, M.; Sassone-Corsi, P. Interplay between Microbes and the Circadian Clock. Cold Spring Harb. Perspect. Biol. 2018, 10, a028365. [Google Scholar] [CrossRef]
- Zarrinpar, A.; Chaix, A.; Yooseph, S.; Panda, S. Diet and Feeding Pattern Affect the Diurnal Dynamics of the Gut Microbiome. Cell Metab. 2014, 20, 1006–1017. [Google Scholar] [CrossRef] [PubMed]
- Thaiss, C.A.; Levy, M.; Korem, T.; Dohnalová, L.; Shapiro, H.; Jaitin, D.A.; David, E.; Winter, D.R.; Gury-BenAri, M.; Tatirovsky, E.; et al. Microbiota Diurnal Rhythmicity Programs Host Transcriptome Oscillations. Cell 2016, 167, 1495–1510.e12. [Google Scholar] [CrossRef] [PubMed]
- Segers, A.; Desmet, L.; Thijs, T.; Verbeke, K.; Tack, J.; Depoortere, I. The Circadian Clock Regulates the Diurnal Levels of Microbial Short-Chain Fatty Acids and Their Rhythmic Effects on Colon Contractility in Mice. Acta Physiol. Oxf. Engl. 2019, 225, e13193. [Google Scholar] [CrossRef]
- Heddes, M.; Altaha, B.; Niu, Y.; Reitmeier, S.; Kleigrewe, K.; Haller, D.; Kiessling, S. The Intestinal Clock Drives the Microbiome to Maintain Gastrointestinal Homeostasis. Nat. Commun. 2022, 13, 6068. [Google Scholar] [CrossRef]
- Voigt, R.M.; Summa, K.C.; Forsyth, C.B.; Green, S.J.; Engen, P.; Naqib, A.; Vitaterna, M.H.; Turek, F.W.; Keshavarzian, A. The Circadian Clock Mutation Promotes Intestinal Dysbiosis. Alcohol. Clin. Exp. Res. 2016, 40, 335–347. [Google Scholar] [CrossRef]
- Wang, H.; Zhang, H.; Su, Y. New Insights into the Diurnal Rhythmicity of Gut Microbiota and Its Crosstalk with Host Circadian Rhythm. Animals 2022, 12, 1677. [Google Scholar] [CrossRef]
- Murakami, M.; Tognini, P. The Circadian Clock as an Essential Molecular Link Between Host Physiology and Microorganisms. Front. Cell. Infect. Microbiol. 2020, 9, 469. [Google Scholar] [CrossRef]
- Mukherji, A.; Kobiita, A.; Ye, T.; Chambon, P. Homeostasis in Intestinal Epithelium Is Orchestrated by the Circadian Clock and Microbiota Cues Transduced by TLRs. Cell 2013, 153, 812–827. [Google Scholar] [CrossRef] [PubMed]
- Leone, V.; Gibbons, S.M.; Martinez, K.; Hutchison, A.L.; Huang, E.Y.; Cham, C.M.; Pierre, J.F.; Heneghan, A.F.; Nadimpalli, A.; Hubert, N.; et al. Effects of Diurnal Variation of Gut Microbes and High-Fat Feeding on Host Circadian Clock Function and Metabolism. Cell Host Microbe 2015, 17, 681–689. [Google Scholar] [CrossRef] [PubMed]
- Reitmeier, S.; Kiessling, S.; Clavel, T.; List, M.; Almeida, E.L.; Ghosh, T.S.; Neuhaus, K.; Grallert, H.; Linseisen, J.; Skurk, T.; et al. Arrhythmic Gut Microbiome Signatures Predict Risk of Type 2 Diabetes. Cell Host Microbe 2020, 28, 258–272.e6. [Google Scholar] [CrossRef] [PubMed]
- Kuang, Z.; Wang, Y.; Li, Y.; Ye, C.; Ruhn, K.A.; Behrendt, C.L.; Olson, E.N.; Hooper, L.V. The Intestinal Microbiota Programs Diurnal Rhythms in Host Metabolism through Histone Deacetylase 3. Science 2019, 365, 1428–1434. [Google Scholar] [CrossRef] [PubMed]
- Wollmuth, E.M.; Angert, E.R. Microbial Circadian Clocks: Host-Microbe Interplay in Diel Cycles. BMC Microbiol. 2023, 23, 124. [Google Scholar] [CrossRef]
- Henao-Mejia, J.; Strowig, T.; Flavell, R.A. Microbiota Keep the Intestinal Clock Ticking. Cell 2013, 153, 741–743. [Google Scholar] [CrossRef]
- Bishehsari, F.; Voigt, R.M.; Keshavarzian, A. Circadian Rhythms and the Gut Microbiota: From the Metabolic Syndrome to Cancer. Nat. Rev. Endocrinol. 2020, 16, 731–739. [Google Scholar] [CrossRef]
- Beli, E.; Prabakaran, S.; Krishnan, P.; Evans-Molina, C.; Grant, M.B. Loss of Diurnal Oscillatory Rhythms in Gut Microbiota Correlates with Changes in Circulating Metabolites in Type 2 Diabetic Db/Db Mice. Nutrients 2019, 11, 2310. [Google Scholar] [CrossRef]
- Parkar, S.G.; Kalsbeek, A.; Cheeseman, J.F. Potential Role for the Gut Microbiota in Modulating Host Circadian Rhythms and Metabolic Health. Microorganisms 2019, 7, 41. [Google Scholar] [CrossRef]
- Allaband, C.; Lingaraju, A.; Martino, C.; Russell, B.; Tripathi, A.; Poulsen, O.; Dantas Machado, A.C.; Zhou, D.; Xue, J.; Elijah, E.; et al. Intermittent Hypoxia and Hypercapnia Alter Diurnal Rhythms of Luminal Gut Microbiome and Metabolome. mSystems 2021, 6, e0011621. [Google Scholar] [CrossRef]
- Costantini, C.; Renga, G.; Sellitto, F.; Borghi, M.; Stincardini, C.; Pariano, M.; Zelante, T.; Chiarotti, F.; Bartoli, A.; Mosci, P.; et al. Microbes in the Era of Circadian Medicine. Front. Cell. Infect. Microbiol. 2020, 10, 30. [Google Scholar] [CrossRef] [PubMed]
- Eelderink-Chen, Z.; Bosman, J.; Sartor, F.; Dodd, A.N.; Kovács, Á.T.; Merrow, M. A Circadian Clock in a Nonphotosynthetic Prokaryote. Sci. Adv. 2021, 7, eabe2086. [Google Scholar] [CrossRef]
- Paulose, J.K.; Wright, J.M.; Patel, A.G.; Cassone, V.M. Human Gut Bacteria Are Sensitive to Melatonin and Express Endogenous Circadian Rhythmicity. PLoS ONE 2016, 11, e0146643. [Google Scholar] [CrossRef] [PubMed]
- Nutman, A.P.; Bennett, V.C.; Friend, C.R.L.; Van Kranendonk, M.J.; Chivas, A.R. Rapid Emergence of Life Shown by Discovery of 3700-Million-Year-Old Microbial Structures. Nature 2016, 537, 535–538. [Google Scholar] [CrossRef]
- Javaux, E.J. Challenges in Evidencing the Earliest Traces of Life. Nature 2019, 572, 451–460. [Google Scholar] [CrossRef]
- Bontognali, T.R.R.; Sessions, A.L.; Allwood, A.C.; Fischer, W.W.; Grotzinger, J.P.; Summons, R.E.; Eiler, J.M. Sulfur Isotopes of Organic Matter Preserved in 3.45-Billion-Year-Old Stromatolites Reveal Microbial Metabolism. Proc. Natl. Acad. Sci. USA 2012, 109, 15146–15151. [Google Scholar] [CrossRef]
- Noffke, N.; Christian, D.; Wacey, D.; Hazen, R.M. Microbially Induced Sedimentary Structures Recording an Ancient Ecosystem in the ca. 3.48 Billion-Year-Old Dresser Formation, Pilbara, Western Australia. Astrobiology 2013, 13, 1103–1124. [Google Scholar] [CrossRef]
- Betts, H.C.; Puttick, M.N.; Clark, J.W.; Williams, T.A.; Donoghue, P.C.J.; Pisani, D. Integrated Genomic and Fossil Evidence Illuminates Life’s Early Evolution and Eukaryote Origins. Nat. Ecol. Evol. 2018, 2, 1556–1562. [Google Scholar] [CrossRef]
- Tashiro, T.; Ishida, A.; Hori, M.; Igisu, M.; Koike, M.; Méjean, P.; Takahata, N.; Sano, Y.; Komiya, T. Early Trace of Life from 3.95 Ga Sedimentary Rocks in Labrador, Canada. Nature 2017, 549, 516–518. [Google Scholar] [CrossRef]
- Mahendrarajah, T.A.; Moody, E.R.R.; Schrempf, D.; Szánthó, L.L.; Dombrowski, N.; Davín, A.A.; Pisani, D.; Donoghue, P.C.J.; Szöllősi, G.J.; Williams, T.A.; et al. ATP Synthase Evolution on a Cross-Braced Dated Tree of Life. bioRxiv 2023. [Google Scholar] [CrossRef]
- West, A.C.; Bechtold, D.A. The Cost of Circadian Desynchrony: Evidence, Insights and Open Questions. Bioessays 2015, 37, 777–788. [Google Scholar] [CrossRef]
- Ouyang, Y.; Andersson, C.R.; Kondo, T.; Golden, S.S.; Johnson, C.H. Resonating Circadian Clocks Enhance Fitness in Cyanobacteria. Proc. Natl. Acad. Sci. USA 1998, 95, 8660–8664. [Google Scholar] [CrossRef] [PubMed]
- Eelderink-Chen, Z.; Olmedo, M.; Bosman, J.; Merrow, M. Using Circadian Entrainment to Find Cryptic Clocks. Methods Enzymol. 2015, 551, 73–93. [Google Scholar] [CrossRef] [PubMed]
- Bodvard, K.; Peeters, K.; Roger, F.; Romanov, N.; Igbaria, A.; Welkenhuysen, N.; Palais, G.; Reiter, W.; Toledano, M.B.; Käll, M.; et al. Light-Sensing via Hydrogen Peroxide and a Peroxiredoxin. Nat. Commun. 2017, 8, 14791. [Google Scholar] [CrossRef] [PubMed]
- Krittika, S.; Yadav, P. Circadian Clocks: An Overview on Its Adaptive Significance. Biol. Rhythm Res. 2020, 51, 1109–1132. [Google Scholar] [CrossRef]
- Vitaterna, M.H.; Takahashi, J.S.; Turek, F.W. Overview of Circadian Rhythms. Alcohol Res. Health 2001, 25, 85–93. [Google Scholar]
- Harmer, S.L.; Panda, S.; Kay, S.A. Molecular Bases of Circadian Rhythms. Annu. Rev. Cell Dev. Biol. 2001, 17, 215–253. [Google Scholar] [CrossRef]
- Paranjpe, D.A.; Sharma, V.K. Evolution of Temporal Order in Living Organisms. J. Circadian Rhythm. 2005, 3, 7. [Google Scholar] [CrossRef]
- Li, W.; Wang, Z.; Cao, J.; Dong, Y.; Chen, Y. Perfecting the Life Clock: The Journey from PTO to TTFL. Int. J. Mol. Sci. 2023, 24, 2402. [Google Scholar] [CrossRef]
- Bhadra, U.; Thakkar, N.; Das, P.; Pal Bhadra, M. Evolution of Circadian Rhythms: From Bacteria to Human. Sleep Med. 2017, 35, 49–61. [Google Scholar] [CrossRef]
- Patke, A.; Young, M.W.; Axelrod, S. Molecular Mechanisms and Physiological Importance of Circadian Rhythms. Nat. Rev. Mol. Cell Biol. 2020, 21, 67–84. [Google Scholar] [CrossRef]
- Kronfeld-Schor, N.; Visser, M.E.; Salis, L.; van Gils, J.A. Chronobiology of Interspecific Interactions in a Changing World. Philos. Trans. R. Soc. B Biol. Sci. 2017, 372, 20160248. [Google Scholar] [CrossRef] [PubMed]
- Vaze, K.M.; Sharma, V.K. On the Adaptive Significance of Circadian Clocks for Their Owners. Chronobiol. Int. 2013, 30, 413–433. [Google Scholar] [CrossRef]
- Fagiani, F.; Di Marino, D.; Romagnoli, A.; Travelli, C.; Voltan, D.; Di Cesare Mannelli, L.; Racchi, M.; Govoni, S.; Lanni, C. Molecular Regulations of Circadian Rhythm and Implications for Physiology and Diseases. Signal Transduct. Target. Ther. 2022, 7, 41. [Google Scholar] [CrossRef]
- Panda, S.; Hogenesch, J.B.; Kay, S.A. Circadian Rhythms from Flies to Human. Nature 2002, 417, 329–335. [Google Scholar] [CrossRef] [PubMed]
- Reddy, A.B.; Rey, G. Metabolic and Nontranscriptional Circadian Clocks: Eukaryotes. Annu. Rev. Biochem. 2014, 83, 165–189. [Google Scholar] [CrossRef] [PubMed]
- Sharma, V.K. Adaptive Significance of Circadian Clocks. Chronobiol. Int. 2003, 20, 901–919. [Google Scholar] [CrossRef]
- Dodd, A.N.; Salathia, N.; Hall, A.; Kévei, E.; Tóth, R.; Nagy, F.; Hibberd, J.M.; Millar, A.J.; Webb, A.A.R. Plant Circadian Clocks Increase Photosynthesis, Growth, Survival, and Competitive Advantage. Science 2005, 309, 630–633. [Google Scholar] [CrossRef] [PubMed]
- Johnson, C.H.; Golden, S.S.; Ishiura, M.; Kondo, T. Circadian Clocks in Prokaryotes. Mol. Microbiol. 1996, 21, 5–11. [Google Scholar] [CrossRef]
- Jabbur, M.L.; Johnson, C.H. Spectres of Clock Evolution: Past, Present, and Yet to Come. Front. Physiol. 2021, 12, 815847. [Google Scholar] [CrossRef]
- Cohen, S.E.; Golden, S.S. Circadian Rhythms in Cyanobacteria. Microbiol. Mol. Biol. Rev. MMBR 2015, 79, 373–385. [Google Scholar] [CrossRef] [PubMed]
- Mackey, S.R.; Golden, S.S.; Ditty, J.L. The Itty-Bitty Time Machine Genetics of the Cyanobacterial Circadian Clock. Adv. Genet. 2011, 74, 13–53. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Tsinoremas, N.F.; Johnson, C.H.; Lebedeva, N.V.; Golden, S.S.; Ishiura, M.; Kondo, T. Circadian Orchestration of Gene Expression in Cyanobacteria. Genes Dev. 1995, 9, 1469–1478. [Google Scholar] [CrossRef] [PubMed]
- Puszynska, A.M.; O’Shea, E.K. Switching of Metabolic Programs in Response to Light Availability Is an Essential Function of the Cyanobacterial Circadian Output Pathway. eLife 2017, 6, e23210. [Google Scholar] [CrossRef] [PubMed]
- Sweeney, B.M.; Borgese, M.B. A Circadian Rhythm in Cell Division in a Prokaryote, the Cyanobacterium Synechococcus Wh78031. J. Phycol. 1989, 25, 183–186. [Google Scholar] [CrossRef]
- Markson, J.S.; Piechura, J.R.; Puszynska, A.M.; O’Shea, E.K. Circadian Control of Global Gene Expression by the Cyanobacterial Master Regulator RpaA. Cell 2013, 155, 1396–1408. [Google Scholar] [CrossRef]
- Mitsui, A.; Kumazawa, S.; Takahashi, A.; Ikemoto, H.; Cao, S.; Arai, T. Strategy by Which Nitrogen-Fixing Unicellular Cyanobacteria Grow Photoautotrophically. Nature 1986, 323, 720–722. [Google Scholar] [CrossRef]
- Diamond, S.; Jun, D.; Rubin, B.E.; Golden, S.S. The Circadian Oscillator in Synechococcus Elongatus Controls Metabolite Partitioning during Diurnal Growth. Proc. Natl. Acad. Sci. USA 2015, 112, E1916–E1925. [Google Scholar] [CrossRef]
- Smith, R.M.; Williams, S.B. Circadian Rhythms in Gene Transcription Imparted by Chromosome Compaction in the Cyanobacterium Synechococcus Elongatus. Proc. Natl. Acad. Sci. USA 2006, 103, 8564–8569. [Google Scholar] [CrossRef]
- Chen, T.H.; Chen, T.L.; Hung, L.M.; Huang, T.C. Circadian Rhythm in Amino Acid Uptake by Synechococcus RF-1. Plant Physiol. 1991, 97, 55–59. [Google Scholar] [CrossRef]
- Dong, G.; Yang, Q.; Wang, Q.; Kim, Y.-I.; Wood, T.L.; Osteryoung, K.W.; van Oudenaarden, A.; Golden, S.S. Elevated ATPase Activity of KaiC Applies a Circadian Checkpoint on Cell Division in Synechococcus Elongatus. Cell 2010, 140, 529–539. [Google Scholar] [CrossRef] [PubMed]
- Dvornyk, V.; Mei, Q. Evolution of kaiA, a Key Circadian Gene of Cyanobacteria. Sci. Rep. 2021, 11, 9995. [Google Scholar] [CrossRef]
- Shultzaberger, R.K.; Boyd, J.S.; Diamond, S.; Greenspan, R.J.; Golden, S.S. Giving Time Purpose: The S. elongatus Clock in a Broader Network Context. Annu. Rev. Genet. 2015, 49, 485–505. [Google Scholar] [CrossRef] [PubMed]
- Sartor, F.; Eelderink-Chen, Z.; Aronson, B.; Bosman, J.; Hibbert, L.E.; Dodd, A.N.; Kovács, Á.T.; Merrow, M. Are There Circadian Clocks in Non-Photosynthetic Bacteria? Biology 2019, 8, 41. [Google Scholar] [CrossRef]
- Merrow, M.; Mazzotta, G.; Chen, Z.; Roenneberg, T. The Right Place at the Right Time: Regulation of Daily Timing by Phosphorylation. Genes Dev. 2006, 20, 2629–2633. [Google Scholar] [CrossRef]
- Welkie, D.G.; Rubin, B.E.; Diamond, S.; Hood, R.D.; Savage, D.F.; Golden, S.S. A Hard Day’s Night: Cyanobacteria in Diel Cycles. Trends Microbiol. 2019, 27, 231–242. [Google Scholar] [CrossRef] [PubMed]
- Ma, P.; Woelfle, M.A.; Johnson, C.H. An Evolutionary Fitness Enhancement Conferred by the Circadian System in Cyanobacteria. Chaos Solitons Fractals 2013, 50, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Woelfle, M.A.; Ouyang, Y.; Phanvijhitsiri, K.; Johnson, C.H. The Adaptive Value of Circadian Clocks: An Experimental Assessment in Cyanobacteria. Curr. Biol. CB 2004, 14, 1481–1486. [Google Scholar] [CrossRef]
- Welkie, D.G.; Rubin, B.E.; Chang, Y.-G.; Diamond, S.; Rifkin, S.A.; LiWang, A.; Golden, S.S. Genome-Wide Fitness Assessment during Diurnal Growth Reveals an Expanded Role of the Cyanobacterial Circadian Clock Protein KaiA. Proc. Natl. Acad. Sci. USA 2018, 115, E7174–E7183. [Google Scholar] [CrossRef]
- Sartor, F.; Xu, X.; Popp, T.; Dodd, A.N.; Kovács, Á.T.; Merrow, M. The Circadian Clock of the Bacterium B. Subtilis Evokes Properties of Complex, Multicellular Circadian Systems. Sci. Adv. 2023, 9, eadh1308. [Google Scholar] [CrossRef]
- Graniczkowska, K.B.; Shaffer, C.L.; Cassone, V.M. Transcriptional Effects of Melatonin on the Gut Commensal Bacterium Klebsiella Aerogenes. Genomics 2022, 114, 110321. [Google Scholar] [CrossRef]
- Paulose, J.K.; Cassone, C.V.; Graniczkowska, K.B.; Cassone, V.M. Entrainment of the Circadian Clock of the Enteric Bacterium Klebsiella Aerogenes by Temperature Cycles. iScience 2019, 19, 1202–1213. [Google Scholar] [CrossRef]
- Paulose, J.K.; Cassone, V.M. The Melatonin-Sensitive Circadian Clock of the Enteric Bacterium Enterobacter Aerogenes. Gut Microbes 2016, 7, 424–427. [Google Scholar] [CrossRef] [PubMed]
- Lefevre, M.; Racedo, S.M.; Denayrolles, M.; Ripert, G.; Desfougères, T.; Lobach, A.R.; Simon, R.; Pélerin, F.; Jüsten, P.; Urdaci, M.C. Safety Assessment of Bacillus Subtilis CU1 for Use as a Probiotic in Humans. Regul. Toxicol. Pharmacol. 2017, 83, 54–65. [Google Scholar] [CrossRef] [PubMed]
- Duanis-Assaf, D.; Steinberg, D.; Shemesh, M. Efficiency of Bacillus Subtilis Metabolism of Sugar Alcohols Governs Its Probiotic Effect against Cariogenic Streptococcus Mutans. Artif. Cells Nanomed. Biotechnol. 2020, 48, 1222–1230. [Google Scholar] [CrossRef] [PubMed]
- Piewngam, P.; Zheng, Y.; Nguyen, T.H.; Dickey, S.W.; Joo, H.-S.; Villaruz, A.E.; Glose, K.A.; Fisher, E.L.; Hunt, R.L.; Li, B.; et al. Pathogen Elimination by Probiotic Bacillus via Signaling Interference. Nature 2018, 562, 532–537. [Google Scholar] [CrossRef]
- Lefevre, M.; Racedo, S.M.; Ripert, G.; Housez, B.; Cazaubiel, M.; Maudet, C.; Jüsten, P.; Marteau, P.; Urdaci, M.C. Probiotic Strain Bacillus Subtilis CU1 Stimulates Immune System of Elderly during Common Infectious Disease Period: A Randomized, Double-Blind Placebo-Controlled Study. Immun. Ageing A 2015, 12, 24. [Google Scholar] [CrossRef]
- Álvarez-Marín, R.; Lepe, J.A.; Gasch-Blasi, O.; Rodríguez-Martínez, J.M.; Calvo-Montes, J.; Lara-Contreras, R.; Martín-Gandul, C.; Tubau-Quintano, F.; Cano-García, M.E.; Rodríguez-López, F.; et al. Clinical Characteristics and Outcome of Bacteraemia Caused by Enterobacter Cloacae and Klebsiella Aerogenes: More Similarities than Differences. J. Glob. Antimicrob. Resist. 2021, 25, 351–358. [Google Scholar] [CrossRef]
- Davin-Regli, A.; Lavigne, J.-P.; Pagès, J.-M. Enterobacter Spp.: Update on Taxonomy, Clinical Aspects, and Emerging Antimicrobial Resistance. Clin. Microbiol. Rev. 2019, 32, e00002-19. [Google Scholar] [CrossRef]
- Mirick, D.K.; Davis, S. Melatonin as a Biomarker of Circadian Dysregulation. Cancer Epidemiol. Biomark. Prev. Publ. Am. Assoc. Cancer Res. Cosponsored Am. Soc. Prev. Oncol. 2008, 17, 3306–3313. [Google Scholar] [CrossRef]
- Martinson, J.N.V.; Walk, S.T. Escherichia coli Residency in the Gut of Healthy Human Adults. EcoSal Plus 2020, 9. [Google Scholar] [CrossRef]
- Tenaillon, O.; Skurnik, D.; Picard, B.; Denamur, E. The Population Genetics of Commensal Escherichia coli. Nat. Rev. Microbiol. 2010, 8, 207–217. [Google Scholar] [CrossRef] [PubMed]
- Richter, T.K.S.; Michalski, J.M.; Zanetti, L.; Tennant, S.M.; Chen, W.H.; Rasko, D.A. Responses of the Human Gut Escherichia coli Population to Pathogen and Antibiotic Disturbances. mSystems 2018, 3, e00047-18. [Google Scholar] [CrossRef]
- Proença, J.T.; Barral, D.C.; Gordo, I. Commensal-to-Pathogen Transition: One-Single Transposon Insertion Results in Two Pathoadaptive Traits in Escherichia coli -Macrophage Interaction. Sci. Rep. 2017, 7, 4504. [Google Scholar] [CrossRef]
- Blount, Z.D. The Unexhausted Potential of E. coli. eLife 2015, 4, e05826. [Google Scholar] [CrossRef]
- Suvarna, K.; Stevenson, D.; Meganathan, R.; Hudspeth, M.E. Menaquinone (Vitamin K2) Biosynthesis: Localization and Characterization of the menA Gene from Escherichia coli. J. Bacteriol. 1998, 180, 2782–2787. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, J.G.; Roth, J.R. Evolution of Coenzyme B(12) Synthesis among Enteric Bacteria: Evidence for Loss and Reacquisition of a Multigene Complex. Genetics 1996, 142, 11–24. [Google Scholar] [CrossRef]
- Leimbach, A.; Hacker, J.; Dobrindt, U.E. Coli as an All-Rounder: The Thin Line between Commensalism and Pathogenicity. Curr. Top. Microbiol. Immunol. 2013, 358, 3–32. [Google Scholar] [CrossRef] [PubMed]
- Nowrouzian, F.; Hesselmar, B.; Saalman, R.; Strannegard, I.-L.; Aberg, N.; Wold, A.E.; Adlerberth, I. Escherichia coli in Infants’ Intestinal Microflora: Colonization Rate, Strain Turnover, and Virulence Gene Carriage. Pediatr. Res. 2003, 54, 8–14. [Google Scholar] [CrossRef]
- Hewitt, O.H.; Shaikh, H.M. The Rhythm of Many: Biological Rhythms in the Marine Environment, From Macro-Scale Planktonic Ecosystems to Micro-Scale Holobionts. Front. Mar. Sci. 2021, 8, 744169. [Google Scholar] [CrossRef]
- Rogers, L.A.; Greenbank, G.R. The Intermittent Growth of Bacterial Cultures. J. Bacteriol. 1930, 19, 181–190. [Google Scholar] [CrossRef] [PubMed]
- Halberg, F.; Conner, R. Circadian Organization and Microbiology: Variance Spectra and a Periodogram on Behavior of Escherichia coli Growing in Fluid Culture. J. Minn. Acad. Sci. 1961, 29, 227–239. [Google Scholar]
- Chen, A.H.; Lubkowicz, D.; Yeong, V.; Chang, R.L.; Silver, P.A. Transplantability of a Circadian Clock to a Noncircadian Organism. Sci. Adv. 2015, 1, e1500358. [Google Scholar] [CrossRef] [PubMed]
- Diallo, A.B.; Mezouar, S.; Boumaza, A.; Fiammingo, O.; Coiffard, B.; Pontarotti, P.; Desnues, B.; Mege, J.-L. RadA, a Key Gene of the Circadian Rhythm of Escherichia coli. Int. J. Mol. Sci. 2022, 23, 6136. [Google Scholar] [CrossRef]
- Kovač, U.; Žužek, Z.; Raspor Dall’Olio, L.; Pohar, K.; Ihan, A.; Moškon, M.; Rozman, D.; Starčič Erjavec, M. Escherichia coli Affects Expression of Circadian Clock Genes in Human Hepatoma Cells. Microorganisms 2021, 9, 869. [Google Scholar] [CrossRef]
- Coiffard, B.; Diallo, A.B.; Culver, A.; Mezouar, S.; Hammad, E.; Vigne, C.; Nicolino-Brunet, C.; Dignat-George, F.; Baumstarck, K.; Boucekine, M.; et al. Circadian Rhythm Disruption and Sepsis in Severe Trauma Patients. Shock Augusta Ga 2019, 52, 29–36. [Google Scholar] [CrossRef]
- Global Burden of Bacterial Antimicrobial Resistance in 2019: A Systematic Analysis. Lancet Lond. Engl. 2022, 399, 629–655. [CrossRef]
- Yang, J.; Pu, J.; Lu, S.; Bai, X.; Wu, Y.; Jin, D.; Cheng, Y.; Zhang, G.; Zhu, W.; Luo, X.; et al. Species-Level Analysis of Human Gut Microbiota With Metataxonomics. Front. Microbiol. 2020, 11, 2029. [Google Scholar] [CrossRef] [PubMed]
- Gómez, M.; Valverde, A.; del Campo, R.; Rodríguez, J.M.; Maldonado-Barragán, A. Phenotypic and Molecular Characterization of Commensal, Community-Acquired and Nosocomial Klebsiella Spp. Microorganisms 2021, 9, 2344. [Google Scholar] [CrossRef]
- Kang, F.; Chai, Z.; Li, B.; Hu, M.; Yang, Z.; Wang, X.; Liu, W.; Ren, H.; Jin, Y.; Yue, J. Characterization and Diversity of Klebsiella Pneumoniae Prophages. Int. J. Mol. Sci. 2023, 24, 9116. [Google Scholar] [CrossRef]
- Calfee, D.P. Recent Advances in the Understanding and Management of Klebsiella Pneumoniae. F1000Research 2017, 6, 1760. [Google Scholar] [CrossRef]
- Sturtevant, R.P. Circadian Variability in Klebsiella Demonstrated by Cosinor Analysis. Int. J. Chronobiol. 1973, 1, 141–146. [Google Scholar] [PubMed]
- Setiawan, A.; Widodo, A.D.W.; Endraswari, P.D. Comparison of Ciprofloxacin, Cotrimoxazole, and Doxycycline on Klebsiella Pneumoniae: Time-Kill Curve Analysis. Ann. Med. Surg. 2022, 84, 104841. [Google Scholar] [CrossRef] [PubMed]
- Munoz-Price, L.S.; Poirel, L.; Bonomo, R.A.; Schwaber, M.J.; Daikos, G.L.; Cormican, M.; Cornaglia, G.; Garau, J.; Gniadkowski, M.; Hayden, M.K.; et al. Clinical Epidemiology of the Global Expansion of Klebsiella Pneumoniae Carbapenemases. Lancet Infect. Dis. 2013, 13, 785–796. [Google Scholar] [CrossRef]
- Chen, L. Notes from the Field: Pan-Resistant New Delhi Metallo-Beta-Lactamase-Producing Klebsiella Pneumoniae—Washoe County, Nevada, 2016. MMWR Morb. Mortal. Wkly. Rep. 2017, 66, 33. [Google Scholar] [CrossRef]
- Ventola, C.L. The Antibiotic Resistance Crisis: Part 1: Causes and Threats. P T Peer-Rev. J. Formul. Manag. 2015, 40, 277–283. [Google Scholar]
- Llor, C.; Bjerrum, L. Antimicrobial Resistance: Risk Associated with Antibiotic Overuse and Initiatives to Reduce the Problem. Ther. Adv. Drug Saf. 2014, 5, 229–241. [Google Scholar] [CrossRef] [PubMed]
- Spang, A.; Mahendrarajah, T.A.; Offre, P.; Stairs, C.W. Evolving Perspective on the Origin and Diversification of Cellular Life and the Virosphere. Genome Biol. Evol. 2022, 14, evac034. [Google Scholar] [CrossRef]
- Da Cunha, V.; Gaïa, M.; Forterre, P. The Expanding Asgard Archaea and Their Elusive Relationships with Eukarya. mLife 2022, 1, 3–12. [Google Scholar] [CrossRef]
- Eme, L.; Spang, A.; Lombard, J.; Stairs, C.W.; Ettema, T.J.G. Archaea and the Origin of Eukaryotes. Nat. Rev. Microbiol. 2017, 15, 711–723. [Google Scholar] [CrossRef]
- Zhou, Z.; Liu, Y.; Li, M.; Gu, J.-D. Two or Three Domains: A New View of Tree of Life in the Genomics Era. Appl. Microbiol. Biotechnol. 2018, 102, 3049–3058. [Google Scholar] [CrossRef] [PubMed]
- Williams, T.A.; Embley, T.M. Changing Ideas about Eukaryotic Origins. Philos. Trans. R. Soc. B Biol. Sci. 2015, 370, 20140318. [Google Scholar] [CrossRef] [PubMed]
- Williams, T.A.; Foster, P.G.; Cox, C.J.; Embley, T.M. An Archaeal Origin of Eukaryotes Supports Only Two Primary Domains of Life. Nature 2013, 504, 231–236. [Google Scholar] [CrossRef] [PubMed]
- Williams, T.A.; Cox, C.J.; Foster, P.G.; Szöllősi, G.J.; Embley, T.M. Phylogenomics Provides Robust Support for a Two-Domains Tree of Life. Nat. Ecol. Evol. 2020, 4, 138–147. [Google Scholar] [CrossRef] [PubMed]
- Tomkins, J. Information Processing Differences between Archaea and Eukaraya—Implications for Homologs and the Myth of Eukaryogenesis. Answers Res. J. 2015, 8, 121–141. [Google Scholar]
- Nobs, S.-J.; MacLeod, F.I.; Wong, H.L.; Burns, B.P. Eukarya the Chimera: Eukaryotes, a Secondary Innovation of the Two Domains of Life? Trends Microbiol. 2022, 30, 421–431. [Google Scholar] [CrossRef]
- Spang, A.; Caceres, E.F.; Ettema, T.J.G. Genomic Exploration of the Diversity, Ecology, and Evolution of the Archaeal Domain of Life. Science 2017, 357, eaaf3883. [Google Scholar] [CrossRef]
- Spang, A.; Saw, J.H.; Jørgensen, S.L.; Zaremba-Niedzwiedzka, K.; Martijn, J.; Lind, A.E.; van Eijk, R.; Schleper, C.; Guy, L.; Ettema, T.J.G. Complex Archaea That Bridge the Gap between Prokaryotes and Eukaryotes. Nature 2015, 521, 173–179. [Google Scholar] [CrossRef]
- Caetano-Anollés, G.; Mughal, F. The Tree of Life Describes a Tripartite Cellular World: Neglected Support from Genome Structure and Codon Usage and the Fallacy of Alignment-Dependent Phylogenetic Interpretations. BioEssays News Rev. Mol. Cell. Dev. Biol. 2021, 43, e2100130. [Google Scholar] [CrossRef]
- Cunha, V.D.; Gaia, M.; Nasir, A.; Forterre, P. Asgard Archaea Do Not Close the Debate about the Universal Tree of Life Topology. PLoS Genet. 2018, 14, e1007215. [Google Scholar] [CrossRef]
- Jüttner, M.; Ferreira-Cerca, S. Looking through the Lens of the Ribosome Biogenesis Evolutionary History: Possible Implications for Archaeal Phylogeny and Eukaryogenesis. Mol. Biol. Evol. 2022, 39, msac054. [Google Scholar] [CrossRef] [PubMed]
- Fournier, G.P.; Poole, A.M. A Briefly Argued Case That Asgard Archaea Are Part of the Eukaryote Tree. Front. Microbiol. 2018, 9, 1896. [Google Scholar] [CrossRef] [PubMed]
- Doolittle, W.F. Evolution: Two Domains of Life or Three? Curr. Biol. 2020, 30, R177–R179. [Google Scholar] [CrossRef] [PubMed]
- Imachi, H.; Nobu, M.K.; Nakahara, N.; Morono, Y.; Ogawara, M.; Takaki, Y.; Takano, Y.; Uematsu, K.; Ikuta, T.; Ito, M.; et al. Isolation of an Archaeon at the Prokaryote-Eukaryote Interface. Nature 2020, 577, 519–525. [Google Scholar] [CrossRef]
- Zhou, Z.; Liu, Y.; Anantharaman, K.; Li, M. The Expanding Asgard Archaea Invoke Novel Insights into Tree of Life and Eukaryogenesis. mLife 2022, 1, 374–381. [Google Scholar] [CrossRef]
- Liu, Y.; Makarova, K.S.; Huang, W.-C.; Wolf, Y.I.; Nikolskaya, A.N.; Zhang, X.; Cai, M.; Zhang, C.-J.; Xu, W.; Luo, Z.; et al. Expanded Diversity of Asgard Archaea and Their Relationships with Eukaryotes. Nature 2021, 593, 553–557. [Google Scholar] [CrossRef]
- Zaremba-Niedzwiedzka, K.; Caceres, E.F.; Saw, J.H.; Bäckström, D.; Juzokaite, L.; Vancaester, E.; Seitz, K.W.; Anantharaman, K.; Starnawski, P.; Kjeldsen, K.U.; et al. Asgard Archaea Illuminate the Origin of Eukaryotic Cellular Complexity. Nature 2017, 541, 353–358. [Google Scholar] [CrossRef]
- Spang, A.; Eme, L.; Saw, J.H.; Caceres, E.F.; Zaremba-Niedzwiedzka, K.; Lombard, J.; Guy, L.; Ettema, T.J.G. Asgard Archaea Are the Closest Prokaryotic Relatives of Eukaryotes. PLoS Genet. 2018, 14, e1007080. [Google Scholar] [CrossRef]
- Heinz, E.; Domman, D. Reshaping the Tree of Life. Nat. Rev. Microbiol. 2017, 15, 322. [Google Scholar] [CrossRef]
- McInerney, J.O.; O’Connell, M.J.; Pisani, D. The Hybrid Nature of the Eukaryota and a Consilient View of Life on Earth. Nat. Rev. Microbiol. 2014, 12, 449–455. [Google Scholar] [CrossRef]
- Maniscalco, M.; Nannen, J.; Sodi, V.; Silver, G.; Lowrey, P.L.; Bidle, K.A. Light-Dependent Expression of Four Cryptic Archaeal Circadian Gene Homologs. Front. Microbiol. 2014, 5, 79. [Google Scholar] [CrossRef] [PubMed]
- Whitehead, K.; Pan, M.; Masumura, K.; Bonneau, R.; Baliga, N.S. Diurnally Entrained Anticipatory Behavior in Archaea. PLoS ONE 2009, 4, e5485. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Whon, T.W.; Lim, M.Y.; Kim, Y.B.; Kim, N.; Kwon, M.-S.; Kim, J.; Lee, S.H.; Choi, H.-J.; Nam, I.-H.; et al. The Human Gut Archaeome: Identification of Diverse Haloarchaea in Korean Subjects. Microbiome 2020, 8, 114. [Google Scholar] [CrossRef]
- Lepp, P.W.; Brinig, M.M.; Ouverney, C.C.; Palm, K.; Armitage, G.C.; Relman, D.A. Methanogenic Archaea and Human Periodontal Disease. Proc. Natl. Acad. Sci. USA 2004, 101, 6176–6181. [Google Scholar] [CrossRef] [PubMed]
- DiBaise, J.K.; Zhang, H.; Crowell, M.D.; Krajmalnik-Brown, R.; Decker, G.A.; Rittmann, B.E. Gut Microbiota and Its Possible Relationship With Obesity. Mayo Clin. Proc. 2008, 83, 460–469. [Google Scholar] [CrossRef] [PubMed]
- Baker, B.J.; De Anda, V.; Seitz, K.W.; Dombrowski, N.; Santoro, A.E.; Lloyd, K.G. Diversity, Ecology and Evolution of Archaea. Nat. Microbiol. 2020, 5, 887–900. [Google Scholar] [CrossRef]
- Bang, C.; Schmitz, R.A. Archaea Associated with Human Surfaces: Not to Be Underestimated. FEMS Microbiol. Rev. 2015, 39, 631–648. [Google Scholar] [CrossRef]
- Chaudhary, P.P.; Conway, P.L.; Schlundt, J. Methanogens in Humans: Potentially Beneficial or Harmful for Health. Appl. Microbiol. Biotechnol. 2018, 102, 3095–3104. [Google Scholar] [CrossRef]
- Zhu, Q.; Mai, U.; Pfeiffer, W.; Janssen, S.; Asnicar, F.; Sanders, J.G.; Belda-Ferre, P.; Al-Ghalith, G.A.; Kopylova, E.; McDonald, D.; et al. Phylogenomics of 10,575 Genomes Reveals Evolutionary Proximity between Domains Bacteria and Archaea. Nat. Commun. 2019, 10, 5477. [Google Scholar] [CrossRef]
- Forterre, P. Carl Woese: Still Ahead of Our Time. mLife 2022, 1, 359–367. [Google Scholar] [CrossRef]
- Martinez-Gutierrez, C.A.; Aylward, F.O. Phylogenetic Signal, Congruence, and Uncertainty across Bacteria and Archaea. Mol. Biol. Evol. 2021, 38, 5514–5527. [Google Scholar] [CrossRef]
- Moody, E.R.R.; Mahendrarajah, T.A.; Dombrowski, N.; Clark, J.W.; Petitjean, C.; Offre, P.; Szöllősi, G.J.; Spang, A.; Williams, T.A. An Estimate of the Deepest Branches of the Tree of Life from Ancient Vertically Evolving Genes. eLife 2022, 11, e66695. [Google Scholar] [CrossRef]
- Ming, H.; Miyazono, K.; Tanokura, M. Cloning, Expression, Purification, Crystallization and Preliminary Crystallographic Analysis of Selenomethionine-Labelled KaiC-like Protein PH0186 from Pyrococcus Horikoshii OT3. Acta Crystallograph. Sect. F Struct. Biol. Cryst. Commun. 2007, 63, 327–329. [Google Scholar] [CrossRef]
- Schmelling, N.M.; Lehmann, R.; Chaudhury, P.; Beck, C.; Albers, S.-V.; Axmann, I.M.; Wiegard, A. Minimal Tool Set for a Prokaryotic Circadian Clock. BMC Evol. Biol. 2017, 17, 169. [Google Scholar] [CrossRef]
- Leipe, D.D.; Aravind, L.; Grishin, N.V.; Koonin, E.V. The Bacterial Replicative Helicase DnaB Evolved from a RecA Duplication. Genome Res. 2000, 10, 5–16. [Google Scholar] [PubMed]
- Dvornyk, V.; Knudsen, B. Functional Divergence of the Circadian Clock Proteins in Prokaryotes. Genetica 2005, 124, 247–254. [Google Scholar] [CrossRef] [PubMed]
- Loza-Correa, M.; Sahr, T.; Rolando, M.; Daniels, C.; Petit, P.; Skarina, T.; Gomez Valero, L.; Dervins-Ravault, D.; Honoré, N.; Savchenko, A.; et al. The Legionella Pneumophila Kai Operon Is Implicated in Stress Response and Confers Fitness in Competitive Environments. Environ. Microbiol. 2014, 16, 359–381. [Google Scholar] [CrossRef] [PubMed]
- Makarova, K.S.; Galperin, M.Y.; Koonin, E.V. Proposed Role for KaiC-Like ATPases as Major Signal Transduction Hubs in Archaea. mBio 2017, 8, e01959-17. [Google Scholar] [CrossRef]
- de Sousa Machado, J.N.; Vollmar, L.; Schimpf, J.; Chaudhury, P.; Kumariya, R.; van der Does, C.; Hugel, T.; Albers, S.-V. Autophosphorylation of the KaiC-like Protein ArlH Inhibits Oligomerization and Interaction with ArlI, the Motor ATPase of the Archaellum. Mol. Microbiol. 2021, 116, 943–956. [Google Scholar] [CrossRef]
- Dvornyk, V.; Vinogradova, O.; Nevo, E. Origin and Evolution of Circadian Clock Genes in Prokaryotes. Proc. Natl. Acad. Sci. USA 2003, 100, 2495–2500. [Google Scholar] [CrossRef]
- Koonin, E.V.; Wolf, Y.I. Genomics of Bacteria and Archaea: The Emerging Dynamic View of the Prokaryotic World. Nucleic Acids Res. 2008, 36, 6688–6719. [Google Scholar] [CrossRef]
- Kawamoto, N.; Ito, H.; Tokuda, I.T.; Iwasaki, H. Damped Circadian Oscillation in the Absence of KaiA in Synechococcus. Nat. Commun. 2020, 11, 2242. [Google Scholar] [CrossRef] [PubMed]
- Gribaldo, S.; Brochier-Armanet, C. The Origin and Evolution of Archaea: A State of the Art. Philos. Trans. R. Soc. B Biol. Sci. 2006, 361, 1007–1022. [Google Scholar] [CrossRef] [PubMed]
- Johnson, C.H.; Zhao, C.; Xu, Y.; Mori, T. Timing the Day: What Makes Bacterial Clocks Tick? Nat. Rev. Microbiol. 2017, 15, 232–242. [Google Scholar] [CrossRef] [PubMed]
- Ma, P.; Mori, T.; Zhao, C.; Thiel, T.; Johnson, C.H. Evolution of KaiC-Dependent Timekeepers: A Proto-Circadian Timing Mechanism Confers Adaptive Fitness in the Purple Bacterium Rhodopseudomonas Palustris. PLoS Genet. 2016, 12, e1005922. [Google Scholar] [CrossRef]
- Loza-Correa, M.; Gomez-Valero, L.; Buchrieser, C. Circadian Clock Proteins in Prokaryotes: Hidden Rhythms? Front. Microbiol. 2010, 1, 130. [Google Scholar] [CrossRef] [PubMed]
- Min, H.; Guo, H.; Xiong, J. Rhythmic Gene Expression in a Purple Photosynthetic Bacterium, Rhodobacter Sphaeroides. FEBS Lett. 2005, 579, 808–812. [Google Scholar] [CrossRef]
- Zinser, E.R.; Lindell, D.; Johnson, Z.I.; Futschik, M.E.; Steglich, C.; Coleman, M.L.; Wright, M.A.; Rector, T.; Steen, R.; McNulty, N.; et al. Choreography of the Transcriptome, Photophysiology, and Cell Cycle of a Minimal Photoautotroph, Prochlorococcus. PLoS ONE 2009, 4, e5135. [Google Scholar] [CrossRef]
- Rocap, G.; Larimer, F.W.; Lamerdin, J.; Malfatti, S.; Chain, P.; Ahlgren, N.A.; Arellano, A.; Coleman, M.; Hauser, L.; Hess, W.R.; et al. Genome Divergence in Two Prochlorococcus Ecotypes Reflects Oceanic Niche Differentiation. Nature 2003, 424, 1042–1047. [Google Scholar] [CrossRef]
- Mullineaux, C.W.; Stanewsky, R. The Rolex and the Hourglass: A Simplified Circadian Clock in Prochlorococcus? J. Bacteriol. 2009, 191, 5333–5335. [Google Scholar] [CrossRef]
- Seki, M.; Ito, H. Evolution of Self-Sustained Circadian Rhythms Is Facilitated by Seasonal Change of Daylight. Proc. R. Soc. B Biol. Sci. 2022, 289, 20220577. [Google Scholar] [CrossRef] [PubMed]
- Gardner, M.J.; Hubbard, K.E.; Hotta, C.T.; Dodd, A.N.; Webb, A.A.R. How Plants Tell the Time. Biochem. J. 2006, 397, 15–24. [Google Scholar] [CrossRef] [PubMed]
- Oravec, M.W.; Greenham, K. The Adaptive Nature of the Plant Circadian Clock in Natural Environments. Plant Physiol. 2022, 190, 968–980. [Google Scholar] [CrossRef] [PubMed]
- Brody, S. Circadian Rhythms in Fungi: Structure/Function/Evolution of Some Clock Components. J. Biol. Rhythm. 2019, 34, 364–379. [Google Scholar] [CrossRef] [PubMed]
- Reppert, S.M.; Weaver, D.R. Coordination of Circadian Timing in Mammals. Nature 2002, 418, 935–941. [Google Scholar] [CrossRef]
- Hurley, J.M.; Loros, J.J.; Dunlap, J.C. Circadian Oscillators: Around the Transcription-Translation Feedback Loop and on to Output. Trends Biochem. Sci. 2016, 41, 834–846. [Google Scholar] [CrossRef]
- Hardin, P.E.; Hall, J.C.; Rosbash, M. Circadian Oscillations in Period Gene mRNA Levels Are Transcriptionally Regulated. Proc. Natl. Acad. Sci. USA 1992, 89, 11711–11715. [Google Scholar] [CrossRef]
- Milev, N.B.; Rhee, S.-G.; Reddy, A.B. Cellular Timekeeping: It’s Redox o’Clock. Cold Spring Harb. Perspect. Biol. 2018, 10, a027698. [Google Scholar] [CrossRef]
- Wulund, L.; Reddy, A.B. A Brief History of Circadian Time: The Emergence of Redox Oscillations as a Novel Component of Biological Rhythms. Perspect. Sci. 2015, 6, 27–37. [Google Scholar] [CrossRef]
- Young, M.W.; Kay, S.A. Time Zones: A Comparative Genetics of Circadian Clocks. Nat. Rev. Genet. 2001, 2, 702–715. [Google Scholar] [CrossRef]
- Shalit-Kaneh, A.; Kumimoto, R.W.; Filkov, V.; Harmer, S.L. Multiple Feedback Loops of the Arabidopsis Circadian Clock Provide Rhythmic Robustness across Environmental Conditions. Proc. Natl. Acad. Sci. USA 2018, 115, 7147–7152. [Google Scholar] [CrossRef]
- Rosbash, M. The Implications of Multiple Circadian Clock Origins. PLoS Biol. 2009, 7, e1000062. [Google Scholar] [CrossRef] [PubMed]
- Nakahira, Y.; Katayama, M.; Miyashita, H.; Kutsuna, S.; Iwasaki, H.; Oyama, T.; Kondo, T. Global Gene Repression by KaiC as a Master Process of Prokaryotic Circadian System. Proc. Natl. Acad. Sci. USA 2004, 101, 881–885. [Google Scholar] [CrossRef]
- Ishiura, M.; Kutsuna, S.; Aoki, S.; Iwasaki, H.; Andersson, C.R.; Tanabe, A.; Golden, S.S.; Johnson, C.H.; Kondo, T. Expression of a Gene Cluster kaiABC as a Circadian Feedback Process in Cyanobacteria. Science 1998, 281, 1519–1523. [Google Scholar] [CrossRef] [PubMed]
- Qin, X.; Byrne, M.; Xu, Y.; Mori, T.; Johnson, C.H. Coupling of a Core Post-Translational Pacemaker to a Slave Transcription/Translation Feedback Loop in a Circadian System. PLoS Biol. 2010, 8, e1000394. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, M.; Imai, K.; Ito, H.; Nishiwaki, T.; Murayama, Y.; Iwasaki, H.; Oyama, T.; Kondo, T. Reconstitution of Circadian Oscillation of Cyanobacterial KaiC Phosphorylation In Vitro. Science 2005, 308, 414–415. [Google Scholar] [CrossRef]
- Pattanayek, R.; Wang, J.; Mori, T.; Xu, Y.; Johnson, C.H.; Egli, M. Visualizing a Circadian Clock Protein: Crystal Structure of KaiC and Functional Insights. Mol. Cell 2004, 15, 375–388. [Google Scholar] [CrossRef]
- Tomita, J.; Nakajima, M.; Kondo, T.; Iwasaki, H. No Transcription-Translation Feedback in Circadian Rhythm of KaiC Phosphorylation. Science 2005, 307, 251–254. [Google Scholar] [CrossRef]
- Abe, J.; Hiyama, T.B.; Mukaiyama, A.; Son, S.; Mori, T.; Saito, S.; Osako, M.; Wolanin, J.; Yamashita, E.; Kondo, T.; et al. Circadian Rhythms. Atomic-Scale Origins of Slowness in the Cyanobacterial Circadian Clock. Science 2015, 349, 312–316. [Google Scholar] [CrossRef]
- O’Neill, J.S.; Reddy, A.B. Circadian Clocks in Human Red Blood Cells. Nature 2011, 469, 498–503. [Google Scholar] [CrossRef]
- O’Neill, J.S.; van Ooijen, G.; Dixon, L.E.; Troein, C.; Corellou, F.; Bouget, F.-Y.; Reddy, A.B.; Millar, A.J. Circadian Rhythms Persist without Transcription in a Eukaryote. Nature 2011, 469, 554–558. [Google Scholar] [CrossRef] [PubMed]
- del Olmo, M.; Kramer, A.; Herzel, H. A Robust Model for Circadian Redox Oscillations. Int. J. Mol. Sci. 2019, 20, 2368. [Google Scholar] [CrossRef]
- Rhee, S.G. Overview on Peroxiredoxin. Mol. Cells 2016, 39, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Perkins, A.; Nelson, K.J.; Parsonage, D.; Poole, L.B.; Karplus, P.A. Peroxiredoxins: Guardians against Oxidative Stress and Modulators of Peroxide Signaling. Trends Biochem. Sci. 2015, 40, 435–445. [Google Scholar] [CrossRef] [PubMed]
- Zeida, A.; Trujillo, M.; Ferrer-Sueta, G.; Denicola, A.; Estrin, D.A.; Radi, R. Catalysis of Peroxide Reduction by Fast Reacting Protein Thiols. Chem. Rev. 2019, 119, 10829–10855. [Google Scholar] [CrossRef]
- Toledano, M.B.; Huang, B. Microbial 2-Cys Peroxiredoxins: Insights into Their Complex Physiological Roles. Mol. Cells 2016, 39, 31–39. [Google Scholar] [CrossRef]
- Wood, Z.A.; Schröder, E.; Robin Harris, J.; Poole, L.B. Structure, Mechanism and Regulation of Peroxiredoxins. Trends Biochem. Sci. 2003, 28, 32–40. [Google Scholar] [CrossRef]
- Nelson, K.J.; Knutson, S.T.; Soito, L.; Klomsiri, C.; Poole, L.B.; Fetrow, J.S. Analysis of the Peroxiredoxin Family: Using Active Site Structure and Sequence Information for Global Classification and Residue Analysis. Proteins 2011, 79, 947–964. [Google Scholar] [CrossRef]
- Soito, L.; Williamson, C.; Knutson, S.T.; Fetrow, J.S.; Poole, L.B.; Nelson, K.J. PREX: PeroxiRedoxin Classification indEX, a Database of Subfamily Assignments across the Diverse Peroxiredoxin Family. Nucleic Acids Res. 2011, 39, D332–D337. [Google Scholar] [CrossRef]
- Bolduc, J.; Koruza, K.; Luo, T.; Malo Pueyo, J.; Vo, T.N.; Ezeriņa, D.; Messens, J. Peroxiredoxins Wear Many Hats: Factors That Fashion Their Peroxide Sensing Personalities. Redox Biol. 2021, 42, 101959. [Google Scholar] [CrossRef]
- Hall, A.; Karplus, P.A.; Poole, L.B. Typical 2-Cys Peroxiredoxins--Structures, Mechanisms and Functions. FEBS J. 2009, 276, 2469–2477. [Google Scholar] [CrossRef]
- Edgar, R.S.; Green, E.W.; Zhao, Y.; van Ooijen, G.; Olmedo, M.; Qin, X.; Xu, Y.; Pan, M.; Valekunja, U.K.; Feeney, K.A.; et al. Peroxiredoxins Are Conserved Markers of Circadian Rhythms. Nature 2012, 485, 459–464. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, K.; Ishikawa, M.; Kaneko, M.; Kamiya, K.; Kato, S.; Nakanishi, S. The Endogenous Redox Rhythm Is Controlled by a Central Circadian Oscillator in Cyanobacterium Synechococcus Elongatus PCC7942. Photosynth. Res. 2019, 142, 203–210. [Google Scholar] [CrossRef]
- Loudon, A.S.I. Circadian Biology: A 2.5 Billion Year Old Clock. Curr. Biol. CB 2012, 22, R570–R571. [Google Scholar] [CrossRef] [PubMed]
- Allen, J.F.; Thake, B.; Martin, W.F. Nitrogenase Inhibition Limited Oxygenation of Earth’s Proterozoic Atmosphere. Trends Plant Sci. 2019, 24, 1022–1031. [Google Scholar] [CrossRef] [PubMed]
- Crowe, S.A.; Døssing, L.N.; Beukes, N.J.; Bau, M.; Kruger, S.J.; Frei, R.; Canfield, D.E. Atmospheric Oxygenation Three Billion Years Ago. Nature 2013, 501, 535–538. [Google Scholar] [CrossRef]
- Hörnlein, C.; Confurius-Guns, V.; Stal, L.J.; Bolhuis, H. Daily Rhythmicity in Coastal Microbial Mats. NPJ Biofilms Microbiomes 2018, 4, 11. [Google Scholar] [CrossRef]
- Rosato, E.; Kyriacou, C.P. Origins of Circadian Rhythmicity. J. Biol. Rhythms 2002, 17, 506–511. [Google Scholar] [CrossRef]
- Pittendrigh, C.S. Temporal Organization: Reflections of a Darwinian Clock-Watcher. Annu. Rev. Physiol. 1993, 55, 16–54. [Google Scholar] [CrossRef]
- Nikaido, S.S.; Johnson, C.H. Daily and Circadian Variation in Survival from Ultraviolet Radiation in Chlamydomonas Reinhardtii. Photochem. Photobiol. 2000, 71, 758–765. [Google Scholar] [CrossRef]
- Cockell, C.S. Ultraviolet Radiation and the Photobiology of Earth’s Early Oceans. Orig. Life Evol. Biosphere J. Int. Soc. Study Orig. Life 2000, 30, 467–499. [Google Scholar] [CrossRef] [PubMed]
- Robertson, J.B.; Davis, C.R.; Johnson, C.H. Visible Light Alters Yeast Metabolic Rhythms by Inhibiting Respiration. Proc. Natl. Acad. Sci. USA 2013, 110, 21130–21135. [Google Scholar] [CrossRef] [PubMed]
- Gehring, W.; Rosbash, M. The Coevolution of Blue-Light Photoreception and Circadian Rhythms. J. Mol. Evol. 2003, 57 (Suppl. 1), S286–S289. [Google Scholar] [CrossRef]
- Baik, L.S.; Recinos, Y.; Chevez, J.A.; Holmes, T.C. Circadian Modulation of Light-Evoked Avoidance/Attraction Behavior in Drosophila. PLoS ONE 2018, 13, e0201927. [Google Scholar] [CrossRef] [PubMed]
- Kawasaki, K.; Iwasaki, H. Involvement of Glycogen Metabolism in Circadian Control of UV Resistance in Cyanobacteria. PLoS Genet. 2020, 16, e1009230. [Google Scholar] [CrossRef]
- Chen, Z.; McKnight, S.L. A Conserved DNA Damage Response Pathway Responsible for Coupling the Cell Division Cycle to the Circadian and Metabolic Cycles. Cell Cycle Georget. Tex 2007, 6, 2906–2912. [Google Scholar] [CrossRef]
- Dietz, K.-J. Peroxiredoxins in Plants and Cyanobacteria. Antioxid. Redox Signal. 2011, 15, 1129–1159. [Google Scholar] [CrossRef]
- Awad, J.; Stotz, H.U.; Fekete, A.; Krischke, M.; Engert, C.; Havaux, M.; Berger, S.; Mueller, M.J. 2-Cysteine Peroxiredoxins and Thylakoid Ascorbate Peroxidase Create a Water-Water Cycle That Is Essential to Protect the Photosynthetic Apparatus under High Light Stress Conditions1. Plant Physiol. 2015, 167, 1592–1603. [Google Scholar] [CrossRef]
- Putker, M.; Crosby, P.; Feeney, K.A.; Hoyle, N.P.; Costa, A.S.H.; Gaude, E.; Frezza, C.; O’Neill, J.S. Mammalian Circadian Period, But Not Phase and Amplitude, Is Robust Against Redox and Metabolic Perturbations. Antioxid. Redox Signal. 2018, 28, 507–520. [Google Scholar] [CrossRef]
- Wong, D.C.; O’Neill, J.S. Non-Transcriptional Processes in Circadian Rhythm Generation. Curr. Opin. Physiol. 2018, 5, 117. [Google Scholar] [CrossRef]
- Causton, H.C. Metabolic Rhythms: A Framework for Coordinating Cellular Function. Eur. J. Neurosci. 2020, 51, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Mastronicola, D.; Falabella, M.; Testa, F.; Pucillo, L.P.; Teixeira, M.; Sarti, P.; Saraiva, L.M.; Giuffrè, A. Functional Characterization of Peroxiredoxins from the Human Protozoan Parasite Giardia Intestinalis. PLoS Negl. Trop. Dis. 2014, 8, e2631. [Google Scholar] [CrossRef] [PubMed]
- Condeles, A.L.; Gomes, F.; de Oliveira, M.A.; Soares Netto, L.E.; Toledo Junior, J.C. Thiol Peroxidases as Major Regulators of Intracellular Levels of Peroxynitrite in Live Saccharomyces Cerevisiae Cells. Antioxidants 2020, 9, 434. [Google Scholar] [CrossRef]
- Chae, H.Z.; Kim, H.J.; Kang, S.W.; Rhee, S.G. Characterization of Three Isoforms of Mammalian Peroxiredoxin That Reduce Peroxides in the Presence of Thioredoxin. Diabetes Res. Clin. Pract. 1999, 45, 101–112. [Google Scholar] [CrossRef]
- Lechardeur, D.; Fernandez, A.; Robert, B.; Gaudu, P.; Trieu-Cuot, P.; Lamberet, G.; Gruss, A. The 2-Cys Peroxiredoxin Alkyl Hydroperoxide Reductase C Binds Heme and Participates in Its Intracellular Availability in Streptococcus Agalactiae. J. Biol. Chem. 2010, 285, 16032–16041. [Google Scholar] [CrossRef] [PubMed]
- Chuang, M.-H.; Wu, M.-S.; Lo, W.-L.; Lin, J.-T.; Wong, C.-H.; Chiou, S.-H. The Antioxidant Protein Alkylhydroperoxide Reductase of Helicobacter Pylori Switches from a Peroxide Reductase to a Molecular Chaperone Function. Proc. Natl. Acad. Sci. USA 2006, 103, 2552–2557. [Google Scholar] [CrossRef]
- Kim, T.K.; Tirloni, L.; Bencosme-Cuevas, E.; Kim, T.H.; Diedrich, J.K.; Yates, J.R.; Mulenga, A. Borrelia Burgdorferi Infection Modifies Protein Content in Saliva of Ixodes Scapularis Nymphs. BMC Genom. 2021, 22, 152. [Google Scholar] [CrossRef]
- Rocha, M.C.; de Godoy, K.F.; Bannitz-Fernandes, R.; Fabri, J.H.T.M.; Barbosa, M.M.F.; de Castro, P.A.; Almeida, F.; Goldman, G.H.; da Cunha, A.F.; Netto, L.E.S.; et al. Analyses of the Three 1-Cys Peroxiredoxins from Aspergillus Fumigatus Reveal That Cytosolic Prx1 Is Central to H2O2 Metabolism and Virulence. Sci. Rep. 2018, 8, 12314. [Google Scholar] [CrossRef]
- Park, S.G.; Cha, M.-K.; Jeong, W.; Kim, I.-H. Distinct Physiological Functions of Thiol Peroxidase Isoenzymes in Saccharomyces Cerevisiae. J. Biol. Chem. 2000, 275, 5723–5732. [Google Scholar] [CrossRef]
- Cha, M.-K.; Kim, H.-K.; Kim, I.-H. Thioredoxin-Linked “Thiol Peroxidase” from Periplasmic Space of Escherichia coli (*). J. Biol. Chem. 1995, 270, 28635–28641. [Google Scholar] [CrossRef]
- Hicks, L.D.; Raghavan, R.; Battisti, J.M.; Minnick, M.F. A DNA-Binding Peroxiredoxin of Coxiella Burnetii Is Involved in Countering Oxidative Stress during Exponential-Phase Growth. J. Bacteriol. 2010, 192, 2077–2084. [Google Scholar] [CrossRef] [PubMed]
- Biteau, B.; Labarre, J.; Toledano, M.B. ATP-Dependent Reduction of Cysteine-Sulphinic Acid by S. Cerevisiae Sulphiredoxin. Nature 2003, 425, 980–984. [Google Scholar] [CrossRef] [PubMed]
- Rhee, S.G.; Jeong, W.; Chang, T.-S.; Woo, H.A. Sulfiredoxin, the Cysteine Sulfinic Acid Reductase Specific to 2-Cys Peroxiredoxin: Its Discovery, Mechanism of Action, and Biological Significance. Kidney Int. Suppl. 2007, 72, S3–S8. [Google Scholar] [CrossRef] [PubMed]
- Jeong, W.; Bae, S.H.; Toledano, M.B.; Rhee, S.G. Role of Sulfiredoxin as a Regulator of Peroxiredoxin Function and Regulation of Its Expression. Free Radic. Biol. Med. 2012, 53, 447–456. [Google Scholar] [CrossRef] [PubMed]
- Veal, E.A.; Underwood, Z.E.; Tomalin, L.E.; Morgan, B.A.; Pillay, C.S. Hyperoxidation of Peroxiredoxins: Gain or Loss of Function? Antioxid. Redox Signal. 2018, 28, 574–590. [Google Scholar] [CrossRef]
- Jang, H.H.; Lee, K.O.; Chi, Y.H.; Jung, B.G.; Park, S.K.; Park, J.H.; Lee, J.R.; Lee, S.S.; Moon, J.C.; Yun, J.W.; et al. Two Enzymes in One; Two Yeast Peroxiredoxins Display Oxidative Stress-Dependent Switching from a Peroxidase to a Molecular Chaperone Function. Cell 2004, 117, 625–635. [Google Scholar] [CrossRef] [PubMed]
- Wood, Z.A.; Poole, L.B.; Karplus, P.A. Peroxiredoxin Evolution and the Regulation of Hydrogen Peroxide Signaling. Science 2003, 300, 650–653. [Google Scholar] [CrossRef]
- Tairum, C.A.; Santos, M.C.; Breyer, C.A.; de Oliveira, A.L.P.; Cabrera, V.I.M.; Toledo-Silva, G.; Mori, G.M.; Toyama, M.H.; Netto, L.E.S.; de Oliveira, M.A. Effects of Serine or Threonine in the Active Site of Typical 2-Cys Prx on Hyperoxidation Susceptibility and on Chaperone Activity. Antioxidants 2021, 10, 1032. [Google Scholar] [CrossRef] [PubMed]
- Bolduc, J.A.; Nelson, K.J.; Haynes, A.C.; Lee, J.; Reisz, J.A.; Graff, A.H.; Clodfelter, J.E.; Parsonage, D.; Poole, L.B.; Furdui, C.M.; et al. Novel Hyperoxidation Resistance Motifs in 2-Cys Peroxiredoxins. J. Biol. Chem. 2018, 293, 11901–11912. [Google Scholar] [CrossRef] [PubMed]
- Rhee, S.G.; Kang, S.W.; Jeong, W.; Chang, T.-S.; Yang, K.-S.; Woo, H.A. Intracellular Messenger Function of Hydrogen Peroxide and Its Regulation by Peroxiredoxins. Curr. Opin. Cell Biol. 2005, 17, 183–189. [Google Scholar] [CrossRef]
- Pizzino, G.; Irrera, N.; Cucinotta, M.; Pallio, G.; Mannino, F.; Arcoraci, V.; Squadrito, F.; Altavilla, D.; Bitto, A. Oxidative Stress: Harms and Benefits for Human Health. Oxid. Med. Cell. Longev. 2017, 2017, 8416763. [Google Scholar] [CrossRef]
- Neumann, C.A.; Krause, D.S.; Carman, C.V.; Das, S.; Dubey, D.P.; Abraham, J.L.; Bronson, R.T.; Fujiwara, Y.; Orkin, S.H.; Van Etten, R.A. Essential Role for the Peroxiredoxin Prdx1 in Erythrocyte Antioxidant Defence and Tumour Suppression. Nature 2003, 424, 561–565. [Google Scholar] [CrossRef] [PubMed]
- Schieber, M.; Chandel, N.S. ROS Function in Redox Signaling and Oxidative Stress. Curr. Biol. CB 2014, 24, R453–R462. [Google Scholar] [CrossRef]
- Antunes, F.; Brito, P.M. Quantitative Biology of Hydrogen Peroxide Signaling. Redox Biol. 2017, 13, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Di Marzo, N.; Chisci, E.; Giovannoni, R. The Role of Hydrogen Peroxide in Redox-Dependent Signaling: Homeostatic and Pathological Responses in Mammalian Cells. Cells 2018, 7, 156. [Google Scholar] [CrossRef] [PubMed]
- Reczek, C.R.; Chandel, N.S. ROS-Dependent Signal Transduction. Curr. Opin. Cell Biol. 2015, 33, 8–13. [Google Scholar] [CrossRef]
- Randall, L.M.; Ferrer-Sueta, G.; Denicola, A. Peroxiredoxins as Preferential Targets in H2O2-Induced Signaling. Methods Enzymol. 2013, 527, 41–63. [Google Scholar] [CrossRef] [PubMed]
- Winterbourn, C.C. The Biological Chemistry of Hydrogen Peroxide. Methods Enzymol. 2013, 528, 3–25. [Google Scholar] [CrossRef]
- Sies, H.; Berndt, C.; Jones, D.P. Oxidative Stress. Annu. Rev. Biochem. 2017, 86, 715–748. [Google Scholar] [CrossRef] [PubMed]
- Winterbourn, C.C.; Hampton, M.B. Thiol Chemistry and Specificity in Redox Signaling. Free Radic. Biol. Med. 2008, 45, 549–561. [Google Scholar] [CrossRef] [PubMed]
- Cox, A.G.; Winterbourn, C.C.; Hampton, M.B. Mitochondrial Peroxiredoxin Involvement in Antioxidant Defence and Redox Signalling. Biochem. J. 2009, 425, 313–325. [Google Scholar] [CrossRef] [PubMed]
- Holmström, K.M.; Finkel, T. Cellular Mechanisms and Physiological Consequences of Redox-Dependent Signalling. Nat. Rev. Mol. Cell Biol. 2014, 15, 411–421. [Google Scholar] [CrossRef] [PubMed]
- Andrés, C.M.C.; Pérez de la Lastra, J.M.; Juan, C.A.; Plou, F.J.; Pérez-Lebeña, E. Chemistry of Hydrogen Peroxide Formation and Elimination in Mammalian Cells, and Its Role in Various Pathologies. Stresses 2022, 2, 256–274. [Google Scholar] [CrossRef]
- Sies, H. Hydrogen Peroxide as a Central Redox Signaling Molecule in Physiological Oxidative Stress: Oxidative Eustress. Redox Biol. 2017, 11, 613–619. [Google Scholar] [CrossRef]
- D’Autréaux, B.; Toledano, M.B. ROS as Signalling Molecules: Mechanisms That Generate Specificity in ROS Homeostasis. Nat. Rev. Mol. Cell Biol. 2007, 8, 813–824. [Google Scholar] [CrossRef] [PubMed]
- Karplus, P.A. A Primer on Peroxiredoxin Biochemistry. Free Radic. Biol. Med. 2015, 80, 183–190. [Google Scholar] [CrossRef] [PubMed]
- Toledano, M.B.; Planson, A.-G.; Delaunay-Moisan, A. Reining in H2O2 for Safe Signaling. Cell 2010, 140, 454–456. [Google Scholar] [CrossRef]
- Barranco-Medina, S.; Lázaro, J.-J.; Dietz, K.-J. The Oligomeric Conformation of Peroxiredoxins Links Redox State to Function. FEBS Lett. 2009, 583, 1809–1816. [Google Scholar] [CrossRef] [PubMed]
- Moon, J.C.; Hah, Y.-S.; Kim, W.Y.; Jung, B.G.; Jang, H.H.; Lee, J.R.; Kim, S.Y.; Lee, Y.M.; Jeon, M.G.; Kim, C.W.; et al. Oxidative Stress-Dependent Structural and Functional Switching of a Human 2-Cys Peroxiredoxin Isotype II That Enhances HeLa Cell Resistance to H2O2-Induced Cell Death. J. Biol. Chem. 2005, 280, 28775–28784. [Google Scholar] [CrossRef] [PubMed]
- Nordzieke, D.E.; Medraño-Fernandez, I. The Plasma Membrane: A Platform for Intra- and Intercellular Redox Signaling. Antioxidants 2018, 7, 168. [Google Scholar] [CrossRef]
- Kil, I.S.; Lee, S.K.; Ryu, K.W.; Woo, H.A.; Hu, M.-C.; Bae, S.H.; Rhee, S.G. Feedback Control of Adrenal Steroidogenesis via H2O2-Dependent, Reversible Inactivation of Peroxiredoxin III in Mitochondria. Mol. Cell 2012, 46, 584–594. [Google Scholar] [CrossRef] [PubMed]
- Stöcker, S.; Van Laer, K.; Mijuskovic, A.; Dick, T.P. The Conundrum of Hydrogen Peroxide Signaling and the Emerging Role of Peroxiredoxins as Redox Relay Hubs. Antioxid. Redox Signal. 2018, 28, 558–573. [Google Scholar] [CrossRef]
- Winterbourn, C.C. Biological Production, Detection, and Fate of Hydrogen Peroxide. Antioxid. Redox Signal. 2018, 29, 541–551. [Google Scholar] [CrossRef]
- Latimer, H.R.; Veal, E.A. Peroxiredoxins in Regulation of MAPK Signalling Pathways; Sensors and Barriers to Signal Transduction. Mol. Cells 2016, 39, 40–45. [Google Scholar] [CrossRef] [PubMed]
- Netto, L.E.S.; Antunes, F. The Roles of Peroxiredoxin and Thioredoxin in Hydrogen Peroxide Sensing and in Signal Transduction. Mol. Cells 2016, 39, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Sobotta, M.C.; Liou, W.; Stöcker, S.; Talwar, D.; Oehler, M.; Ruppert, T.; Scharf, A.N.D.; Dick, T.P. Peroxiredoxin-2 and STAT3 Form a Redox Relay for H2O2 Signaling. Nat. Chem. Biol. 2015, 11, 64–70. [Google Scholar] [CrossRef]
- Fomenko, D.E.; Koc, A.; Agisheva, N.; Jacobsen, M.; Kaya, A.; Malinouski, M.; Rutherford, J.C.; Siu, K.-L.; Jin, D.-Y.; Winge, D.R.; et al. Thiol Peroxidases Mediate Specific Genome-Wide Regulation of Gene Expression in Response to Hydrogen Peroxide. Proc. Natl. Acad. Sci. USA 2011, 108, 2729–2734. [Google Scholar] [CrossRef] [PubMed]
- Delaunay, A.; Pflieger, D.; Barrault, M.B.; Vinh, J.; Toledano, M.B. A Thiol Peroxidase Is an H2O2 Receptor and Redox-Transducer in Gene Activation. Cell 2002, 111, 471–481. [Google Scholar] [CrossRef]
- Talwar, D.; Messens, J.; Dick, T.P. A Role for Annexin A2 in Scaffolding the Peroxiredoxin 2–STAT3 Redox Relay Complex. Nat. Commun. 2020, 11, 4512. [Google Scholar] [CrossRef] [PubMed]
- Klomsiri, C.; Karplus, P.A.; Poole, L.B. Cysteine-Based Redox Switches in Enzymes. Antioxid. Redox Signal. 2011, 14, 1065–1077. [Google Scholar] [CrossRef]
- Travasso, R.D.M.; Sampaio dos Aidos, F.; Bayani, A.; Abranches, P.; Salvador, A. Localized Redox Relays as a Privileged Mode of Cytoplasmic Hydrogen Peroxide Signaling. Redox Biol. 2017, 12, 233–245. [Google Scholar] [CrossRef]
- Brandes, N.; Schmitt, S.; Jakob, U. Thiol-Based Redox Switches in Eukaryotic Proteins. Antioxid. Redox Signal. 2009, 11, 997–1014. [Google Scholar] [CrossRef]
- Stöcker, S.; Maurer, M.; Ruppert, T.; Dick, T.P. A Role for 2-Cys Peroxiredoxins in Facilitating Cytosolic Protein Thiol Oxidation. Nat. Chem. Biol. 2018, 14, 148–155. [Google Scholar] [CrossRef] [PubMed]
- Bozonet, S.M.; Findlay, V.J.; Day, A.M.; Cameron, J.; Veal, E.A.; Morgan, B.A. Oxidation of a Eukaryotic 2-Cys Peroxiredoxin Is a Molecular Switch Controlling the Transcriptional Response to Increasing Levels of Hydrogen Peroxide. J. Biol. Chem. 2005, 280, 23319–23327. [Google Scholar] [CrossRef]
- Calabrese, G.; Peker, E.; Amponsah, P.S.; Hoehne, M.N.; Riemer, T.; Mai, M.; Bienert, G.P.; Deponte, M.; Morgan, B.; Riemer, J. Hyperoxidation of Mitochondrial Peroxiredoxin Limits H2O2-induced Cell Death in Yeast. EMBO J. 2019, 38, e101552. [Google Scholar] [CrossRef] [PubMed]
- Winterbourn, C.C.; Hampton, M.B. Redox Biology: Signaling via a Peroxiredoxin Sensor. Nat. Chem. Biol. 2015, 11, 5–6. [Google Scholar] [CrossRef]
- van Dam, L.; Pagès-Gallego, M.; Polderman, P.E.; van Es, R.M.; Burgering, B.M.T.; Vos, H.R.; Dansen, T.B. The Human 2-Cys Peroxiredoxins Form Widespread, Cysteine-Dependent- and Isoform-Specific Protein-Protein Interactions. Antioxidants 2021, 10, 627. [Google Scholar] [CrossRef]
- Brown, J.D.; Day, A.M.; Taylor, S.R.; Tomalin, L.E.; Morgan, B.A.; Veal, E.A. A Peroxiredoxin Promotes H2O2 Signaling and Oxidative Stress Resistance by Oxidizing a Thioredoxin Family Protein. Cell Rep. 2013, 5, 1425–1435. [Google Scholar] [CrossRef] [PubMed]
- Day, A.M.; Brown, J.D.; Taylor, S.R.; Rand, J.D.; Morgan, B.A.; Veal, E.A. Inactivation of a Peroxiredoxin by Hydrogen Peroxide Is Critical for Thioredoxin-Mediated Repair of Oxidized Proteins and Cell Survival. Mol. Cell 2012, 45, 398–408. [Google Scholar] [CrossRef] [PubMed]
- Vivancos, A.P.; Castillo, E.A.; Biteau, B.; Nicot, C.; Ayté, J.; Toledano, M.B.; Hidalgo, E. A Cysteine-Sulfinic Acid in Peroxiredoxin Regulates H2O2-Sensing by the Antioxidant Pap1 Pathway. Proc. Natl. Acad. Sci. USA 2005, 102, 8875–8880. [Google Scholar] [CrossRef]
- Cox, A.G.; Pearson, A.G.; Pullar, J.M.; Jönsson, T.J.; Lowther, W.T.; Winterbourn, C.C.; Hampton, M.B. Mitochondrial Peroxiredoxin 3 Is More Resilient to Hyperoxidation than Cytoplasmic Peroxiredoxins. Biochem. J. 2009, 421, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Jajere, S.M. A Review of Salmonella Enterica with Particular Focus on the Pathogenicity and Virulence Factors, Host Specificity and Antimicrobial Resistance Including Multidrug Resistance. Vet. World 2019, 12, 504–521. [Google Scholar] [CrossRef] [PubMed]
- Scharff, R.L. Food Attribution and Economic Cost Estimates for Meat- and Poultry-Related Illnesses. J. Food Prot. 2020, 83, 959–967. [Google Scholar] [CrossRef]
- Balasubramanian, R.; Im, J.; Lee, J.-S.; Jeon, H.J.; Mogeni, O.D.; Kim, J.H.; Rakotozandrindrainy, R.; Baker, S.; Marks, F. The Global Burden and Epidemiology of Invasive Non-Typhoidal Salmonella Infections. Hum. Vaccines Immunother. 2019, 15, 1421–1426. [Google Scholar] [CrossRef]
- Alikhan, N.-F.; Moreno, L.Z.; Castellanos, L.R.; Chattaway, M.A.; McLauchlin, J.; Lodge, M.; O’Grady, J.; Zamudio, R.; Doughty, E.; Petrovska, L.; et al. Dynamics of Salmonella Enterica and Antimicrobial Resistance in the Brazilian Poultry Industry and Global Impacts on Public Health. PLoS Genet. 2022, 18, e1010174. [Google Scholar] [CrossRef] [PubMed]
- Aljahdali, N.H.; Sanad, Y.M.; Han, J.; Foley, S.L. Current Knowledge and Perspectives of Potential Impacts of Salmonella Enterica on the Profile of the Gut Microbiota. BMC Microbiol. 2020, 20, 353. [Google Scholar] [CrossRef]
- Yang, H.; Villani, R.M.; Wang, H.; Simpson, M.J.; Roberts, M.S.; Tang, M.; Liang, X. The Role of Cellular Reactive Oxygen Species in Cancer Chemotherapy. J. Exp. Clin. Cancer Res. CR 2018, 37, 266. [Google Scholar] [CrossRef]
- Nash, A.K.; Auchtung, T.A.; Wong, M.C.; Smith, D.P.; Gesell, J.R.; Ross, M.C.; Stewart, C.J.; Metcalf, G.A.; Muzny, D.M.; Gibbs, R.A.; et al. The Gut Mycobiome of the Human Microbiome Project Healthy Cohort. Microbiome 2017, 5, 153. [Google Scholar] [CrossRef] [PubMed]
- Pérez, J.C. Fungi of the Human Gut Microbiota: Roles and Significance. Int. J. Med. Microbiol. 2021, 311, 151490. [Google Scholar] [CrossRef] [PubMed]
- Fiers, W.D.; Gao, I.H.; Iliev, I.D. Gut Mycobiota under Scrutiny: Fungal Symbionts or Environmental Transients? Curr. Opin. Microbiol. 2019, 50, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Li, X.V.; Leonardi, I.; Iliev, I.D. Gut Mycobiota in Immunity and Inflammatory Disease. Immunity 2019, 50, 1365–1379. [Google Scholar] [CrossRef]
- Hoffmann, C.; Dollive, S.; Grunberg, S.; Chen, J.; Li, H.; Wu, G.D.; Lewis, J.D.; Bushman, F.D. Archaea and Fungi of the Human Gut Microbiome: Correlations with Diet and Bacterial Residents. PLoS ONE 2013, 8, e66019. [Google Scholar] [CrossRef]
- Hallen-Adams, H.E.; Suhr, M.J. Fungi in the Healthy Human Gastrointestinal Tract. Virulence 2016, 8, 352–358. [Google Scholar] [CrossRef]
- Cavalieri, D.; McGovern, P.E.; Hartl, D.L.; Mortimer, R.; Polsinelli, M. Evidence for S. cerevisiae Fermentation in Ancient Wine. J. Mol. Evol. 2003, 57 (Suppl. 1), S226–S232. [Google Scholar] [CrossRef] [PubMed]
- Ghannoum, M.A.; Jurevic, R.J.; Mukherjee, P.K.; Cui, F.; Sikaroodi, M.; Naqvi, A.; Gillevet, P.M. Characterization of the Oral Fungal Microbiome (Mycobiome) in Healthy Individuals. PLoS Pathog. 2010, 6, e1000713. [Google Scholar] [CrossRef] [PubMed]
- Scanlan, P.D.; Marchesi, J.R. Micro-Eukaryotic Diversity of the Human Distal Gut Microbiota: Qualitative Assessment Using Culture-Dependent and -Independent Analysis of Faeces. ISME J. 2008, 2, 1183–1193. [Google Scholar] [CrossRef] [PubMed]
- Angebault, C.; Djossou, F.; Abélanet, S.; Permal, E.; Ben Soltana, M.; Diancourt, L.; Bouchier, C.; Woerther, P.-L.; Catzeflis, F.; Andremont, A.; et al. Candida Albicans Is Not Always the Preferential Yeast Colonizing Humans: A Study in Wayampi Amerindians. J. Infect. Dis. 2013, 208, 1705–1716. [Google Scholar] [CrossRef]
- Sokol, H.; Leducq, V.; Aschard, H.; Pham, H.-P.; Jegou, S.; Landman, C.; Cohen, D.; Liguori, G.; Bourrier, A.; Nion-Larmurier, I.; et al. Fungal Microbiota Dysbiosis in IBD. Gut 2017, 66, 1039–1048. [Google Scholar] [CrossRef]
- Findley, K.; Oh, J.; Yang, J.; Conlan, S.; Deming, C.; Meyer, J.A.; Schoenfeld, D.; Nomicos, E.; Park, M.; NIH Intramural Sequencing Center Comparative Sequencing Program; et al. Topographic Diversity of Fungal and Bacterial Communities in Human Skin. Nature 2013, 498, 367–370. [Google Scholar] [CrossRef]
- Esposito, M.M.; Patsakos, S.; Borruso, L. The Role of the Mycobiome in Women’s Health. J. Fungi 2023, 9, 348. [Google Scholar] [CrossRef] [PubMed]
- Sabbatini, S.; Monari, C.; Ballet, N.; Mosci, P.; Decherf, A.C.; Pélerin, F.; Perito, S.; Scarpelli, P.; Vecchiarelli, A. Saccharomyces Cerevisiae–Based Probiotic as Novel Anti-Microbial Agent for Therapy of Bacterial Vaginosis. Virulence 2018, 9, 954–966. [Google Scholar] [CrossRef] [PubMed]
- Sivignon, A.; de Vallée, A.; Barnich, N.; Denizot, J.; Darcha, C.; Pignède, G.; Vandekerckove, P.; Darfeuille-Michaud, A. Saccharomyces Cerevisiae CNCM I-3856 Prevents Colitis Induced by AIEC Bacteria in the Transgenic Mouse Model Mimicking Crohn’s Disease. Inflamm. Bowel Dis. 2015, 21, 276–286. [Google Scholar] [CrossRef] [PubMed]
- Hatoum, R.; Labrie, S.; Fliss, I. Antimicrobial and Probiotic Properties of Yeasts: From Fundamental to Novel Applications. Front. Microbiol. 2012, 3, 421. [Google Scholar] [CrossRef]
- Anoop, V.; Rotaru, S.; Shwed, P.S.; Tayabali, A.F.; Arvanitakis, G. Review of Current Methods for Characterizing Virulence and Pathogenicity Potential of Industrial Saccharomyces Cerevisiae Strains towards Humans. FEMS Yeast Res. 2015, 15, fov057. [Google Scholar] [CrossRef]
- Murphy, A.; Kavanagh, K. Emergence of Saccharomyces Cerevisiae as a Human Pathogen: Implications for Biotechnology. Enzyme Microb. Technol. 1999, 25, 551–557. [Google Scholar] [CrossRef]
- Pérez-Torrado, R.; Querol, A. Opportunistic Strains of Saccharomyces Cerevisiae: A Potential Risk Sold in Food Products. Front. Microbiol. 2015, 6, 1522. [Google Scholar] [CrossRef] [PubMed]
- Eelderink-Chen, Z.; Mazzotta, G.; Sturre, M.; Bosman, J.; Roenneberg, T.; Merrow, M. A Circadian Clock in Saccharomyces Cerevisiae. Proc. Natl. Acad. Sci. USA 2010, 107, 2043–2047. [Google Scholar] [CrossRef]
- Silverman, S.J.; Petti, A.A.; Slavov, N.; Parsons, L.; Briehof, R.; Thiberge, S.Y.; Zenklusen, D.; Gandhi, S.J.; Larson, D.R.; Singer, R.H.; et al. Metabolic Cycling in Single Yeast Cells from Unsynchronized Steady-State Populations Limited on Glucose or Phosphate. Proc. Natl. Acad. Sci. USA 2010, 107, 6946–6951. [Google Scholar] [CrossRef]
- Tu, B.P.; McKnight, S.L. The Yeast Metabolic Cycle: Insights into the Life of a Eukaryotic Cell. Cold Spring Harb. Symp. Quant. Biol. 2007, 72, 339–343. [Google Scholar] [CrossRef]
- Tu, B.P.; Kudlicki, A.; Rowicka, M.; McKnight, S.L. Logic of the Yeast Metabolic Cycle: Temporal Compartmentalization of Cellular Processes. Science 2005, 310, 1152–1158. [Google Scholar] [CrossRef]
- Mellor, J. The Molecular Basis of Metabolic Cycles and Their Relationship to Circadian Rhythms. Nat. Struct. Mol. Biol. 2016, 23, 1035–1044. [Google Scholar] [CrossRef] [PubMed]
- Lloyd, D. Respiratory Oscillations in Yeasts. In Cellular Oscillatory Mechanisms; Maroto, M., Monk, N.A.M., Eds.; Advances in Experimental Medicine and Biology; Springer: New York, NY, USA, 2009; pp. 118–140. ISBN 978-0-387-09794-7. [Google Scholar]
- Causton, H.C.; Feeney, K.A.; Ziegler, C.A.; O’Neill, J.S. Metabolic Cycles in Yeast Share Features Conserved among Circadian Rhythms. Curr. Biol. 2015, 25, 1056–1062. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, J.S. Redox-Coupled Rhythm and Brews. Nat. Chem. Biol. 2021, 17, 373–374. [Google Scholar] [CrossRef] [PubMed]
- Murray, D.B.; Klevecz, R.R.; Lloyd, D. Generation and Maintenance of Synchrony in Saccharomyces Cerevisiae Continuous Culture. Exp. Cell Res. 2003, 287, 10–15. [Google Scholar] [CrossRef]
- Beuse, M.; Bartling, R.; Kopmann, A.; Diekmann, H.; Thoma, M. Effect of the Dilution Rate on the Mode of Oscillation in Continuous Cultures of Saccharomyces Cerevisiae. J. Biotechnol. 1998, 61, 15–31. [Google Scholar] [CrossRef]
- Lloyd, D. The Ultradian Clock: Not to Be Confused with the Cell Cycle. Nat. Rev. Mol. Cell Biol. 2006, 7, 1. [Google Scholar] [CrossRef]
- Lloyd, D.; Murray, D.B. Redox Rhythmicity: Clocks at the Core of Temporal Coherence. BioEssays News Rev. Mol. Cell. Dev. Biol. 2007, 29, 465–473. [Google Scholar] [CrossRef]
- Klevecz, R.R.; Bolen, J.; Forrest, G.; Murray, D.B. A Genomewide Oscillation in Transcription Gates DNA Replication and Cell Cycle. Proc. Natl. Acad. Sci. USA 2004, 101, 1200–1205. [Google Scholar] [CrossRef]
- Wang, G.-Z.; Hickey, S.L.; Shi, L.; Huang, H.-C.; Nakashe, P.; Koike, N.; Tu, B.P.; Takahashi, J.S.; Konopka, G. Cycling Transcriptional Networks Optimize Energy Utilization on a Genome Scale. Cell Rep. 2015, 13, 1868–1880. [Google Scholar] [CrossRef]
- Slavov, N.; Macinskas, J.; Caudy, A.; Botstein, D. Metabolic Cycling without Cell Division Cycling in Respiring Yeast. Proc. Natl. Acad. Sci. USA 2011, 108, 19090–19095. [Google Scholar] [CrossRef]
- Burnetti, A.J.; Aydin, M.; Buchler, N.E. Cell Cycle Start Is Coupled to Entry into the Yeast Metabolic Cycle across Diverse Strains and Growth Rates. Mol. Biol. Cell 2016, 27, 64–74. [Google Scholar] [CrossRef]
- Johnson, C.H. Circadian Clocks and Cell Division. Cell Cycle 2010, 9, 3864–3873. [Google Scholar] [CrossRef]
- Papagiannakis, A.; Niebel, B.; Wit, E.C.; Heinemann, M. Autonomous Metabolic Oscillations Robustly Gate the Early and Late Cell Cycle. Mol. Cell 2017, 65, 285–295. [Google Scholar] [CrossRef]
- Chen, Z.; Odstrcil, E.A.; Tu, B.P.; McKnight, S.L. Restriction of DNA Replication to the Reductive Phase of the Metabolic Cycle Protects Genome Integrity. Science 2007, 316, 1916–1919. [Google Scholar] [CrossRef]
- O’Neill, J.S.; Hoyle, N.P.; Robertson, J.B.; Edgar, R.S.; Beale, A.D.; Peak-Chew, S.Y.; Day, J.; Costa, A.S.H.; Frezza, C.; Causton, H.C. Eukaryotic Cell Biology Is Temporally Coordinated to Support the Energetic Demands of Protein Homeostasis. Nat. Commun. 2020, 11, 4706. [Google Scholar] [CrossRef]
- Küenzi, M.T.; Fiechter, A. Changes in Carbohydrate Composition and Trehalase-Activity during the Budding Cycle of Saccharomyces Cerevisiae. Arch. Mikrobiol. 1969, 64, 396–407. [Google Scholar] [CrossRef]
- Murray, D.B.; Beckmann, M.; Kitano, H. Regulation of Yeast Oscillatory Dynamics. Proc. Natl. Acad. Sci. USA 2007, 104, 2241–2246. [Google Scholar] [CrossRef] [PubMed]
- Slavov, N.; Botstein, D. Coupling among Growth Rate Response, Metabolic Cycle, and Cell Division Cycle in Yeast. Mol. Biol. Cell 2011, 22, 1997–2009. [Google Scholar] [CrossRef]
- Lloyd, D.; Murray, D.B.; Aon, M.A.; Cortassa, S.; Roussel, M.R.; Beckmann, M.; Poole, R.K. Temporal Metabolic Partitioning of the Yeast and Protist Cellular Networks: The Cell Is a Global Scale-Invariant (Fractal or Self-Similar) Multioscillator. J. Biomed. Opt. 2019, 24, 051404. [Google Scholar] [CrossRef] [PubMed]
- Murray, D.B.; Amariei, C.; Sasidharan, K.; Machné, R.; Aon, M.A.; Lloyd, D. Temporal Partitioning of the Yeast Cellular Network. In Systems Biology of Metabolic and Signaling Networks: Energy, Mass and Information Transfer; Aon, M.A., Saks, V., Schlattner, U., Eds.; Springer Series in Biophysics; Springer: Berlin/Heidelberg, Germany, 2014; pp. 323–349. ISBN 978-3-642-38505-6. [Google Scholar]
- Murray, D.B. The Respiratory Oscillation in Yeast Phase Definitions and Periodicity. Nat. Rev. Mol. Cell Biol. 2006, 7, 1. [Google Scholar] [CrossRef]
- Takhaveev, V.; Özsezen, S.; Smith, E.N.; Zylstra, A.; Chaillet, M.L.; Chen, H.; Papagiannakis, A.; Milias-Argeitis, A.; Heinemann, M. Temporal Segregation of Biosynthetic Processes Is Responsible for Metabolic Oscillations during the Budding Yeast Cell Cycle. Nat. Metab. 2023, 5, 294–313. [Google Scholar] [CrossRef]
- Murray, D.B.; Lloyd, D. Multiple Rediscoveries and Misconceptions; the Yeast Metabolic Oscillation. Function 2021, 2, zqab039. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Wang, X.; Yang, X.; Liu, S.; Jiang, L.; Qu, Y.; Hu, L.; Ouyang, Q.; Tang, C. Reliable Cell Cycle Commitment in Budding Yeast Is Ensured by Signal Integration. eLife 2015, 4, e03977. [Google Scholar] [CrossRef] [PubMed]
- Laxman, S.; Sutter, B.M.; Tu, B.P. Behavior of a Metabolic Cycling Population at the Single Cell Level as Visualized by Fluorescent Gene Expression Reporters. PLoS ONE 2010, 5, e12595. [Google Scholar] [CrossRef]
- Aon, M.A.; Cortassa, S.; Lemar, K.M.; Hayes, A.J.; Lloyd, D. Single and Cell Population Respiratory Oscillations in Yeast: A 2-Photon Scanning Laser Microscopy Study. FEBS Lett. 2007, 581, 8–14. [Google Scholar] [CrossRef]
- Amponsah, P.S.; Yahya, G.; Zimmermann, J.; Mai, M.; Mergel, S.; Mühlhaus, T.; Storchova, Z.; Morgan, B. Peroxiredoxins Couple Metabolism and Cell Division in an Ultradian Cycle. Nat. Chem. Biol. 2021, 17, 477–484. [Google Scholar] [CrossRef] [PubMed]
- Lloyd, D.; Murray, D.B. The Temporal Architecture of Eukaryotic Growth. FEBS Lett. 2006, 580, 2830–2835. [Google Scholar] [CrossRef]
- Lloyd, D.; Murray, D.B. Ultradian Metronome: Timekeeper for Orchestration of Cellular Coherence. Trends Biochem. Sci. 2005, 30, 373–377. [Google Scholar] [CrossRef]
- Kippert, F. Circadian Control of Heat Tolerance in Stationary Phase Cultures of Schizosaccharomyces Pombe. Arch. Microbiol. 1989, 151, 177–179. [Google Scholar] [CrossRef]
- Lloyd, D.; Kippert, F. Intracellular Coordination by the Ultradian Clock. Cell Biol. Int. 1993, 17, 1047–1052. [Google Scholar] [CrossRef]
- Kippert, F.; Lloyd, D. A Temperature-Compensated Ultradian Clock Ticks in Schizosaccharomyces Pombe. Microbiol. Read. Engl. 1995, 141 Pt 4, 883–890. [Google Scholar] [CrossRef]
- Hanzén, S.; Vielfort, K.; Yang, J.; Roger, F.; Andersson, V.; Zamarbide-Forés, S.; Andersson, R.; Malm, L.; Palais, G.; Biteau, B.; et al. Lifespan Control by Redox-Dependent Recruitment of Chaperones to Misfolded Proteins. Cell 2016, 166, 140–151. [Google Scholar] [CrossRef]
- Llopis, S.; Querol, A.; Heyken, A.; Hube, B.; Jespersen, L.; Fernández-Espinar, M.T.; Pérez-Torrado, R. Transcriptomics in Human Blood Incubation Reveals the Importance of Oxidative Stress Response in Saccharomyces Cerevisiae Clinical Strains. BMC Genom. 2012, 13, 419. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez-Escobedo, G.; Hernández-Carreón, O.; Morales-Rojano, B.; Revuelta-Rodríguez, B.; Vázquez-Franco, N.; Castaño, I.; De Las Peñas, A. Candida Glabrata Peroxiredoxins, Tsa1 and Tsa2, and Sulfiredoxin, Srx1, Protect against Oxidative Damage and Are Necessary for Virulence. Fungal Genet. Biol. FG B 2020, 135, 103287. [Google Scholar] [CrossRef]
- Roger, F.; Picazo, C.; Reiter, W.; Libiad, M.; Asami, C.; Hanzén, S.; Gao, C.; Lagniel, G.; Welkenhuysen, N.; Labarre, J.; et al. Peroxiredoxin Promotes Longevity and H2O2-Resistance in Yeast through Redox-Modulation of Protein Kinase A. eLife 2020, 9, e60346. [Google Scholar] [CrossRef] [PubMed]
- Molin, M.; Yang, J.; Hanzén, S.; Toledano, M.B.; Labarre, J.; Nyström, T. Life Span Extension and H2O2 Resistance Elicited by Caloric Restriction Require the Peroxiredoxin Tsa1 in Saccharomyces Cerevisiae. Mol. Cell 2011, 43, 823–833. [Google Scholar] [CrossRef] [PubMed]
- Kritsiligkou, P.; Nowicki-Osuch, K.; Carter, Z.; Kershaw, C.J.; Creamer, D.R.; Weids, A.J.; Grant, C.M. Tolerance to Nascent Protein Misfolding Stress Requires Fine-Tuning of the cAMP/PKA Pathway. J. Biol. Chem. 2021, 296, 100690. [Google Scholar] [CrossRef] [PubMed]
- Ghaemmaghami, S.; Huh, W.-K.; Bower, K.; Howson, R.W.; Belle, A.; Dephoure, N.; O’Shea, E.K.; Weissman, J.S. Global Analysis of Protein Expression in Yeast. Nature 2003, 425, 737–741. [Google Scholar] [CrossRef]
- Gaucher, J.; Montellier, E.; Sassone-Corsi, P. Molecular Cogs: Interplay between Circadian Clock and Cell Cycle. Trends Cell Biol. 2018, 28, 368–379. [Google Scholar] [CrossRef]
- Farshadi, E.; van der Horst, G.T.J.; Chaves, I. Molecular Links between the Circadian Clock and the Cell Cycle. J. Mol. Biol. 2020, 432, 3515–3524. [Google Scholar] [CrossRef]
- Yao, J.; He, C.; Zhao, W.; Hu, N.; Long, D. Circadian Clock and Cell Cycle: Cancer and Chronotherapy. Acta Histochem. 2021, 123, 151816. [Google Scholar] [CrossRef] [PubMed]
- Klevecz, R.R.; Li, C.M. Evolution of the Clock from Yeast to Man by Period-Doubling Folds in the Cellular Oscillator. Cold Spring Harb. Symp. Quant. Biol. 2007, 72, 421–429. [Google Scholar] [CrossRef] [PubMed]
- Roenneberg, T.; Merrow, M. Life before the Clock: Modeling Circadian Evolution. J. Biol. Rhythms 2002, 17, 495–505. [Google Scholar] [CrossRef]
- Brown, S.A.; Kowalska, E.; Dallmann, R. (Re)Inventing the Circadian Feedback Loop. Dev. Cell 2012, 22, 477–487. [Google Scholar] [CrossRef] [PubMed]
- Salichos, L.; Rokas, A. The Diversity and Evolution of Circadian Clock Proteins in Fungi. Mycologia 2010, 102, 269–278. [Google Scholar] [CrossRef]
- Camponeschi, I.; Montanari, A.; Mazzoni, C.; Bianchi, M.M. Light Stress in Yeasts: Signaling and Responses in Creatures of the Night. Int. J. Mol. Sci. 2023, 24, 6929. [Google Scholar] [CrossRef]
- Garmendia-Torres, C.; Goldbeter, A.; Jacquet, M. Nucleocytoplasmic Oscillations of the Yeast Transcription Factor Msn2: Evidence for Periodic PKA Activation. Curr. Biol. CB 2007, 17, 1044–1049. [Google Scholar] [CrossRef]
- Kuang, Z.; Pinglay, S.; Ji, H.; Boeke, J.D. Msn2/4 Regulate Expression of Glycolytic Enzymes and Control Transition from Quiescence to Growth. eLife 2017, 6, e29938. [Google Scholar] [CrossRef]
- Hirayama, J.; Cho, S.; Sassone-Corsi, P. Circadian Control by the Reduction/Oxidation Pathway: Catalase Represses Light-Dependent Clock Gene Expression in the Zebrafish. Proc. Natl. Acad. Sci. USA 2007, 104, 15747–15752. [Google Scholar] [CrossRef]
- Homma, T.; Okano, S.; Lee, J.; Ito, J.; Otsuki, N.; Kurahashi, T.; Kang, E.S.; Nakajima, O.; Fujii, J. SOD1 Deficiency Induces the Systemic Hyperoxidation of Peroxiredoxin in the Mouse. Biochem. Biophys. Res. Commun. 2015, 463, 1040–1046. [Google Scholar] [CrossRef]
- Ślesak, I.; Ślesak, H.; Kruk, J. Oxygen and Hydrogen Peroxide in the Early Evolution of Life on Earth: In Silico Comparative Analysis of Biochemical Pathways. Astrobiology 2012, 12, 775–784. [Google Scholar] [CrossRef]
- He, H.; Wu, X.; Xian, H.; Zhu, J.; Yang, Y.; Lv, Y.; Li, Y.; Konhauser, K.O. An Abiotic Source of Archean Hydrogen Peroxide and Oxygen That Pre-Dates Oxygenic Photosynthesis. Nat. Commun. 2021, 12, 6611. [Google Scholar] [CrossRef]
- He, H.; Wu, X.; Zhu, J.; Lin, M.; Lv, Y.; Xian, H.; Yang, Y.; Lin, X.; Li, S.; Li, Y.; et al. A Mineral-Based Origin of Earth’s Initial Hydrogen Peroxide and Molecular Oxygen. Proc. Natl. Acad. Sci. USA 2023, 120, e2221984120. [Google Scholar] [CrossRef]
- Haqq-Misra, J.; Kasting, J.F.; Lee, S. Availability of O2 and H2O2 on Pre-Photosynthetic Earth. Astrobiology 2011, 11, 293–302. [Google Scholar] [CrossRef]
- Borda, M.J.; Elsetinow, A.R.; Schoonen, M.A.; Strongin, D.R. Pyrite-Induced Hydrogen Peroxide Formation as a Driving Force in the Evolution of Photosynthetic Organisms on an Early Earth. Astrobiology 2001, 1, 283–288. [Google Scholar] [CrossRef]
- Inupakutika, M.A.; Sengupta, S.; Devireddy, A.R.; Azad, R.K.; Mittler, R. The Evolution of Reactive Oxygen Species Metabolism. J. Exp. Bot. 2016, 67, 5933–5943. [Google Scholar] [CrossRef] [PubMed]
- Rutter, J.; Reick, M.; McKnight, S.L. Metabolism and the Control of Circadian Rhythms. Annu. Rev. Biochem. 2002, 71, 307–331. [Google Scholar] [CrossRef] [PubMed]
- Putker, M.; O’Neill, J.S. Reciprocal Control of the Circadian Clock and Cellular Redox State—A Critical Appraisal. Mol. Cells 2016, 39, 6–19. [Google Scholar] [CrossRef] [PubMed]
- Tu, B.P.; McKnight, S.L. Metabolic Cycles as an Underlying Basis of Biological Oscillations. Nat. Rev. Mol. Cell Biol. 2006, 7, 696–701. [Google Scholar] [CrossRef]
- Reddy, A.B.; O’Neill, J.S. Metaclocks. EMBO Rep. 2011, 12, 612. [Google Scholar] [CrossRef]
- O’Neill, J.S.; Feeney, K.A. Circadian Redox and Metabolic Oscillations in Mammalian Systems. Antioxid. Redox Signal. 2014, 20, 2966–2981. [Google Scholar] [CrossRef] [PubMed]
- Lakin-Thomas, P.L. Transcriptional Feedback Oscillators: Maybe, Maybe Not. J. Biol. Rhythms 2006, 21, 83–92. [Google Scholar] [CrossRef]
- Rutter, J.; Reick, M.; Wu, L.C.; McKnight, S.L. Regulation of Clock and NPAS2 DNA Binding by the Redox State of NAD Cofactors. Science 2001, 293, 510–514. [Google Scholar] [CrossRef] [PubMed]
- Kojima, S.; Shingle, D.L.; Green, C.B. Post-Transcriptional Control of Circadian Rhythms. J. Cell Sci. 2011, 124, 311–320. [Google Scholar] [CrossRef]
- Rey, G.; Reddy, A.B. Connecting Cellular Metabolism to Circadian Clocks. Trends Cell Biol. 2013, 23, 234–241. [Google Scholar] [CrossRef] [PubMed]
- Bass, J. Circadian Topology of Metabolism. Nature 2012, 491, 348–356. [Google Scholar] [CrossRef]
- Alamoudi, A.A. Why Do Cancer Cells Break from Host Circadian Rhythm? Insights from Unicellular Organisms. BioEssays News Rev. Mol. Cell. Dev. Biol. 2021, 43, e2000205. [Google Scholar] [CrossRef]
- Sandbichler, A.M.; Jansen, B.; Peer, B.A.; Paulitsch, M.; Pelster, B.; Egg, M. Metabolic Plasticity Enables Circadian Adaptation to Acute Hypoxia in Zebrafish Cells. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2018, 46, 1159–1174. [Google Scholar] [CrossRef]
- Kitayama, Y.; Nishiwaki, T.; Terauchi, K.; Kondo, T. Dual KaiC-Based Oscillations Constitute the Circadian System of Cyanobacteria. Genes Dev. 2008, 22, 1513–1521. [Google Scholar] [CrossRef]
- Rey, G.; Reddy, A.B. Interplay between Cellular Redox Oscillations and Circadian Clocks. Diabetes Obes. Metab. 2015, 17 (Suppl. 1), 55–64. [Google Scholar] [CrossRef]
- Cho, C.-S.; Yoon, H.J.; Kim, J.Y.; Woo, H.A.; Rhee, S.G. Circadian Rhythm of Hyperoxidized Peroxiredoxin II Is Determined by Hemoglobin Autoxidation and the 20S Proteasome in Red Blood Cells. Proc. Natl. Acad. Sci. USA 2014, 111, 12043. [Google Scholar] [CrossRef]
- O’Neill, J.S.; Maywood, E.S.; Hastings, M.H. Cellular Mechanisms of Circadian Pacemaking: Beyond Transcriptional Loops. Handb. Exp. Pharmacol. 2013, 217, 67–103. [Google Scholar] [CrossRef]
- van Ooijen, G.; Dixon, L.E.; Troein, C.; Millar, A.J. Proteasome Function Is Required for Biological Timing throughout the Twenty-Four Hour Cycle. Curr. Biol. 2011, 21, 869–875. [Google Scholar] [CrossRef] [PubMed]
- Stincone, A.; Prigione, A.; Cramer, T.; Wamelink, M.M.C.; Campbell, K.; Cheung, E.; Olin-Sandoval, V.; Grüning, N.-M.; Krüger, A.; Alam, M.T.; et al. The Return of Metabolism: Biochemistry and Physiology of the Pentose Phosphate Pathway. Biol. Rev. Camb. Philos. Soc. 2015, 90, 927–963. [Google Scholar] [CrossRef]
- Rey, G.; Valekunja, U.K.; Feeney, K.A.; Wulund, L.; Milev, N.B.; Stangherlin, A.; Ansel-Bollepalli, L.; Velagapudi, V.; O’Neill, J.S.; Reddy, A.B. The Pentose Phosphate Pathway Regulates the Circadian Clock. Cell Metab. 2016, 24, 462–473. [Google Scholar] [CrossRef]
- Tu, B.P.; Mohler, R.E.; Liu, J.C.; Dombek, K.M.; Young, E.T.; Synovec, R.E.; McKnight, S.L. Cyclic Changes in Metabolic State during the Life of a Yeast Cell. Proc. Natl. Acad. Sci. USA 2007, 104, 16886–16891. [Google Scholar] [CrossRef] [PubMed]
- Ch, R.; Rey, G.; Ray, S.; Jha, P.K.; Driscoll, P.C.; Dos Santos, M.S.; Malik, D.M.; Lach, R.; Weljie, A.M.; MacRae, J.I.; et al. Rhythmic Glucose Metabolism Regulates the Redox Circadian Clockwork in Human Red Blood Cells. Nat. Commun. 2021, 12, 377. [Google Scholar] [CrossRef]
- Paulose, J.K.; Rucker, E.B.; Cassone, V.M. Toward the Beginning of Time: Circadian Rhythms in Metabolism Precede Rhythms in Clock Gene Expression in Mouse Embryonic Stem Cells. PLoS ONE 2012, 7, e49555. [Google Scholar] [CrossRef]
- Rey, G.; Milev, N.B.; Valekunja, U.K.; Ch, R.; Ray, S.; Silva Dos Santos, M.; Nagy, A.D.; Antrobus, R.; MacRae, J.I.; Reddy, A.B. Metabolic Oscillations on the Circadian Time Scale in Drosophila Cells Lacking Clock Genes. Mol. Syst. Biol. 2018, 14, e8376. [Google Scholar] [CrossRef]
- Swan, J.A.; Golden, S.S.; LiWang, A.; Partch, C.L. Structure, Function, and Mechanism of the Core Circadian Clock in Cyanobacteria. J. Biol. Chem. 2018, 293, 5026–5034. [Google Scholar] [CrossRef]
- Moreno-Cabezuelo, J.Á.; Gómez-Baena, G.; Díez, J.; García-Fernández, J.M. Integrated Proteomic and Metabolomic Analyses Show Differential Effects of Glucose Availability in Marine Synechococcus and Prochlorococcus. Microbiol. Spectr. 2023, 11, e0327522. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.-I.; Vinyard, D.J.; Ananyev, G.M.; Dismukes, G.C.; Golden, S.S. Oxidized Quinones Signal Onset of Darkness Directly to the Cyanobacterial Circadian Oscillator. Proc. Natl. Acad. Sci. USA 2012, 109, 17765–17769. [Google Scholar] [CrossRef] [PubMed]
- Rust, M.J.; Golden, S.S.; O’Shea, E.K. Light-Driven Changes in Energy Metabolism Directly Entrain the Cyanobacterial Circadian Oscillator. Science 2011, 331, 220–223. [Google Scholar] [CrossRef] [PubMed]
- Pattanayak, G.K.; Phong, C.; Rust, M.J. Rhythms in Energy Storage Control the Ability of the Cyanobacterial Circadian Clock to Reset. Curr. Biol. CB 2014, 24, 1934–1938. [Google Scholar] [CrossRef]
- Shinde, S.; Zhang, X.; Singapuri, S.P.; Kalra, I.; Liu, X.; Morgan-Kiss, R.M.; Wang, X. Glycogen Metabolism Supports Photosynthesis Start through the Oxidative Pentose Phosphate Pathway in Cyanobacteria. Plant Physiol. 2020, 182, 507–517. [Google Scholar] [CrossRef]
- Pattanayak, G.K.; Lambert, G.; Bernat, K.; Rust, M.J. Controlling the Cyanobacterial Clock by Synthetically Rewiring Metabolism. Cell Rep. 2015, 13, 2362–2367. [Google Scholar] [CrossRef]
- Reddy, A.B. Redox and Metabolic Oscillations in the Clockwork. In A Time for Metabolism and Hormones; Sassone-Corsi, P., Christen, Y., Eds.; Springer: Cham, Switzerland, 2016. [Google Scholar] [CrossRef]
- Lakin-Thomas, P. Circadian Rhythms, Metabolic Oscillators, and the Target of Rapamycin (TOR) Pathway: The Neurospora Connection. Curr. Genet. 2019, 65, 339–349. [Google Scholar] [CrossRef]
- Eskandari, R.; Ratnayake, L.; Lakin-Thomas, P.L. Shared Components of the FRQ-Less Oscillator and TOR Pathway Maintain Rhythmicity in Neurospora. J. Biol. Rhythms 2021, 36, 329–345. [Google Scholar] [CrossRef]
- Ratnayake, L.; Adhvaryu, K.K.; Kafes, E.; Motavaze, K.; Lakin-Thomas, P. A Component of the TOR (Target Of Rapamycin) Nutrient-Sensing Pathway Plays a Role in Circadian Rhythmicity in Neurospora Crassa. PLoS Genet. 2018, 14, e1007457. [Google Scholar] [CrossRef]
- Roenneberg, T.; Merrow, M. The Network of Time: Understanding the Molecular Circadian System. Curr. Biol. CB 2003, 13, R198–R207. [Google Scholar] [CrossRef]
- Roenneberg, T.; Merrow, M. Molecular Circadian Oscillators: An Alternative Hypothesis. J. Biol. Rhythms 1998, 13, 167–179. [Google Scholar] [CrossRef]
- Golden, S.S.; Canales, S.R. Cyanobacterial Circadian Clocks--Timing Is Everything. Nat. Rev. Microbiol. 2003, 1, 191–199. [Google Scholar] [CrossRef]
- Huang, T.C.; Tu, J.; Chow, T.J.; Chen, T.H. Circadian Rhythm of the Prokaryote Synechococcus Sp. RF-1. Plant Physiol. 1990, 92, 531–533. [Google Scholar] [CrossRef]
- Grobbelaar, N.; Huang, T.C.; Lin, H.Y.; Chow, T.J. Dinitrogen-Fixing Endogenous Rhythm in Synechococcus RF-1. FEMS Microbiol. Lett. 1986, 37, 173–177. [Google Scholar] [CrossRef]
- Iwasaki, H. A Retrospective: On Disproving the Transcription–Translation Feedback Loop Model in Cyanobacteria. In Circadian Rhythms in Bacteria and Microbiomes; Johnson, C.H., Rust, M.J., Eds.; Springer International Publishing: Cham, Switzerland, 2021; pp. 53–65. [Google Scholar] [CrossRef]
- Teng, S.-W.; Mukherji, S.; Moffitt, J.R.; de Buyl, S.; O’Shea, E.K. Robust Circadian Oscillations in Growing Cyanobacteria Require Transcriptional Feedback. Science 2013, 340, 737–740. [Google Scholar] [CrossRef]
- Zwicker, D.; Lubensky, D.K.; ten Wolde, P.R. Robust Circadian Clocks from Coupled Protein-Modification and Transcription-Translation Cycles. Proc. Natl. Acad. Sci. USA 2010, 107, 22540–22545. [Google Scholar] [CrossRef] [PubMed]
- Mihalcescu, I.; Hsing, W.; Leibler, S. Resilient Circadian Oscillator Revealed in Individual Cyanobacteria. Nature 2004, 430, 81–85. [Google Scholar] [CrossRef] [PubMed]
- Roenneberg, T.; Merrow, M. Circadian Systems and Metabolism. J. Biol. Rhythms 1999, 14, 449–459. [Google Scholar] [CrossRef]
- Axmann, I.M.; Dühring, U.; Seeliger, L.; Arnold, A.; Vanselow, J.T.; Kramer, A.; Wilde, A. Biochemical Evidence for a Timing Mechanism in Prochlorococcus. J. Bacteriol. 2009, 191, 5342–5347. [Google Scholar] [CrossRef]
- Ray, S.; Valekunja, U.K.; Stangherlin, A.; Howell, S.A.; Snijders, A.P.; Damodaran, G.; Reddy, A.B. Circadian Rhythms in the Absence of the Clock Gene Bmal1. Science 2020, 367, 800–806. [Google Scholar] [CrossRef]
- Abruzzi, K.C.; Gobet, C.; Naef, F.; Rosbash, M. Comment on “Circadian Rhythms in the Absence of the Clock Gene Bmal1”. Science 2021, 372, eabf0922. [Google Scholar] [CrossRef] [PubMed]
- Ness-Cohn, E.; Allada, R.; Braun, R. Comment on “Circadian Rhythms in the Absence of the Clock Gene Bmal1”. Science 2021, 372, eabe9230. [Google Scholar] [CrossRef] [PubMed]
- Ray, S.; Valekunja, U.K.; Stangherlin, A.; Howell, S.A.; Snijders, A.P.; Damodaran, G.; Reddy, A.B. Response to Comment on “Circadian Rhythms in the Absence of the Clock Gene Bmal1”. Science 2021, 372, eabf1941. [Google Scholar] [CrossRef] [PubMed]
- Ray, S.; Valekunja, U.K.; Stangherlin, A.; Howell, S.A.; Snijders, A.P.; Damodaran, G.; Reddy, A.B. Response to Comment on “Circadian Rhythms in the Absence of the Clock Gene Bmal1”. Science 2021, 372, eabf1930. [Google Scholar] [CrossRef] [PubMed]
- Byndloss, M.X.; Olsan, E.E.; Rivera-Chávez, F.; Tiffany, C.R.; Cevallos, S.A.; Lokken, K.L.; Torres, T.P.; Byndloss, A.J.; Faber, F.; Gao, Y.; et al. Microbiota-Activated PPAR-γ-Signaling Inhibits Dysbiotic Enterobacteriaceae Expansion. Science 2017, 357, 570–575. [Google Scholar] [CrossRef]
- Miller, B.M.; Liou, M.J.; Zhang, L.F.; Nguyen, H.; Litvak, Y.; Schorr, E.-M.; Jang, K.K.; Tiffany, C.R.; Butler, B.P.; Bäumler, A.J. Anaerobic Respiration of NOX1-Derived Hydrogen Peroxide Licenses Bacterial Growth at the Colonic Surface. Cell Host Microbe 2020, 28, 789–797.e5. [Google Scholar] [CrossRef]
- Sommer, F.; Bäckhed, F. The Gut Microbiota--Masters of Host Development and Physiology. Nat. Rev. Microbiol. 2013, 11, 227–238. [Google Scholar] [CrossRef]
- Herfindal, A.M.; Rocha, S.D.C.; Papoutsis, D.; Bøhn, S.K.; Carlsen, H. The ROS-Generating Enzyme NADPH Oxidase 1 Modulates the Colonic Microbiota but Offers Minor Protection against Dextran Sulfate Sodium-Induced Low-Grade Colon Inflammation in Mice. Free Radic. Biol. Med. 2022, 188, 298–311. [Google Scholar] [CrossRef]
- Yang, Y.; Yu, P.; Lu, Y.; Gao, C.; Sun, Q. Disturbed Rhythmicity of Intestinal Hydrogen Peroxide Alters Gut Microbial Oscillations in BMAL1-Deficient Monkeys. Cell Rep. 2023, 42, 112183. [Google Scholar] [CrossRef]
- Lee, Y.; Wisor, J.P. Multi-Modal Regulation of Circadian Physiology by Interactive Features of Biological Clocks. Biology 2021, 11, 21. [Google Scholar] [CrossRef]
- Qian, H.; Hu, B.; Yu, S.; Pan, X.; Wu, T.; Fu, Z. The Effects of Hydrogen Peroxide on the Circadian Rhythms of Microcystis Aeruginosa. PLoS ONE 2012, 7, e33347. [Google Scholar] [CrossRef]
- Huus, K.E.; Ley, R.E. Blowing Hot and Cold: Body Temperature and the Microbiome. mSystems 2021, 6, e00707-21. [Google Scholar] [CrossRef] [PubMed]
- Hylander, B.L.; Repasky, E.A. Temperature as a Modulator of the Gut Microbiome: What Are the Implications and Opportunities for Thermal Medicine? Int. J. Hyperth. Off. J. Eur. Soc. Hyperthermic Oncol. N. Am. Hyperth. Group 2019, 36, 83–89. [Google Scholar] [CrossRef]
- Bongers, K.S.; Chanderraj, R.; Woods, R.J.; McDonald, R.A.; Adame, M.D.; Falkowski, N.R.; Brown, C.A.; Baker, J.M.; Winner, K.M.; Fergle, D.J.; et al. The Gut Microbiome Modulates Body Temperature Both in Sepsis and Health. Am. J. Respir. Crit. Care Med. 2023, 207, 1030–1041. [Google Scholar] [CrossRef] [PubMed]
- Kullberg, R.F.J.; Wiersinga, W.J. Good Bug, Bad Bug: What Is Influencing the Fever Response during Infection? Am. J. Respir. Crit. Care Med. 2022, 207, 967–969. [Google Scholar] [CrossRef] [PubMed]
- Paulose, J.K.; Cassone, C.V.; Cassone, V.M. Aging, Melatonin Biosynthesis, and Circadian Clockworks in the Gastrointestinal System of the Laboratory Mouse. Physiol. Genom. 2019, 51, 1–9. [Google Scholar] [CrossRef]
- Konturek, S.J.; Konturek, P.C.; Brzozowska, I.; Pawlik, M.; Sliwowski, Z.; Cześnikiewicz-Guzik, M.; Kwiecień, S.; Brzozowski, T.; Bubenik, G.A.; Pawlik, W.W. Localization and Biological Activities of Melatonin in Intact and Diseased Gastrointestinal Tract (GIT). J. Physiol. Pharmacol. Off. J. Pol. Physiol. Soc. 2007, 58, 381–405. [Google Scholar]
- Yin, J.; Li, Y.; Han, H.; Ma, J.; Liu, G.; Wu, X.; Huang, X.; Fang, R.; Baba, K.; Bin, P.; et al. Administration of Exogenous Melatonin Improves the Diurnal Rhythms of the Gut Microbiota in Mice Fed a High-Fat Diet. mSystems 2020, 5, e00002-20. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Siebieszuk, A.; Sejbuk, M.; Witkowska, A.M. Studying the Human Microbiota: Advances in Understanding the Fundamentals, Origin, and Evolution of Biological Timekeeping. Int. J. Mol. Sci. 2023, 24, 16169. https://doi.org/10.3390/ijms242216169
Siebieszuk A, Sejbuk M, Witkowska AM. Studying the Human Microbiota: Advances in Understanding the Fundamentals, Origin, and Evolution of Biological Timekeeping. International Journal of Molecular Sciences. 2023; 24(22):16169. https://doi.org/10.3390/ijms242216169
Chicago/Turabian StyleSiebieszuk, Adam, Monika Sejbuk, and Anna Maria Witkowska. 2023. "Studying the Human Microbiota: Advances in Understanding the Fundamentals, Origin, and Evolution of Biological Timekeeping" International Journal of Molecular Sciences 24, no. 22: 16169. https://doi.org/10.3390/ijms242216169