
Diminished Ovarian Reserve in Endometriosis: Insights from In Vitro, In Vivo, and Human Studies—A Systematic Review

Abstract
:1. Introduction
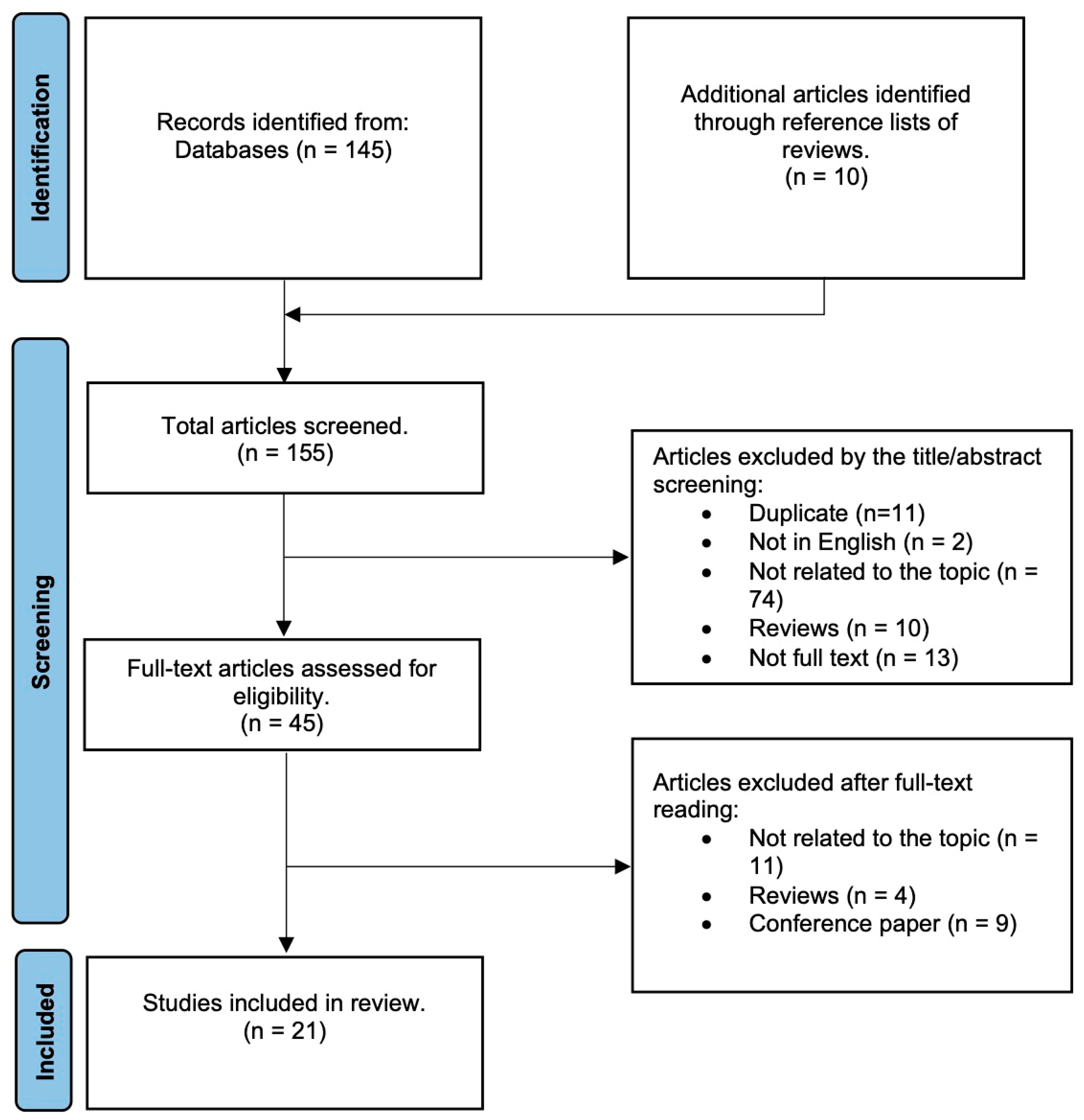
2. Methods
3. Results and Discussion
3.1. Characteristics of Studies on the Effect of Endometriosis and Its Therapeutic Interventions on Ovarian Reserve
3.2. The Prediction of Ovarian Reserve
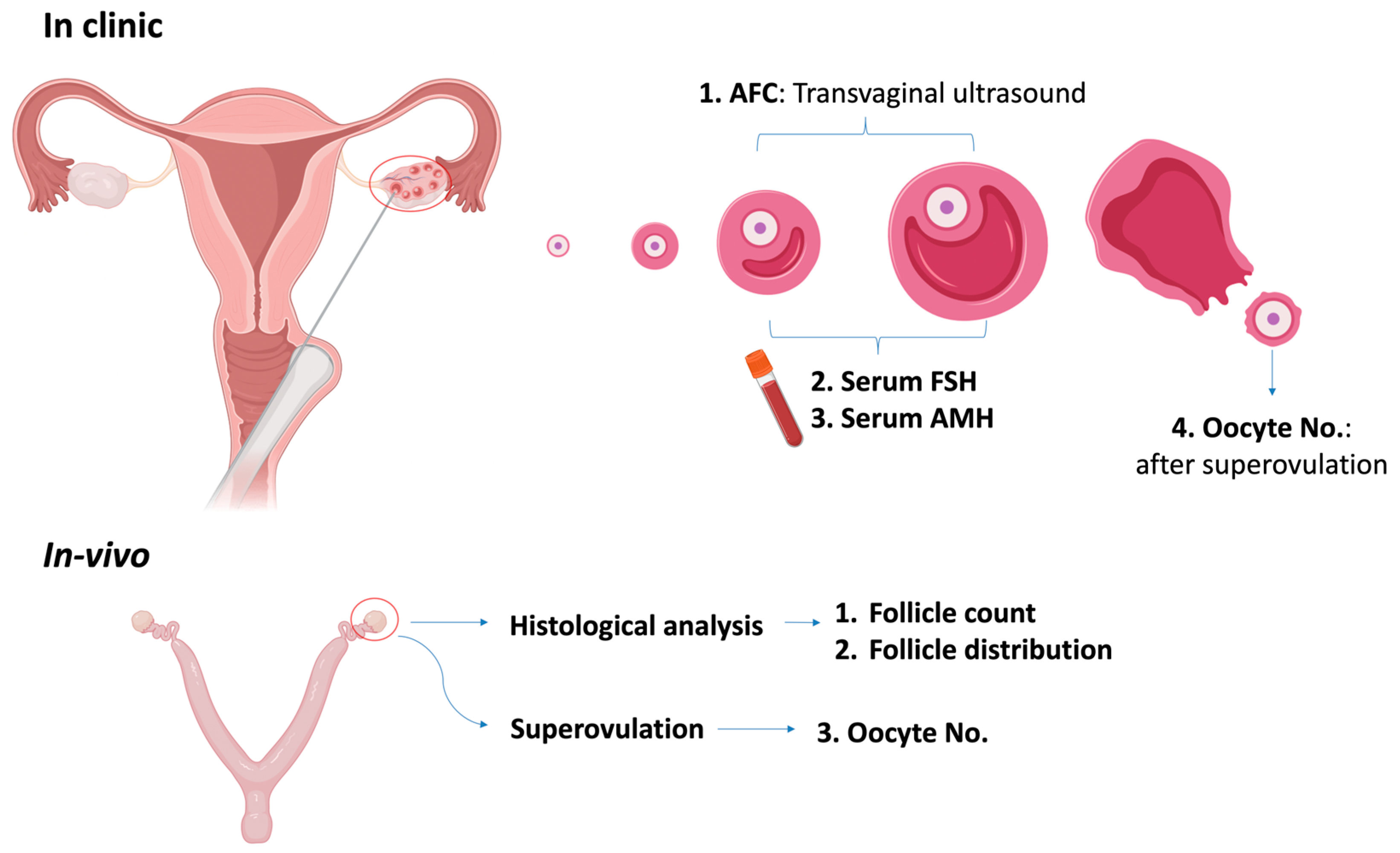
3.2.1. Quantitative Assessment
3.2.2. Qualitative Assessment
3.3. The Link of Endometriosis with Reduced Ovarian Reserve
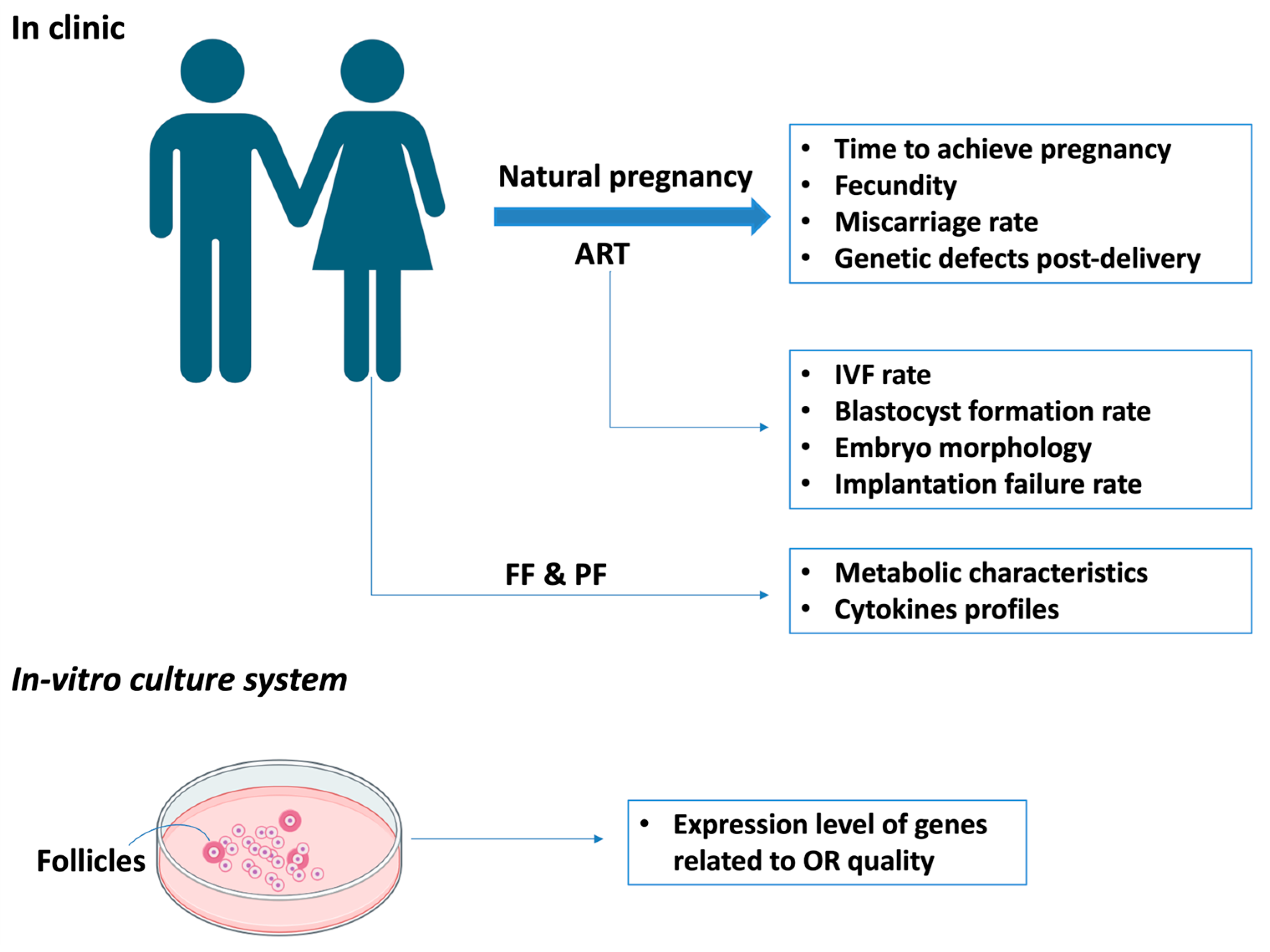
3.3.1. The Clinical Relevance of Endometriosis and Ovarian Reserve from Human Studies
Endometriosis Per Se
Endometriosis Treatments
- Medical Treatment
- Non-Steroidal Anti-Inflammatory Drugs (NSAIDs)
- Hormone Treatments
- 2.
- Surgery
3.3.2. The Relevance of Endometriosis and Ovarian Reserve from In Vitro and In Vivo Studies
Decreased Ovarian Reserve in In Vitro Studies of Endometriosis
Decreased Ovarian Reserve in In Vivo Studies of Endometriosis
3.4. Potential Mechanisms Underlying Diminished Ovarian Reserve in Endometriosis Based on In Vitro, In Vivo, and Human Studies
3.4.1. Inflammation, Oxidative Stress, Fibrosis, and DNA Damage
In Human Studies
In Vitro and In Vivo Studies
3.4.2. Dysregulated Ovulation and Ovarian Steroids Secretion
In Human Studies
In Vitro and In Vivo Studies
3.4.3. Hyperactivation of Primordial Follicle
In Human Studies
In Vitro and In Vivo Studies
3.5. Potential Therapeutic Interventions
3.5.1. Hyperactivation of Primordial Follicle
3.5.2. The Choice of Different Surgical Options Regarding Endometriosis
3.5.3. Therapeutic Targets to the Signaling Pathways Leading to Reduced OR in Women with Endometriosis
3.5.4. The Natural Products to Treat Endometriosis That Will Not Lead to Ovulation Inhibition
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Parasar, P.; Ozcan, P.; Terry, K.L. Endometriosis: Epidemiology, Diagnosis and Clinical Management. Curr. Obs. Gynecol. Rep. 2017, 6, 34–41. [Google Scholar] [CrossRef] [PubMed]
- Bulun, S.E. Endometriosis. N. Engl. J. Med. 2009, 360, 268–279. [Google Scholar] [CrossRef] [PubMed]
- The Practice Committee of the American Society for Reproductive Medicine. Testing and interpreting measures of ovarian reserve: A committee opinion. Fertil. Steril. 2012, 98, 1407–1415. [Google Scholar] [CrossRef] [PubMed]
- The Practice Committee of the American Society for Reproductive Medicine. Testing and interpreting measures of ovarian reserve: A committee opinion. Fertil. Steril. 2020, 114, 1151–1157. [Google Scholar] [CrossRef]
- Dunlop, C.E.; Anderson, R.A. The regulation and assessment of follicular growth. Scand. J. Clin. Lab. Investig. 2014, 244, 13–17; discussion 17. [Google Scholar] [CrossRef]
- Jones, R.E.; Lopez, K.H. Human Reproductive Biology, 4th ed.; Academic Press: Cambridge, MA, USA, 2013; pp. 1–381. [Google Scholar]
- Wang, X.; Wang, L.; Xiang, W. Mechanisms of ovarian aging in women: A review. J. Ovarian Res. 2023, 16, 67. [Google Scholar] [CrossRef]
- Tan, Z.; Gong, X.; Li, Y.; Hung, S.W.; Huang, J.; Wang, C.C.; Chung, J.P.W. Impacts of endometrioma on ovarian aging from basic science to clinical management. Front. Endocrinol. 2022, 13, 1073261. [Google Scholar] [CrossRef]
- Azhar, E.; Seifer, D.B.; Melzer, K.; Ahmed, A.; Weedon, J.; Minkoff, H. Knowledge of ovarian reserve and reproductive choices. J. Assist. Reprod. Genet. 2015, 32, 409–415. [Google Scholar] [CrossRef]
- Seyhan, A.; Ata, B.; Uncu, G. The Impact of Endometriosis and Its Treatment on Ovarian Reserve. Semin. Reprod. Med. 2015, 33, 422–428. [Google Scholar] [CrossRef]
- Moher, D.; Liberati, A.; Tetzlaff, J.; Altman, D.G. Preferred reporting items for systematic reviews and meta-analyses: The PRISMA statement. PLoS Med. 2009, 6, e1000097. [Google Scholar] [CrossRef]
- Tan, Z.; Hung, S.W.; Zheng, X.; Wang, C.C.; Chung, J.P.; Zhang, T. What We Have Learned from Animal Models to Understand the Etiology and Pathology of Endometrioma-Related Infertility. Biomedicines 2022, 10, 1483. [Google Scholar] [CrossRef]
- Kanellopoulos, D.; Karagianni, D.; Pergialiotis, V.; Nikiteas, N.; Lazaris, A.C.; Iliopoulos, D. The effect of endometriosis on fertility in an animal model. J. Med. Life 2022, 15, 1170–1175. [Google Scholar] [CrossRef]
- Park, Y.; Cho, Y.J.; Sung, N.; Park, M.J.; Guan, X.; Gibbons, W.E.; O’Malley, B.W.; Han, S.J. Oleuropein suppresses endometriosis progression and improves the fertility of mice with endometriosis. J. Biomed. Sci. 2022, 29, 100. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, S.; Nakamura, T.; Motooka, Y.; Ito, F.; Jiang, L.; Akatsuka, S.; Iwase, A.; Kajiyama, H.; Kikkawa, F.; Toyokuni, S. Novel ovarian endometriosis model causes infertility via iron-mediated oxidative stress in mice. Redox Biol. 2020, 37, 101726. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, A.; Koga, K.; Satake, E.; Makabe, T.; Taguchi, A.; Miyashita, M.; Takamura, M.; Harada, M.; Hirata, T.; Hirota, Y.; et al. Endometriosis Triggers Excessive Activation of Primordial Follicles via PI3K-PTEN-Akt-Foxo3 Pathway. J. Clin. Endocrinol. Metab. 2019, 104, 5547–5554. [Google Scholar] [CrossRef]
- Kacan, T.; Yildiz, C.; Baloglu Kacan, S.; Seker, M.; Ozer, H.; Cetin, A. Everolimus as an mTOR Inhibitor Suppresses Endometriotic Implants: An Experimental Rat Study. Geburtshilfe Frauenheilkde 2017, 77, 66–72. [Google Scholar] [CrossRef]
- Ozer, H.; Boztosun, A.; Acmaz, G.; Atilgan, R.; Akkar, O.B.; Kosar, M.I. The efficacy of bevacizumab, sorafenib, and retinoic acid on rat endometriosis model. Reprod. Sci. 2013, 20, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Stilley, J.A.; Birt, J.A.; Nagel, S.C.; Sutovsky, M.; Sutovsky, P.; Sharpe-Timms, K.L. Neutralizing TIMP1 restores fecundity in a rat model of endometriosis and treating control rats with TIMP1 causes anomalies in ovarian function and embryo development. Biol. Reprod. 2010, 83, 185–194. [Google Scholar] [CrossRef]
- Stilley, J.A.; Woods-Marshall, R.; Sutovsky, M.; Sutovsky, P.; Sharpe-Timms, K.L. Reduced fecundity in female rats with surgically induced endometriosis and in their daughters: A potential role for tissue inhibitors of metalloproteinase 1. Biol. Reprod. 2009, 80, 649–656. [Google Scholar] [CrossRef]
- D’Hooghe, T.M.; Bambra, C.S.; Raeymaekers, B.M.; Riday, A.M.; Suleman, M.A.; Koninckx, P.R. The cycle pregnancy rate is normal in baboons with stage I endometriosis but decreased in primates with stage II and stage III–IV disease. Fertil. Steril. 1996, 66, 809–813. [Google Scholar] [CrossRef] [PubMed]
- Barragan, J.C.; Brotons, J.; Ruiz, J.A.; Acien, P. Experimentally induced endometriosis in rats: Effect on fertility and the effects of pregnancy and lactation on the ectopic endometrial tissue. Fertil. Steril. 1992, 58, 1215–1219. [Google Scholar] [CrossRef]
- Schenken, R.S.; Williams, R.F.; Hodgen, G.D. Experimental endometriosis in monkeys. Ann. N. Y. Acad. Sci. 1991, 622, 242–255. [Google Scholar] [CrossRef]
- Muraoka, A.; Osuka, S.; Yabuki, A.; Bayasula; Yoshihara, M.; Tanaka, H.; Sonehara, R.; Miyake, N.; Murakami, M.; Yoshita, S.; et al. Impact of perioperative use of GnRH agonist or dienogest on ovarian reserve after cystectomy for endometriomas: A randomized controlled trial. Reprod. Biol. Endocrinol. 2021, 19, 179. [Google Scholar] [CrossRef] [PubMed]
- Sireesha, M.U.; Chitra, T.; Subbaiah, M.; Nandeesha, H. Effect of Laparoscopic Ovarian Cystectomy on Ovarian Reserve in Benign Ovarian Cysts. J. Hum. Reprod. Sci. 2021, 14, 56–60. [Google Scholar]
- Barra, F.; Lagana, A.S.; Scala, C.; Garzon, S.; Ghezzi, F.; Ferrero, S. Pretreatment with dienogest in women with endometriosis undergoing IVF after a previous failed cycle. Reprod. Biomed. Online 2020, 41, 859–868. [Google Scholar] [CrossRef]
- Karadag, C.; Yoldemir, T.; Demircan Karadag, S.; Turgut, A. The effects of endometrioma size and bilaterality on ovarian reserve. J. Obstet. Gynaecol. 2020, 40, 531–536. [Google Scholar] [CrossRef]
- Candiani, M.; Ottolina, J.; Posadzka, E.; Ferrari, S.; Castellano, L.M.; Tandoi, I.; Pagliardini, L.; Nocun, A.; Jach, R. Assessment of ovarian reserve after cystectomy versus ‘one-step’ laser vaporization in the treatment of ovarian endometrioma: A small randomized clinical trial. Hum. Reprod. 2018, 33, 2205–2211. [Google Scholar] [CrossRef]
- Kasapoglu, I.; Ata, B.; Uyaniklar, O.; Seyhan, A.; Orhan, A.; Yildiz Oguz, S.; Uncu, G. Endometrioma-related reduction in ovarian reserve (ERROR): A prospective longitudinal study. Fertil. Steril. 2018, 110, 122–127. [Google Scholar] [CrossRef] [PubMed]
- Muller, V.; Kogan, I.; Yarmolinskaya, M.; Niauri, D.; Gzgzyan, A.; Aylamazyan, E. Dienogest treatment after ovarian endometrioma removal in infertile women prior to IVF. Gynecol. Endocrinol. 2017, 33 (Suppl. S1), 18–21. [Google Scholar] [CrossRef]
- Taniguchi, F.; Enatsu, A.; Ota, I.; Toda, T.; Arata, K.; Harada, T. Effects of low dose oral contraceptive pill containing drospirenone/ethinylestradiol in patients with endometrioma. Eur. J. Obstet. Gynecol. Reprod. Biol. 2015, 191, 116–120. [Google Scholar] [CrossRef] [PubMed]
- Lima, M.L.; Martins, W.P.; Coelho Neto, M.A.; Nastri, C.O.; Ferriani, R.A.; Navarro, P.A. Assessment of ovarian reserve by antral follicle count in ovaries with endometrioma. Ultrasound Obstet. Gynecol. 2015, 46, 239–242. [Google Scholar] [CrossRef] [PubMed]
- Li, C.Z.; Liu, B.; Wen, Z.Q.; Sun, Q. The impact of electrocoagulation on ovarian reserve after laparoscopic excision of ovarian cysts: A prospective clinical study of 191 patients. Fertil. Steril. 2009, 92, 1428–1435. [Google Scholar] [CrossRef] [PubMed]
- Khan, H.L.; Bhatti, S.; Suhail, S.; Gul, R.; Awais, A.; Hamayun, H.; Enver, F.; Abbas, S.; Hassan, Z.; Nisar, R.; et al. Antral follicle count (AFC) and serum anti-Müllerian hormone (AMH) are the predictors of natural fecundability have similar trends irrespective of fertility status and menstrual characteristics among fertile and infertile women below the age of 40 years. Reprod. Biol. Endocrinol. 2019, 17, 20. [Google Scholar] [CrossRef]
- Coelho Neto, M.A.; Ludwin, A.; Borrell, A.; Benacerraf, B.; Dewailly, D.; da Silva Costa, F.; Condous, G.; Alcazar, J.L.; Jokubkiene, L.; Guerriero, S.; et al. Counting ovarian antral follicles by ultrasound: A practical guide. Ultrasound Obstet. Gynecol. 2018, 51, 10–20. [Google Scholar] [CrossRef]
- Halis, G.; Arici, A. Endometriosis and inflammation in infertility. Ann. N. Y. Acad. Sci. 2004, 1034, 300–315. [Google Scholar] [CrossRef] [PubMed]
- Riccabona, M. Basics, principles, techniques and modern methods in paediatric ultrasonography. Eur. J. Radiol. 2014, 83, 1487–1494. [Google Scholar] [CrossRef]
- Park, S.R.; Kim, S.R.; Kim, S.K.; Park, J.R.; Hong, I.S. A novel role of follicle-stimulating hormone (FSH) in various regeneration-related functions of endometrial stem cells. Exp. Mol. Med. 2022, 54, 1524–1535. [Google Scholar] [CrossRef]
- Kessel, B.; Jia, X.-C.; Davoren, J.B.; Hsueh, A.J.W. The Ovarian Granulosa Cell as a Follicle-Stimulating Hormone Target Tissue. In Endocrinology and Physiology of Reproduction; Leung, P.C.K., Armstrong, D.T., Ruf, K.B., Moger, W.H., Friesen, H.G., Eds.; Springer: Boston, MA, USA, 1987; pp. 163–176. [Google Scholar]
- Allan, C.M.; Wang, Y.; Jimenez, M.; Marshan, B.; Spaliviero, J.; Illingworth, P.; Handelsman, D.J. Follicle-stimulating hormone increases primordial follicle reserve in mature female hypogonadal mice. J. Endocrinol. 2006, 188, 549–557. [Google Scholar] [CrossRef]
- Streuli, I.; Fraisse, T.; Pillet, C.; Ibecheole, V.; Bischof, P.; de Ziegler, D. Serum antimüllerian hormone levels remain stable throughout the menstrual cycle and after oral or vaginal administration of synthetic sex steroids. Fertil. Steril. 2008, 90, 395–400. [Google Scholar] [CrossRef]
- Arslan, A.A.; Zeleniuch-Jacquotte, A.; Lukanova, A.; Rinaldi, S.; Kaaks, R.; Toniolo, P. Reliability of follicle-stimulating hormone measurements in serum. Reprod. Biol. Endocrinol. 2003, 1, 49. [Google Scholar] [CrossRef] [PubMed]
- Jirge, P.R. Ovarian reserve tests. J. Hum. Reprod. Sci. 2011, 4, 108–113. [Google Scholar] [CrossRef]
- Ata, B.; Seyhan, A.; Seli, E. Diminished ovarian reserve versus ovarian aging: Overlaps and differences. Curr. Opin. Obstet. Gynecol. 2019, 31, 139–147. [Google Scholar] [CrossRef]
- Weenen, C.; Laven, J.S.; Von Bergh, A.R.; Cranfield, M.; Groome, N.P.; Visser, J.A.; Kramer, P.; Fauser, B.C.; Themmen, A.P. Anti-Müllerian hormone expression pattern in the human ovary: Potential implications for initial and cyclic follicle recruitment. Mol. Hum. Reprod. 2004, 10, 77–83. [Google Scholar] [CrossRef]
- Jeppesen, J.V.; Anderson, R.A.; Kelsey, T.W.; Christiansen, S.L.; Kristensen, S.G.; Jayaprakasan, K.; Raine-Fenning, N.; Campbell, B.K.; Yding Andersen, C. Which follicles make the most anti-Mullerian hormone in humans? Evidence for an abrupt decline in AMH production at the time of follicle selection. Mol. Hum. Reprod. 2013, 19, 519–527. [Google Scholar] [CrossRef]
- Durlinger, A.L.; Gruijters, M.J.; Kramer, P.; Karels, B.; Ingraham, H.A.; Nachtigal, M.W.; Uilenbroek, J.T.; Grootegoed, J.A.; Themmen, A.P. Anti-Müllerian hormone inhibits initiation of primordial follicle growth in the mouse ovary. Endocrinology 2002, 143, 1076–1084. [Google Scholar] [CrossRef]
- Cook, C.L.; Siow, Y.; Taylor, S.; Fallat, M.E. Serum müllerian-inhibiting substance levels during normal menstrual cycles. Fertil. Steril. 2000, 73, 859–861. [Google Scholar] [CrossRef]
- Van Disseldorp, J.; Lambalk, C.B.; Kwee, J.; Looman, C.W.; Eijkemans, M.J.; Fauser, B.C.; Broekmans, F.J. Comparison of inter- and intra-cycle variability of anti-Mullerian hormone and antral follicle counts. Hum. Reprod. 2010, 25, 221–227. [Google Scholar] [CrossRef]
- Broer, S.L.; Dólleman, M.; van Disseldorp, J.; Broeze, K.A.; Opmeer, B.C.; Bossuyt, P.M.; Eijkemans, M.J.; Mol, B.W.; Broekmans, F.J. Prediction of an excessive response in in vitro fertilization from patient characteristics and ovarian reserve tests and comparison in subgroups: An individual patient data meta-analysis. Fertil. Steril. 2013, 100, 420–429.e7. [Google Scholar] [CrossRef]
- De Vet, A.; Laven, J.S.; de Jong, F.H.; Themmen, A.P.; Fauser, B.C. Antimullerian hormone serum levels: A putative marker for ovarian aging. Fertil. Steril. 2002, 77, 357–362. [Google Scholar] [CrossRef]
- Iliodromiti, S.; Anderson, R.A.; Nelson, S.M. Technical and performance characteristics of anti-Müllerian hormone and antral follicle count as biomarkers of ovarian response. Hum. Reprod. Update 2015, 21, 698–710. [Google Scholar] [CrossRef]
- Rustamov, O.; Smith, A.; Roberts, S.A.; Yates, A.P.; Fitzgerald, C.; Krishnan, M.; Nardo, L.G.; Pemberton, P.W. Anti-Mullerian hormone: Poor assay reproducibility in a large cohort of subjects suggests sample instability. Hum. Reprod. 2012, 27, 3085–3091. [Google Scholar] [CrossRef]
- Findlay, J.K.; Hutt, K.J.; Hickey, M.; Anderson, R.A. What is the “ovarian reserve”? Fertil. Steril. 2015, 103, 628–630. [Google Scholar] [CrossRef]
- Kevenaar, M.E.; Meerasahib, M.F.; Kramer, P.; van de Lang-Born, B.M.N.; de Jong, F.H.; Groome, N.P.; Themmen, A.P.N.; Visser, J.A. Serum Anti-Müllerian Hormone Levels Reflect the Size of the Primordial Follicle Pool in Mice. Endocrinology 2006, 147, 3228–3234. [Google Scholar] [CrossRef] [PubMed]
- Myers, M.; Britt, K.L.; Wreford, N.G.; Ebling, F.J.; Kerr, J.B. Methods for quantifying follicular numbers within the mouse ovary. Reproduction 2004, 127, 569–580. [Google Scholar] [CrossRef] [PubMed]
- Sonigo, C.; Jankowski, S.; Yoo, O.; Trassard, O.; Bousquet, N.; Grynberg, M.; Beau, I.; Binart, N. High-throughput ovarian follicle counting by an innovative deep learning approach. Sci. Rep. 2018, 8, 13499. [Google Scholar] [CrossRef]
- Fortune, J.E.; Yang, M.Y.; Allen, J.J.; Herrick, S.L. Triennial Reproduction Symposium: The ovarian follicular reserve in cattle: What regulates its formation and size? J. Anim. Sci. 2013, 91, 3041–3050. [Google Scholar] [CrossRef] [PubMed]
- Nie, X.; Dai, Y.; Zheng, Y.; Bao, D.; Chen, Q.; Yin, Y.; Fu, H.; Hou, D. Establishment of a Mouse Model of Premature Ovarian Failure Using Consecutive Superovulation. Cell Physiol. Biochem. 2018, 51, 2341–2358. [Google Scholar] [CrossRef]
- Tulandi, T.G.R. Preservation of Fertility; Taylor and Francis: London, UK, 2004; p. 3. [Google Scholar]
- Storr, A.; Venetis, C.A.; Cooke, S.; Kilani, S.; Ledger, W. Inter-observer and intra-observer agreement between embryologists during selection of a single Day 5 embryo for transfer: A multicenter study. Hum. Reprod. 2017, 32, 307–314. [Google Scholar] [CrossRef]
- Minasi, M.G.; Colasante, A.; Riccio, T.; Ruberti, A.; Casciani, V.; Scarselli, F.; Spinella, F.; Fiorentino, F.; Varricchio, M.T.; Greco, E. Correlation between aneuploidy, standard morphology evaluation and morphokinetic development in 1730 biopsied blastocysts: A consecutive case series study. Hum. Reprod. 2016, 31, 2245–2254. [Google Scholar] [CrossRef]
- Zhang, Y.; Yan, Z.; Qin, Q.; Nisenblat, V.; Chang, H.M.; Yu, Y.; Wang, T.; Lu, C.; Yang, M.; Yang, S.; et al. Transcriptome Landscape of Human Folliculogenesis Reveals Oocyte and Granulosa Cell Interactions. Mol. Cell 2018, 72, 1021–1034.e4. [Google Scholar] [CrossRef]
- Singh, A.K.; Dutta, M.; Chattopadhyay, R.; Chakravarty, B.; Chaudhury, K. Intrafollicular interleukin-8, interleukin-12, and adrenomedullin are the promising prognostic markers of oocyte and embryo quality in women with endometriosis. J. Assist. Reprod. Genet. 2016, 33, 1363–1372. [Google Scholar] [CrossRef]
- Ilhan, G.; Bacanakgil, B.H.; Vuruskan, A.K.; Eken, M.K.; Karasu, A.F.G.; Bilgic, B.E.; Kucukyurt, A.K. The effect of individual oocyte matched follicular fluid oxidant, antioxidant status, and pro- and anti-inflammatory cytokines on IVF outcomes of patients with diminished ovarian reserve. Medicine 2023, 102, e32757. [Google Scholar] [CrossRef] [PubMed]
- Tian, Z.; Zhang, Y.; Zhang, C.; Wang, Y.; Zhu, H.L. Antral follicle count is reduced in the presence of endometriosis: A systematic review and meta-analysis. Reprod. Biomed. Online 2021, 42, 237–247. [Google Scholar] [CrossRef]
- Muzii, L.; Di Tucci, C.; Di Feliciantonio, M.; Marchetti, C.; Perniola, G.; Panici, P.B. The effect of surgery for endometrioma on ovarian reserve evaluated by antral follicle count: A systematic review and meta-analysis. Hum. Reprod. 2014, 29, 2190–2198. [Google Scholar] [CrossRef]
- Muzii, L.; Di Tucci, C.; Di Feliciantonio, M.; Galati, G.; Di Donato, V.; Musella, A.; Palaia, I.; Panici, P.B. Antimullerian hormone is reduced in the presence of ovarian endometriomas: A systematic review and meta-analysis. Fertil. Steril. 2018, 110, 932–940.e1. [Google Scholar] [CrossRef]
- Horne, A.W.; Missmer, S.A. Pathophysiology, diagnosis, and management of endometriosis. BMJ 2022, 379, e070750. [Google Scholar] [CrossRef] [PubMed]
- Taylor, H.S.; Kotlyar, A.M.; Flores, V.A. Endometriosis is a chronic systemic disease: Clinical challenges and novel innovations. Lancet 2021, 397, 839–852. [Google Scholar] [CrossRef] [PubMed]
- NICE. Endometriosis: Diagnosis and Management; National Institute for Health and Care Excellence: London, UK, 2017. [Google Scholar]
- Becker, C.M.; Bokor, A.; Heikinheimo, O.; Horne, A.; Jansen, F.; Kiesel, L.; King, K.; Kvaskoff, M.; Nap, A.; Petersen, K.; et al. ESHRE guideline: Endometriosis. Hum. Reprod. Open 2022, 2022, hoac009. [Google Scholar] [CrossRef]
- Norman, R.J. Reproductive consequences of COX-2 inhibition. Lancet 2001, 358, 1287–1288. [Google Scholar] [CrossRef]
- Brouwer, J.; Hazes, J.M.; Laven, J.S.; Dolhain, R.J. Fertility in women with rheumatoid arthritis: Influence of disease activity and medication. Ann. Rheum. Dis. 2015, 74, 1836–1841. [Google Scholar] [CrossRef]
- Micu, M.C.; Micu, R.; Ostensen, M. Luteinized unruptured follicle syndrome increased by inactive disease and selective cyclooxygenase 2 inhibitors in women with inflammatory arthropathies. Arthritis Care Res. 2011, 63, 1334–1338. [Google Scholar] [CrossRef] [PubMed]
- Tsafriri, A. Ovulation as a tissue remodelling process. Proteolysis and cumulus expansion. Adv. Exp. Med. Biol. 1995, 377, 121–140. [Google Scholar] [PubMed]
- Smith, G.; Roberts, R.; Hall, C.; Nuki, G. Reversible ovulatory failure associated with the development of luteinized unruptured follicles in women with inflammatory arthritis taking non-steroidal anti-inflammatory drugs. Br. J. Rheumatol. 1996, 35, 458–462. [Google Scholar] [CrossRef]
- Brown, J.; Crawford, T.J.; Datta, S.; Prentice, A. Oral contraceptives for pain associated with endometriosis. Cochrane Database Syst. Rev. 2018, 5, CD001019. [Google Scholar] [CrossRef]
- Brown, J.; Kives, S.; Akhtar, M. Progestagens and anti-progestagens for pain associated with endometriosis. Cochrane Database Syst. Rev. 2012, 2012, CD002122. [Google Scholar] [CrossRef]
- Wan, L.S.; Ganguly, M.; Weiss, G. Pituitary response to LHRH stimulation in women on oral contraceptives: A followup dose response study. Contraception 1981, 24, 229–234. [Google Scholar] [CrossRef] [PubMed]
- Lidegaard, O.; Nielsen, L.H.; Skovlund, C.W.; Skjeldestad, F.E.; Lokkegaard, E. Risk of venous thromboembolism from use of oral contraceptives containing different progestogens and oestrogen doses: Danish cohort study, 2001–2009. BMJ 2011, 343, d6423. [Google Scholar] [CrossRef]
- Ferrero, S.; Barra, F.; Leone Roberti Maggiore, U. Current and Emerging Therapeutics for the Management of Endometriosis. Drugs 2018, 78, 995–1012. [Google Scholar] [CrossRef]
- Deb, S.; Campbell, B.K.; Pincott-Allen, C.; Clewes, J.S.; Cumberpatch, G.; Raine-Fenning, N.J. Quantifying effect of combined oral contraceptive pill on functional ovarian reserve as measured by serum anti-Mullerian hormone and small antral follicle count using three-dimensional ultrasound. Ultrasound Obstet. Gynecol. 2012, 39, 574–580. [Google Scholar] [CrossRef]
- Kallio, S.; Puurunen, J.; Ruokonen, A.; Vaskivuo, T.; Piltonen, T.; Tapanainen, J.S. Antimullerian hormone levels decrease in women using combined contraception independently of administration route. Fertil. Steril. 2013, 99, 1305–1310. [Google Scholar] [CrossRef]
- Landersoe, S.K.; Birch Petersen, K.; Sorensen, A.L.; Larsen, E.C.; Martinussen, T.; Lunding, S.A.; Kroman, M.S.; Nielsen, H.S.; Nyboe Andersen, A. Ovarian reserve markers after discontinuing long-term use of combined oral contraceptives. Reprod. Biomed. Online 2020, 40, 176–186. [Google Scholar] [CrossRef]
- Johnson, L.N.; Sammel, M.D.; Dillon, K.E.; Lechtenberg, L.; Schanne, A.; Gracia, C.R. Antimullerian hormone and antral follicle count are lower in female cancer survivors and healthy women taking hormonal contraception. Fertil. Steril. 2014, 102, 774–781.e3. [Google Scholar] [CrossRef]
- Birch Petersen, K.; Hvidman, H.W.; Forman, J.L.; Pinborg, A.; Larsen, E.C.; Macklon, K.T.; Sylvest, R.; Andersen, A.N. Ovarian reserve assessment in users of oral contraception seeking fertility advice on their reproductive lifespan. Hum. Reprod. 2015, 30, 2364–2375. [Google Scholar] [CrossRef]
- Landersoe, S.K.; Forman, J.L.; Birch Petersen, K.; Larsen, E.C.; Nohr, B.; Hvidman, H.W.; Nielsen, H.S.; Nyboe Andersen, A. Ovarian reserve markers in women using various hormonal contraceptives. Eur. J. Contracept. Reprod. Health Care 2020, 25, 65–71. [Google Scholar] [CrossRef]
- Hariton, E.; Shirazi, T.N.; Douglas, N.C.; Hershlag, A.; Briggs, S.F. Anti-Mullerian hormone levels among contraceptive users: Evidence from a cross-sectional cohort of 27,125 individuals. Am. J. Obstet. Gynecol. 2021, 225, 515.e1–515.e10. [Google Scholar] [CrossRef] [PubMed]
- Amer, S.; James, C.; Al-Hussaini, T.K.; Mohamed, A.A. Assessment of Circulating Anti-Mullerian Hormone in Women Using Hormonal Contraception: A Systematic Review. J. Womens Health 2020, 29, 100–110. [Google Scholar] [CrossRef] [PubMed]
- Vandever, M.A.; Kuehl, T.J.; Sulak, P.J.; Witt, I.; Coffee, A.; Wincek, T.J.; Reape, K.Z. Evaluation of pituitary-ovarian axis suppression with three oral contraceptive regimens. Contraception 2008, 77, 162–170. [Google Scholar] [CrossRef]
- van Heusden, A.M.; Fauser, B.C. Residual ovarian activity during oral steroid contraception. Hum. Reprod. Update 2002, 8, 345–358. [Google Scholar] [CrossRef]
- Landersoe, S.K.; Larsen, E.C.; Forman, J.L.; Birch Petersen, K.; Kroman, M.S.; Frederiksen, H.; Juul, A.; Nohr, B.; Lossl, K.; Nielsen, H.S.; et al. Ovarian reserve markers and endocrine profile during oral contraception: Is there a link between the degree of ovarian suppression and AMH? Gynecol. Endocrinol. 2020, 36, 1090–1095. [Google Scholar] [CrossRef] [PubMed]
- Gougeon, A. Regulation of ovarian follicular development in primates: Facts and hypotheses. Endocr. Rev. 1996, 17, 121–155. [Google Scholar] [CrossRef]
- Barra, F.; Scala, C.; Ferrero, S. Current understanding on pharmacokinetics, clinical efficacy and safety of progestins for treating pain associated to endometriosis. Expert Opin. Drug Metab. Toxicol. 2018, 14, 399–415. [Google Scholar] [CrossRef] [PubMed]
- Bizzarri, N.; Remorgida, V.; Leone Roberti Maggiore, U.; Scala, C.; Tafi, E.; Ghirardi, V.; Salvatore, S.; Candiani, M.; Venturini, P.L.; Ferrero, S. Dienogest in the treatment of endometriosis. Expert Opin. Pharmacother. 2014, 15, 1889–1902. [Google Scholar] [CrossRef]
- Ichioka, M.; Mita, S.; Shimizu, Y.; Imada, K.; Kiyono, T.; Bono, Y.; Kyo, S. Dienogest, a synthetic progestin, down-regulates expression of CYP19A1 and inflammatory and neuroangiogenesis factors through progesterone receptor isoforms A and B in endometriotic cells. J. Steroid Biochem. Mol. Biol. 2015, 147, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Oktay, K.; Newton, H.; Mullan, J.; Gosden, R.G. Development of human primordial follicles to antral stages in SCID/hpg mice stimulated with follicle stimulating hormone. Hum. Reprod. 1998, 13, 1133–1138. [Google Scholar] [CrossRef]
- Hung, S.W.; Zhang, R.; Tan, Z.; Chung, J.P.W.; Zhang, T.; Wang, C.C. Pharmaceuticals targeting signaling pathways of endometriosis as potential new medical treatment: A review. Med. Res. Rev. 2021, 41, 2489–2564. [Google Scholar] [CrossRef]
- Mohamed, K.A.; Davies, W.A.; Lashen, H. Antimullerian hormone and pituitary gland activity after prolonged down-regulation with goserelin acetate. Fertil. Steril. 2006, 86, 1515–1517. [Google Scholar] [CrossRef] [PubMed]
- Marschalek, J.; Ott, J.; Husslein, H.; Kuessel, L.; Elhenicky, M.; Mayerhofer, K.; Franz, M.B. The impact of GnRH agonists in patients with endometriosis on prolactin and sex hormone levels: A pilot study. Eur. J. Obstet. Gynecol. Reprod. Biol. 2015, 195, 156–159. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Tarrega, E.; Monzo, A.M.; Quiroga, R.; Polo-Sanchez, P.; Fernandez-Colom, P.; Monterde-Estrada, M.; Novella-Maestre, E.; Pellicer, A. Effect of GnRH agonist before IVF on outcomes in infertile endometriosis patients: A randomized controlled trial. Reprod. Biomed. Online 2020, 41, 653–662. [Google Scholar] [CrossRef]
- Ma, C.; Qiao, J.; Liu, P.; Chen, G. Ovarian suppression treatment prior to in-vitro fertilization and embryo transfer in Chinese women with stage III or IV endometriosis. Int. J. Gynaecol. Obstet. 2008, 100, 167–170. [Google Scholar] [CrossRef]
- Georgiou, E.X.; Melo, P.; Baker, P.E.; Sallam, H.N.; Arici, A.; Garcia-Velasco, J.A.; Abou-Setta, A.M.; Becker, C.; Granne, I.E. Long-term GnRH agonist therapy before in vitro fertilisation (IVF) for improving fertility outcomes in women with endometriosis. Cochrane Database Syst. Rev. 2019, 2019, CD013240. [Google Scholar] [CrossRef]
- Taylor, H.S.; Giudice, L.C.; Lessey, B.A.; Abrao, M.S.; Kotarski, J.; Archer, D.F.; Diamond, M.P.; Surrey, E.; Johnson, N.P.; Watts, N.B.; et al. Treatment of Endometriosis-Associated Pain with Elagolix, an Oral GnRH Antagonist. N. Engl. J. Med. 2017, 377, 28–40. [Google Scholar] [CrossRef] [PubMed]
- Donnez, J.; Taylor, H.S.; Taylor, R.N.; Akin, M.D.; Tatarchuk, T.F.; Wilk, K.; Gotteland, J.P.; Lecomte, V.; Bestel, E. Treatment of endometriosis-associated pain with linzagolix, an oral gonadotropin-releasing hormone-antagonist: A randomized clinical trial. Fertil. Steril. 2020, 114, 44–55. [Google Scholar] [CrossRef] [PubMed]
- Giudice, L.C.; As-Sanie, S.; Arjona Ferreira, J.C.; Becker, C.M.; Abrao, M.S.; Lessey, B.A.; Brown, E.; Dynowski, K.; Wilk, K.; Li, Y.; et al. Once daily oral relugolix combination therapy versus placebo in patients with endometriosis-associated pain: Two replicate phase 3, randomised, double-blind, studies (SPIRIT 1 and 2). Lancet 2022, 399, 2267–2279. [Google Scholar] [CrossRef]
- Hart, R.J.; Hickey, M.; Maouris, P.; Buckett, W. Excisional surgery versus ablative surgery for ovarian endometriomata. Cochrane Database Syst. Rev. 2008, 16, CD004992. [Google Scholar] [CrossRef]
- Somigliana, E.; Berlanda, N.; Benaglia, L.; Vigano, P.; Vercellini, P.; Fedele, L. Surgical excision of endometriomas and ovarian reserve: A systematic review on serum antimullerian hormone level modifications. Fertil. Steril. 2012, 98, 1531–1538. [Google Scholar] [CrossRef]
- Raffi, F.; Metwally, M.; Amer, S. The impact of excision of ovarian endometrioma on ovarian reserve: A systematic review and meta-analysis. J. Clin. Endocrinol. Metab. 2012, 97, 3146–3154. [Google Scholar] [CrossRef] [PubMed]
- Kuroda, M.; Kuroda, K.; Arakawa, A.; Fukumura, Y.; Kitade, M.; Kikuchi, I.; Kumakiri, J.; Matsuoka, S.; Brosens, I.A.; Brosens, J.J.; et al. Histological assessment of impact of ovarian endometrioma and laparoscopic cystectomy on ovarian reserve. J. Obstet. Gynaecol. Res. 2012, 38, 1187–1193. [Google Scholar] [CrossRef] [PubMed]
- Matsuzaki, S.; Houlle, C.; Darcha, C.; Pouly, J.L.; Mage, G.; Canis, M. Analysis of risk factors for the removal of normal ovarian tissue during laparoscopic cystectomy for ovarian endometriosis. Hum. Reprod. 2009, 24, 1402–1406. [Google Scholar] [CrossRef]
- Muzii, L.; Marana, R.; Angioli, R.; Bianchi, A.; Cucinella, G.; Vignali, M.; Benedetti Panici, P.; Busacca, M. Histologic analysis of specimens from laparoscopic endometrioma excision performed by different surgeons: Does the surgeon matter? Fertil. Steril. 2011, 95, 2116–2119. [Google Scholar] [CrossRef] [PubMed]
- Carmona, F.; Martinez-Zamora, M.A.; Rabanal, A.; Martinez-Roman, S.; Balasch, J. Ovarian cystectomy versus laser vaporization in the treatment of ovarian endometriomas: A randomized clinical trial with a five-year follow-up. Fertil. Steril. 2011, 96, 251–254. [Google Scholar] [CrossRef]
- Ding, W.; Li, M.; Teng, Y. The impact on ovarian reserve of haemostasis by bipolar coagulation versus suture following surgical stripping of ovarian endometrioma: A meta-analysis. Reprod. Biomed. Online 2015, 30, 635–642. [Google Scholar] [CrossRef]
- Ata, B.; Turkgeldi, E.; Seyhan, A.; Urman, B. Effect of hemostatic method on ovarian reserve following laparoscopic endometrioma excision; comparison of suture, hemostatic sealant, and bipolar dessication. A systematic review and meta-analysis. J. Minim. Invasive Gynecol. 2015, 22, 363–372. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; An, D.; Ye, J.; Fu, R.; Zhao, A. Effect of early inflammatory reaction on ovarian reserve after laparoscopic cystectomy for ovarian endometriomas. J. Obstet. Gynaecol. 2022, 42, 3124–3128. [Google Scholar] [CrossRef]
- Burns, K.A.; Pearson, A.M.; Slack, J.L.; Por, E.D.; Scribner, A.N.; Eti, N.A.; Burney, R.O. Endometriosis in the Mouse: Challenges and Progress Toward a ‘Best Fit’ Murine Model. Front. Physiol. 2021, 12, 806574. [Google Scholar] [CrossRef] [PubMed]
- Yotova, I.; Hudson, Q.J.; Pauler, F.M.; Proestling, K.; Haslinger, I.; Kuessel, L.; Perricos, A.; Husslein, H.; Wenzl, R. LINC01133 Inhibits Invasion and Promotes Proliferation in an Endometriosis Epithelial Cell Line. Int. J. Mol. Sci. 2021, 22, 8385. [Google Scholar] [CrossRef] [PubMed]
- Pastor, D.M.; Poritz, L.S.; Olson, T.L.; Kline, C.L.; Harris, L.R.; Koltun, W.A.; Chinchilli, V.M.; Irby, R.B. Primary cell lines: False representation or model system? a comparison of four human colorectal tumors and their coordinately established cell lines. Int. J. Clin. Exp. Med. 2010, 3, 69–83. [Google Scholar]
- Gu, Z.Y.; Jia, S.Z.; Leng, J.H. Establishment of endometriotic models: The past and future. Chin. Med. J. 2020, 133, 1703–1710. [Google Scholar] [CrossRef]
- Lin, X.; Dai, Y.; Tong, X.; Xu, W.; Huang, Q.; Jin, X.; Li, C.; Zhou, F.; Zhou, H.; Lin, X.; et al. Excessive oxidative stress in cumulus granulosa cells induced cell senescence contributes to endometriosis-associated infertility. Redox Biol. 2020, 30, 101431. [Google Scholar] [CrossRef]
- Dumesic, D.A.; Meldrum, D.R.; Katz-Jaffe, M.G.; Krisher, R.L.; Schoolcraft, W.B. Oocyte environment: Follicular fluid and cumulus cells are critical for oocyte health. Fertil. Steril. 2015, 103, 303–316. [Google Scholar] [CrossRef]
- Li, Y.; Li, R.; Ouyang, N.; Dai, K.; Yuan, P.; Zheng, L.; Wang, W. Investigating the impact of local inflammation on granulosa cells and follicular development in women with ovarian endometriosis. Fertil. Steril. 2019, 112, 882–891.e1. [Google Scholar] [CrossRef]
- Chen, Y.J.; Wu, H.H.; Liau, W.T.; Tsai, C.Y.; Tsai, H.W.; Chao, K.C.; Sung, Y.J.; Li, H.Y. A tumor necrosis factor-α inhibitor reduces the embryotoxic effects of endometriotic peritoneal fluid. Fertil. Steril. 2013, 100, 1476–1485. [Google Scholar] [CrossRef] [PubMed]
- Piromlertamorn, W.; Saeng-anan, U.; Vutyavanich, T. Effects of ovarian endometriotic fluid exposure on fertilization rate of mouse oocytes and subsequent embryo development. Reprod. Biol. Endocrinol. 2013, 11, 4. [Google Scholar] [CrossRef]
- Giorgi, V.S.I.; Ferriani, R.A.; Navarro, P.A. Follicular Fluid from Infertile Women with Mild Endometriosis Impairs In Vitro Bovine Embryo Development: Potential Role of Oxidative Stress. Rev. Bras. De Ginecol. E Obs. 2021, 43, 119–125. [Google Scholar] [CrossRef]
- D’Hooghe, T.M.; Kyama, C.M.; Chai, D.; Fassbender, A.; Vodolazkaia, A.; Bokor, A.; Mwenda, J.M. Nonhuman primate models for translational research in endometriosis. Reprod. Sci. 2009, 16, 152–161. [Google Scholar] [CrossRef]
- Ami, Y.; Suzaki, Y.; Goto, N. Endometriosis in cynomolgus monkeys retired from breeding. J. Vet. Med. Sci. 1993, 55, 7–11. [Google Scholar] [CrossRef]
- Nishimoto-Kakiuchi, A.; Netsu, S.; Okabayashi, S.; Taniguchi, K.; Tanimura, H.; Kato, A.; Suzuki, M.; Sankai, T.; Konno, R. Spontaneous endometriosis in cynomolgus monkeys as a clinically relevant experimental model. Hum. Reprod. 2018, 33, 1228–1236. [Google Scholar] [CrossRef]
- Simitsidellis, I.; Gibson, D.A.; Saunders, P.T.K. Animal models of endometriosis: Replicating the aetiology and symptoms of the human disorder. Best. Pr. Res. Clin. Endocrinol. Metab. 2018, 32, 257–269. [Google Scholar] [CrossRef] [PubMed]
- Yamanaka, A.; Kimura, F.; Takebayashi, A.; Kita, N.; Takahashi, K.; Murakami, T. Primate model research for endometriosis. Tohoku J. Exp. Med. 2012, 226, 95–99. [Google Scholar] [CrossRef] [PubMed]
- D’Hooghe, T.M.; Bambra, C.S.; Raeymaekers, B.M.; De Jonge, I.; Lauweryns, J.M.; Koninckx, P.R. Intrapelvic injection of menstrual endometrium causes endometriosis in baboons (Papio cynocephalus and Papio anubis). Am. J. Obstet. Gynecol. 1995, 173, 125–134. [Google Scholar] [CrossRef]
- Fazleabas, A.T.; Brudney, A.; Gurates, B.; Chai, D.; Bulun, S. A modified baboon model for endometriosis. Ann. N. Y. Acad. Sci. 2002, 955, 308–317; Discussion 340–342, 396–406. [Google Scholar] [CrossRef]
- Grummer, R. Animal models in endometriosis research. Hum. Reprod. Update 2006, 12, 641–649. [Google Scholar] [CrossRef]
- Sharpe-Timms, K.L. Using rats as a research model for the study of endometriosis. Ann. N. Y. Acad. Sci. 2002, 955, 318–327; Discussion 340–342, 396–406. [Google Scholar] [CrossRef] [PubMed]
- Tirado-Gonzalez, I.; Barrientos, G.; Tariverdian, N.; Arck, P.C.; Garcia, M.G.; Klapp, B.F.; Blois, S.M. Endometriosis research: Animal models for the study of a complex disease. J. Reprod. Immunol. 2010, 86, 141–147. [Google Scholar] [CrossRef] [PubMed]
- Dinulescu, D.M.; Ince, T.A.; Quade, B.J.; Shafer, S.A.; Crowley, D.; Jacks, T. Role of K-ras and Pten in the development of mouse models of endometriosis and endometrioid ovarian cancer. Nat. Med. 2005, 11, 63–70. [Google Scholar] [CrossRef]
- Schenken, R.S.; Asch, R.H. Surgical induction of endometriosis in the rabbit: Effects on fertility and concentrations of peritoneal fluid prostaglandins. Fertil. Steril. 1980, 34, 581–587. [Google Scholar] [CrossRef]
- Moon, C.E.; Bertero, M.C.; Curry, T.E.; London, S.N.; Muse, K.N.; Sharpe, K.L.; Vernon, M.W. The presence of luteinized unruptured follicle syndrome and altered folliculogenesis in rats with surgically induced endometriosis. Am. J. Obstet. Gynecol. 1993, 169, 676–682. [Google Scholar] [CrossRef]
- Pal, A.K.; Biswas, S.; Goswami, S.K.; Kabir, S.N. Effect of pelvic endometrial implants on overall reproductive functions of female rats. Biol. Reprod. 1999, 60, 954–958. [Google Scholar] [CrossRef] [PubMed]
- Elsherbini, M.; Koga, K.; Maki, E.; Kumasawa, K.; Satake, E.; Taguchi, A.; Makabe, T.; Takeuchi, A.; Izumi, G.; Takamura, M.; et al. Impact of Chronic Exposure to Endometriosis on Perinatal Outcomes: Establishment of a Mouse Model. Biomedicines 2022, 10, 2627. [Google Scholar] [CrossRef] [PubMed]
- Bengochea, T.S.O.C.; Ricci, A.G.; Abramovich, D.; Parborell, F.; Meresman, G.F.; Barañao, R.I.; Bilotas, M.A. Endometriosis Impairs Ovarian Reserve and Ovulation in a Rat Model. In XVIII Jornadas Anuales Multidisciplinarias; De la Sociedad Argentina de Biología: Buenos Aires, Argentina, 2016; p. A43. [Google Scholar]
- Syrop, C.H.; Halme, J. Cyclic changes of peritoneal fluid parameters in normal and infertile patients. Obstet. Gynecol. 1987, 69, 416–418. [Google Scholar] [PubMed]
- Pellicer, A.; Albert, C.; Mercader, A.; Bonilla-Musoles, F.; Remohi, J.; Simon, C. The follicular and endocrine environment in women with endometriosis: Local and systemic cytokine production. Fertil. Steril. 1998, 70, 425–431. [Google Scholar] [CrossRef]
- Carlberg, M.; Nejaty, J.; Froysa, B.; Guan, Y.; Soder, O.; Bergqvist, A. Elevated expression of tumour necrosis factor alpha in cultured granulosa cells from women with endometriosis. Hum. Reprod. 2000, 15, 1250–1255. [Google Scholar] [CrossRef]
- Harada, T.; Iwabe, T.; Terakawa, N. Role of cytokines in endometriosis. Fertil. Steril. 2001, 76, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.S.; Joo, B.S.; Na, Y.J.; Yoon, M.S.; Choi, O.H.; Kim, W.W. Relationships between concentrations of tumor necrosis factor-alpha and nitric oxide in follicular fluid and oocyte quality. J. Assist. Reprod. Genet. 2000, 17, 222–228. [Google Scholar] [CrossRef] [PubMed]
- Cacciottola, L.; Donnez, J.; Dolmans, M.-M. Can Endometriosis-Related Oxidative Stress Pave the Way for New Treatment Targets? Int. J. Mol. Sci. 2021, 22, 7138. [Google Scholar] [CrossRef] [PubMed]
- Qiao, J.; Wang, Z.B.; Feng, H.L.; Miao, Y.L.; Wang, Q.; Yu, Y.; Wei, Y.C.; Yan, J.; Wang, W.H.; Shen, W.; et al. The root of reduced fertility in aged women and possible therapentic options: Current status and future perspects. Mol. Asp. Med. 2014, 38, 54–85. [Google Scholar] [CrossRef]
- Komatsu, K.; Masubuchi, S. Increased supply from blood vessels promotes the activation of dormant primordial follicles in mouse ovaries. J. Reprod. Dev. 2020, 66, 105–113. [Google Scholar] [CrossRef]
- Saito, A.; Iwase, A.; Nakamura, T.; Osuka, S.; Bayasula; Murase, T.; Kato, N.; Ishida, C.; Takikawa, S.; Goto, M.; et al. Involvement of mesosalpinx in endometrioma is a possible risk factor for decrease of ovarian reserve after cystectomy: A retrospective cohort study. Reprod. Biol. Endocrinol. 2016, 14, 72. [Google Scholar] [CrossRef] [PubMed]
- Jiang, D.; Nie, X. Effect of endometrioma and its surgical excision on fertility (Review). Exp. Ther. Med. 2020, 20, 114. [Google Scholar] [CrossRef]
- Fauser, B.C.; Van Heusden, A.M. Manipulation of human ovarian function: Physiological concepts and clinical consequences. Endocr. Rev. 1997, 18, 71–106. [Google Scholar]
- Drummond, A.E. The role of steroids in follicular growth. Reprod. Biol. Endocrinol. 2006, 4, 16. [Google Scholar] [CrossRef]
- Chang, E.M.; Kim, Y.S.; Won, H.J.; Yoon, T.K.; Lee, W.S. Association Between Sex Steroids, Ovarian Reserve, and Vitamin D Levels in Healthy Nonobese Women. J. Clin. Endocrinol. Metab. 2014, 99, 2526–2532. [Google Scholar] [CrossRef]
- Marquardt, R.M.; Kim, T.H.; Shin, J.H.; Jeong, J.W. Progesterone and Estrogen Signaling in the Endometrium: What Goes Wrong in Endometriosis? Int. J. Mol. Sci. 2019, 20, 3822. [Google Scholar] [CrossRef] [PubMed]
- Chantalat, E.; Valera, M.C.; Vaysse, C.; Noirrit, E.; Rusidze, M.; Weyl, A.; Vergriete, K.; Buscail, E.; Lluel, P.; Fontaine, C.; et al. Estrogen Receptors and Endometriosis. Int. J. Mol. Sci. 2020, 21, 2815. [Google Scholar] [CrossRef] [PubMed]
- Kitajima, M.; Matsumoto, K.; Kajimura, I.; Harada, A.; Miyashita, N.; Matsumura, A.; Kitajima, Y.; Miura, K. The Effects of Endometriosis on Ovarian Functions. Endocrines 2021, 2, 142–149. [Google Scholar] [CrossRef]
- Stilley, J.A.W.; Birt, J.A.; Sharpe-Timms, K.L. Cellular and molecular basis for endometriosis-associated infertility. Cell Tissue Res. 2012, 349, 849–862. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, C.R.; Eddy, C.A.; Olive, D.L.; Schenken, R.S. Effect of ovarian endometriosis on ovulation in rabbits. Am. J. Obstet. Gynecol. 1989, 160, 40–44. [Google Scholar] [CrossRef]
- King, C.M.; Barbara, C.; Prentice, A.; Brenton, J.D.; Charnock-Jones, D.S. Models of endometriosis and their utility in studying progression to ovarian clear cell carcinoma. J. Pathol. 2016, 238, 185–196. [Google Scholar] [CrossRef]
- Chou, C.H.; Chen, M.J. The Effect of Steroid Hormones on Ovarian Follicle Development. Vitam. Horm. 2018, 107, 155–175. [Google Scholar]
- Chauvin, S.; Cohen-Tannoudji, J.; Guigon, C.J. Estradiol Signaling at the Heart of Folliculogenesis: Its Potential Deregulation in Human Ovarian Pathologies. Int. J. Mol. Sci. 2022, 23, 512. [Google Scholar] [CrossRef]
- Burns, K.A.; Rodriguez, K.F.; Hewitt, S.C.; Janardhan, K.S.; Young, S.L.; Korach, K.S. Role of Estrogen Receptor Signaling Required for Endometriosis-Like Lesion Establishment in a Mouse Model. Endocrinology 2012, 153, 3960–3971. [Google Scholar] [CrossRef]
- Patiño-García, D.; Palomino, J.; Pomés, C.; Celle, C.; Torres-Estay, V.; Orellana, R. Estetrol Increases Progesterone Genetic Response without Triggering Common Estrogenic Effects in Endometriotic Cell Lines and Primary Cultures. Biomedicines 2023, 11, 1169. [Google Scholar] [CrossRef]
- Cahill, D.J.; Hull, M.G. Pituitary-ovarian dysfunction and endometriosis. Hum. Reprod. Update 2000, 6, 56–66. [Google Scholar] [CrossRef]
- Kitajima, M.; Dolmans, M.M.; Donnez, O.; Masuzaki, H.; Soares, M.; Donnez, J. Enhanced follicular recruitment and atresia in cortex derived from ovaries with endometriomas. Fertil. Steril. 2014, 101, 1031–1037. [Google Scholar] [CrossRef]
- Maidarti, M.; Anderson, R.A.; Telfer, E.E. Crosstalk between PTEN/PI3K/Akt Signalling and DNA Damage in the Oocyte: Implications for Primordial Follicle Activation, Oocyte Quality and Ageing. Cells 2020, 9, 200. [Google Scholar] [CrossRef] [PubMed]
- Jagarlamudi, K.; Liu, L.; Adhikari, D.; Reddy, P.; Idahl, A.; Ottander, U.; Lundin, E.; Liu, K. Oocyte-specific deletion of Pten in mice reveals a stage-specific function of PTEN/PI3K signaling in oocytes in controlling follicular activation. PLoS ONE 2009, 4, e6186. [Google Scholar] [CrossRef] [PubMed]
- Kawamura, K.; Kawamura, N.; Hsueh, A.J. Activation of dormant follicles: A new treatment for premature ovarian failure? Curr. Opin. Obstet. Gynecol. 2016, 28, 217–222. [Google Scholar] [CrossRef] [PubMed]
- De Felici, M.; Klinger, F.G. PI3K/PTEN/AKT Signaling Pathways in Germ Cell Development and Their Involvement in Germ Cell Tumors and Ovarian Dysfunctions. Int. J. Mol. Sci. 2021, 22, 9838. [Google Scholar] [CrossRef]
- Zhang, M.; Su, Y.Q.; Sugiura, K.; Xia, G.; Eppig, J.J. Granulosa cell ligand NPPC and its receptor NPR2 maintain meiotic arrest in mouse oocytes. Science 2010, 330, 366–369. [Google Scholar] [CrossRef]
- Pelosi, E.; Omari, S.; Michel, M.; Ding, J.; Amano, T.; Forabosco, A.; Schlessinger, D.; Ottolenghi, C. Constitutively active Foxo3 in oocytes preserves ovarian reserve in mice. Nat. Commun. 2013, 4, 1843. [Google Scholar] [CrossRef]
- Andrade, G.M.; da Silveira, J.C.; Perrini, C.; Del Collado, M.; Gebremedhn, S.; Tesfaye, D.; Meirelles, F.V.; Perecin, F. The role of the PI3K-Akt signaling pathway in the developmental competence of bovine oocytes. PLoS ONE 2017, 12, e0185045. [Google Scholar] [CrossRef] [PubMed]
- Al-Jabri, S.; Tulandi, T. Management and prevention of pelvic adhesions. Semin. Reprod. Med. 2011, 29, 130–137. [Google Scholar] [CrossRef]
- Halwagy, A.S.E.; Gergawy, A.A.A.; Dawood, A.S.; Shehata, A. Reduction of Postoperative Adhesions after Laparoscopic Surgery for Endometriosis by Using a Novel Anti-Fibrotic Drug Pirfenidone: A Randomized Double Blind Study. Gynecol. Obstet. 2017, 7, 1000422. [Google Scholar] [CrossRef]
- Kramer, B.; Andress, J.; Neis, F.; Hoffmann, S.; Brucker, S.; Kommoss, S.; Holler, A. Adhesion prevention after endometriosis surgery—Results of a randomized, controlled clinical trial with second-look laparoscopy. Langenbecks Arch. Surg. 2021, 406, 2133–2143. [Google Scholar] [CrossRef] [PubMed]
- Kramer, B.; Andress, J.; Neis, F.; Hoffmann, S.; Brucker, S.; Kommoss, S.; Holler, A. Improvement in Fertility and Pain after Endometriosis Resection and Adhesion Prevention with 4DryField® PH: Follow-up of a Randomized Controlled Clinical Trial. J. Clin. Med. 2023, 12, 3597. [Google Scholar] [CrossRef]
- Keshavarzi, S.; MacDougall, M.; Lulic, D.; Kasasbeh, A.; Levy, M. Clinical experience with the surgicel family of absorbable hemostats (oxidized regenerated cellulose) in neurosurgical applications: A review. Wounds 2013, 25, 160–167. [Google Scholar] [PubMed]
- Wu, Q.; Yang, Q.; Lin, Y.; Wu, L.; Lin, T. The optimal time for laparoscopic excision of ovarian endometrioma: A prospective randomized controlled trial. Reprod. Biol. Endocrinol. 2023, 21, 59. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Zhang, Y.; Lian, X.; Li, F.; Wang, C.; Zhu, F.; Qiu, Y.; Chen, Y. Therapeutic target database update 2022: Facilitating drug discovery with enriched comparative data of targeted agents. Nucleic Acids Res. 2022, 50, D1398–D1407. [Google Scholar] [CrossRef] [PubMed]
- Bilusic, M.; Heery, C.R.; Collins, J.M.; Donahue, R.N.; Palena, C.; Madan, R.A.; Karzai, F.; Marté, J.L.; Strauss, J.; Gatti-Mays, M.E.; et al. Phase I trial of HuMax-IL8 (BMS-986253), an anti-IL-8 monoclonal antibody, in patients with metastatic or unresectable solid tumors. J. Immunother. Cancer 2019, 7, 240. [Google Scholar] [CrossRef] [PubMed]
- Thaker, P.H.; Brady, W.E.; Lankes, H.A.; Odunsi, K.; Bradley, W.H.; Moore, K.N.; Muller, C.Y.; Anwer, K.; Schilder, R.J.; Alvarez, R.D.; et al. A phase I trial of intraperitoneal GEN-1, an IL-12 plasmid formulated with PEG-PEI-cholesterol lipopolymer, administered with pegylated liposomal doxorubicin in patients with recurrent or persistent epithelial ovarian, fallopian tube or primary peritoneal cancers: An NRG Oncology/Gynecologic Oncology Group study. Gynecol. Oncol. 2017, 147, 283–290. [Google Scholar] [PubMed]
- Petroski, N. Advax Adjuvant: A Potent and Safe Immunopotentiator Composed of Delta Inulin. In Immunopotentiators in Modern Vaccines; National Library of Medicine: Bethesda, MD, USA, 2017; pp. 199–210. [Google Scholar] [CrossRef]
- Mayoclinic Tafenoquine (Oral Route). Available online: https://www.mayoclinic.org/drugs-supplements/tafenoquine-oral-route/side-effects/drg-20443911 (accessed on 1 February 2023).
- Mayoclinic Drugs and Supplements, Sirolimus (Oral Route). Available online: https://www.mayoclinic.org/drugs-supplements/sirolimus-oral-route/side-effects/drg-20068199 (accessed on 30 July 2023).
- Le Tutour, B.; Guedon, D. Antioxidative activities of Olea europaea leaves and related phenolic compounds. Phytochemistry 1992, 31, 1173–1178. [Google Scholar] [CrossRef]
- Schwertner, A.; Conceicao Dos Santos, C.C.; Costa, G.D.; Deitos, A.; de Souza, A.; de Souza, I.C.; Torres, I.L.; da Cunha Filho, J.S.; Caumo, W. Efficacy of melatonin in the treatment of endometriosis: A phase II, randomized, double-blind, placebo-controlled trial. Pain 2013, 154, 874–881. [Google Scholar] [CrossRef]
- Cetinkaya, N.; Attar, R.; Yildirim, G.; Ficicioglu, C.; Ozkan, F.; Yilmaz, B.; Yesildaglar, N. The effects of different doses of melatonin treatment on endometrial implants in an oophorectomized rat endometriosis model. Arch. Gynecol. Obstet. 2015, 291, 591–598. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Hung, S.W.; Zhang, R.; Man, G.C.; Zhang, T.; Chung, J.P.; Fang, L.; Wang, C.C. Melatonin in Endometriosis: Mechanistic Understanding and Clinical Insight. Nutrients 2022, 14, 4087. [Google Scholar] [CrossRef] [PubMed]
- Barchas, J.; DaCosta, F.; Spector, S. Acute pharmacology of melatonin. Nature 1967, 214, 919–920. [Google Scholar] [CrossRef]
- Zhao, X.; Wang, D.; Wu, Z.; Pan, B.; Yang, H.; Zeng, C.; Zhang, M.; Liu, G.; Han, H.; Zhou, G. Female Reproductive Performance in the Mouse: Effect of Oral Melatonin. Molecules 2018, 23, 1845. [Google Scholar] [CrossRef]
- Eryilmaz, O.G.; Devran, A.; Sarikaya, E.; Aksakal, F.N.; Mollamahmutoglu, L.; Cicek, N. Melatonin improves the oocyte and the embryo in IVF patients with sleep disturbances, but does not improve the sleeping problems. J. Assist. Reprod. Genet. 2011, 28, 815–820. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Author, Year | Study Type | Disease Subtype | Interventions | OR Maker |
|---|---|---|---|---|
| Kanellopoulos D et al., 2022 [13] | In vivo/rat | OMA/diffuse intraperitoneal endometriosis/subcutaneous endometriosis | - | The number of luteinized unruptured follicles ↑ |
| Park Y et al., 2020 [14] | In vivo/mouse | Diffuse intraperitoneal endometriosis | Oleuropein (25 mg/kg/day, p.o.) for 21 days | Pregnancy rate (vehicle vs. oleuropein vs. no endometriosis = 70%:100%:100%) |
| Hayashi S et al., 2020 [15] | In vivo/mouse | OMA | - | The number of pups ↓ |
| Takeuchi A et al., 2019 [16] | In vivo/mouse | Diffuse intraperitoneal endometriosis | - | Primordial follicle count ↓ |
| Kacan T et al., 2017 [17] | In vivo/rat | Peritoneal endometriosis | Everolimus (1.5 mg/kg/day, p.o.) for 14 days | Follicle number ↑ |
| Ozer H et al., 2013 [18] | In vivo/rat | Peritoneal endometriosis | Retinoic acid (10 mg/kg/day, p.o.) for 10 days | Primordial follicle count ↑ |
| Stilley JA et al., 2010 [19] | In vivo/rat, in vitro | DIE | TIMP1-blocking antibody (32μg/kg, i.p., every third day for a total of three injections) | Follicle number ↑ Embryo quality ↑ |
| Stilley JA et al., 2009 [20] | In vivo/rat, in vitro | DIE | - | Antral follicle number ↓ Oocyte number ↓ Oocyte quality ↓ Zygotes number ↓ Embryo quality ↓ Fecundity ↓ |
| D’Hooghe TM et al., 1996 [21] | In vivo/baboon | Diffuse intraperitoneal endometriosis | - | Cycle pregnancy rate (stage II and stage III-IV) ↓ |
| Barragán JC et al., 1992 [22] | In vivo/rat | Peritoneal endometriosis/DIE | - | Pregnancy rate ↓ |
| Schenken RS et al., 1991 [23] | In vivo/monkey | Peritoneal endometriosis | - | Pregnancy rate (moderate or severe disease) ↓ |
| Muraoka A et al., 2021 [24] | Human study | OMA | Laparoscopic ovarian cystectomy with dienogest (2 mg/day) or buserelin acetate (1.8 mg/month) for 2 months preoperatively and 2 months postoperatively | AMH after 1-year follow-up (dienogest group ↑ vs. buserelin group ↓) |
| Sireesha MU et al., 2021 [25] | Human study | OMA | Laparoscopic ovarian cystectomy | AMH (1 week and 3 months postoperatively) ↓ |
| Barra F et al., 2020 [26] | Human study, in vitro | OMA | Dienogest (2 mg/day, p.o.) for 3 months | Implantation rate ↑ Pregnancy rate ↑ Live birth rate ↑ The number of oocytes retrieved, two-pronuclear embryos and blastocysts ↑ |
| Karadağ C et al., 2020 [27] | Human study | OMA | - | AMH ↓ AFC ↓ |
| Candiani M et al., 2018 [28] | Human study | OMA | CO2 laser vaporization or laparoscopic ovarian cystectomy | AFC (laser vaporization group ↑ vs. cystectomy group ↓) and AMH (laser vaporization group ↓ vs. cystectomy group↑) at 3-month follow-up |
| Kasapoglu I et al., 2018 [29] | Human study | OMA | - | AMH ↓ |
| Muller V et al., 2017 [30] | Human study, in vitro | OMA | Laparoscopic ovarian cystectomy followed by dienogest (2 mg/day, p.o.) treatment for 6 months | Pregnancy rate ↑ Live birth rate ↑ The number of oocytes retrieved and A-class embryos ↑ |
| Taniguchi F et al., 2015 [31] | Human study | OMA | OCP (DRSP/EE 3 mg/20μg, 24/4 regimen, 6 cycles) | AMH (after 6 cycles) ↓ |
| Lima ML et al., 2015 [32] | Human study | OMA | - | AFC ↓ The number of oocytes retrieved (-) |
| Li CZ et al., 2009 [33] | Human study | OMA | Electrocoagulation or suture after excision of OMA | AFC and ovarian diameter at 3-, 6-, 12-month follow-up (electrocoagulation ↓ vs. suture ↑) |
| Measurement | Side of Ovary | Follicle Stage | Menstrual Cycle-Sensitive | Expense |
|---|---|---|---|---|
| AFC | One-by-one | Antral follicle | No | ** |
| FSH | Both | Developing follicles | Yes | * |
| AMH | Both | Preantral and antral | No | * |
| Oocyte retrieval after superovulation | Both/Unilateral | / | / | *** |
| Signaling Pathway/Molecules | Drug 1 | Drug Type | Indication | Clinical Phase 2 | Pregnancy Category | Potential Long-Term Implications 3 |
|---|---|---|---|---|---|---|
| IL-8 | BMS-986253 | Monoclonal antibody | Coronavirus Disease 2019 | Phase 2 | Unknown | Grade 1/2 [183] |
| IL-12 | GEN-1 | Gene therapy | Ovarian cancer | Phase 2 | Unknown | Grade 1, 2, 3 [184] |
| TNF-α | ADVAX | Highly potent adjuvants | Human immunodeficiency virus infection | Phase 1 | B | Grade 1 [185] |
| ROS | Tafenoquine | Small molecular drug | Malaria; Plasmodium vivax malaria | Approved | C | Grade 1, 2 [186] |
| PI3K/AKT/mTOR pathway (PAm pathway) | Sirolimus | Small molecular drug | Multiple myeloma; Organ transplant rejection | Approved | C | Grade 1, 2, 3 [187] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tan, Z.; Gong, X.; Wang, C.C.; Zhang, T.; Huang, J. Diminished Ovarian Reserve in Endometriosis: Insights from In Vitro, In Vivo, and Human Studies—A Systematic Review. Int. J. Mol. Sci. 2023, 24, 15967. https://doi.org/10.3390/ijms242115967
Tan Z, Gong X, Wang CC, Zhang T, Huang J. Diminished Ovarian Reserve in Endometriosis: Insights from In Vitro, In Vivo, and Human Studies—A Systematic Review. International Journal of Molecular Sciences. 2023; 24(21):15967. https://doi.org/10.3390/ijms242115967
Chicago/Turabian StyleTan, Zhouyurong, Xue Gong, Chi Chiu Wang, Tao Zhang, and Jin Huang. 2023. "Diminished Ovarian Reserve in Endometriosis: Insights from In Vitro, In Vivo, and Human Studies—A Systematic Review" International Journal of Molecular Sciences 24, no. 21: 15967. https://doi.org/10.3390/ijms242115967