
Identification of HuSWEET Family in Pitaya (Hylocereus undatus) and Key Roles of HuSWEET12a and HuSWEET13d in Sugar Accumulation
 and
and
Abstract
:1. Introduction
2. Results
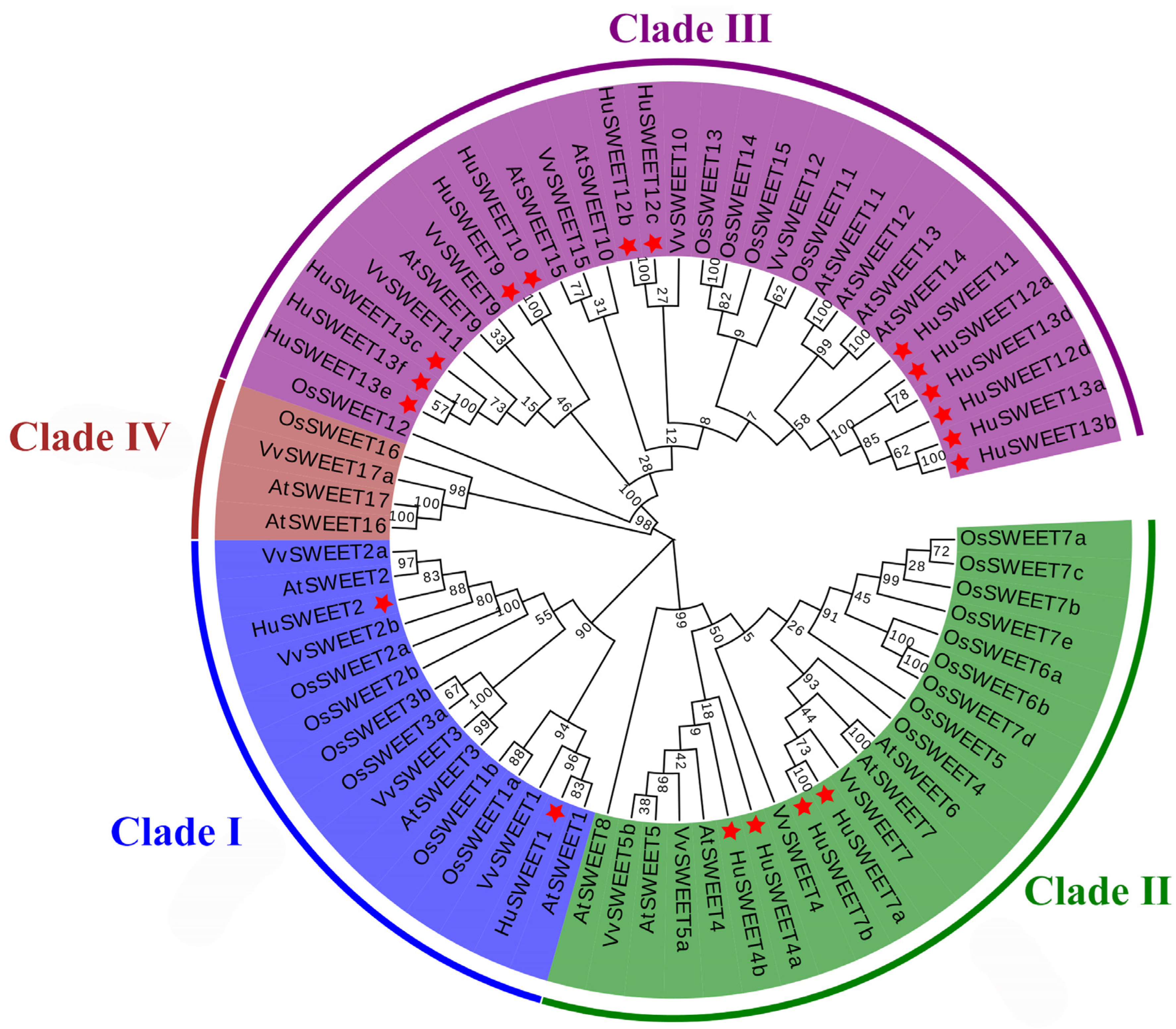
2.1. Identification and Phylogenetic Analyses of the HuSWEET Family
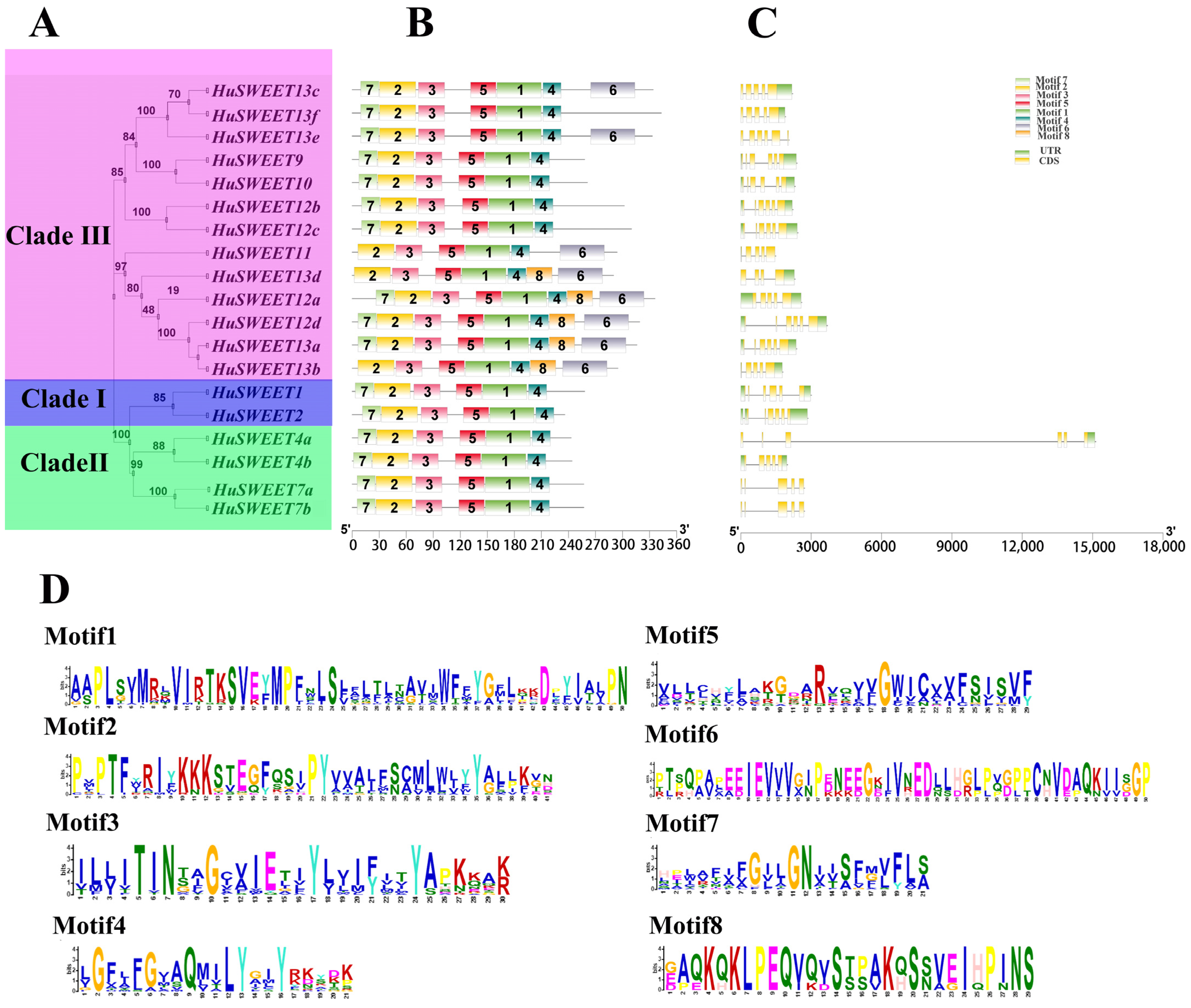
2.2. Conserved Motif and Gene Structure Analyses of HuSWEETs
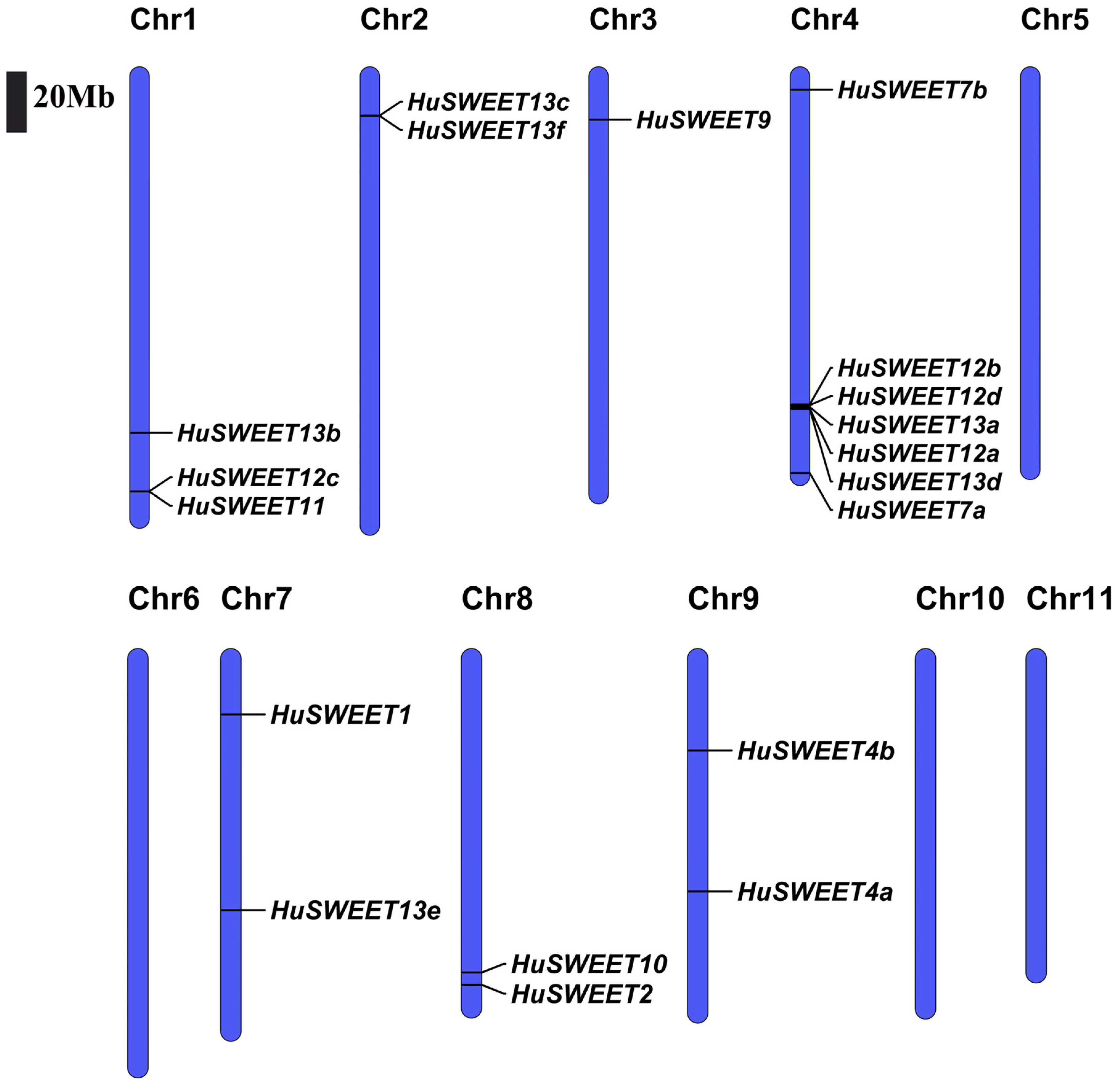
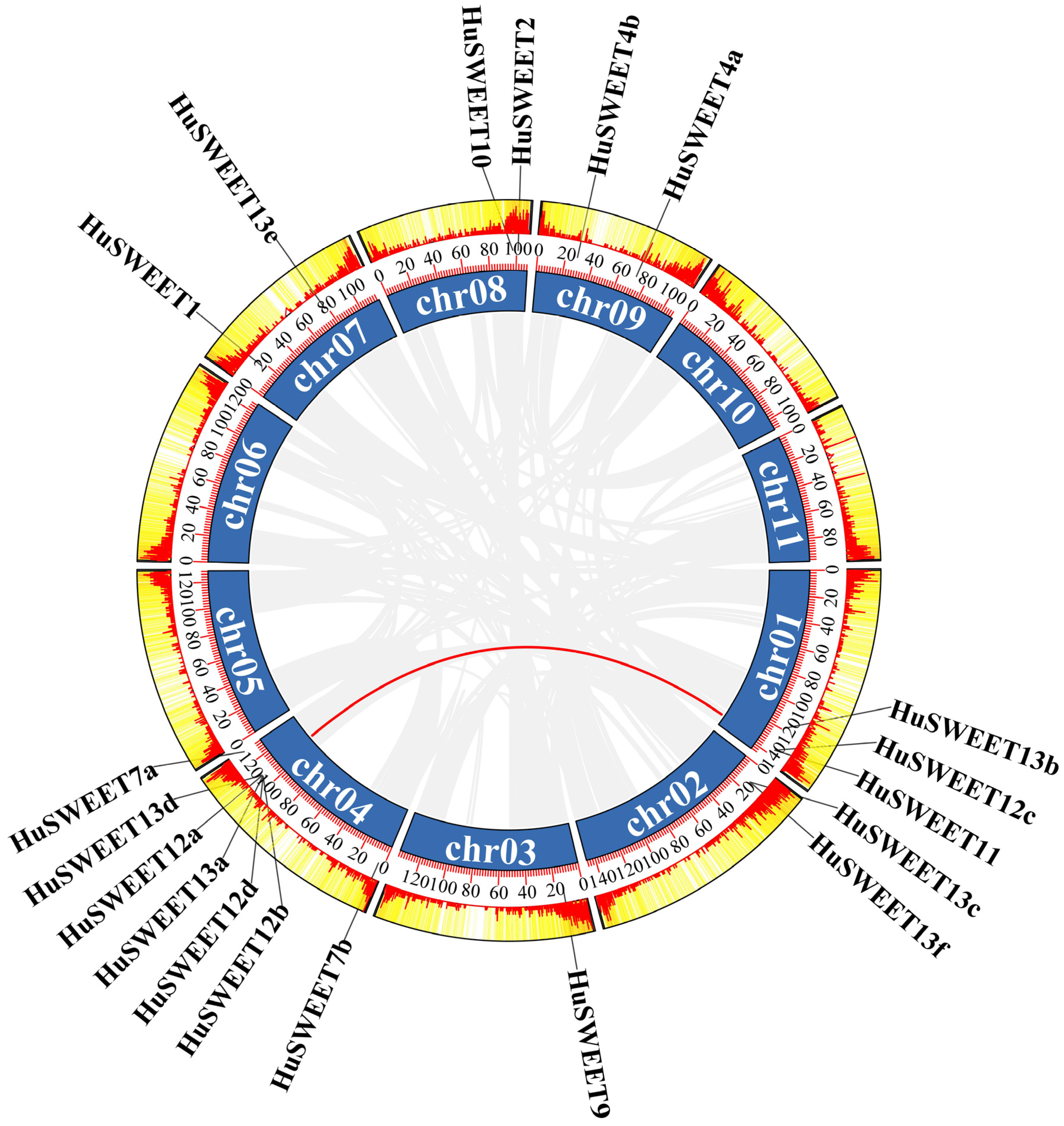
2.3. Chromosomal Localizations and Synteny Analyses of HuSWEETs
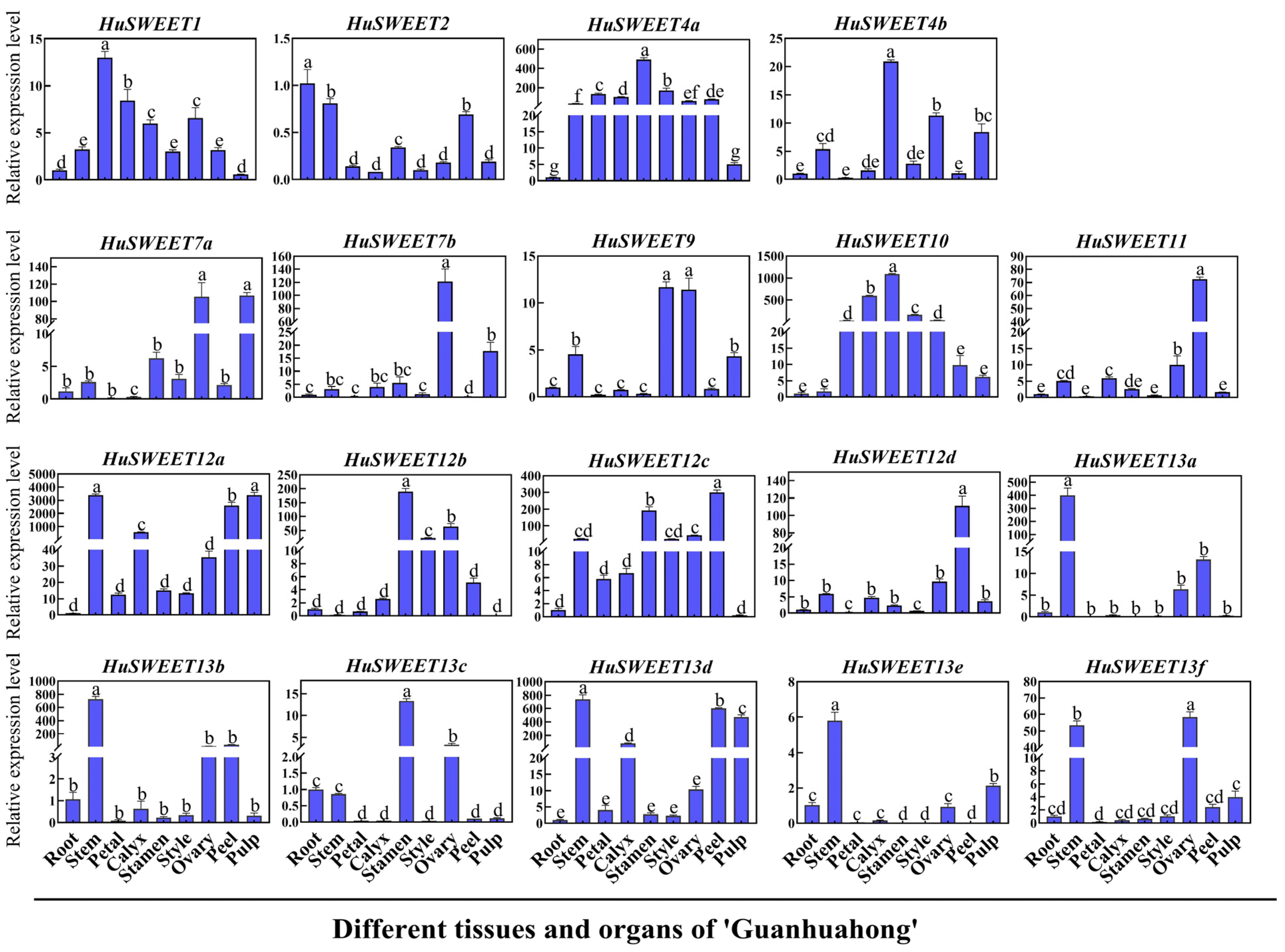
2.4. Expression Profiles of HuSWEETs in Different Pitaya Tissues and Organs
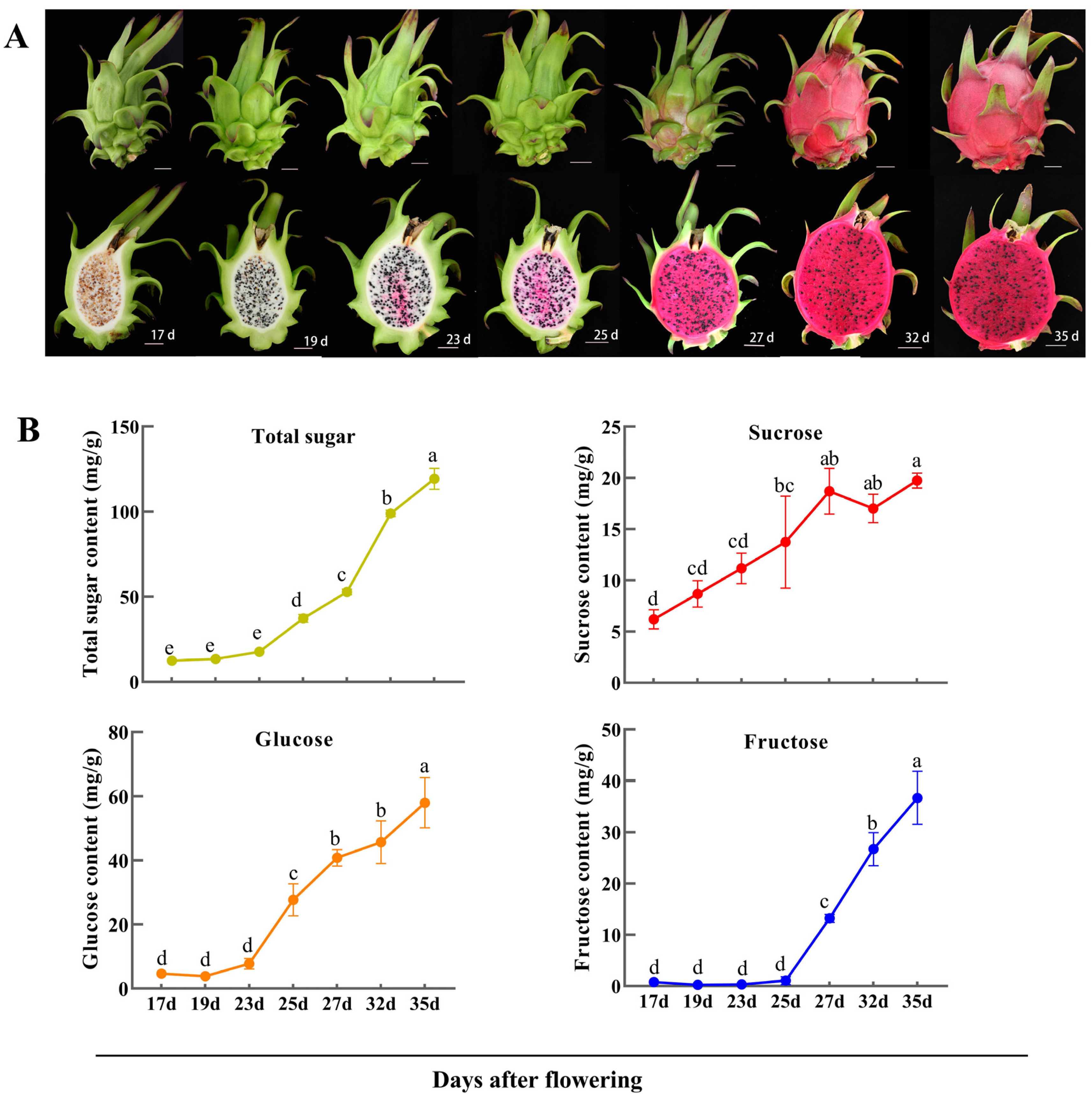
2.5. Changes in Soluble Sugars during Pitaya Fruit Development
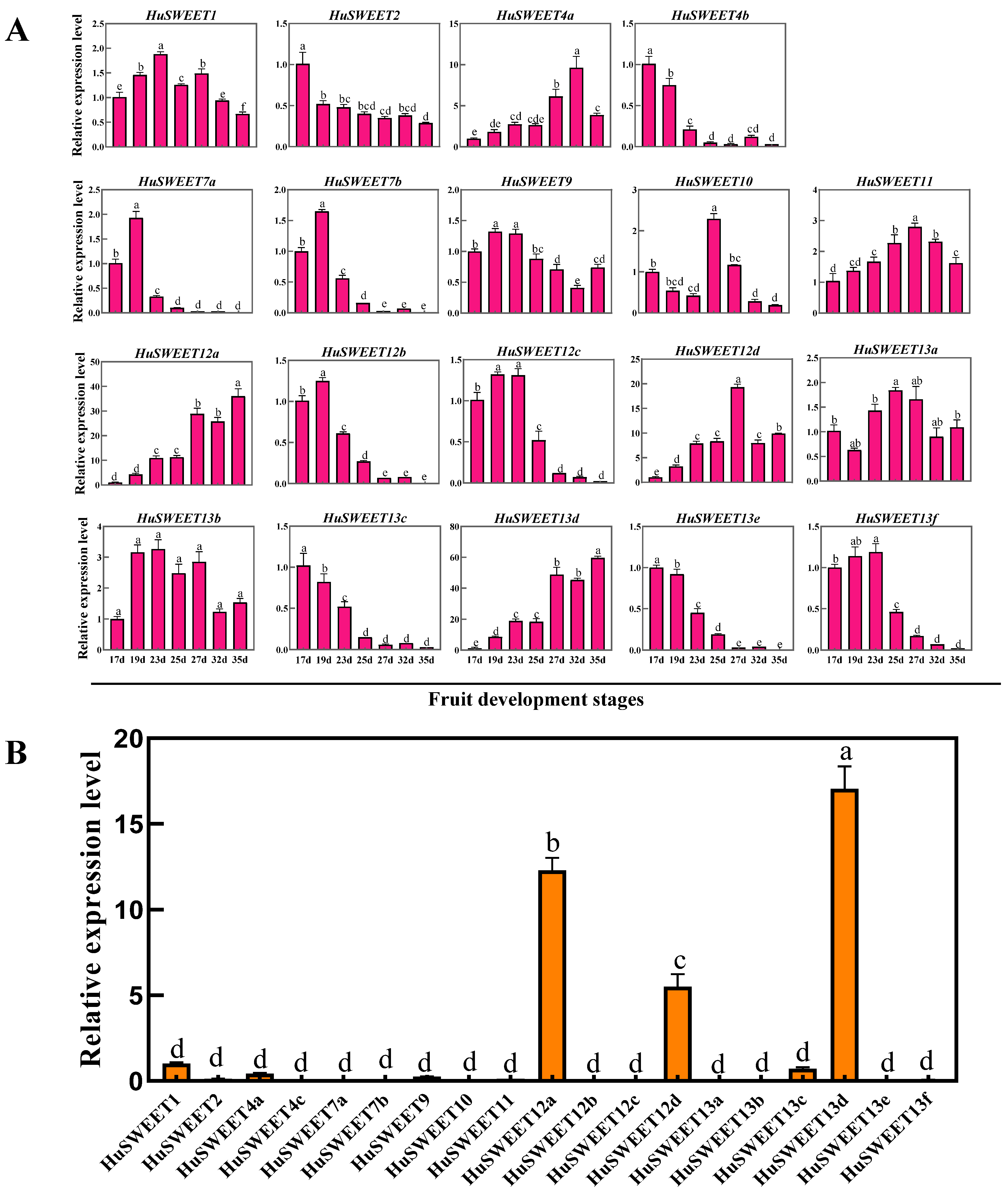
2.6. Expression Analyses of HuSWEETs in Pitaya Fruits
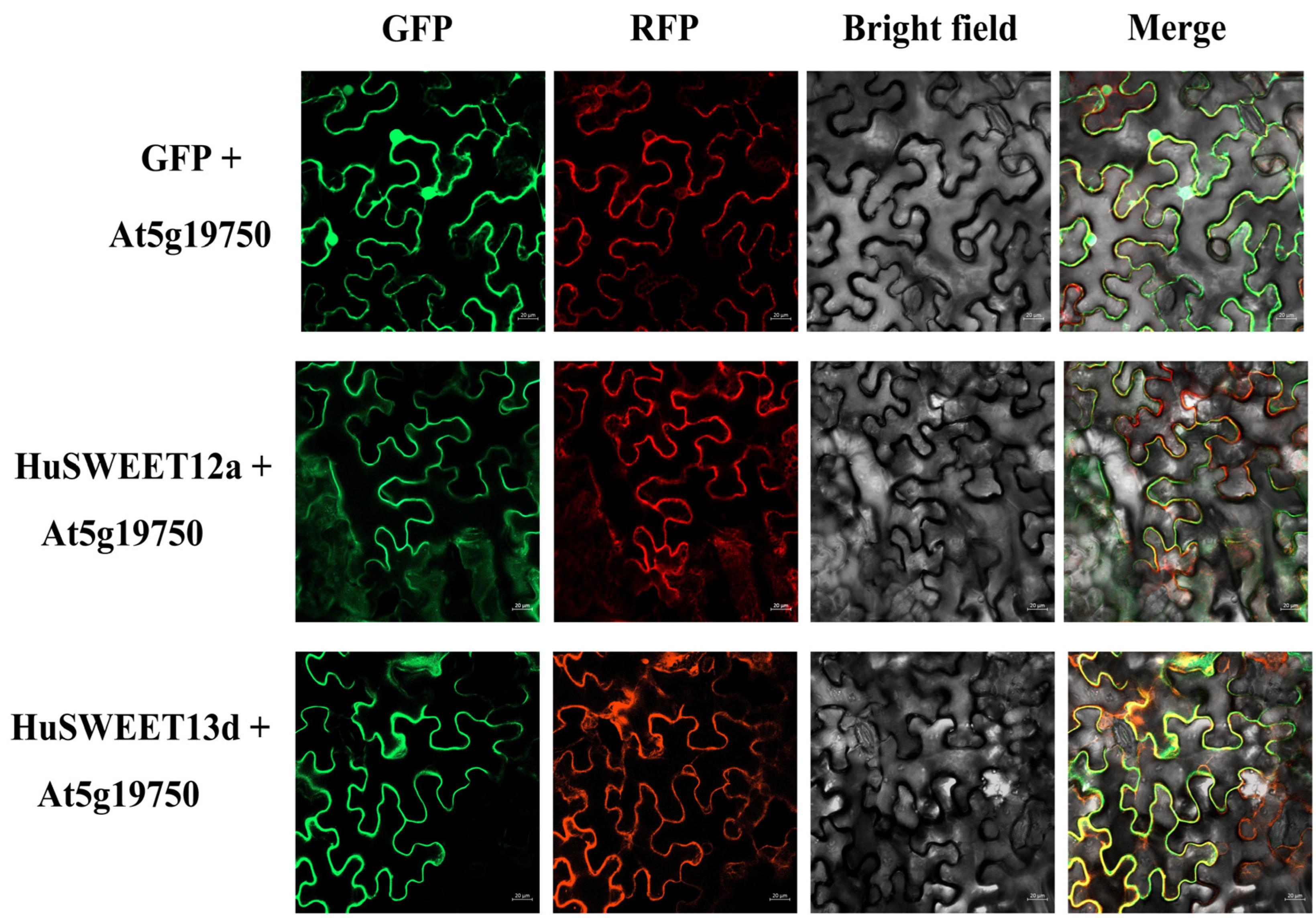
2.7. Plasma Membrane-Localized HuSWEET12a and HuSWEET13d Proteins
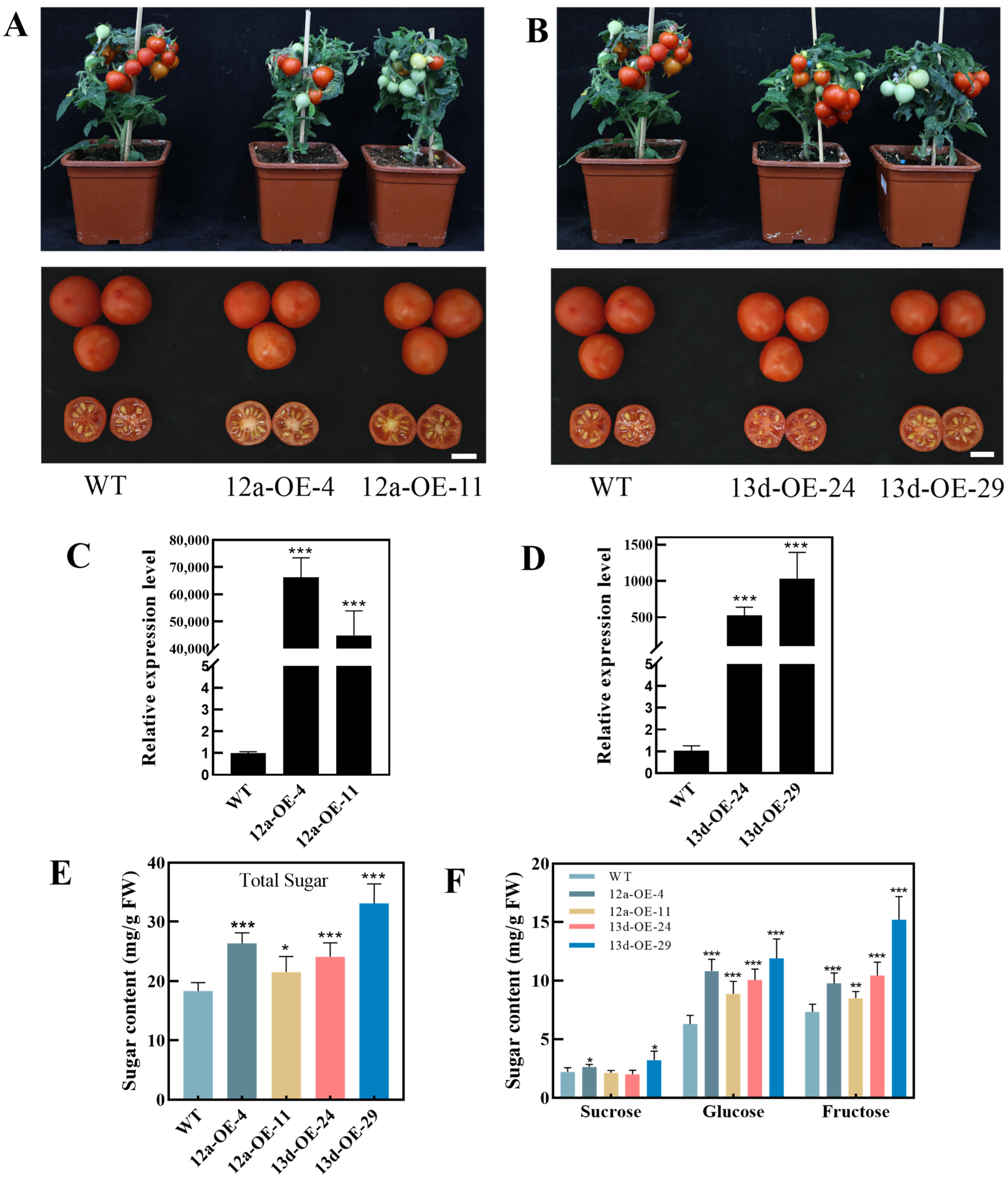
2.8. Overexpression of HuSWEET12a and HuSWEET13d Could Increase Sugar Contents of Tomato Fruits
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Measurement of Soluble Sugars
4.3. Identification of HuSWEET Family Genes in Pitaya
4.4. Gene Structure and Conserved Motifs Analyses of HuSWEETs
4.5. Locations in the Chromosome, Phylogenetic, and Duplication Pattern of HuSWEETs
4.6. RNA Isolation and Gene Expression Analyses
4.7. Subcellular Localization
4.8. Genetic Transformation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chen, L.Q.; Cheung, L.S.; Feng, L.; Tanner, W.; Frommer, W.B. Transport of sugars. Annu. Rev. Biochem. 2015, 84, 865–894. [Google Scholar] [CrossRef] [PubMed]
- Ruan, Y.L. Sucrose metabolism: Gateway to diverse carbon use and sugar signaling. Annu. Rev. Plant Biol. 2014, 65, 33–67. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.Q. SWEET sugar transporters for phloem transport and pathogen nutrition. New Phytol. 2014, 201, 1150–1155. [Google Scholar] [CrossRef]
- Lalonde, S.; Wipf, D.; Frommer, W.B. Transport mechanisms for organic forms of carbon and nitrogen between source and sink. Annu. Rev. Plant Biol. 2004, 55, 341–372. [Google Scholar] [CrossRef] [PubMed]
- Eom, J.S.; Chen, L.Q.; Sosso, D.; Julius, B.T.; Lin, I.W.; Qu, X.Q.; Braun, D.M.; Frommer, W.B. SWEETs, transporters for intracellular and intercellular sugar translocation. Curr. Opin. Plant Biol. 2015, 25, 53–62. [Google Scholar] [CrossRef]
- Chen, L.Q.; Hou, B.H.; Lalonde, S.; Takanaga, H.; Hartung, M.L.; Qu, X.Q.; Guo, W.J.; Kim, J.G.; Underwood, W.; Chaudhuri, B.; et al. Sugar transporters for intercellular exchange and nutrition of pathogens. Nature 2010, 468, 527–532. [Google Scholar] [CrossRef]
- Faham, S.; Watanabe, A.; Besserer, G.M.; Cascio, D.; Specht, A.; Hirayama, B.A.; Wright, E.M.; Abramson, J. The crystal structure of a sodium galactose transporter reveals mechanistic insights into Na+/sugar symport. Science 2008, 321, 810–814. [Google Scholar] [CrossRef]
- Xuan, Y.H.; Hu, Y.B.; Chen, L.Q.; Sosso, D.; Ducat, D.C.; Hou, B.H.; Frommer, W.B. Functional role of oligomerization for bacterial and plant SWEET sugar transporter family. Proc. Natl. Acad. Sci. USA 2013, 110, E3685–E3694. [Google Scholar] [CrossRef]
- Baker, R.F.; Leach, K.A.; Braun, D.M. SWEET as sugar: New sucrose effluxers in plants. Mol. Plant 2012, 5, 766–768. [Google Scholar] [CrossRef]
- Braun, D.M. Plant science. SWEET! The pathway is complete. Science 2012, 335, 173–174. [Google Scholar] [CrossRef]
- Yuan, M.; Wang, S. Rice MtN3/saliva/SWEET family genes and their homologs in cellular organisms. Mol. Plant 2013, 6, 665–674. [Google Scholar] [CrossRef]
- Patil, G.; Valliyodan, B.; Deshmukh, R.; Prince, S.; Nicander, B.; Zhao, M.; Sonah, H.; Song, L.; Lin, L.; Chaudhary, J.; et al. Soybean (Glycine max) SWEET gene family: Insights through comparative genomics, transcriptome profiling and whole genome re-sequence analysis. BMC Genom. 2015, 16, 520. [Google Scholar] [CrossRef]
- Chong, J.; Piron, M.C.; Meyer, S.; Merdinoglu, D.; Bertsch, C.; Mestre, P. The SWEET family of sugar transporters in grapevine: VvSWEET4 is involved in the interaction with Botrytis cinerea. J. Exp. Bot. 2014, 65, 6589–6601. [Google Scholar] [CrossRef]
- Zheng, Q.M.; Tang, Z.; Xu, Q.; Deng, X.X. Isolation, phylogenetic relationship and expression profiling of sugar transporter genes in sweet orange (Citrus sinensis). Plant Cell Tiss. Org. 2014, 119, 609–624. [Google Scholar] [CrossRef]
- Wei, X.; Liu, F.; Chen, C.; Ma, F.; Li, M. The Malus domestica sugar transporter gene family: Identifications based on genome and expression profiling related to the accumulation of fruit sugars. Front Plant Sci. 2014, 5, 569. [Google Scholar] [CrossRef] [PubMed]
- Feng, L.; Frommer, W.B. Structure and function of SemiSWEET and SWEET sugar transporters. Trends Biochem. Sci. 2015, 40, 480–486. [Google Scholar] [CrossRef]
- Xie, H.H.; Wang, D.; Qin, Y.Q.; Ma, A.N.; Fu, J.X.; Qin, Y.H.; Hu, G.B.; Zhao, J.T. Genome-wide identification and expression analysis of SWEET gene family in Litchi chinensis reveal the involvement of LcSWEET2a/3b in early seed development. BMC Plant Biol. 2019, 19, 499. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Ali, M.M.; Guo, T.; Alam, S.M.; Gull, S.; Iftikhar, J.; Yousef, A.F.; Mosa, W.F.A.; Chen, F. Genome-wide identification, in silico analysis and expression profiling of SWEET gene family in loquat (Eriobotrya japonica Lindl.). Agriculture 2022, 12, 1312. [Google Scholar] [CrossRef]
- Fatima, U.; Anjali, A.; Senthil-Kumar, M. AtSWEET11 and AtSWEET12: The twin traders of sucrose. Trends Plant Sci. 2022, 27, 958–960. [Google Scholar] [CrossRef]
- Ge, Y.X.; Angenent, G.C.; Dahlhaus, E.; Franken, J.; Peters, J.; Wullems, G.J.; Creemers-Molenaar, J. Partial silencing of the NEC1 gene results in early opening of anthers in Petunia hybrida. Mol. Genet. Genom. 2001, 265, 414–423. [Google Scholar] [CrossRef]
- Ge, Y.X.; Angenent, G.C.; Wittich, P.E.; Peters, J.; Franken, J.; Busscher, M.; Zhang, L.M.; Dahlhaus, E.; Kater, M.M.; Wullems, G.J.; et al. NEC1, a novel gene, highly expressed in nectary tissue of Petunia hybrida. Plant J. 2000, 24, 725–734. [Google Scholar] [CrossRef] [PubMed]
- Lin, I.W.; Sosso, D.; Chen, L.Q.; Gase, K.; Kim, S.G.; Kessler, D.; Klinkenberg, P.M.; Gorder, M.K.; Hou, B.H.; Qu, X.Q.; et al. Nectar secretion requires sucrose phosphate synthases and the sugar transporter SWEET9. Nature 2014, 508, 546–549. [Google Scholar] [CrossRef]
- Chen, L.Q.; Lin, I.W.; Qu, X.Q.; Sosso, D.; McFarlane, H.E.; Londoño, A.; Samuels, A.L.; Frommer, W.B. A cascade of sequentially expressed sucrose transporters in the seed coat and endosperm provides nutrition for the Arabidopsis embryo. Plant Cell 2015, 27, 607–619. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Luo, D.; Yang, B.; Frommer, W.B.; Eom, J.S. SWEET11 and 15 as key players in seed filling in rice. New Phytol. 2018, 218, 604–615. [Google Scholar] [CrossRef] [PubMed]
- Durand, M.; Porcheron, B.; Hennion, N.; Maurousset, L.; Lemoine, R.; Pourtau, N. Water deficit enhances C export to the roots in Arabidopsis thaliana plants with contribution of sucrose transporters in both shoot and roots. Plant Physiol. 2016, 170, 1460–1479. [Google Scholar] [CrossRef] [PubMed]
- Hir, R.L.; Spinner, L.; Klemens, P.A.; Chakraborti, D.; de Marco, F.; Vilaine, F.; Wolff, N.; Lemoine, R.; Porcheron, B.; Géry, C.; et al. Disruption of the sugar transporters AtSWEET11 and AtSWEET12 affects vascular development and freezing tolerance in Arabidopsis. Mol. Plant 2015, 8, 1687–1690. [Google Scholar] [CrossRef]
- Jing, L.U.; Sun, M.H.; Qi-Jun, M.A.; Kang, H.; Liu, Y.J.; Hao, Y.J.; You, C.X. MdSWEET17,a sugar transporter in apple, enhances drought tolerance in tomato. J. Integr. Agric. 2019, 18, 11. [Google Scholar] [CrossRef]
- Sun, M.X.; Huang, X.Y.; Yang, J.; Guan, Y.F.; Yang, Z.N. Arabidopsis RPG1 is important for primexine deposition and functions redundantly with RPG2 for plant fertility at the late reproductive stage. Plant Reprod. 2013, 26, 83–91. [Google Scholar] [CrossRef]
- Zhang, X.S.; Feng, C.Y.; Wang, M.N.; Li, T.L.; Liu, X.; Jiang, J. Plasma membrane-localized SlSWEET7a and SlSWEET14 regulate sugar transport and storage in tomato fruits. Hortic. Res. 2021, 8, 186. [Google Scholar] [CrossRef]
- Zhang, C.; Bian, Y.; Hou, S.; Li, X. Sugar transport played a more important role than sugar biosynthesis in fruit sugar accumulation during Chinese jujube domestication. Planta 2018, 248, 1187–1199. [Google Scholar] [CrossRef]
- Ren, Y.; Li, M.; Guo, S.; Sun, H.; Zhao, J.; Zhang, J.; Liu, G.; He, H.; Tian, S.; Yu, Y.; et al. Evolutionary gain of oligosaccharide hydrolysis and sugar transport enhanced carbohydrate partitioning in sweet watermelon fruits. Plant Cell 2021, 33, 1554–1573. [Google Scholar] [CrossRef] [PubMed]
- Fang, T.; Peng, Y.; Rao, Y.; Li, S.; Zeng, L. Genome-wide identification and expression analysis of sugar transporter (ST) gene family in longan (Dimocarpus longan L.). Plants 2020, 9, 342. [Google Scholar] [CrossRef] [PubMed]
- Shammai, A.; Petreikov, M.; Yeselson, Y.; Faigenboim, A.; Moy-Komemi, M.; Cohen, S.; Cohen, D.; Besaulov, E.; Efrati, A.; Houminer, N.; et al. Natural genetic variation for expression of a SWEET transporter among wild species of Solanum lycopersicum (tomato) determines the hexose composition of ripening tomato fruit. Plant J. 2018, 96, 343–357. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Liu, H.; Yao, X.; Wang, J.; Feng, S.; Sun, L.; Ma, S.; Xu, K.; Chen, L.Q.; Sui, X. Hexose transporter CsSWEET7a in cucumber mediates phloem unloading in companion cells for fruit development. Plant Physiol. 2021, 186, 640–654. [Google Scholar] [CrossRef]
- Zhang, Z.; Zou, L.; Ren, C.; Ren, F.; Wang, Y.; Fan, P.; Li, S.H.; Liang, Z.C. VvSWEET10 mediates sugar accumulation in grapes. Genes 2019, 10, 255. [Google Scholar] [CrossRef]
- Liu, D.D.; Chao, W.M.; Turgeon, R. Transport of sucrose, not hexose, in the phloem. J. Exp. Bot. 2012, 63, 4315–4320. [Google Scholar] [CrossRef]
- Hu, X.; Li, S.; Lin, X.; Fang, H.; Shi, Y.; Grierson, D.; Chen, K. Transcription factor CitERF16 is Involved in citrus fruit sucrose accumulation by activating CitSWEET11d. Front Plant Sci. 2021, 12, 809619. [Google Scholar] [CrossRef]
- Li, X.; Guo, W.; Li, J.; Yue, P.; Bu, H.; Jiang, J.; Liu, W.; Xu, Y.; Yuan, H.; Li, T.; et al. Histone acetylation at the promoter for the transcription factor PuWRKY31 affects sucrose accumulation in pear fruit. Plant Physiol. 2020, 182, 2035–2046. [Google Scholar] [CrossRef]
- Ibrahim, S.; Mohamed, G.A.; Khedr, A.; Zayed, M.F.; El-Kholy, A.A.E.S. Genus Hylocereus: Beneficial phytochemicals, nutritional importance, and biological relevance—A review. J. Food Biochem. 2018, 42, e12491. [Google Scholar] [CrossRef]
- Hua, Q.; Chen, C.; Tel Zur, N.; Wang, H.; Wu, J.; Chen, J.; Zhang, Z.; Zhao, J.; Hu, G.; Qin, Y. Metabolomic characterization of pitaya fruit from three red-skinned cultivars with different pulp colors. Plant Physiol. Biochem. 2018, 126, 117–125. [Google Scholar] [CrossRef]
- Xie, F.; Chen, C.; Chen, J.; Yuan, Y.; Hua, Q.; Zhang, Z.; Zhao, J.; Hu, G.; Chen, J.; Qin, Y. Metabolic profiling of sugars and organic acids, and expression analyses of metabolism-associated genes in two yellow-peel pitaya species. Plants 2022, 11, 694. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.K.; Xing, Y.M.; Ramakrishnan, M.; Chen, C.B.; Xie, F.F.; Hua, Q.Z.; Chen, J.Y.; Zhang, R.; Zhao, J.T.; Hu, G.B.; et al. Transcriptomic-based identification and characterization of genes related to sugar metabolism in ‘Hongshuijing’ pitaya. Hortic. Plant J. 2022, 8, 450–460. [Google Scholar] [CrossRef]
- Katz, E.; Boo, K.H.; Kim, H.Y.; Eigenheer, R.A.; Phinney, B.S.; Shulaev, V.; Negre-Zakharov, F.; Sadka, A.; Blumwald, E. Label-free shotgun proteomics and metabolite analysis reveal a significant metabolic shift during citrus fruit development. J. Exp. Bot. 2011, 62, 5367–5384. [Google Scholar] [CrossRef] [PubMed]
- Wei, W.; Cheng, M.N.; Ba, L.J.; Zeng, R.X.; Luo, D.L.; Qin, Y.H.; Liu, Z.L.; Kuang, J.F.; Lu, W.J.; Chen, J.Y.; et al. Pitaya HpWRKY3 is associated with fruit sugar accumulation by transcriptionally modulating sucrose metabolic genes HpINV2 and HpSuSy1. Int. J. Mol. Sci. 2019, 20, 1890. [Google Scholar] [CrossRef]
- Chen, J.Y.; Xie, F.F.; Cui, Y.Z.; Chen, C.B.; Lu, W.J.; Hu, X.D.; Hua, Q.Z.; Zhao, J.; Wu, Z.J.; Gao, D.; et al. A chromosome-scale genome sequence of pitaya (Hylocereus undatus) provides novel insights into the genome evolution and regulation of betalain biosynthesis. Hortic. Res. 2021, 8, 164. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Chen, C.J.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.H.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Gautam, T.; Saripalli, G.; Gahlaut, V.; Kumar, A.; Sharma, P.K.; Balyan, H.S.; Gupta, P.K. Further studies on sugar transporter (SWEET) genes in wheat (Triticum aestivum L.). Mol. Biol. Rep. 2019, 46, 2327–2353. [Google Scholar] [CrossRef]
- Yuan, M.; Zhao, J.; Huang, R.; Li, X.; Xiao, J.; Wang, S. Rice MtN3/saliva/SWEET gene family: Evolution, expression profiling, and sugar transport. J. Integr. Plant Biol. 2014, 56, 559–570. [Google Scholar] [CrossRef]
- Jia, B.; Zhu, X.F.; Pu, Z.J.; Duan, Y.X.; Hao, L.J.; Zhang, J.; Chen, L.Q.; Jeon, C.O.; Xuan, Y.H. Integrative view of the diversity and evolution of SWEET and SemiSWEET sugar transporters. Front Plant Sci. 2017, 8, 2178. [Google Scholar] [CrossRef]
- Miao, H.; Sun, P.; Liu, Q.; Miao, Y.; Liu, J.; Zhang, K.; Hu, W.; Zhang, J.; Wang, J.; Wang, Z.; et al. Genome-wide analyses of SWEET family proteins reveal involvement in fruit development and abiotic/biotic stress responses in banana. Sci. Rep. 2017, 7, 3536. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.P.; Zhang, F.; Song, S.H.; Tang, X.W.; Xu, H.; Liu, G.M.; Wang, Y.; He, H.J. Genome-wide identification, characterization, and expression analysis of the SWEET gene family in cucumber. J. Integr. Agric. 2017, 16, 1486–1501. [Google Scholar] [CrossRef]
- Nie, P.; Xu, G.; Yu, B.; Lyu, D.; Xue, X.; Qin, S. Genome-wide identification and expression profiling reveal the potential functions of the SWEET gene family during the sink organ development period in apple (Malus × domestica Borkh.). Agronomy 2022, 12, 1747. [Google Scholar] [CrossRef]
- Xu, G.; Guo, C.; Shan, H.; Kong, H. Divergence of duplicate genes in exon-intron structure. Proc. Natl. Acad. Sci. USA 2012, 109, 1187–1192. [Google Scholar] [CrossRef]
- Jeena, G.S.; Kumar, S.; Shukla, R.K. Structure, evolution and diverse physiological roles of SWEET sugar transporters in plants. Plant Mol. Biol. 2019, 100, 351–365. [Google Scholar] [CrossRef]
- Moriyama, E.N.; Strope, P.K.; Opiyo, S.O.; Chen, Z.; Jones, A.M. Mining the Arabidopsis thaliana genome for highly-divergent seven transmembrane receptors. Genome Biol. 2006, 7, R96. [Google Scholar] [CrossRef]
- Wang, P.; Wei, P.; Niu, F.; Liu, X.; Zhang, H.; Lyu, M.; Yuan, Y.; Wu, B. Cloning and functional assessments of floral-expressed SWEET transporter genes from Jasminum sambac. Int. J. Mol. Sci. 2019, 20, 4001. [Google Scholar] [CrossRef]
- Chen, H.Y.; Huh, J.H.; Yu, Y.C.; Ho, L.H.; Chen, L.Q.; Tholl, D.; Frommer, W.B.; Guo, W.J. The Arabidopsis vacuolar sugar transporter SWEET2 limits carbon sequestration from roots and restricts Pythium infection. Plant J. 2015, 83, 1046–1058. [Google Scholar] [CrossRef]
- Zhen, Q.L.; Fang, T.; Peng, Q.; Liao, L.; Zhao, L.; Owiti, A.; Han, Y.P. Developing gene-tagged molecular markers for evaluation of genetic association of apple SWEET genes with fruit sugar accumulation. Hortic. Res. 2018, 5, 14. [Google Scholar] [CrossRef]
- Guo, C.; Li, H.; Xia, X.; Liu, X.; Yang, L. Functional and evolution characterization of SWEET sugar transporters in Ananas comosus. Biochem. Biophys. Res. Commun. 2018, 496, 407–414. [Google Scholar] [CrossRef]
- Ye, Y.X.; Hu, G.B.; Li, J.X.; Qin, Y.H.; Gu, W.Q.; Zhai, X.; Liu, Z.X.; Li, J.X. A new red-fleshed pitaya cultivar ‘Guanhuahong’. Acta Hortic. Sinica 2018, 45, 2267–2268. [Google Scholar] [CrossRef]
- Finn, R.D.; Clements, J.; Eddy, S.R. HMMER web server: Interactive sequence similarity searching. Nucleic Acids Res. 2011, 39, W29–W37. [Google Scholar] [CrossRef] [PubMed]
- Finn, R.D.; Mistry, J.; Tate, J.; Coggill, P.; Heger, A.; Pollington, J.E.; Gavin, O.L.; Gunasekaran, P.; Ceric, G.; Forslund, K.; et al. The Pfam protein families database. Nucleic Acids Res. 2010, 38, D211–D222. [Google Scholar] [CrossRef]
- Zhang, H.; Gao, S.; Lercher, M.J.; Hu, S.; Chen, W.H. EvolView, an online tool for visualizing, annotating and managing phylogenetic trees. Nucleic Acids Res. 2012, 40, W569–W572. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.B.; Wu, J.Y.; Hua, Q.Z.; Tel-Zur, N.; Xie, F.F.; Zhang, Z.K.; Chen, J.Y.; Zhang, R.; Hu, G.B.; Zhao, J.T.; et al. Identification of reliable reference genes for quantitative real-time PCR normalization in pitaya. Plant Methods 2019, 15, 70. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 4, 402–408. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID | Gene Names | Length of Open Reading Frames (bp) | No. Amino Acids (aa) | Isoelectric Points | Molecular Weights (kDa) | Hydrophilicity | Instability Index |
|---|---|---|---|---|---|---|---|
| HU07G00905.1 | HuSWEET1 | 774 | 257 | 9.32 | 28.01 | 0.477 | 27.48 |
| HU08G01881.1 | HuSWEET2 | 708 | 235 | 9.17 | 26.30 | 0.87 | 56.04 |
| HU09G00864.1 | HuSWEET4a | 729 | 242 | 9.62 | 27.04 | 0.555 | 36.88 |
| HU09G00626.1 | HuSWEET4b | 732 | 243 | 9.32 | 27.03 | 0.53 | 28.03 |
| HU04G02229.1 | HuSWEET7a | 771 | 256 | 9.24 | 27.83 | 0.783 | 44.92 |
| HU04G00351.1 | HuSWEET7b | 771 | 256 | 9.28 | 27.86 | 0.788 | 46.82 |
| HU03G01299.1 | HuSWEET9 | 774 | 257 | 9.47 | 28.33 | 0.557 | 35.88 |
| HU08G01549.1 | HuSWEET10 | 783 | 260 | 9.32 | 28.57 | 0.635 | 36.59 |
| HU01G02213.1 | HuSWEET11 | 882 | 293 | 9.12 | 33.12 | 0.34 | 48.54 |
| HU04G01326.1 | HuSWEET12a | 1008 | 335 | 7.02 | 37.37 | 0.542 | 41.98 |
| HU04G01322.1 | HuSWEET12b | 906 | 301 | 9.14 | 34.13 | 0.772 | 41.03 |
| HU01G02212.1 | HuSWEET12c | 930 | 309 | 9.45 | 34.74 | 0.593 | 39.71 |
| HU04G01323.1 | HuSWEET12d | 957 | 318 | 8.74 | 35.53 | 0.501 | 42.51 |
| HU04G01325.1 | HuSWEET13a | 948 | 315 | 8.96 | 34.87 | 0.534 | 37.13 |
| HU01G01645.1 | HuSWEET13b | 885 | 294 | 8.85 | 32.44 | 0.486 | 38.55 |
| HU02G01006.1 | HuSWEET13c | 1002 | 333 | 5.71 | 37.57 | 0.368 | 107.12 |
| HU04G01327.1 | HuSWEET13d | 870 | 289 | 8.32 | 32.14 | 0.391 | 46.8 |
| HU07G01376.1 | HuSWEET13e | 999 | 332 | 8.18 | 37.39 | 0.387 | 39.42 |
| HU02G01007.1 | HuSWEET13f | 1029 | 342 | 8.15 | 38.39 | 0.356 | 36.9 |
| Genes | Soluble Sugar | Glucose | Fructose | Sucrose |
|---|---|---|---|---|
| HuSWEET1 | −0.689 | −0.598 | −0.709 | −0.332 |
| HuSWEET2 | −0.587 | −0.701 | −0.545 | −0.840 * |
| HuSWEET4a | 0.684 | 0.689 | 0.637 | 0.704 |
| HuSWEET4c | −0.583 | −0.763 | −0.532 | −0.885 ** |
| HuSWEET7a | −0.591 | −0.75 | −0.549 | −0.775 * |
| HuSWEET7b | −0.668 | −0.838 * | −0.629 | −0.851 * |
| HuSWEET9 | −0.771 * | −0.839 * | −0.741 | −0.729 |
| HuSWEET10 | −0.462 | −0.161 | −0.497 | −0.164 |
| HuSWEET11 | 0.322 | 0.588 | 0.267 | 0.728 |
| HuSWEET12a | 0.905 ** | 0.955 ** | 0.898 ** | 0.972 ** |
| HuSWEET12b | −0.762 * | −0.915 ** | −0.725 | −0.934 ** |
| HuSWEET12c | −0.839 * | −0.961 ** | −0.821 * | −0.891 ** |
| HuSWEET12d | 0.387 | 0.63 | 0.371 | 0.803 * |
| HuSWEET13a | −0.089 | 0.258 | −0.144 | 0.345 |
| HuSWEET13b | −0.411 | −0.333 | −0.432 | −0.097 |
| HuSWEET13c | −0.727 | −0.898 ** | −0.682 | −0.959 ** |
| HuSWEET13d | 0.908 ** | 0.951 ** | 0.900 ** | 0.972 ** |
| HuSWEET13e | −0.739 | −0.897 ** | −0.695 | −0.957 ** |
| HuSWEET13f | −0.854 * | −0.973 ** | −0.834 * | −0.910 ** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, R.; Wu, L.; Zeng, J.; Shah, K.; Zhang, R.; Hu, G.; Qin, Y.; Zhang, Z. Identification of HuSWEET Family in Pitaya (Hylocereus undatus) and Key Roles of HuSWEET12a and HuSWEET13d in Sugar Accumulation. Int. J. Mol. Sci. 2023, 24, 12882. https://doi.org/10.3390/ijms241612882
Jiang R, Wu L, Zeng J, Shah K, Zhang R, Hu G, Qin Y, Zhang Z. Identification of HuSWEET Family in Pitaya (Hylocereus undatus) and Key Roles of HuSWEET12a and HuSWEET13d in Sugar Accumulation. International Journal of Molecular Sciences. 2023; 24(16):12882. https://doi.org/10.3390/ijms241612882
Chicago/Turabian StyleJiang, Rui, Liangfang Wu, Jianmei Zeng, Kamran Shah, Rong Zhang, Guibing Hu, Yonghua Qin, and Zhike Zhang. 2023. "Identification of HuSWEET Family in Pitaya (Hylocereus undatus) and Key Roles of HuSWEET12a and HuSWEET13d in Sugar Accumulation" International Journal of Molecular Sciences 24, no. 16: 12882. https://doi.org/10.3390/ijms241612882