
Evaluation of the Antiseizure Activity of Endemic Plant Halfordia kendack Guillaumin and Its Main Constituent, Halfordin, on a Zebrafish Pentylenetetrazole (PTZ)-Induced Seizure Model
 , , , , , , and
, , , , , , and
Abstract
:1. Introduction
2. Results
2.1. LLC Isolation of Halfordin
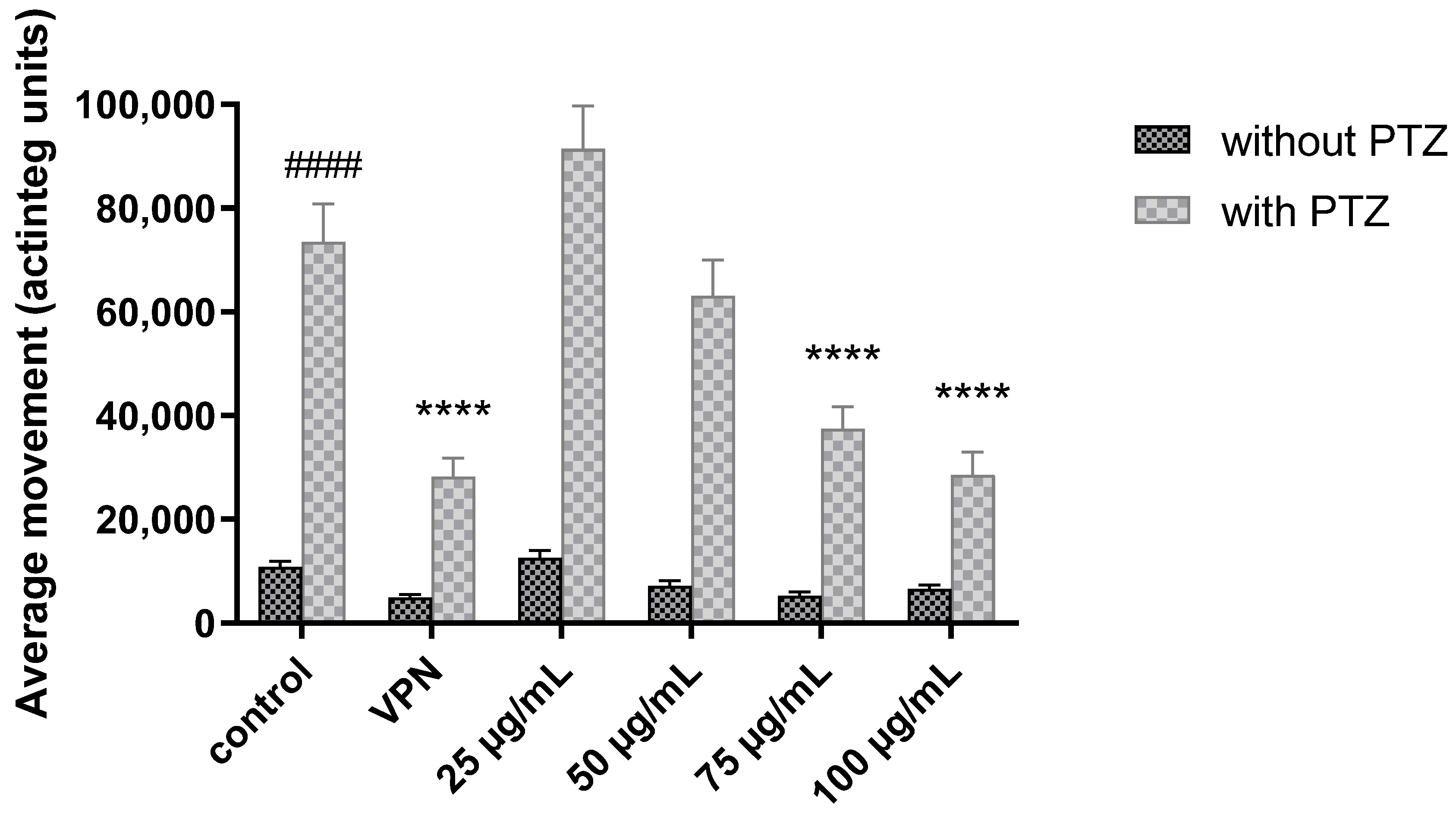
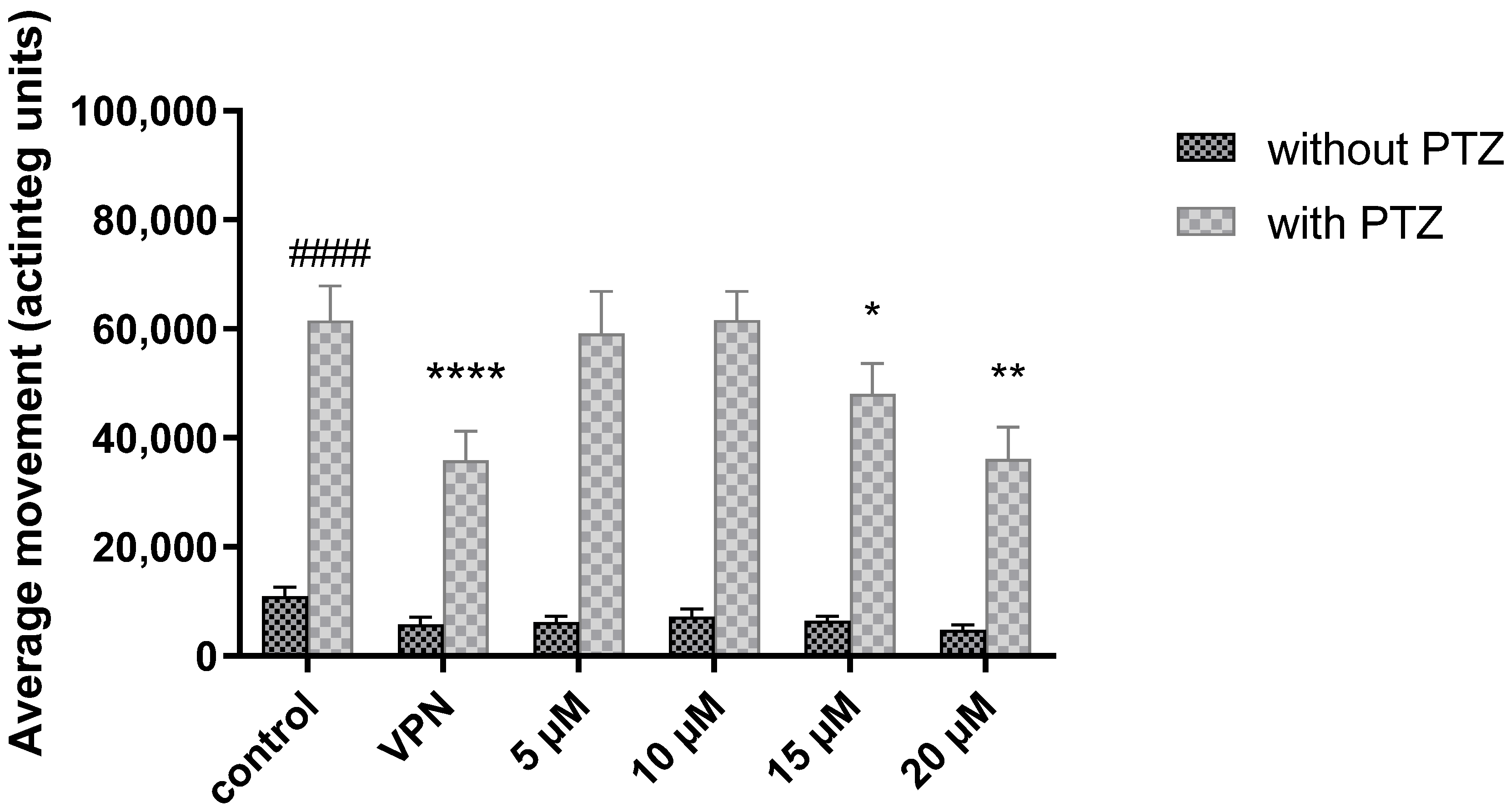
2.2. Zebrafish Locomotor Assay
2.3. LFP Recordings
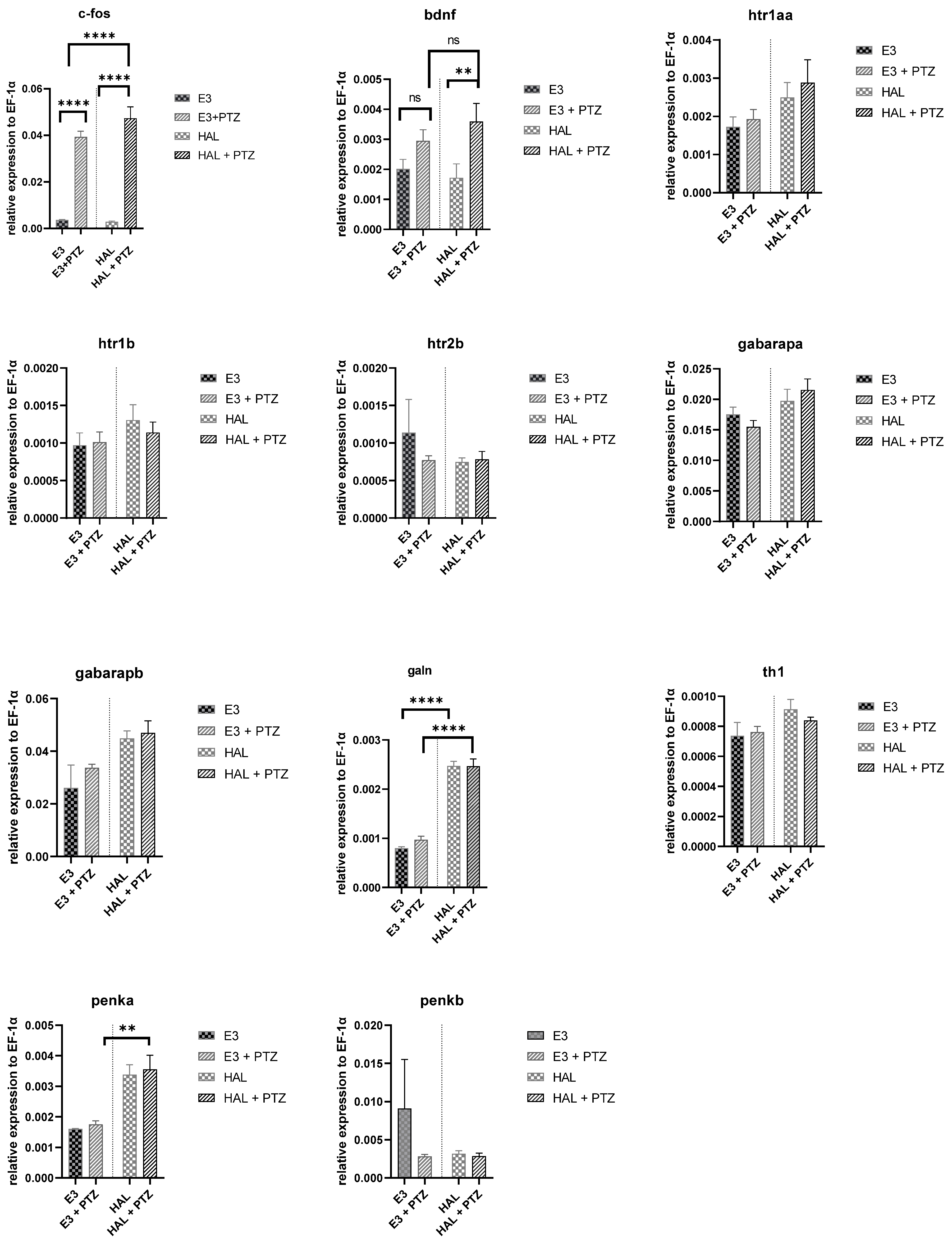
2.4. Gene Expression
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Plant Material and Extraction
4.3. Liquid-Liquid Chromatography Isolation
4.3.1. Liquid-Liquid Chromatography (LLC) Unit
4.3.2. Selection of Biphasic Solvent System (Shake Flask Experiments)
4.3.3. Preparation of the Biphasic Solvent System
4.3.4. LLC Separation
4.3.5. HPLC-DAD Analysis
4.3.6. Structure Elucidation
4.4. Zebrafish Experiments
4.4.1. Maintaining Zebrafish
4.4.2. Toxicity Assay
4.4.3. Antiepileptic Evaluation
4.4.4. LFP Recordings
4.4.5. RNA Isolation, Reverse Transcription and Quantitative PCR
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- WHO. Epilepsy: A Public Health Imperative; WHO: Geneva, Switzerland, 2019. [Google Scholar]
- Avalos, J.C.; Silva, B.A.; Tevés Echazu, M.F.; Rosso, B.; Besocke, A.G.; Del Carmen Garcia, M. Quality of life in patients with epilepsy or psychogenic nonepileptic seizures and the contribution of psychiatric comorbidities. Epilepsy Behav. 2020, 112, 107447. [Google Scholar] [CrossRef] [PubMed]
- Bialer, M.; Johannessen, S.I.; Levy, R.H.; Perucca, E.; Tomson, T.; White, H.S. Progress report on new antiepileptic drugs: A summary of the Ninth Eilat Conference (EILAT IX). Epilepsy Res. 2009, 83, 1–43. [Google Scholar] [CrossRef] [PubMed]
- Bialer, M.; Johannessen, S.I.; Levy, R.H.; Perucca, E.; Tomson, T.; White, H.S. Progress report on new antiepileptic drugs: A summary of the Twelfth Eilat Conference (EILAT XII). Epilepsy Res. 2015, 111, 85–141. [Google Scholar] [CrossRef] [PubMed]
- Golub, V.; Reddy, D.S. Cannabidiol Therapy for Refractory Epilepsy and Seizure Disorders. Adv. Exp. Med. Biol. 2021, 1264, 93–110. [Google Scholar] [PubMed]
- Kozioł, E.; Luca, S.V.; Marcourt, L.; Nour, M.; Hnawia, E.; Jakubowicz-Gil, J.; Paduch, R.; Mroczek, T.; Wolfender, J.-L.; Skalicka-Woźniak, K. Efficient extraction and isolation of skimmianine from New Caledonian plant Medicosma leratii and evaluation of its effects on apoptosis, necrosis, and autophagy. Phytochem. Lett. 2019, 30, 224–230. [Google Scholar] [CrossRef]
- Sultana, N.; Skelton, B.W.; Waterman, P.G.; White, A.H. 3,5-Dimethoxy-3-(1,1-dimethylprop-2-enyl)-3,4-dihydropsoralen-4-one: An unusual furanocoumarin derivative from Halfordia kendack. Phytochemistry 2000, 55, 675–678. [Google Scholar] [CrossRef]
- Sultana, N.; Waterman, P.G. 3-Monoterpenyl-2,4-dioxygenated quinoline alkaloids from the aerial parts of Halfordia kendack. Phytochemistry 2001, 58, 329–332. [Google Scholar] [CrossRef]
- Zagaja, M.; Pyrka, D.; Skalicka-Wozniak, K.; Glowniak, K.; Florek-Luszczki, M.; Glensk, M.; Luszczki, J.J. Effect of xanthotoxin (8-methoxypsoralen) on the anticonvulsant activity of classical antiepileptic drugs against maximal electroshock-induced seizures in mice. Fitoterapia 2015, 105, 1–6. [Google Scholar] [CrossRef]
- Kowalczyk, J.; Nakos-Bimpos, M.; Polissidis, A.; Dalla, C.; Kokras, N.; Skalicka-Wozniak, K.; Budzynska, B. Xanthotoxin affects depression-related behavior and neurotransmitters content in a sex-dependent manner in mice. Behav. Brain Res. 2021, 399, 112985. [Google Scholar] [CrossRef]
- Budzynska, B.; Boguszewska-Czubara, A.; Kruk-Slomka, M.; Skalicka-Wozniak, K.; Michalak, A.; Musik, I.; Biala, G. Effects of imperatorin on scopolamine-induced cognitive impairment and oxidative stress in mice. Psychopharmacology 2015, 232, 931–942. [Google Scholar] [CrossRef]
- Liu, L.; Bao, L.; Wang, L.; Ma, K.; Han, J.; Yang, Y.; Liu, R.; Ren, J.; Yin, W.; Wang, W.; et al. Asperorydines A-M: Prenylated Tryptophan-Derived Alkaloids with Neurotrophic Effects from Aspergillus oryzae. J. Org. Chem. 2018, 83, 812–822. [Google Scholar] [CrossRef] [PubMed]
- Gulsun, T.; Ucar, B.; Sahin, S.; Humpel, C. The Organic Cation Transporter 2 Inhibitor Quinidine Modulates the Neuroprotective Effect of Nerve Growth Factor and Memantine on Cholinergic Neurons of the Basal Nucleus of Meynert in Organotypic Brain Slices. Pharmacology 2021, 106, 390–399. [Google Scholar] [CrossRef] [PubMed]
- Fukuoka, M.; Kuki, I.; Kawawaki, H.; Okazaki, S.; Kim, K.; Hattori, Y.; Tsuji, H.; Nukui, M.; Inoue, T.; Yoshida, Y.; et al. Quinidine therapy for West syndrome with KCNTI mutation: A case report. Brain Dev. 2017, 39, 80–83. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Zhang, D.; Ren, J.; Yang, M. Skimmianine, a furoquinoline alkaloid from Zanthoxylum nitidum as a potential acetylcholinesterase inhibitor. Med. Chem. Res. 2012, 21, 722–725. [Google Scholar] [CrossRef]
- Afrikanova, T.; Serruys, A.S.K.; Buenafe, O.E.M.; Clinckers, R.; Smolders, I.; de Witte, P.A.M.; Crawford, A.D.; Esguerra, C.V. Validation of the Zebrafish Pentylenetetrazol Seizure Model: Locomotor versus Electrographic Responses to Antiepileptic Drugs. PLoS ONE 2013, 8, e54166. [Google Scholar] [CrossRef] [Green Version]
- Orellana-Paucar, A.M.; Serruys, A.S.K.; Afrikanova, T.; Maes, J.; De Borggraeve, W.; Alen, J.; León-Tamariz, F.; Wilches-Arizábala, I.M.; Crawford, A.D.; de Witte, P.A.; et al. Anticonvulsant activity of bisabolene sesquiterpenoids of Curcuma longa in zebrafish and mouse seizure models. Epilepsy Behav. 2012, 24, 14–22. [Google Scholar] [CrossRef]
- MacRae, C.A.; Peterson, R.T. Zebrafish as tools for drug discovery. Nat. Rev. Drug Discov. 2015, 14, 721–731. [Google Scholar] [CrossRef]
- Baraban, S.C.; Taylor, M.R.; Castro, P.A.; Baier, H. Pentylenetetrazole induced changes in zebrafish behavior, neural activity and c-fos expression. Neuroscience 2005, 131, 759–768. [Google Scholar] [CrossRef]
- Ito, Y. Countercurrent chromatography. J. Biochem. Biophys. Methods 1981, 17, 65–143. [Google Scholar] [CrossRef]
- Kedzierski, B.; Kukuła-Koch, W.; Głowniak, K. Application of CPC and related methods for the isolation of natural substances--a review. Acta Pol. Pharm. 2014, 71, 223–227. [Google Scholar]
- Widelski, J.; Luca, S.V.; Skiba, A.; Maciąg, M.; Budzyńska, B.; Marcourt, L.; Wolfender, J.-L.; Skalicka-Woźniak, K. Coumarins from Seseli devenyense Simonk: Isolation by Liquid-Liquid Chromatography and Potential Anxiolytic Activity Using an In Vivo Zebrafish Larvae Model. Int. J. Mol. Sci. 2021, 22, 1829. [Google Scholar] [CrossRef] [PubMed]
- Ammar, S.; Abidi, J.; Vlad Luca, S.; Boumendjel, M.; Skalicka-Woźniak, K.; Bouaziz, M. Untargeted metabolite profiling and phytochemical analysis based on RP-HPLC-DAD-QTOF-MS and MS/MS for discovering new bioactive compounds in Rumex algeriensis flowers and stems. Phytochem. Anal. 2020, 31, 616–635. [Google Scholar] [CrossRef] [PubMed]
- Garrard, I.J. Simple Approach to the Development of a CCC Solvent Selection Protocol Suitable for Automation. J. Liq. Chromatogr. Relat. Technol. 2005, 28, 1923–1935. [Google Scholar] [CrossRef]
- Hegarty, M.P.; Lahey, F.N. The coumarins of Halfordia scleroxyla F. Muell. Aust. J. Chem. 1956, 9, 120–131. [Google Scholar] [CrossRef]
- Quek, A.; Kassim, N.K.; Lim, P.C.; Tan, D.C.; Mohammad Latif, M.A.; Ismail, A.; Shaari, K.; Awang, K. α-Amylase and dipeptidyl peptidase-4 (DPP-4) inhibitory effects of Melicope latifolia bark extracts and identification of bioactive constituents using in vitro and in silico approaches. Pharm. Biol. 2021, 59, 964–973. [Google Scholar] [CrossRef]
- Koziol, E.; Jóźwiak, K.; Budzyńska, B.; de Witte, P.A.M.; Copmans, D.; Skalicka-woźniak, K. Comparative antiseizure analysis of diverse natural coumarin derivatives in zebrafish. Int. J. Mol. Sci. 2021, 22, 11420. [Google Scholar] [CrossRef]
- Luszczki, J.J.; Andres-Mach, M.; Cisowski, W.; Mazol, I.; Glowniak, K.; Czuczwar, S.J. Osthole suppresses seizures in the mouse maximal electroshock seizure model. Eur. J. Pharmacol. 2009, 607, 107–109. [Google Scholar] [CrossRef]
- Luszczki, J.J.; Glowniak, K.; Czuczwar, S.J. Imperatorin enhances the protective activity of conventional antiepileptic drugs against maximal electroshock-induced seizures in mice. Eur. J. Pharmacol. 2007, 574, 133–139. [Google Scholar] [CrossRef]
- Zhu, H.-L.; Wan, J.-B.; Wang, Y.-T.; Li, B.-C.; Xiang, C.; He, J.; Li, P. Medicinal compounds with antiepileptic/anticonvulsant activities. Epilepsia 2014, 55, 3–16. [Google Scholar] [CrossRef]
- Kozioł, E.; Deniz, F.S.S.; Orhan, I.E.; Marcourt, L.; Budzyńska, B.; Wolfender, J.L.; Crawford, A.D.; Skalicka-Woźniak, K. High-performance counter-current chromatography isolation and initial neuroactivity characterization of furanocoumarin derivatives from Peucedanum alsaticum L (Apiaceae). Phytomedicine 2019, 54, 259–264. [Google Scholar] [CrossRef]
- Burrows, D.R.W.; Samarut Liu, J.; Baraban, S.C.; Richardson, M.P.; Meyer, M.P.; Rosch, R.E. Imaging epilepsy in larval zebrafish. Eur. J. Paediatr. Neurol. 2020, 24, 70–80. [Google Scholar] [CrossRef] [PubMed]
- Kumari, S.; Sharma, P.; Mazumder, A.G.; Rana, A.K.; Sharma, S.; Singh, D. Development and validation of chemical kindling in adult zebrafish: A simple and improved chronic model for screening of antiepileptic agents. J. Neurosci. Methods 2020, 346, 108916. [Google Scholar] [CrossRef] [PubMed]
- Jia, Y.; Yan, Q.; Zheng, Y.; Li, L.; Zhang, B.; Chang, Z.; Wang, Z.; Tang, H.; Qin, Y.; Guan, X.-Y. Long non-coding RNA NEAT1 mediated RPRD1B stability facilitates fatty acid metabolism and lymph node metastasis via c-Jun/c-Fos/SREBP1 axis in gastric cancer. J. Exp. Clin. Cancer Res. 2022, 41, 287. [Google Scholar] [CrossRef]
- Corbett, B.F.; You, J.C.; Zhang, X.; Pyfer, M.S.; Tosi, U.; Iascone, D.M.; Petrof, I.; Hazra, A.; Fu, C.-H.; Stephens, G.S.; et al. ΔFosB Regulates Gene Expression and Cognitive Dysfunction in a Mouse Model of Alzheimer’s Disease. Cell Rep. 2017, 20, 344–355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cacialli, P.; Gueguen, M.-M.; Coumailleau, P.; D’Angelo, L.; Kah, O.; Lucini, C.; Pellegrini, E. BDNF Expression in Larval and Adult Zebrafish Brain: Distribution and Cell Identification. PLoS ONE 2016, 11, e0158057. [Google Scholar] [CrossRef] [Green Version]
- De Felice, E.; Porreca, I.; Alleva, E.; De Girolamo, P.; Ambrosino, C.; Ciriaco, E.; Germanà, A.; Sordino, P. Localization of BDNF expression in the developing brain of zebrafish. J. Anat. 2014, 224, 564–574. [Google Scholar] [CrossRef] [PubMed]
- Jin, M.; Sheng, W.; Han, L.; He, Q.; Ji, X.; Liu, K. Activation of BDNF-TrkB signaling pathway-regulated brain inflammation in pentylenetetrazole-induced seizures in zebrafish. Fish Shellfish Immunol. 2018, 83, 26–36. [Google Scholar] [CrossRef]
- Iughetti, L.; Lucaccioni, L.; Fugetto, F.; Predieri, B.; Berardi, A.; Ferrari, F. Brain-derived neurotrophic factor and epilepsy: A systematic review. Neuropeptides 2018, 72, 23–29. [Google Scholar] [CrossRef]
- Hökfelt, T.; Xu, Z.Q.; Shi, T.J.; Holmberg, K.; Zhang, X. Galanin in ascending systems. Focus on coexistence with 5-hydroxytryptamine and noradrenaline. Ann. N. Y. Acad. Sci. 1998, 863, 252–263. [Google Scholar] [CrossRef]
- Podlasz, P.; Jakimiuk, A.; Chmielewska-Krzesinska, M.; Kasica, N.; Nowik, N.; Kaleczyc, J. Galanin regulates blood glucose level in the zebrafish: A morphological and functional study. Histochem. Cell Biol. 2016, 145, 105–117. [Google Scholar] [CrossRef] [Green Version]
- Podlasz, P.; Sallinen, V.; Chen, Y.-C.; Kudo, H.; Fedorowska, N.; Panula, P. Galanin gene expression and effects of its knock-down on the development of the nervous system in larval zebrafish. J. Comp. Neurol. 2012, 520, 3846–3862. [Google Scholar] [CrossRef] [PubMed]
- Heinemann, U.; Beck, H.; Dreier, J.P.; Ficker, E.; Stabel, J.; Zhang, C.L. The dentate gyrus as a regulated gate for the propagation of epileptiform activity. Epilepsy Res. 1992, 7, 273–280. [Google Scholar]
- Kokaia, M.; Holmberg, K.; Nanobashvili, A.; Xu, Z.-Q.D.; Kokaia, Z.; Lendahl, U.; Hilke, S.; Theodorsson, E.; Kahl, U.; Bartfai, T.; et al. Suppressed kindling epileptogenesis in mice with ectopic overexpression of galanin. Proc. Natl. Acad. Sci. USA 2001, 98, 14006–14011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazarati, A.M.; Hohmann, J.G.; Bacon, A.; Liu, H.; Sankar, R.; Steiner, R.A.; Wynick, D.; Wasterlain, C.G. Modulation of hippocampal excitability and seizures by galanin. J. Neurosci. 2000, 20, 6276–6281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zini, S.; Roisin, M.P.; Langel, U.; Bartfai, T.; Ben-Ari, Y. Galanin reduces release of endogenous excitatory amino acids in the rat hippocampus. Eur. J. Pharmacol. 1993, 245, 1–7. [Google Scholar] [CrossRef]
- Mazarati, A.M.; Halászi, E.; Telegdy, G. Anticonvulsive effects of galanin administered into the central nervous system upon the picrotoxin-kindled seizure syndrome in rats. Brain Res. 1992, 589, 164–166. [Google Scholar] [CrossRef]
- Lerner, J.T.; Sankar, R.; Mazarati, A.M. Galanin and Epilepsy. In Galanin; Hökfelt, T., Ed.; Springer: Basel, Switzerland, 2010; pp. 183–194. [Google Scholar]
- Podlasz, P.; Jakimiuk, A.; Kasica-Jarosz, N.; Czaja, K.; Wasowicz, K. Neuroanatomical Localization of Galanin in Zebrafish Telencephalon and Anticonvulsant Effect of Galanin Overexpression. ACS Chem. Neurosci. 2018, 9, 3049–3059. [Google Scholar] [CrossRef]
- Christiansen, S.H. Regulation of the galanin system in the brainstem and hypothalamus by electroconvulsive stimulation in mice. Neuropeptides 2011, 45, 337–341. [Google Scholar] [CrossRef]
- Gonzalez Nuñez, V.; Gonzalez Sarmiento, R.; Rodriguez, R.E. Characterization of zebrafish proenkephalin reveals novel opioid sequences. Mol. Brain Res. 2003, 114, 31–39. [Google Scholar] [CrossRef]
- König, M.; Zimmer, A.M.; Steiner, H.; Holmes, P.V.; Crawley, J.N.; Brownstein, M.J.; Zimmer, A. Pain responses, anxiety and aggression in mice deficient in pre-proenkephalin. Nature 1996, 383, 535–538. [Google Scholar] [CrossRef]
- Burtscher, J.; Bean, C.; Zangrandi, L.; Kmiec, I.; Agostinho, A.; Scorrano, L.; Gnaiger, E.; Schwarzer, C. Proenkephalin Derived Peptides Are Involved in the Modulation of Mitochondrial Respiratory Control During Epileptogenesis. Front. Mol. Neurosci. 2018, 11, 351. [Google Scholar] [CrossRef] [PubMed]
- Henry, M.S.; Bisht, K.; Vernoux, N.; Gendron, L.; Torres-Berrio, A.; Drolet, G.; Tremblay, M.-È. Delta Opioid Receptor Signaling Promotes Resilience to Stress Under the Repeated Social Defeat Paradigm in Mice. Front. Mol. Neurosci. 2018, 11, 100. [Google Scholar] [CrossRef] [PubMed]
- Piao, X.L.; Park, I.H.; Baek, S.H.; Kim, H.Y.; Park, M.K.; Park, J.H. Antioxidative activity of furanocoumarins isolated from Angelicae dahuricae. J. Ethnopharmacol. 2004, 93, 243–246. [Google Scholar] [CrossRef]
- Fetcho, J.R.; Higashijima, S.-i.; McLean, D.L. Zebrafish and motor control over the last decade. Brain Res. Rev. 2008. [CrossRef] [Green Version]
- Pietri, T.; Manalo, E.; Ryan, J.; Saint-Amant, L.; Washbourne, P. Glutamate drives the touch response through a rostral loop in the spinal cord of zebrafish embryos. Dev. Neurobiol. 2009, 69, 780–795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, V.P. In Situ Detection of MicroRNA Expression with RNAscope Probes. Methods Mol. Biol. 2018, 1649, 197–208. [Google Scholar] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | HEMWat | KD-Halfordin |
|---|---|---|
| 1 | 1/1/1/1 | 2.54 |
| 2 | 6/5/6/5 | 1.20 |
| 3 | 3/2/3/2 | 0.69 |
| 4 | 2/1/2/1 | 0.36 |
| 5 | 5/2/5/2 | 0.24 |
| 6 | 3/1/3/1 | 0.20 |
| 7 | 4/1/4/1 | 0.14 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Skiba, A.; Kozioł, E.; Luca, S.V.; Budzyńska, B.; Podlasz, P.; Van Der Ent, W.; Shojaeinia, E.; Esguerra, C.V.; Nour, M.; Marcourt, L.; et al. Evaluation of the Antiseizure Activity of Endemic Plant Halfordia kendack Guillaumin and Its Main Constituent, Halfordin, on a Zebrafish Pentylenetetrazole (PTZ)-Induced Seizure Model. Int. J. Mol. Sci. 2023, 24, 2598. https://doi.org/10.3390/ijms24032598
Skiba A, Kozioł E, Luca SV, Budzyńska B, Podlasz P, Van Der Ent W, Shojaeinia E, Esguerra CV, Nour M, Marcourt L, et al. Evaluation of the Antiseizure Activity of Endemic Plant Halfordia kendack Guillaumin and Its Main Constituent, Halfordin, on a Zebrafish Pentylenetetrazole (PTZ)-Induced Seizure Model. International Journal of Molecular Sciences. 2023; 24(3):2598. https://doi.org/10.3390/ijms24032598
Chicago/Turabian StyleSkiba, Adrianna, Ewelina Kozioł, Simon Vlad Luca, Barbara Budzyńska, Piotr Podlasz, Wietske Van Der Ent, Elham Shojaeinia, Camila V. Esguerra, Mohammed Nour, Laurence Marcourt, and et al. 2023. "Evaluation of the Antiseizure Activity of Endemic Plant Halfordia kendack Guillaumin and Its Main Constituent, Halfordin, on a Zebrafish Pentylenetetrazole (PTZ)-Induced Seizure Model" International Journal of Molecular Sciences 24, no. 3: 2598. https://doi.org/10.3390/ijms24032598