
Virulence Factors of Candida spp. and Host Immune Response Important in the Pathogenesis of Vulvovaginal Candidiasis



Abstract
:1. Introduction
2. Overview of Candida spp.
2.1. Risk Factors Associated with Superficial and Invasive Candida Infections
2.2. Most Commonly Isolated Species
2.3. Infections Caused by Fungi of the Genus Candida
3. Vulvovaginal Candidiasis
4. Pathogenicity of Fungi of the Genus Candida spp. Related to Vaginal Infections—Virulence Factors
4.1. Adhesion
4.2. Hydrolytic Enzymes
4.3. Adaptability to Changing Environmental Conditions, Dimorphism and Phenotypic Switching
4.4. Biofilm
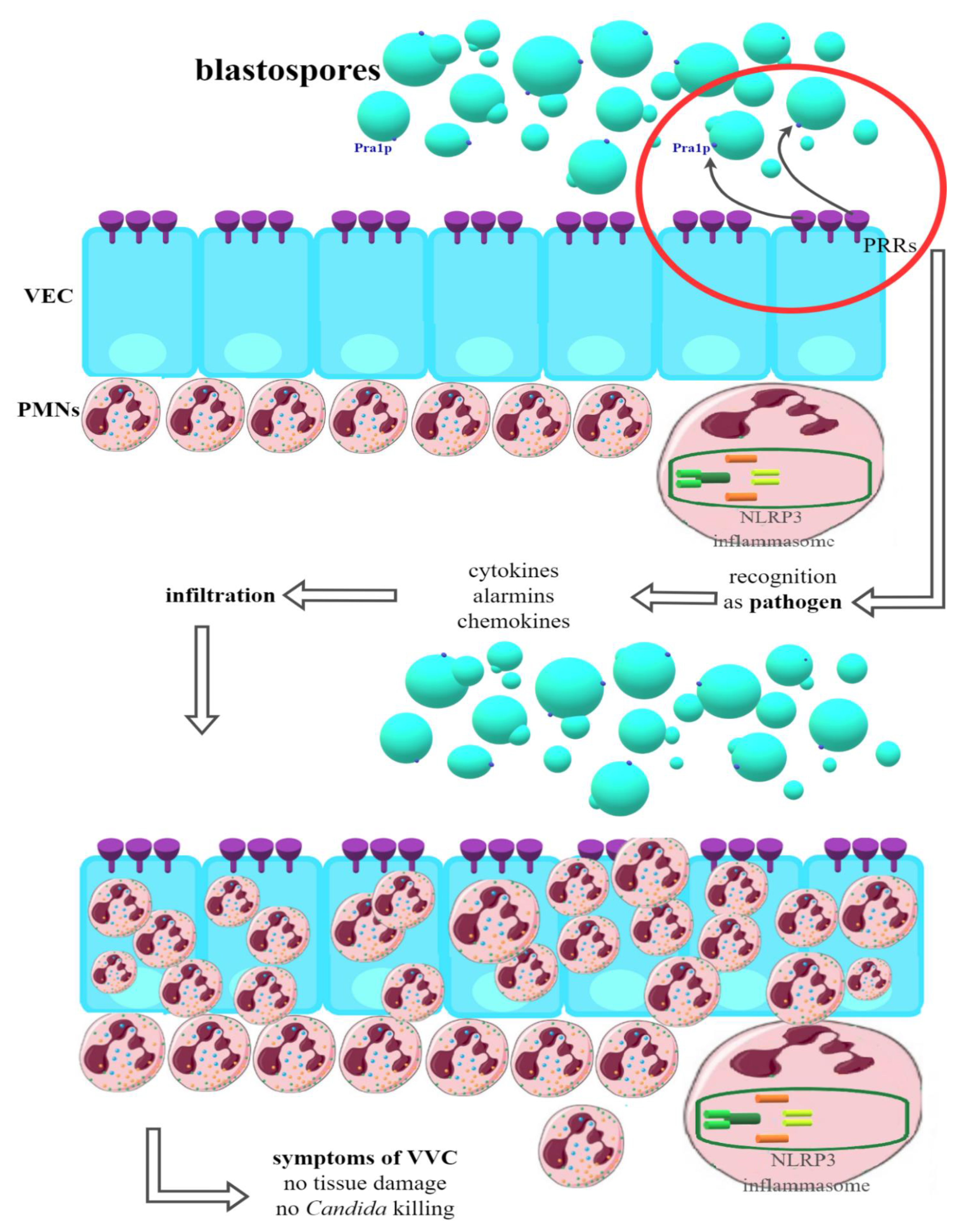
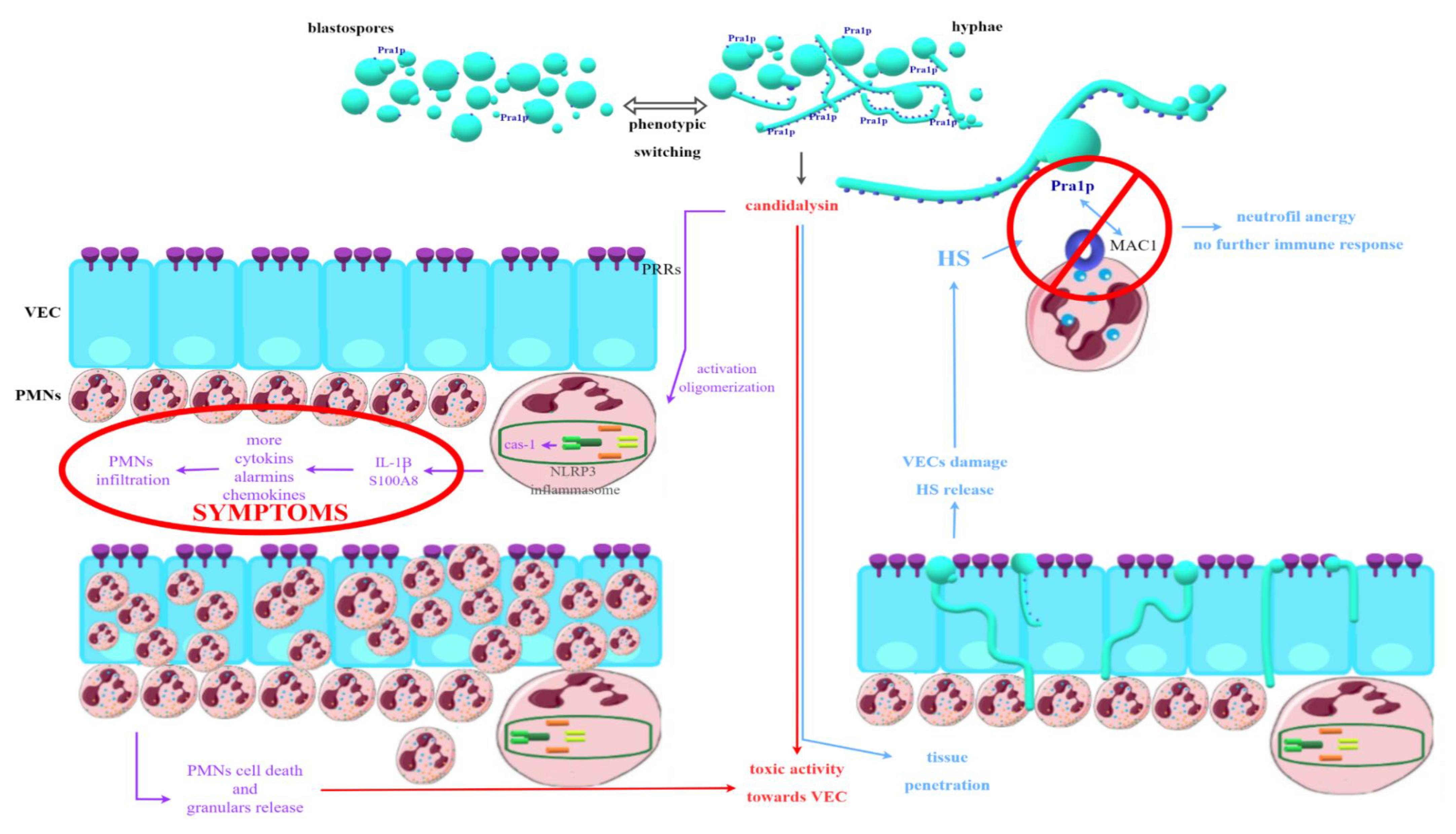
5. Pathogenicity of Fungi of the Genus Candida spp. Associated with Vaginal Infections—Host Immune Response
6. Summary
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yang, B.; Rao, R. Emerging Pathogens of the Candida Species. Intech Open Sci. 2012, 1, 3–18. [Google Scholar] [CrossRef] [Green Version]
- Singh, D.K.; Tóth, R.; Gácser, A. Mechanisms of Pathogenic Candida Species to Evade the Host Complement Attack. Front. Cell. Infect. Microbiol. 2020, 10, 94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodrigues, C.F.; Rodrigues, M.E.; Henriques, M. Candida sp. Infections in patients with diabetes mellitus. J. Clin. Med. 2019, 8, 76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, S.; Fatima, Z.; Hameed, S. Predisposing factors endorsing Candida infections. Infez. Med. 2015, 23, 211–223. [Google Scholar] [PubMed]
- Wróblewska, M.; Sulik-Tyszka, B. Candida auris—Epidemiology and laboratory diagnostics of infections. Diagnostyka Lab. 2017, 53, 235–240. [Google Scholar] [CrossRef]
- Alberto Cortés, J.; Fernanda Corrales, I. Invasive Candidiasis: Epidemiology and Risk Factors. In Fungal Infection; IntechOpen: London, UK, 2018; pp. 33–46. [Google Scholar]
- Vila, T.; Sultan, A.S.; Montelongo-Jauregui, D.; Jabra-Rizk, M.A. Oral candidiasis: A disease of opportunity. J. Fungi 2020, 6, 15. [Google Scholar] [CrossRef] [Green Version]
- Sikora, M. Czynniki wirulencji grzybów z rodzaju candida istotne w patogenezie zakażeń występujących u pacjentów żywionych pozajelitowo. Postep. Mikrobiol. 2015, 3, 224–234. [Google Scholar]
- Arsenault, A.B.; Bliss, J.M. Neonatal Candidiasis: New Insights into an Old Problem at a Unique Host-Pathogen Interface. Curr. Fungal Infect. Rep. 2015, 9, 246–252. [Google Scholar] [CrossRef] [Green Version]
- Flevari, A.; Theodorakopoulou, M.; Velegraki, A.; Armaganidis, A.; Dimopoulos, G. Treatment of invasive candidiasis in the elderly: A review. Clin. Interv. Aging 2013, 8, 1199–1208. [Google Scholar] [CrossRef] [Green Version]
- Dekkers, B.G.J.; Veringa, A.; Marriott, D.J.E.; Boonstra, J.M.; van der Elst, K.C.M.; Doukas, F.F.; McLachlan, A.J.; Alffenaar, J.W.C. Invasive Candidiasis in the Elderly: Considerations for Drug Therapy. Drugs Aging 2018, 35, 781–789. [Google Scholar] [CrossRef] [Green Version]
- Mayer, F.L.; Wilson, D.; Hube, B. Candida albicans pathogenicity mechanisms. Virulence 2013, 4, 119–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopes, J.P.; Lionakis, M.S. Pathogenesis and virulence of Candida albicans. Virulence 2022, 13, 89–121. [Google Scholar] [CrossRef] [PubMed]
- Taei, M.; Chadeganipour, M.; Mohammadi, R. An alarming rise of non-albicans Candida species and uncommon yeasts in the clinical samples; a combination of various molecular techniques for identification of etiologic agents. BMC Res. Notes 2019, 12, 779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aydemir, Ö.; Demiray, T.; Köroğlu, M.; Aydemir, Y.; Altındiş, M. Emerge of non-albicans Candida species; evaluation of Candida species and antifungal susceptibilities according to years. Biomed. Res. 2017, 28, 2820–2825. [Google Scholar]
- Lindberg, E.; Hammarström, H.; Ataollahy, N.; Kondori, N. Species distribution and antifungal drug susceptibilities of yeasts isolated from the blood samples of patients with candidemia. Sci. Rep. 2019, 9, 12–17. [Google Scholar] [CrossRef] [Green Version]
- Das, K.H.; Mangayarkarasi, V.; Sen, M. Antifungal resistant in non-albicans candida species are emerging as a threat to antenatal women with vulvovaginal candidiasis. Biomed. Pharmacol. J. 2019, 12, 1369–1378. [Google Scholar] [CrossRef]
- Liu, F.; Zhong, L.; Zhou, F.; Zheng, C.; Zhang, K.; Cai, J.; Zhou, H.; Tang, K.; Dong, Z.; Cui, W.; et al. Clinical features, strain distribution, antifungal resistance and prognosis of patients with non-albicans candidemia: A retrospective observational study. Infect. Drug Resist. 2021, 14, 3233–3246. [Google Scholar] [CrossRef]
- Ahmad, S.; Alfouzan, W. Candida auris: Epidemiology, diagnosis, pathogenesis, antifungal susceptibility, and infection control measures to combat the spread of infections in healthcare facilities. Microorganisms 2021, 9, 807. [Google Scholar] [CrossRef]
- Grondalska, D.; Kmieciak, W. Candida auris—Nowy patogen grzybiczy. Post. Mikrobiol 2019, 191, E865. [Google Scholar]
- Garcia-Bustos, V.; Ruiz-Saurí, A.; Ruiz-Gaitán, A.; Sigona-Giangreco, I.A.; Cabañero-Navalon, M.D.; Sabalza-Baztán, O.; Salavert-Lletí, M.; Tormo, M.Á.; Pemán, J. Characterization of the Differential Pathogenicity of Candida auris in a Galleria mellonella Infection Model. Microbiol. Spectr. 2021, 9, e0001321. [Google Scholar] [CrossRef]
- Keighley, C.; Garnham, K.; Harch, S.A.J.; Robertson, M.; Chaw, K.; Teng, J.C.; Chen, S.C.A. Candida auris: Diagnostic Challenges and Emerging Opportunities for the Clinical Microbiology Laboratory. Curr. Fungal Infect. Rep. 2021, 15, 116–126. [Google Scholar] [CrossRef] [PubMed]
- Talapko, J.; Juzbašić, M.; Matijević, T.; Pustijanac, E.; Bekić, S.; Kotris, I.; Škrlec, I. Candida albicans-the virulence factors and clinical manifestations of infection. J. Fungi 2021, 7, 79. [Google Scholar] [CrossRef] [PubMed]
- Staniszewska, M.; Bondaryk, M.; Kowalska, M.; Magda, U.; Łuka, M.; Ochal, Z.; Kurzątkowski, W. Pathogenesis and treatment of fungal infections by Candida spp. [Patogeneza i leczenie zakażeń Candida spp.]. Postep. Mikrobiol. 2014, 53, 229–240. [Google Scholar]
- Khan, F.; Bamunuarachchi, N.I.; Pham, D.T.N.; Tabassum, N.; Khan, M.S.A.; Kim, Y.M. Mixed biofilms of pathogenic Candida-bacteria: Regulation mechanisms and treatment strategies. Crit. Rev. Microbiol. 2021, 45, 699–727. [Google Scholar] [CrossRef] [PubMed]
- Ramage, G.; Martínez, J.P.; López-Ribot, J.L. Candida biofilms on implanted biomaterials: A clinically significant problem. FEMS Yeast Res. 2006, 6, 979–986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vera-González, N.; Shukla, A. Advances in Biomaterials for the Prevention and Disruption of Candida Biofilms. Front. Microbiol. 2020, 11, 2251. [Google Scholar] [CrossRef]
- Peng, Z.; Tang, J. Intestinal Infection of Candida albicans: Preventing the Formation of Biofilm by C. albicans and Protecting the Intestinal Epithelial Barrier. Front. Microbiol. 2022, 12, 783010. [Google Scholar] [CrossRef]
- Zeng, X.; Zhang, Y.; Zhang, T.; Xue, Y.; Xu, H.; An, R. Risk Factors of Vulvovaginal Candidiasis among Women of Reproductive Age in Xi’an: A Cross-Sectional Study. Biomed Res. Int. 2018, 2018, 9703754. [Google Scholar] [CrossRef] [Green Version]
- Achkar, J.M.; Fries, B.C. Candida infections of the genitourinary tract. Clin. Microbiol. Rev. 2010, 23, 253–273. [Google Scholar] [CrossRef] [Green Version]
- Aniebue, U.U.; Nwankwo, T.O.; Nwafor, M.I. Vulvovaginal candidiasis in reproductive age women in Enugu Nigeria, clinical versus laboratory-assisted diagnosis. Niger. J. Clin. Pract. 2018, 21, 1017–1022. [Google Scholar] [CrossRef]
- Auriemma, R.S.; Scairati, R.; del Vecchio, G.; Liccardi, A.; Verde, N.; Pirchio, R.; Pivonello, R.; Ercolini, D.; Colao, A. The Vaginal Microbiome: A Long Urogenital Colonization Throughout Woman Life. Front. Cell. Infect. Microbiol. 2021, 11, 613. [Google Scholar] [CrossRef] [PubMed]
- Lema, V.M. Recurrent Vulvo-Vaginal Candidiasis: Diagnostic and Management Challenges in a Developing Country Context. Obstet. Gynecol. Int. J. 2017, 7, 00260. [Google Scholar] [CrossRef]
- Hoffmann, J.N.; You, H.M.; Hedberg, E.C.; Jordan, J.A.; McClintock, M.K. Prevalence of bacterial vaginosis and Candida among postmenopausal women in the United States. J. Gerontol. Ser. B Psychol. Sci. Soc. Sci. 2014, 69, S205–S214. [Google Scholar] [CrossRef] [PubMed]
- McClelland, R.S.; Richardson, B.A.; Hassan, W.M.; Graham, S.M.; Kiarie, J.; Baeten, J.M.; Mandaliya, K.; Jaoko, W.; Ndinya-Achola, J.O.; Holmes, K.K. Prospective study of vaginal bacteria flora and other risk factors for vulvovaginal candidiasis. J. Infect. Dis. 2009, 199, 1883–1890. [Google Scholar] [CrossRef] [Green Version]
- Ceccarani, C.; Foschi, C.; Parolin, C.; D’Antuono, A.; Gaspari, V.; Consolandi, C.; Laghi, L.; Camboni, T.; Vitali, B.; Severgnini, M.; et al. Diversity of vaginal microbiome and metabolome during genital infections. Sci. Rep. 2019, 9, 14095. [Google Scholar] [CrossRef] [Green Version]
- Konadu, D.G.; Owusu-Ofori, A.; Yidana, Z.; Boadu, F.; Iddrisu, L.F.; Adu-Gyasi, D.; Dosoo, D.; Awuley, R.L.; Owusu-Agyei, S.; Asante, K.P. Prevalence of vulvovaginal candidiasis, bacterial vaginosis and trichomoniasis in pregnant women attending antenatal clinic in the middle belt of Ghana. BMC Pregnancy Childbirth 2019, 19, 1–10. [Google Scholar] [CrossRef]
- Ang, X.Y.; Mageswaran, U.M.; Chung, Y.L.F.; Lee, B.K.; Azhar, S.N.A.; Roslan, N.S.; Saufian, I.F.B.; Mustaffa, N.S.; Kalam, E.M.; Ibrahim, A.F.; et al. Probiotics Reduce Vaginal Candidiasis in Pregnant Women via Modulating Abundance of Candida and Lactobacillus in Vaginal and Cervicovaginal Regions. Microorganisms 2022, 10, 285. [Google Scholar] [CrossRef]
- Żyrek, D.; Wajda, A.; Czechowicz, P.; Nowicka, J.; Jaśkiewicz, M.; Neubauer, D.; Kamysz, W. The antimicrobial activity of omiganan alone and in combination against candida isolated from vulvovaginal candidiasis and bloodstream infections. Antibiotics 2021, 10, 1001. [Google Scholar] [CrossRef]
- Swidsinski, A.; Guschin, A.; Tang, Q.; Dörffel, Y.; Verstraelen, H.; Tertychnyy, A.; Khayrullina, G.; Luo, X.; Sobel, J.D.; Jiang, X. Vulvovaginal candidiasis: Histologic lesions are primarily polymicrobial and invasive and do not contain biofilms. Am. J. Obstet. Gynecol. 2019, 220, e1–e91. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Cerdeira, C.; Gregorio, M.C.; Molares-Vila, A.; López-Barcenas, A.; Fabbrocini, G.; Bardhi, B.; Sinani, A.; Sánchez-Blanco, E.; Arenas-Guzmán, R.; Hernandez-Castro, R. Biofilms and vulvovaginal candidiasis. Colloids Surf. B Biointerfaces 2019, 174, 110–125. [Google Scholar] [CrossRef]
- Silva, S.; Negri, M.; Henriques, M.; Oliveira, R.; Williams, D.W.; Azeredo, J. Adherence and biofilm formation of non-Candida albicans Candida species. Trends Microbiol. 2011, 19, 241–247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peters, B.M.; Yano, J.; Noverr, M.C.; Fidel, P.L. Candida Vaginitis: When Opportunism Knocks, the Host Responds. PLoS Pathog. 2014, 10, e1003965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nobile, C.J.; Johnson, A.D. Candida albicans Biofilms and Human Disease. Annu. Rev. Microbiol. 2015, 69, 71–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hacioglu, M.; Guzel, C.B.; Savage, P.B.; Seher Birteksoz Tan, A. Antifungal susceptibilities, in vitro production of virulence factors and activities of ceragenins against Candida spp. Isolated from vulvovaginal candidiasis. Med. Mycol. 2019, 57, 291–299. [Google Scholar] [CrossRef] [PubMed]
- Tulasidas, S.; Rao, P.; Bhat, S.; Manipura, R. A study on biofilm production and antifungal drug resistance among Candida species from vulvovaginal and bloodstream infections. Infect. Drug Resist. 2018, 11, 2443–2448. [Google Scholar] [CrossRef] [Green Version]
- Gonçalves, B.; Ferreira, C.; Alves, C.T.; Henriques, M.; Azeredo, J.; Silva, S. Vulvovaginal candidiasis: Epidemiology, microbiology and risk factors. Crit. Rev. Microbiol. 2016, 42, 905–927. [Google Scholar] [CrossRef] [Green Version]
- Zhu, W.; Filler, S.G. Interactions of Candida albicans with epithelial cells. Cell. Microbiol. 2010, 12, 273–282. [Google Scholar] [CrossRef] [Green Version]
- Paiva, L.C.F.; Vidigal, P.G.; Donatti, L.; Svidzinski, T.I.E.; Consolaro, M.E.L. Assessment of in vitro biofilm formation by Candida species isolates from vulvovaginal candidiasis and ultrastructural characteristics. Micron 2012, 43, 497–502. [Google Scholar] [CrossRef]
- Silva, S.; Negri, M.; Henriques, M.; Oliveira, R.; Williams, D.W.; Azeredo, J. Candida glabrata, Candida parapsilosis and Candida tropicalis: Biology, epidemiology, pathogenicity and antifungal resistance. FEMS Microbiol. Rev. 2012, 36, 288–305. [Google Scholar] [CrossRef] [Green Version]
- Ielasi, F.S.; Decanniere, K.; Willaert, R.G. The epithelial adhesin 1 (Epa1p) from the human-pathogenic yeast Candida glabrata: Structural and functional study of the carbohydrate-binding domain. Acta Crystallogr. Sect. D Biol. Crystallogr. 2012, 68, 210–217. [Google Scholar] [CrossRef]
- Staniszewska, M.; Bondaryk, M.; Siennicka, K.; Piłat, J.; Schaller, M.; Kurza̧tkowski, W. Role of aspartic proteinases in Candida albicans virulence. Part I. Substrate specificity of Aspartic proteinases and Candida albicans pathogenesis (Postepy Mikrobiologii (2012) (131)). Postep. Mikrobiol. 2012, 51, 236. [Google Scholar]
- Vylkova, S.; Lorenz, M.C. Encounters with Mammalian Cells: Survival Strategies of Candida Species. In Candida and Candidiasis, 2nd. ed.; Calderone, R.A., Clancy, C.J., Eds.; American Society of Microbiology: Washington, DC, USA, 2012; pp. 261–282. [Google Scholar]
- Sikora, M.; Dabkowska, M.; Swoboda-Kopec, E.; Jarzynka, S.; Netsvyetayeva, I.; Jaworska-Zaremba, M.; Pertkiewicz, M.; Mlynarczyk, G. Differences in proteolytic activity and gene profiles of fungal strains isolated from the total parenteral nutrition patients. Folia Microbiol. 2011, 56, 143–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tellapragada, C.; Eshwara, V.K.; Johar, R.; Shaw, T.; Malik, N.; Bhat, P.V.; Kamath, A.; Mukhopadhyay, C. Antifungal Susceptibility Patterns, In Vitro Production of Virulence Factors, and Evaluation of Diagnostic Modalities for the Speciation of Pathogenic Candida from Blood Stream Infections and Vulvovaginal Candidiasis. J. Pathog. 2014, 2014, 142864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sobel, J.D. Vulvovaginal candidosis. Lancet 2007, 369, 1961–1971. [Google Scholar] [CrossRef]
- Jafari, M.; Salari, S.; Pakshir, K.; Zomorodian, K. Exoenzyme activity and possibility identification of Candida dubliniensis among Candida albicans species isolated from vaginal candidiasis. Microb. Pathog. 2017, 110, 73–77. [Google Scholar] [CrossRef]
- Naglik, J.R.; Rodgers, C.A.; Shirlaw, P.J.; Dobbie, J.L.; Fernandes-Naglik, L.L.; Greenspan, D.; Agabian, N.; Challacombe, S.J. Differential Expression of Candida albicans Secreted Aspartyl Proteinase and Phospholipase B Genes in Humans Correlates with Active Oral and Vaginal Infections. J. Infect. Dis. 2003, 188, 469–479. [Google Scholar] [CrossRef] [Green Version]
- Calderone, R.A.; Fonzi, W. A Virulence factors in Candida albicans. TRENDS Microbiol. 2001, 9, 327–335. [Google Scholar] [CrossRef]
- Cassone, A. Vulvovaginal Candida albicans infections: Pathogenesis, immunity and vaccine prospects. BJOG Int. J. Obstet. Gynaecol. 2015, 122, 785–794. [Google Scholar] [CrossRef]
- Sudbery, P.; Gow, N.; Berman, J. The distinct morphogenic states of Candida albicans. Trends Microbiol. 2004, 12, 317–324. [Google Scholar] [CrossRef]
- Moran, G.; Coleman, D.; Sullivan, D. An Introduction to the Medically Important Candida Species. In Candida and Candidiasis, 2nd ed.; Calderone, R.A., Clancy, C.J., Eds.; American Society of Microbiology: Washington, DC, USA, 2012; pp. 11–25. [Google Scholar]
- Consolaro, M.E.L.; Albertoni, T.A.; Svidzinski, A.E.; Peralta, R.M.; Svidzinski, T.I.E. Vulvovaginal candidiasis is associated with the production of germ tubes by Candida albicans. Mycopathologia 2005, 159, 501–507. [Google Scholar] [CrossRef]
- Sudbery, P.E. Growth of Candida albicans hyphae. Nat. Rev. Microbiol. 2011, 9, 737–748. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Yu, F.; Huang, L.; Hu, Z. The changes of antifungal susceptibilities caused by the phenotypic switching of Candida species in 229 patients with vulvovaginal candidiasis. J. Clin. Lab. Anal. 2019, 33, e22644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lastauskienė, E.; Čeputytė, J.; Girkontaitė, I.; Zinkevičienė, A. Phenotypic Switching of Candida guilliermondii is Associated with Pseudohyphae Formation and Antifungal Resistance. Mycopathologia 2015, 179, 205–211. [Google Scholar] [CrossRef] [PubMed]
- Harriott, M.M.; Noverr, M.C. Importance of Candida-bacterial polymicrobial biofilms in disease. Trends Microbiol. 2011, 19, 557–563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, S.; Henriques, M.; Martins, A.; Oliveira, R.; Williams, D.; Azeredo, J. Biofilms of non-Candida albicans Candida species: Quantification, structure and matrix composition. Med. Mycol. 2009, 47, 681–689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harriott, M.M.; Lilly, E.A.; Rodriguez, T.E.; Fidel, P.L.; Noverr, M.C. Candida albicans forms biofilms on the vaginal mucosa. Microbiology 2010, 156, 3635–3644. [Google Scholar] [CrossRef] [Green Version]
- Muzny, C.A.; Schwebke, J.R. Biofilms: An underappreciated mechanism of treatment failure and recurrence in vaginal infections. Clin. Infect. Dis. 2015, 61, 601–606. [Google Scholar] [CrossRef] [Green Version]
- Desai, J.V.; Mitchell, A.P. Candida albicans biofilm development and its genetic controls. Microbiol. Spectr. 2015, 3, 139–148. [Google Scholar] [CrossRef] [Green Version]
- McKloud, E.; Delaney, C.; Sherry, L.; Kean, R.; Williams, S.; Metcalfe, R.; Thomas, R.; Richardson, R.; Gerasimidis, K.; Nile, C.J.; et al. Recurrent Vulvovaginal Candidiasis: A Dynamic Interkingdom Biofilm Disease of Candida and Lactobacillus. mSystems 2021, 6, e00622-21. [Google Scholar] [CrossRef]
- Thein, Z.M.; Seneviratne, C.J.; Samaranayake, Y.H.; Samaranayake, L.P. Community lifestyle of Candida in mixed biofilms: A mini review. Mycoses 2009, 52, 467–475. [Google Scholar] [CrossRef]
- Rodrigues, M.E.; Gomes, F.; Rodrigues, C.F. Candida spp./bacteria mixed biofilms. J. Fungi 2020, 6, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lohse, M.B.; Gulati, M.; Johnson, A.D.; Nobile, C.J. Development and regulation of single-and multi-species Candida albicans biofilms. Nat. Rev. Microbiol. 2018, 16, 19–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomaszewski, J. The statement of polish gynecological society experts on the etiology and treatment of recurrent vulvovaginal candidiasis. Ginekol. Pol. 2011, 82, 869–873. [Google Scholar]
- Sherry, L.; Kean, R.; Mckloud, E.; Donnell, L.E.O.; Metcalfe, R.; Jones, B.L. Biofilms formed by isolates from patients are heterogeneous and insensitive to fluconazole. Antimicrob Agents Chemother 2017, 61, e01065-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Auler, M.E.; Morreira, D.; Rodrigues, F.F.O.; Abrão, M.S.; Margarido, P.F.R.; Matsumoto, F.E.; Silva, E.G.; Silva, B.C.M.; Schneider, R.P.; Paula, C.R. Biofilm formation on intrauterine devices in patients with recurrent vulvovaginal candidiasis Biofilm formation on IUDs in patients with vulvovaginal candidiasis. Med. Mycol. 2010, 48, 211–216. [Google Scholar] [CrossRef] [Green Version]
- Sobel, J.D. Editorial Commentary: Vaginal Biofilm: Much Ado about Nothing, or a New Therapeutic Challenge? Clin. Infect. Dis. 2015, 61, 607–608. [Google Scholar] [CrossRef] [Green Version]
- Yano, J.; Peters, B.M.; Noverr, M.C.; Fidel, P.L. Novel Mechanism behind the Immunopathogenesis of Vulvovaginal Candidiasis: “Neutrophil Anergy. ” Infect. Immun. 2018, 86, e00684-17. [Google Scholar] [CrossRef] [Green Version]
- Willems, H.M.E.; Ahmed, S.S.; Liu, J.; Xu, Z.; Peters, B.M. Vulvovaginal candidiasis: A current understanding and burning questions. J. Fungi 2020, 6, 27. [Google Scholar] [CrossRef] [Green Version]
- Fidel, P.L. History and update on host defense against vaginal candidiasis. Am. J. Reprod. Immunol. 2007, 57, 2–12. [Google Scholar] [CrossRef]
- Roselletti, E.; Perito, S.; Sabbatini, S.; Monari, C.; Vecchiarelli, A. Vaginal Epithelial Cells discriminate between yeast and hyphae of C. albicans in women who are colonized with or have vaginal candidiasis. J. Infect. Dis. 2018, 10, 1645–1654. [Google Scholar] [CrossRef]
- Richardson, J.P.; Willems, H.M.E.; Moyes, D.L.; Shoaie, S.; Barker, K.S.; Tan, L.; Palmer, G.E.; Hube, B.; Naglik, J.R.; Peters, M. Candidalysin Drives Epithelial Signaling, Neutrophil Recruitment, and Immunopathology at the Vaginal Mucosa. Infect. Immun. 2018, 86, e00645-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roselletti, E.; Perito, S.; Gabrielli, E.; Mencacci, A.; Pericolini, E.; Sabbatini, S.; Cassone, A.; Vecchiarelli, A. NLRP3 inflammasome is a key player in human vulvovaginal disease caused by Candida albicans. Sci. Rep. 2017, 7, 17877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bruno, V.M.; Shetty, A.C.; Yano, J.; Fidel, P.L.; Noverr, M.C.; Peters, B.M. Transcriptomic Analysis of Vulvovaginal Candidiasis Identifies a Role for the NLRP3 Inflammasome. mBio 2015, 6, e00182-15. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Factor | Genes | Candida Strains | Function in Biofilm Formation during VVC | References |
|---|---|---|---|---|
| Als proteins | family of ALS | C. albicans C. parapsilosis C. tropicalis C. dubliniensis C. guillermondii C. lusitaniae | adhesion to VECs | [8,42,47,48] |
| Epa protein | EPA | C. albicans C. glabrata | adhesion to VECs | [8,42,51] |
| Hwp1 protein | HWP1 | Candida spp. | adhesion to VECs | [8,41,44,48] |
| Sap enzymes | family of SAP | C. albicans C. parapsilosis C. tropicalis C. dubliniensis | adhesion to VECs, phenotypic switching, formation of hyphae and its adhesion and penetration into VECs | [24,42,47,50,54] |
| yapsins | YPS | C. glabrata | adhesion to VECs, phenotypic switching, formation of hyphae and its adhesion and penetration into VEC | [8,53,54] |
| lipases | LIP | C. albicans C. parapsilosis C. tropicalis | increasing adhesion to VECs | [42,50,53,59] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Czechowicz, P.; Nowicka, J.; Gościniak, G. Virulence Factors of Candida spp. and Host Immune Response Important in the Pathogenesis of Vulvovaginal Candidiasis. Int. J. Mol. Sci. 2022, 23, 5895. https://doi.org/10.3390/ijms23115895
Czechowicz P, Nowicka J, Gościniak G. Virulence Factors of Candida spp. and Host Immune Response Important in the Pathogenesis of Vulvovaginal Candidiasis. International Journal of Molecular Sciences. 2022; 23(11):5895. https://doi.org/10.3390/ijms23115895
Chicago/Turabian StyleCzechowicz, Paulina, Joanna Nowicka, and Grażyna Gościniak. 2022. "Virulence Factors of Candida spp. and Host Immune Response Important in the Pathogenesis of Vulvovaginal Candidiasis" International Journal of Molecular Sciences 23, no. 11: 5895. https://doi.org/10.3390/ijms23115895