
Anti-Hepatocellular Carcinoma Biomolecules: Molecular Targets Insights

 , ,
, ,
Abstract
:1. Introduction
2. HCC Targeted Therapy
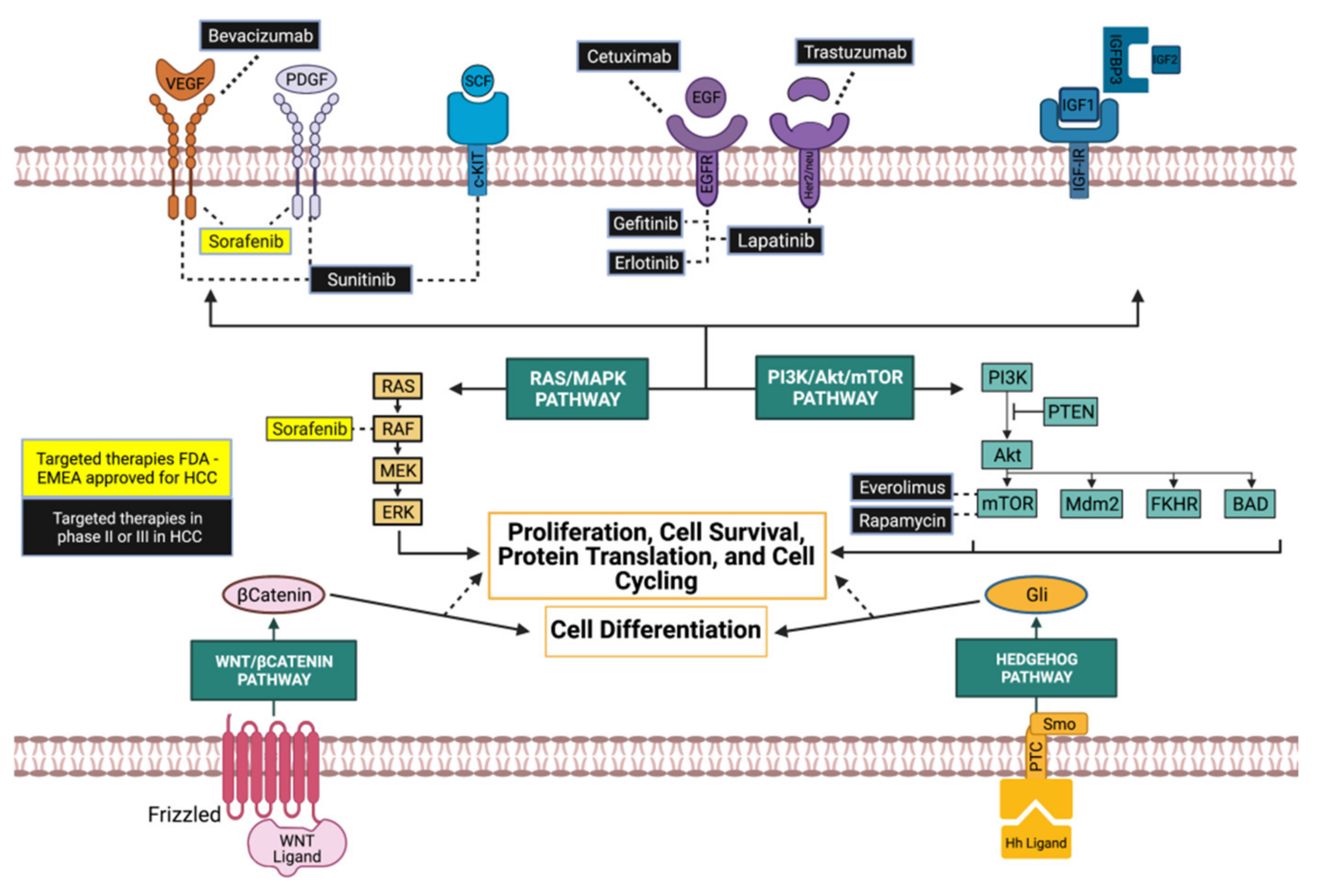
2.1. Tyrosine Kinase Inhibitors
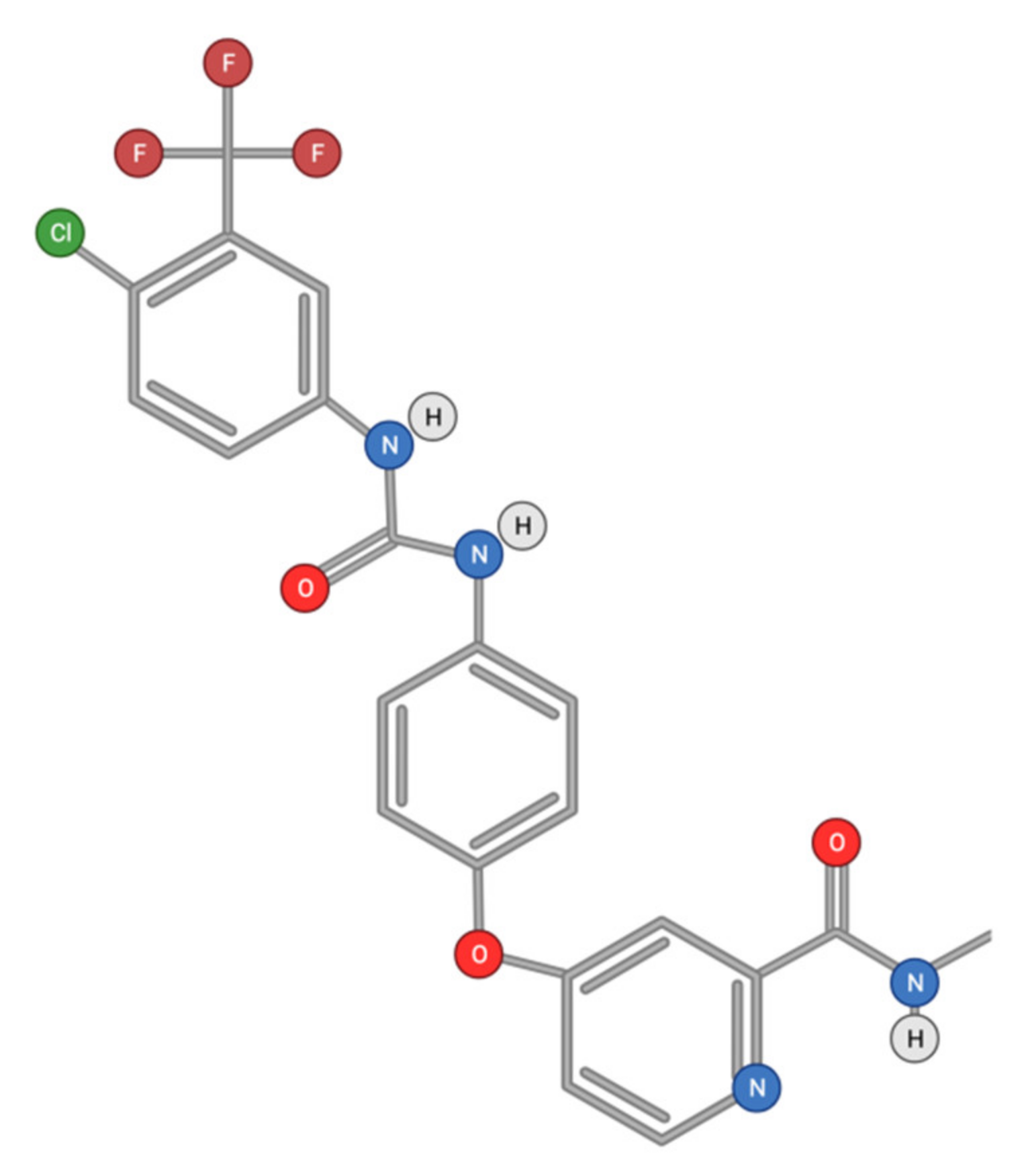
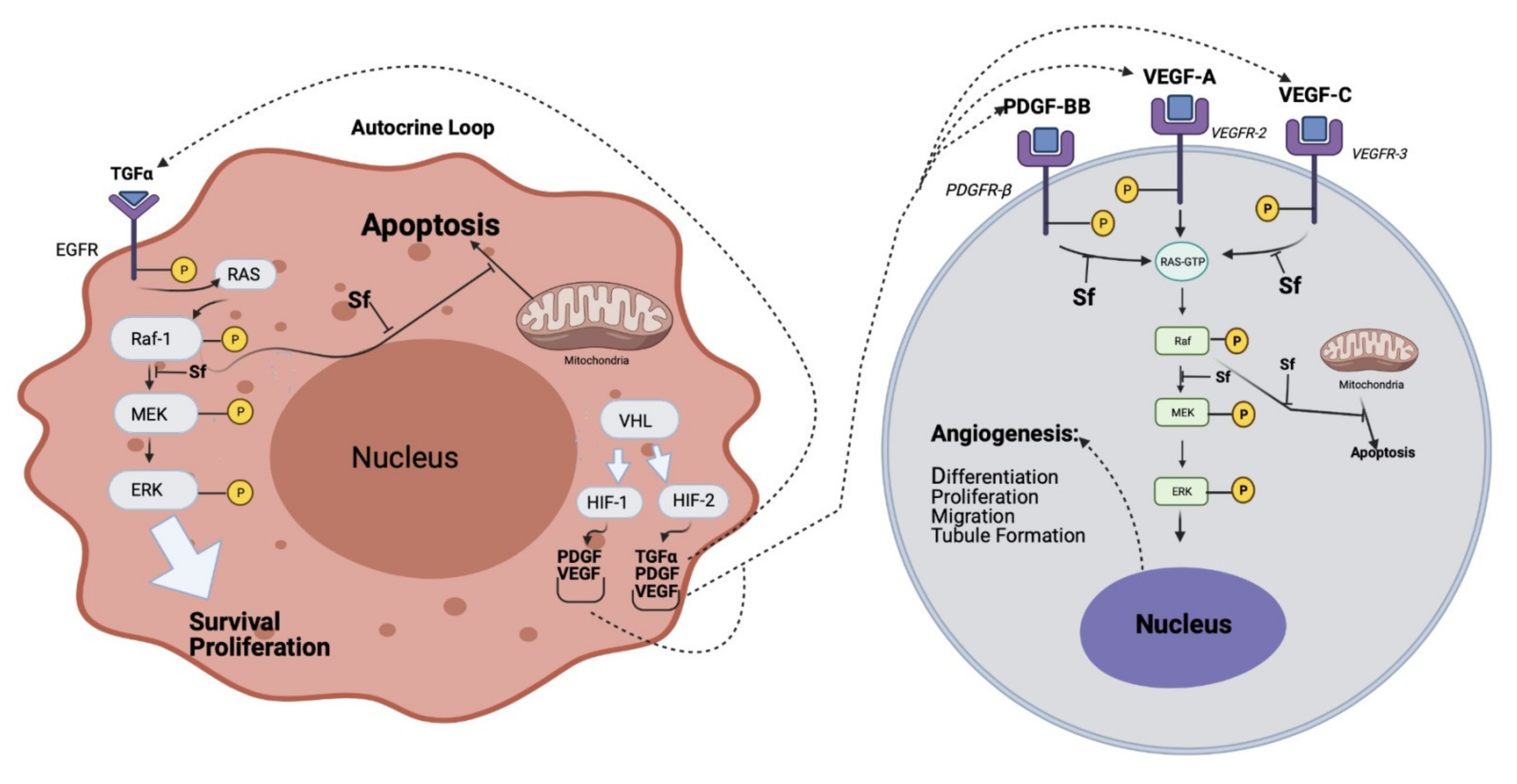
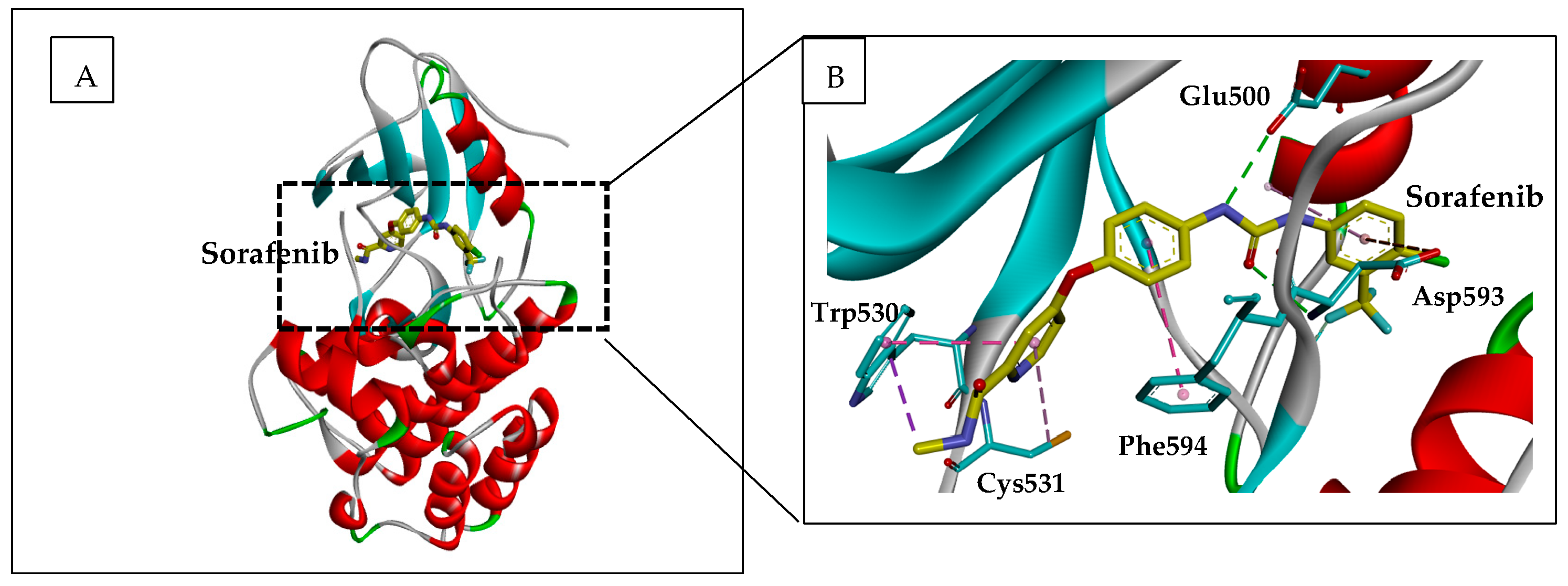
2.1.1. Sorafenib
2.1.2. Other Tyrosine Kinase Inhibitors
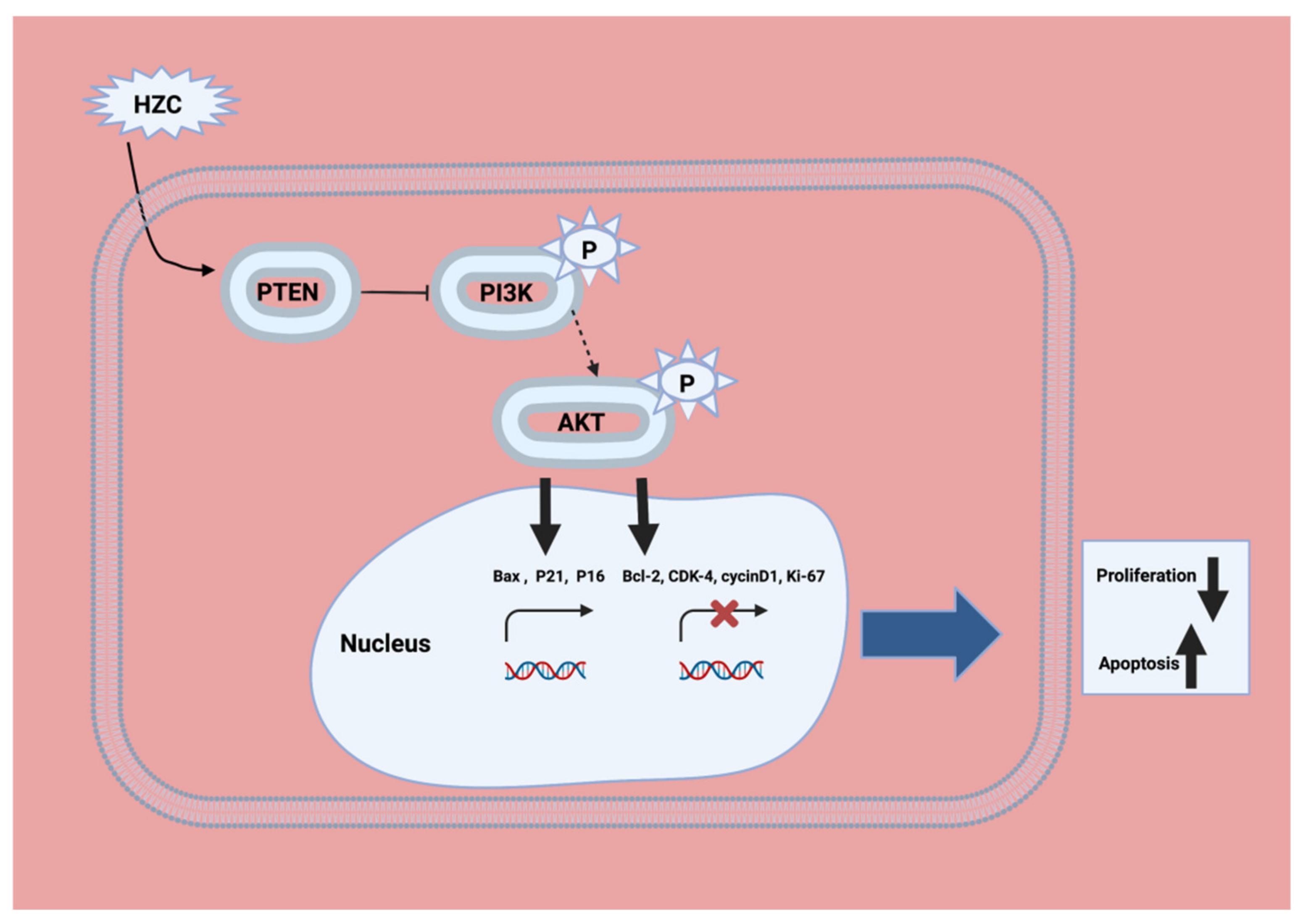
2.2. PI3K/Akt/mTOR Inhibition
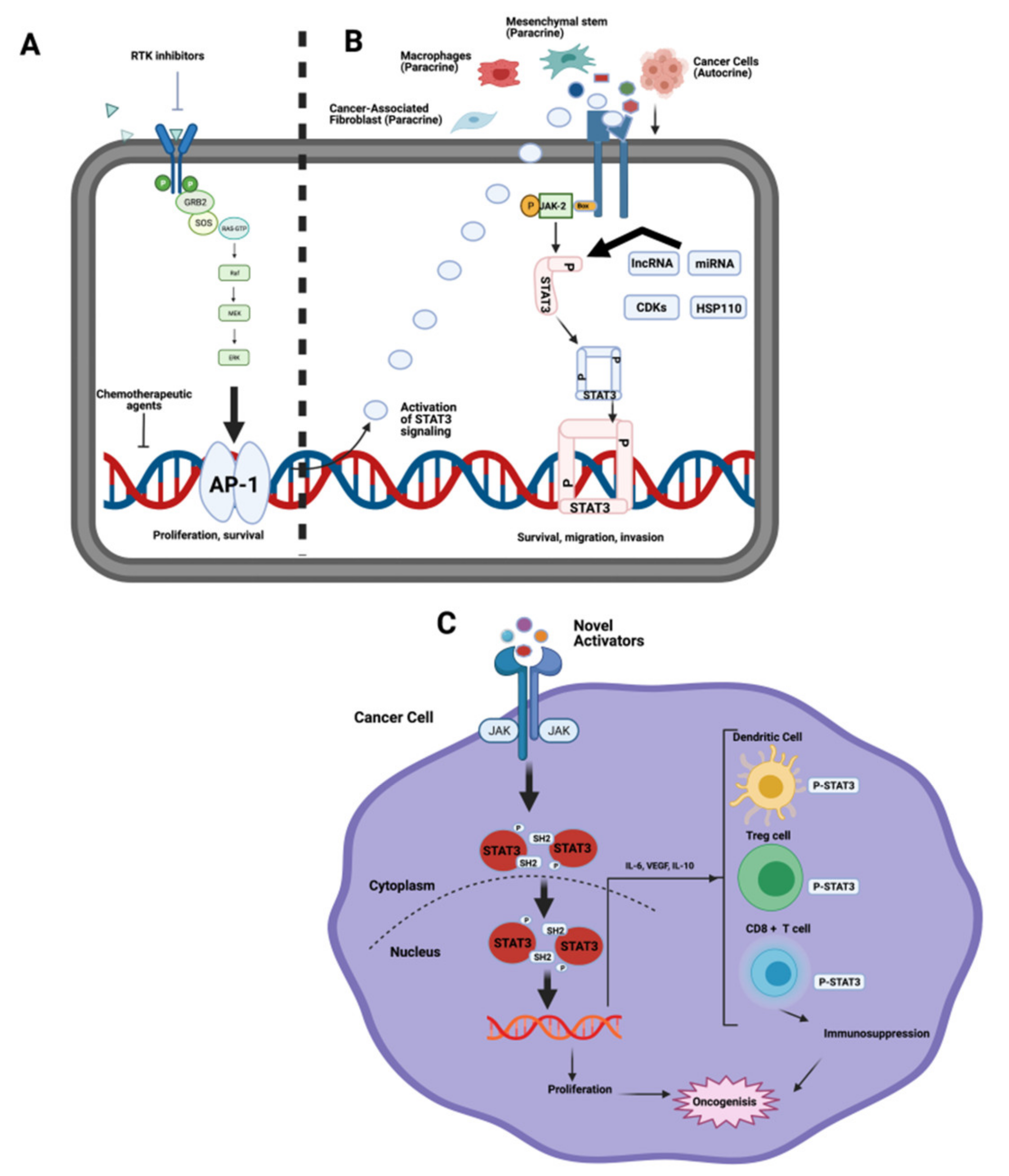
2.3. JAK/STAT Pathway
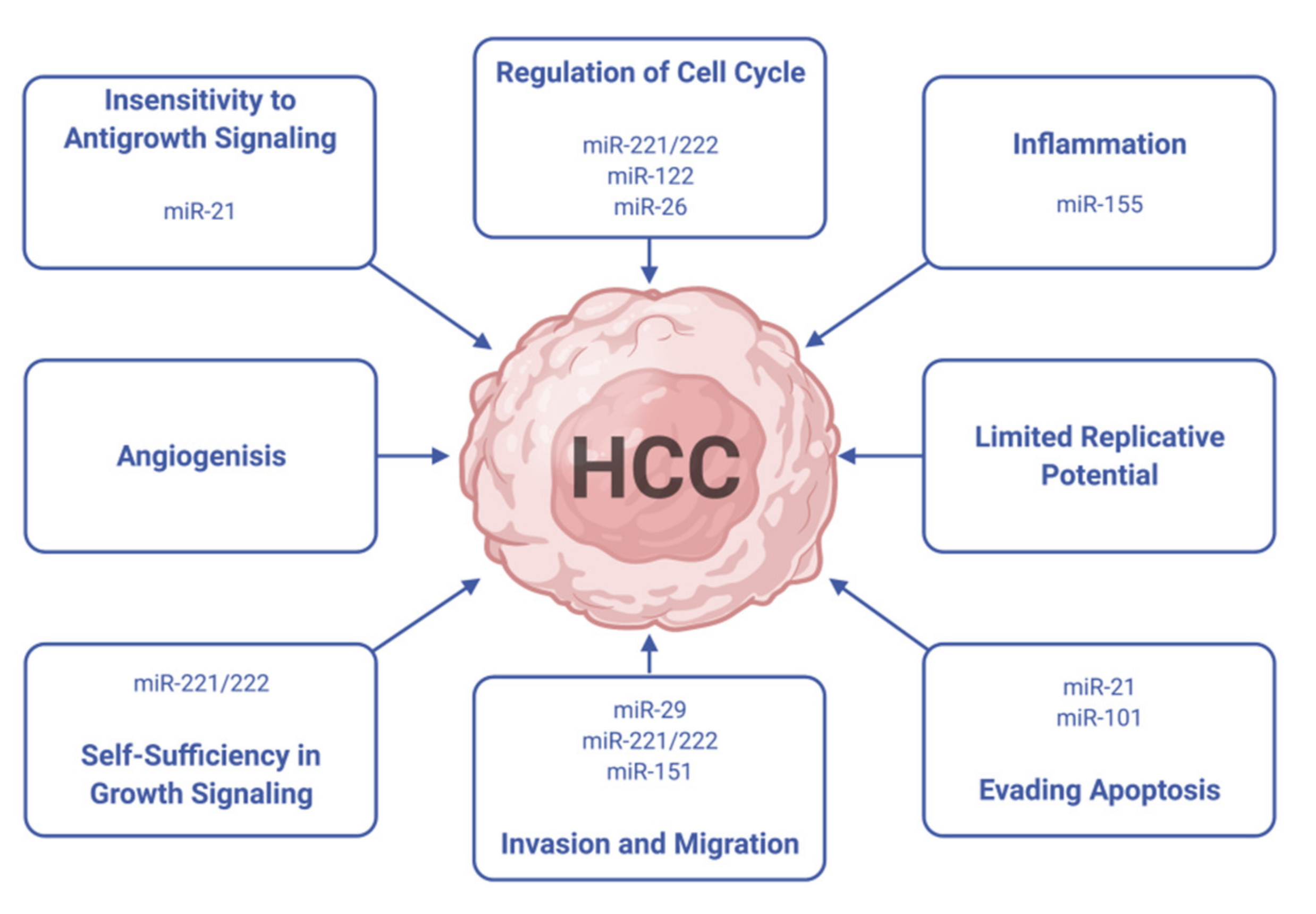
3. Micro RNAs (miRNAs) in HCC Treatment
4. Phytochemicals as Potential Anti-HCC Biomolecules
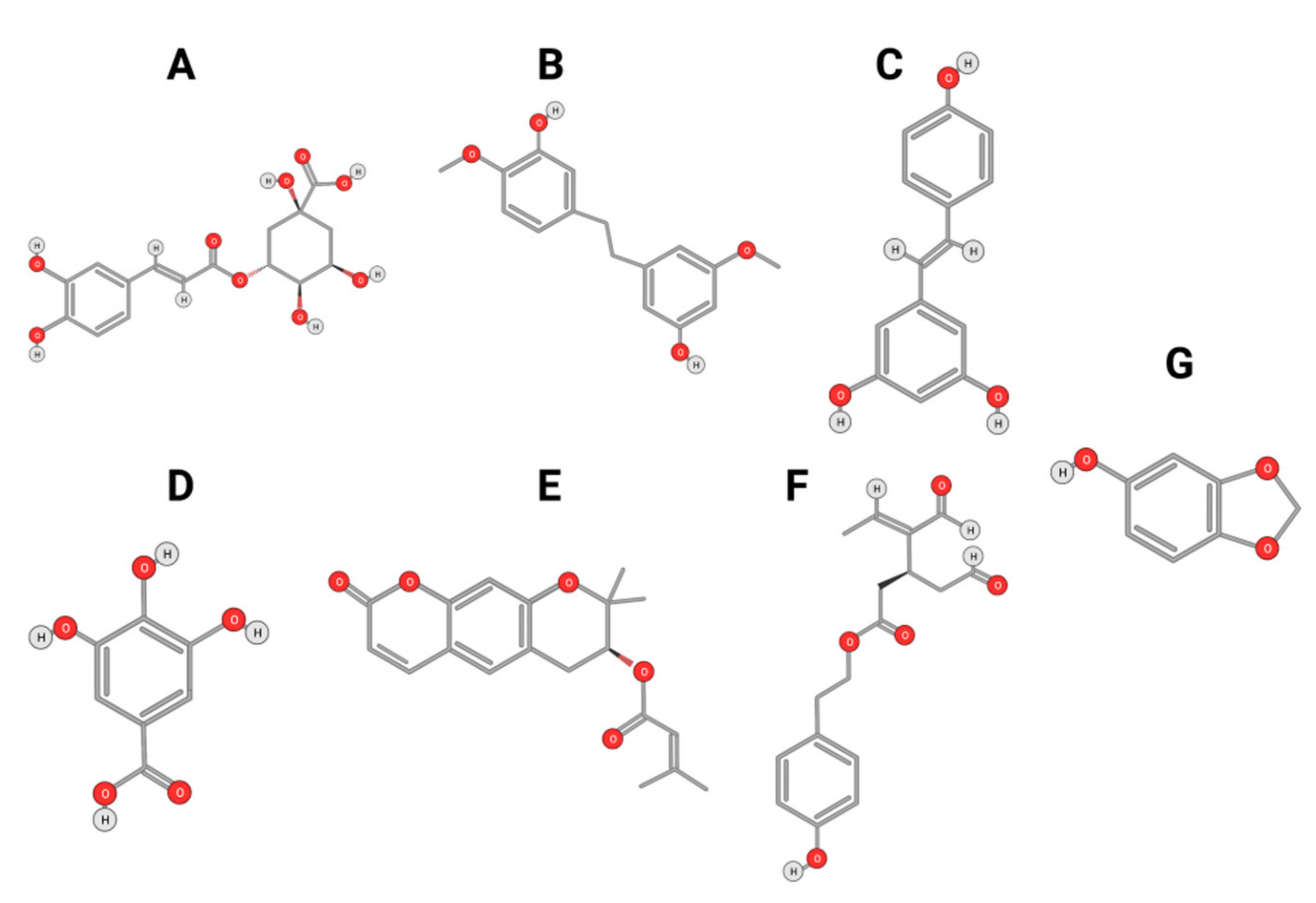
4.1. Polyphenols
4.1.1. Phenolic Compounds
4.1.2. Flavonoids
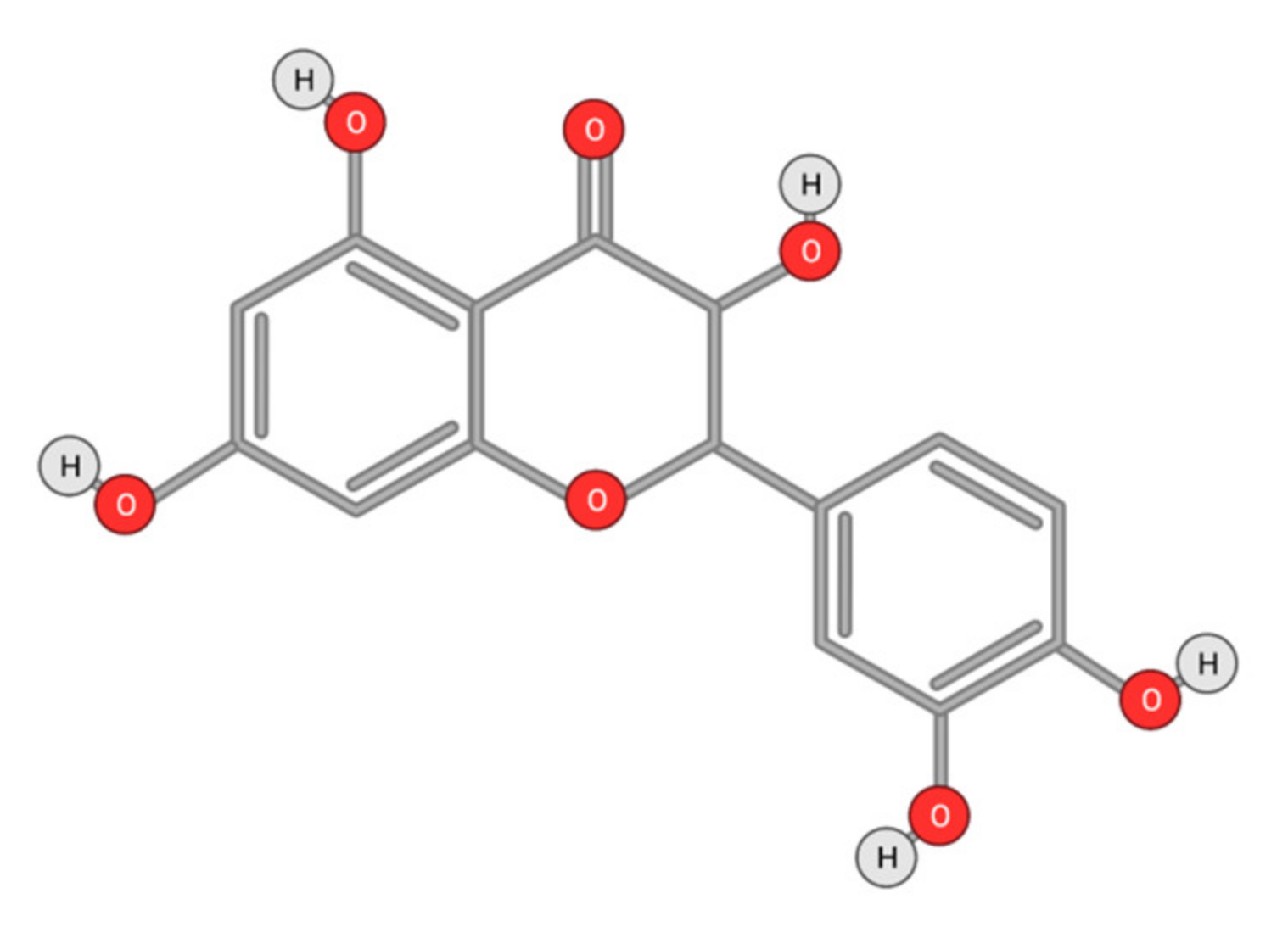
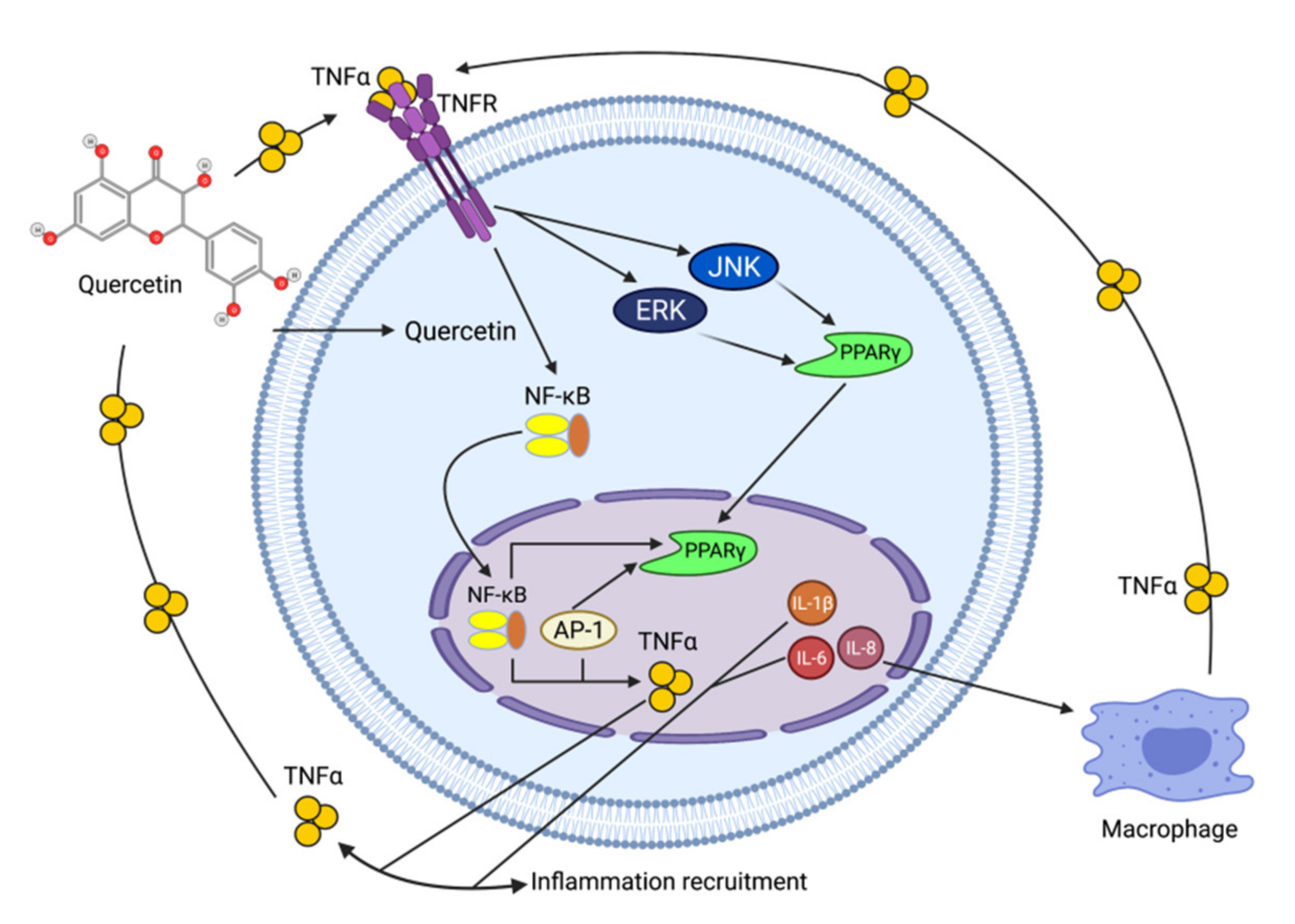
Quercetin
Other Flavonoids
4.2. Carotenoids and Alkaloids
4.2.1. Crocin
4.2.2. Capsaicin
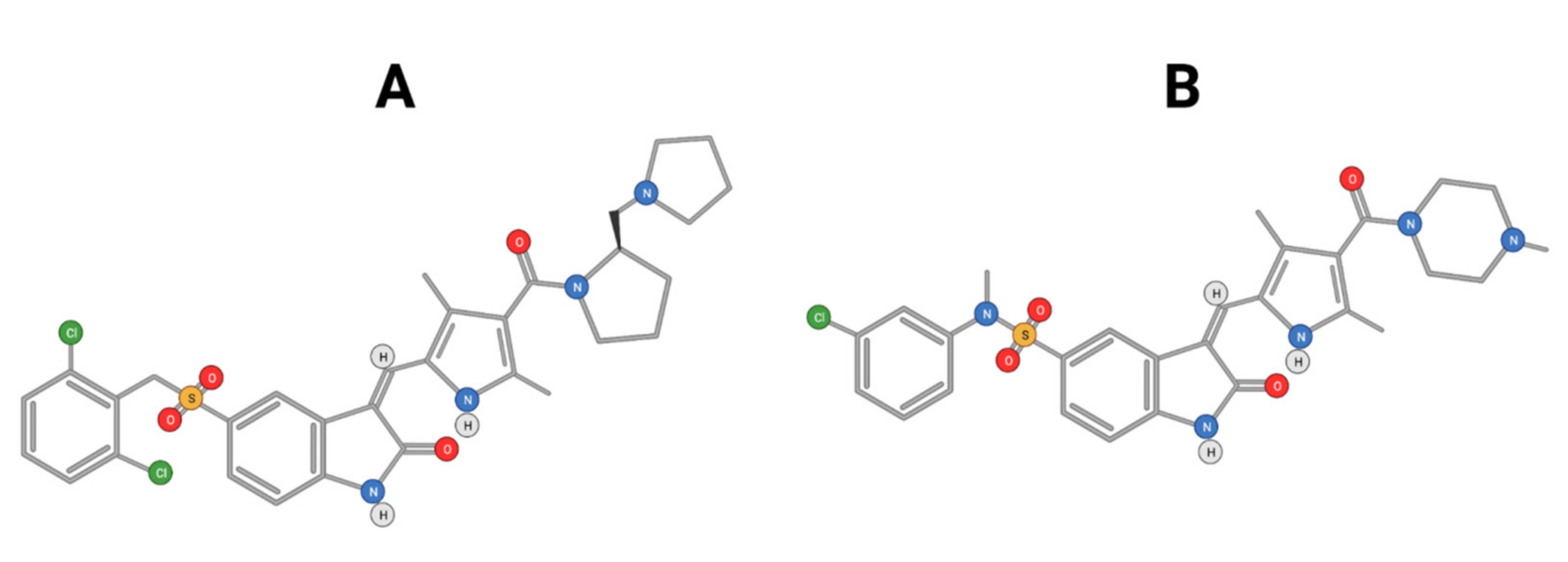
4.3. Phytochemicals and Drug Design
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wan, P.T.; Garnett, M.J.; Roe, S.M.; Lee, S.; Niculescu-Duvaz, D.; Good, V.M.; Jones, C.M.; Marshall, C.J.; Springer, C.J.; Barford, D.; et al. Mechanism of activation of the RAF-ERK signaling pathway by oncogenic mutations of B-RAF. Cell 2004, 116, 855–867. [Google Scholar] [CrossRef] [Green Version]
- Singal, A.G.; Lampertico, P.; Nahon, P. Epidemiology and surveillance for hepatocellular carcinoma: New trends. J. Hepatol. 2020, 72, 250–261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petrick, J.L.; Braunlin, M.; Laversanne, M.; Valery, P.C.; Bray, F.; McGlynn, K.A. International trends in liver cancer incidence, overall and by histologic subtype, 1978–2007. Int. J. Cancer 2016, 139, 1534–1545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chow, A.K.-M.; Yau, S.W.-L.; Ng, L. Novel molecular targets in hepatocellular carcinoma. World J. Clin. Oncol. 2020, 11, 589–605. [Google Scholar] [CrossRef]
- Clark, R.; Lee, S.-H. Anticancer properties of capsaicin against human cancer. Anticancer Res. 2016, 36, 837–843. [Google Scholar]
- Okuda, H. Hepatocellular carcinoma development in cirrhosis. Best Pract. Res. Clin. Gastroenterol. 2007, 21, 161–173. [Google Scholar] [CrossRef]
- Salem, R.; Lewandowski, R.J. Chemoembolization and radioembolization for hepatocellular carcinoma. Clin. Gastroenterol. Hepatol. 2013, 11, 604–611. [Google Scholar] [CrossRef] [Green Version]
- Wilkes, G.M. Targeted therapy: Attacking cancer with molecular and immunological targeted agents. Asia-Pacific J. Oncol. Nurs. 2018, 5, 137–155. [Google Scholar] [CrossRef]
- Parveen, A.; Subedi, L.; Kim, H.W.; Khan, Z.; Zahra, Z.; Farooqi, M.Q.; Kim, S.Y. Phytochemicals targeting VEGF and VEGF-related multifactors as anticancer therapy. J. Clin. Med. 2019, 8, 350. [Google Scholar] [CrossRef] [Green Version]
- Gauthier, A.; Ho, M. Role of sorafenib in the treatment of advanced hepatocellular carcinoma: An update. Hepatol. Res. 2013, 43, 147–154. [Google Scholar] [CrossRef] [Green Version]
- Peer, C.J.; Sissung, T.M.; Kim, A.; Jain, L.; Woo, S.; Gardner, E.R.; Kirkland, C.T.; Troutman, S.M.; English, B.C.; Richardson, E.D.; et al. Sorafenib is an inhibitor of UGT1A1 but is metabolized by UGT1A9: Implications of genetic variants on pharmacokinetics and hyperbilirubinemia. Clin. Cancer Res. 2012, 18, 2099–2107. [Google Scholar] [CrossRef] [Green Version]
- Wilhelm, S.M.; Adnane, L.; Newell, P.; Villanueva, A.; Llovet, J.M.; Lynch, M. Preclinical overview of sorafenib, a multikinase inhibitor that targets both Raf and VEGF and PDGF receptor tyrosine kinase signaling. Mol. Cancer Ther. 2008, 7, 3129–3140. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.; Loecher, M.; Iyer, R. Immunomodulation in hepatocellular cancer. J. Gastrointest. Oncol. 2018, 9, 208–219. [Google Scholar] [CrossRef] [Green Version]
- Raoul, J.-L.; Bruix, J.; Greten, T.F.; Sherman, M.; Mazzaferro, V.M.; Hilgard, P.; Scherubl, H.; Scheulen, M.E.; Germanidis, G.; Dominguez, S.; et al. Relationship between baseline hepatic status and outcome, and effect of sorafenib on liver function: SHARP trial subanalyses. J. Hepatol. 2012, 56, 1080–1088. [Google Scholar] [CrossRef]
- Cuestas, M.L.; Oubiña, J.R.; Mathet, V.L. Hepatocellular carcinoma and multidrug resistance: Past, present and new challenges for therapy improvement. World J. Pharmacol. 2015, 4, 96–116. [Google Scholar] [CrossRef]
- Keating, G.M. Sorafenib: A review in hepatocellular carcinoma. Proteins 2017, 4, 5. [Google Scholar] [CrossRef]
- Chen, K.-F.; Chen, H.-L.; Tai, W.-T.; Feng, W.-C.; Hsu, C.-H.; Chen, P.-J.; Cheng, A.-L. Activation of phosphatidylin-ositol 3-kinase/Akt signaling pathway mediates acquired resistance to sorafenib in hepatocellular carcinoma cells. J. Pharmacol. Exp. Ther. 2011, 337, 155–161. [Google Scholar] [CrossRef] [Green Version]
- Toh, T.B.; Lim, J.J.; Hooi, L.; Rashid, M.B.M.A.; Chow, E.K.-H. Targeting Jak/Stat pathway as a therapeutic strategy against SP/CD44+ tumorigenic cells in Akt/β-catenin-driven hepatocellular carcinoma. J. Hepatol. 2020, 72, 104–118. [Google Scholar] [CrossRef]
- Koeberle, D.; Dufour, J.-F.; Demeter, G.; Li, Q.; Ribi, K.; Samaras, P.; Saletti, P.; Roth, A.; Horber, D.; Buehlmann, M.; et al. Sorafenib with or without everolimus in patients with advanced hepatocellular carcinoma (HCC): A randomized multicenter, multinational phase II trial (SAKK 77/08 and SASL 29). Ann. Oncol. 2016, 27, 856–861. [Google Scholar] [CrossRef]
- Breuleux, M.; Klopfenstein, M.; Stephan, C.; Doughty, C.A.; Barys, L.; Maira, S.-M.; Kwiatkowski, D.; Lane, H.A. In-creased AKT S473 phosphorylation after mTORC1 inhibition is rictor dependent and does not predict tumor cell response to PI3K/mTOR inhibition. Mol. Cancer Ther. 2009, 8, 742–753. [Google Scholar] [CrossRef] [Green Version]
- Jiang, H.; Wu, D.; Xu, D.; Yu, H.; Zhao, Z.; Ma, D.; Jin, J. Eupafolin exhibits potent anti-angiogenic and antitumor ac-tivity in hepatocellular carcinoma. Int. J. Biol. Sci. 2017, 13, 701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lowinger, T.; Riedl, B.; Dumas, J.; Smith, R. Design and discovery of small molecules targeting Raf-1 kinase. Curr. Pharm. Des. 2002, 8, 2269–2278. [Google Scholar] [CrossRef] [PubMed]
- Nagar, B.; Bornmann, W.G.; Pellicena, P.; Schindler, T.; Veach, D.R.; Miller, W.T.; Clarkson, B.; Kuriyan, J. Crystal structures of the kinase domain of c-Abl in complex with the small molecule inhibitors PD173955 and imatinib (STI-571). Cancer Res. 2002, 62, 4236–4243. [Google Scholar] [PubMed]
- Pargellis, C.; Tong, L.; Churchill, L.; Cirillo, P.F.; Gilmore, T.; Graham, A.G.; Grob, P.M.; Hickey, E.R.; Moss, N.; Pav, S.; et al. Inhibition of p38 MAP kinase by utilizing a novel allosteric binding site. Nat. Struct. Biol. 2002, 9, 268–272. [Google Scholar] [CrossRef]
- Wu, C.; Wang, M.; Tang, Q.; Luo, R.; Chen, L.; Zheng, P.; Zhu, W. Design, synthesis, activity and docking study of sorafenib analogs bearing sulfonylurea unit. Molecules 2015, 20, 19361–19371. [Google Scholar] [CrossRef] [Green Version]
- Yao, J.; Chen, J.; He, Z.; Sun, W.; Xu, W. Design, synthesis and biological activities of thiourea containing sorafenib analogs as antitumor agents. Bioorg. Med. Chem. 2012, 20, 2923–2929. [Google Scholar] [CrossRef]
- Wang, M.; Xu, S.; Wu, C.; Liu, X.; Tao, H.; Huang, Y.; Liu, Y.; Zheng, P.; Zhu, W. Design, synthesis and activity of novel sorafenib analogues bearing chalcone unit. Bioorg. Med. Chem. Lett. 2016, 26, 5450–5454. [Google Scholar] [CrossRef]
- Finn, R.S.; Kang, Y.-K.; Mulcahy, M.; Polite, B.N.; Lim, H.Y.; Walters, I.; Baudelet, C.; Manekas, D.; Park, J.-W. Phase II, open-label study of brivanib as second-line therapy in patients with advanced hepatocellular carcinoma. Clin. Cancer Res. 2012, 18, 2090–2098. [Google Scholar] [CrossRef] [Green Version]
- Huynh, H.; Ngo, V.C.; Fargnoli, J.; Ayers, M.; Soo, K.C.; Koong, H.N.; Thng, C.H.; Ong, H.S.; Chung, A.; Chow, P.; et al. Brivanib alaninate, a dual inhibitor of vascular endothelial growth factor receptor and fibroblast growth factor receptor tyrosine kinases, induces growth inhibition in mouse models of human hepatocellular carcinoma. Clin. Cancer Res. 2008, 14, 6146–6153. [Google Scholar] [CrossRef] [Green Version]
- Alberts, S.R.; Fitch, T.R.; Kim, G.P.; Morlan, B.W.; Dakhil, S.R.; Gross, H.M.; Nair, S. Cediranib (AZD2171) in patients with advanced hepatocellular carcinoma: A phase II north central cancer treatment group (NCCTG) Clinical Trial. Am. J. Clin. Oncol. 2012, 35, 329. [Google Scholar] [CrossRef]
- Huynh, H.; Chow, P.; Tai, W.M.; Choo, S.P.; Chung, A.Y.F.; Ong, H.S.; Soo, K.C.; Ong, R.; Linnartz, R.; Shi, M.M. Dovitinib demonstrates antitumor and antimetastatic activities in xenograft models of hepatocellular carcinoma. J. Hepatol. 2012, 56, 595–601. [Google Scholar] [CrossRef]
- Cheng, A.-L.; Thongprasert, S.; Lim, H.Y.; Sukeepaisarnjaroen, W.; Yang, T.-S.; Wu, C.-C.; Chao, Y.; Chan, S.; Kudo, M.; Ikeda, M.; et al. Randomized, open-label phase 2 study comparing frontline dovitinib versus sorafenib in patients with advanced hepatocellular carcinoma. Hepatology 2016, 64, 774–784. [Google Scholar] [CrossRef]
- Philip, P.A.; Mahoney, M.R.; Allmer, C.; Thomas, J.; Pitot, H.C.; Kim, G.; Donehower, R.C.; Fitch, T.; Picus, J.; Erlichman, C. Phase II study of erlotinib (OSI-774) in patients with advanced hepatocellular cancer. J. Clin. Oncol. 2005, 23, 6657–6663. [Google Scholar] [CrossRef]
- Thomas, M.B.; Chadha, R.; Glover, K.; Wang, X.; Morris, J.; Brown, T.; Rashid, A.; Dancey, J.; Abbruzzese, J.L. Phase 2 study of erlotinib in patients with unresectable hepatocellular carcinoma. Cancer 2007, 110, 1059–1067. [Google Scholar] [CrossRef]
- Zhu, B.-D.; Yuan, S.-J.; Zhao, Q.-C.; Li, X.; Li, Y.; Lu, Q.-Y. Antitumor effect of Gefitinib, an epidermal growth factor receptor tyrosine kinase inhibitor, combined with cytotoxic agent on murine hepatocellular carcinoma. World J. Gastroenterol. 2005, 11, 1382–1386. [Google Scholar] [CrossRef]
- O’Neil, B.H.; Goff, L.W.; Kauh, J.S.W.; Strosberg, J.R.; Bekaii-Saab, T.S.; Lee, R.-m.; Kazi, A.; Moore, D.T.; Learoyd, M.; Lush, R.M.; et al. Phase II study of the mitogen-activated protein kinase 1/2 inhibitor selumetinib in patients with advanced hepa-tocellular carcinoma. J. Clin. Oncol. 2011, 29, 2350. [Google Scholar] [CrossRef]
- Yi, T.; Cho, S.-G.; Yi, Z.; Pang, X.; Rodriguez, M.; Wang, Y.; Sethi, G.; Aggarwal, B.B.; Liu, M. Thymoquinone inhibits tumor angiogenesis and tumor growth through suppressing AKT and extracellular signal-regulated kinase signaling pathways. Mol. Cancer Ther. 2008, 7, 1789–1796. [Google Scholar] [CrossRef] [Green Version]
- Cainap, C.; Qin, S.; Huang, W.-T.; Chung, I.J.; Pan, H.; Cheng, Y.; Kudo, M.; Kang, Y.-K.; Chen, P.-J.; Toh, H.-C. Li-nifanib versus Sorafenib in patients with advanced hepatocellular carcinoma: Results of a randomized phase III trial. J. Clin. Oncol. 2015, 33, 172. [Google Scholar] [CrossRef]
- Toh, H.C.; Chen, P.-J.; Carr, B.; Knox, J.J.; Gill, S.; Ansell, P.; McKeegan, E.M.; Dowell, B.; Pedersen, M.; Qin, Q.; et al. Phase 2 trial of linifanib (ABT-869) in patients with unresectable or metastatic hepatocellular carcinoma. Cancer 2013, 119, 380–387. [Google Scholar] [CrossRef]
- Jasinghe, V.J.; Xie, Z.; Zhou, J.; Khng, J.; Poon, L.-F.; Senthilnathan, P.; Glaser, K.B.; Albert, D.H.; Davidsen, S.K.; Chen, C.-S. ABT-869, a multi-targeted tyrosine kinase inhibitor, in combination with rapamycin is effective for subcutaneous hepa-tocellular carcinoma xenograft. J. Hepatol. 2008, 49, 985–997. [Google Scholar] [CrossRef]
- Zhu, A.X.; Sahani, D.V.; Duda, D.G.; di Tomaso, E.; Ancukiewicz, M.; Catalano, O.A.; Sindhwani, V.; Blaszkowsky, L.S.; Yoon, S.S.; Lahdenranta, J.; et al. Efficacy, Safety, and Potential Biomarkers of Sunitinib Monotherapy in Advanced Hepatocellular Carcinoma: A Phase II Study. J. Clin. Oncol. 2009, 27, 3027–3035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanai, F.; Yoshida, H.; Tateishi, R.; Sato, S.; Kawabe, T.; Obi, S.; Kondo, Y.; Taniguchi, M.; Tagawa, K.; Ikeda, M.; et al. A phase I/II trial of the oral antiangiogenic agent TSU-68 in patients with advanced hepatocellular carcinoma. Cancer Chemother. Pharmacol. 2010, 67, 315–324. [Google Scholar] [CrossRef]
- Kudo, M.; Cheng, A.-L.; Park, J.-W.; Park, J.H.; Liang, P.-C.; Hidaka, H.; Izumi, N.; Heo, J.; Lee, Y.J.; Sheen, I.-S.; et al. Orantinib versus placebo combined with transcatheter arterial chemoembolisation in patients with unresectable hepatocellular carcinoma (ORIENTAL): A randomised, double-blind, placebo-controlled, multicentre, phase 3 study. Lancet Gastroenterol. Hepatol. 2018, 3, 37–46. [Google Scholar] [CrossRef]
- Siegel, A.B.; Cohen, E.I.; Ocean, A.; Lehrer, D.; Goldenberg, A.; Knox, J.J.; Chen, H.; Clark-Garvey, S.; Weinberg, A.; Mandeli, J.; et al. Phase II trial evaluating the clinical and biologic effects of bevacizumab in unresectable hepatocellular carcinoma. J. Clin. Oncol. 2008, 26, 2992–2998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kubota, M.; Shimizu, M.; Baba, A.; Ohno, T.; Kochi, T.; Shirakami, Y.; Moriwaki, H. Combination of bevacizumab and acyclic retinoid inhibits the growth of hepatocellular carcinoma xenografts. J. Nutr. Sci. Vitaminol. 2014, 60, 357–362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, A.X.; Stuart, K.; Blaszkowsky, L.S.; Ma, A.M.; Reitberg, D.P.; Clark, J.W.; Enzinger, P.C.; Bhargava, P.; Meyerhardt, J.A.; Horgan, K.; et al. Phase 2 study of cetuximab in patients with advanced hepatocellular carcinoma. Cancer Interdiscip. Int. J. Am. Cancer Soc. 2007, 110, 581–589. [Google Scholar] [CrossRef] [PubMed]
- Llovet, J.M.; Bruix, J. Novel advancements in the management of hepatocellular carcinoma in 2008. J. Hepatol. 2008, 48, S20–S37. [Google Scholar] [CrossRef] [PubMed]
- Peters, B.M.L.; Miksad, R.A. Cabozantinib in the treatment of hepatocellular carcinoma. Future Oncol. 2017, 13, 1915–1929. [Google Scholar] [CrossRef]
- Cao, H.; Phan, H.; Yang, L.-X. Improved chemotherapy for hepatocellular carcinoma. Anticancer Res. 2012, 32, 1379–1386. [Google Scholar]
- You, H.; Ding, W.; Dang, H.; Jiang, Y.; Rountree, C.B. c-Met represents a potential therapeutic target for personalized treatment in hepatocellular carcinoma. Hepatology 2011, 54, 879–889. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.-G.; Yin, X.-C.; Liu, X.-F.; Meng, K.-W.; Tang, K.; Huang, F.-L.; Xu, G.; Gao, J. Puerarin induces hepatocellular car-cinoma cell apoptosis modulated by MAPK signaling pathways in a dose-dependent manner. Anticancer. Res. 2017, 37, 4425–4431. [Google Scholar]
- Stephens, L.; Eguinoa, A.; Erdjument-Bromage, H.; Lui, M.; Cooke, F.; Coadwell, J.; Smrcka, A.; Thelen, M.; Cadwallader, K.; Tempst, P.; et al. The Gβγ sensitivity of a PI3K is dependent upon a tightly associated adaptor, p101. Cell 1997, 89, 105–114. [Google Scholar] [CrossRef] [Green Version]
- Franke, T.F.; Yang, S.-I.; Chan, T.; Datta, K.; Kazlauskas, A.; Morrison, D.K.; Kaplan, D.R.; Tsichlis, P.N. The protein kinase encoded by the Akt proto-oncogene is a target of the PDGF-activated phosphatidylinositol 3-kinase. Cell 1995, 81, 727–736. [Google Scholar] [CrossRef] [Green Version]
- Klippel, A.; Kavanaugh, W.M.; Pot, D.; Williams, L.T. A specific product of phosphatidylinositol 3-kinase directly activates the protein kinase Akt through its pleckstrin homology domain. Mol. Cell. Biol. 1997, 17, 338–344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matter, M.; Decaens, T.; Andersen, J.B.; Thorgeirsson, S.S. Targeting the mTOR pathway in hepatocellular carcinoma: Current state and future trends. J. Hepatol. 2014, 60, 855–865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kunter, I.; Erdal, E.; Nart, D.; Yilmaz, F.; Karademir, S.; Sagol, O.; Atabey, N. Active form of AKT controls cell proliferation and response to apoptosis in hepatocellular carcinoma. Oncol. Rep. 2014, 31, 573–580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; Zhang, M.; Zhang, Z.; Jia, Y.; Zhang, C.; Peng, L. Hydrazinocurcumin and 5-fluorouracil enhance apoptosis and restrain tumorigenicity of HepG2 cells via disrupting the PTEN-mediated PI3K/Akt signaling pathway. Biomed. Pharmacother. 2020, 129, 109851. [Google Scholar] [CrossRef]
- Viollet, B.; Guigas, B.; Leclerc, J.; Hébrard, S.; Lantier, L.; Mounier, R.; Andreelli, F.; Foretz, M. AMP-activated protein kinase in the regulation of hepatic energy metabolism: From physiology to therapeutic perspectives. Acta Physiol. 2009, 196, 81–98. [Google Scholar] [CrossRef] [Green Version]
- Fasolo, A.; Sessa, C. Targeting mTOR pathways in human malignancies. Curr. Pharm. Des. 2012, 18, 2766–2777. [Google Scholar] [CrossRef]
- Wang, Z.; Fan, J.; Zhou, J.; Wu, Z.-Q.; Qiu, S.-J.; Yu, Y.; Huang, X.-W.; Tang, Z.-Y. Inhibition of growth and metastasis of hepatocellular carcinoma by rapamycin: Experiment with mice. Zhonghua Yi Xue Za Zhi 2006, 86, 1666–1670. [Google Scholar]
- Piguet, A.-C.; Saar, B.; Hlushchuk, R.; St-Pierre, M.V.; McSheehy, P.M.J.; Radojevic, V.; Afthinos, M.; Terracciano, L.; Djonov, V.; Dufour, J.-F. everolimus augments the effects of sorafenib in a syngeneic orthotopic model of hepatocellular carcinoma. Mol. Cancer Ther. 2011, 10, 1007–1017. [Google Scholar] [CrossRef] [Green Version]
- Huynh, H.; Chow, K.P.; Soo, K.C.; Toh, H.C.; Choo, S.P.; Foo, K.F.; Poon, D.; Ngo, V.C.; Tran, E. RAD001 (everolimus) inhibits tumour growth in xenograft models of human hepatocellular carcinoma. J. Cell. Mol. Med. 2009, 13, 1371–1380. [Google Scholar] [CrossRef]
- Zhou, P.; Yang, X.-L.; Wang, X.-G.; Hu, B.; Zhang, L.; Zhang, W.; Si, H.-R.; Zhu, Y.; Li, B.; Huang, C.-L.; et al. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature 2020, 579, 270–273. [Google Scholar] [CrossRef] [Green Version]
- Thomas, S.J.; Snowden, J.A.; Zeidler, M.; Danson, S. The role of JAK/STAT signalling in the pathogenesis, prognosis and treatment of solid tumours. Br. J. Cancer 2015, 113, 365–371. [Google Scholar] [CrossRef] [Green Version]
- Svinka, J.; Mikulits, W.; Eferl, R. STAT3 in hepatocellular carcinoma: New perspectives. Hepatic Oncol. 2014, 1, 107–120. [Google Scholar] [CrossRef]
- Rosmorduc, O.; Desbois-Mouthon, C. Targeting STAT3 in hepatocellular carcinoma: Sorafenib again. J. Hepatol. 2011, 55, 957–959. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Liao, X.; Yu, T.; Gong, Y.; Zhang, L.; Huang, J.; Yang, C.; Han, C.; Yu, L.; Zhu, G.; et al. Analysis of clinical significance and prospective molecular mechanism of main elements of the JAK/STAT pathway in hepatocellular carcinoma. Int. J. Oncol. 2019, 55, 805–822. [Google Scholar] [CrossRef] [Green Version]
- Calvisi, D.F.; Ladu, S.; Gorden, A.; Farina, M.; Conner, E.A.; Lee, J.; Factor, V.M.; Thorgeirsson, S.S. Ubiquitous activation of Ras and Jak/Stat pathways in human HCC. Gastroenterology 2006, 130, 1117–1128. [Google Scholar] [CrossRef]
- Tang, J.J.H.; Thng, D.K.H.; Lim, J.J.; Toh, T.B. JAK/STAT signaling in hepatocellular carcinoma. Hepatic Oncol. 2020, 7, HEP18. [Google Scholar] [CrossRef] [Green Version]
- Wonganan, O.; He, Y.-J.; Shen, X.-F.; Wongkrajang, K.; Suksamrarn, A.; Zhang, G.-L.; Wang, F. 6-Hydroxy-3-O-methyl-kaempferol 6-O-glucopyranoside potentiates the anti-proliferative effect of interferon α/β by promoting activation of the JAK/STAT signaling by inhibiting SOCS3 in hepatocellular carcinoma cells. Toxicol. Appl. Pharmacol. 2017, 336, 31–39. [Google Scholar] [CrossRef]
- He, G.; Karin, M. NF-κB and STAT3–key players in liver inflammation and cancer. Cell. Res. 2011, 21, 159–168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, L.; Lin, S.; Xu, L.; Lin, J.; Zhao, C.; Huang, X. Novel activators and small-molecule inhibitors of STAT3 in cancer. Cytokine Growth Factor Rev. 2019, 49, 10–22. [Google Scholar] [CrossRef] [PubMed]
- Xu, G.; Zhu, L.; Wang, Y.; Shi, Y.; Gong, A.; Wu, C. Stattic Enhances Radiosensitivity and Reduces Radio-Induced Migration and Invasion in HCC Cell Lines through an Apoptosis Pathway. BioMed Res. Int. 2017, 2017, 1832494. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Yao, J.; Xie, M.; Liu, Y.; Zheng, M. Exosomal miRNAs in hepatocellular carcinoma development and clinical responses. J. Hematol. Oncol. 2018, 11, 1–9. [Google Scholar] [CrossRef]
- Xie, M.; A Steitz, J. Versatile microRNA biogenesis in animals and their viruses. RNA Biol. 2014, 11, 673–681. [Google Scholar] [CrossRef] [Green Version]
- Wong, C.-M.; Tsang, F.H.; Ng, I.O. Non-coding RNAs in hepatocellular carcinoma: Molecular functions and pathological implications. Nat. Rev. Gastroenterol. Hepatol. 2018, 15, 137–151. [Google Scholar] [CrossRef]
- Braconi, C.; Valeri, N.; Kogure, T.; Gasparini, P.; Huang, N.; Nuovo, G.J.; Terracciano, L.; Croce, C.M.; Patel, T. Expression and functional role of a transcribed noncoding RNA with an ultraconserved element in hepatocellular carcinoma. Proc. Natl. Acad. Sci. USA 2011, 108, 786–791. [Google Scholar] [CrossRef] [Green Version]
- Kogure, T.; Lin, W.-L.; Yan, I.K.; Braconi, C.; Patel, T. Intercellular nanovesicle-mediated microRNA transfer: A mechanism of environmental modulation of hepatocellular cancer cell growth. Hepatology 2011, 54, 1237–1248. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Huang, Y.; Huang, J.; Lin, L.; Wei, G. Gigantol attenuates the proliferation of human liver cancer HepG2 cells through the PI3K/Akt/NF-κB signaling pathway. Oncol. Rep. 2017, 37, 865–870. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.-Y.; Zhang, K.; Chen, D.-Q.; Chen, J.; Feng, B.; Song, H.; Chen, Y.; Zhu, Z.; Lu, L.; De, W.; et al. MicroRNA-451: Epitheli-al-mesenchymal transition inhibitor and prognostic biomarker of hepatocelluar carcinoma. Oncotarget 2015, 6, 18613. [Google Scholar] [CrossRef] [Green Version]
- Hsu, S.-H.; Wang, B.; Kota, J.; Yu, J.; Costinean, S.; Kutay, H.; Yu, L.; Bai, S.; La Perle, K.; Chivukula, R.R.; et al. Essential metabolic, anti-inflammatory, and anti-tumorigenic functions of miR-122 in liver. J. Clin. Investig. 2012, 122, 2871–2883. [Google Scholar] [CrossRef] [Green Version]
- Zeisel, M.B.; Felmlee, D.J.; Baumert, T.F. Hepatitis C virus entry. Hepat. C Virus Mol. Virol. Antivir. Ther. 2013, 369, 87–112. [Google Scholar]
- Chen, C.-L.; Wu, J.-C.; Chen, G.-Y.; Yuan, P.-H.; Tseng, Y.-W.; Li, K.-C.; Hwang, S.-M.; Hu, Y.-C. Baculovirus-mediated miRNA regulation to suppress hepatocellular carcinoma tumorigenicity and metastasis. Mol. Ther. 2015, 23, 79–88. [Google Scholar] [CrossRef] [Green Version]
- Wang, F.; Li, L.; Piontek, K.; Sakaguchi, M.; Selaru, F.M. Exosome miR-335 as a novel therapeutic strategy in hepatocellular carcinoma. Hepatology 2018, 67, 940–954. [Google Scholar] [CrossRef]
- Tang, J.; Li, Y.; Liu, K.; Zhu, Q.; Yang, W.-H.; Xiong, L.-K.; Guo, D.-L. Exosomal miR-9-3p suppresses HBGF-5 expression and is a functional biomarker in hepatocellular carcinoma. Minerva Med. 2017, 109, 15–23. [Google Scholar] [CrossRef]
- Xiong, L.; Zhen, S.; Yu, Q.; Gong, Z. HCV-E2 inhibits hepatocellular carcinoma metastasis by stimulating mast cells to secrete exosomal shuttle microRNAs. Oncol. Lett. 2017, 14, 2141–2146. [Google Scholar] [CrossRef] [Green Version]
- Jin, F.; Wang, Y.; Li, M.; Zhu, Y.; Liang, H.; Wang, C.; Wang, F.; Zhang, C.-Y.; Zen, K.; Li, L. MiR-26 enhances chemosensitivity and promotes apoptosis of hepatocellular carcinoma cells through inhibiting autophagy. Cell Death Dis. 2018, 8, e2540. [Google Scholar] [CrossRef]
- Afrin, R.; Arumugam, S.; Rahman, A.; Wahed, M.I.I.; Karuppagounder, V.; Harima, M.; Suzuki, H.; Miyashita, S.; Suzuki, K.; Yoneyama, H.; et al. Curcumin ameliorates liver damage and progression of NASH in NASH-HCC mouse model possibly by mod-ulating HMGB1-NF-κB translocation. Int. Immunopharmacol. 2017, 44, 174–182. [Google Scholar] [CrossRef]
- Yan, Y.; Liu, N.; Hou, N.; Dong, L.; Li, J. Chlorogenic acid inhibits hepatocellular carcinoma in vitro and in vivo. J. Nutr. Biochem. 2017, 46, 68–73. [Google Scholar] [CrossRef]
- Jagan, S.; Ramakrishnan, G.; Anandakumar, P.; Kamaraj, S.; Devaki, T. Antiproliferative potential of gallic acid against di-ethylnitrosamine-induced rat hepatocellular carcinoma. Mol. Cell. Biochem. 2008, 319, 51–59. [Google Scholar] [CrossRef]
- Chen, Y.; Zhou, B.; Xu, L.; Fan, H.; Xie, J.; Wang, D. MicroRNA-146a promotes gastric cancer cell apoptosis by targeting transforming growth factor β-activated kinase 1. Mol. Med. Rep. 2017, 16, 755–763. [Google Scholar] [CrossRef] [Green Version]
- Yu, H.; Pan, C.; Zhao, S.; Wang, Z.; Zhang, H.; Wu, W. Resveratrol inhibits tumor necrosis factor-α-mediated matrix metal-loproteinase-9 expression and invasion of human hepatocellular carcinoma cells. Biomed. Pharmacother. 2008, 62, 366–372. [Google Scholar] [CrossRef]
- Pei, T.; Meng, Q.; Han, J.; Sun, H.; Li, L.; Song, R.; Sun, B.; Pan, S.; Liang, D.; Liu, L. (−)-Oleocanthal inhibits growth and metastasis by blocking activation of STAT3 in human hepatocellular carcinoma. Oncotarget 2016, 7, 43475–43491. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Qin, S.; Zheng, Y.; Han, L.; Zhang, M.; Luo, N.; Liu, Z.; Gu, N.; Gu, X.; Yin, X. Molecular targeting of VEGF/VEGFR signaling by the anti-VEGF monoclonal antibody BD0801 inhibits the growth and induces apoptosis of human hepatocellular carcinoma cells in vitro and in vivo. Cancer Biol. Ther. 2017, 18, 166–176. [Google Scholar] [CrossRef] [Green Version]
- Agullo, G.; Gamet-Payrastre, L.; Manenti, S.; Viala, C.; Rémésy, C.; Chap, H.; Payrastre, B. Relationship between flavonoid structure and inhibition of phosphatidylinositol 3-kinase: A comparison with tyrosine kinase and protein kinase C inhibition. Biochem. Pharmacol. 1997, 53, 1649–1657. [Google Scholar] [CrossRef]
- Peet, G.W.; Li, J. IκB kinases α and β show a random sequential kinetic mechanism and are inhibited by staurosporine and quercetin. J. Biol. Chem. 1999, 274, 32655–32661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maurya, A.K.; Vinayak, M. Anticarcinogenic action of quercetin by downregulation of phosphatidylinositol 3-kinase (PI3K) and protein kinase C (PKC) via induction of p53 in hepatocellular carcinoma (HepG2) cell line. Mol. Biol. Rep. 2015, 42, 1419–1429. [Google Scholar] [CrossRef]
- Li, Y.; Yao, J.; Han, C.; Yang, J.; Chaudhry, M.T.; Wang, S.; Liu, H.; Yin, Y. Quercetin, Inflammation and Immunity. Nutrients 2016, 8, 167. [Google Scholar] [CrossRef]
- Sur, S.; Pal, D.; Roy, R.; Barua, A.; Roy, A.; Saha, P.; Panda, C.K. Tea polyphenols EGCG and TF restrict tongue and liver carcinogenesis simultaneously induced by N-nitrosodiethylamine in mice. Toxicol. Appl. Pharmacol. 2016, 300, 34–46. [Google Scholar] [CrossRef]
- Li, N.; Men, W.; Zheng, Y.; Wang, H.; Meng, X. Oroxin B Induces Apoptosis by Down-Regulating MicroRNA-221 Resulting in the Inactivation of the PTEN/PI3K/AKT Pathway in Liver Cancer. Molecules 2019, 24, 4384. [Google Scholar] [CrossRef] [Green Version]
- Thirusangu, P.; Vigneshwaran, V.; Avin, B.V.; Rakesh, H.; Vikas, H.; Prabhakar, B. Scutellarein antagonizes the tumorigenesis by modulating cytokine VEGF mediated neoangiogenesis and DFF-40 actuated nucleosomal degradation. Biochem. Biophys. Res. Commun. 2017, 484, 85–92. [Google Scholar] [CrossRef]
- Yang, H.; Wang, J.; Fan, J.-H.; Zhang, Y.-Q.; Zhao, J.-X.; Dai, X.-J.; Liu, Q.; Shen, Y.-J.; Liu, C.; Sun, W.-D.; et al. Ilexgenin A exerts anti-inflammation and anti-angiogenesis effects through inhibition of STAT3 and PI3K pathways and exhibits synergistic effects with Sorafenib on hepatoma growth. Toxicol. Appl. Pharmacol. 2017, 315, 90–101. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Cheng, X.; Chen, C.; Huijuan, W.; Zhao, H.; Liu, W.; Xiang, Z.; Wang, Q. Apigenin, a flavonoid constituent derived from P. villosa, inhibits hepatocellular carcinoma cell growth by CyclinD1/CDK4 regulation via p38 MAPK-p21 signaling. Pathol. Res. Pract. 2020, 216, 152701. [Google Scholar] [CrossRef] [PubMed]
- Lim, W.C.; Kim, H.; Ko, H. Delphinidin inhibits epidermal growth factor-induced epithelial-to-mesenchymal transition in hepatocellular carcinoma cells. J. Cell. Biochem. 2019, 120, 9887–9899. [Google Scholar] [CrossRef] [PubMed]
- Amin, A.; Hamza, A.A.; Bajbouj, K.; Ashraf, S.S.; Daoud, S. Saffron: A potential candidate for a novel anticancer drug against hepatocellular carcinoma. Hepatology 2011, 54, 857–867. [Google Scholar] [CrossRef]
- Amin, A.; Awad, B. Crocin-Sorafenib Combination Therapy for Liver Cancer. U.S. Patent 10,933,076, 2 March 2021. [Google Scholar]
- Amin, A.; Al Mansoori, A.; Baig, B. Safranal-Sorafenib Combination Therapy for Liver Cancer. U.S. Patent 10,568,873, 25 February 2020. [Google Scholar]
- Al-Hrout, A.; Chaiboonchoe, A.; Khraiwesh, B.; Murali, C.; Baig, B.; El-Awady, R.; Tarazi, H.; Alzahmi, A.; Nelson, D.R.; Greish, Y.E.; et al. Safranal induces DNA double-strand breakage and ER-stress-mediated cell death in hepatocellular carcinoma cells. Sci. Rep. 2018, 8, 1–15. [Google Scholar] [CrossRef] [Green Version]
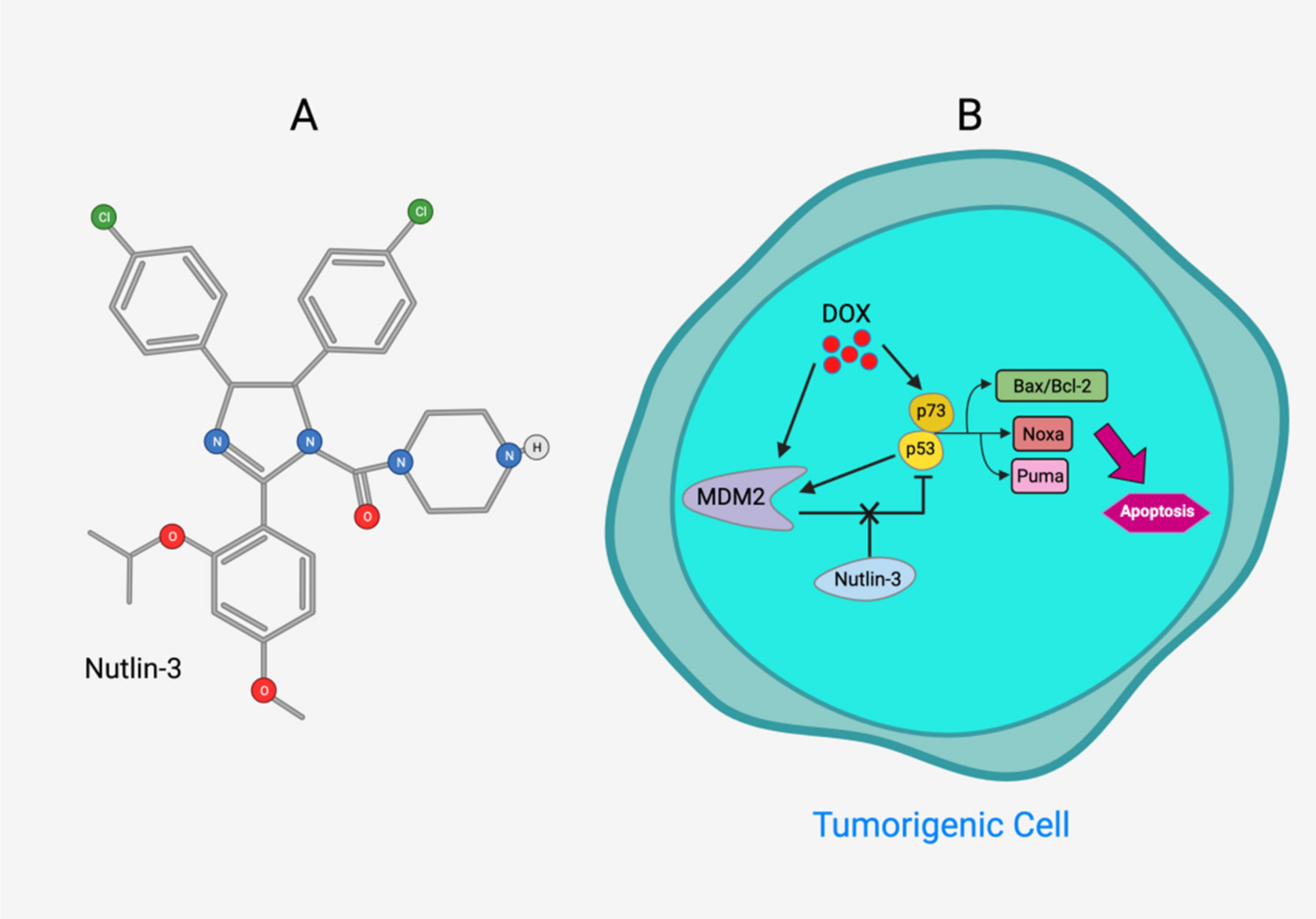
- Zheng, T.; Wang, J.; Song, X.; Meng, X.; Pan, S.; Jiang, H.; Liu, L. Nutlin-3 cooperates with doxorubicin to induce apoptosis of human hepatocellular carcinoma cells through p53 or p73 signaling pathways. J. Cancer Res. Clin. Oncol. 2010, 136, 1597–1604. [Google Scholar] [CrossRef]
- Shimizu, M.; Shirakami, Y.; Sakai, H.; Tatebe, H.; Nakagawa, T.; Hara, Y.; Weinstein, I.B.; Moriwaki, H. EGCG inhibits acti-vation of the insulin-like growth factor (IGF)/IGF-1 receptor axis in human hepatocellular carcinoma cells. Cancer Lett. 2008, 262, 10–18. [Google Scholar] [CrossRef]
- Shao, J.; Meng, Q.; Li, Y. Theaflavins suppress tumor growth and metastasis via the blockage of the STAT3 pathway in hepatocellular carcinoma. OncoTargets Ther. 2016, 9, 4265–4275. [Google Scholar] [CrossRef] [Green Version]
- Banjerdpongchai, R.; Wudtiwai, B.; Khaw-On, P.; Rachakhom, W.; Duangnil, N.; Kongtawelert, P. Hesperidin from citrus seed induces human hepatocellular carcinoma HepG2 cell apoptosis via both mitochondrial and death receptor pathways. Tumor Biol. 2016, 37, 227–237. [Google Scholar] [CrossRef] [Green Version]
- Shu, G.; Yang, J.; Zhao, W.; Xu, C.; Hong, Z.; Mei, Z.; Yang, X. Kurarinol induces hepatocellular carcinoma cell apoptosis through suppressing cellular signal transducer and activator of transcription 3 signaling. Toxicol. Appl. Pharmacol. 2014, 281, 157–165. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.-T.; Lin, G.-B.; Lin, S.-H.; Lu, C.-H.; Hsieh, C.-H.; Ma, B.-L.; Chao, C.-Y. Static magnetic field enhances the anticancer efficacy of capsaicin on HepG2 cells via capsaicin receptor TRPV1. PLoS ONE 2018, 13, e0191078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, Y.-J.; Lee, E.-J.; Kim, H.-R.; Hwang, K.-A. Molecular mechanisms of luteolin-7-O-glucoside-induced growth inhibition on human liver cancer cells: G2/M cell cycle arrest and caspase-independent apoptotic signaling pathways. BMB Rep. 2013, 46, 611–616. [Google Scholar] [CrossRef] [Green Version]
- Lin, Z.; Carvalho, B.R.; Kahn, E.; Lv, R.; Rao, R.; Terrones, H.; A Pimenta, M.; Terrones, M. Defect engineering of two-dimensional transition metal dichalcogenides. 2D Mater. 2016, 3, 022002. [Google Scholar] [CrossRef]
- Huang, C.; Wei, Y.-X.; Shen, M.-C.; Tu, Y.-H.; Wang, C.-C.; Huang, H.-C. Chrysin, Abundant in Morinda citrifolia Fruit Water–EtOAc Extracts, Combined with Apigenin Synergistically Induced Apoptosis and Inhibited Migration in Human Breast and Liver Cancer Cells. J. Agric. Food Chem. 2016, 64, 4235–4245. [Google Scholar] [CrossRef]
- Zou, M.; Lu, N.; Hu, C.; Liu, W.; Sun, Y.; Wang, X.; You, Q.; Gu, C.; Xi, T.; Guo, Q. Beclin 1-mediated autophagy in hepato-cellular carcinoma cells: Implication in anticancer efficiency of oroxylin A via inhibition of mTOR signaling. Cell. Signal. 2012, 24, 1722–1732. [Google Scholar] [CrossRef]
- Xu, M.; Lu, N.; Zhang, H.; Dai, Q.; Wei, L.; Li, Z.; You, Q.; Guo, Q. Wogonin induced cytotoxicity in human hepatocellular carcinoma cells by activation of unfolded protein response and inactivation of AKT. Hepatol. Res. 2013, 43, 890–905. [Google Scholar] [CrossRef]
- Liang, R.R.; Zhang, S.; Qi, J.A.; Wang, Z.D.; Li, J.; Liu, P.J.; Huang, C.; Le, X.F.; Yang, J.; Li, Z.F. Preferential inhibition of hepatocellular carcinoma by the flavonoid Baicalein through blocking MEK-ERK signaling. Int. J. Oncol. 2012, 41, 969–978. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.-F.; Li, T.; Tang, Z.-H.; Chang, L.-L.; Zhu, H.; Chen, X.-P.; Wang, Y.-T.; Lu, J.-J. Baicalein triggers autophagy and inhibits the protein kinase B/Mammalian target of rapamycin pathway in hepatocellular carcinoma HepG2 Cells. Phytother. Res. 2015, 29, 674–679. [Google Scholar] [CrossRef]
- Wang, F.; Wang, Y.-H.; Wang, J.-J.; Xu, H.-L.; Wang, C.-M. Eriodictyol-induced anti-cancer and apoptotic effects in human hepatocellular carcinoma cells are associated with cell cycle arrest and modulation of apoptosis-related proteins. Bangladesh J. Pharmacol. 2016, 11, 285–291. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Song, J.; Wu, D.; Wang, J.; Dong, W. Hesperetin induces the apoptosis of hepatocellular carcinoma cells via mito-chondrial pathway mediated by the increased intracellular reactive oxygen species, ATP and calcium. Med. Oncol. 2015, 32, 101. [Google Scholar] [CrossRef]
- Su, L.; Chen, X.; Wu, J.; Lin, B.; Zhang, H.; Lan, L.; Luo, H. Galangin inhibits proliferation of hepatocellular carcinoma cells by inducing endoplasmic reticulum stress. Food Chem. Toxicol. 2013, 62, 810–816. [Google Scholar] [CrossRef]
- Zhang, H.-T.; Luo, H.; Wu, J.; Lan, L.-B.; Fan, D.-H.; Zhu, K.-D.; Chen, X.-Y.; Wen, M.; Liu, H.-M. Galangin induces apoptosis of hepatocellular carcinoma cells via the mitochondrial pathway. World J. Gastroenterol. 2010, 16, 3377. [Google Scholar] [CrossRef]
- Nishikawa, T.; Nakajima, T.; Moriguchi, M.; Jo, M.; Sekoguchi, S.; Ishii, M.; Takashima, H.; Katagishi, T.; Kimura, H.; Minami, M.; et al. A green tea polyphenol, epigalocatechin-3-gallate, induces apoptosis of human hepatocellular carcinoma, possibly through inhibition of Bcl-2 family proteins. J. Hepatol. 2006, 44, 1074–1082. [Google Scholar] [CrossRef]
- Huang, X.; Lian, T.; Guan, X.; Liu, B.; Hao, S.; Zhang, J.; Bao, S.; Tan, X.; Zhu, R.; Song, Z. Dihydromyricetin reduces TGF-β via P53 activation-dependent mechanism in hepatocellular carcinoma HepG2 cells. Protein Pept. Lett. 2017, 24, 419–424. [Google Scholar] [CrossRef]
- Wang, Z.; Zhang, H.; Zhou, J.; Zhang, X.; Chen, L.; Chen, K.; Huang, Z. Eriocitrin from lemon suppresses the proliferation of human hepatocellular carcinoma cells through inducing apoptosis and arresting cell cycle. Cancer Chemother. Pharmacol. 2016, 78, 1143–1150. [Google Scholar] [CrossRef]
- Youns, M.; Hegazy, W.A.H. The natural flavonoid fisetin inhibits cellular proliferation of hepatic, colorectal, and pancreatic cancer cells through modulation of multiple signaling pathways. PLoS ONE 2017, 12, e0169335. [Google Scholar] [CrossRef]
- Mylonis, I.; Lakka, A.; Tsakalof, A.; Simos, G. The dietary flavonoid kaempferol effectively inhibits HIF-1 activity and hepatoma cancer cell viability under hypoxic conditions. Biochem. Biophys. Res. Commun. 2010, 398, 74–78. [Google Scholar] [CrossRef]
- Liu, C.; Yang, S.; Wang, K.; Bao, X.; Liu, Y.; Zhou, S.; Liu, H.; Qiu, Y.; Wang, T.; Yu, H. Alkaloids from traditional Chinese medicine against hepatocellular carcinoma. Biomed. Pharmacother. 2019, 120, 109543. [Google Scholar] [CrossRef]
- Shi, L.; Yang, F.; Luo, F.; Liu, Y.; Zhang, F.; Zou, M.; Liu, Q. Evodiamine exerts anti-tumor effects against hepatocellular car-cinoma through inhibiting β-catenin-mediated angiogenesis. Tumor Biol. 2016, 37, 12791–12803. [Google Scholar] [CrossRef]
- Li, C.; Cai, G.; Song, D.; Gao, R.; Teng, P.; Zhou, L.; Ji, Q.; Sui, H.; Cai, J.; Li, Q.; et al. Development of EGFR-targeted evodiamine nanoparticles for the treatment of colorectal cancer. Biomater. Sci. 2019, 7, 3627–3639. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Xu, K.; Jiang, R.; Li, D.-Y.; Guo, X.-X.; Zhou, P.; Tang, J.-F.; Li, L.-S.; Zeng, D.; Hu, L.; et al. Evodiamine inhibits proliferation and promotes apoptosis of hepatocellular carcinoma cells via the Hippo-Yes-Associated Protein signaling pathway. Life Sci. 2020, 251, 117424. [Google Scholar] [CrossRef] [PubMed]
- Wiltberger, G.; Wu, Y.; Lange, U.; Hau, H.-M.; Tapper, E.; Krenzien, F.; Atanasov, G.; Benzing, C.; Feldbrügge, L.; Csizmadia, E.; et al. Protective effects of coffee consumption following liver transplantation for hepatocellular carcinoma in cirrhosis. Aliment. Pharmacol. Ther. 2019, 49, 779–788. [Google Scholar] [CrossRef] [PubMed]
- Tong, D.; Qu, H.; Meng, X.; Jiang, Y.; Liu, D.; Ye, S.; Chen, H.; Jin, Y.; Fu, S.; Geng, J. S-allylmercaptocysteine promotes MAPK inhibitor-induced apoptosis by activating the TGF-β signaling pathway in cancer cells. Oncol. Rep. 2014, 32, 1124–1132. [Google Scholar] [CrossRef]
- Xiao, J.; Xing, F.; Liu, Y.; Lv, Y.; Wang, X.; Ling, M.-T.; Gao, H.; Ouyang, S.; Yang, M.; Zhu, J.; et al. Garlic-derived compound S-allylmercaptocysteine inhibits hepatocarcinogenesis through targeting LRP6/Wnt pathway. Acta Pharm. Sin. B 2018, 8, 575–586. [Google Scholar] [CrossRef]
- Amin, A.; Hamza, A.A.; Daoud, S.; Khazanehdari, K.; Hrout, A.A.; Baig, B.; Chaiboonchoe, A.; Adrian, T.E.; Zaki, N.; Salehi-Ashtiani, K. Saffron-based crocin prevents early lesions of liver cancer: In vivo, in vitro and network analyses. Recent Pat. Anti-Cancer Drug Discov. 2016, 11, 121–133. [Google Scholar] [CrossRef]
- El-Kharrag, R.; Amin, A.; Hisaindee, S.; Greish, Y.; Karam, S.M. Development of a therapeutic model of precancerous liver using crocin-coated magnetite nanoparticles. Int. J. Oncol. 2017, 50, 212–222. [Google Scholar] [CrossRef] [Green Version]
- Kim, B.; Park, B. Saffron carotenoids inhibit STAT3 activation and promote apoptotic progression in IL-6-stimulated liver cancer cells. Oncol. Rep. 2018, 39, 1883–1891. [Google Scholar] [CrossRef] [Green Version]
- Yao, C.; Liu, B.-B.; Qian, X.-D.; Li, L.-Q.; Cao, H.-B.; Guo, Q.-S.; Zhou, G.-F. Crocin induces autophagic apoptosis in hepato-cellular carcinoma by inhibiting Akt/mTOR activity. Onco Targets Ther. 2018, 11, 2017. [Google Scholar] [CrossRef]
- Sawant, A.V.; Srivastava, S.; Prassanawar, S.S.; Bhattacharyya, B.; Panda, D. Crocin, a carotenoid, suppresses spindle mi-crotubule dynamics and activates the mitotic checkpoint by binding to tubulin. Biochem. Pharmacol. 2019, 163, 32–45. [Google Scholar] [CrossRef]
- Amin, A. Prevention of Liver Cancer with Safranal-Based Formulations. U.S. Patent 10,912,741, 9 February 2021. [Google Scholar]
- Noureini, S.K.; Wink, M. Antiproliferative effects of crocin in HepG2 Cells by telomerase inhibition and hTERT down-regulation. Asian Pac. J. Cancer Prev. 2012, 13, 2305–2309. [Google Scholar] [CrossRef] [Green Version]
- Tharehalli, U.; Günes, C.; Lechel, A. Telomeres and telomerase in the development of liver cancer. Cancers 2020, 12, 2048. [Google Scholar] [CrossRef]
- Amin, A.; Bajbouj, K.; Koch, A.; Gandesiri, M.; Schneider-Stock, R. Defective autophagosome formation in p53-null colorectal cancer reinforces crocin-induced apoptosis. Int. J. Mol. Sci. 2015, 16, 1544–1561. [Google Scholar] [CrossRef] [Green Version]
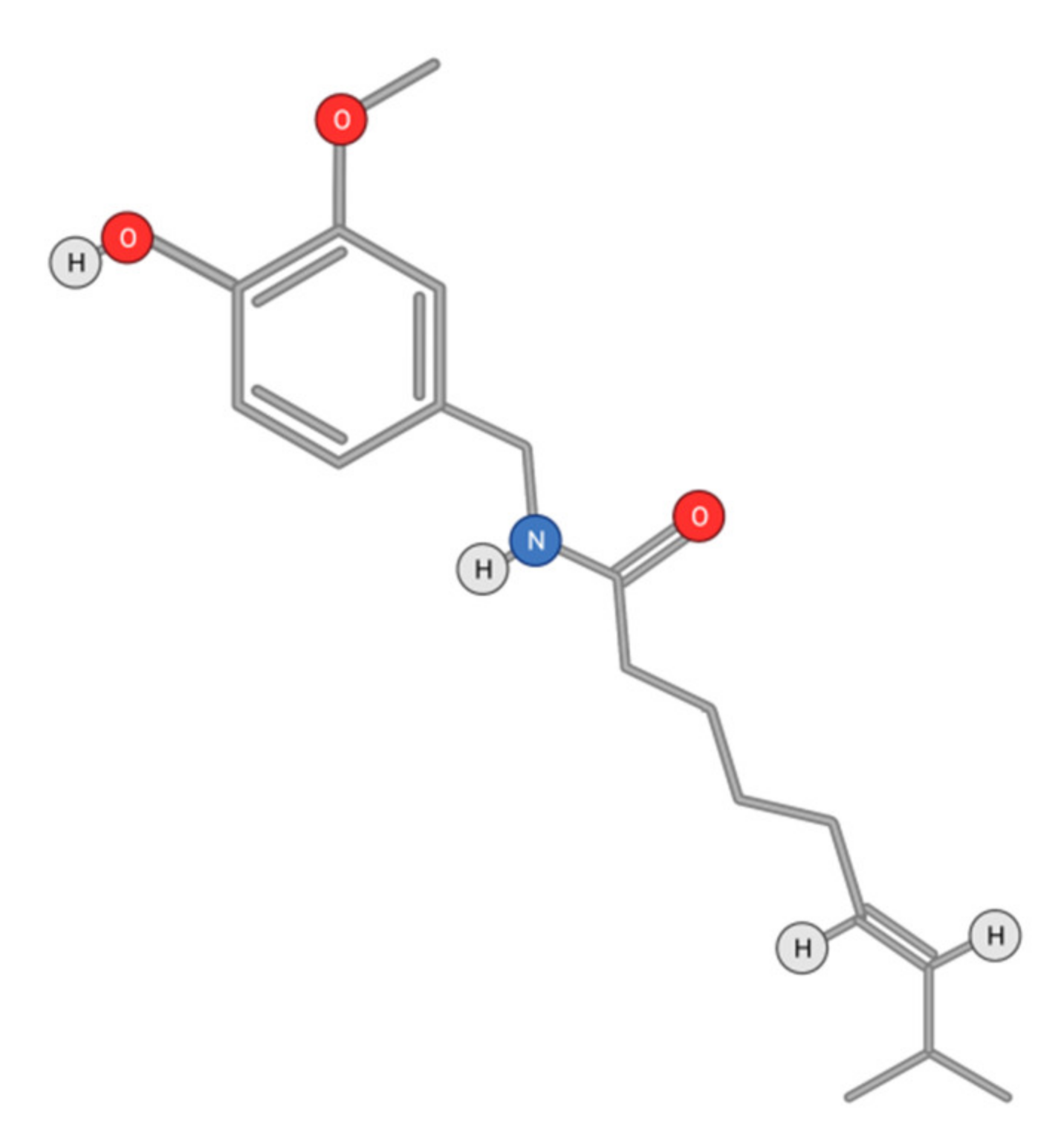
- Scheau, C.; Badarau, I.A.; Caruntu, C.; Mihai, G.L.; Didilescu, A.C.; Constantin, C.; Neagu, M. Capsaicin: Effects on the pathogenesis of hepatocellular carcinoma. Molecules 2019, 24, 2350. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.-T.; Wang, H.-C.; Hsu, Y.-C.; Cho, C.-L.; Yang, M.-Y.; Chien, C.-Y. Capsaicin Induces Autophagy and Apoptosis in Human Nasopharyngeal Carcinoma Cells by Downregulating the PI3K/AKT/mTOR Pathway. Int. J. Mol. Sci. 2017, 18, 1343. [Google Scholar] [CrossRef] [Green Version]
- Min, J.-K.; Han, K.-Y.; Kim, E.-C.; Kim, Y.-M.; Lee, S.-W.; Kim, O.-H.; Kim, K.-W.; Gho, Y.S.; Kwon, Y.-G. Capsaicin inhibits in vitro and in vivo angiogenesis. Cancer Res. 2004, 64, 644–651. [Google Scholar] [CrossRef] [Green Version]
- Kato, K.; Ogura, T.; Kishimoto, A.; Minegishi, Y.; Nakajima, N.; Miyazaki, M.; Esumi, H. Critical roles of AMP-activated protein kinase in constitutive tolerance of cancer cells to nutrient deprivation and tumor formation. Oncogene 2002, 21, 6082–6090. [Google Scholar] [CrossRef] [Green Version]
- Fattori, V.; Hohmann, M.S.; Rossaneis, A.C.; Pinho-Ribeiro, F.A.; Verri, W.A. Capsaicin: Current understanding of its mecha-nisms and therapy of pain and other pre-clinical and clinical uses. Molecules 2016, 21, 844. [Google Scholar] [CrossRef] [Green Version]
- Bort, A.; Sánchez, B.G.; Spínola, E.; Mateos-Gomez, P.A.; Rodríguez-Henche, N.; Díaz-Laviada, I. The red pepper’s spicy ingredient capsaicin activates AMPK in HepG2 cells through CaMKKβ. PLoS ONE 2019, 14, e0211420. [Google Scholar] [CrossRef] [Green Version]
- Bort, A.; Spínola, E.; Rodríguez-Henche, N.; Díaz-Laviada, I. Capsaicin exerts synergistic antitumor effect with sorafenib in hepatocellular carcinoma cells through AMPK activation. Oncotarget 2017, 8, 87684–87698. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Wang, L.; Wang, R.; Luo, X.; Li, Y.; Li, J.; Li, Y.; Chen, Z. Phenolic contents, cellular antioxidant activity and anti-proliferative capacity of different varieties of oats. Food Chem. 2018, 239, 260–267. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.S.; Ni, Y.H.; Zhao, C.R.; Qiao, Z.; Yu, H.X.; Wang, L.Y.; Sun, J.Y.; Du, C.; Zhang, J.H.; Dong, L.Y.; et al. Capsaicin en-hances the antitumor activity of sorafenib in hepatocellular carcinoma cells and mouse xenograft tumors through increased ERK signaling. Acta Pharmacol. Sin. 2018, 39, 438–448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Q.; Li, L.; Wang, F.; Chen, J.; Zhao, Y.; Wang, P.; Nilius, B.; Liu, D.; Zhu, Z. Dietary capsaicin prevents nonalcoholic fatty liver disease through transient receptor potential vanilloid 1-mediated peroxisome proliferator-activated receptor δ activation. Pflügers Arch. Eur. J. Physiol. 2013, 465, 1303–1316. [Google Scholar] [CrossRef] [PubMed]
- Munson, M.J.; Ganley, I.G. MTOR, PIK3C3, and autophagy: Signaling the beginning from the end. Autophagy 2015, 11, 2375–2376. [Google Scholar] [CrossRef]
- Chen, X.; Tan, M.; Xie, Z.-Q.; Feng, B.; Zhao, Z.; Yang, K.; Hu, C.; Liao, N.; Wang, T.; Chen, D.; et al. Inhibiting ROS-STAT3-dependent autophagy enhanced capsaicin–induced apoptosis in human hepatocellular carcinoma cells. Free Radic. Res. 2016, 50, 744–755. [Google Scholar] [CrossRef]
- Jung, M.-Y.; Kang, H.-J.; Moon, A. Capsaicin-induced apoptosis in SK-Hep-1 hepatocarcinoma cells involves Bcl-2 downreg-ulation and caspase-3 activation. Cancer Lett. 2001, 165, 139–145. [Google Scholar] [CrossRef]
- Huang, S.-P.; Chen, J.-C.; Wu, C.-C.; Chen, C.-T.; Tang, N.-Y.; Ho, Y.-T.; Lo, C.; Lin, J.-P.; Chung, J.-G.; Lin, J.-G. Capsaicin-induced apoptosis in human hepatoma HepG2 cells. Anticancer Res. 2009, 29, 165–174. [Google Scholar]
- Kim, J.-A.; Kang, Y.S.; Lee, Y.S. A phospholipase C-dependent intracellular Ca 2+ release pathway mediates the capsai-cin-induced apoptosis in HepG2 human hepatoma cells. Arch. Pharmacal. Res. 2005, 28, 73–80. [Google Scholar] [CrossRef]
- Waterhouse, N.J.; Goldstein, J.C.; Von Ahsen, O.; Schuler, M.; Newmeyer, D.D.; Green, D.R. Cytochrome c maintains mito-chondrial transmembrane potential and ATP generation after outer mitochondrial membrane permeabilization during the apoptotic process. J. Cell Biol. 2001, 153, 319–328. [Google Scholar] [CrossRef]
- Brenner, C.; Kroemer, G. Mitochondria—the death signal integrators. Science 2000, 289, 1150–1151. [Google Scholar] [CrossRef]
- Kapetanovic, I. Computer-aided drug discovery and development (CADDD): In silico-chemico-biological approach. Chem. -Biol. Interact. 2008, 171, 165–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mabrouk, M.S. Discovering best candidates for Hepatocellular Carcinoma (HCC) by in-silico techniques and tools. Int. J. Bioinform. Res. Appl. 2012, 8, 141–152. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, G.S.; Rajagopal, V.; Joseph, S.R.A.; Sebastian, D.; Savarimuthu, I.; Selvaraj, K.R.N.; Thobias, A.F. In vitro, in silico and in vivo antitumor activity of crude methanolic extract of tetilla dactyloidea (Carter, 1869) on DEN Induced HCC in a Rat Model. Biomed. Pharmacother. 2017, 95, 795–807. [Google Scholar] [CrossRef] [PubMed]
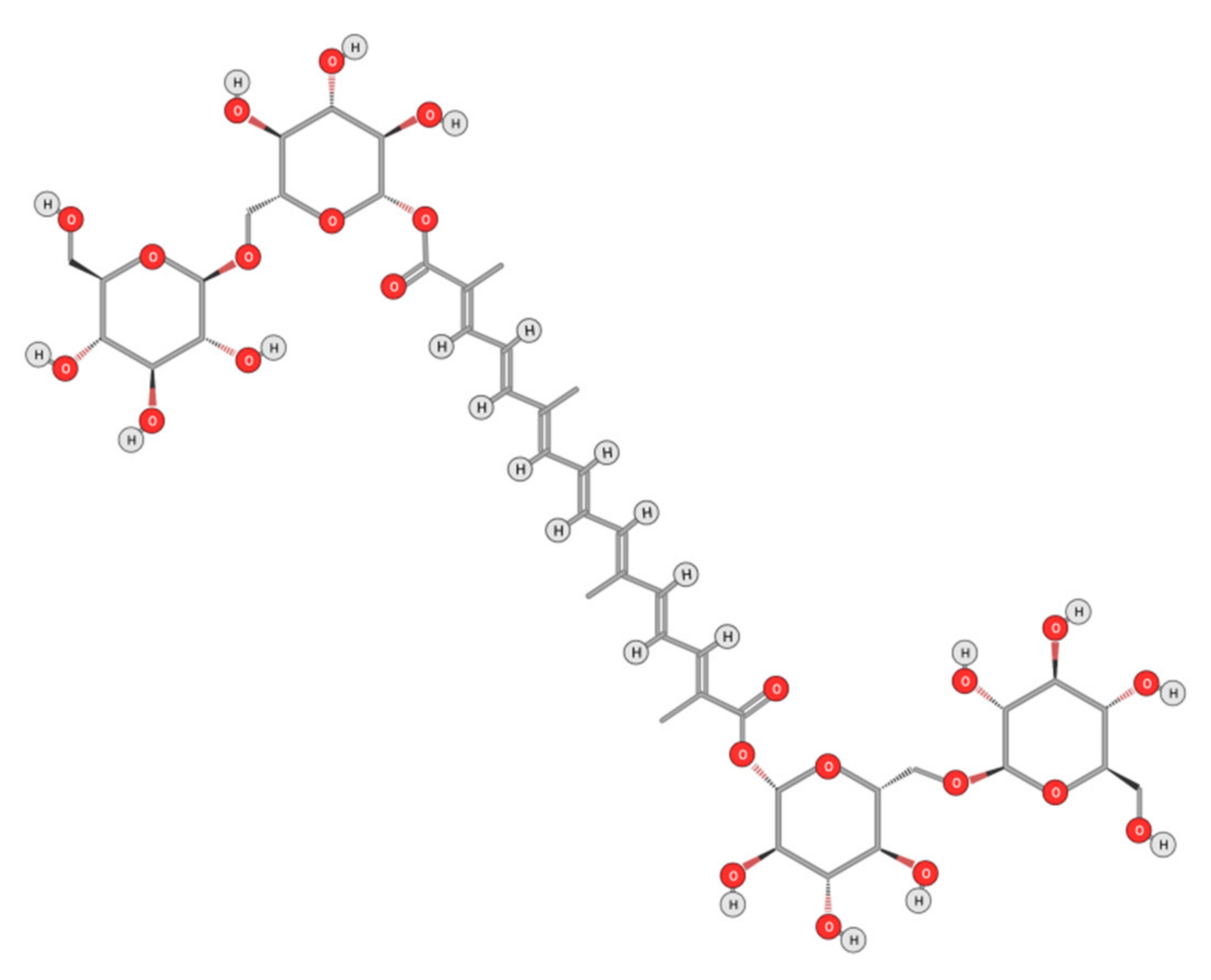
- Wang, W.; Xiong, X.; Li, X.; Zhang, Q.; Yang, W.; Du, L. In silico investigation of the anti-tumor mechanisms of epigallocate-chin-3-gallate. Molecules 2019, 24, 1445. [Google Scholar] [CrossRef] [Green Version]
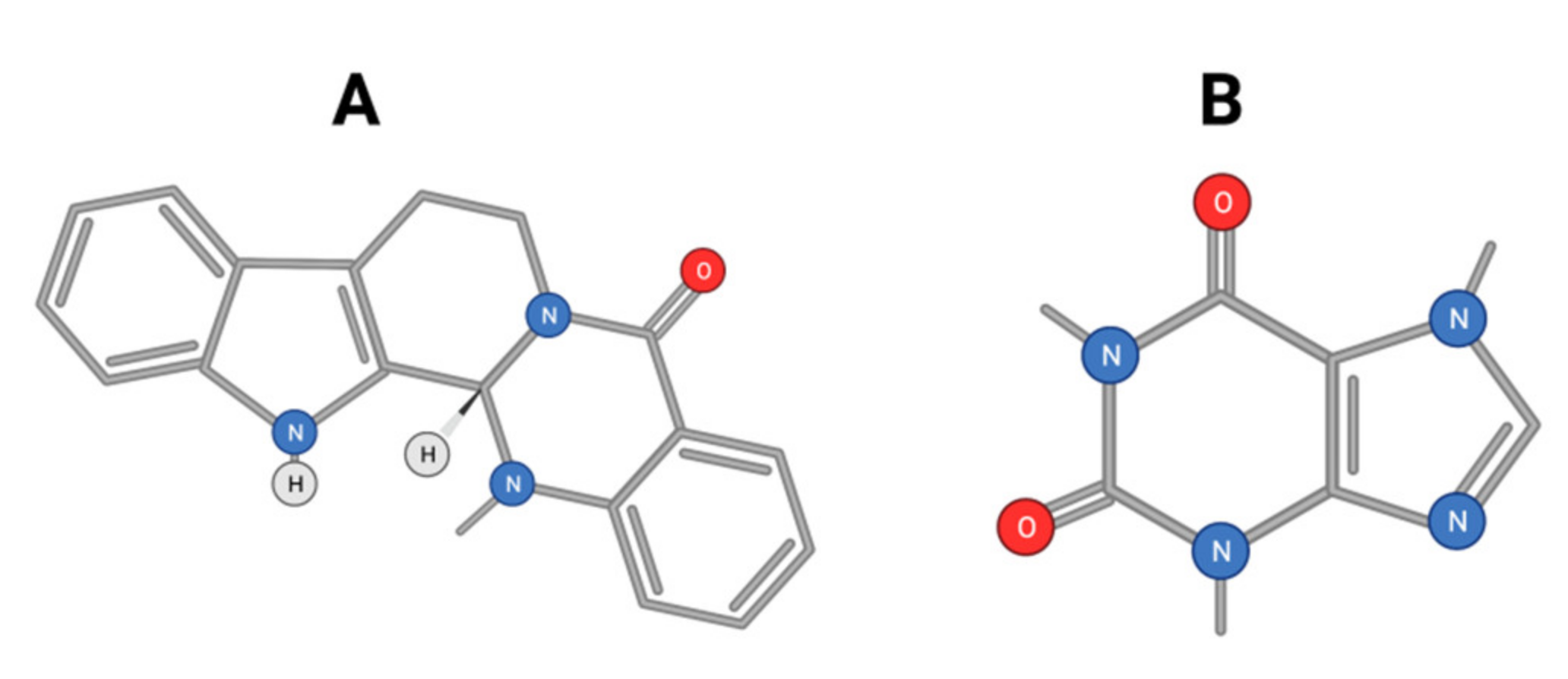
- Song, L.; Luo, Y.; Wang, X.; Almutairi, M.M.; Pan, H.; Li, W.; Liu, Y.; Wang, Q.; Hong, M. Exploring the active mechanism of berberine against HCC by systematic pharmacology and experimental validation. Mol. Med. Rep. 2019, 20, 4654–4664. [Google Scholar] [CrossRef] [Green Version]
- Lagoa, R.; Silva, J.; Rodrigues, J.R.; Bishayee, A. Advances in phytochemical delivery systems for improved anticancer activity. Biotechnol. Adv. 2020, 38, 107382. [Google Scholar] [CrossRef]
- Zi, C.-T.; Wang, Z.-H.; Shi, J.; Shi, B.-Y.; Zhang, N.; Wu, Y.-L.; Xie, Y.-R.; Zhou, L.; Xiao, C.; Wang, X.-J.; et al. Synthesis, cytotoxicity, and molecular docking of methylated (–)-epigallocatechin-3-gallate-4β-triazolopodophyllotoxin derivatives as novel antitumor agents. J. Chem. Res. 2021, 17475198211027328. [Google Scholar] [CrossRef]
















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Drug | Targets | Descriptions | Reference |
|---|---|---|---|
Cediranib  | VEGFR | Shows high toxicity and is ineffective for patients with unresectable or metastatic HCC | [30] |
Dovitinib  | c-KIT, Flt-3, FGFR, VEGFR | Significantly prolongs survival and inhibits primary tumour growth and lung metastasis in HCC xenograft models | [31,32] |
Erlotinib  | EGFR | Shows modest prolonged progression-free survival and overall survival in patients with unresectable HCC | [33,34] |
Gefitinib  | EGFR | Inhibits the tumor growth of HCC xenografts in a mouse model | [35] |
Selumetinib  | MEK1 | Suppresses tumour growth of HCC xenografts in mouse model Shows inadequate antitumour activity with short progression-free survival in patients with locally advanced or metastatic HCC | [36,37] |
Brivanib  | FGFR, VEGFR | Increases apoptosis, reduces microvessel density, and decreases VEGFR phosphorylation Shows promising antitumour activity in patients with advanced HCC | [28,29] |
Linifanib  | PDGFR, VEGFR | Inhibits tumour growth of HCC xenografts in the mouse model Shows similar overall survival to sorafenib in patients with advanced HCC | [38,39,40] |
Sunitinib  | c-Kit, Flt-3, PDGFP, VEGFR | Increases apoptosis and reduces microvessel density of HCC xenografts Displays poor overall survival in patients with advanced HCC and has severe toxicity | [41] |
Orantinib  | FGFR, PDGFR, VEGFR | Suppresses the tumour growth of subcutaneously co-injected HCC cell line xenografts Shows no improvement in overall survival in patients with unresectable HCC | [42,43] |
| Bevacizumab | VEGF | Inhibits tumour growth of HCC cell lines or patient-derived HCC xenografts | [44,45] |
| Cetuximab | EGFR | Shows no obvious response in patients with advanced HCC | [46] |
| Compound | Main Origin and Structure | Signaling Pathway | HCC Model | Reference |
|---|---|---|---|---|
Luteolin  | Celery, green pepper, parsley, thyme, dandelion | ROS-mediated pathway Caspase Activation | Hep G2 cells | [114] |
Luteolin-7-O-glucoside  | Dandelion, coffee and Cynara scolymus | Arrest G2/M phases of cell by JNK and Caspase activation | HepG2 cells | [115] |
Isoorientin  | Passion flower, Vitex negundo, Terminalia myriocarpa | Regulation of Cell cycle-related genes ROS-mediated pathway Caspase-3 and caspase-9 activation | Hep G2 cells | [116] |
Chrysin  | Honey, propolis, passion flowers, and Passiflora caerulea | Downregulation of Skp2 and LRP6 Activation of the p53/Bcl-2/caspase-9 | Hep G2 cells | [117] |
Oroxylin A  | Scutellaria baicalensis and Oroxylum indicum | Suppression of PI3K-PTEN-Akt-mTOR signaling pathway Activation of the ERK-eIF2α-ATF4-CHOP branch of the UPR pathway | HepG2 cells | [118] |
Wogonin  | Scutellaria baicalensis | Activation of the UPR pathway and inactivation of Akt signaling | HepG2, SMMC7721, and Hep3B cells | [119] |
Baicalein  | Roots of Scutellaria baicalensis and Scutellaria lateriflora | Inhibition of the PKB/mTOR pathway Blocking MEK-ERK signaling | HepG2 cells Xenograft in mice | [120,121] |
Eriodictyol  | Eriodictyon californicum | Upregulation of Bax and PARP downregulation of Bcl-2 | HepG2 cells | [122] |
Hesperidin  | Citrus fruits | Regulation of mitochondrial pathway Death receptor pathway; increases the levels of intracellular ROS, ATP, and Ca2+ | HepG2 cells Xenograft in mice | [112,123] |
Puerarin  | Root of Pueraria | Regulation of MAPK pathways | SMMC-7721 cells | [51] |
Galangin  | Alpinia officinarum and Helichrysum aureonitens | Pro apoptotic Mitochondrial pathway mediated by Bax | HepG2, HepG2, Hep3B, and PLC/PRF/5 cells | [124,125] |
Epigallocatechin gallate  | Tea leaves | Inhibition of tyrosine kinase receptors Downregulation of PI3K/Akt activity; downregulating Bcl-2 alpha and Bcl-xl by inactivation of NF-κB Hypoxia | SMMC7721, SKhep1, HLE, HepG2, HuH-7, PLC/PRF/5 cells; Xenograft in mice | [110,126] |
Dihydromyricetin  | Ampelopsis japonica, Hovenia dulcis | Reduction of TGFβ via p53- dependent pathway | HepG2 cells | [127] |
Kurarinol  | Roots of Sophora flavescens | Suppressing STAT3 signaling | HepG2, Huh-7, and H22 cells; Xenograft in mice | [113] |
Eriocitrin  | Lemon fruits | Upregulation of p53, cyclin A, cyclin D3, and CDK6 through activation of mitochondrial pathway | HepG2 cells | [128] |
Fisetin  | Strawberries, apples, persimmons, onions, and cucumbers | Regulation of CDK5 signaling NRF2-mediated oxidative stress response Glucocorticoid signaling ERK/MAPK signaling | HepG2 cells | [129] |
Kaempferol  | kale, beans, tea, spinach, and broccoli. | Inhibition of MAPK and HIF-1 | Huh7 cells | [130] |
Theaflavins  | Black tea | Activating the caspase pathwayBlockage of STAT3 pathway | HepG2 cells Xenograft in mice | [111] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Juaid, N.; Amin, A.; Abdalla, A.; Reese, K.; Alamri, Z.; Moulay, M.; Abdu, S.; Miled, N. Anti-Hepatocellular Carcinoma Biomolecules: Molecular Targets Insights. Int. J. Mol. Sci. 2021, 22, 10774. https://doi.org/10.3390/ijms221910774
Juaid N, Amin A, Abdalla A, Reese K, Alamri Z, Moulay M, Abdu S, Miled N. Anti-Hepatocellular Carcinoma Biomolecules: Molecular Targets Insights. International Journal of Molecular Sciences. 2021; 22(19):10774. https://doi.org/10.3390/ijms221910774
Chicago/Turabian StyleJuaid, Nouf, Amr Amin, Ali Abdalla, Kevin Reese, Zaenah Alamri, Mohamed Moulay, Suzan Abdu, and Nabil Miled. 2021. "Anti-Hepatocellular Carcinoma Biomolecules: Molecular Targets Insights" International Journal of Molecular Sciences 22, no. 19: 10774. https://doi.org/10.3390/ijms221910774