
Overview of Human HtrA Family Proteases and Their Distinctive Physiological Roles and Unique Involvement in Diseases, Especially Cancer and Pregnancy Complications


Abstract
:1. Introduction
2. Protein Domain Architecture, Tissue Distribution and Key Molecular Characteristics of Human HtrAs
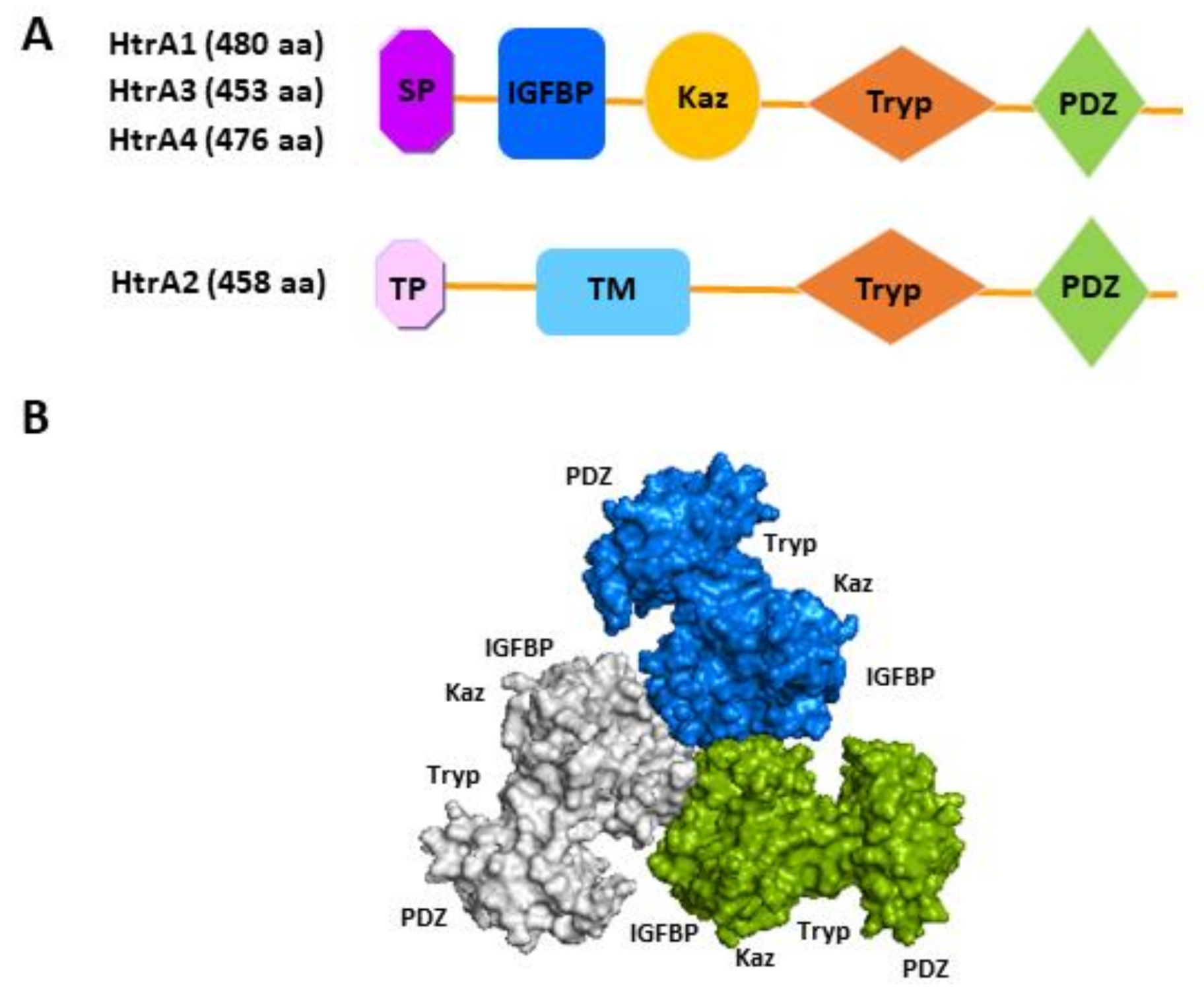
2.1. Domain Architecture
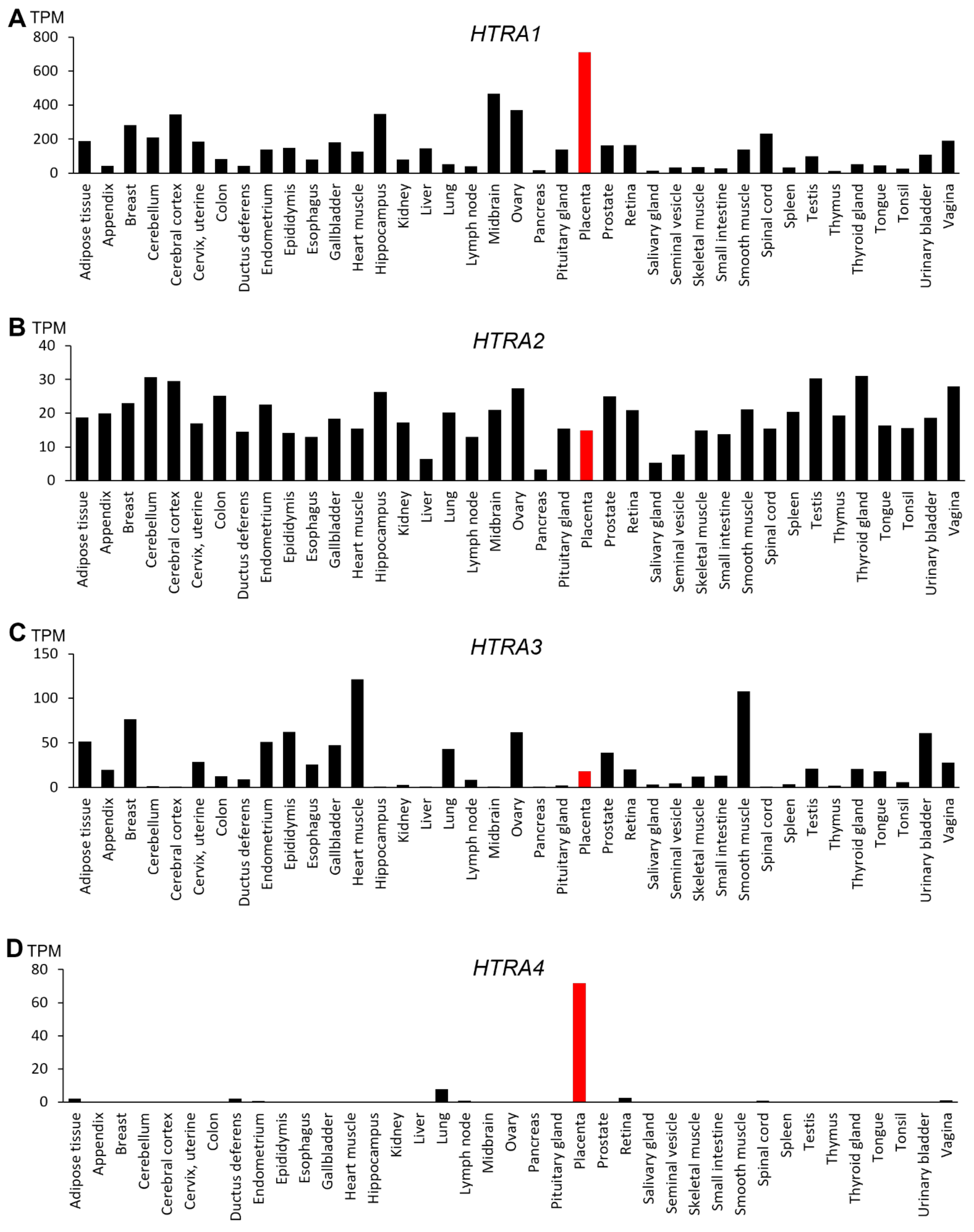
2.2. Tissue Distribution
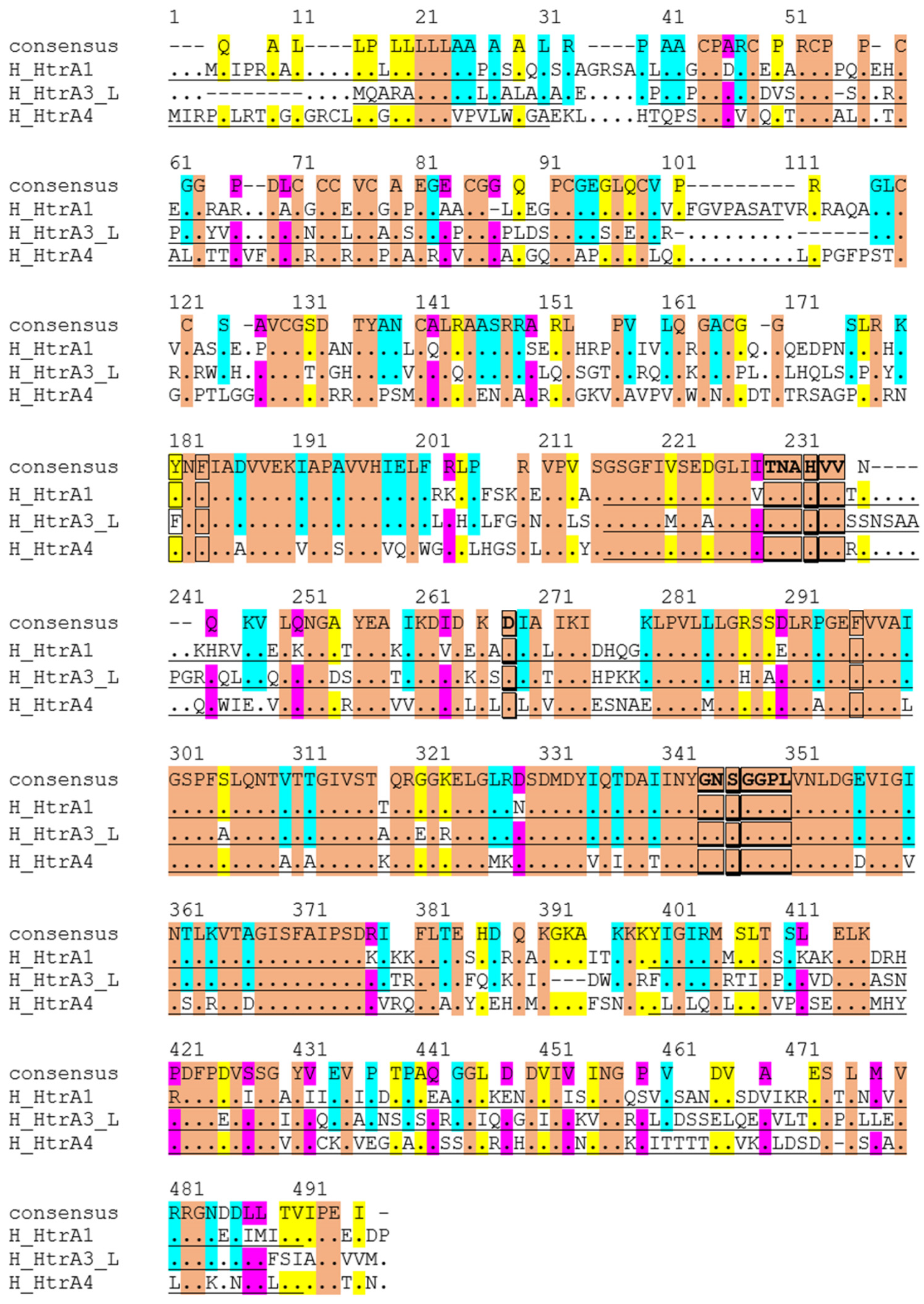
2.3. Key Molecular Characteristics
3. Function, Regulation, and Potential Substrates of Human HtrAs and Their Involvement of Various Diseases
3.1. HtrA1
3.2. HtrA2
3.3. HtrA3
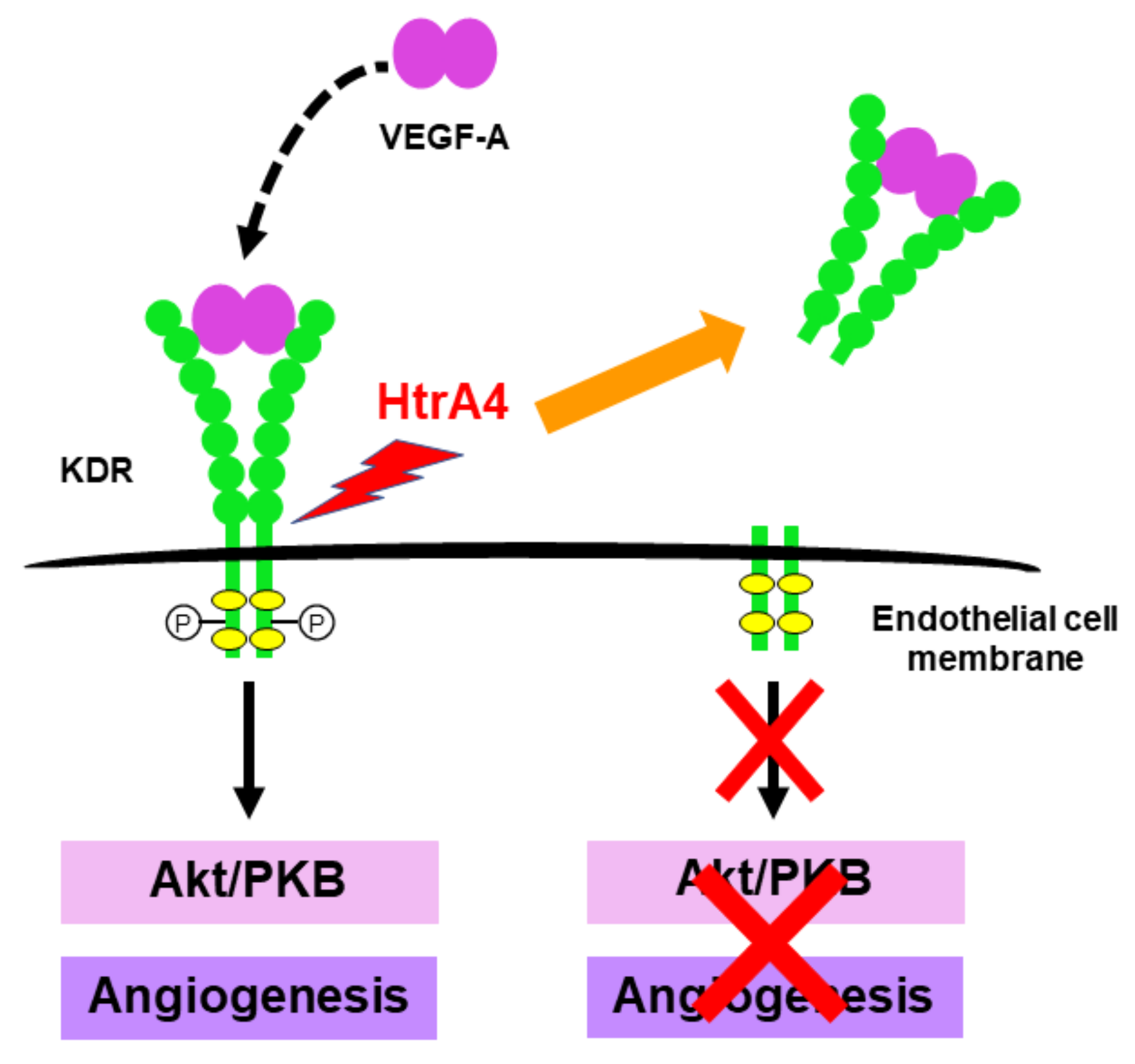
3.4. HtrA4
4. HtrAs and Various Cancers
4.1. HtrA1
4.2. HtrA2

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cancer Type | mRNA Levels in FPKM | |||
|---|---|---|---|---|
| HTRA1 | HTRA2 | HTRA3 | HTRA4 | |
| Glioma (n = 153) | 192.4 | 12.2 | 2.3 | 0.1 |
| Thyroid cancer (n = 501) | 42.8 | 7.1 | 3.2 | 0.2 |
| Lung cancer (n = 994) | 36.2 | 8 | 15.9 | 0.5 |
| Colorectal cancer (n = 597) | 23.4 | 7.9 | 11.9 | 0.1 |
| Head and neck cancer (n = 499) | 58.3 | 7.5 | 12.6 | 0.1 |
| Stomach cancer (n = 354) | 34.2 | 5 | 14.2 | 0.2 |
| Liver cancer (n = 365) | 57.5 | 5.6 | 0.8 | 0 |
| Pancreatic cancer (n = 176) | 93.5 | 7.2 | 56.9 | 0.4 |
| Renal cancer (n = 877) | 79.6 | 8.4 | 1.2 | 0.4 |
| Urothelial cancer (n = 406) | 36.5 | 8.3 | 9.1 | 0.1 |
| Prostate cancer (n = 494) | 23.4 | 7.2 | 5.7 | 0.1 |
| Testis cancer (n = 134) | 10.4 | 7.8 | 3.3 | 0.6 |
| Breast cancer (n = 1075) | 102.4 | 7.6 | 24.5 | 0.2 |
| Cervical cancer (n = 291) | 32.2 | 7.7 | 5.3 | 0.2 |
| Endometrial cancer (n = 541) | 46.4 | 7.7 | 6.4 | 0.2 |
| Ovarian cancer (n = 373) | 47.8 | 9.1 | 4.3 | 0.1 |
| Melanoma (n = 102) | 26.8 | 9.5 | 3 | 0.1 |
4.3. HtrA3
| HtrA Member | Type of Cancer | Changes in Expression | References |
|---|---|---|---|
| HtrA1 | Ovarian cancer | ↓ | [19,92,99] |
| Metastatic melanomas | ↓ | [93] | |
| Breast carcinoma | ↓ | [94,118] | |
| Metastatic prostate cancer | ↓ | [95] | |
| Lung cancer | ↓ | [96] | |
| Endometrial cancer | ↓ | [20,97] | |
| Mesotheliomas | ↓ | [119] | |
| HtrA2 | Ovarian cancer | ↑ | [100] |
| Prostate cancer | Inconsistent | [102,104] | |
| Stomach cancer | ↑ | [103,105] | |
| Breast cancer | ↓ | [105] | |
| HtrA3 | Ovarian cancer | ↓ | [19,106,107] |
| Endometrial cancer | ↓ | [20,109] | |
| Breast cancer | ↓ | [111] | |
| Lung cancer | ↓ | [80,113] | |
| Oesophageal adenocarcinoma | ↑ | [114] | |
| Pancreatic adenocarcinoma | ↑ | [115] | |
| Testicular seminoma | ↑ | [116] | |
| Haematologic cancer | Variable | [117] | |
| HtrA4 | Not reported | Not reported |
4.4. HtrA4
5. HtrAs in Placental Development and Pregnancy Complications
5.1. HtrA1
5.2. HtrA3
5.3. HtrA4
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Clausen, T.; Southan, C.; Ehrmann, M. The HtrA family of proteases: Implications for protein composition and cell fate. Mol. Cell 2002, 10, 443–455. [Google Scholar] [CrossRef]
- Clausen, T.; Kaiser, M.; Huber, R.; Ehrmann, M. HTRA proteases: Regulated proteolysis in protein quality control. Nat. Rev. Mol. Cell Biol. 2011, 12, 152–162. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.J.; Zheng, J.J. PDZ domains and their binding partners: Structure, specificity, and modification. Cell Commun. Signal. 2010, 8, 8. [Google Scholar] [CrossRef] [Green Version]
- Lipinska, B.; Zylicz, M.; Georgopoulos, C. The HtrA (DegP) protein, essential for Escherichia coli survival at high temperatures, is an endopeptidase. J. Bacteriol. 1990, 172, 1791–1797. [Google Scholar] [CrossRef] [Green Version]
- Chang, Z. The function of the DegP (HtrA) protein: Protease versus chaperone. IUBMB Life 2016, 68, 904–907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilken, C.; Kitzing, K.; Kurzbauer, R.; Ehrmann, M.; Clausen, T. Crystal structure of the DegS stress sensor: How a PDZ domain recognizes misfolded protein and activates a protease. Cell 2004, 117, 483–494. [Google Scholar] [CrossRef] [Green Version]
- Krojer, T.; Sawa, J.; Schafer, E.; Saibil, H.R.; Ehrmann, M.; Clausen, T. Structural basis for the regulated protease and chaperone function of DegP. Nature 2008, 453, 885–890. [Google Scholar] [CrossRef] [PubMed]
- Spiess, C.; Beil, A.; Ehrmann, M. A temperature-dependent switch from chaperone to protease in a widely conserved heat shock protein. Cell 1999, 97, 339–347. [Google Scholar] [CrossRef] [Green Version]
- Israeli, M.A.; Elia, U.; Rotem, S.; Cohen, H.; Tidhar, A.; Bercovich-Kinori, A.; Cohen, O.; Chitlaru, T. Distinct Contribution of the HtrA Protease and PDZ Domains to Its Function in Stress Resilience and Virulence of Bacillus anthracis. Front. Microbiol. 2019, 10, 255. [Google Scholar] [CrossRef] [PubMed]
- Ingmer, H.; Brondsted, L. Proteases in bacterial pathogenesis. Res. Microbiol. 2009, 160, 704–710. [Google Scholar] [CrossRef]
- Sgouras, D.; Tegtmeyer, N.; Wessler, S. Activity and Functional Importance of Helicobacter pylori Virulence Factors. Adv. Exp. Med. Biol. 2019, 1149, 35–56. [Google Scholar]
- Hansen, G.; Hilgenfeld, R. Architecture and regulation of HtrA-family proteins involved in protein quality control and stress response. Cell. Mol. Life Sci. CMLS 2013, 70, 761–775. [Google Scholar] [CrossRef] [PubMed]
- Zaide, G.; Elia, U.; Cohen-Gihon, I.; Israeli, M.A.; Rotem, S.; Israeli, O.; Ehrlich, S.; Cohen, H.; Lazar, S.; Beth-Din, A.; et al. Comparative Analysis of the Global Transcriptomic Response to Oxidative Stress of Bacillus anthracis htrA-Disrupted and Parental Wild Type Strains. Microorganisms 2020, 8, 1896. [Google Scholar] [CrossRef] [PubMed]
- Figaj, D.; Ambroziak, P.; Przepiora, T.; Skorko-Glonek, J. The Role of Proteases in the Virulence of Plant Pathogenic Bacteria. Int. J. Mol. Sci. 2019, 20, 672. [Google Scholar] [CrossRef] [Green Version]
- Backert, S.; Bernegger, S.; Skórko-Glonek, J.; Wessler, S. Extracellular HtrA serine proteases: An emerging new strategy in bacterial pathogenesis. Cell. Microbiol. 2018, 20, e12845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zurawa-Janicka, D.; Wenta, T.; Jarzab, M.; Skorko-Glonek, J.; Glaza, P.; Gieldon, A.; Ciarkowski, J.; Lipinska, B. Structural insights into the activation mechanisms of human HtrA serine proteases. Arch. Biochem. Biophys. 2017, 621, 6–23. [Google Scholar] [CrossRef]
- Zurawa-Janicka, D.; Skorko-Glonek, J.; Lipinska, B. HtrA proteins as targets in therapy of cancer and other diseases. Expert Opin. Ther. Targets 2010, 14, 665–679. [Google Scholar] [CrossRef] [PubMed]
- Welsh, J.B.; Zarrinkar, P.P.; Sapinoso, L.M.; Kern, S.G.; Behling, C.A.; Monk, B.J.; Lockhart, D.J.; Burger, R.A.; Hampton, G.M. Analysis of gene expression profiles in normal and neoplastic ovarian tissue samples identifies candidate molecular markers of epithelial ovarian cancer. Proc. Natl. Acad. Sci. USA 2001, 98, 1176–1181. [Google Scholar] [CrossRef] [Green Version]
- Narkiewicz, J.; Klasa-Mazurkiewicz, D.; Zurawa-Janicka, D.; Skorko-Glonek, J.; Emerich, J.; Lipinska, B. Changes in mRNA and protein levels of human HtrA1, HtrA2 and HtrA3 in ovarian cancer. Clin. Biochem. 2008, 41, 561–569. [Google Scholar] [CrossRef]
- Narkiewicz, J.; Lapinska-Szumczyk, S.; Zurawa-Janicka, D.; Skorko-Glonek, J.; Emerich, J.; Lipinska, B. Expression of human HtrA1, HtrA2, HtrA3 and TGF-beta1 genes in primary endometrial cancer. Oncol. Rep. 2009, 21, 1529–1537. [Google Scholar]
- Lorenzi, T.; Marzioni, D.; Giannubilo, S.; Quaranta, A.; Crescimanno, C.; De Luca, A.; Baldi, A.; Todros, T.; Tranquilli, A.L.; Castellucci, M. Expression patterns of two serine protease HtrA1 forms in human placentas complicated by preeclampsia with and without intrauterine growth restriction. Placenta 2009, 30, 35–40. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Puryer, M.; Lin, E.; Hale, K.; Salamonsen, L.A.; Manuelpillai, U.; Tong, S.; Chan, W.; Wallace, E.M.; Nie, G. Placental HtrA3 is regulated by oxygen tension and serum levels are altered during early pregnancy in women destined to develop preeclampsia. J. Clin. Endocrinol. Metab. 2011, 96, 403–411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teoh, S.S.; Zhao, M.; Wang, Y.; Chen, Q.; Nie, G. Serum HtrA1 is differentially regulated between early-onset and late-onset preeclampsia. Placenta 2015, 36, 990–995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, H.; Zhao, M.; Chen, Q.; Wang, Y.; Li, Y.; Kaitu’u-Lino, T.J.; Tong, S.; Nie, G. Human HtrA4 Expression Is Restricted to the Placenta, Is Significantly Up-Regulated in Early-Onset Preeclampsia, and High Levels of HtrA4 Cause Endothelial Dysfunction. J. Clin. Endocrinol. Metab. 2015, 100, E936–E945. [Google Scholar] [CrossRef] [Green Version]
- Singh, H.; Nero, T.L.; Wang, Y.; Parker, M.W.; Nie, G. Activity-modulating monoclonal antibodies to the human serine protease HtrA3 provide novel insights into regulating HtrA proteolytic activities. PLoS ONE 2014, 9, e108235. [Google Scholar] [CrossRef]
- Nie, G.Y.; Hampton, A.; Li, Y.; Findlay, J.K.; Salamonsen, L.A. Identification and cloning of two isoforms of human high-temperature requirement factor A3 (HtrA3), characterization of its genomic structure and comparison of its tissue distribution with HtrA1 and HtrA2. Biochem. J. 2003, 371 Pt 1, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Nie, G.Y.; Li, Y.; Minoura, H.; Batten, L.; Ooi, G.T.; Findlay, J.K.; Salamonsen, L.A. A novel serine protease of the mammalian HtrA family is up-regulated in mouse uterus coinciding with placentation. Mol. Hum. Reprod. 2003, 9, 279–290. [Google Scholar] [CrossRef] [Green Version]
- Bowden, M.; Drummond, A.E.; Salamonsen, L.A.; Findlay, J.K.; Nie, G. Evolutionary conservation of mammalian HTRA3 and its developmental regulation in the rat ovary. J. Exp. Zool. Part B Mol. Dev. Evol. 2009, 312, 701–713. [Google Scholar] [CrossRef] [PubMed]
- Eigenbrot, C.; Ultsch, M.; Lipari, M.T.; Moran, P.; Lin, S.J.; Ganesan, R.; Quan, C.; Tom, J.; Sandoval, W.; van Lookeren Campagne, M.; et al. Structural and functional analysis of HtrA1 and its subdomains. Structure 2012, 20, 1040–1050. [Google Scholar] [CrossRef] [Green Version]
- De Luca, A.; De Falco, M.; Severino, A.; Campioni, M.; Santini, D.; Baldi, F.; Paggi, M.G.; Baldi, A. Distribution of the serine protease HtrA1 in normal human tissues. J. Histochem. Cytochem. Off. J. Histochem. Soc. 2003, 51, 1279–1284. [Google Scholar] [CrossRef] [Green Version]
- De Luca, A.; De Falco, M.; Fedele, V.; Cobellis, L.; Mastrogiacomo, A.; Laforgia, V.; Tuduce, I.L.; Campioni, M.; Giraldi, D.; Paggi, M.G.; et al. The serine protease HtrA1 is upregulated in the human placenta during pregnancy. J. Histochem. Cytochem. Off. J. Histochem. Soc. 2004, 52, 885–892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nie, G.; Hale, K.; Li, Y.; Manuelpillai, U.; Wallace, E.M.; Salamonsen, L.A. Distinct expression and localization of serine protease HtrA1 in human endometrium and first-trimester placenta. Dev. Dyn. Off. Publ. Am. Assoc. Anat. 2006, 235, 3448–3455. [Google Scholar] [CrossRef]
- Nie, G.; Li, Y.; Hale, K.; Okada, H.; Manuelpillai, U.; Wallace, E.M.; Salamonsen, L.A. Serine peptidase HTRA3 is closely associated with human placental development and is elevated in pregnancy serum. Biol. Reprod. 2006, 74, 366–374. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Li, Y.; Hoh, J. Generation and characterization of mice with a conditional null allele of the HtrA4 gene. Mol. Med. Rep. 2015, 12, 6768–6774. [Google Scholar] [CrossRef] [Green Version]
- Inagaki, A.; Nishizawa, H.; Ota, S.; Suzuki, M.; Inuzuka, H.; Miyamura, H.; Sekiya, T.; Kurahashi, H.; Udagawa, Y. Upregulation of HtrA4 in the placentas of patients with severe pre-eclampsia. Placenta 2012, 33, 919–926. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Nie, G. High levels of HtrA4 observed in preeclamptic circulation drastically alter endothelial gene expression and induce inflammation in human umbilical vein endothelial cells. Placenta 2016, 47, 46–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, C.; Xing, F.; He, Y.; Zong, S.; Luo, C.; Li, C.; Duan, T.; Wang, K.; Zhou, Q. Elevated HTRA1 and HTRA4 in severe preeclampsia and their roles in trophoblast functions. Mol. Med. Rep. 2018, 18, 2937–2944. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Lim, R.; Nie, G. HtrA4 may play a major role in inhibiting endothelial repair in pregnancy complication preeclampsia. Sci. Rep. 2019, 9, 2728. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; La, M.; Pham, T.; Lovrecz, G.O.; Nie, G. High levels of HtrA4 detected in preeclamptic circulation may disrupt endothelial cell function by cleaving the main VEGFA receptor KDR. FASEB J. 2019, 33, 5058–5066. [Google Scholar] [CrossRef]
- Tseng, E.; Yee Teoh, S.S.; Wang, Y.; Nie, G. Elevated protease HtrA4 in the maternal circulation of preeclampsia may contribute to endothelial barrier disruption by cleaving key junctional protein VE-cadherin. Placenta 2019, 76, 51–53. [Google Scholar] [CrossRef] [PubMed]
- Zumbrunn, J.; Trueb, B. Primary structure of a putative serine protease specific for IGF-binding proteins. FEBS Lett. 1996, 398, 187–192. [Google Scholar] [CrossRef] [Green Version]
- Clawson, G.A.; Bui, V.; Xin, P.; Wang, N.; Pan, W. Intracellular localization of the tumor suppressor HtrA1/Prss11 and its association with HPV16 E6 and E7 proteins. J. Cell. Biochem. 2008, 105, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Grau, S.; Baldi, A.; Bussani, R.; Tian, X.; Stefanescu, R.; Przybylski, M.; Richards, P.; Jones, S.A.; Shridhar, V.; Clausen, T.; et al. Implications of the serine protease HtrA1 in amyloid precursor protein processing. Proc. Natl. Acad. Sci. USA 2005, 102, 6021–6026. [Google Scholar] [CrossRef] [Green Version]
- Grau, S.; Richards, P.J.; Kerr, B.; Hughes, C.; Caterson, B.; Williams, A.S.; Junker, U.; Jones, S.A.; Clausen, T.; Ehrmann, M. The role of human HtrA1 in arthritic disease. J. Biol. Chem. 2006, 281, 6124–6129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Z.; Camp, N.J.; Sun, H.; Tong, Z.; Gibbs, D.; Cameron, D.J.; Chen, H.; Zhao, Y.; Pearson, E.; Li, X.; et al. A variant of the HTRA1 gene increases susceptibility to age-related macular degeneration. Science 2006, 314, 992–993. [Google Scholar] [CrossRef]
- Hara, K.; Shiga, A.; Fukutake, T.; Nozaki, H.; Miyashita, A.; Yokoseki, A.; Kawata, H.; Koyama, A.; Arima, K.; Takahashi, T.; et al. Association of HTRA1 mutations and familial ischemic cerebral small-vessel disease. N. Engl. J. Med. 2009, 360, 1729–1739. [Google Scholar] [CrossRef]
- Tiaden, A.N.; Richards, P.J. The emerging roles of HTRA1 in musculoskeletal disease. Am. J. Pathol. 2013, 182, 1482–1488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Truebestein, L.; Tennstaedt, A.; Monig, T.; Krojer, T.; Canellas, F.; Kaiser, M.; Clausen, T.; Ehrmann, M. Substrate-induced remodeling of the active site regulates human HTRA1 activity. Nat. Struct. Mol. Biol. 2011, 18, 386–388. [Google Scholar] [CrossRef]
- Runyon, S.T.; Zhang, Y.; Appleton, B.A.; Sazinsky, S.L.; Wu, P.; Pan, B.; Wiesmann, C.; Skelton, N.J.; Sidhu, S.S. Structural and functional analysis of the PDZ domains of human HtrA1 and HtrA3. Protein Sci. 2007, 16, 2454–2471. [Google Scholar] [CrossRef] [Green Version]
- Murwantoko; Yano, M.; Ueta, Y.; Murasaki, A.; Kanda, H.; Oka, C.; Kawaichi, M. Binding of proteins to the PDZ domain regulates proteolytic activity of HtrA1 serine protease. Biochem. J. 2004, 381 Pt 3, 895–904. [Google Scholar] [CrossRef] [Green Version]
- Oka, C.; Tsujimoto, R.; Kajikawa, M.; Koshiba-Takeuchi, K.; Ina, J.; Yano, M.; Tsuchiya, A.; Ueta, Y.; Soma, A.; Kanda, H.; et al. HtrA1 serine protease inhibits signaling mediated by Tgfbeta family proteins. Development 2004, 131, 1041–1053. [Google Scholar] [CrossRef] [Green Version]
- Lin, M.K.; Yang, J.; Hsu, C.W.; Gore, A.; Bassuk, A.G.; Brown, L.M.; Colligan, R.; Sengillo, J.D.; Mahajan, V.B.; Tsang, S.H. HTRA1, an age-related macular degeneration protease, processes extracellular matrix proteins EFEMP1 and TSP1. Aging Cell 2018, 17, e12710. [Google Scholar] [CrossRef] [PubMed]
- Vierkotten, S.; Muether, P.S.; Fauser, S. Overexpression of HTRA1 leads to ultrastructural changes in the elastic layer of Bruch’s membrane via cleavage of extracellular matrix components. PLoS ONE 2011, 6, e22959. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beaufort, N.; Scharrer, E.; Kremmer, E.; Lux, V.; Ehrmann, M.; Huber, R.; Houlden, H.; Werring, D.; Haffner, C.; Dichgans, M. Cerebral small vessel disease-related protease HtrA1 processes latent TGF-beta binding protein 1 and facilitates TGF-beta signaling. Proc. Natl. Acad. Sci. USA 2014, 111, 16496–16501. [Google Scholar] [CrossRef] [Green Version]
- Yu, Z.; Cao, S.; Wu, A.; Yue, H.; Zhang, C.; Wang, J.; Xia, M.; Wu, J. Genetically Confirmed CARASIL: Case Report with Novel HTRA1 Mutation and Literature Review. World Neurosurg. 2020, 143, 121–128. [Google Scholar] [CrossRef] [PubMed]
- Graham, J.R.; Chamberland, A.; Lin, Q.; Li, X.J.; Dai, D.; Zeng, W.; Ryan, M.S.; Rivera-Bermudez, M.A.; Flannery, C.R.; Yang, Z. Serine protease HTRA1 antagonizes transforming growth factor-beta signaling by cleaving its receptors and loss of HTRA1 in vivo enhances bone formation. PLoS ONE 2013, 8, e74094. [Google Scholar] [CrossRef] [Green Version]
- Tennstaedt, A.; Popsel, S.; Truebestein, L.; Hauske, P.; Brockmann, A.; Schmidt, N.; Irle, I.; Sacca, B.; Niemeyer, C.M.; Brandt, R.; et al. Human high temperature requirement serine protease A1 (HTRA1) degrades tau protein aggregates. J. Biol. Chem. 2012, 287, 20931–20941. [Google Scholar] [CrossRef] [Green Version]
- He, X.; Khurana, A.; Maguire, J.L.; Chien, J.; Shridhar, V. HtrA1 sensitizes ovarian cancer cells to cisplatin-induced cytotoxicity by targeting XIAP for degradation. Int. J. Cancer 2012, 130, 1029–1035. [Google Scholar] [CrossRef] [Green Version]
- Chien, J.; Ota, T.; Aletti, G.; Shridhar, R.; Boccellino, M.; Quagliuolo, L.; Baldi, A.; Shridhar, V. Serine protease HtrA1 associates with microtubules and inhibits cell migration. Mol. Cell Biol. 2009, 29, 4177–4187. [Google Scholar] [CrossRef] [Green Version]
- Ajayi, F.; Kongoasa, N.; Gaffey, T.; Asmann, Y.W.; Watson, W.J.; Baldi, A.; Lala, P.; Shridhar, V.; Brost, B.; Chien, J. Elevated expression of serine protease HtrA1 in preeclampsia and its role in trophoblast cell migration and invasion. Am. J. Obstet. Gynecol. 2008, 199, 557.e1–557.e10. [Google Scholar] [CrossRef]
- Moisoi, N.; Klupsch, K.; Fedele, V.; East, P.; Sharma, S.; Renton, A.; Plun-Favreau, H.; Edwards, R.E.; Teismann, P.; Esposti, M.D.; et al. Mitochondrial dysfunction triggered by loss of HtrA2 results in the activation of a brain-specific transcriptional stress response. Cell Death Differ. 2009, 16, 449–464. [Google Scholar] [CrossRef] [Green Version]
- Jones, J.M.; Datta, P.; Srinivasula, S.M.; Ji, W.; Gupta, S.; Zhang, Z.; Davies, E.; Hajnoczky, G.; Saunders, T.L.; Van Keuren, M.L.; et al. Loss of Omi mitochondrial protease activity causes the neuromuscular disorder of mnd2 mutant mice. Nature 2003, 425, 721–727. [Google Scholar] [CrossRef] [Green Version]
- Park, H.J.; Kim, S.S.; Seong, Y.M.; Kim, K.H.; Goo, H.G.; Yoon, E.J.; Do, S.M.; Kang, S.; Rhim, H. Beta-amyloid precursor protein is a direct cleavage target of HtrA2 serine protease. Implications for the physiological function of HtrA2 in the mitochondria. J. Biol. Chem. 2006, 281, 34277–34287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, Y.; Imai, Y.; Nakayama, H.; Takahashi, K.; Takio, K.; Takahashi, R. A serine protease, HtrA2, is released from the mitochondria and interacts with XIAP, inducing cell death. Mol. Cell 2001, 8, 613–621. [Google Scholar] [CrossRef]
- Verhagen, A.M.; Silke, J.; Ekert, P.G.; Pakusch, M.; Kaufmann, H.; Connolly, L.M.; Day, C.L.; Tikoo, A.; Burke, R.; Wrobel, C.; et al. HtrA2 promotes cell death through its serine protease activity and its ability to antagonize inhibitor of apoptosis proteins. J. Biol. Chem. 2002, 277, 445–454. [Google Scholar] [CrossRef] [Green Version]
- Cilenti, L.; Soundarapandian, M.M.; Kyriazis, G.A.; Stratico, V.; Singh, S.; Gupta, S.; Bonventre, J.V.; Alnemri, E.S.; Zervos, A.S. Regulation of HAX-1 anti-apoptotic protein by Omi/HtrA2 protease during cell death. J. Biol. Chem. 2004, 279, 50295–50301. [Google Scholar] [CrossRef] [Green Version]
- Trencia, A.; Fiory, F.; Maitan, M.A.; Vito, P.; Barbagallo, A.P.; Perfetti, A.; Miele, C.; Ungaro, P.; Oriente, F.; Cilenti, L.; et al. Omi/HtrA2 promotes cell death by binding and degrading the anti-apoptotic protein ped/pea-15. J. Biol. Chem. 2004, 279, 46566–46572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chien, J. WT1 as a substrate of HtrA2: A potential pathway for therapeutic targeting by HtrA proteases. Future Oncol. 2010, 6, 1233–1235. [Google Scholar] [CrossRef] [Green Version]
- Hartkamp, J.; Carpenter, B.; Roberts, S.G. The Wilms’ tumor suppressor protein WT1 is processed by the serine protease HtrA2/Omi. Mol. Cell 2010, 37, 159–171. [Google Scholar] [CrossRef]
- Tocharus, J.; Tsuchiya, A.; Kajikawa, M.; Ueta, Y.; Oka, C.; Kawaichi, M. Developmentally regulated expression of mouse HtrA3 and its role as an inhibitor of TGF-beta signaling. Dev. Growth Differ. 2004, 46, 257–274. [Google Scholar] [CrossRef]
- Wenta, T.; Zurawa-Janicka, D.; Rychlowski, M.; Jarzab, M.; Glaza, P.; Lipinska, A.; Bienkowska-Szewczyk, K.; Herman-Antosiewicz, A.; Skorko-Glonek, J.; Lipinska, B. HtrA3 is a cellular partner of cytoskeleton proteins and TCP1alpha chaperonin. J. Proteom. 2018, 177, 88–111. [Google Scholar] [CrossRef] [PubMed]
- Wenta, T.; Rychlowski, M.; Jurewicz, E.; Jarzab, M.; Zurawa-Janicka, D.; Filipek, A.; Lipinska, B. The HtrA3 protease promotes drug-induced death of lung cancer cells by cleavage of the X-linked inhibitor of apoptosis protein (XIAP). FEBS J. 2019, 286, 4579–4596. [Google Scholar] [CrossRef] [PubMed]
- Dynon, K.; Heng, S.; Puryer, M.; Li, Y.; Walton, K.; Endo, Y.; Nie, G. HtrA3 as an early marker for preeclampsia: Specific monoclonal antibodies and sensitive high-throughput assays for serum screening. PLoS ONE 2012, 7, e45956. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.-J.; Cheong, M.-L.; Lee, Y.-S.; Lee, M.-T.; Chen, H. High-temperature requirement protein A4 (HtrA4) suppresses the fusogenic activity of syncytin-1 and promotes trophoblast invasion. Mol. Cell. Biol. 2012, 32, 3707–3717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vande Walle, L.; Lamkanfi, M.; Vandenabeele, P. The mitochondrial serine protease HtrA2/Omi: An overview. Cell Death Differ. 2008, 15, 453–460. [Google Scholar] [CrossRef] [Green Version]
- Bogaerts, V.; Nuytemans, K.; Reumers, J.; Pals, P.; Engelborghs, S.; Pickut, B.; Corsmit, E.; Peeters, K.; Schymkowitz, J.; De Deyn, P.P.; et al. Genetic variability in the mitochondrial serine protease HTRA2 contributes to risk for Parkinson disease. Hum. Mutat. 2008, 29, 832–840. [Google Scholar] [CrossRef]
- Kruger, R.; Sharma, M.; Riess, O.; Gasser, T.; Van Broeckhoven, C.; Theuns, J.; Aasly, J.; Annesi, G.; Bentivoglio, A.R.; Brice, A.; et al. A large-scale genetic association study to evaluate the contribution of Omi/HtrA2 (PARK13) to Parkinson’s disease. Neurobiol. Aging 2011, 32, 548.e9–548.e18. [Google Scholar] [CrossRef] [Green Version]
- Gray, C.W.; Ward, R.V.; Karran, E.; Turconi, S.; Rowles, A.; Viglienghi, D.; Southan, C.; Barton, A.; Fantom, K.G.; West, A.; et al. Characterization of human HtrA2, a novel serine protease involved in the mammalian cellular stress response. Eur. J. Biochem. 2000, 267, 5699–5710. [Google Scholar] [CrossRef] [Green Version]
- Cilenti, L.; Kyriazis, G.A.; Soundarapandian, M.M.; Stratico, V.; Yerkes, A.; Park, K.M.; Sheridan, A.M.; Alnemri, E.S.; Bonventre, J.V.; Zervos, A.S. Omi/HtrA2 protease mediates cisplatin-induced cell death in renal cells. Am. J. Physiol. Ren. Physiol. 2005, 288, F371–F379. [Google Scholar] [CrossRef] [Green Version]
- Beleford, D.; Rattan, R.; Chien, J.; Shridhar, V. High temperature requirement A3 (HtrA3) promotes etoposide- and cisplatin-induced cytotoxicity in lung cancer cell lines. J. Biol. Chem. 2010, 285, 12011–12027. [Google Scholar] [CrossRef] [Green Version]
- Glaza, P.; Osipiuk, J.; Wenta, T.; Zurawa-Janicka, D.; Jarzab, M.; Lesner, A.; Banecki, B.; Skorko-Glonek, J.; Joachimiak, A.; Lipinska, B. Structural and Functional Analysis of Human HtrA3 Protease and Its Subdomains. PLoS ONE 2015, 10, e0131142. [Google Scholar] [CrossRef] [PubMed]
- Singh, H.; Makino, S.; Endo, Y.; Li, Y.; Stephens, A.N.; Nie, G. Application of the wheat-germ cell-free translation system to produce high temperature requirement A3 (HtrA3) proteases. BioTechniques 2012, 52, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.Y.; Chuang, P.Y.; Chen, C.P.; Chiu, Y.H.; Lo, H.F.; Cheong, M.L.; Huang, J.Y.; Kuo, P.L.; Chen, H. Functional antagonism between high temperature requirement protein A (HtrA) family members regulates trophoblast invasion. J. Biol. Chem. 2014, 289, 22958–22968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Damsky, C.H.; Librach, C.; Lim, K.H.; Fitzgerald, M.L.; McMaster, M.T.; Janatpour, M.; Zhou, Y.; Logan, S.K.; Fisher, S.J. Integrin switching regulates normal trophoblast invasion. Development 1994, 120, 3657–3666. [Google Scholar] [CrossRef]
- Chiu, Y.H.; Chen, H. GATA3 inhibits GCM1 activity and trophoblast cell invasion. Sci. Rep. 2016, 6, 21630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mansilla, M.; Wang, Y.; Lim, R.; Palmer, K.; Nie, G. HtrA4 is up-regulated during trophoblast syncytialization and BeWo cells fail to syncytialize without HtrA4. Sci. Rep. 2021, 11, 14363. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, Q.; Zhao, M.; Walton, K.; Harrison, C.; Nie, G. Multiple Soluble TGF-beta Receptors in Addition to Soluble Endoglin Are Elevated in Preeclamptic Serum and They Synergistically Inhibit TGF-beta Signaling. J. Clin. Endocrinol. Metab. 2017, 102, 3065–3074. [Google Scholar] [CrossRef]
- Wenta, T.; Rychlowski, M.; Jarzab, M.; Lipinska, B. HtrA4 Protease Promotes Chemotherapeutic-Dependent Cancer Cell Death. Cells 2019, 8, 1112. [Google Scholar] [CrossRef] [Green Version]
- Kummari, R.; Dutta, S.; Chaganti, L.K.; Bose, K. Discerning the mechanism of action of HtrA4: A serine protease implicated in the cell death pathway. Biochem. J. 2019, 476, 1445–1463. [Google Scholar] [CrossRef]
- Wenta, T.; Jarzab, M.; Rychlowski, M.; Borysiak, M.; Latala, A.; Zurawa-Janicka, D.; Filipek, A.; Lipinska, B. Cellular substrates and pro-apoptotic function of the human HtrA4 protease. J. Proteom. 2019, 209, 103505. [Google Scholar] [CrossRef]
- Chien, J.; Campioni, M.; Shridhar, V.; Baldi, A. HtrA serine proteases as potential therapeutic targets in cancer. Curr. Cancer Drug Targets 2009, 9, 451–468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chien, J.; Staub, J.; Hu, S.I.; Erickson-Johnson, M.R.; Couch, F.J.; Smith, D.I.; Crowl, R.M.; Kaufmann, S.H.; Shridhar, V. A candidate tumor suppressor HtrA1 is downregulated in ovarian cancer. Oncogene 2004, 23, 1636–1644. [Google Scholar] [CrossRef] [Green Version]
- Baldi, A.; De Luca, A.; Morini, M.; Battista, T.; Felsani, A.; Baldi, F.; Catricala, C.; Amantea, A.; Noonan, D.M.; Albini, A.; et al. The HtrA1 serine protease is down-regulated during human melanoma progression and represses growth of metastatic melanoma cells. Oncogene 2002, 21, 6684–6688. [Google Scholar] [CrossRef]
- Sotiriou, C.; Wirapati, P.; Loi, S.; Harris, A.; Fox, S.; Smeds, J.; Nordgren, H.; Farmer, P.; Praz, V.; Haibe-Kains, B.; et al. Gene expression profiling in breast cancer: Understanding the molecular basis of histologic grade to improve prognosis. J. Natl. Cancer Inst. 2006, 98, 262–272. [Google Scholar] [CrossRef] [PubMed]
- Welsh, J.B.; Sapinoso, L.M.; Su, A.I.; Kern, S.G.; Wang-Rodriguez, J.; Moskaluk, C.A.; Frierson, H.F., Jr.; Hampton, G.M. Analysis of gene expression identifies candidate markers and pharmacological targets in prostate cancer. Cancer Res. 2001, 61, 5974–5978. [Google Scholar] [PubMed]
- Esposito, V.; Campioni, M.; De Luca, A.; Spugnini, E.P.; Baldi, F.; Cassandro, R.; Mancini, A.; Vincenzi, B.; Groeger, A.; Caputi, M.; et al. Analysis of HtrA1 serine protease expression in human lung cancer. Anticancer Res. 2006, 26, 3455–3459. [Google Scholar]
- Mullany, S.A.; Moslemi-Kebria, M.; Rattan, R.; Khurana, A.; Clayton, A.; Ota, T.; Mariani, A.; Podratz, K.C.; Chien, J.; Shridhar, V. Expression and functional significance of HtrA1 loss in endometrial cancer. Clin. Cancer Res. 2011, 17, 427–436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, X.; Ota, T.; Liu, P.; Su, C.; Chien, J.; Shridhar, V. Downregulation of HtrA1 promotes resistance to anoikis and peritoneal dissemination of ovarian cancer cells. Cancer Res. 2010, 70, 3109–3118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chien, J.; Aletti, G.; Baldi, A.; Catalano, V.; Muretto, P.; Keeney, G.L.; Kalli, K.R.; Staub, J.; Ehrmann, M.; Cliby, W.A.; et al. Serine protease HtrA1 modulates chemotherapy-induced cytotoxicity. J. Clin. Investig. 2006, 116, 1994–2004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Diao, Y.-C.; Jiao, J.-W.; Wang, C.; Yu, J.; Li, B.; Dai, S.-Z. HtrA2/Omi protease in epithelial ovarian carcinoma: Clinical significance and antineoplastic efficacy of its overexpression on subcutaneous transplanted tumor in nude mice model. Int. J. Clin. Exp. Patho. 2016, 9, 3161–3169. [Google Scholar]
- Soyama, H.; Miyamoto, M.; Takano, M.; Aoyama, T.; Matsuura, H.; Sakamoto, T.; Takasaki, K.; Kuwahara, M.; Kato, K.; Yoshikawa, T.; et al. Ovarian serous carcinomas acquire cisplatin resistance and increased invasion through downregulation of the high-temperature-required protein A2 (HtrA2), following repeated treatment with cisplatin. Med. Oncol. 2017, 34, 201. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Chen, X.; Ping, H.; Chen, Z.; Zeng, F.; Lu, G. Immunohistochemical analysis of Omi/HtrA2 expression in prostate cancer and benign prostatic hyperplasia. J. Huazhong Univ. Sci. Technology. Med. Sci. Hua Zhong Ke Ji Da Xue Xue Bao Yi Xue Ying De Wen Ban Huazhong Keji Daxue Xuebao Yixue Yingdewen Ban 2005, 25, 671–673. [Google Scholar] [CrossRef]
- Lee, S.H.; Lee, J.W.; Kim, H.S.; Kim, S.Y.; Park, W.S.; Kim, S.H.; Lee, J.Y.; Yoo, N.J. Immunohistochemical analysis of Omi/HtrA2 expression in stomach cancer. APMIS Acta Pathol. Microbiol. Immunol. Scand. 2003, 111, 586–590. [Google Scholar] [CrossRef]
- Dhanasekaran, S.M.; Barrette, T.R.; Ghosh, D.; Shah, R.; Varambally, S.; Kurachi, K.; Pienta, K.J.; Rubin, M.A.; Chinnaiyan, A.M. Delineation of prognostic biomarkers in prostate cancer. Nature 2001, 412, 822–826. [Google Scholar] [CrossRef]
- Sorlie, T.; Tibshirani, R.; Parker, J.; Hastie, T.; Marron, J.S.; Nobel, A.; Deng, S.; Johnsen, H.; Pesich, R.; Geisler, S.; et al. Repeated observation of breast tumor subtypes in independent gene expression data sets. Proc. Natl. Acad. Sci. USA 2003, 100, 8418–8423. [Google Scholar] [CrossRef] [Green Version]
- Bowden, M.A.; Drummond, A.E.; Fuller, P.J.; Salamonsen, L.A.; Findlay, J.K.; Nie, G. High-temperature requirement factor A3 (Htra3): A novel serine protease and its potential role in ovarian function and ovarian cancers. Mol. Cell. Endocrinol. 2010, 327, 13–18. [Google Scholar] [CrossRef]
- Zhao, M.; Ding, J.X.; Nie, G.Y.; Wei, J.; Li, Y.; Yin, X.Y.; Chen, Q. HTRA3 is reduced in ovarian cancers regardless of stage. Cancer Investig. 2014, 32, 464–469. [Google Scholar] [CrossRef] [PubMed]
- Singh, H.; Li, Y.; Fuller, P.J.; Harrison, C.; Rao, J.; Stephens, A.N.; Nie, G. HtrA3 Is Downregulated in Cancer Cell Lines and Significantly Reduced in Primary Serous and Granulosa Cell Ovarian Tumors. J. Cancer 2013, 4, 152–164. [Google Scholar] [CrossRef] [Green Version]
- Bowden, M.A.; Di Nezza-Cossens, L.A.; Jobling, T.; Salamonsen, L.A.; Nie, G. Serine proteases HTRA1 and HTRA3 are down-regulated with increasing grades of human endometrial cancer. Gynecol. Oncol. 2006, 103, 253–260. [Google Scholar] [CrossRef] [PubMed]
- Lv, Q.; Yang, B.; Ning, C.; Xie, B.; Nie, G.; Chen, X.; Chen, Q. Hypoxia is involved in the reduction of HtrA3 in patients with endometrial hyperplasia and cancer. Biochem. Biophys. Res. Commun. 2018, 503, 2918–2923. [Google Scholar] [CrossRef] [PubMed]
- Yin, Y.; Wu, M.; Nie, G.; Wang, K.; Wei, J.; Zhao, M.; Chen, Q. HtrA3 is negatively correlated with lymph node metastasis in invasive ductal breast cancer. Tumour Biol. J. Int. Soc. Oncodevelopmental Biol. Med. 2013, 34, 3611–3617. [Google Scholar] [CrossRef]
- Beleford, D.; Liu, Z.; Rattan, R.; Quagliuolo, L.; Boccellino, M.; Baldi, A.; Maguire, J.; Staub, J.; Molina, J.; Shridhar, V. Methylation induced gene silencing of HtrA3 in smoking-related lung cancer. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2010, 16, 398–409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, J.; Zhang, J.; Zhang, X.; Feng, M.; Qu, J. High temperature requirement A3 (HTRA3) expression predicts postoperative recurrence and survival in patients with non-small-cell lung cancer. Oncotarget 2016, 7, 40725–40734. [Google Scholar] [CrossRef] [Green Version]
- Hao, Y.; Triadafilopoulos, G.; Sahbaie, P.; Young, H.S.; Omary, M.B.; Lowe, A.W. Gene expression profiling reveals stromal genes expressed in common between Barrett’s esophagus and adenocarcinoma. Gastroenterology 2006, 131, 925–933. [Google Scholar] [CrossRef] [Green Version]
- Iacobuzio-Donahue, C.A.; Maitra, A.; Olsen, M.; Lowe, A.W.; van Heek, N.T.; Rosty, C.; Walter, K.; Sato, N.; Parker, A.; Ashfaq, R.; et al. Exploration of global gene expression patterns in pancreatic adenocarcinoma using cDNA microarrays. Am. J. Pathol. 2003, 162, 1151–1162. [Google Scholar] [CrossRef]
- Korkola, J.E.; Houldsworth, J.; Chadalavada, R.S.; Olshen, A.B.; Dobrzynski, D.; Reuter, V.E.; Bosl, G.J.; Chaganti, R.S. Down-regulation of stem cell genes, including those in a 200-kb gene cluster at 12p13.31, is associated with in vivo differentiation of human male germ cell tumors. Cancer Res. 2006, 66, 820–827. [Google Scholar] [CrossRef] [Green Version]
- Andersson, A.; Ritz, C.; Lindgren, D.; Eden, P.; Lassen, C.; Heldrup, J.; Olofsson, T.; Rade, J.; Fontes, M.; Porwit-Macdonald, A.; et al. Microarray-based classification of a consecutive series of 121 childhood acute leukemias: Prediction of leukemic and genetic subtype as well as of minimal residual disease status. Leukemia 2007, 21, 1198–1203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van‘t Veer, L.J.; Dai, H.; van de Vijver, M.J.; He, Y.D.; Hart, A.A.; Mao, M.; Peterse, H.L.; van der Kooy, K.; Marton, M.J.; Witteveen, A.T.; et al. Gene expression profiling predicts clinical outcome of breast cancer. Nature 2002, 415, 530–536. [Google Scholar] [CrossRef] [Green Version]
- Baldi, A.; Mottolese, M.; Vincenzi, B.; Campioni, M.; Mellone, P.; Di Marino, M.; di Crescenzo, V.G.; Visca, P.; Menegozzo, S.; Spugnini, E.P.; et al. The serine protease HtrA1 is a novel prognostic factor for human mesothelioma. Pharmacogenomics 2008, 9, 1069–1077. [Google Scholar] [CrossRef] [PubMed]
- Hasan, M.Z.; Ikawati, M.; Tocharus, J.; Kawaichi, M.; Oka, C. Abnormal development of placenta in HtrA1-deficient mice. Dev. Biol. 2015, 397, 89–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nie, G.; Li, Y.; Salamonsen, L.A. Serine protease HtrA1 is developmentally regulated in trophoblast and uterine decidual cells during placental formation in the mouse. Dev. Dyn. 2005, 233, 1102–1109. [Google Scholar] [CrossRef]
- Zong, L.; Wang, L.; Huang, P.; Shao, W.; Song, Y.; Gou, W. High temperature requirement A1 in placental tissues and serum from pre-eclamptic pregnancies with or without fetal growth restriction. Arch. Med. Sci. AMS 2013, 9, 690–696. [Google Scholar] [CrossRef]
- Skinner, S.; Rolnik, D.L.; Wang, Y.; Nie, G.; Syngelaki, A.; Nicolaides, K.H.; da Silva Costa, F. Second and third trimester serum levels of HtrA1 in pregnancies affected by pre-eclampsia. Placenta 2021, 106, 1–6. [Google Scholar] [CrossRef]
- Bowden, M.A.; Li, Y.; Liu, Y.X.; Findlay, J.K.; Salamonsen, L.A.; Nie, G. HTRA3 expression in non-pregnant rhesus monkey ovary and endometrium, and at the maternal-fetal interface during early pregnancy. Reprod. Biol. Endocrinol. RBE 2008, 6, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nie, G.; Li, Y.; He, H.; Findlay, J.K.; Salamonsen, L.A. HtrA3, a serine protease possessing an IGF-binding domain, is selectively expressed at the maternal-fetal interface during placentation in the mouse. Placenta 2006, 27, 491–501. [Google Scholar] [CrossRef]
- Singh, H.; Makino, S.I.; Endo, Y.; Nie, G. Inhibition of HTRA3 stimulates trophoblast invasion during human placental development. Placenta 2010, 31, 1085–1092. [Google Scholar] [CrossRef]
- Singh, H.; Endo, Y.; Nie, G. Decidual HtrA3 negatively regulates trophoblast invasion during human placentation. Hum. Reprod. 2011, 26, 748–757. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Salamonsen, L.A.; Hyett, J.; Costa, F.D.S.; Nie, G. Maternal HtrA3 optimizes placental development to influence offspring birth weight and subsequent white fat gain in adulthood. Sci. Rep. 2017, 7, 4627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teoh, S.S.Y.; Wang, Y.; Li, Y.; Leemaqz, S.Y.; Dekker, G.A.; Roberts, C.T.; Nie, G. Low Serum Levels of HtrA3 at 15 Weeks of Gestation Are Associated with Late-Onset Preeclampsia Development and Small for Gestational Age Birth. Fetal Diagn. Ther. 2019, 46, 392–401. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Li, Y.; Hyett, J.; da Silva Costa, F.; Nie, G. HtrA3 Isoform-Specific ELISAs for Early Detection of Preeclampsia. J. Biomol. Screen. 2016, 23, 1092–1099. [Google Scholar] [CrossRef]
- Nishizawa, H.; Ota, S.; Suzuki, M.; Kato, T.; Sekiya, T.; Kurahashi, H.; Udagawa, Y. Comparative gene expression profiling of placentas from patients with severe pre-eclampsia and unexplained fetal growth restriction. Reprod. Biol. Endocrinol. RBE 2011, 9, 107. [Google Scholar] [CrossRef] [Green Version]
- Lapaire, O.; Grill, S.; Lalevee, S.; Kolla, V.; Hosli, I.; Hahn, S. Microarray screening for novel preeclampsia biomarker candidates. Fetal Diagn. Ther. 2012, 31, 147–153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaartokallio, T.; Cervera, A.; Kyllonen, A.; Laivuori, K.; Kere, J.; Laivuori, H. Gene expression profiling of pre-eclamptic placentae by RNA sequencing. Sci. Rep. 2015, 5, 14107. [Google Scholar] [CrossRef] [PubMed]
- Brew, O.; Sullivan, M.H.; Woodman, A. Comparison of Normal and Pre-Eclamptic Placental Gene Expression: A Systematic Review with Meta-Analysis. PLoS ONE 2016, 11, e0161504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Lim, R.; Nie, G. Elevated circulating HtrA4 in preeclampsia may alter endothelial expression of senescence genes. Placenta 2020, 90, 71–81. [Google Scholar] [CrossRef]
- Wang, Y.; Gu, Y.; Granger, D.N.; Roberts, J.M.; Alexander, J.S. Endothelial junctional protein redistribution and increased monolayer permeability in human umbilical vein endothelial cells isolated during preeclampsia. Am. J. Obstet. Gynecol. 2002, 186, 214–220. [Google Scholar] [CrossRef]
- Prochazka, M.; Prochazkova, J.; Lubusky, M.; Pilka, R.; Ulehlova, J.; Michalec, I.; Polak, P.; Kacerovsky, M.; Slavik, L. Markers of endothelial activation in preeclampsia. Clin. Lab. 2015, 61, 39–46. [Google Scholar] [CrossRef]
- Rousseau, A.; Favier, R.; Van Dreden, P. Elevated circulating soluble thrombomodulin activity, tissue factor activity and circulating procoagulant phospholipids: New and useful markers for pre-eclampsia? Eur. J. Obstet. Gynecol. Reprod. Biol. 2009, 146, 46–49. [Google Scholar] [CrossRef]
- Stenczer, B.; Molvarec, A.; Szabo, G.; Szarka, A.; Fugedi, G.; Szijarto, J.; Rigo, J., Jr. Circulating levels of thrombospondin-1 are decreased in HELLP syndrome. Thromb. Res. 2012, 129, 470–473. [Google Scholar] [CrossRef]
- Steinmetz, M.; Nickenig, G.; Werner, N. Endothelial-regenerating cells: An expanding universe. Hypertension 2010, 55, 593–599. [Google Scholar] [CrossRef] [Green Version]




| HtrA1 | HtrA2 | HtrA3 | HtrA4 | |
|---|---|---|---|---|
| Approved name | HtrA serine peptidase 1 | HtrA serine peptidase 2 | HtrA serine peptidase 3 | HtrA serine peptidase 4 |
| Gene name | HTRA1 | HTRA2 | HTRA3 | HTRA4 |
| Gene Synonyms | ARMD7, CARASIL, CADASIL2, HtrA, IGFBP5-protease, L56, ORF480, PRSS11 | MGCA8, OMI, PARK13, PRSS25 | TASP, PRSP | |
| Entrez Gene ID | 5654 | 27,429 | 94,031 | 203,100 |
| HGNC ID | 9476 | 14,348 | 30,406 | 26,909 |
| Ensembl ID | ENSG00000166033 | ENSG00000115317 | ENSG00000170801 | ENSG00000169495 |
| MEROPS ID | S01.277 | S01.278 | S01.284 | S01.285 |
| Chromosoml location | 10q26.13 (122,461,553–122,514,907) Plus strand | 2p13.1 (74,529,405–74,533,556) Plus strand | 4p16.1 (8,269,712–8,307,098) Plus strand | 8p11.22 (38,974,228–38,988,663) Plus strand |
| Gene size (bases) | 53,355 | 4152 | 37,387 | 14,436 |
| RefSeq | NM_002775 NP_002766 | NM_013247 NP_037379 | NM_053044 NP_444272 | NM_153692 NP_710159 |
| mRNA (bases) | 2091 | 1785 | 2539 | 2032 |
| Number of exons | 9 | 8 | 10 | 9 |
| Alternative splicing | 2 additional variants predicted | 9 additional varaints predicted with some confirmed | 1 additional variant confirmed | unknown |
| UniProtKB/Swiss-Prot ID | Q92743 | O43464 | P83110 | P83105 |
| Full lenght Protein (aa) | 480 | 458 | 453 | 476 |
| Transient peptide | N/A | 1–31 (31aa) | N/A | N/A |
| Signal peptide | 1–22 (22aa) | N/A | 1–17 (17aa) | 1–31 (31aa) |
| Propeptide | N/A | 32–133 (102) | N/A | N/A |
| Mature peptide Mol_wt (dalton) | 23–480 (458aa) 49,048 | 134–458 (325aa) 34,981 | 18–453 (436aa) 46,945 | 32–476 (445aa) 47,685 |
| Transmembrane | N/A | 105–125 (21aa) | N/A | N/A |
| IGF-binding domain | 33–100 (68aa) | N/A | 21–77 (57aa) | 36–99 (64aa) |
| Kazal-like domain | 98–157 (60aa) | N/A | 64–128 (65aa) | 88–154 (67aa) |
| Serine protease domain | 204–364 (161aa) | 166–342 (177aa) | 175–340 (166aa) | 202–362 (161aa) |
| PDZ domain | 365–467 (103aa) | 364–445 (82aa) | 359–444 (86aa) | 383–474 (92aa) |
| Catalytic triad Histidine, Aspartic acid and Serine | H220 D250 S328 | H198 D228 S306 | H191 D227 S305 | H218 D248 S326 |
| Trimer stabilization sites | Y169 F171 F278 | F149 | Likely F140 F142 F255 | Likely Y167 F169 F276 |
| Protein binding sites | 382..385, 387, 444..445, 448...449 | 361..364, 366, 423..424, 427..428 | 356...359, 361, 418..419, 422..423 | 380..383, 385, 442–443, 446..447 |
| IAP-binding motif | unknown | 134-137 | unknown | unknown |
| Potential phosphorylation sites | S195, T237, Y238, T365, S367, S456 | T51, T242, T54, T326, S96, S330, T157, S400, S212,T453 | S214, S334, T363 | S310, Y314, S424 |
| Comparision between HtrA Members | HtrA1 vs. HtrA3 | HtrA1 vs. HtrA4 | HtrA3 vs. HtrA4 | ||||
|---|---|---|---|---|---|---|---|
| Identity | Similarity | Identity | Similarity | Identity | Similarity | ||
| Overall | 58% | 73% | 54% | 70% | 51% | 68% | |
| Individual domains | Signal peptide | 50% | 50% | 89% | 89% | − | − |
| IGFB | 52% | 58% | 45% | 50% | 50% | 58% | |
| Kazal | 58% | 72% | 52% | 68% | 48% | 67% | |
| Trypsin | 77% | 89% | 73% | 88% | 69% | 81% | |
| PDZ | 41% | 71% | 41% | 68% | 41% | 70% | |
| HtrA Member | Functions and Substrates | Associated Diseases | References |
|---|---|---|---|
| HtrA1 | Degrading ECM proteins—fibronectin, type I collagen and decorin | Musculoskeletal diseases | [47] |
| Processing ECM proteins—EFEMP1 and TSP1 | Age-related macular degeneration | [52,53] | |
| Processing ECM proteins—LTBP-1 | Cerebral small vessel disease CARASIL | [46,54] | |
| Degrading APP and tau protein aggregates | Alzheimer’s disease | [43,57] | |
| Degrading XIAP to activate caspase activity Disruption of microtubules to inhibit cell migration | Cancer | [58,59] | |
| Processing ECM proteins or growth factors involved in trophoblast migration and invasion | Preeclampsia | [31,37,60] | |
| HtrA2 | Degrading unfolded or misfolded proteins | Parkinson’s disease | [61,62] |
| Breaking down APP in mitochondria to maintain normal cellular function | Alzheimer’s disease | [63] | |
| Binding to and degrading IAPs to facilitate caspase activities | Cancer and chemoresistance | [64,65] | |
| Degrading HAX-1 to promote cell death | Mitochondria-related dysfunction | [66] | |
| Degrading Ped-Pea15 to promote cell death | Environmental stressor-induced cellular dysfunction | [67] | |
| Degrading WTP1 to increase c-Myc and JunB to promote apoptosis | Cancer | [68,69] | |
| HtrA3 | Cleaving ECM proteins—decorin and biglycan | Osteoarthritis and cancer | [70] |
| Cleaving cytoskeleton proteins—actin, β-tubulin and vimentin | Destabilization of cytoskeleton dynamics in cancer treatments | [71] | |
| Acting as a chaperone by interacting with TCP1α chaperonin | Cancer and alteration of cancer cells | [71] | |
| Cleaving XIAP to promote drug-induced apoptosis | Cancer and chemoresistance | [72] | |
| Binding to BMP4, TGF-β1, TGF-β2 and GDF5 to inhibit their functions | Placental development and Preeclampsia | [22,70,73] | |
| HtrA4 | Degrading fibronectin to impede trophoblast invasion | Preeclampsia | [74] |
| Cleaving endothelial junction protein VE-Cadherin to increase permeability | Preeclampsia | [40] | |
| Cleaving VEGF-A receptor KDR to inhibit angiogenesis | Endothelial dysfunction in preeclampsia | [39] |
| Impact | Molecular Mechanisms of HtrA4 Action | References |
|---|---|---|
| Stimulation of endothelial cell proinflammation | Upregulating proinflammatory factors | [36] |
| Inhibition of endothelial cell proliferation | Downregulating cell cycle genes, including cell proliferation marker Ki67 | [38] |
| Inhibition of proliferation and differentiation of endothelial progenitor cells | Downregulating cell cycle genes and inhibiting tube formation | [38] |
| Promotion of cellular senescence | Upregulating senescence genes and downregulating genes involved in DNA repair | [135] |
| Inhibition of angiogenesis | Cleaving VEGF-A receptor KDR to inhibit VEGF-A action | [39] |
| Enhancement of cellular permeability | Cleaving VE-Cadherin to disrupt cell contact integrity | [24,40] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Nie, G. Overview of Human HtrA Family Proteases and Their Distinctive Physiological Roles and Unique Involvement in Diseases, Especially Cancer and Pregnancy Complications. Int. J. Mol. Sci. 2021, 22, 10756. https://doi.org/10.3390/ijms221910756
Wang Y, Nie G. Overview of Human HtrA Family Proteases and Their Distinctive Physiological Roles and Unique Involvement in Diseases, Especially Cancer and Pregnancy Complications. International Journal of Molecular Sciences. 2021; 22(19):10756. https://doi.org/10.3390/ijms221910756
Chicago/Turabian StyleWang, Yao, and Guiying Nie. 2021. "Overview of Human HtrA Family Proteases and Their Distinctive Physiological Roles and Unique Involvement in Diseases, Especially Cancer and Pregnancy Complications" International Journal of Molecular Sciences 22, no. 19: 10756. https://doi.org/10.3390/ijms221910756