
Exploring the Therapeutic Significance of microRNAs and lncRNAs in Kidney Diseases

 ,
,  ,
,  , and
, and 
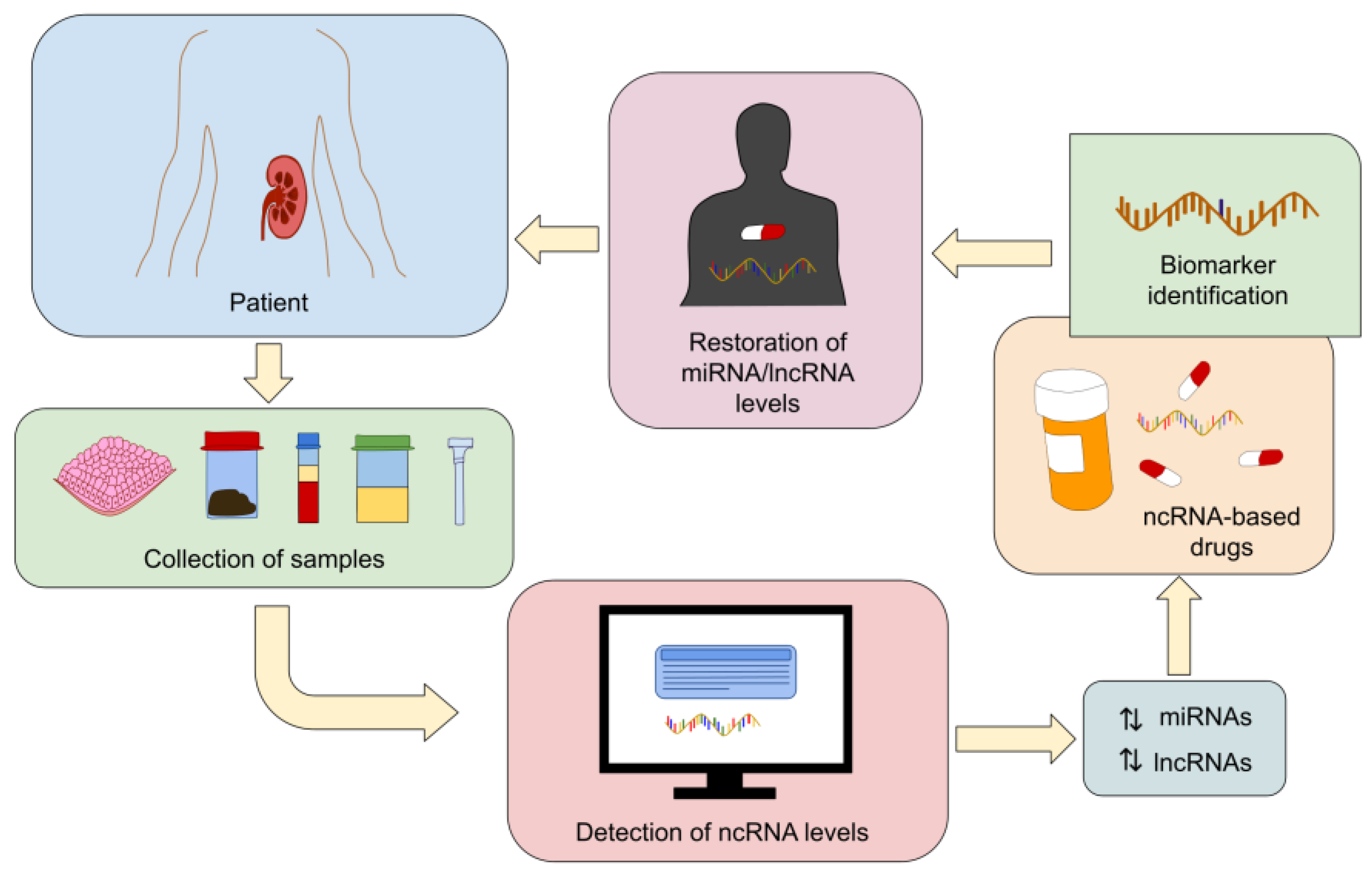{kind=link}
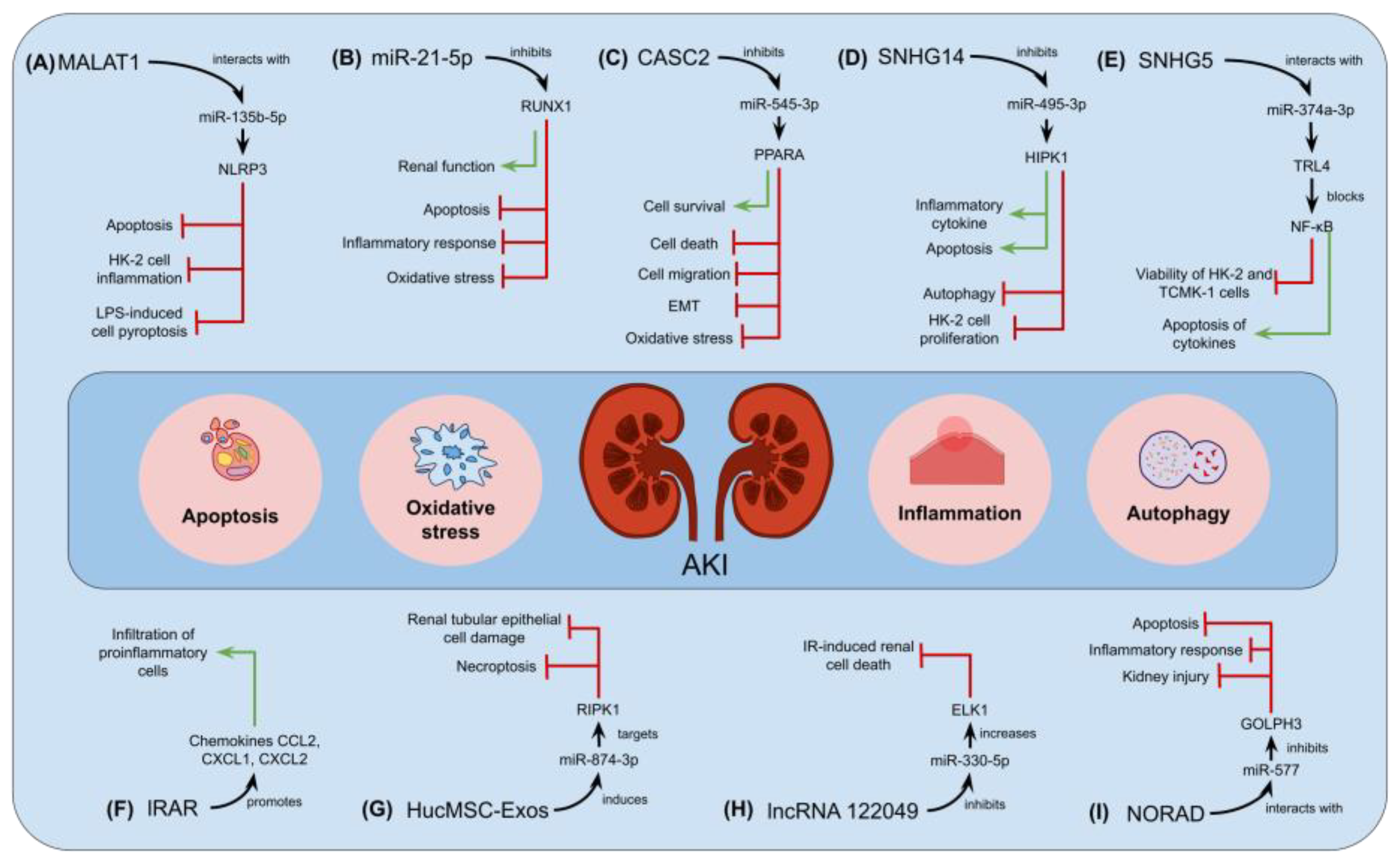{kind=link}
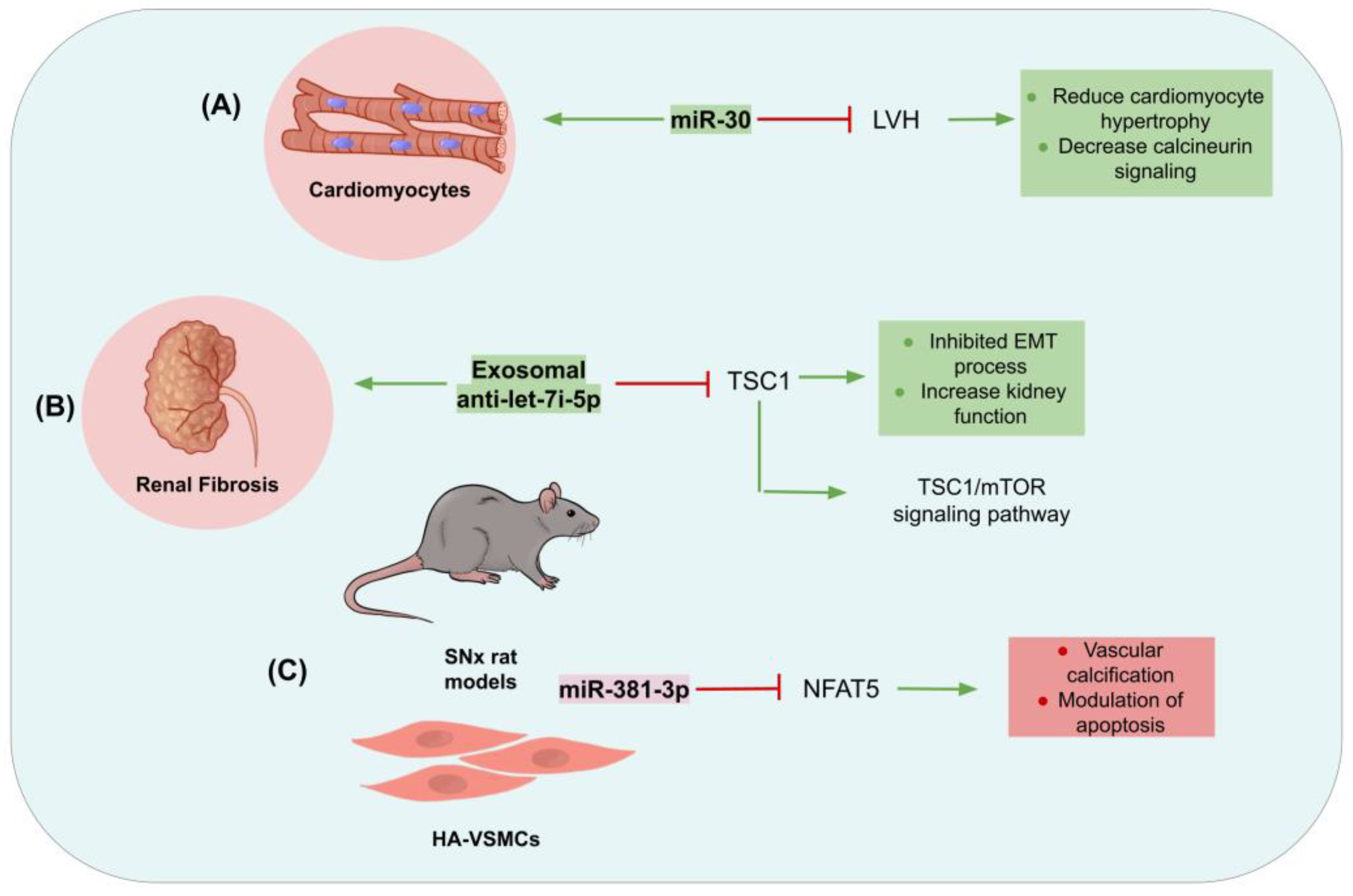{kind=link}
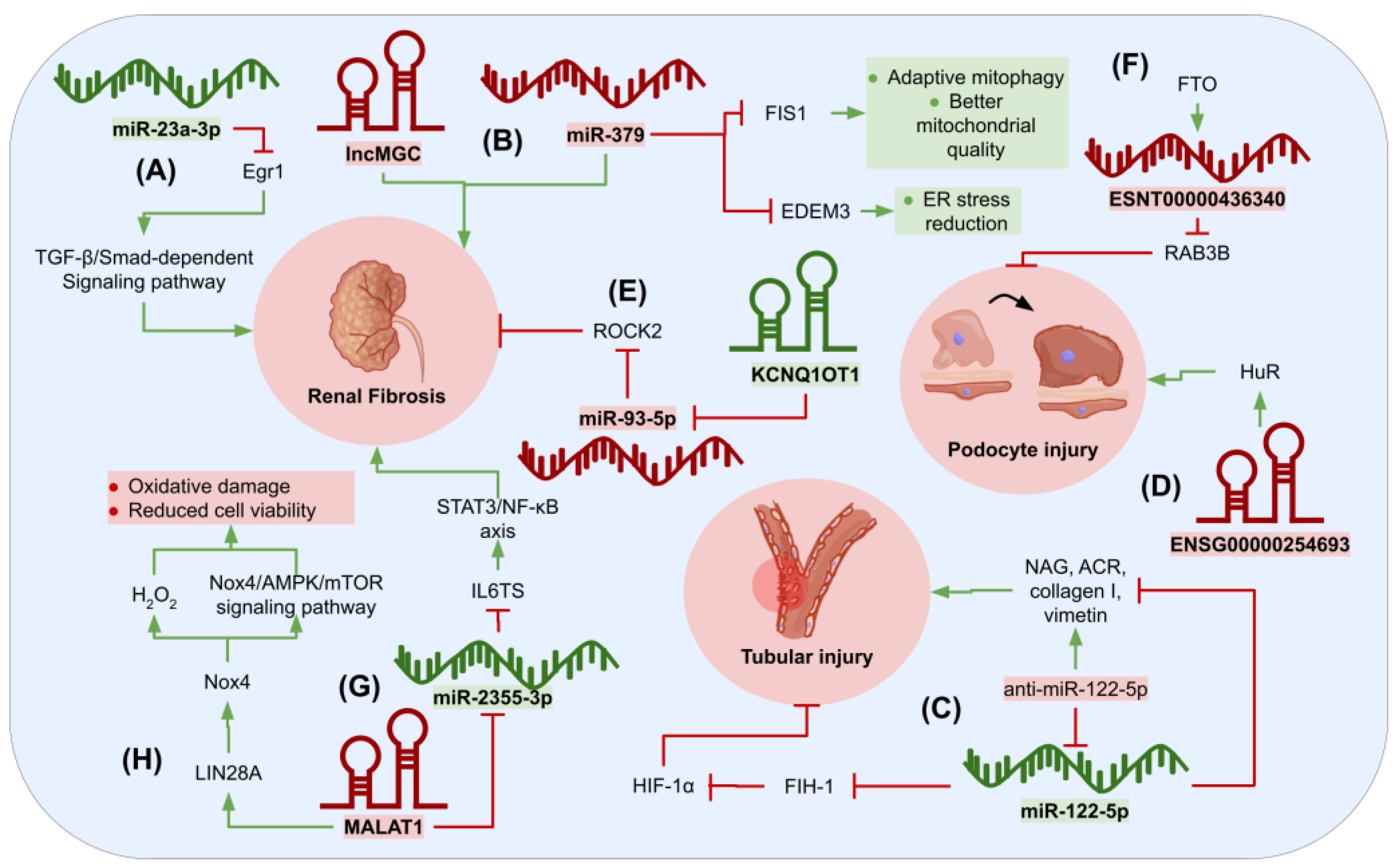{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Therapeutic Inferences of miRNAs and lncRNAs in Acute Kidney Injury
3. Therapeutic Inferences of miRNAs and lncRNAs in Chronic Kidney Disease
4. Therapeutic Inferences of miRNAs and lncRNAs in Diabetic Kidney Disease
5. Therapeutic Inferences of miRNAs and lncRNAs in Renal Fibrosis
6. Therapeutic Inferences of miRNAs and lncRNAs in Autosomal Dominant Polycystic Kidney Disease
7. Therapeutic Inferences of miRNAs and lncRNAs in Renal Cancer
8. Concluding Remarks
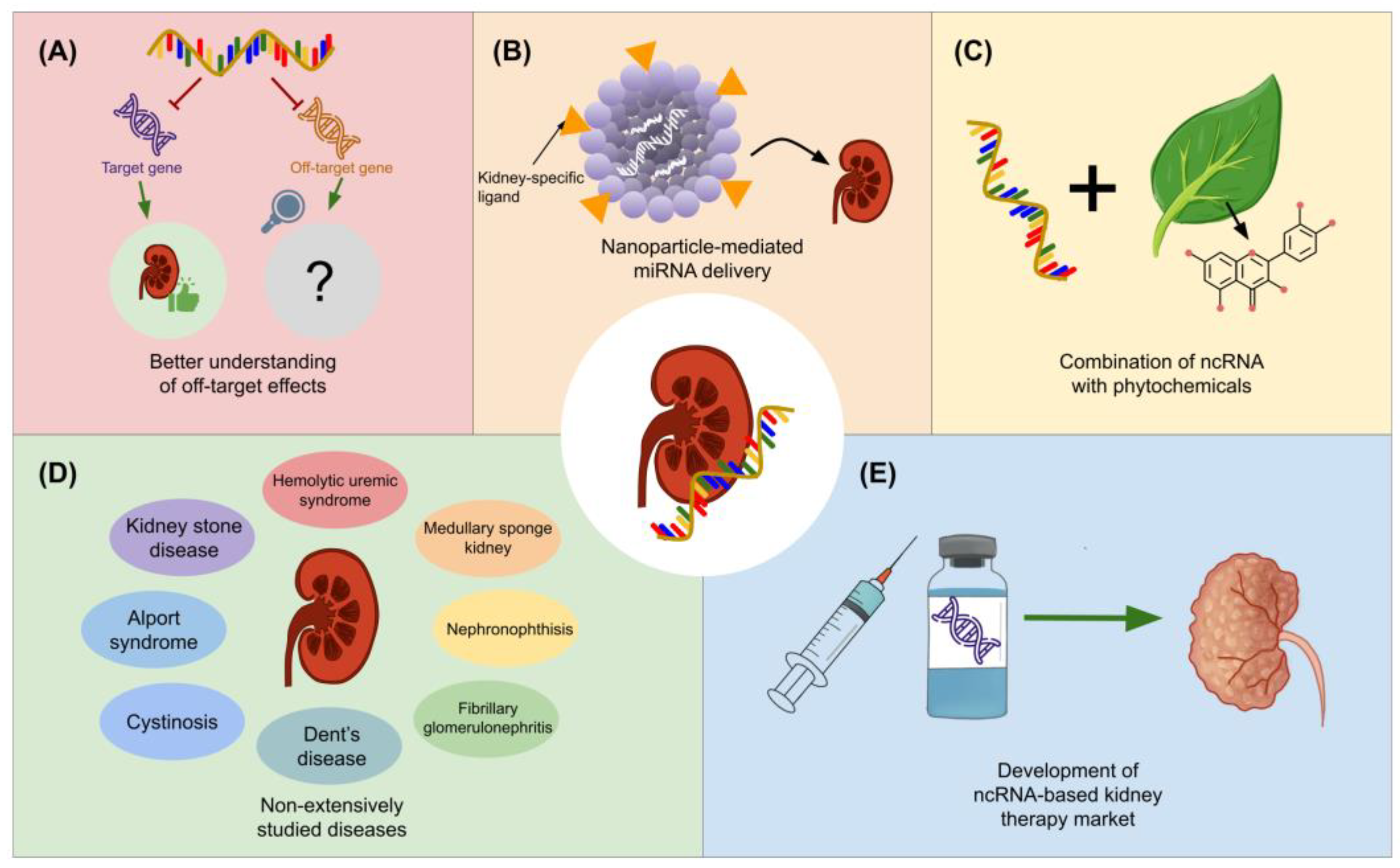
9. Future Insights
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Jourde-Chiche, N.; Fakhouri, F.; Dou, L.; Bellien, J.; Burtey, S.; Frimat, M.; Jarrot, P.A.; Kaplanski, G.; Le Quintrec, M.; Pernin, V.; et al. Endothelium structure and function in kidney health and disease. Nat. Rev. Nephrol. 2019, 15, 87–108. [Google Scholar] [CrossRef]
- Levey, A.S.; Coresh, J. Chronic kidney disease. Lancet 2012, 379, 165–180. [Google Scholar] [CrossRef]
- Syed-Ahmed, M.; Narayanan, M. Immune Dysfunction and Risk of Infection in Chronic Kidney Disease. Adv. Chronic Kidney Dis. 2019, 26, 8–15. [Google Scholar] [CrossRef]
- Cañadas-Garre, M.; Anderson, K.; Cappa, R.; Skelly, R.; Smyth, L.J.; McKnight, A.J.; Maxwell, A.P. Genetic susceptibility to chronic kidney disease—Some more pieces for the heritability puzzle. Front. Genet. 2019, 10, 453. [Google Scholar] [CrossRef]
- Knauf, F.; Brewer, J.R.; Flavell, R.A. Immunity, microbiota and kidney disease. Nat. Rev. Nephrol. 2019, 15, 263–274. [Google Scholar] [CrossRef]
- Ingrosso, D.; Perna, A.F. DNA Methylation Dysfunction in Chronic Kidney Disease. Genes 2020, 11, 811. [Google Scholar] [CrossRef]
- Luan, Z.L.; Zhang, C.; Ming, W.H.; Huang, Y.Z.; Guan, Y.F.; Zhang, X.Y. Nuclear receptors in renal health and disease. eBioMedicine 2022, 76, 103855. [Google Scholar] [CrossRef]
- Peired, A.J.; Lazzeri, E.; Guzzi, F.; Anders, H.J.; Romagnani, P. From kidney injury to kidney cancer. Kidney Int. 2021, 100, 55–66. [Google Scholar] [CrossRef]
- Faria, J.; Ahmed, S.; Gerritsen, K.G.F.; Mihaila, S.M.; Masereeuw, R. Kidney-based in vitro models for drug-induced toxicity testing. Arch. Toxicol. 2019, 93, 3397–3418. [Google Scholar] [CrossRef]
- Nimmagadda, S.M.; Agasthi, S.S.; Shai, A.; Khandavalli, D.K.R.; Vatti, J.R. Kidney Failure Detection and Predictive Analytics for ckd Using Machine Learning Procedures. Arch. Comput. Methods Eng. 2023, 30, 2341–2354. [Google Scholar] [CrossRef]
- Lea-Henry, T.N.; Carland, J.E.; Stocker, S.L.; Sevastos, J.; Roberts, D.M. Clinical pharmacokinetics in kidney disease: Fundamental principles. Clin. J. Am. Soc. Nephrol. 2018, 13, 1085–1095. [Google Scholar] [CrossRef]
- Evans, M.; Lopau, K. The transition clinic in chronic kidney disease care. Nephrol. Dial. Transplant. 2020, 35, ii4–ii10. [Google Scholar] [CrossRef]
- Ramaswamy, K.; Madariaga, H.M.; Thomas, B.S.; Lerma, E.V. Kidney transplantation for the primary care provider. Disease-a-Month 2020, 66, 100869. [Google Scholar] [CrossRef]
- Huang, X.; Ma, Y.; Li, Y.; Han, F.; Lin, W. Targeted Drug Delivery Systems for Kidney Diseases. Front. Bioeng. Biotechnol. 2021, 9, 683247. [Google Scholar] [CrossRef]
- Bian, J.; Liebert, A.; Bicknell, B.; Chen, X.M.; Huang, C.; Pollock, C.A. Faecal Microbiota Transplantation and Chronic Kidney Disease. Nutrients 2022, 14, 2528. [Google Scholar] [CrossRef]
- Borkar, P.; Yadav, V.; Tiwari, R.R.; Samarth, R.M. A systematic review of potential candidates of herbal medicine in treatment of chronic kidney disease. Phytomed. Plus 2022, 2, 100361. [Google Scholar] [CrossRef]
- Cao, Y.L.; Lin, J.H.; Hammes, H.P.; Zhang, C. Flavonoids in Treatment of Chronic Kidney Disease. Molecules 2022, 27, 2365. [Google Scholar] [CrossRef]
- Akinnibosun, O.A.; Maier, M.C.; Eales, J.; Tomaszewski, M.; Charchar, F.J. Telomere therapy for chronic kidney disease. Epigenomics 2022, 14, 1039–1054. [Google Scholar] [CrossRef]
- Tanemoto, F.; Mimura, I. Therapies Targeting Epigenetic Alterations in Acute Kidney Injury-to-Chronic Kidney Disease Transition. Pharmaceuticals 2022, 15, 123. [Google Scholar] [CrossRef]
- Foster, M.H.; Ord, J.R. Emerging immunotherapies for autoimmune kidney disease. Hum. Vaccines Immunother. 2019, 15, 876–890. [Google Scholar] [CrossRef]
- Fioretto, P.; Pontremoli, R. Expanding the therapy options for diabetic kidney disease. Nat. Rev. Nephrol. 2022, 18, 78–79. [Google Scholar] [CrossRef]
- Alicic, R.Z.; Johnson, E.J.; Tuttle, K.R. SGLT2 Inhibition for the Prevention and Treatment of Diabetic Kidney Disease: A Review. Am. J. Kidney Dis. 2018, 72, 267–277. [Google Scholar] [CrossRef]
- Bondue, T.; van den Heuvel, L.; Levtchenko, E.; Brock, R. The potential of RNA-based therapy for kidney diseases. Pediatr. Nephrol. 2022, 38, 327–344. [Google Scholar] [CrossRef]
- Kato, M. Noncoding RNAs as therapeutic targets in early stage diabetic kidney disease. Kidney Res. Clin. Pract. 2018, 37, 197–209. [Google Scholar] [CrossRef]
- Zheng, Q.; Reid, G.; Eccles, M.R.; Stayner, C. Non-coding RNAs as potential biomarkers and therapeutic targets in polycystic kidney disease. Front. Physiol. 2022, 13, 1006427. [Google Scholar] [CrossRef]
- van Zonneveld, A.J.; Zhao, Q.; Rotmans, J.I.; Bijkerk, R. Circulating non-coding RNAs in chronic kidney disease and its complications. Nat. Rev. Nephrol. 2023, 19, 573–586. [Google Scholar] [CrossRef]
- Yin, Y.; Shen, X. Noncoding RNA-chromatin association: Functions and mechanisms. Fundam. Res. 2023, 3, 665–675. [Google Scholar] [CrossRef]
- Paul, S.; Bravo Vázquez, L.A.; Pérez Uribe, S.; Manzanero Cárdenas, L.A.; Ruíz Aguilar, M.F.; Chakraborty, S.; Sharma, A. Roles of microRNAs in carbohydrate and lipid metabolism disorders and their therapeutic potential. Biochimie 2021, 187, 83–93. [Google Scholar] [CrossRef]
- Bravo-Vázquez, L.A.; Moreno Becerril, M.Y.; Mora Hernández, E.O.; de León Carmona, G.G.; Aguirre Padilla, M.E.; Chakraborty, S.; Bandyopadhyay, A.; Paul, S. The Emerging Role of MicroRNAs in Bone Diseases and Their Therapeutic Potential. Molecules 2022, 27, 211. [Google Scholar] [CrossRef]
- Paul, S.; Bravo Vázquez, L.A.; Reyes-Pérez, P.R.; Estrada-Meza, C.; Aponte Alburquerque, R.A.; Pathak, S.; Banerjee, A.; Bandyopadhyay, A.; Chakraborty, S.; Srivastava, A. The role of microRNAs in solving COVID-19 puzzle from infection to therapeutics: A mini-review. Virus Res. 2022, 308, 198631. [Google Scholar] [CrossRef]
- Estrada-Meza, C.; Torres-Copado, A.; Loreti González-Melgoza, L.; Ruiz-Manriquez, L.M.; De Donato, M.; Sharma, A.; Pathak, S.; Banerjee, A.; Paul, S. Recent insights into the microRNA and long non-coding RNA-mediated regulation of stem cell populations. 3 Biotech 2022, 12, 270. [Google Scholar] [CrossRef]
- Bravo-Vázquez, L.A.; Frías-Reid, N.; Ramos-Delgado, A.G.; Osorio-Pérez, S.M.; Zlotnik-Chávez, H.R.; Pathak, S.; Banerjee, A.; Bandyopadhyay, A.; Duttaroy, A.K.; Paul, S. MicroRNAs and long non-coding RNAs in pancreatic cancer: From epigenetics to potential clinical applications. Transl. Oncol. 2023, 27, 101579. [Google Scholar] [CrossRef]
- Bravo-Vázquez, L.A.; Méndez-García, A.; Rodríguez, A.L.; Sahare, P.; Pathak, S.; Banerjee, A.; Duttaroy, A.K.; Paul, S. Applications of nanotechnologies for miRNA-based cancer therapeutics: Current advances and future perspectives. Front. Bioeng. Biotechnol. 2023, 11, 1208547. [Google Scholar] [CrossRef]
- Samami, E.; Pourali, G.; Arabpour, M.; Fanipakdel, A.; Shahidsales, S.; Javadinia, S.A.; Hassanian, S.M.; Mohammadparast, S.; Avan, A. The Potential Diagnostic and Prognostic Value of Circulating MicroRNAs in the Assessment of Patients With Prostate Cancer: Rational and Progress. Front. Oncol. 2022, 11, 716831. [Google Scholar] [CrossRef]
- Grammatikakis, I.; Lal, A. Significance of lncRNA abundance to function. Mamm. Genome 2022, 33, 271–280. [Google Scholar] [CrossRef]
- Li, M.; Zhao, Y.; Li, H.; Deng, X.; Sheng, M. Application value of circulating LncRNA in diagnosis, treatment, and prognosis of breast cancer. Funct. Integr. Genom. 2023, 23, 61. [Google Scholar] [CrossRef]
- Wu, Y.; Li, Q.; Zhang, R.; Dai, X.; Chen, W.; Xing, D. Circulating microRNAs: Biomarkers of disease. Clin. Chim. Acta 2021, 516, 46–54. [Google Scholar] [CrossRef]
- López-Jiménez, E.; Andrés-León, E. The Implications of ncRNAs in the Development of Human Diseases. Non-Coding RNA 2021, 7, 17. [Google Scholar] [CrossRef]
- Bayraktar, E.; Bayraktar, R.; Oztatlici, H.; Lopez-Berestein, G.; Amero, P.; Rodriguez-Aguayo, C. Targeting miRNAs and Other Non-Coding RNAs as a Therapeutic Approach: An Update. Non-Coding RNA 2023, 9, 27. [Google Scholar] [CrossRef]
- Kolanska, K.; Bendifallah, S.; Canlorbe, G.; Mekinian, A.; Touboul, C.; Aractingi, S.; Chabbert-Buffet, N.; Daraï, E. Role of miRNAs in Normal Endometrium and in Endometrial Disorders: Comprehensive Review. J. Clin. Med. 2021, 10, 3457. [Google Scholar] [CrossRef]
- Smolarz, B.; Durczyński, A.; Romanowicz, H.; Szyłło, K.; Hogendorf, P. MiRNAs in Cancer (Review of Literature). Int. J. Mol. Sci. 2022, 23, 2805. [Google Scholar] [CrossRef] [PubMed]
- Lobera, E.S.; Varela, M.A.; Jimenez, R.L.; Moreno, R.B. MiRNA as biomarker in lung cancer. Mol. Biol. Rep. 2023, 50, 9521–9527. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Moya, J.M.; Vilella, F.; Simón, C. MicroRNA: Key gene expression regulators. Fertil. Steril. 2014, 101, 1516–1523. [Google Scholar] [CrossRef] [PubMed]
- Saliminejad, K.; Khorram Khorshid, H.R.; Soleymani Fard, S.; Ghaffari, S.H. An overview of microRNAs: Biology, functions, therapeutics, and analysis methods. J. Cell. Physiol. 2019, 234, 5451–5465. [Google Scholar] [CrossRef] [PubMed]
- Sen, C.K.; Ghatak, S. MiRNA Control of Tissue Repair and Regeneration. Am. J. Pathol. 2015, 185, 2629–2640. [Google Scholar] [CrossRef] [PubMed]
- Nejad, C.; Stunden, H.J.; Gantier, M.P. A guide to miRNAs in inflammation and innate immune responses. FEBS J. 2018, 285, 3695–3716. [Google Scholar] [CrossRef]
- Salas-Huetos, A.; James, E.R.; Aston, K.I.; Carrell, D.T.; Jenkins, T.G.; Yeste, M. The role of miRNAs in male human reproduction: A systematic review. Andrology 2020, 8, 7–26. [Google Scholar] [CrossRef]
- Salas-Huetos, A.; James, E.R.; Aston, K.I.; Jenkins, T.G.; Carrell, D.T.; Yeste, M. The Expression of miRNAs in Human Ovaries, Oocytes, Extracellular Vesicles, and Early Embryos: A Systematic Review. Cells 2019, 8, 1564. [Google Scholar] [CrossRef]
- Paul, S.; Bravo Vázquez, L.A.; Pérez Uribe, S.; Reyes-Pérez, P.R.; Sharma, A. Current Status of microRNA-Based Therapeutic Approaches in Neurodegenerative Disorders. Cells 2020, 9, 1698. [Google Scholar] [CrossRef]
- Bravo-Vázquez, L.A.; Medina-Ríos, I.; Márquez-Gallardo, L.D.; Reyes-Muñoz, J.; Serrano-Cano, F.I.; Pathak, S.; Banerjee, A.; Bandyopadhyay, A.; Duttaroy, A.K.; Paul, S. Functional Implications and Clinical Potential of MicroRNAs in Irritable Bowel Syndrome: A Concise Review. Dig. Dis. Sci. 2023, 68, 38–53. [Google Scholar] [CrossRef]
- Subramanian, S.; Steer, C.J. Special issue: MicroRNA regulation in health and disease. Genes 2019, 10, 457. [Google Scholar] [CrossRef] [PubMed]
- Peters, L.J.F.; Floege, J.; Biessen, E.A.L.; Jankowski, J.; van der Vorst, E.P.C. Micrornas in chronic kidney disease: Four candidates for clinical application. Int. J. Mol. Sci. 2020, 21, 6547. [Google Scholar] [CrossRef] [PubMed]
- Askari, H.; Raeis-Abdollahi, E.; Abazari, M.F.; Akrami, H.; Vakili, S.; Savardashtaki, A.; Tajbakhsh, A.; Sanadgol, N.; Azarnezhad, A.; Rahmati, L.; et al. Recent findings on the role of microRNAs in genetic kidney diseases. Mol. Biol. Rep. 2022, 49, 7039–7056. [Google Scholar] [CrossRef] [PubMed]
- Mahtal, N.; Lenoir, O.; Tinel, C.; Anglicheau, D.; Tharaux, P.L. MicroRNAs in kidney injury and disease. Nat. Rev. Nephrol. 2022, 18, 643–662. [Google Scholar] [CrossRef] [PubMed]
- Sun, P.; Wang, J.; Ilyasova, T.; Shumadalova, A.; Agaverdiev, M.; Wang, C. The function of miRNAs in the process of kidney development. Non-Coding RNA Res. 2023, 8, 593–601. [Google Scholar] [CrossRef] [PubMed]
- Schena, F.P.; Serino, G.; Sallustio, F. MicroRNAs in kidney diseases: New promising biomarkers for diagnosis and monitoring. Nephrol. Dial. Transplant. 2014, 29, 755–763. [Google Scholar] [CrossRef]
- Wang, C.; Wang, L.; Ding, Y.; Lu, X.; Zhang, G.; Yang, J.; Zheng, H.; Wang, H.; Jiang, Y.; Xu, L. LncRNA Structural Characteristics in Epigenetic Regulation. Int. J. Mol. Sci. 2017, 18, 2659. [Google Scholar] [CrossRef]
- Giroud, M.; Scheideler, M. Long Non-Coding RNAs in Metabolic Organs and Energy Homeostasis. Int. J. Mol. Sci. 2017, 18, 2578. [Google Scholar] [CrossRef]
- Ransohoff, J.D.; Wei, Y.; Khavari, P.A. The functions and unique features of long intergenic non-coding RNA. Nat. Rev. Mol. Cell Biol. 2018, 19, 143–157. [Google Scholar] [CrossRef]
- Bhartiya, D.; Jalali, S.; Ghosh, S.; Scaria, V. Distinct Patterns of Genetic Variations in Potential Functional Elements in Long Noncoding RNAs. Hum. Mutat. 2014, 35, 192–201. [Google Scholar] [CrossRef]
- Borkiewicz, L.; Kalafut, J.; Dudziak, K.; Przybyszewska-podstawka, A.; Telejko, I. Decoding LncRNAs. Cancers 2021, 13, 2643. [Google Scholar] [CrossRef] [PubMed]
- Fatica, A.; Bozzoni, I. Long non-coding RNAs: New players in cell differentiation and development. Nat. Rev. Genet. 2014, 15, 7–21. [Google Scholar] [CrossRef] [PubMed]
- Jiang, N.; Zhang, X.; Gu, X.; Li, X.; Shang, L. Progress in understanding the role of lncRNA in programmed cell death. Cell Death Discov. 2021, 7, 30. [Google Scholar] [CrossRef]
- Chen, Y.G.; Satpathy, A.T.; Chang, H.Y. Gene regulation in the immune system by long noncoding RNAs. Nat. Immunol. 2017, 18, 962–972. [Google Scholar] [CrossRef] [PubMed]
- Patil, S.; Dang, K.; Zhao, X.; Gao, Y.; Qian, A. Role of LncRNAs and CircRNAs in Bone Metabolism and Osteoporosis. Front. Genet. 2020, 11, 584118. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Fang, Z.; Hong, M.; Yang, D.; Xie, W. Long-noncoding RNAs (lncRNAs) in drug metabolism and disposition, implications in cancer chemo-resistance. Acta Pharm. Sin. B 2020, 10, 105–112. [Google Scholar] [CrossRef]
- Kamalidehghan, B.; Habibi, M.; Afjeh, S.S.; Shoai, M.; Alidoost, S.; Ghale, R.A.; Eshghifar, N.; Pouresmaeili, F. The Importance of Small Non-Coding RNAs in Human Reproduction: A Review Article. Appl. Clin. Genet. 2020, 13, 1–11. [Google Scholar] [CrossRef]
- Leti, F.; DiStefano, J.K. Long Noncoding RNAs as Diagnostic and Therapeutic Targets in Type 2 Diabetes and Related Complications. Genes 2017, 8, 207. [Google Scholar] [CrossRef]
- Good, D.J. Non-Coding RNAs in Human Health and Diseases. Genes 2023, 14, 1429. [Google Scholar] [CrossRef]
- Ignarski, M.; Islam, R.; Müller, R.-U. Long Non-Coding RNAs in Kidney Disease. Int. J. Mol. Sci. 2019, 20, 3276. [Google Scholar] [CrossRef]
- Gu, Y.Y.; Lu, F.H.; Huang, X.R.; Zhang, L.; Mao, W.; Yu, X.Q.; Liu, X.S.; Lan, H.Y. Non-Coding RNAs as Biomarkers and Therapeutic Targets for Diabetic Kidney Disease. Front. Pharmacol. 2021, 11, 583528. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.N.; Yang, C.E.; Zhang, D.D.; Chen, Y.Y.; Yu, X.Y.; Zhao, Y.Y.; Miao, H. Long non-coding RNAs: A double-edged sword in aging kidney and renal disease. Chem. Biol. Interact. 2021, 337, 109396. [Google Scholar] [CrossRef] [PubMed]
- Moreno, J.A.; Hamza, E.; Guerrero-Hue, M.; Rayego-Mateos, S.; García-Caballero, C.; Vallejo-Mudarra, M.; Metzinger, L.; Metzinger-Le Meuth, V. Non-Coding RNAs in Kidney Diseases: The Long and Short of Them. Int. J. Mol. Sci. 2021, 22, 6077. [Google Scholar] [CrossRef] [PubMed]
- Mao, M.; Zhang, J.; Xiang, Y.; Gong, M.; Deng, Y.; Ye, D. Role of exosomal competitive endogenous RNA (ceRNA) in diagnosis and treatment of malignant tumors. Bioengineered 2022, 13, 12156–12168. [Google Scholar] [CrossRef] [PubMed]
- Braga, E.A.; Fridman, M.V.; Burdennyy, A.M.; Filippova, E.A.; Loginov, V.I.; Pronina, I.V.; Dmitriev, A.A.; Kushlinskii, N.E. Regulation of the Key Epithelial Cancer Suppressor miR-124 Function by Competing Endogenous RNAs. Int. J. Mol. Sci. 2022, 23, 13620. [Google Scholar] [CrossRef] [PubMed]
- Ma, B.; Wang, S.; Wu, W.; Shan, P.; Chen, Y.; Meng, J.; Xing, L.; Yun, J.; Hao, L.; Wang, X.; et al. Mechanisms of circRNA/lncRNA-miRNA interactions and applications in disease and drug research. Biomed. Pharmacother. 2023, 162, 114672. [Google Scholar] [CrossRef] [PubMed]
- Kapadia, C.H.; Luo, B.; Dang, M.N.; Irvin-Choy, N.; Valcourt, D.M.; Day, E.S. Polymer nanocarriers for MicroRNA delivery. J. Appl. Polym. Sci. 2020, 137, 48651. [Google Scholar] [CrossRef]
- Szczepanek, J.; Skorupa, M.; Jarkiewicz-Tretyn, J.; Cybulski, C.; Tretyn, A. Harnessing Epigenetics for Breast Cancer Therapy: The Role of DNA Methylation, Histone Modifications, and MicroRNA. Int. J. Mol. Sci. 2023, 24, 7235. [Google Scholar] [CrossRef]
- Garbo, S.; Maione, R.; Tripodi, M.; Battistelli, C. Next RNA Therapeutics: The Mine of Non-Coding. Int. J. Mol. Sci. 2022, 23, 7471. [Google Scholar] [CrossRef]
- Haemmerle, M.; Gutschner, T. Long Non-Coding RNAs in Cancer and Development: Where Do We Go from Here? Int. J. Mol. Sci. 2015, 16, 1395–1405. [Google Scholar] [CrossRef]
- Makris, K.; Spanou, L. Acute Kidney Injury: Definition, Pathophysiology and Clinical Phenotypes. Clin. Biochem. Rev. 2016, 37, 85–98. [Google Scholar]
- Zuk, A.; Bonventre, J.V. Recent Advances on Acute Kidney Injury and its Consequences and Impact on Chronic Kidney Disease. Curr. Opin. Nephrol. Hypertens. 2019, 28, 397–405. [Google Scholar] [CrossRef] [PubMed]
- Ge, Q.M.; Huang, C.M.; Zhu, X.Y.; Bian, F.; Pan, S.M. Differentially expressed miRNAs in sepsisinduced acute kidney injury target oxidative stress and mitochondrial dysfunction pathways. PLoS ONE 2017, 12, e0173292. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Wang, B.; Ma, L.; Fu, P. An Update of Long-Noncoding RNAs in Acute Kidney Injury. Front. Physiol. 2022, 13, 849403. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Li, Y.; Zhu, X.; Jiang, C. MicroRNA as an early diagnostic biomarker for contrast-induced acute kidney injury. Drug Chem. Toxicol. 2022, 45, 1552–1557. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Zhao, Y.; Wang, L.; Zhang, W.; Liu, C.; Yin, A. MicroRNA-194 overexpression protects against hypoxia/reperfusion-induced HK-2 cell injury through direct targeting Rheb. J. Cell. Biochem. 2019, 120, 8311–8318. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Huang, H.; Liu, W.; Liu, S.; Wang, X.Y.; Diao, Z.L.; Zhang, A.H.; Guo, W.; Han, X.; Dong, X.; et al. Endothelial progenitor cells-derived exosomal microRNA-21-5p alleviates sepsis-induced acute kidney injury by inhibiting RUNX1 expression. Cell Death Dis. 2021, 12, 335. [Google Scholar] [CrossRef] [PubMed]
- Jia, P.; Xu, S.; Ren, T.; Pan, T.; Wang, X.; Zhang, Y.; Zou, Z.; Guo, M.; Zeng, Q.; Shen, B.; et al. LncRNA IRAR regulates chemokines production in tubular epithelial cells thus promoting kidney ischemia-reperfusion injury. Cell Death Dis. 2022, 13, 562. [Google Scholar] [CrossRef]
- Yang, N.; Wang, H.; Zhang, L.; Lv, J.; Niu, Z.; Liu, J.; Zhang, Z. Long non-coding RNA SNHG14 aggravates LPS-induced acute kidney injury through regulating miR-495-3p/HIPK1. Acta Biochim. Biophys. Sin. 2021, 53, 719–728. [Google Scholar] [CrossRef]
- Wang, M.; Wei, J.; Shang, F.; Zang, K.; Zhang, P. Down-regulation of lncRNA SNHG5 relieves sepsis-induced acute kidney injury by regulating the miR-374a-3p/TLR4/NF-κB pathway. J. Biochem. 2021, 169, 575–583. [Google Scholar] [CrossRef]
- Hu, Q.; Zen, W.; Zhang, M.; Wang, Z.; Cui, W.; Liu, Y.; Xu, B. Long Non-Coding RNA CASC2 Overexpression Ameliorates Sepsis-Associated Acute Kidney Injury by Regulating MiR-545-3p/PPARA axis. J. Surg. Res. 2021, 265, 223–232. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Li, X.; Qiu, S.; Wang, Y.; Zhang, D. LncRNA 122049 suppresses apoptosis of renal tubular epithelial cells in ischemic AKI by targeting the miR-330-5p/ELK1 axis. FASEB J. 2022, 36, e22395. [Google Scholar] [CrossRef] [PubMed]
- Xie, Z.; Wei, L.; Chen, J.; Chen, Z. LncRNA NORAD deficiency alleviates kidney injury in mice and decreases the inflammatory response and apoptosis of lipopolysaccharide-stimulated HK-2 cells via the miR-577/GOLPH3 axis. Cytokine 2022, 153, 155844. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Chen, M.; Guo, Q.; Shen, L.; Liu, X.; Pan, J.; Zhang, Y.; Xu, T.; Zhang, D.; Wei, G. Human umbilical cord mesenchymal stem cell exosome-derived miR-874-3p targeting RIPK1/PGAM5 attenuates kidney tubular epithelial cell damage. Cell. Mol. Biol. Lett. 2023, 28, 12. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Xu, C. LncRNA MALAT1-deficiency restrains lipopolysaccharide (LPS)-induced pyroptotic cell death and inflammation in HK-2 cells by releasing microRNA-135b-5p. Ren. Fail. 2021, 43, 1288–1297. [Google Scholar] [CrossRef] [PubMed]
- Younes, S.; Mourad, N.; Safwan, J.; Dabbous, M.; Rahal, M.; Al Nabulsi, M.; Sakr, F. Chronic kidney disease awareness among the general population: Tool validation and knowledge assessment in a developing country. BMC Nephrol. 2022, 23, 266. [Google Scholar] [CrossRef]
- Webster, A.C.; Nagler, E.V.; Morton, R.L.; Masson, P. Chronic Kidney Disease. Lancet 2017, 389, 1238–1252. [Google Scholar] [CrossRef]
- Noble, R.; Taal, M.W. Epidemiology and causes of chronic kidney disease. Medicine 2019, 47, 562–566. [Google Scholar] [CrossRef]
- Shelke, V.; Kale, A.; Sankrityayan, H.; Anders, H.J.; Gaikwad, A.B. Long non-coding RNAs as emerging regulators of miRNAs and epigenetics in diabetes-related chronic kidney disease. Arch. Physiol. Biochem. 2022. [Google Scholar] [CrossRef]
- Bao, J.; Lu, Y.; She, Q.; Dou, W.; Tang, R.; Xu, X.; Zhang, M.; Zhu, L.; Zhou, Q.; Li, H.; et al. MicroRNA-30 regulates left ventricular hypertrophy in chronic kidney disease. JCI Insight 2021, 6, e138027. [Google Scholar] [CrossRef]
- Jin, J.; Qian, F.; Zheng, D.; He, W.; Gong, J.; He, Q. Mesenchymal Stem Cells Attenuate Renal Fibrosis via Exosomes-Mediated Delivery of microRNA Let-7i-5p Antagomir. Int. J. Nanomed. 2021, 16, 3565–3578. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Guo, Y.; Bao, S.; Huang, H.; Liu, W.; Guo, W. Bone marrow mesenchymal stem cell-derived exosomal microRNA-381-3p alleviates vascular calcification in chronic kidney disease by targeting NFAT5. Cell Death Dis. 2022, 13, 278. [Google Scholar] [CrossRef] [PubMed]
- Hussain, S.; Chand Jamali, M.; Habib, A.; Hussain, M.S.; Akhtar, M.; Najmi, A.K. Diabetic kidney disease: An overview of prevalence, risk factors, and biomarkers. Clin. Epidemiol. Glob. Health 2021, 9, 2–6. [Google Scholar] [CrossRef]
- Levey, A.S.; Grams, M.E.; Inker, L.A. Uses of GFR and Albuminuria Level in Acute and Chronic Kidney Disease. N. Engl. J. Med. 2022, 386, 2120–2128. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Fernandez, N.; Jacobs-Cachá, C.; Mora-Gutiérrez, J.M.; Vergara, A.; Orbe, J.; Soler, M.J. Matrix Metalloproteinases in Diabetic Kidney Disease. J. Clin. Med. 2020, 9, 472. [Google Scholar] [CrossRef] [PubMed]
- Lv, J.; Wu, Y.; Mai, Y.; Bu, S. Noncoding RNAs in Diabetic Nephropathy: Pathogenesis, Biomarkers, and Therapy. J. Diabetes Res. 2020, 2020, 3960857. [Google Scholar] [CrossRef]
- Srivastava, S.P.; Goodwin, J.E.; Tripathi, P.; Kanasaki, K.; Koya, D. Interactions among Long Non-Coding RNAs and microRNAs Influence Disease Phenotype in Diabetes and Diabetic Kidney Disease. Int. J. Mol. Sci. 2021, 22, 6027. [Google Scholar] [CrossRef]
- Sheng, S.; Zou, M.; Yang, Y.; Guan, M.; Ren, S.; Wang, X.; Wang, L.; Xue, Y. MiR-23a-3p regulates the inflammatory response and fibrosis in diabetic kidney disease by targeting early growth response 1. Vitr. Cell. Dev. Biol. Anim. 2021, 57, 763–774. [Google Scholar] [CrossRef]
- Kato, M.; Abdollahi, M.; Tunduguru, R.; Tsark, W.; Chen, Z.; Wu, X.; Wang, J.; Chen, Z.B.; Lin, F.M.; Lanting, L.; et al. MiR-379 deletion ameliorates features of diabetic kidney disease by enhancing adaptive mitophagy via FIS1. Commun. Biol. 2021, 4, 30. [Google Scholar] [CrossRef]
- Cheng, L.; Qiu, X.; He, L.; Liu, L. MicroRNA-122-5p ameliorates tubular injury in diabetic nephropathy via FIH-1/HIF-1α pathway. Ren. Fail. 2022, 44, 293–303. [Google Scholar] [CrossRef]
- Yu, Q.; Lin, J.; Ma, Q.; Li, Y.; Wang, Q.; Chen, H.; Liu, Y.; Liu, B. Long Noncoding RNA ENSG00000254693 Promotes Diabetic Kidney Disease via Interacting with HuR. J. Diabetes Res. 2022, 2022, 8679548. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.; Fu, P.; Ma, L. Kidney fibrosis: From mechanisms to therapeutic medicines. Signal Transduct. Target. Ther. 2023, 8, 129. [Google Scholar] [CrossRef] [PubMed]
- Kalantar-Zadeh, K.; Jafar, T.H.; Nitsch, D.; Neuen, B.L.; Perkovic, V. Chronic kidney disease. Lancet 2021, 398, 786–802. [Google Scholar] [CrossRef]
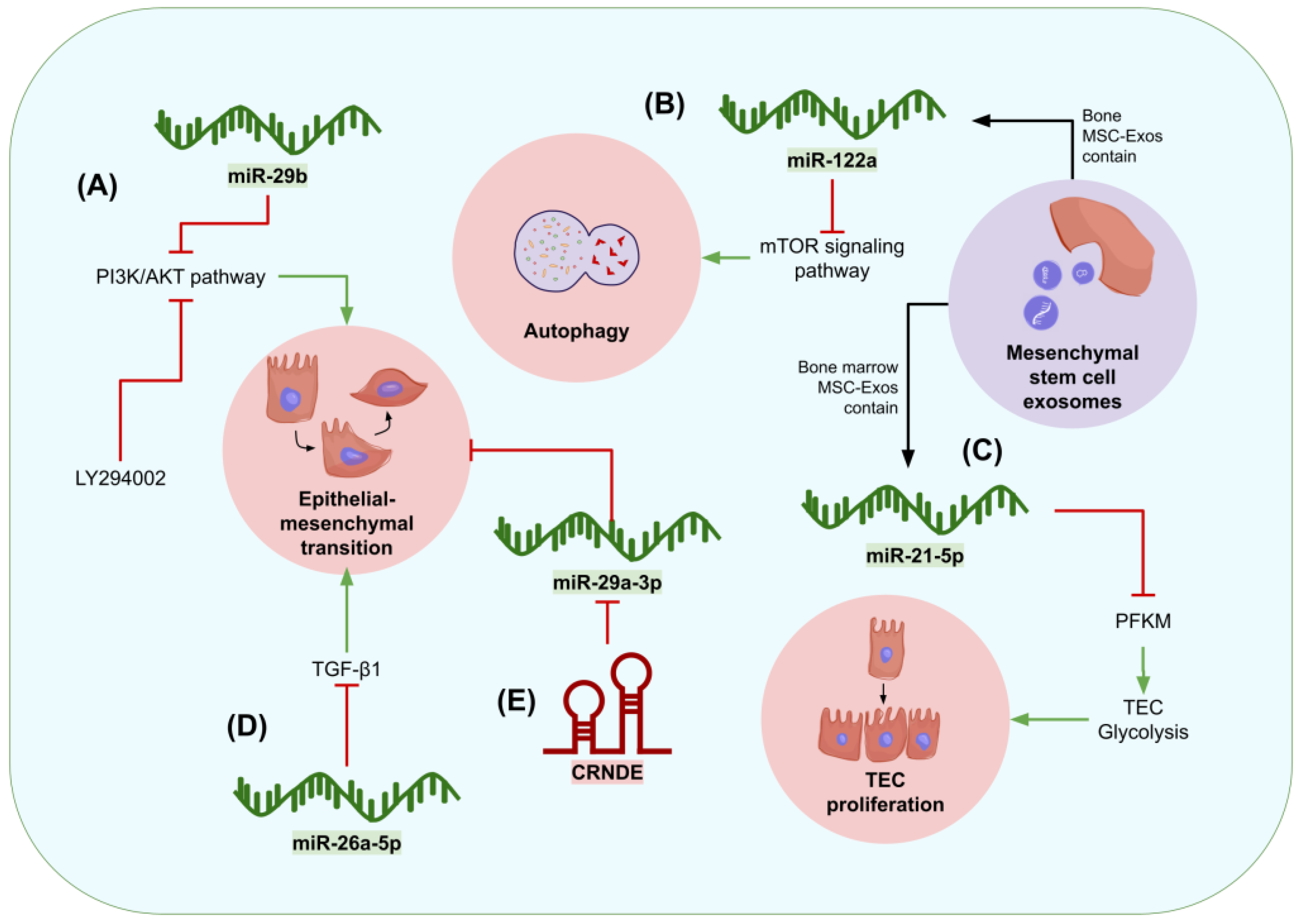
- Hu, S.; Hu, H.; Wang, R.; He, H.; Shui, H. MicroRNA-29b prevents renal fibrosis by attenuating renal tubular epithelial cell–mesenchymal transition through targeting the PI3K/AKT pathway. Int. Urol. Nephrol. 2021, 53, 1941–1950. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Qu, J.; Yuan, X.; Zhuang, S.; Wu, H.; Chen, R.; Wu, J.; Zhang, M.; Ying, L. Mesenchymal Stem Cells Alleviate Renal Fibrosis and Inhibit Autophagy via Exosome Transfer of miRNA-122a. Stem Cells Int. 2022, 2022, 1981798. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Cheuk, Y.C.; Jia, Y.; Chen, T.; Chen, J.; Luo, Y.; Cao, Y.; Guo, J.; Dong, L.; Zhang, Y.; et al. Bone marrow mesenchymal stem cell-derived exosomal miR-21a-5p alleviates renal fibrosis by attenuating glycolysis by targeting PFKM. Cell Death Dis. 2022, 13, 876. [Google Scholar] [CrossRef]
- Zhao, M.; Li, N.; Wan, C.; Zhang, Q.; Wang, H.; Jiang, C. LncRNA CRNDE is involved in the pathogenesis of renal fibrosis by regulating renal epithelial cell mesenchymal-epithelial transition via targeting miR-29a-3p. Mutat. Res. Fundam. Mol. Mech. Mutagen. 2023, 826, 111817. [Google Scholar] [CrossRef]
- Chung, Y.H.; Huang, G.K.; Kang, C.H.; Cheng, Y.T.; Kao, Y.H.; Chien, Y.S. MicroRNA-26a-5p Restoration Ameliorates Unilateral Ureteral Obstruction-Induced Renal Fibrosis in Mice Through Modulating TGF-β Signaling. Lab. Investig. 2023, 103, 100131. [Google Scholar] [CrossRef]
- Balsam, L.B. Commentary: Antifibrotic medications and wound healing after cardiac surgery. JTCVS Tech. 2021, 6, 104–105. [Google Scholar] [CrossRef]
- Malekshahabi, T.; Khoshdel Rad, N.; Serra, A.L.; Moghadasali, R. Autosomal dominant polycystic kidney disease: Disrupted pathways and potential therapeutic interventions. J. Cell. Physiol. 2019, 234, 12451–12470. [Google Scholar] [CrossRef]
- Reiterová, J.; Tesař, V. Autosomal Dominant Polycystic Kidney Disease: From Pathophysiology of Cystogenesis to Advances in the Treatment. Int. J. Mol. Sci. 2022, 23, 3317. [Google Scholar] [CrossRef] [PubMed]
- Simms, R.J.; Thong, K.M.; Dworschak, G.C.; Ong, A.C.M. Increased psychosocial risk, depression and reduced quality of life living with autosomal dominant polycystic kidney disease. Nephrol. Dial. Transplant. 2016, 31, 1130–1140. [Google Scholar] [CrossRef] [PubMed]
- Eriksson, D.; Karlsson, L.; Eklund, O.; Dieperink, H.; Honkanen, E.; Melin, J.; Selvig, K.; Lundberg, J. Health-related quality of life across all stages of autosomal dominant polycystic kidney disease. Nephrol. Dial. Transplant. 2017, 32, 2106–2111. [Google Scholar] [CrossRef] [PubMed]
- Yarlioglu, A.M.; Oguz, E.G.; Gundogmus, A.G.; Atilgan, K.G.; Sahin, H.; Ayli, M.D. The relationship between depression, anxiety, quality of life levels, and the chronic kidney disease stage in the autosomal dominant polycystic kidney disease. Int. Urol. Nephrol. 2023, 55, 983–992. [Google Scholar] [CrossRef] [PubMed]
- Bais, T.; Gansevoort, R.T.; Meijer, E. Drugs in Clinical Development to Treat Autosomal Dominant Polycystic Kidney Disease. Drugs 2022, 82, 1095–1115. [Google Scholar] [CrossRef] [PubMed]
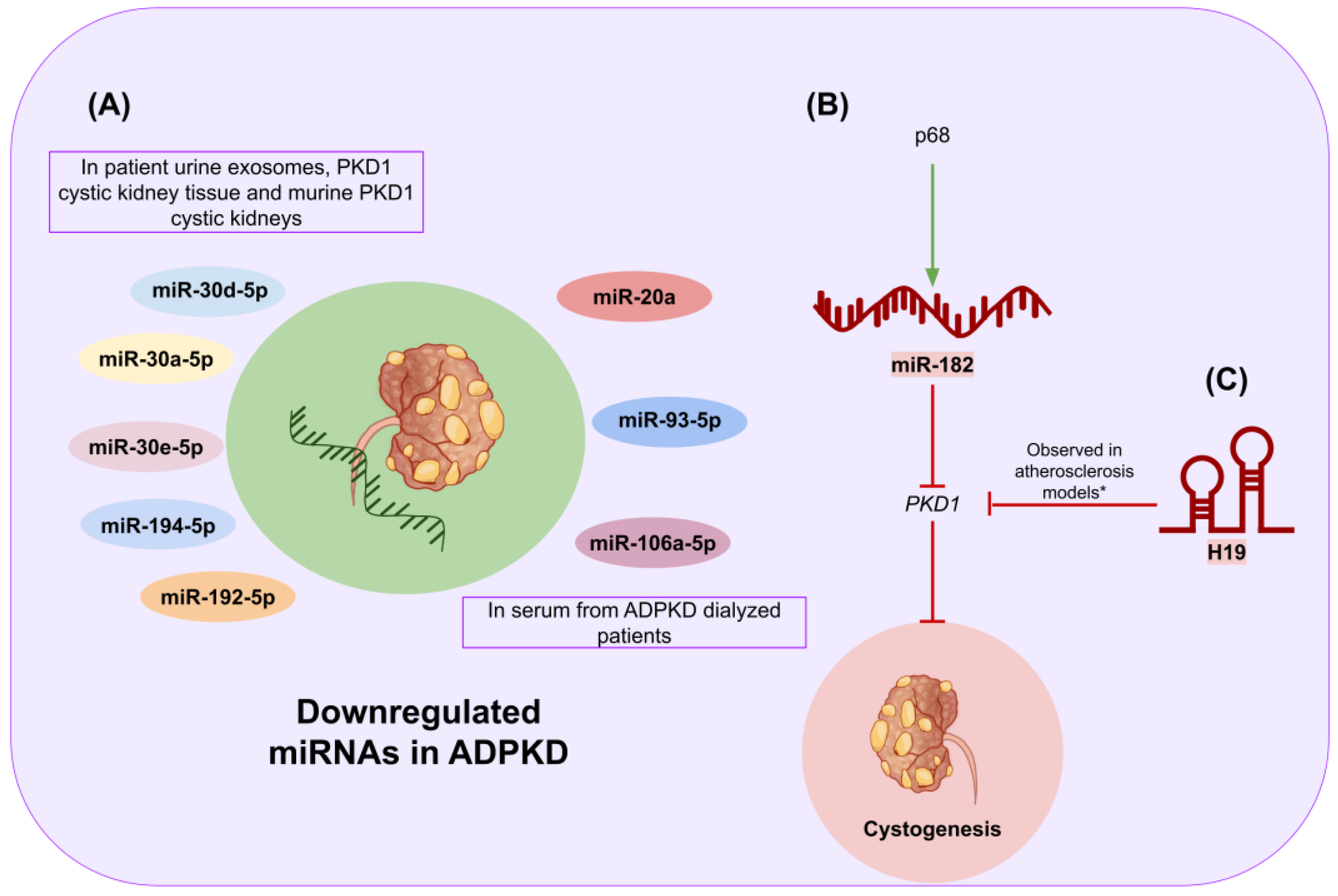
- Magayr, T.A.; Song, X.; Streets, A.J.; Vergoz, L.; Chang, L.; Valluru, M.K.; Yap, H.L.; Lannoy, M.; Haghighi, A.; Simms, R.J.; et al. Global microRNA profiling in human urinary exosomes reveals novel disease biomarkers and cellular pathways for autosomal dominant polycystic kidney disease. Kidney Int. 2020, 98, 420–435. [Google Scholar] [CrossRef] [PubMed]
- Małachowska, B.; Tkaczyk, M.; Chrul, S.; Zwiech, R.; Młynarski, W.; Fendler, W. Serum microRNA profiles in patients with autosomal dominant polycystic kidney disease show systematic dysregulation partially reversible by hemodialysis. Arch. Med. Sci. 2019, 17, 1730–1741. [Google Scholar] [CrossRef]
- Zhang, L.; Li, L.X.; Zhou, J.X.; Harris, P.C.; Calvet, J.P.; Li, X. RNA helicase p68 inhibits the transcription and post-transcription of Pkd1 in ADPKD. Theranostics 2020, 10, 8281–8297. [Google Scholar] [CrossRef]
- Yang, Y.; Tang, F.; Wei, F.; Yang, L.; Kuang, C.; Zhang, H.; Deng, J.; Wu, Q. Silencing of long non-coding RNA H19 downregulates CTCF to protect against atherosclerosis by upregulating PKD1 expression in ApoE knockout mice. Aging 2019, 11, 10016–10030. [Google Scholar] [CrossRef]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA. Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef]
- Hsieh, J.J.; Purdue, M.P.; Signoretti, S.; Swanton, C.; Albiges, L.; Schmidinger, M.; Heng, D.Y.; Larkin, J.; Ficarra, V. Renal cell carcinoma. Nat. Rev. Dis. Prim. 2017, 3, 17009. [Google Scholar] [CrossRef] [PubMed]
- Bergerot, C.D.; Clark, K.L.; Ashing, K.T.; Bergerot, P.G.; Obenchain, R.; Dizman, N.; Hsu, J.A.; Philip, E.; Loscalzo, M.; Pal, S.K. Biopsychosocial distress and clinical outcome in metastatic renal cell carcinoma. Palliat. Support. Care 2019, 17, 353–355. [Google Scholar] [CrossRef] [PubMed]
- Vartolomei, L.; Cotruș, A.; Stanciu, C.; Delcea, C.; Tozzi, M.; Lievore, E.; Crocetto, F.; Del Giudice, F.; Lucarelli, G.; Muto, M.; et al. Quality of Life and Psychological Distress among Patients with Small Renal Masses. J. Clin. Med. 2022, 11, 3944. [Google Scholar] [CrossRef] [PubMed]
- He, Y.H.; Chen, C.; Shi, Z. The biological roles and clinical implications of microRNAs in clear cell renal cell carcinoma. J. Cell. Physiol. 2018, 233, 4458–4465. [Google Scholar] [CrossRef]
- Liu, X.; Hao, Y.; Yu, W.; Yang, X.; Luo, X.; Zhao, J.; Li, J.; Hu, X.; Li, L. Long Non-Coding RNA Emergence during Renal Cell Carcinoma Tumorigenesis. Cell. Physiol. Biochem. 2018, 47, 735–746. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Jin, S.; Xiao, W.; Zhu, X.; Jia, C.; Lin, Z. Long noncoding RNA LINC00641 promotes renal cell carcinoma progression via sponging microRNA-340-5p. Cancer Cell Int. 2021, 21, 210. [Google Scholar] [CrossRef] [PubMed]
- Guo, R.; Zou, B.; Liang, Y.; Bian, J.; Xu, J.; Zhou, Q.; Zhang, C.; Chen, T.; Yang, M.; Wang, H.; et al. LncRNA RCAT1 promotes tumor progression and metastasis via miR-214-5p/E2F2 axis in renal cell carcinoma. Cell Death Dis. 2021, 12, 689. [Google Scholar] [CrossRef]
- Hu, G.; Ma, J.; Zhang, J.; Chen, Y.; Liu, H.; Huang, Y.; Zheng, J.; Xu, Y.; Xue, W.; Zhai, W. Hypoxia-induced lncHILAR promotes renal cancer metastasis via ceRNA for the miR-613/206/1-1-3p/Jagged-1/Notch/CXCR4 signaling pathway. Mol. Ther. 2021, 29, 2979–2994. [Google Scholar] [CrossRef]
- Qin, J.; Zhu, T.; Wu, W.; Chen, H.; He, Y. Long Non-Coding RNA PCED1B-AS1 Promotes the Progression of Clear Cell Renal Cell Carcinoma Through miR-484/ZEB1 Axis. Onco. Targets. Ther. 2021, 14, 393–402. [Google Scholar] [CrossRef]
- Wang, D.; Zhu, X.; Siqin, B.; Ren, C.; Yi, F. Long non-coding RNA CYTOR modulates cancer progression through miR-136-5p/MAT2B axis in renal cell carcinoma. Toxicol. Appl. Pharmacol. 2022, 447, 116067. [Google Scholar] [CrossRef]
- Hueso, M.; Mallén, A.; Suñé-Pou, M.; Aran, J.M.; Suñé-Negre, J.M.; Navarro, E. NcRNAs in Therapeutics: Challenges and Limitations in Nucleic Acid-Based Drug Delivery. Int. J. Mol. Sci. 2021, 22, 11596. [Google Scholar] [CrossRef] [PubMed]
- Herkt, M.; Thum, T. Pharmacokinetics and Proceedings in Clinical Application of Nucleic Acid Therapeutics. Mol. Ther. 2021, 29, 521–539. [Google Scholar] [CrossRef] [PubMed]
- Bartolucci, D.; Pession, A.; Hrelia, P.; Tonelli, R. Precision Anti-Cancer Medicines by Oligonucleotide Therapeutics in Clinical Research Targeting Undruggable Proteins and Non-Coding RNAs. Pharmaceutics 2022, 14, 1453. [Google Scholar] [CrossRef] [PubMed]
- Xing, T.; He, H. Epigenomics of clear cell renal cell carcinoma: Mechanisms and potential use in molecular pathology. Chin. J. Cancer Res. 2016, 28, 80–91. [Google Scholar] [CrossRef] [PubMed]
- Blokhin, I.; Khorkova, O.; Hsiao, J.; Wahlestedt, C. Developments in lncRNA drug discovery: Where are we heading? Expert Opin. Drug Discov. 2018, 13, 837–849. [Google Scholar] [CrossRef] [PubMed]
- Alzhrani, R.; Alsaab, H.O.; Petrovici, A.; Bhise, K.; Vanamala, K.; Sau, S.; Krinock, M.J.; Iyer, A.K. Improving the therapeutic efficiency of noncoding RNAs in cancers using targeted drug delivery systems. Drug Discov. Today 2020, 25, 718–730. [Google Scholar] [CrossRef] [PubMed]
- Yu, A.M.; Choi, Y.H.; Tu, M.J. RNA Drugs and RNA Targets for Small Molecules: Principles, Progress, and Challenges. Pharmacol. Rev. 2020, 72, 862–898. [Google Scholar] [CrossRef]
- Huang, Y.; Jiang, K.; Zhang, X.; Chung, E.J. The effect of size, charge, and peptide ligand length on kidney targeting by small, organic nanoparticles. Bioeng. Transl. Med. 2020, 5, e10173. [Google Scholar] [CrossRef]
- Ma, Y.; Cai, F.; Li, Y.; Chen, J.; Han, F.; Lin, W. A review of the application of nanoparticles in the diagnosis and treatment of chronic kidney disease. Bioact. Mater. 2020, 5, 732–743. [Google Scholar] [CrossRef]
- Liu, C.P.; Hu, Y.; Lin, J.C.; Fu, H.L.; Lim, L.Y.; Yuan, Z.X. Targeting strategies for drug delivery to the kidney: From renal glomeruli to tubules. Med. Res. Rev. 2019, 39, 561–578. [Google Scholar] [CrossRef]
- Kang, J.Y.; Kim, H.; Mun, D.; Yun, N.; Joung, B. Co-delivery of curcumin and miRNA-144-3p using heart-targeted extracellular vesicles enhances the therapeutic efficacy for myocardial infarction. J. Control. Release 2021, 331, 62–73. [Google Scholar] [CrossRef] [PubMed]
- Basist, P.; Parveen, B.; Zahiruddin, S.; Gautam, G.; Parveen, R.; Khan, M.A.; Krishnan, A.; Shahid, M.; Ahmad, S. Potential nephroprotective phytochemicals: Mechanism and future prospects. J. Ethnopharmacol. 2022, 283, 114743. [Google Scholar] [CrossRef] [PubMed]
- Hashem, K.S.; Abdelazem, A.Z.; Mohammed, M.A.; Nagi, A.M.; Aboulhoda, B.E.; Mohammed, E.T.; Abdel-Daim, M.M. Thymoquinone alleviates mitochondrial viability and apoptosis in diclofenac-induced acute kidney injury (AKI) via regulating Mfn2 and miR-34a mRNA expressions. Environ. Sci. Pollut. Res. 2021, 28, 10100–10113. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Wang, Q.; Xun, Y.; Li, C.; Wang, S. The Preliminary Exploration of What Role miRNAs Derived From Urinary Exosomes Play in Kidney Stone Formation. Urology 2022, 166, 104–110. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Shi, Y.; Hu, J.; Ding, J.; Guo, Z.; Yu, B. Integrated analysis of mRNA-seq and miRNA-seq reveals the potential roles of Egr1, Rxra and Max in kidney stone disease. Urolithiasis 2023, 51, 13. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.; Lai, Y.; Wu, W.; Chen, D.; Zhong, F.; Huang, J.; Zeng, T.; Duan, X.; Huang, Y.; Zhang, S.; et al. LncRNA-miRNA-mRNA expression variation profile in the urine of calcium oxalate stone patients. BMC Med. Genom. 2019, 12, 57. [Google Scholar] [CrossRef]
- Wang, B.; He, G.; Xu, G.; Wen, J.; Yu, X. MiRNA-34a inhibits cell adhesion by targeting CD44 in human renal epithelial cells: Implications for renal stone disease. Urolithiasis 2020, 48, 109–116. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhang, Y.; Han, S.; Chen, H.; Chen, C.; Ji, L.; Gao, B. Overexpression of miR-30c-5p reduces cellular cytotoxicity and inhibits the formation of kidney stones through ATG5. Int. J. Mol. Med. 2020, 45, 375–384. [Google Scholar] [CrossRef]
- Xie, Z.; Chen, J.; Chen, Z. MicroRNA-204 attenuates oxidative stress damage of renal tubular epithelial cells in calcium oxalate kidney-stone formation via MUC4-mediated ERK signaling pathway. Urolithiasis 2022, 50, 1–10. [Google Scholar] [CrossRef]
- Song, Z.; Zhang, Y.; Gong, B.; Xu, H.; Hao, Z.; Liang, C. Long noncoding RNA LINC00339 promotes renal tubular epithelial pyroptosis by regulating the miR-22-3p/NLRP3 axis in calcium oxalate–induced kidney stone. J. Cell. Biochem. 2019, 120, 10452–10462. [Google Scholar] [CrossRef]
- Xi, J.; Chen, Y.; Jing, J.; Qi, W.; Zhang, Y. LncRNA LINC01197 inhibited the formation of calcium oxalate-induced kidney stones by regulating miR-516b-5p/SIRT3/FOXO1 signaling pathway. Cell Tissue Res. 2023, 392, 553–563. [Google Scholar] [CrossRef]
- Rubel, D.; Boulanger, J.; Craciun, F.; Xu, E.Y.; Zhang, Y.; Phillips, L.; Callahan, M.; Weber, W.; Song, W.; Ngai, N.; et al. Anti-microRNA-21 Therapy on Top of ACE Inhibition Delays Renal Failure in Alport Syndrome Mouse Models. Cells 2022, 11, 594. [Google Scholar] [CrossRef] [PubMed]
- Lorenzen, J.M.; Menne, J.; Schmidt, B.M.W.; Schmidt, M.; Martino, F.; Dietrich, R.; Samiri, S.; Worthmann, H.; Heeren, M.; Weissenborn, K.; et al. Circulating microRNAs in Patients with Shiga-Toxin-Producing E. coli O104:H4 Induced Hemolytic Uremic Syndrome. PLoS ONE 2012, 7, e47215. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.K. RNA therapy: Rich history, various applications and unlimited future prospects. Exp. Mol. Med. 2022, 54, 455–465. [Google Scholar] [CrossRef] [PubMed]
- Bajan, S.; Hutvagner, G. RNA-Based Therapeutics: From Antisense Oligonucleotides to miRNAs. Cells 2020, 9, 137. [Google Scholar] [CrossRef] [PubMed]
- Torra, R.; Furlano, M. New therapeutic options for Alport syndrome. Nephrol. Dial. Transplant. 2019, 34, 1272–1279. [Google Scholar] [CrossRef]
- Lee, E.C.; Valencia, T.; Allerson, C.; Schairer, A.; Flaten, A.; Yheskel, M.; Kersjes, K.; Li, J.; Gatto, S.; Takhar, M.; et al. Discovery and preclinical evaluation of anti-miR-17 oligonucleotide RGLS4326 for the treatment of polycystic kidney disease. Nat. Commun. 2019, 10, 4148. [Google Scholar] [CrossRef]
- Ramalingam, H.; Yheskel, M.; Patel, V. Modulation of polycystic kidney disease by non-coding RNAs. Cell. Signal. 2020, 71, 109548. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bravo-Vázquez, L.A.; Paul, S.; Colín-Jurado, M.G.; Márquez-Gallardo, L.D.; Castañón-Cortés, L.G.; Banerjee, A.; Pathak, S.; Duttaroy, A.K. Exploring the Therapeutic Significance of microRNAs and lncRNAs in Kidney Diseases. Genes 2024, 15, 123. https://doi.org/10.3390/genes15010123
Bravo-Vázquez LA, Paul S, Colín-Jurado MG, Márquez-Gallardo LD, Castañón-Cortés LG, Banerjee A, Pathak S, Duttaroy AK. Exploring the Therapeutic Significance of microRNAs and lncRNAs in Kidney Diseases. Genes. 2024; 15(1):123. https://doi.org/10.3390/genes15010123
Chicago/Turabian StyleBravo-Vázquez, Luis Alberto, Sujay Paul, Miriam Guadalupe Colín-Jurado, Luis David Márquez-Gallardo, Luis Germán Castañón-Cortés, Antara Banerjee, Surajit Pathak, and Asim K. Duttaroy. 2024. "Exploring the Therapeutic Significance of microRNAs and lncRNAs in Kidney Diseases" Genes 15, no. 1: 123. https://doi.org/10.3390/genes15010123