
Characterization of Autochthonous Lactic Acid Bacteria Isolated from a Traditional Ethiopian Beverage, Tella
by
 , and
, and
Gashaw Assefa Yehuala
1,2,3,
Nurelegne Tefera Shibeshi
3, 
Su-Hyeon Kim
1,4 and
Mi-Kyung Park
1,4,* 1
School of Food Science and Biotechnology, Kyungpook National University, Daegu 41566, Republic of Korea
2
College of Biological and Chemical Engineering, Addis Ababa Science and Technology University, Addis Ababa P.O. Box 16417, Ethiopia
3
School of Chemical and Bio-Engineering, Addis Ababa Institute of Technology, Addis Ababa University, Addis Ababa P.O. Box 385, Ethiopia
4
Food and Bio-Industry Institute, Kyungpook National University, Daegu 41566, Republic of Korea
*
Author to whom correspondence should be addressed.
Foods 2024, 13(4), 575; https://doi.org/10.3390/foods13040575
Submission received: 11 January 2024
/
Revised: 29 January 2024
/
Accepted: 8 February 2024
/
Published: 14 February 2024
(This article belongs to the Section Food Microbiology)
Abstract
:This study aimed to isolate lactic acid bacteria (LAB) from a traditional Ethiopian fermented product, Tella, and evaluate their functional properties. Of forty-three isolates, seven LAB were screened and identified as Pediococcus pentosaceus, Latilactobacillus curvatus, Leuconostoc mesenteroides, and Lactiplantibacillus plantarum species. The isolates were tested for their alcohol tolerance, acid and bile resistance, auto-aggregation, co-aggregation, hydrophobicity, antibacterial activity, and antibiotic susceptibility. LAB isolates, specifically P. pentosaceus TAA01, L. mesenteroides TDB22, and L. plantarum TDM41, showed a higher degree of alcohol tolerance in 8% and 10% (w/v) ethanol concentrations. Additionally, these three isolates displayed survival rates >85% in both acidic pH and bile environments. Among the isolates, L. plantarum TDM41 demonstrated the highest auto-aggregation, co-aggregation, and hydrophobicity with (44.9 ± 1.7)%, (41.4 ± 0.2)%, and (52.1 ± 0.1)% values, respectively. The cell-free supernatant of the isolates exhibited antibacterial activity against foodborne pathogens of Escherichia coli, Salmonella Enteritidis, and Staphylococcus aureus. Each isolate exhibited various levels of resistance and susceptibility to seven antibiotics and resistance was observed against four of the antibiotics tested. After performing a principal component analysis, Pediococcus pentosaceus TAA01, L. mesenteroides TDB22, and L. plantarum TDM41 were selected as the most promising ethanol-tolerant probiotic isolates.
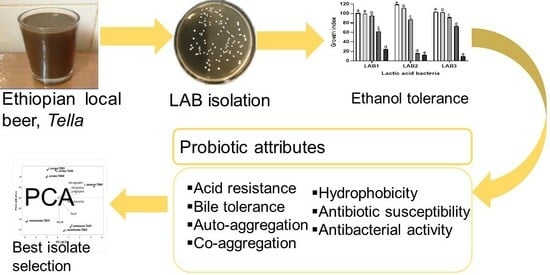
1. Introduction
Several traditional cereal-fermented beverages are produced in Ethiopia, such as Tella, Borde, Cheka, Korefe, Keribo, and Shamita [1,2]. Tella is the most popular fermented alcoholic beverage (2–8%) with an opaque appearance and light yellow to dark brown color [3]. It contains polyphenolic and flavonoid compounds, which offer various biological advantages including protection from free radicals, cancer, and aging [4]. Tella is made from various cereals such as barley, maize, wheat, millet, sorghum, and teff, depending on localities and their tradition [5,6].
Tella fermentation relies on microorganisms sourced from raw ingredients comprising yeasts (Saccharomyces), Lactobacillus, Bacillus, and other bacteria [3,7]. The traditional Tella preparation method usually comprises four phases, namely, the making of “tejet”, “tenses”, “difdif”, and finally Tella [3,8]. The starting cereals pass through different preparation methods such as soaking, germinating, roasting, grinding, and baking. The Tella preparation process is initiated by soaking barley and germinating it, followed by drying and grinding it into malt flour locally called “bikil” flour. In parallel, “gesho” (Rhamnus prinoides) leaves and stems are sun-dried and ponded. Then, “bikil” flour and gesho powder are blended and allowed to ferment for 96 h to create a fermenting mass known as “tejet” [8]. Following this, equal proportions of sorghum, millet, and teff flour are combined with water to produce a dough. The dough is subsequently baked to create unleavened bread, locally known as “kita”, which is then incorporated into the previously prepared “tejet”. The mixture is then sealed tightly and left to ferment for 96 h to turn into “tenses” [8]. While the “tenses” is fermenting, maize grain is soaked in water for about 72 h, and then, it is dried, roasted, and ground to make a dark maize flour called “Asharo”. “Asharo” is then added to the earlier produced “tenses” and fermented for a period of 96 h [8]. After this duration of fermentation, a thick mixture locally known as “difdif” is formed. Water is incorporated into “difdif” and allowed to ferment for a duration of 48 h [3]. Finally, solid residues are filtered out and the resulting liquid is served to consumers as Tella [1].
Probiotic bacterial strains contribute to the promotion of good nutrition by aiding in health maintenance through the prevention, control, and treatment of diseases [9,10,11,12]. In Tella and similar cereal fermentations, lactic acid bacteria (LAB) play a crucial role in shaping sensory attributes, ensuring safety, and enhancing functionality [7,9,12,13]. LAB are recognized as a key group of probiotic organisms, with numerous strains known for primarily residing in the gastrointestinal tract (GIT) and showing resilience against challenging conditions like low pH, bile salts, natural growth inhibitors, and interactions with other microbes [9]. Functional beverages can be developed through the incorporation of probiotics into food matrices. However, these probiotic microorganisms must exhibit resistance to various stresses encountered during the production process, such as high ethanol content and heat exposure [14].
Dairy products, especially yogurt, have been the common vehicles for probiotics due to the favorable conditions that milk and its derivatives create for these microorganisms [15]. However, there is a significant interest in developing new non-dairy food matrices for probiotic delivery. This trend is driven by a growing consumer preference for plant-based options to address issues like lactose intolerance, cholesterol concerns, and allergic reactions to milk proteins. Consumers highly appreciate food matrices, particularly cereal beverages endowed with functional properties, for their nutritional benefits. Consequently, probiotic-enriched alcoholic beverages have emerged as an innovative solution for delivering beneficial microorganisms. However, navigating the complex alcoholic beverage environment presents challenges in effectively cultivating specific probiotic strains [14].
In the context of probiotic beer development, Tella could be the best alternative and readily available source for alcohol-tolerant LAB strains with functional properties, particularly probiotic characteristics. The previous studies of Tella were primarily focused on outlining its traditional processing methodologies, physicochemical characteristics, and microbial profiles during fermentation [3,7,16]. Remarkably, there were no studies on the characterizations of LAB isolates such as alcohol tolerance, probiotic properties, and selection of LAB for use as starters in industrial applications. Therefore, the principal objective of this study was isolation and characterization of LAB from Tella samples for the selection of potential functional autochthonous starters with better alcohol-tolerant properties. The assessment and selection of the LAB candidates was carried out through a methodical approach encompassing the evaluation of their alcohol tolerance, survival rates and adhesion potential within the GIT, and antibacterial activities. In addition, the overall safety of confirmed LAB isolates was evaluated by an antibiotic susceptibility test to the selected commercial antibiotics.
2. Materials and Methods
2.1. Tella Samples
Fifteen Tella samples were purchased in August from traditional Tella breweries in Addis Ababa (AA), Debre Birhan (DB), and Debre Markos (DM), cities in Ethiopia. The capital, Addis Ababa, and the two nearby cities in the northern part of Ethiopia, DB and DM, are well known for the high rate of traditional Tella consumption and marketing [7]. Specific Tella vendors were selected based on the recommendation of customers to access good-quality Tella. Samples were carefully collected using sterile screw glass bottles and transported in an icebox. All samples were kept in a refrigerator until microbial analysis.
2.2. LAB Isolation
Lactic acid bacteria isolation was carried out in anaerobic conditions since probiotic strains are required to function in anaerobic conditions during cereal fermentation and in the GIT. Serially diluted Tella samples were spread onto De Man–Rogosa–Sharpe (MRS, Kisan Bio Company, Limited (Co., Ltd.), Seoul, Republic of Korea) agar plates supplemented with 50 mg/L cycloheximide (Sigma-Aldrich Co., Saint Louis, MO, USA). After incubation under anaerobic conditions at 30 °C for 48 h, cream to pale yellow colonies displaying circular shapes were randomly selected. These colonies were purified through at least five repetitive streaking cycles on the MRS agar medium. The purified isolates were maintained at −28 °C in MRS broth with 15% glycerol for further phenotypic analyses [17].
2.3. Phenotypic Identification of Purified Isolates
The Gram staining, catalase activity, and acidification ability of each LAB isolate were assessed for phenotypic identification. Gram staining was conducted by following the method described in [18]. For the catalase activity of purified single isolates, 3% hydrogen peroxide (Duksan Pure Chemicals Co., Ltd., Ansan, Republic of Korea) was mixed separately on a clean microscope slide with the pure isolates. Positive reactions were evidenced by immediate effervescence (bubble formation) due to the catalase hydrolyzing the hydrogen peroxide [19]. The acidification ability of each purified isolate was assessed by inoculation (6 log CFU/mL) in MRS broth and incubation at 30 °C for 24 h, then measuring its pH [20].
2.4. Genotypic Identification of Presumptive LAB Isolates
Presumptive LAB isolates were identified genotypically following the previous method [21]. Briefly, genomic DNA of seven presumptive LAB isolates was extracted and purified using a commercial Genomic DNA Isolation Kit (Solgent Co. Ltd., Daejeon, Republic of Korea). The 16S rRNA gene was amplified with the universal bacterial primer pair 27F and 1492R and the purified PCR products were sequenced by Solgent Co., Daejeon, Republic of Korea. The obtained sequences were analyzed using the basic local alignment search tool (BLAST) and aligned using the multiple alignment software ClustalW algorithm [22]. A phylogenetic tree was constructed using the maximum likelihood method with 100 bootstrap values via MEGA 11 software program [23].
2.5. Alcohol Tolerance of LAB Isolates
The alcohol tolerance of each LAB isolate was determined by assessing its ability to withstand challenges posed by ethanol at various concentrations during the fermentation process. The assessments were conducted following established methodologies as described in previous studies [20,24]. The overnight culture of each LAB isolate was individually inoculated at 6 log CFU/mL in MRS broth with varying concentrations of ethanol (2%, 4%, 6%, 8%, and 10%). The measurement of microbial growth was performed after 48 h by reading the absorbance at 600 nm. To serve as controls, samples of unmodified MRS broth inoculated with each LAB isolate were incubated at a temperature of 30°C. For each case, the data were expressed as growth index (GI), a relative measure comparing the growth in an experimental condition (As) relative to the control (Ac). GI was expressed using the following formula:
GI = As/Ac × 100,
2.6. Probiotic Characterization of LAB Isolates
2.6.1. Preparation of LAB and Bacterial Culture
The seven confirmed LAB isolates were cultured with MRS broth at 37 °C for 18 h under an anaerobic condition to evaluate their probiotic properties. Pathogenic strains of Escherichia coli ATCC 43895, Salmonella Enteritidis ATCC 13076, and Staphylococcus aureus ATCC 25923 were cultured in tryptic soy broth (TSB, Difco Laboratories Incorporated (Inc.), Sparks, MD, USA) at 37 °C for 18 h. After the specified incubation periods, each LAB isolate and pathogenic bacterial culture was harvested through centrifugation at 6000× g for 15 min. The harvested cells were then washed twice with phosphate-buffered saline (PBS, pH 7, Welgene Inc., Gyeongsan, Republic of Korea). Finally, the pellet was resuspended in PBS for further property tests. Cell-free supernatant (CFS) of LAB was prepared by filtering the supernatant obtained post-centrifugation of the overnight LAB culture through a microfilter (0.22 μm, GVS Co. Ltd., Panorama, Los Angeles, CA, USA).
2.6.2. Acid Resistance
The acid resistance of each LAB isolate was evaluated according to the method described by Vijayalakshmi et al. [25], with minor modifications. Each LAB culture was adjusted to pH 3.0 and incubated at 37 °C for 3 h. Sample aliquots were taken at time 0 and after 3 h of incubation, plated on MRS agar plates, and incubated anaerobically at 37 °C for 48 h to determine the survival rate after exposure to low pH. Cultures of each LAB isolate adjusted to pH 7.2 were used as controls. The survival rate was calculated as follows:
2.6.3. Bile Salt Tolerance
Bile salt tolerance of each LAB isolate was evaluated based on the methods described by Mallappa et al. [26], with minor modifications. Briefly, 1% LAB culture was inoculated in MRS broth supplemented with 0.3% (w/v) bile salt (Sigma-Aldrich Co., Ltd., USA) and incubated anaerobically at 37 °C for 4 h. Sample aliquots were taken at time 0 and after 4 h of incubation, plated on MRS agar plates, and incubated anaerobically at 37 °C for 48 h to determine the survival rate after exposure to bile salts. Cultures of each LAB isolate without bile salt were used as controls. The survival rate was calculated as follows:
2.6.4. Cell Auto-Aggregation
Auto-aggregation of each LAB isolate was determined following the method described by Mallappa et al. [26]. The absorbance (A0) of each LAB culture was adjusted with PBS to approximately 0.8 at 600 nm. After incubation at 37 °C for 4 h, the absorbance (At) of the upper fraction of incubated culture suspension was measured and the percentage of auto-aggregation was determined as follows:
Cell auto-aggregation (%) = (1 − At/A0) × 100,
2.6.5. Co-Aggregation
The co-aggregation property of each LAB isolate with the pathogenic strains was assessed following the method outlined by Mallappa et al. [26]. The initial absorbances of each LAB culture (Alac) and pathogenic bacterial culture (Apath) were adjusted to 0.8 ± 0.05 (8 log CFU/mL) and 0.3 ± 0.05 (8 log CFU/mL) at 600 nm, respectively. Then, equal volumes (1.5 mL) of each LAB culture and pathogenic bacterial culture were vortexed for 10 sec prior to incubation. After 4 h incubation at 37 °C, the absorbance of the mixture (Amix) was measured to calculate the co-aggregation rate:
Co-aggregation (%) = [((Alac + Apath)/2 − Amix)/(Alac + Apath)/2] × 100,
2.6.6. Cell Surface Hydrophobicity
Each isolate’s hydrophobicity was determined by its adhesion ability to organic solvents, based on the procedure described by Muñoz-Provencio et al. [27], with some modifications. Each LAB culture was adjusted to 8 log CFU/mL and the initial absorbance at 600 nm was measured (A0). A total of 3 mL of each LAB culture was separately mixed with 1 mL of xylene (Samchun Pure Chemical Co., Ltd., Pyeongtaek, Republic of Korea) and chloroform (Duksan Pure Chemicals Co., Ltd., Ansan-si, Republic of Korea). The mixtures were incubated at 37 °C for 10 min, vortexed thoroughly, and incubated again at 37 °C for 4 h. After incubation, the aqueous phase was removed, and the absorbance (At) was measured at 600 nm for the calculation of cell surface hydrophobicity using the following formula:
Cell surface hydrophobicity (%) = (1 − At/A0) × 100,
2.7. Antibiotic Susceptibility
The disc diffusion method [28,29] was employed to assess the antibiotic susceptibility of LAB isolates using eleven commercial antibiotics (Oxoid Ltd., Basingstoke, UK), including ampicillin, gentamicin, kanamycin, streptomycin, erythromycin, tetracycline, chloramphenicol, penicillin G, trimethoprim/sulfamethoxazole, ciprofloxacin, and azithromycin. A total of 100 μL of each LAB isolate suspension (8 log CFU/mL) was evenly spread on an MRS agar plate prior to the placement of each antibiotic disc. The antibiotic concentration on the discs is specified in Table S1. After anaerobic incubation at 37 °C for 24 h, each inhibition zone was measured for the determination of its susceptibility following the Clinical and Laboratory Standards [30] criteria.
2.8. Antibacterial Activity
Each LAB isolate’s antibacterial activity was tested against three pathogens, E. coli ATCC 43895, S. Enteritidis ATCC 13076, and S. aureus ATCC 25923, using the well diffusion method described by Sakoui et al. [31]. Briefly, 1% (v/v) of each pathogenic bacterial culture (8 log CFU/mL) was inoculated separately into molten Luria–Bertani agar (Difco, Sparks, MD, USA). After solidification, wells of 7 mm in diameter were prepared, and 100 μL of CFS was added to each well. Following diffusion for 4 h at 4 °C, the plates were incubated at 37 °C for 24 h. As a control, fresh MRS broth was utilized instead of CFS. The diameter (mm) of the clear zone around the wells was measured to compare antibacterial activity.
2.9. Statistical Analysis
All of the tests were performed in three independent experiments. The experimental results were expressed as mean ± standard deviation. The data were subjected to a one-way analysis of variance using GraphPad Prism 8.3.0 (GraphPad Software Inc., La Jolla, CA, USA) with p < 0.05 for statistical significance. Probiotic properties, including resistance to low pH and bile salts, auto-aggregation and co-aggregation, hydrophobicity, and antibacterial activity, were subjected to a principal component analysis (PCA) using Minitab 19.2 Statistical Software (Minitab Inc., State College, PA, USA) to select the best LAB isolates.
3. Results and Discussion
3.1. Isolation and Phenotypic Characterization of Presumptive LAB Isolates
Based on disparities in macroscopic characteristics, 43 bacterial colonies were isolated from 15 Tella samples (Table 1). Out of the 43 initial colonies, 33 were Gram-positive, 19 were catalase-negative, and 15 were both Gram-positive and catalase-negative. The cell morphology of these fifteen isolates consisted of four cocci, nine bacilli, one streptococcus, and one streptobacillus (Table 2). Based on the criteria defined by Amelia et al. [32], LAB encompass Gram-positive and catalase-negative bacteria. Consequently, 15 isolates with both characteristics were selected for further physiological tests, specifically evaluating their acidification ability, as detailed in Table 2. Seven isolates, including TAA01, TAA04, TDB19, TDB21, TDB22, TDM40, and TDM41, were able to reduce the pH of the MRS broth by >1.0. Therefore, these isolates were considered presumptive LAB with potential application for Tella fermentation and were subjected to genotypic identification.
3.2. Genotypic Identification
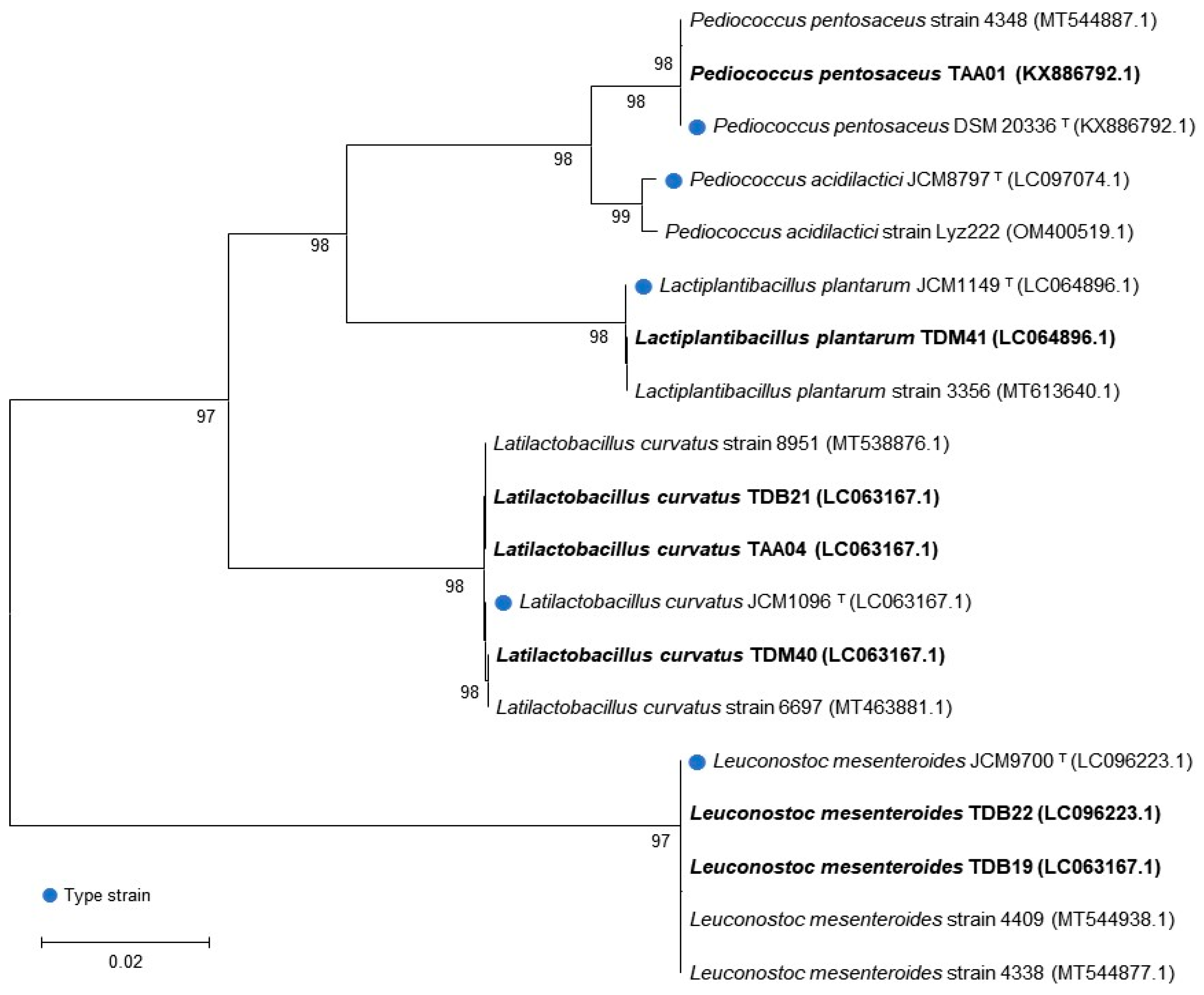
The genetic analysis of the presumptive LAB isolates identified the following genera: Pediococcus (one isolate), Latilactobacillus (three isolates), Leuconostoc (two isolates), and Lactiplantibacillus (one isolate) (Table 2). The BLAST hit displaying the maximum identity and query length was used as the closest relative of the isolates whose sequences were acquired. All isolates exhibited a similarity of >99% with the nucleotide sequence database of the National Center for Biotechnology Information (NCBI). The genetic sequence of each isolate was deposited in the NCBI GenBank database, and accession numbers were obtained. The identification and classification of the isolates were further confirmed by phylogenetic tree analysis (Figure 1), where each LAB isolate (written in bold) was classified with type strains (indicated by blue dots and “T” superscripts) and other similar strains in the database.
Pediococcus pentosaceus was isolated from AA Tella samples, two Leuconostoc mesenteroides were isolated from DB samples, and Lactiplantibacillus plantarum isolate was isolated from DM. On the other hand, Latilactobacillus curvatus isolates were consistently present in Tella samples collected from all sites. LAB’s predominant presence in Tella has been previously reported [3,7], although it is important to note that variations in seasonal conditions and processing practices may contribute to differences in the identified species of isolates. Following identification, all seven isolates underwent a series of tests specifically designed to evaluate their alcohol tolerance and probiotic characteristics. This comprehensive evaluation could be used to identify the most promising functional isolates for Tella fermentation and other functional food industry applications within the group.
3.3. Alcohol Tolerance Ability
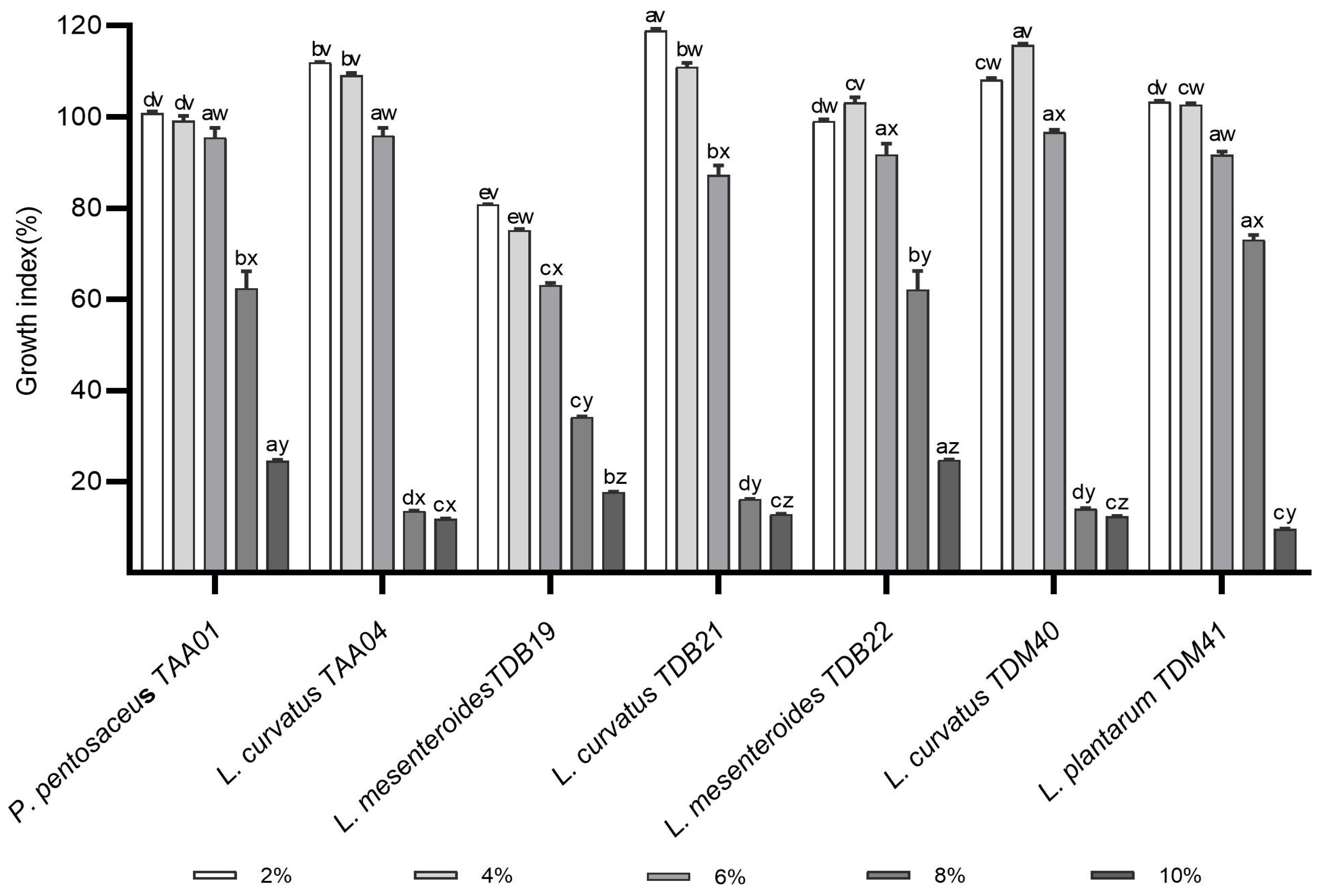
The alcohol content of Tella varies between 2% and 8% v/v [3]. High alcohol concentration affects both the growth and activity of LAB. When the concentration of ethanol surpasses a specific threshold, LAB are unable to autonomously sustain cellular stability, potentially leading to cell death. This outcome is attributed to intracellular metabolic imbalance resulting from the disruption of cell membrane functions [33]. Therefore, starter LAB strains to be employed in Tella fermentation as well as other food fermentations should have better resistance to alcohol stresses. In the present study, the alcohol tolerance of each LAB isolate was determined for ethanol concentrations ranging from 2 to 10% (Figure 2). As shown in Figure 2, there is a decreasing trend of GI of LAB isolates with the increase in ethanol concentration. The average GI was 104.2 ± 13.5%, 102.3 ± 13.2%, 88.9 ± 12.1%, 39.4 ± 26.0%, and 16.3 ± 6.2% for ethanol concentrations of 2%, 4%, 6%, 8%, and 10%, respectively. Notably, the GI values for certain isolates at 2% and 4% exceeded those of the control, indicating a potential promotion of their growth at lower alcohol concentrations. Such growth promotion at the low alcohol concentration may be due to the adaptation of the isolates to alcoholic conditions, where the LAB isolate may have developed different ethanol tolerance mechanisms. Among the isolates, P. pentosaceus TAA01 and L. curvatus TAA04 were not significantly inhibited by the increase in ethanol from 2% to 4%. At a 6% ethanol concentration, the GI of LAB isolates varied in the following order from highest to lowest: L. curvatus TDM40 (96.7 ± 0.5)%, L. curvatus TAA04 (95.9 ± 1.7)%, P. pentosaceus TAA01 (95.5 ± 2.1)%, L. mesenteroides TDB22 (91.8 ± 2.4)%, L. plantarum TDM41 (91.7 ± 0.7)%, L. curvatus TDB21 (87.3 ± 2.0)%, and L. mesenteroides TDB19 (63.1 ± 0.5)%. In this ranking, there is no significant GI difference (p > 0.05) among the top five isolates (Figure 2). As the ethanol concentration rose to 8% and 10%, the tolerance capabilities of LAB isolates exhibited more pronounced variations. P. pentosaceus TAA01, L. mesenteroides TDB22, and L. plantarum TDM41 were the most tolerant in 8% ethanol, and P. pentosaceus TAA01, L. mesenteroides TDB19, and L. mesenteroides TDB22 were the most tolerant at a 10% ethanol concentration. These findings closely agree with the study of [33,34]. Another study by Jin et al. [35] reported that L. paracasei isolates displayed a 5% alcohol tolerance. Compared to these previous studies, three isolates in the present study including P. pentosaceus TAA01, L. mesenteroides TDB22, and L. plantarum TDM41 exhibited a better GI (above 62%) under an 8% alcohol level. Remarkably, when subjected to the 8% ethanol concentration, L. plantarum TDM41 demonstrated a better GI of 73.08%, surpassing the earlier reported GI value of L. plantarum LTJ12 (59.01%) by Wang et al. [34]. Isolates of LAB with better alcohol tolerance have the potential to improve fermentation efficiency and stay viable during storage. This is particularly significant because such tolerant LAB strains could be incorporated into products delivered to consumers, ensuring viable probiotic counts and delivering health benefits. The findings of the current study, particularly those regarding the top three alcohol-tolerant isolates, indicate promising opportunities for developing functional cereal-based alcoholic beverages that cater to consumers seeking non-dairy options. However, further studies using Tella fermentation models are required to establish more robust conclusions.
3.4. Probiotic Properties
“Probiotics are living microorganisms that impart benefits to the host when administered in adequate quantities” [36]. They are essential in preparing functional beverages. Commonly, probiotics have been selected for their stress-resistant phenotypes, ensuring their survival through the GIT and subsequent establishment in the gut. However, the biological properties responsible for their health-promoting effects (e.g., antioxidant activities) and safety characteristics (e.g., antibiotic susceptibility) remain too important [37]. This study identified functional isolates among the screened LAB and evaluated their probiotic properties, including resistance to low pH and bile salts, auto-aggregation and co-aggregation, hydrophobicity, antibacterial activity, and antibiotic susceptibility.
3.4.1. Resistance to Low pH and Bile Salts
Acid and bile salt resistance are among the crucial criteria used to select isolates with probiotic functionality. Acid tolerance is essential to withstand the unfavorable conditions of the GIT and enables the strain to survive longer in high-acid beverages, such as yogurt and Tella [7,38]. To thrive and colonize in the human GIT successfully, probiotics ideally should possess traits that enable them to withstand the acidity of the stomach and endure exposure to bile in the upper portion of the intestine [26,31]. The majority of in vitro assays have been designed to identify strains capable of enduring the harsh conditions of the gastrointestinal tract, such as low pH values ranging from 2.0 to 3.0 and bile concentrations of 0.3% [26,39,40]. In this study, LAB isolates underwent evaluation in conditions reflective of the stomach environment, encompassing a pH of 3.0, a 0.3% bile salt concentration, and an exposure time of 3 h. However, it is worth checking the survival ability of the isolates at various pH levels in the range to ensure their versatile application.
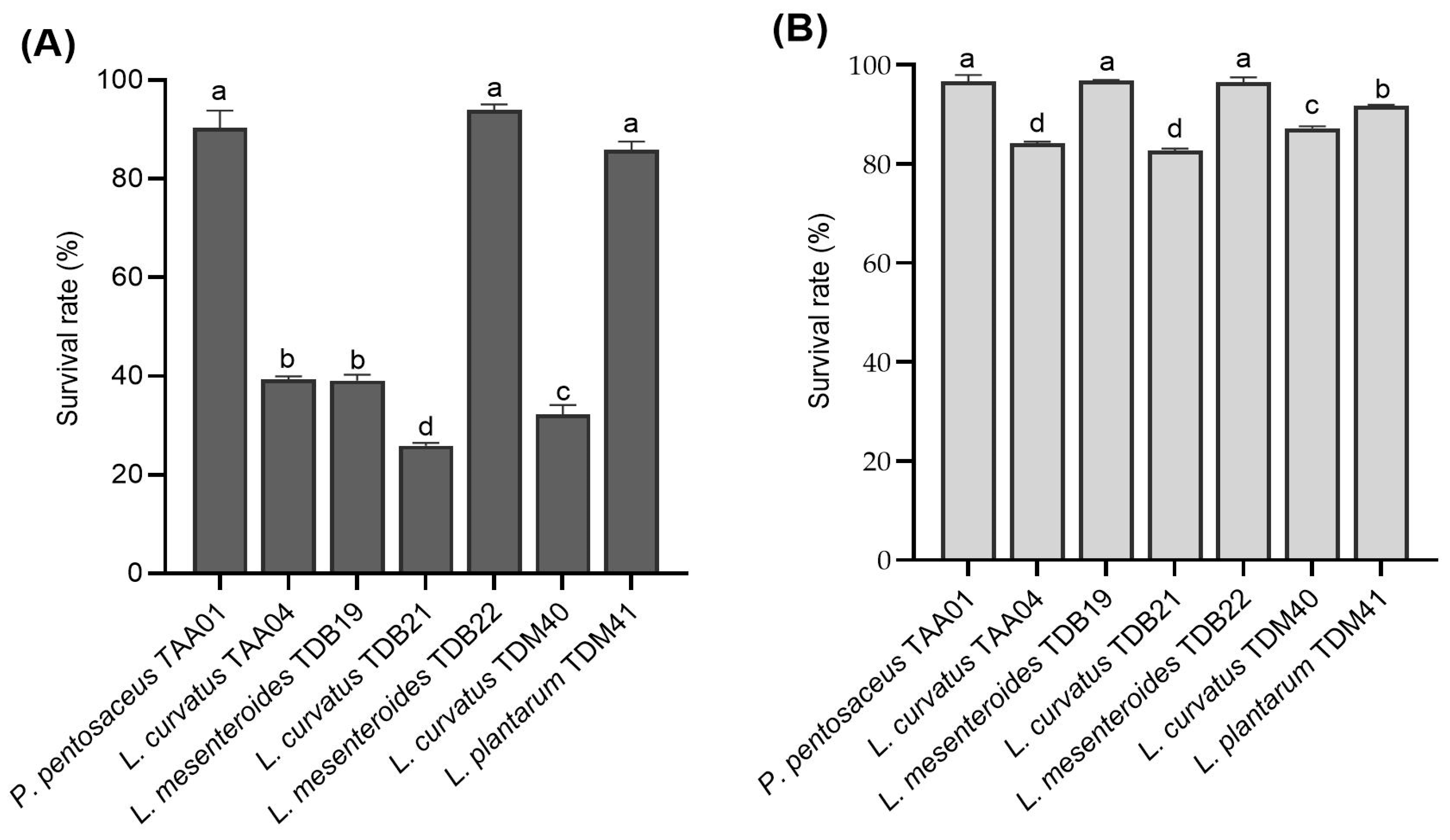
The LAB isolates displayed various resistance levels to low pH (3.0) and bile salt concentration (0.3%) ranging between (25.93 ± 0.6)% and (94.03 ± 1.1)% and between (82.71 ± 0.4)% and (96.92 ± 0.1)%, respectively (Figure 3A,B). The control samples in both acid and bile salt tolerance experiments did not show significant viability changes between the initial and final counts. Isolates P. pentosaceus TAA01, L. mesenteroides TDB22, and L. plantarum TDM41 displayed the highest resistance rates to low pH, with survival rates of (90.33 ± 3.52)%, (94.03 ± 1.06)%, and (85.93 ± 1.64)%, respectively. Regarding bile salt conditions, P. pentosaceus TAA01, L. mesenteroides TDB19, and L. mesenteroides TDB22 displayed the highest resistance, with survival rates of (96.75 ± 1.26)%, (96.92 ± 0.09)%, and (96.47 ± 1.07)%, respectively. The results are in close agreement with previous studies [20,26,41]. Notably, similar species, including L. mesenteroides (TDB19 and TDB22) and L. curvatus (TAA04, TDB21, and TDM40), displayed significantly different (p < 0.05) survival rates for both low pH and bile salt resistance, underscoring the crucial factor of strain specificity in the selection of probiotic strains [9,42]. TDB19 and TDB22, both belonging to L. mesenteroides, exhibited differences in survival rates, with TDB22 demonstrating superior survival in both acid and bile salt treatments. Similarly, within the L. curvatus species, TAA04, TDB21, and TDM40 displayed distinct acid resistance abilities, with TDM40 showing enhanced bile tolerance compared to TAA04 and TDB21. Isolates displaying strong acid resistance, like P. pentosaceus TAA01, L. mesenteroides TDB22, and L. plantarum TDM41, indicate their ability to remain viable in acidic environments, both during fermentation and exposure to gastrointestinal fluids. Using these isolates as starter cultures for fermenting cereal beverages, such as Tella, offers an exciting opportunity for effectively delivering probiotics. However, evaluating the survival capacity of these isolates in vivo remains prudent.
3.4.2. Auto-Aggregation and Co-Aggregation Properties
The probiotic ability to self-aggregate and co-aggregate with pathogenic bacteria is a good indicator of gut colonization [43]. The aggregation ability of probiotics is related to their cell adherence properties and specifies their capability to survive and persist in the GIT, beneficially affecting their host [9]. Among the tested LAB isolates, L. plantarum TDM41 displayed the highest auto-aggregation ability (44.9 ± 1.7)% and L. mesenteroides TDB19 the lowest (24.5 ± 1.9%) (Table 3). The auto-aggregation properties of the LAB isolates in the current study displayed higher values than those reported by Vijayalakshmi et al. [25] and lower values than those of Sakoui et al. [31]. Additionally, similar isolates of L. mesenteroides (TDB19 and TDB22) displayed significantly different auto-aggregation properties, indicating that auto-aggregation properties are also strain-specific [26].
A co-aggregation assay between the LAB isolates and pathogenic strains, including E. coli, S. Enteritidis, and S. aureus, was performed to evaluate interbacterial adherence. The LAB isolates displayed co-aggregation values between (19.5 ± 0.3)% (L. mesenteroides TDB19 with E. coli ATCC 43895) and (41.4 ± 0.2)% (L. plantarum TDM41 with S. aureus ATCC 25923) (Table 3). Wider ranges (7.0–70.0)% of LAB co-aggregation properties with other pathogens, including E. coli ATCC8539, L. monocytogenes ATCC19115, S. aureus ATCC 114, and S. Typhimurium LT2, have been previously reported [26,31,44], indicating that the co-aggregation properties in this study were moderate. Furthermore, our findings indicate that LAB co-aggregation with pathogens and their ability to adhere to the epithelial cell surface are strain-specific [26]. This specificity was demonstrated by the significantly different (p < 0.05) co-aggregation ability of L. curvatus isolates to E. coli ATCC 43895, L. mesenteroides isolates to S. Enteritidis ATCC 13076, and L. mesenteroides isolates to S. aureus ATCC 25923. Such variability can be explained by the presence of specific molecules on the surface of the LAB isolates acting as ligands for pathogen binding [45]. The co-aggregation of probiotics with pathogenic bacteria could also form a defensive barrier that may inhibit pathogens from colonizing the gut [46]. Good co-aggregation properties, along with auto-aggregation, can be used as a guide for selecting probiotic strains [47,48].
3.4.3. Hydrophobicity
Cell surface hydrophobicity is an essential probiotic property that influences bacterial adhesion and interaction with the host cells in the GIT [49,50]. Here, all isolates were evaluated for their ability to adhere to two organic solvents: chloroform and xylene. The isolates’ hydrophobicity ranged from (17.0 ± 0.4)% in xylene to (52.1 ± 0.1)% in chloroform (Table 3). The hydrophobicity of LAB isolates was generally higher in chloroform than in xylene. L. plantarum TDM41 displayed the highest hydrophobicity toward xylene (45.4 ± 0.1)% and chloroform (52.1 ± 0.1)%. On the other hand, L. mesenteroides TDB19 displayed the lowest values toward xylene (17.0% ± 0.4%) and chloroform (18.0 ± 0.1)%. Many studies have reported a hydrophobicity ranging from (2.0 ± 0.2)% to (88.0 ± 0.2)% [9,26,31,51]. Compared to such studies, our LAB isolates had a moderate hydrophobicity, indicating their ability to competitively attach to the epithelial cells and promote health [52].
3.5. Antibiotic Susceptibility
The food matrix can influence the spread of antimicrobial resistance by providing a suitable environment for the survival of resistant and multiresistant bacteria [53]. The proliferation of bacteria resistant to antimicrobial agents poses a significant threat to human health [54]. Therefore, evaluating antimicrobial resistance in indigenous LAB isolates is a crucial safety measure, given that fermented foods can disseminate these resistant microorganisms. In addition, horizontal gene transfer of antibiotic-resistant genes from probiotics to pathogenic bacteria is possible; however, LAB have been “generally recognized as safe” [51]. In this study, all seven LAB isolates were sensitive to erythromycin, tetracycline, chloramphenicol, and azithromycin but resistant to ampicillin, gentamycin, kanamycin, and streptomycin (Table 4). Gentamycin, kanamycin, and streptomycin are antibiotics that generally target Gram-negative bacteria. Due to their thick cell wall, Gram-positive LAB are resistant to these antibiotics [55]. In addition, resistance to aminoglycosides, including gentamicin, kanamycin, and streptomycin, is intrinsic to the Lactobacillus genus and cannot present safety problems [56,57]. Most LAB isolates displayed intermediate susceptibility to penicillin, while the three L. curvatus isolates were susceptible. All isolates displayed resistance to trimethoprim/sulfamethoxazole, except L. plantarum TDM41. Furthermore, four isolates displayed resistance to ciprofloxacin, while the other three exhibited intermediate susceptibility. The findings align with prior research, indicating that LAB isolates exhibit sensitivity to antibiotics that impede protein synthesis, such as chloramphenicol, erythromycin, and tetracycline. Conversely, resistance was observed against aminoglycosides, including gentamycin, kanamycin, and streptomycin [9,58]. The different susceptibility levels of the LAB isolates toward penicillin and ciprofloxacin indicate strain specificity [59]. Probiotic isolates should be susceptible to at least two clinically relevant antibiotics [60]; therefore, the isolates in this study meet the safety requirements in terms of phenotypic resistance evaluation. Additionally, isolates can harbor truncated antimicrobial resistance genes without expressing the phenotypic resistance pattern [61] or they can show phenotypic resistance to multiple antimicrobial compounds without presenting commonly evaluated resistance genes. Hence, it is imperative to examine the key resistance genes even when an isolate does not exhibit phenotypic resistance. If isolates carry any resistance gene, it becomes crucial to assess both the expression of that gene and the existence of mobile genetic elements capable of transferring these resistance determinants to other bacteria [62].
3.6. Antibacterial Activity
The antibacterial activity of LAB is essential for their selection as candidates for starter culture development and probiotic application [63,64,65]. Selecting a starter culture candidate with better antagonistic properties toward harmful pathogens in the fermentation medium and GIT could help produce improved quality products: the so-called functional beverages. The antibacterial activity levels of the CFS against test pathogens, including E. coli ATCC 43895, S. Enteritidis ATCC 13076, and S. aureus ATCC 25923, are presented in Table 5. The CFS from all LAB isolates displayed antibacterial activity against all of the test pathogens, except the non-detectable effect of L. curvatus TAA04 and TDM40 on S. aureus ATCC 25923 and L. mesenteroides TDB19 on E. coli ATCC 43895 and S. Enteritidis ATCC 13076. E. coli ATCC 43895 and S. Enteritidis ATCC 13076 were inhibited by six LAB isolates, whereas S. aureus ATCC 25923 was inhibited by five. The highest inhibition zone (17.0 ± 1 mm) was observed on S. Enteritidis ATCC 13076 by L. mesenteroides TDB22, and the lowest (9.0 ± 1.0 mm) was on S. Enteritidis ATCC 13076 by L. curvatus TDM40. Overall, P. pentosaceus TAA01, L. mesenteroides TDB22, and L. plantarum TDM41 exhibited better antibacterial activity than other isolates. Similar findings have been reported in several studies [31,43]. The production of organic acids, hydrogen peroxide, and bacteriocins by LAB is responsible for the inhibition effect on pathogens [63,65,66]. In general, the results demonstrated that the LAB isolates had a more effective antibacterial activity against E. coli ATCC 43895 and S. Enteritidis ATCC 13076 than against S. aureus ATCC 25923.
3.7. Functional Starter Selection
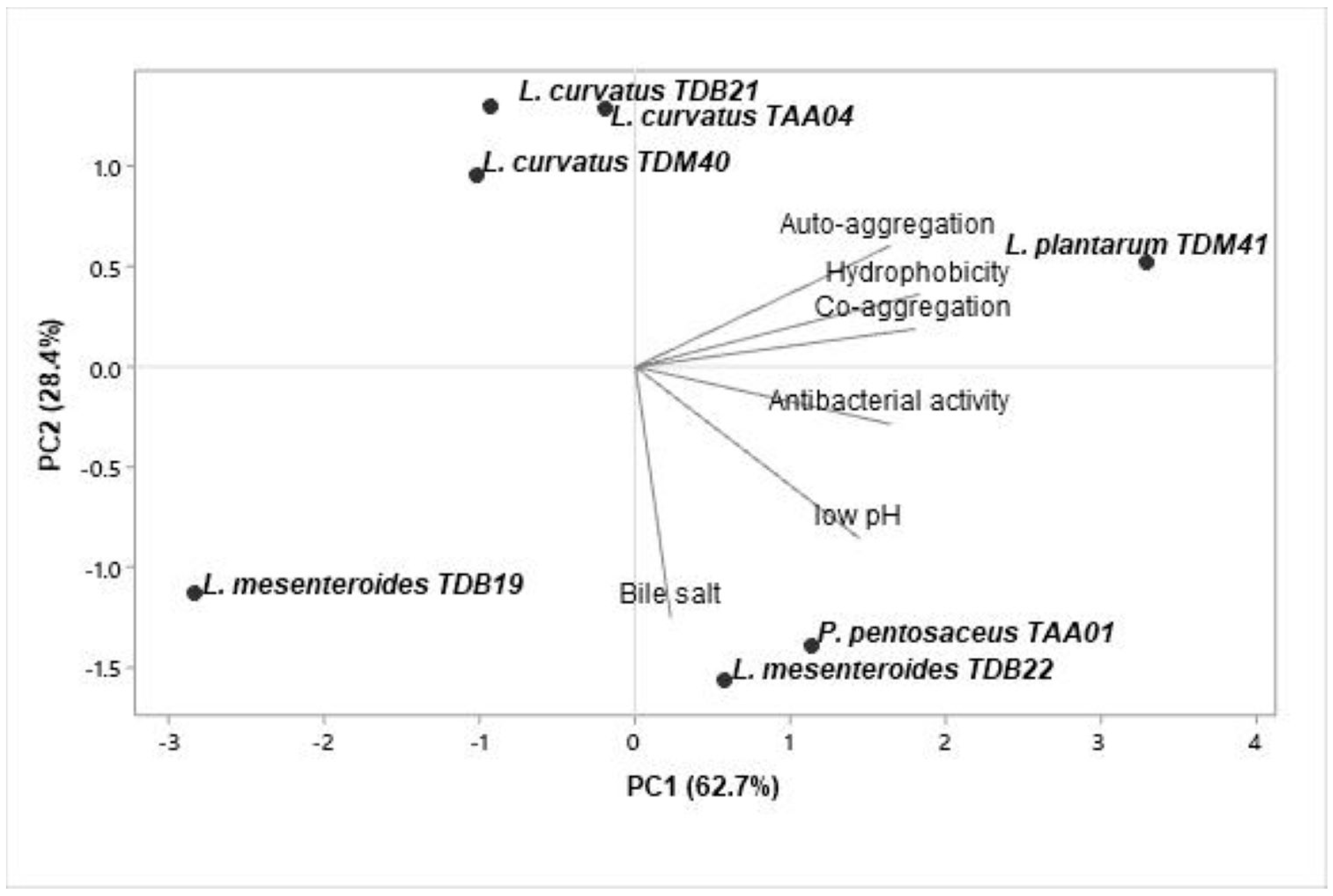
The obtained data were subjected to a multivariate PCA using Minitab 19.2 software to evaluate the similarity and variability between the functional properties of the isolates and select potential starter candidates. The analyzed data include acid and bile salt resistance, auto-aggregation and co-aggregation, cell surface hydrophobicity, and antimicrobial activity. The first two PCs accounted for 91.1% of the total variance (Figure 4). PC1 and PC2 contributed 62.7% and 28.4%, respectively. The biplot based on PC1 and PC2 differentiated the isolates into four quadrants. Isolates in quadrants II and IV displayed a better correlation to PC1, indicating that P. pentosaceus TAA01, L. mesenteroides TDB22, and L. plantarum TDM41 are good candidates for potential industrial applications.
4. Conclusions
It is undeniable that probiotic microbes are incredibly important for our health and well-being. The LAB isolated from Tella, especially P. pentosaceus TAA01, L. mesenteroides TDB22, and L. plantarum TDM41, showed remarkable resilience, tolerating ethanol concentrations up to 8%. These isolates also demonstrated superior survival under conditions simulating the GIT. Moreover, all of the isolated strains displayed moderate to high adhesion properties, with L. plantarum TDM41 standing out as the top performer in terms of auto-aggregation, co-aggregation, and hydrophobicity. While susceptibility to several antibiotics was observed in all strains, they also exhibited antimicrobial potential. The present study’s findings highlight the effectiveness of PCA in selecting the most promising probiotic isolates, with P. pentosaceus TAA01, L. mesenteroides TDB22, and L. plantarum TDM41 showing promising functional properties among the LAB isolates. However, there is a need for further exploration, particularly in establishing Tella model systems to assess technological properties, the impact on organoleptic qualities, and in vivo investigation of the probiotic properties. This additional evaluation will pave the way for implementing an indigenous starter culture in Tella fermentation, contributing to the development of functional beverages.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/foods13040575/s1. Table S1: Antibiotic sensitivity interpretative standard table.
Author Contributions
Conceptualization, G.A.Y., N.T.S., S.-H.K. and M.-K.P.; methodology, G.A.Y.; software, G.A.Y.; validation, G.A.Y.; formal analysis, G.A.Y.; investigation, G.A.Y.; resources, M.-K.P.; data curation, G.A.Y.; writing—original draft preparation, G.A.Y.; writing—review and editing, G.A.Y., N.T.S., S.-H.K. and M.-K.P.; visualization, G.A.Y. and S.-H.K.; supervision, N.T.S. and M.-K.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The original contributions presented in the study are included in the article/supplementary material, further inquiries can be directed to the corresponding author/s.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Fentie, E.G.; Emire, S.A.; Demsash, H.D.; Dadi, D.W.; Shin, J.-H. Cereal-and fruit-based Ethiopian traditional fermented alcoholic beverages. Foods 2020, 9, 1781. [Google Scholar] [CrossRef] [PubMed]
- Yohannes, T.; Melak, F.; Siraj, K. Preparation and physicochemical analysis of some Ethiopian traditional alcoholic beverages. Afr. J. Food Sci. 2013, 7, 399–403. [Google Scholar] [CrossRef]
- Samuel, S.; Berhanu, A. The microbiology of Tella fermentation. Sinet 1991, 14, 81–92. [Google Scholar]
- Shewakena, S.; Chandravanshi, B.; Debebe, A. Levels of total polyphenol, flavonoid, tannin and antioxidant activity of selected Ethiopian fermented traditional beverages. Int. Food Res. J. 2017, 24, 2033–2040. [Google Scholar] [CrossRef]
- Andualem, B.; Shiferaw, M.; Berhane, N. Isolation and characterization of Saccaromyces cervisiae yeasts isolates from Tella for beer production. Annu. Res. Rev. Biol. 2017, 15, 1–12. [Google Scholar] [CrossRef]
- Ashenafi, M. A review on the microbiology of indigenous fermented foods and beverages of Ethiopia. Ethiop. J. Biol. Sci. 2006, 5, 189–245. [Google Scholar] [CrossRef]
- Tekle, B.; Anuradha Jabasingh, S.; Fantaw, D.; Gebreslassie, T.; Ram Mohan Rao, S.; Baraki, H.; Gebregziabher, K. An insight into the Ethiopian traditional alcoholic beverage: Tella processing, fermentation kinetics, microbial profiling and nutrient analysis. LWT Food Sci. Technol. 2019, 107, 9–15. [Google Scholar] [CrossRef]
- Berhanu, A. Microbial profile of Tella and the role of “gesho” (Rhamnus prinoides) as bittering and antimicrobial agent in traditional Tella (beer) production. Int. Food Res. 2014, 21, 357–365. [Google Scholar]
- Gupta, S.; Mohanty, U.; Majumdar, R.K. Isolation and characterization of lactic acid bacteria from traditional fermented fish product Shidal of India with reference to their probiotic potential. LWT Food Sci. Technol. 2021, 146, 111641. [Google Scholar] [CrossRef]
- Holzapfel, W.H. Appropriate starter culture technologies for small-scale fermentation in developing countries. Int. J. Food Microbiol. 2002, 75, 197–212. [Google Scholar] [CrossRef]
- Hotessa, N.; Robe, J. Ethiopian indigenous traditional fermented beverage: The role of the microorganisms toward nutritional and safety value of fermented beverage. Int. J. Microbiol. 2020, 2020, 8891259. [Google Scholar] [CrossRef] [PubMed]
- Leroy, F.; De Vuyst, L. Lactic acid bacteria as functional starter cultures for the food fermentation industry. Trends Food Sci. Technol. 2004, 15, 67–78. [Google Scholar] [CrossRef]
- Enujiugha, V.N.; Badejo, A.A. Probiotic potentials of cereal-based beverages. Crit. Rev. Food Sci. Nutr. 2017, 57, 790–804. [Google Scholar] [CrossRef] [PubMed]
- Hinojosa-Avila, C.R.; García-Gamboa, R.; Chedraui-Urrea, J.J.; García-Cayuela, T. Exploring the potential of probiotic-enriched beer: Microorganisms, fermentation strategies, sensory attributes, and health implications. Food Res. Int. 2023, 175, 113717. [Google Scholar] [CrossRef] [PubMed]
- Min, M.; Bunt, C.R.; Mason, S.L.; Hussain, M.A. Non-dairy probiotic food products: An emerging group of functional foods. Crit. Rev. Food Sci. Nutr. 2019, 59, 2626–2641. [Google Scholar] [CrossRef] [PubMed]
- Alemu, F.; Amhaselassie, T.; Kelbessa, U.; Elias, S. Methanol, fusel oil, and ethanol contents of some Ethiopian traditional alcoholic beverages. Sinet 1991, 14, 19–27. [Google Scholar]
- Lacerda, I.C.; Miranda, R.L.; Borelli, B.M.; Nunes, Á.C.; Nardi, R.M.; Lachance, M.-A.; Rosa, C.A. Lactic acid bacteria and yeasts associated with spontaneous fermentations during the production of sour cassava starch in Brazil. Int. J. Food Microbiol. 2005, 105, 213–219. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.C.; Hussey, M.A. Gram Stain Protocols, 3rd ed.; American Society for Microbiology: Washington, DC, USA, 2005; pp. 113–144. [Google Scholar]
- Reiner, K. Catalase Test Protocol; American Society for Microbiology: Washington, DC, USA, 2010; pp. 1–7. [Google Scholar]
- Speranza, B.; Racioppo, A.; Beneduce, L.; Bevilacqua, A.; Sinigaglia, M.; Corbo, M.R. Autochthonous lactic acid bacteria with probiotic aptitudes as starter cultures for fish-based products. Food Microbiol. 2017, 65, 244–253. [Google Scholar] [CrossRef]
- Singh, H.; Du, J.; Singh, P.; Yi, T.H. Extracellular synthesis of silver nanoparticles by Pseudomonas sp. THG-LS1. 4 and their antimicrobial application. J. Pharm. Anal. 2018, 8, 258–264. [Google Scholar] [CrossRef]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Bevilacqua, A.; Altieri, C.; Corbo, M.R.; Sinigaglia, M.; Ouoba, L.I.I. Characterization of lactic acid bacteria isolated from Italian Bella di Cerignola table olives: Selection of potential multifunctional starter cultures. J. Food Sci. 2010, 75, M536–M544. [Google Scholar] [CrossRef] [PubMed]
- Vijayalakshmi, S.; Adeyemi, D.E.; Choi, I.Y.; Sultan, G.; Madar, I.H.; Park, M.-K. Comprehensive in silico analysis of lactic acid bacteria for the selection of desirable probiotics. LWT Food Sci. Technol. 2020, 130, 109617. [Google Scholar] [CrossRef]
- Mallappa, R.H.; Singh, D.K.; Rokana, N.; Pradhan, D.; Batish, V.K.; Grover, S. Screening and selection of probiotic Lactobacillus strains of Indian gut origin based on assessment of desired probiotic attributes combined with principal component and heatmap analysis. LWT Food Sci. Technol. 2019, 105, 272–281. [Google Scholar] [CrossRef]
- Muñoz-Provencio, D.; Llopis, M.; Antolín, M.; De Torres, I.; Guarner, F.; Pérez-Martínez, G.; Monedero, V. Adhesion properties of Lactobacillus casei strains to resected intestinal fragments and components of the extracellular matrix. Arch. Microbiol. 2009, 191, 153–161. [Google Scholar] [CrossRef] [PubMed]
- Haghshenas, B.; Nami, Y.; Abdullah, N.; Radiah, D.; Rosli, R.; Khosroushahi, A.Y. Anti-proliferative effects of Enterococcus strains isolated from fermented dairy products on different cancer cell lines. J. Funct. Foods 2014, 11, 363–374. [Google Scholar] [CrossRef]
- Won, S.-M.; Chen, S.; Park, K.W.; Yoon, J.-H. Isolation of lactic acid bacteria from kimchi and screening of Lactobacillus sakei ADM14 with anti-adipogenic effect and potential probiotic properties. LWT Food Sci. Technol. 2020, 126, 109296. [Google Scholar] [CrossRef]
- CLSI. Performance Standards for Antimicrobial Disk Susceptibility Tests, 30th ed.; Clinical and Laboratory Standards Institute: Wayne, NJ, USA, 2020; pp. 1–16. [Google Scholar]
- Sakoui, S.; Derdak, R.; Addoum, B.; Pop, O.L.; Vodnar, D.C.; Suharoschi, R.; Soukri, A.; El Khalfi, B. The first study of probiotic properties and biological activities of lactic acid bacteria isolated from Bat guano from Er-rachidia, Morocco. LWT Food Sci. Technol. 2022, 159, 113224. [Google Scholar] [CrossRef]
- Amelia, R.; Philip, K.; Pratama, Y.E.; Purwati, E. Characterization and probiotic potential of lactic acid bacteria isolated from dadiah sampled in West Sumatra. Food Sci. Technol. 2020, 41, 746–752. [Google Scholar] [CrossRef]
- Chen, X.; Wang, T.; Jin, M.; Tan, Y.; Liu, L.; Liu, L.; Li, C.; Yang, Y.; Du, P. Metabolomics analysis of growth inhibition of Lactobacillus plantarum under ethanol stress. Int. J. Food Sci. Technol. 2020, 55, 3441–3454. [Google Scholar] [CrossRef]
- Wang, J.; Lu, C.; Xu, Q.; Li, Z.; Song, Y.; Zhou, S.; Zhang, T.; Luo, X. Bacterial diversity and lactic acid bacteria with high alcohol tolerance in the fermented grains of soy sauce aroma type baijiu in North China. Foods 2022, 11, 1794. [Google Scholar] [CrossRef] [PubMed]
- Jin, G.; Jiranek, V.; Hayes, A.M.; Grbin, P.R. Isolation and characterization of high-ethanol-tolerance lactic acid bacteria from Australian Wine. Foods 2022, 11, 1231. [Google Scholar] [CrossRef] [PubMed]
- Evaluation of Health and Nutritional Properties of Powder Milk and Live Lactic Acid Bacteria; FAO/WHO: Cordoba, Argentina, 2001; pp. 1–34.
- de Melo Pereira, G.V.; de Oliveira Coelho, B.; Magalhães Júnior, A.I.; Thomaz-Soccol, V.; Soccol, C.R. How to select a probiotic? A review and update of methods and criteria. Biotechnol. Adv. 2018, 36, 2060–2076. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.-Q.; Meng, X.-C.; Zhang, B.-R.; Wang, Y.; Shang, Y.-L. Influence of cell surface properties on adhesion ability of bifidobacteria. World J. Microbiol. Biotechnol. 2010, 26, 1999–2007. [Google Scholar] [CrossRef]
- Chandran, A.; Duary, R.K.; Grover, S.; Batish, V.K. Relative expression of bacterial and host specific genes associated with probiotic survival and viability in the mice gut fed with Lactobacillus plantarum Lp91. Microbiol. Res. 2013, 168, 555–562. [Google Scholar] [CrossRef] [PubMed]
- Kaushik, J.K.; Kumar, A.; Duary, R.K.; Mohanty, A.K.; Grover, S.; Batish, V.K. Functional and probiotic attributes of an indigenous isolate of Lactobacillus plantarum. PLoS ONE 2009, 4, e8099. [Google Scholar] [CrossRef] [PubMed]
- Maldonado, N.C.; de Ruiz, C.S.; Otero, M.C.; Sesma, F.; Nader-Macías, M.E. Lactic acid bacteria isolated from young calves—Characterization and potential as probiotics. Res. Vet. Sci. 2012, 92, 342–349. [Google Scholar] [CrossRef] [PubMed]
- Gu, R.-X.; Yang, Z.-Q.; Li, Z.-H.; Chen, S.-L.; Luo, Z.-L. Probiotic properties of lactic acid bacteria isolated from stool samples of longevous people in regions of Hotan, Xinjiang and Bama, Guangxi, China. Anaerobe 2008, 14, 313–317. [Google Scholar] [CrossRef]
- Cesena, C.; Morelli, L.; Alander, M.; Siljander, T.; Tuomola, E.; Salminen, S.; Mattila-Sandholm, T.; Vilpponen-Salmela, T.; von Wright, A. Lactobacillus crispatus and its nonaggregating mutant in human colonization trials. J. Dairy Sci. 2001, 84, 1001–1010. [Google Scholar] [CrossRef]
- Rokana, N.; Mallappa, R.H.; Batish, V.K.; Grover, S. Interaction between putative probiotic Lactobacillus strains of Indian gut origin and Salmonella: Impact on intestinal barrier function. LWT Food Sci. Technol. 2017, 84, 851–860. [Google Scholar] [CrossRef]
- Campana, R.; van Hemert, S.; Baffone, W. Strain-specific probiotic properties of lactic acid bacteria and their interference with human intestinal pathogens invasion. Gut Pathog. 2017, 9, 12. [Google Scholar] [CrossRef]
- Vidhyasagar, V.; Jeevaratnam, K. Evaluation of Pediococcus pentosaceus strains isolated from Idly batter for probiotic properties in vitro. J. Funct. Food. 2013, 5, 235–243. [Google Scholar] [CrossRef]
- Ferreira, C.L.; Grześkowiak, L.; Collado, M.C.; Salminen, S. In vitro evaluation of Lactobacillus gasseri strains of infant origin on adhesion and aggregation of specific pathogens. J. Food Prot. 2011, 74, 1482–1487. [Google Scholar] [CrossRef] [PubMed]
- Vlková, E.; Rada, V.; Šmehilová, M.; Killer, J. Auto-aggregation and co-aggregation ability in Bifidobacteria and Clostridia. Folia Microbiol. 2008, 53, 263–269. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Alcántara, A.M.; Wacher, C.; Llamas, M.G.; López, P.; Pérez-Chabela, M.L. Probiotic properties and stress response of thermotolerant lactic acid bacteria isolated from cooked meat products. LWT Food Sci. Technol. 2018, 91, 249–257. [Google Scholar] [CrossRef]
- Xu, H.; Jeong, H.; Lee, H.; Ahn, J. Assessment of cell surface properties and adhesion potential of selected probiotic strains. Lett. Appl. Microbiol. 2009, 49, 434–442. [Google Scholar] [CrossRef] [PubMed]
- Oh, A.; Daliri, E.B.-M.; Oh, D.H. Screening for potential probiotic bacteria from Korean fermented soybean paste: In vitro and Caenorhabditis elegans model testing. LWT Food Sci. Technol. 2018, 88, 132–138. [Google Scholar] [CrossRef]
- Schillinger, U.; Guigas, C.; Heinrich Holzapfel, W. In vitro adherence and other properties of lactobacilli used in probiotic yoghurt-like products. Int. Dairy J. 2005, 15, 1289–1297. [Google Scholar] [CrossRef]
- Bhargava, K.; Zhang, Y. Multidrug-resistant coagulase-negative Staphylococci in food animals. J. Appl. Microbiol. 2012, 113, 1027–1036. [Google Scholar] [CrossRef]
- Chajęcka-Wierzchowska, W.; Zadernowska, A.; Nalepa, B.; Sierpińska, M.; Łaniewska-Trokenheim, Ł. Coagulase-negative staphylococci (CoNS) isolated from ready-to-eat food of animal origin–phenotypic and genotypic antibiotic resistance. Food Microbiol. 2015, 46, 222–226. [Google Scholar] [CrossRef]
- Han, J.; Chen, D.; Li, S.; Li, X.; Zhou, W.-W.; Zhang, B.; Jia, Y. Antibiotic susceptibility of potentially probiotic Lactobacillus strains. Ital. J. Food Sci. 2015, 27, 282–289. [Google Scholar]
- Botta, C.; Langerholc, T.; Cencič, A.; Cocolin, L. In vitro selection and characterization of new probiotic candidates from table olive microbiota. PLoS ONE 2014, 9, e94457. [Google Scholar] [CrossRef] [PubMed]
- Elkins, C.A.; Mullis, L.B. Bile-mediated aminoglycoside sensitivity in Lactobacillus species likely results from increased membrane permeability attributable to cholic acid. Appl. Environ. Microbiol. 2004, 70, 7200–7209. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Pillidge, C.; Gopal, P.; Gill, H. Antibiotic susceptibility profiles of new probiotic Lactobacillus and Bifidobacterium strains. Int. J. Food Microbiol. 2005, 98, 211–217. [Google Scholar] [CrossRef] [PubMed]
- Danielsen, M.; Simpson, P.; O’connor, E.; Ross, R.; Stanton, C. Susceptibility of Pediococcus spp. to antimicrobial agents. J. Appl. Microbiol. 2007, 102, 384–389. [Google Scholar] [CrossRef] [PubMed]
- Borriello, S.; Hammes, W.; Holzapfel, W.; Marteau, P.; Schrezenmeir, J.; Vaara, M.; Valtonen, V. Safety of probiotics that contain Lactobacilli or Bifidobacteria. Clin. Infect. Dis. 2003, 36, 775–780. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, W.; Wang, J.; Wu, C.; Shen, Z.; Fu, X.; Yan, Y.; Zhang, Q.; Schwarz, S.; Shen, J. Distribution of the multidrug resistance gene cfr in Staphylococcus species isolates from swine farms in China. Antimicrob. Agents Chemother. 2012, 56, 1485–1490. [Google Scholar] [CrossRef] [PubMed]
- dos Santos Cruxen, C.E.; Funck, G.D.; Haubert, L.; da Silva Dannenberg, G.; de Lima Marques, J.; Chaves, F.C.; da Silva, W.P.; Fiorentini, Â.M. Selection of native bacterial starter culture in the production of fermented meat sausages: Application potential, safety aspects, and emerging technologies. Food Res. Int. 2019, 122, 371–382. [Google Scholar] [CrossRef]
- Fei, Y.; Li, L.; Zheng, Y.; Liu, D.; Zhou, Q.; Fu, L. Characterization of Lactobacillus amylolyticus L6 as potential probiotics based on genome sequence and corresponding phenotypes. LWT Food Sci. Technol. 2018, 90, 460–468. [Google Scholar] [CrossRef]
- Holzapfel, W. Use of starter cultures in fermentation on a household scale. Food Control 1997, 8, 241–258. [Google Scholar] [CrossRef]
- Servin, A.L. Antagonistic activities of lactobacilli and bifidobacteria against microbial pathogens. FEMS Microbiol. Rev. 2004, 28, 405–440. [Google Scholar] [CrossRef]
- Shah, N.P. Functional cultures, and health benefits. Int. Dairy J. 2007, 17, 1262–1277. [Google Scholar] [CrossRef]
Figure 1.
Phylogenic analysis of autochthonous lactic acid bacteria isolates based on 16S rRNA gene sequencing. The number next to the branches indicates the percentage of replicate trees in which the associated taxa clustered together in the bootstrap test with 100 replicates.
Figure 1.
Phylogenic analysis of autochthonous lactic acid bacteria isolates based on 16S rRNA gene sequencing. The number next to the branches indicates the percentage of replicate trees in which the associated taxa clustered together in the bootstrap test with 100 replicates.

Figure 2.
Alcohol tolerance abilities of autochthonous lactic acid bacteria isolates exposed to 2%, 4%, 6%, 8%, and 10% ethanol (v/v) in MRS broth. Values are expressed as mean ± SD (n = 3). Different superscript letters (a–e) represent significant differences (p < 0.05) between different isolates under the same treatment and (v–z) represent significant differences within the same isolates across different treatments.
Figure 2.
Alcohol tolerance abilities of autochthonous lactic acid bacteria isolates exposed to 2%, 4%, 6%, 8%, and 10% ethanol (v/v) in MRS broth. Values are expressed as mean ± SD (n = 3). Different superscript letters (a–e) represent significant differences (p < 0.05) between different isolates under the same treatment and (v–z) represent significant differences within the same isolates across different treatments.

Figure 3.
Survival rates of autochthonous lactic acid bacteria isolates following exposure to (A) low pH (3.0) for 3 h and (B) bile salt (0.3%) for 4 h (count, log CFU/mL). Values are expressed as mean ± SD (n = 3). Different superscript letters (a–d) indicate a significant difference (p < 0.05).
Figure 3.
Survival rates of autochthonous lactic acid bacteria isolates following exposure to (A) low pH (3.0) for 3 h and (B) bile salt (0.3%) for 4 h (count, log CFU/mL). Values are expressed as mean ± SD (n = 3). Different superscript letters (a–d) indicate a significant difference (p < 0.05).

Figure 4.
Principal component analysis of probiotic properties (low pH resistance, bile salt resistance, auto-aggregation, co-aggregation, cell surface hydrophobicity, and antibacterial activity) of seven autochthonous lactic acid bacteria isolates. The graph is a biplot displaying the projection of variables and LAB isolates formed by the two major principal components.
Figure 4.
Principal component analysis of probiotic properties (low pH resistance, bile salt resistance, auto-aggregation, co-aggregation, cell surface hydrophobicity, and antibacterial activity) of seven autochthonous lactic acid bacteria isolates. The graph is a biplot displaying the projection of variables and LAB isolates formed by the two major principal components.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Isolation and distribution of presumptive lactic acid bacteria (LAB) in Tella samples from three different areas of Ethiopia.
Table 1.
Isolation and distribution of presumptive lactic acid bacteria (LAB) in Tella samples from three different areas of Ethiopia.
| Collection Cites | No. of LAB -Positive Samples | No. of Single Isolates | Isolate Code 1 | Physiological Test | No. of Presumptive LAB Isolates 2 | |
|---|---|---|---|---|---|---|
| No. of Gram-Positive Isolates | No. of Catalase -Negative Isolates | |||||
| Addis Ababa (n = 5) | 5 | 16 | TAA01–TAA16 | 13 | 6 | 3 |
| Debre Birhan (n = 5) | 3 | 14 | TDB17–TDB30 | 9 | 6 | 6 |
| Debre Markos (n = 5) | 5 | 13 | TDM31–TDM43 | 11 | 7 | 6 |
1 T, Tella; AA, Addis Ababa; DB, Debre Birhan; DM, Debre Markos. 2 Presumptive LAB isolates are Gram-positive and catalase-negative isolates.
Table 2.
Phenotypic characterization of presumptive LAB isolates and their genotypic identification using 16S rRNA sequencing.
Table 2.
Phenotypic characterization of presumptive LAB isolates and their genotypic identification using 16S rRNA sequencing.
| Isolate Code | Cell Morphology | Acidification (ΔpH > 1) 1 | Genotype Identification | |||
|---|---|---|---|---|---|---|
| The Closest Type Strain | Query Length (bp) | Identity (%) | Accession Number | |||
| TAA01 | Coccus | + | Pediococcus pentosaceus DSM 20336 T | 1441 | 100.0 | KX886792.1 |
| TAA04 | Bacillus | + | Lactiplantibacillus curvatus JCM 1096 T | 1440 | 100.0 | LC063167.1 |
| TAA14 | Bacillus | − | ND | ND | ND | ND |
| TDB18 | Streptococcus | − | ND | ND | ND | ND |
| TDB19 | Coccus | + | Leuconostoc mesenteroides JCM 9700 T | 1418 | 100.0 | LC063167.1 |
| TDB21 | Bacillus | + | Lactiplantibacillus curvatus JCM 1096 T | 1435 | 99.9 | LC063167.1 |
| TDB22 | Coccus | + | Leuconostoc mesenteroides JCM 9700 T | 1417 | 100.0 | LC096223.1 |
| TDB23 | Coccus | − | ND | ND | ND | ND |
| TDB24 | Bacillus | − | ND | ND | ND | ND |
| TDM32 | Bacillus | − | ND | ND | ND | ND |
| TDM34 | Streptobacillus | − | ND | ND | ND | ND |
| TDM35 | Bacillus | − | ND | ND | ND | ND |
| TDM38 | Bacillus | − | ND | ND | ND | ND |
| TDM40 | Bacillus | + | Lactiplantibacillus curvatus JCM 1096 T | 1440 | 100.0 | LC063167.1 |
| TDM41 | Bacillus | + | Lactiplantibacillus plantarum JCM 1149 T | 1438 | 100.0 | LC064896.1 |
1 Acidification ability (ΔpH): the difference in pH was recorded before and after inoculation with individual isolates and incubation at 30 °C for 4 h. T, type strain. +, ΔpH > 1. −, ΔpH < 1. ND, not determined.
Table 3.
Auto-aggregation, hydrophobicity, and co-aggregation properties of autochthonous lactic acid bacteria (LAB) isolates.
Table 3.
Auto-aggregation, hydrophobicity, and co-aggregation properties of autochthonous lactic acid bacteria (LAB) isolates.
| LAB Isolates | Auto- Aggregation (%) | Co-Aggregation (%) | Hydrophobicity (%) | |||
|---|---|---|---|---|---|---|
| E. coli ATCC 43895 | S. Enteritidis ATCC 13076 | S. aureus ATCC 25923 | Xylene | Chloroform | ||
| P. pentosaceus TAA01 | 31.7 ± 0.1 b | 23.3 ± 0.1 c | 29.4 ± 0.8 a | 32.0 ± 0.1 b | 32.2 ± 0.6 c | 35.3 ± 0.3 c |
| L. curvatus TAA04 | 33.4 ± 1.1 b | 26.5 ± 0.4 b | 26.6 ± 0.1 b | 27.0 ± 0.4 c | 31.7 ± 0.6 c | 39.8 ± 0.6 b |
| L. mesenteroides TDB19 | 24.5 ± 1.9 c | 19.5 ± 0.3 f | 20.7 ± 0.2 d | 21.6 ± 0.1 e | 17.0 ± 0.4 e | 18.0 ± 0.1 g |
| L. curvatus TDB21 | 33.8 ± 1.4 b | 23.8 ± 0.2 c | 21.2 ± 0.1 d | 24.2 ± 0.4 d | 28.1 ± 0.9 d | 25.9 ± 0.6 f |
| L. mesenteroides TDB22 | 32.3 ± 0.5 b | 20.5 ± 0.5 e | 29.9 ± 0.8 a | 20.2 ± 0.1 f | 34.5 ± 0.2 b | 28.1 ± 0.1 e |
| L. curvatus TDM40 | 34.5 ± 1.8 b | 21.9 ± 0.4 d | 23.2 ± 0.4 c | 24.0 ± 0.4 d | 29.2 ± 1.2 d | 31.6 ± 0.6 d |
| L. plantarum TDM41 | 44.9 ± 1.7 a | 34.0 ± 0.5 a | 23.5 ± 0.5 c | 41.4 ± 0.2 a | 45.4 ± 0.1 a | 52.1 ± 0.1 a |
Data represent the mean ± standard deviation of three independent experiments. Different superscript letters (a–g) represent significant differences (p < 0.05) among the means within the same column.
Table 4.
Antibiotic susceptibilities of autochthonous lactic acid bacteria (LAB) isolates.
| LAB Isolates | Antibiotics 1 | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| AMP | GEN | KAN | STR | ERY | TET | CHL | PEN | SXT | CIP | AZM | |
| P. pentosaceus TAA01 | R | R | R | R | S | S | S | I | R | R | S |
| L. curvatus TAA04 | R | R | R | R | S | S | S | S | R | I | S |
| L. mesenteroides TDB19 | R | R | R | R | S | S | S | I | R | I | S |
| L. curvatus TDB21 | R | R | R | R | S | S | S | S | R | R | S |
| L. mesenteroides TDB22 | R | R | R | R | S | S | S | I | R | R | S |
| L. curvatus TDM40 | R | R | R | R | S | S | S | S | R | I | S |
| L. plantarum TDM41 | R | R | R | R | S | S | S | I | S | R | S |
1 AMP, ampicillin (10 µg); GEN, gentamicin (10 µg); KAN, kanamycin (30 µg); STR, streptomycin (10 µg); ERY, erythromycin (15 µg); TET, tetracycline (30 µg); CHL, chloramphenicol (30 µg); PEN, penicillin G (10 µg); SXT, trimethoprim/sulfamethoxazole (110 µg); CIP, ciprofloxacin (5 µg); and AZM, azithromycin (10 µg). R, resistance; S, susceptibility; I, moderate susceptibility.
Table 5.
Antibacterial activity of autochthonous lactic acid bacteria (LAB) isolates against three foodborne pathogens.
Table 5.
Antibacterial activity of autochthonous lactic acid bacteria (LAB) isolates against three foodborne pathogens.
| LAB Isolates | Inhibition Zone (mm) | ||
|---|---|---|---|
| E. coli ATCC 43895 | S. Enteritidis ATCC 13076 | S. aureus ATCC 25923 | |
| P. pentosaceus TAA01 | 16.0 ± 0.7 | 16.3 ± 0.8 | 12.7 ± 0.5 |
| L. curvatus TAA04 | 12.0 ± 0.7 | 13.0 ± 1.0 | ND |
| L. mesenteroides TDB19 | ND | ND | 9.7 ± 0.5 |
| L. curvatus TDB21 | 11.0 ± 1.4 | 9.7 ± 0.6 | 13.5 ± 0.7 |
| L. mesenteroides TDB22 | 14.0 ± 0.7 | 17.0 ± 1.0 | 14.0 ± 1.0 |
| L. curvatus TDM40 | 11.3 ± 1.2 | 9.0 ± 1.0 | ND |
| L. plantarum TDM41 | 14.7 ± 1.1 | 16.0 ± 1.0 | 14.0 ± 1.7 |
Data represent the mean ± standard deviation of three independent experiments. ND, not detected.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Yehuala, G.A.; Shibeshi, N.T.; Kim, S.-H.; Park, M.-K. Characterization of Autochthonous Lactic Acid Bacteria Isolated from a Traditional Ethiopian Beverage, Tella. Foods 2024, 13, 575. https://doi.org/10.3390/foods13040575
AMA Style
Yehuala GA, Shibeshi NT, Kim S-H, Park M-K. Characterization of Autochthonous Lactic Acid Bacteria Isolated from a Traditional Ethiopian Beverage, Tella. Foods. 2024; 13(4):575. https://doi.org/10.3390/foods13040575
Chicago/Turabian StyleYehuala, Gashaw Assefa, Nurelegne Tefera Shibeshi, Su-Hyeon Kim, and Mi-Kyung Park. 2024. "Characterization of Autochthonous Lactic Acid Bacteria Isolated from a Traditional Ethiopian Beverage, Tella" Foods 13, no. 4: 575. https://doi.org/10.3390/foods13040575
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.