

Lignocellulose Degrading Weizmannia coagulans Capable of Enantiomeric L-Lactic Acid Production via Consolidated Bioprocessing
 , ,
, ,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Culture Media
2.2. Isolation and Screening of Presumptive Bacillus coagulans (Weizmannia coagulans) Strains from Soil Samples
2.3. Identification and Biochemical Characterization
2.4. Potential Evaluation of Weizmannia coagulans Strains for Lignocellulolytic Enzyme Production
2.5. Effect of Different Agricultural Residues on Lignocellulolytic Enzyme Production
2.6. CBP of Lignocellulosic Feedstock to Lactic Acid
2.6.1. Sources of Lignocellulose
2.6.2. CBP of Lignocellulosic Feedstocks to Lactic Acid
2.6.3. CBP of Pretreated Lignocellulosic Feedstocks to Lactic Acid
2.7. Analytical Methods
2.7.1. Enzyme Activity Assay
2.8. Lactic Acid Production Analysis
3. Results and Discussion
3.1. Isolation of Presumptive Weizmannia coagulans
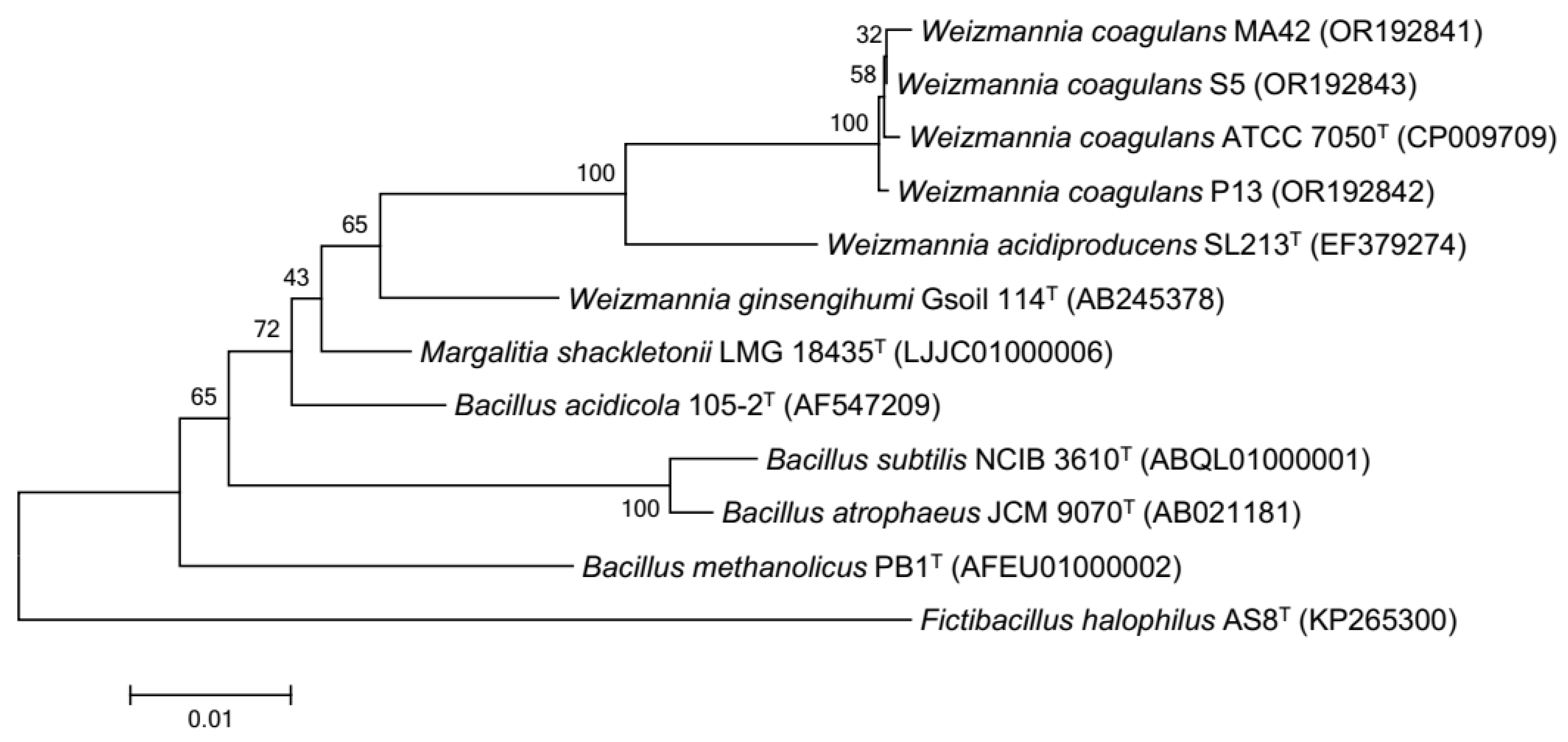
3.2. Biochemcial Characterization and Molecular Identification of the Presumptive Weizmannia coagulans
3.3. Selection of Weizmannia coagulans Strain for CBP of Lactic Acid Production Based on Lignocellulosic Enzyme Production
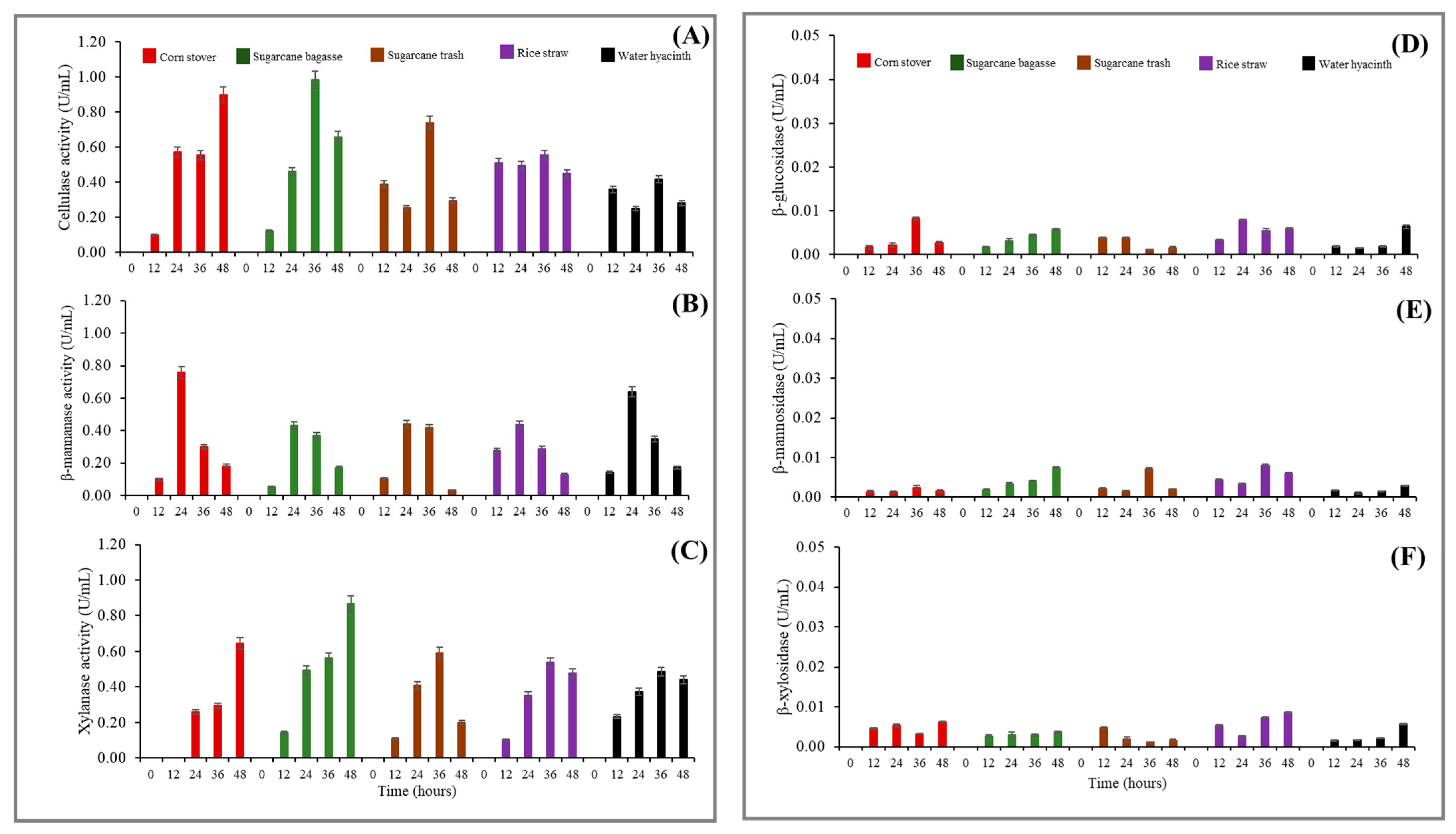
3.4. Effect of Different Agricultural Residues on Lignocellulolytic Enzyme Production
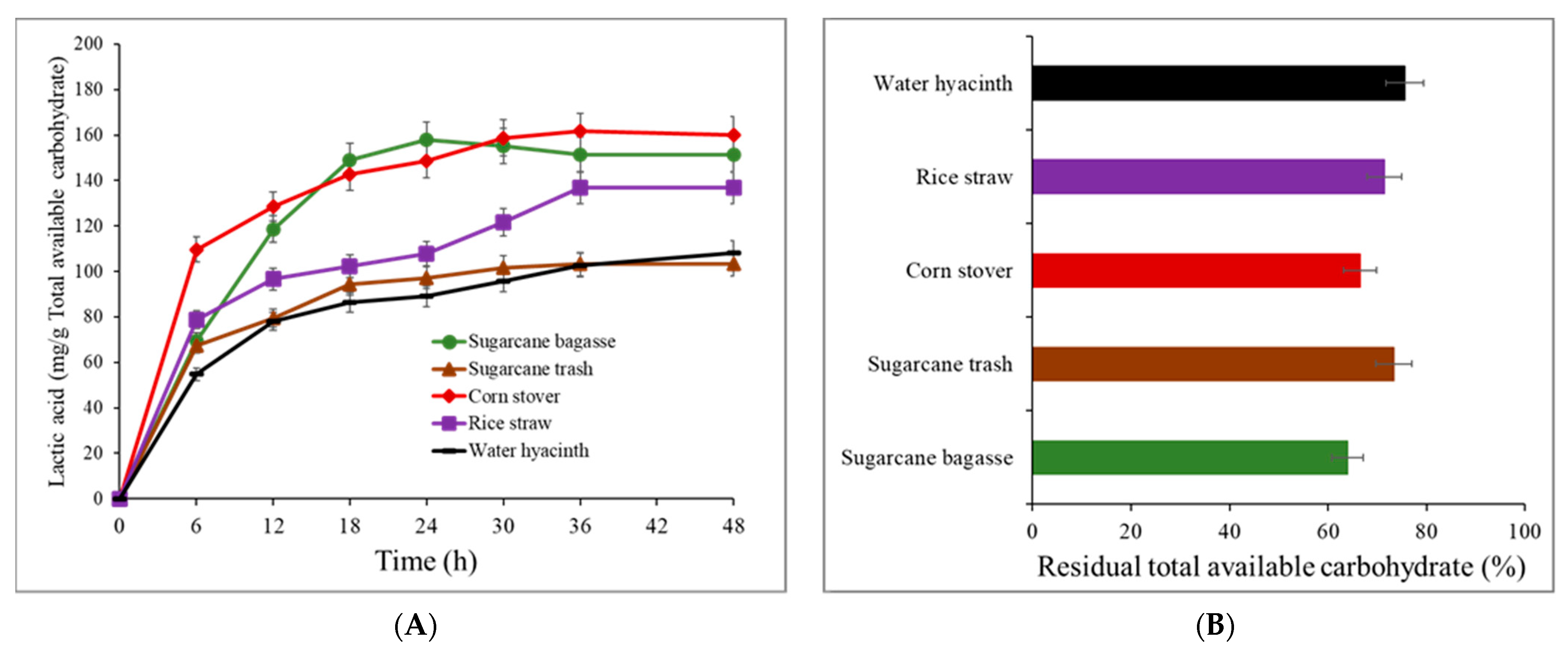
3.5. CBP Lactic Acid Production from Lignocellulose Materials
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vijayakumar, J.; Aravindan, R.; Viruthagiri, T. Recent trends in the production, purification and application of lactic acid. Chem. Biochem. Eng. Q. 2008, 22, 245–264. [Google Scholar]
- Poudel, P.; Tashiro, Y.; Sakai, K. New application of Bacillus strains for optically pure l-lactic acid production: General overview and future prospects. Biosci. Biotechnol. Biochem. 2016, 80, 642–654. [Google Scholar] [CrossRef] [PubMed]
- Farah, S.; Anderson, D.G.; Langer, R. Physical and mechanical properties of PLA, and their functions in widespread applications-a comprehensive review. Adv. Drug Deliv. Rev. 2016, 107, 367–392. [Google Scholar] [PubMed]
- Christensen, C.H.; Rass-Hansen, J.; Marsden, C.C.; Taarning, E.; Egeblad, K. The renewable chemicals industry. In ChemSusChem: Chemistry & Sustainability Energy & Materials; Claire, M.C., Ed.; Wiley-VCH GmbH: Weinheim, Germany, 2008; Volume 1, pp. 283–289. [Google Scholar]
- Patel, A.; Shah, A.R. Integrated lignocellulosic biorefinery: Gateway for production of second-generation ethanol and value added products. J. Bioresour. Bioprod. 2021, 6, 108–128. [Google Scholar]
- Aulitto, M.; Fusco, S.; Bartolucci, S.; Franzén, C.J.; Contursi, P. Bacillus coagulans MA-13: A promising thermophilic and cellulolytic strain for the production of lactic acid from lignocellulosic hydrolysate. Biotechnol. Biofuels 2017, 10, 210. [Google Scholar] [CrossRef] [PubMed]
- Ye, L.; Zhou, X.; Hudari, M.S.B.; Li, Z.; Wu, J.C. Highly efficient production of L-lactic acid from xylose by newly isolated Bacillus coagulans C106. Bioresour. Technol. 2013, 132, 38–44. [Google Scholar] [CrossRef] [PubMed]
- Chundawat, S.P.; Beckham, G.T.; Himmel, M.E.; Dale, B.E. Deconstruction of lignocellulosic biomass to fuels and chemicals. Annu. Rev. Chem. Biomol. Eng. 2011, 2, 121–145. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, M.; Meyer, A.S. Lignocellulose pretreatment severity–relating pH to biomatrix opening. New Biotechnol. 2010, 27, 739–750. [Google Scholar] [CrossRef]
- Ouyang, J.; Liu, B.; Zhang, M.; Zheng, Z.; Yu, H. Enzymatic hydrolysis, adsorption, and recycling during hydrolysis of bagasse sulfite pulp. Bioresour. Technol. 2013, 146, 288–293. [Google Scholar] [CrossRef]
- Van Zyl, W.H.; Lynd, L.R.; den Haan, R.; McBride, J.E. Consolidated bioprocessing for bioethanol production using Saccharomyces cerevisiae. In Biofuels; Scheper, T., Ed.; Springer: Berlin/Heidelberg, Germany, 2007; pp. 205–235. [Google Scholar]
- Olguin-Maciel, E.; Singh, A.; Chable-Villacis, R.; Tapia-Tussell, R.; Ruiz, H.A. Consolidated bioprocessing, an innovative strategy towards sustainability for biofuels production from crop residues: An overview. Agron. J. 2020, 10, 1834. [Google Scholar] [CrossRef]
- Van Dyk, J.; Pletschke, B. A review of lignocellulose bioconversion using enzymatic hydrolysis and synergistic cooperation between enzymes-factors affecting enzymes, conversion and synergy. Bioresour. Adv. 2012, 30, 1458–1480. [Google Scholar] [CrossRef]
- Rhee, M.S.; Moritz, B.E.; Xie, G.; Glavina Del Rio, T.; Dalin, E.; Tice, H.; Bruce, D.; Goodwin, L.; Chertkov, O.; Brettin, T. Complete genome sequence of a thermotolerant sporogenic lactic acid bacterium, Bacillus coagulans strain 36D1. Stand. Genom. Sci. 2011, 5, 331–340. [Google Scholar] [CrossRef] [PubMed]
- Yao, K.; Zhou, Q.-x.; Liu, D.-m.; Chen, S.-m.; Yuan, K. Comparative proteomics of the metabolic pathways involved in l-lactic acid production in Bacillus coagulans BCS13002 using different carbon sources. LWT-Food Sci. Technol. 2019, 116, 108445. [Google Scholar] [CrossRef]
- Maas, R.H.; Bakker, R.R.; Jansen, M.L.; Visser, D.; De Jong, E.; Eggink, G.; Weusthuis, R.A. Lactic acid production from lime-treated wheat straw by Bacillus coagulans: Neutralization of acid by fed-batch addition of alkaline substrate. Appl. Microbiol. Biotechnol. 2008, 78, 751–758. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Zhang, Z.; Lin, Y.; Zhao, S.; Mei, Y.; Liang, Y.; Peng, N. High-titer lactic acid production from NaOH-pretreated corn stover by Bacillus coagulans LA204 using fed-batch simultaneous saccharification and fermentation under non-sterile condition. Bioresour. Technol. 2015, 182, 251–257. [Google Scholar] [CrossRef]
- Fu, J.; Wang, Z.; Miao, H.; Yu, C.; Zheng, Z.; Ouyang, J. Rapid adaptive evolution of Bacillus coagulans to undetoxified corncob hydrolysates for lactic acid production and new insights into its high phenolic degradation. Bioresour. Technol. 2023, 383, 129246. [Google Scholar] [CrossRef] [PubMed]
- Nakajo, M.; Moriyama, Y. Development of selective medium and counting method for heat resistant spores of Bacillus coagulan. Nippon Shokuhin Kogyo Gakkaishi 1994, 41, 281–286. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A User-Friendly Biological Sequence Alignment Editor and Analysis Program for Windows 95/98/NT; Nucleic Acids Symposium Series; Oxford University Press: Oxford, UK, 1999; Volume 41, pp. 95–98. [Google Scholar]
- Tamura, K.; Dudley, J.; Nei, M.; Kumar, S. MEGA4: Molecular evolutionary genetics analysis (MEGA) software version 4.0. Mol. Biol. Evol. 2007, 24, 1596–1599. [Google Scholar] [CrossRef]
- Unban, K.; Khatthongngam, N.; Pattananandecha, T.; Saenjum, C.; Shetty, K.; Khanongnuch, C. Microbial community dynamics during the non-filamentous fungi growth-based fermentation process of Miang, a traditional fermented tea of north Thailand and their product characterizations. Front. Microbiol. 2020, 11, 1515. [Google Scholar] [CrossRef]
- DuBois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.T.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Wongputtisin, P.; Khanongnuch, C.; Khongbantad, W.; Niamsup, P.; Lumyong, S. Screening and selection of Bacillus spp. for fermented corticate soybean meal production. J. Appl. Microbiol. 2012, 113, 798–806. [Google Scholar] [CrossRef]
- Miller, G.L. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Thompson, P.J. Thermophilic organisms involved in food spoilage: Aciduric flat-sour sporeforming aerobes. J. Food Prot. 1981, 44, 154–156. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.; Xu, P.; Tao, F. L-lactic acid production by Bacillus coagulans through simultaneous saccharification and fermentation of lignocellulosic corncob residue. Bioresour. Technol. Rep. 2019, 6, 131–137. [Google Scholar] [CrossRef]
- Zou, L.; Ouyang, S.; Hu, Y.; Zheng, Z.; Ouyang, J. Efficient lactic acid production from dilute acid-pretreated lignocellulosic biomass by a synthetic consortium of engineered Pseudomonas putida and Bacillus coagulans. Biotechnol. Biofuels 2021, 14, 227. [Google Scholar] [CrossRef]
- Tri, C.L.; Kamei, I. The improvement of sodium hydroxide pretreatment in bioethanol production from Japanese bamboo Phyllostachys edulis using the white rot fungus Phlebia sp. MG-60. Int. Biodeterior. Biodegrad. 2018, 133, 86–92. [Google Scholar] [CrossRef]
- Wang, Y.; Cao, W.; Luo, J.; Wan, Y. Exploring the potential of lactic acid production from lignocellulosic hydrolysates with various ratios of hexose versus pentose by Bacillus coagulans IPE22. Bioresour. Technol. 2018, 261, 342–349. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.S.; Patel, S.; Saini, N.; Chen, S. Robust demarcation of 17 distinct Bacillus species clades, proposed as novel Bacillaceae genera, by phylogenomics and comparative genomic analyses: Description of Robertmurraya kyonggiensis sp. nov. and proposal for an emended genus Bacillus limiting it only to the members of the Subtilis and Cereus clades of species. Int. J. Syst. Evol. Microbiol. 2020, 70, 5753–5798. [Google Scholar]
- Konuray Altun, G.; Erginkaya, Z. Identification and characterization of Bacillus coagulans strains for probiotic activity and safety. LWT-Food Sci. Technol. 2021, 151, 112233. [Google Scholar] [CrossRef]
- Abada, E.A.E.-m. Isolation and characterization of a antimicrobial compound from Bacillus coagulans. Anim. Cells Syst. 2008, 12, 41–46. [Google Scholar] [CrossRef]
- Cubas-Cano, E.; López-Gómez, J.P.; González-Fernández, C.; Ballesteros, I.; Tomás-Pejó, E. Towards sequential bioethanol and l-lactic acid co-generation: Improving xylose conversion to l-lactic acid in presence of lignocellulosic ethanol with an evolved Bacillus coagulans. Renew. Energy 2020, 153, 759–765. [Google Scholar] [CrossRef]
- Lynd, L.R.; Weimer, P.J.; Van Zyl, W.H.; Pretorius, I.S. Microbial cellulose utilization: Fundamentals and biotechnology. Microbiol. Mol. Biol. Rev. 2002, 66, 506–577. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Fan, H.; Mou, H.; Liu, J.; Huang, J.; Dong, X.; Song, H. Preparation and characterization of xylan by an efficient approach with mechanical pretreatments. Ind. Crop. Prod. 2021, 165, 113420. [Google Scholar] [CrossRef]
- Sachslehner, A.; Nidetzky, B.; Kulbe, K.D.; Haltrich, D. Induction of mannanase, xylanase, and endoglucanase activities in Sclerotium rolfsii. Appl. Environ. Microbiol. 1998, 64, 594–600. [Google Scholar] [CrossRef] [PubMed]
- MacKenzie, C.R.; Bilous, D.; Schneider, H.; Johnson, K.G. Induction of cellulolytic and xylanolytic enzyme systems in Streptomyces spp. Appl. Environ. Microbiol. 1987, 53, 2835–2839. [Google Scholar] [CrossRef]
- Kanwar, S.S.; Ghazi, I.A.; Chimni, S.S.; Joshi, G.K.; Rao, G.V.; Kaushal, R.K.; Gupta, R.; Punj, V. Purification and properties of a novel extra-cellular thermotolerant metallolipase of Bacillus coagulans MTCC-6375 isolate. Protein Expr. Purif. 2006, 46, 421–428. [Google Scholar] [CrossRef]
- Babu, K.R.; Satyanarayana, T. α-Amylase production by thermophilic Bacillus coagulans in solid state fermentation. Process Biochem. 1995, 30, 305–309. [Google Scholar] [CrossRef]
- Choudhury, B.; Chauhan, S.; Singh, S.; Ghosh, P. Production of xylanase of Bacillus coagulans and its bleaching potential. World J. Microbiol. Biotechnol. 2006, 22, 283–288. [Google Scholar] [CrossRef]
- ABBAS, H.; Rizal, Y.; Dharma, A.; Kompiang, I.P. Selection and identification of cellulase-producing bacteria isolated from the litter of mountain and swampy forest. Microbiol. Indones. 2007, 1, 7. [Google Scholar]
- Acharya, S.; Chaudhary, A. Optimization of fermentation conditions for cellulases production by Bacillus licheniformis MVS1 and Bacillus sp. MVS3 isolated from Indian hot spring. Braz. Arch. Biol. Technol. 2012, 55, 497–503. [Google Scholar] [CrossRef]
- Bukola, A.-T.; Temitope, E.; Gbemisola, A.; Iyadunni, O. Screening and production of mannanase by Bacillus strains isolated from fermented food condiments. Innov. Rom. Food Biotechnol. 2013, 13, 53–62. [Google Scholar]
- Chaipoot, S.; Phongphisutthinant, R.; Sriwattana, S.; Ounjaijean, S.; Wiriyacharee, P. Preparation of isoflavone glucosides from Soy germ and β-Glucosidase from Bacillus coagulans PR03 for isoflavone aglycones production. Chiang Mai Univ. J. Nat. Sci. 2019, 18, 479–497. [Google Scholar] [CrossRef]
- Svetlitchnyi, V.A.; Svetlichnaya, T.P.; Falkenhan, D.A.; Swinnen, S.; Knopp, D.; Läufer, A. Direct conversion of cellulose to L-lactic acid by a novel thermophilic Caldicellulosiruptor strain. Biotechnol. Biofuels Bioprod. 2022, 15, 44. [Google Scholar] [CrossRef]
- Gandini, C.; Tarraran, L.; Kalemasi, D.; Pessione, E.; Mazzoli, R. Recombinant Lactococcus lactis for efficient conversion of cellodextrins into L-lactic acid. Biotechnol. Bioeng. 2017, 114, 2807–2817. [Google Scholar] [CrossRef]
- Mazzoli, R.; Olson, D.G.; Concu, A.M.; Holwerda, E.K.; Lynd, L.R. In vivo evolution of lactic acid hyper-tolerant Clostridium thermocellum. New Biotechnol. 2022, 67, 12–22. [Google Scholar] [CrossRef] [PubMed]
- Tomás-Pejó, E.; Alvira, P.; Ballesteros, M.; Negro, M.J. Chapter 7—Pretreatment technologies for lignocellulose-to-bioethanol conversion. In Biofuels; Pandey, A., Larroche, C., Ricke, S.C., Dussap, C.-G., Gnansounou, E., Eds.; Academic Press: Amsterdam, The Netherlands, 2011; pp. 149–176. [Google Scholar]
- Schroedter, L.; Schneider, R.; Remus, L.; Venus, J. L-(+)-lactic acid from reed: Comparing various resources for the nutrient provision of B. coagulans. Resources 2020, 9, 89. [Google Scholar] [CrossRef]
- de Oliveira, R.A.; Schneider, R.; Rossell, C.E.V.; Maciel Filho, R.; Venus, J. Polymer grade l-lactic acid production from sugarcane bagasse hemicellulosic hydrolysate using Bacillus coagulans. Bioresour. Technol. Rep. 2019, 6, 26–31. [Google Scholar] [CrossRef]
- Schroedter, L.; Streffer, F.; Streffer, K.; Unger, P.; Venus, J. Biorefinery concept employing Bacillus coagulans: LX-lignin and L-(+)-lactic acid from lignocellulose. Microorganisms 2021, 9, 1810. [Google Scholar] [CrossRef]
- Abdel-Rahman, M.A.; Hassan, S.E.; Roushdy, M.M.; Azab, M.S.; Gaber, M.A. Free-nutrient supply and thermos-alkaline conditions for direct lactic acid production from mixed lignocellulosic and food waste materials. Bioresour. Technol. Rep. 2019, 7, 100256. [Google Scholar] [CrossRef]
- Hassan, S.E.-D.; Abdel-Rahman, M.A.; Roushdy, M.M.; Azab, M.S.; Gaber, M.A. Effective biorefinery approach for lactic acid production based on co-fermentation of mixed organic wastes by Enterococcus durans BP130. Biocatal. Agric. Biotechnol. 2019, 20, 101203. [Google Scholar] [CrossRef]
- Shahab, R.L.; Luterbacher, J.S.; Brethauer, S.; Studer, M.H. Consolidated bioprocessing of lignocellulosic biomass to lactic acid by a synthetic fungal-bacterial consortium. Biotechnol. Bioeng. 2018, 115, 1207–1215. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, J.; Cai, C.; Chen, H.; Jiang, T.; Zheng, Z. Efficient non-sterilized fermentation of biomass-derived xylose to lactic acid by a thermotolerant Bacillus coagulans NL01. Appl. Biochem. Biotechnol. 2012, 168, 2387–2397. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Rahman, M.A.; Hassan, S.E.-D.; Alrefaey, H.M.; Elsakhawy, T. Efficient co-utilization of biomass-derived mixed sugars for lactic acid production by Bacillus coagulans Azu-10. Fermentation 2021, 7, 28. [Google Scholar] [CrossRef]
- Tolieng, V.; Tanaka, N.; Shiwa, Y.; Thitiprasert, S.; Kanchanasin, P.; Phongsopitanun, W.; Booncharoen, A.; Thongchul, N.; Tanasupawat, S. Weizmannia acidilactici sp. nov., a lactic acid producing bacterium isolated from soils. Syst. Appl. Microbiol. 2023, 46, 126389. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| NO. | Isolate | Source | D-Lactic Acid (g/L) | L-Lactic Acid (g/L) | Total Lactic Acid (g/L) | Optical Purity of L-Lactic Acid (%) |
|---|---|---|---|---|---|---|
| 1 | S1 | San Sai-1 | 0.06 ± 0.01 | 0.84 ± 0.21 | 0.90 ± 0.11 | 93.0 |
| 2 | S3 | San Sai-1 | 0.13 ± 0.05 | 0.81 ± 0.21 | 0.94 ± 0.13 | 86.2 |
| 3 | S4 | San Sai-1 | 0.16 ± 0.02 | 1.10 ± 0.21 | 1.26 ± 0.12 | 87.3 |
| 4 | S5 | San Sai-1 | 0 | 1.43 ± 0.16 | 1.43 ± 0.16 | 100.0 |
| 5 | P10 | Arboretum | 0.32 ± 0.05 | 0.84 ± 0.20 | 1.16 ± 0.13 | 72.4 |
| 6 | P12 | Arboretum | 0.03 ± 0.01 | 0.61 ± 0.21 | 0.64 ± 0.11 | 95.3 |
| 7 | P13 | Arboretum | 0 | 1.74 ± 0.07 | 1.74 ± 0.07 | 100.0 |
| 8 | MA26 | Mae Aen-2 | 0.22 ± 0.02 | 1.97 ± 0.22 | 2.19 ± 0.12 | 90.0 |
| 9 | MA42 | Mae Aen-1 | 0 | 1.60 ± 0.21 | 1.60 ± 0.21 | 100.0 |
| 10 | MA47 | Mae Aen-1 | 0.22 ± 0.05 | 1.40 ± 0.30 | 1.62 ± 0.18 | 86.4 |
| 11 | MA48 | Mae Aen-1 | 0.29 ± 0.01 | 1.00 ± 0.05 | 1.29 ± 0.03 | 77.5 |
| 12 | R55 | Rice field | 0.35 ± 0.08 | 1.94 ± 0.04 | 2.29 ± 0.06 | 84.7 |
| 13 | SS-59 | SanSai-2 | 0.16 ± 0.02 | 0.81 ± 0.21 | 0.97 ± 0.12 | 83.5 |
| 14 | SS-60 | SanSai-2 | 0.06 ± 0.01 | 0.78 ± 0.21 | 0.84 ± 0.11 | 92.9 |
| 15 | SS-68 | SanSai-2 | 0.03 ± 0.01 | 0.87 ± 0.20 | 0.90 ± 0.10 | 96.7 |
| Characteristics | MA42 | P13 | S5 |
|---|---|---|---|
| Shape | Rod | Rod | Rod |
| Gram staining | + | + | + |
| Endospore formation | + | + | + |
| Growth under aerobic condition * | + | + | + |
| Growth under anaerobic condition * | + | + | + |
| Growth in carbon source ** | |||
| D-glucose | + | + | + |
| D-xylose | + | + | + |
| Carboxyl methyl cellulose (CMC) | + | + | + |
| Locust bean gum (LBG) | + | + | + |
| Beechwood xylan | + | + | − |
| Optimal temperature for growth (°C) * | 50 | 50 | 45 |
| Optimal initial pH for growth * | 5–7 | 6–7 | 5–7 |
| Enzyme production | |||
| Cellulase (NA + CMC) | + | + | + |
| β-Mannanase (NA + LBG) | + | + | + |
| Xylanase (NA + xylan) | + | + | − |
| Lactic acid production from xylose (g/L) * | |||
| Control pH | 7.88 ± 0.20 | 6.76 ± 0.13 | 7.13 ± 0.16 |
| Non-control pH | 1.73 ± 0.12 | 1.54 ± 0.10 | 1.63 ± 0.11 |
| Isolate | Cellulase Activity (U/mL) | β-Mannanase Activity (U/mL) | Xylanase Activity (U/mL) |
|---|---|---|---|
| MA42 | 0.370 ± 0.01 b | 0.473 ± 0.01 a | 0.299 ± 0.01 a |
| P13 | 0.473 ± 0.01 a | 0.424 ± 0.01 b | 0.094 ± 0.01 b |
| S5 | 0.009 ± 0.00 c | 0.004 ± 0.00 c | ND |
| Materials | Treatment | Lactic Acid Production Efficiency | Increase (%) | Total Available Carbohydrate Retained in Solid Residue | |
|---|---|---|---|---|---|
| (mg/g Substrate) | (mg/g Total Available Substrate) | ||||
| Sugarcane bagasse | Untreated | 77.6 ± 1.1 d | 148.4 ± 3.9 e | - | 64.0 |
| 0.25 M H2SO4 | 149.2 ± 1.2 a | 553.4 ± 2.9 a | 92.4 | 44.0 | |
| 0.05 M H2SO4 | 111.1 ± 2.1 b | 230.6 ± 1.0 b | 43.2 | 50.3 | |
| 0.25 M NaOH | 100.8 ± 3.2 c | 216.8 ± 1.6 c | 29.9 | 48.1 | |
| 0.05 M NaOH | 81.4 ± 2.2 d | 170.1 ± 1.0 d | 4.9 | 58.7 | |
| Sugarcane trash | Untreated | 44.6 ± 2.2 d | 101.8 ± 1.2 d | - | 73.3 |
| 0.25 M H2SO4 | 91.8 ± 1.1 a | 325.4 ± 4.1 a | 105.8 | 46.8 | |
| 0.05 M H2SO4 | 64.6 ± 3.3 c | 134.8 ± 1.9 c | 44.9 | 60.9 | |
| 0.25 M NaOH | 86.0 ± 2.2 b | 318.9 ± 4.1 a | 92.8 | 50.3 | |
| 0.05 M NaOH | 62.0 ± 1.1 c | 173.7 ± 2.2 b | 39.1 | 64.1 | |
| Corn stover | Untreated | 71.8 ± 1.1 d | 160.7 ± 1.7 d | - | 66.5 |
| 0.25 M H2SO4 | 117.6 ± 4.2 a | 326.6 ± 4.4 a | 64.0 | 46.8 | |
| 0.05 M H2SO4 | 78.9 ± 3.3 c | 176.0 ± 2.3 c | 9.9 | 64.8 | |
| 0.25 M NaOH | 109.9 ± 1.1 b | 295.0 ± 3.8 b | 53.2 | 47.2 | |
| 0.05 M NaOH | 74.0 ± 2.1 cd | 176.1 ± 2.2 c | 3.1 | 68.5 | |
| Rice straw | Untreated | 64.0 ± 2.2 d | 136.8 ± 1.5 d | - | 71.4 |
| 0.25 M H2SO4 | 114.3 ± 4.1 a | 528.0 ± 7.2 a | 78.7 | 45.5 | |
| 0.05 M H2SO4 | 78.8 ± 1.1 c | 175.9 ± 2.1 c | 23.1 | 54.2 | |
| 0.25 M NaOH | 101.4 ± 3.3 b | 219.2 ± 2.9 b | 58.5 | 50.8 | |
| 0.05 M NaOH | 67.2 ± 2.2 d | 138.9 ± 1.8 d | 4.9 | 61.2 | |
| Water hyacinth | Untreated | 51.1 ± 2.2 e | 110.2 ± 1.3 e | - | 75.6 |
| 0.25 M H2SO4 | 129.9 ± 2.2 a | 547.0 ± 2.2 a | 154.4 | 44.7 | |
| 0.05 M H2SO4 | 88.6 ± 1.1 c | 254.6 ± 3.0 c | 73.4 | 60.0 | |
| 0.25 M NaOH | 98.9 ± 4.3 b | 486.5 ± 6.4 b | 93.6 | 47.9 | |
| 0.05 M NaOH | 67.2 ± 3.2 d | 183.8 ± 2.5 d | 31.6 | 63.5 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pamueangmun, P.; Abdullahi, A.D.; Kabir, M.H.; Unban, K.; Kanpiengjai, A.; Venus, J.; Shetty, K.; Saenjum, C.; Khanongnuch, C. Lignocellulose Degrading Weizmannia coagulans Capable of Enantiomeric L-Lactic Acid Production via Consolidated Bioprocessing. Fermentation 2023, 9, 761. https://doi.org/10.3390/fermentation9080761
Pamueangmun P, Abdullahi AD, Kabir MH, Unban K, Kanpiengjai A, Venus J, Shetty K, Saenjum C, Khanongnuch C. Lignocellulose Degrading Weizmannia coagulans Capable of Enantiomeric L-Lactic Acid Production via Consolidated Bioprocessing. Fermentation. 2023; 9(8):761. https://doi.org/10.3390/fermentation9080761
Chicago/Turabian StylePamueangmun, Punnita, Aliyu Dantani Abdullahi, Md. Humayun Kabir, Kridsada Unban, Apinun Kanpiengjai, Joachim Venus, Kalidas Shetty, Chalermpong Saenjum, and Chartchai Khanongnuch. 2023. "Lignocellulose Degrading Weizmannia coagulans Capable of Enantiomeric L-Lactic Acid Production via Consolidated Bioprocessing" Fermentation 9, no. 8: 761. https://doi.org/10.3390/fermentation9080761