
Predicting Suitable Habitats for China’s Endangered Plant Handeliodendron bodinieri (H. Lév.) Rehder

1
College of Chemistry and Bioengineering, Guangxi Normal University for Nationalities, Chongzuo 532200, China
2
Biology Department of Sciences College, Qiongtai Normal University, Haikou 571127, China
3
College of Mathematics, Physics and Electronic Information Engineering, Guangxi Normal University for Nationalities, Chongzuo 532200, China
4
Department of Social Sciences, Education University of Hong Kong, Tai Po, Hong Kong, China
*
Authors to whom correspondence should be addressed.
Diversity 2023, 15(10), 1033; https://doi.org/10.3390/d15101033
Submission received: 7 July 2023
/
Revised: 15 September 2023
/
Accepted: 22 September 2023
/
Published: 25 September 2023
(This article belongs to the Section Biogeography and Macroecology)
Abstract
:Demarcating a plant species’ actual and potential biogeographical distribution is crucial for understanding the key environmental variables shaping its habitat conditions. We used MaxEnt and species distribution modeling to predict the likely range of China’s endangered species, Handeliodendron bodinieri (H. Lév.) Rehder, based on forty-four validated distribution records and eight selected environmental variables. Combined with percentage contribution and permutation importance, the jackknife statistical method was applied to test and evaluate pertinent factors restricting the potential distribution of H. bodinieri. The response curves of critical bioclimatic factors were employed to determine the potential species range. The generated MaxEnt model was confirmed to have excellent simulation accuracy. The current core potential distribution areas are concentrated in the Guangxi and Guizhou provinces of Southwest China, with a significant inter-regional difference. The precipitation of the warmest quarter (Bio18) and minimum temperature of the coldest month (Bio6) had the greatest impact on the distribution area of H. bodinieri. The findings could provide useful information and a reasonable reference for managers to enhance the protection of this declining species.
1. Introduction
Climate change has become the biggest threat to protecting and maintaining global biodiversity [1,2]. Global warming and drought constitute the main climate change stressors on biota. The fundamental abiotic alterations disturb or damage species’ habitats by changing the cardinal environmental attributes of temperature and precipitation [3,4]. Nature responds by modifying the structure, function and composition of biological communities and species distribution patterns at different scales [5,6]. In extreme cases, the diversity and stability of local and regional ecosystems could be disrupted and endangered [7].
Climate change has wrought far-reaching impacts on a broad range of plant functions and physiological processes. They include regeneration, growth pattern, morphology, anatomy, and reproductive behavior, which are key determinants of plant species distribution on a large scale [8,9]. These harmful and aberrant influences have accelerated shifting, contraction and fragmentation in the global geographical distribution patterns of many plant species. The changes have induced migration, reproductive failure and a raised mortality rate, thus increasing the number of globally endangered plant species [10]. Moreover, the survival of existing endangered plant species could be jeopardized due to straining beyond the tipping point [11].
The cumulative and serious impacts of climate change have disrupted the stability and operation of ecosystems [12,13]. For example, drastic short-term changes could kill individuals with weak adaptability and poor dissemination ability. The affected species would experience range shrinkage and local extinction [14]. Therefore, understanding the intricate relationships between potential suitable distribution patterns of endangered plants and environmental factors is critical for conservation. The knowledge can foster the monitoring and restoration of declining populations, improve resource management, and assess and abate anthropogenic impacts [15,16].
Species distribution models (e.g., MaxEnt, Garp, Bioclim, and Climex) are important tools to assess climate change’s impacts on altering the distribution of suitable areas for plant species [17,18]. These models can establish a connection between a species’ geographical distribution and environmental variables through statistical response functions. The analysis can predict potential suitable areas and future species distribution at different times in climate change scenarios [19,20]. Among these modeling approaches, the Maximum Entropy (MaxEnt) model is one of the most popular for modeling plant species distributions and their environmental niches. Its common adoption can be attributed to parsimonious data requirements, using only categorical and presence data as input variables for the species in question [21,22]. In addition, even with small sample sizes and data gaps, its forecast precision can remain stable and reliable [23,24]. The method can directly generate a spatially open habitat suitability map and evaluate the significance level of individual environmental variables using a built-in jackknife test [25,26]. The MaxEnt has been widely applied to different research fields, including conservation, ecology and evolution. More importantly, it has been employed to identify ecological niches and compare niche similarities between species pairs in geographic and environmental space [27,28].
Handeliodendron bodinieri (H. Lév.) Rehder (Sapindaceae) is a rare native monotypic plant species endemic to China. The range of the endangered small tree is confined to the karst region of the northwest Guangxi and south Guizhou provinces in Southwest China at a 500–900 m altitude [29]. This species is a semi-deciduous broad-leaf tree reaching up to 20 m in height. Its flowers are unisexual, with an aborted stamen or pistil in dioecious trees [30]. Although female plants bear fruits every year, the fruit abortion rate is high, resulting in poor natural regeneration. The oil-rich seeds are heavily consumed as food by granivorous wild animals, including invertebrates and rodents, which constitutes an important reproductive deprivation [31]. Another cause of population decline is excessive human seed collection to meet industrial and food uses, including for biodiesel extraction, as a source of protein, and as edible oil [30]. It is also harvested for its fine timber to make high-quality furniture. The natural and human pressures have jointly contributed to its continual decline.
In addition, the karst terrain is characterized by a finely divided mosaic of steep rocky hills and outcrops, with skeletal soil interrupted by depressions with thick and fertile soils. The physical environment comprises a varied habitat patchwork marked by considerable spatial heterogeneity in soil, water, and nutrients. The considerable habitat fragmentation is unfavorable to seed germination, rendering the species vulnerable to extinction. With a shrinking fragmented range, a small declining wild population, and intensification of negative forces [32], the endangered tree has been designated a second-level protected species under the List of National Key Protected Wild Plants (2021) in China (https://www.gov.cn/zhengce/zhengceku/2021-09/09/content_5636409.htm, 1 July 2023). The species has received extensive scientific investigations regarding population structure, spatial distribution, phytochemistry, reproductive ecology, genetic diversity, propagation, and population conservation [30,33]. Habitat fragmentation and drought stress have been increasingly recognized as important factors in protecting endangered species in China’s karst landscape. Excessive human seed collection has decimated living plants and degraded the quantity and quality of soil seed banks. Therefore, it is necessary and important to predict how global climate change will impact the potentially suitable distribution of H. bodinieri in China.
This study aimed to explore the suitable habitat distribution of H. bodinieri at the regional scale and focused on the following aims: (1) to define and demarcate the potential spatial distribution pattern; (2) to determine the correlation between the potential suitable distribution pattern and environmental factors.
2. Materials and Methods
2.1. Establishing Species Occurrence Records
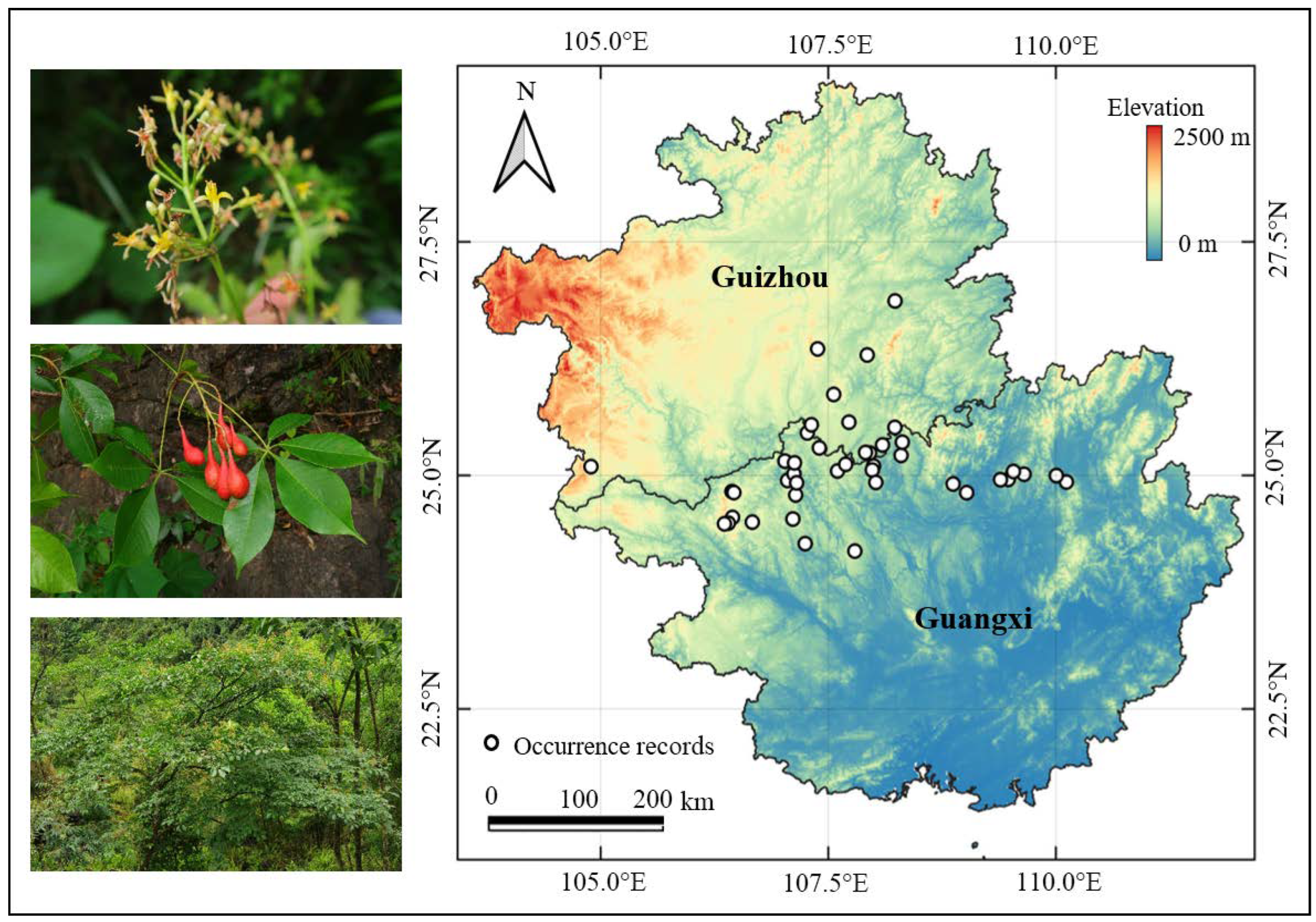
The research data are the occurrence records of H. bodinieri from 1930 to 2019. They were collected from the Chinese Virtual Herbarium (http://www.cvh.ac.cn (accessed on 6 July 2023)), the Plant Photo Bank of China (http://www.Plantphotophoto.cn (accessed on 6 July 2023)), the National Specimen Information Infrastructure (http://www.n-sii.org.cn/ (accessed on 6 July 2023)), and the published literature. Each record was scrutinized for data quality and suitability. Duplicated records and those with unclear latitude and longitude information were rejected. Finally, 44 valid occurrence records of H. bodinieri were included in this study. The longitude and latitude data of geographic distribution points were obtained from ArcGIS (10.2). The distribution points were stored in a CSV file sorted by species name and longitude, and latitude of the distribution points (Figure 1).
2.2. Selecting Environmental Variables
Nineteen bioclimatic variables, including bioclimatic parameters, monthly temperature and precipitation data, were extracted from the WorldClim database, version 2.0 (http://www.worldclim.org (accessed on 6 July 2023)) using the ‘Extract value by points’ function in DIVA-GIS 7.5 [34] (Table 1). The annual temperature and precipitation variables, considered too general, were deleted from the dataset [35,36]. Pearson correlation coefficients (r) between environmental variables were calculated. The strongly associated environmental factors |r > 0.8| were removed to reduce model overfitting due to multicollinearity and improve simulation accuracy [37,38]. Ultimately, eight of the nineteen variables were retained as evaluator variables.
2.3. MaxEnt Modeling of Species Distribution
The eight selected environmental variables and species occurrence records of H. bodinieri were loaded into MaxEnt 3.3. The package “ENMeval” of R 4.02 was used to create and evaluate the models [39,40]. For diverse environmental variables used in prediction, the jackknife test yielded training, test, and area under the curve (AUC) gains for three scenarios (without variables, with only one variable and with all variables). Then, the AUC of the receiver operating characteristic curve (ROC) was calculated to measure the accuracy of the generated models [36]. The AUC value has a range of 0–1. A higher AUC value indicates a considerable deviation in the geographic distribution of the simulation object from a random distribution. It also means a stronger correlation between the simulation result and the environmental variables, i.e., the model has a better prediction accuracy [41]. The AUC statistic is classified into five performance categories: excellent (0.9–1.0), good (0.8–0.9), fair (0.7–0.8), poor (0.6–0.7), and fail (0.5–0.6) [42]. Based on the optimal threshold of environmental factors (occurrence-probability) generated by MaxEnt, the final output with a 0–1 range was divided into five potential distribution areas using the reclassify tool of ArcGIS software: fail (0–0.15), poor (0.15–0.3), fair (0.3–0.45), good (0.45–0.6), and excellent (>0.6) [38,41].
3. Results
3.1. Evaluating Model Performance
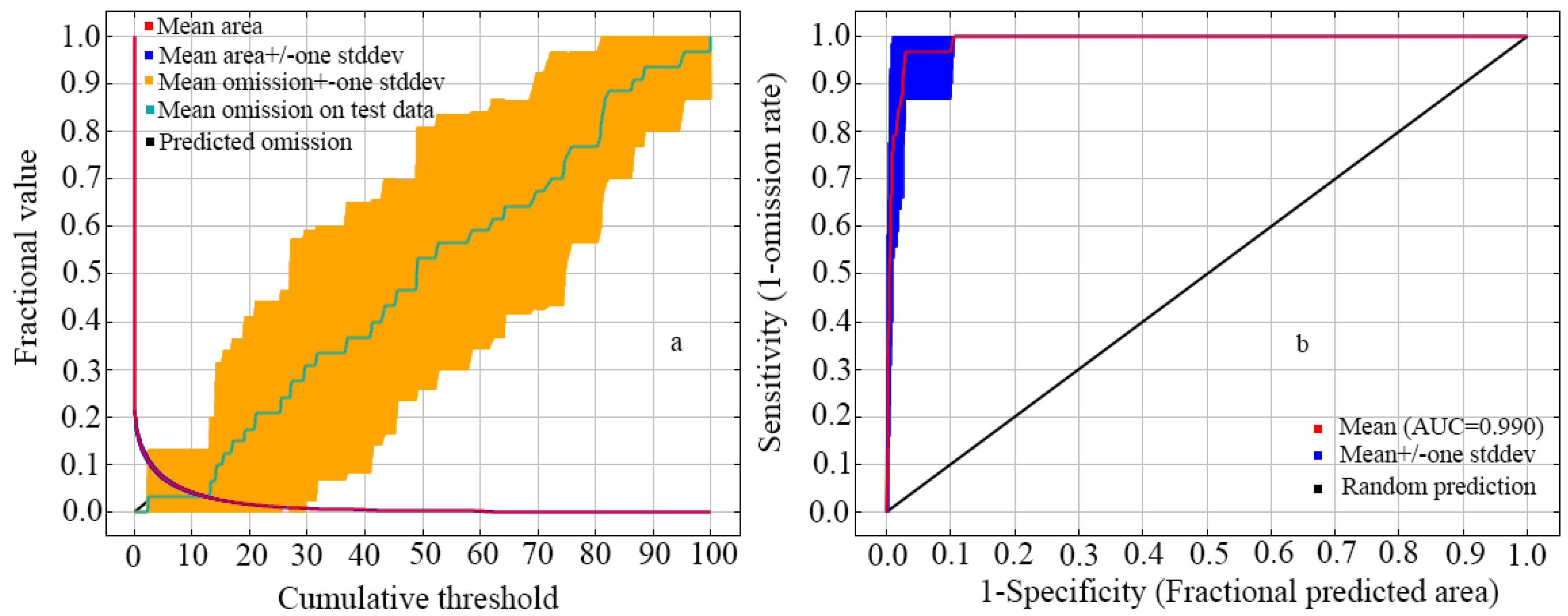
The lines of omission from the training data were close to forecasted omission rates in the model, signifying the correct fitting of the training data, and the test and training data were unique (Figure 2a). The AUC value for the reconstructed MaxEnt was 0.99 (Figure 2b), indicating excellent prediction accuracy of the model, including the selected variables. This model is assessed to be reliable for defining suitable areas for introducing H. bodinieri in China.
3.2. Key Environmental Factors and Validating Modeling Results
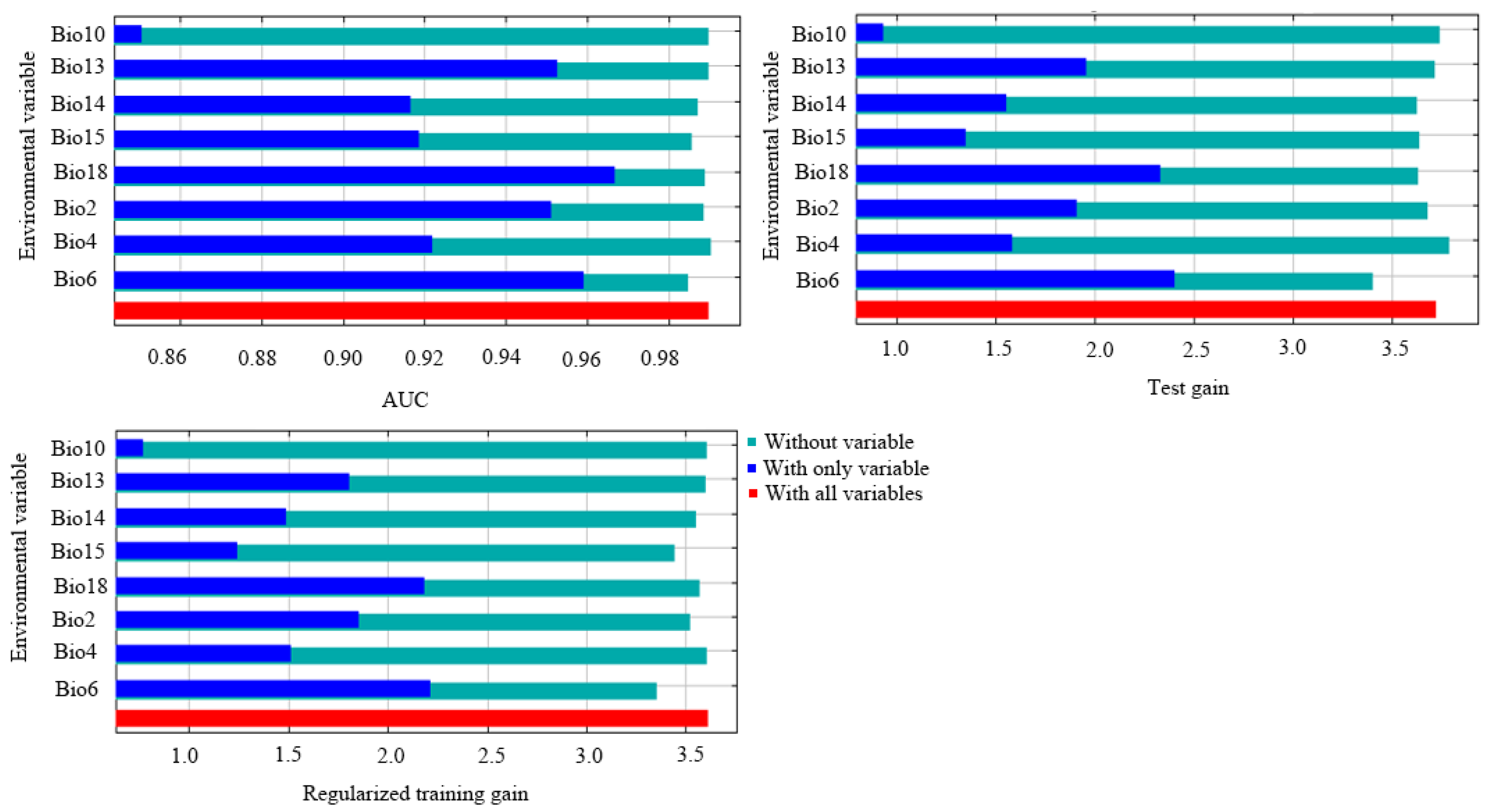
Eight variables, after removing collinearity, were determined as key environmental variables affecting the potential distribution of H. bodinieri, according to the contributions to the MaxEnt modeling process by the jackknife test (Table 2). Of these, Bio18 (precipitation of the warmest quarter), Bio6 (minimum temperature of the coldest month), and Bio2 (mean diurnal range) had the highest contributions of 32.7%, 26.3%, and 20.5%, respectively (total contribution exceeding 79.5%; Table 2). Meanwhile, Bio6 (75.7%), Bio2 (13%), and Bio18 (6.9%) had the highest permutation importance values (Table 2). Therefore, these three bioclimatic factors (Bio18, Bio6, Bio2) were the main drivers of the modern geographical distribution of H. bodinieri.
Bio6 demonstrated the highest gain in regularized training and testing. Bio18 had the highest gain in AUC, indicating its leading contribution to the distribution of H. bodinieri (Figure 3). Bio2 and Bio13 had secondary contributions to the H. bodinieri distribution under the three gain patterns. In contrast, Bio10 had the lowest gain and the least importance, with little effect on predicting species distribution (Figure 3). The results showed that temperature exerted a greater effect on H. bodinieri distribution than precipitation (Figure 3, Table 2).
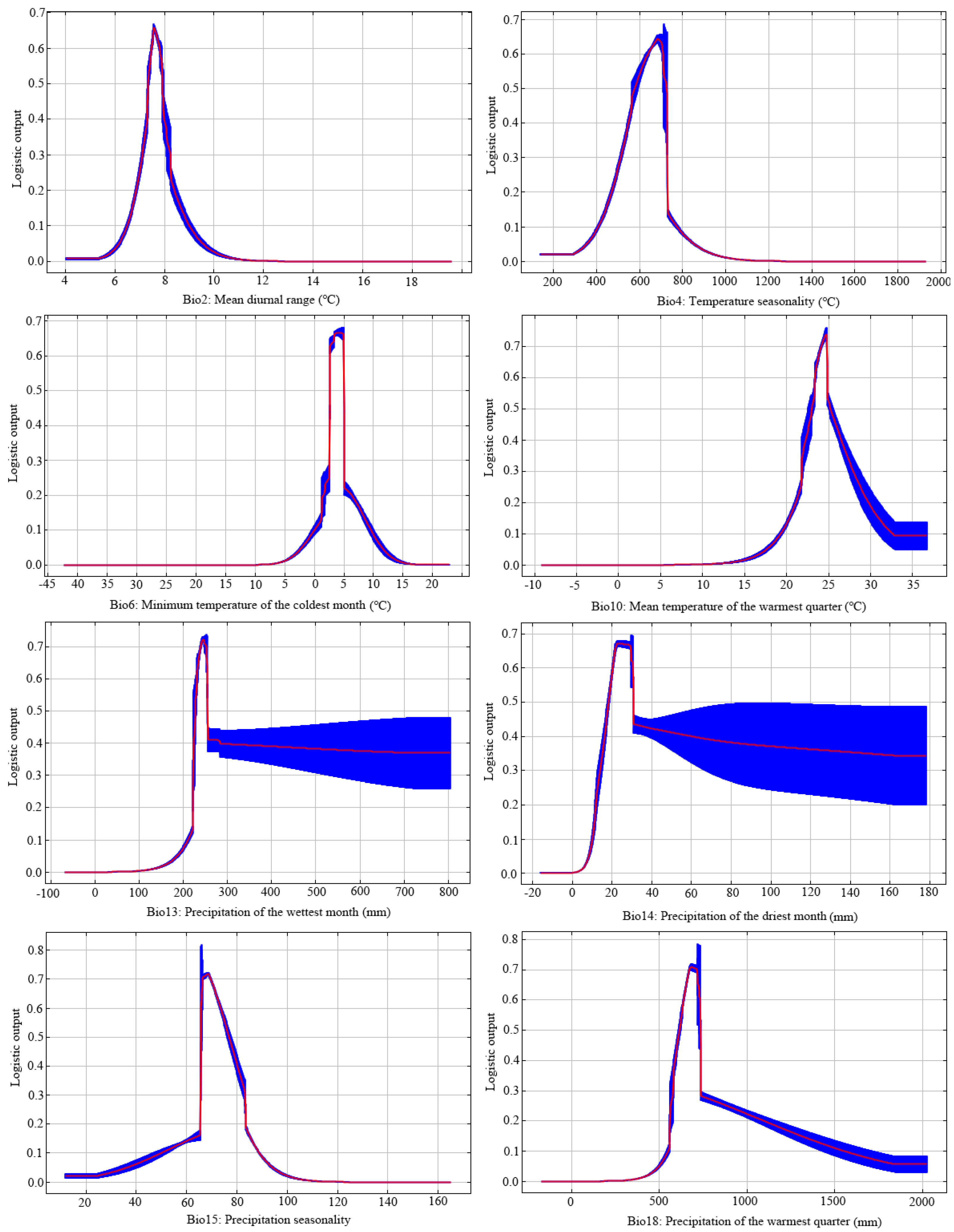
The response curves built by MaxEnt between environmental variables and the probability of species presence indicated the influence of environmental stresses on the target species occurrence (Figure 4). Bio2, Bio4, Bio6, Bio10, Bio15, and Bio18 showed single-peaked curves, indicating that H. bodinieri had significantly adapted to these environmental variables. The response curves of Bio6 (minimum temperature of the coldest month) and Bio18 (precipitation in the wettest quarter) illustrated the effect of changing bioclimatic values on the distribution probability of H. bodinieri (Figure 4). Below −5 °C, the distribution probability was almost zero, and the suitable range of the minimum temperature of the coldest month (Bio6) for H. bodinieri was 2–5 °C. This result provides a strong basis for defining the extreme temperature range of H. bodinieri during winter in China. Moreover, when the precipitation in the wettest quarter (Bio18) was 580–720 mm, the distribution probability of H. bodinieri reached a peak. This result indicates the most suitable rainfall amount and temperature for its survival (Figure 4).
3.3. Predicting the Suitable Habitat of H. Bodinieri in China
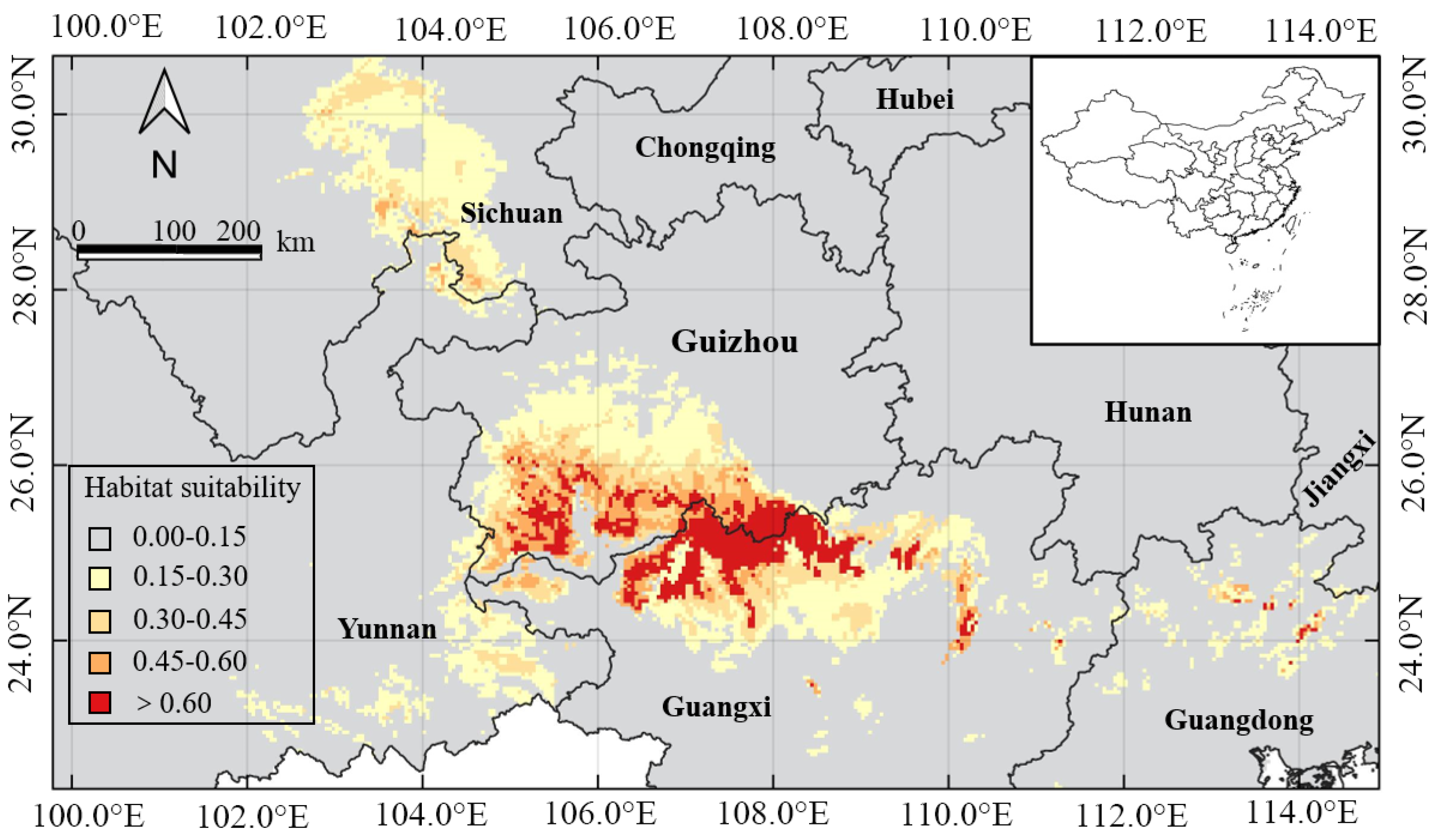
The potential suitable distribution of H. bodinieri predicted by MaxEnt showed that the suitable area for the present climatic conditions is mainly located in the Southwest China provinces of Sichuan, Guizhou, Yunnan, Guangxi, and Guangdong (Figure 5). By habitat suitability, the total areas for the fail, poor, fair, good, and excellent suitability were 116.4 × 104 km2, 8.35 × 104 km2, 3.22 × 104 km2, 1.85 × 104 km2, and 1.93 × 104 km2, respectively, comprising 12.13%, 0.87%, 0.34%, 0.19%, and 0.20% of China’s total land area (960 × 104 km2) (Table 3). Guangxi and Guizhou provinces had relatively large areas with excellent suitability at 1.19 × 104 km2 and 0.71 × 104 km2 (Table 3). Notably, the size of the current potential distribution is significantly larger than the present occurrence of H. bodinieri in China.
4. Discussion
The response relationship between species distribution and environmental factors is one of the important research topics in ecology to predict the species’ geographical distributions and abundance distribution patterns [2,43]. This study was the first to explore the impacts of climate change on the geographical range and environmental suitability of the habitat of H. bodinieri in China using MaxEnt modeling. In this work, 44 valid occurrence records were used to conduct the analysis (Figure 1), which effectively solved the overfitting phenomenon in small samples and ensured the accuracy and stability of the simulation. Eight key environmental factors were chosen based on the contribution rate of each factor to the MaxEnt (Table 2). The final threshold-independent AUC reached 0.99 (Figure 2b), indicating that the MaxEnt predicted a high degree of fit between the habitat range and the actual suitable habitat of H. bodinieri. The results also demonstrated that MaxEnt could avoid overfitting and achieve a good prediction of species distribution areas despite a small sample size. It could provide an effective tool to protect and manage endangered species with an extremely small population [44].
The climate offers the most important factors shaping the physical geography of habitats and species distribution [45]. This main factor influences changes in constituent factors such as annual average temperature, extreme temperature, and annual average precipitation. Such changes can lead to the loss and fragmentation of species’ habitats, seriously threatening endangered species with confined natural distribution and increasing their extinction risk [1,46].
Our results identified Bio18 (precipitation of the warmest quarter) as the main factor molding the potential distribution of H. bodinieri. The results pinpointed 580–720 mm as the most suitable range for precipitation of the warmest quarter (Figure 4), which may be related to the physiological characteristics of H. bodinieri. Based on existing distribution data, this endangered plant is mainly found in karst habitats with deficient surface runoff and sporadic occurrences of small soil pockets interspersed among many rock outcrops [47]. On fine and hot summer days, the strong sunshine vaporizes a large amount of soil water. Therefore, plenty of water is needed in the warm season to meet transpiration consumption and maintain normal physiology in such fragmented and harsh karst habitats.
Moreover, June to August is a critical period for ripening H. bodinieri fruits and seeds [48]. The higher precipitation of the warmest quarter benefits plant growth and development. It can also raise atmospheric humidity and soil moisture content to foster fruit growth and maturation and improve seed quality [49,50]. Proper seed development is critical for the species’ reproductive prospects. If the seeds fail to mature, it will depress the population’s natural regeneration ability, reducing species distribution and diffusion to exacerbate the population’s decline.
Temperature substantially determines plant life activities and biochemical processes and plays an important role in shaping plant distribution [51,52]. Temperature can expand the plant’s adaptability amplitude to habitat conditions and environmental changes within a certain range [37]. However, when a given temperature peak is exceeded, the habitat range shrinks or disappears [53]. The optimum minimum temperature of the coldest month (Bio6) is about 2–5 °C, indicating that the growth and development of H. bodinieri demand a certain heat level. Other studies showed that temperature could significantly favor the dormancy release of seeds, and appropriately low temperatures in the coldest month can break seed dormancy and promote subsequent germination [54]. Nevertheless, too low a temperature can cause injurious freezing, frost damage, and even inactivation of seeds [55], thus trimming the viable seeds for subsequent germination.
Our results predicted the potential distribution area that H. bodinieri may disseminate and spread in China beyond its current range (Figure 1 and Figure 5). The spatial discrepancy between actual and potential ranges might be related to the limited scope of current research on H. bodinieri, focusing on biological characteristics [30,33]. Moreover, research data that accurately describe species’ environmental requirements are scarce. Most individuals of H. bodinieri are distributed in disjointed, harsh, and mountainous karst habitats with largely inaccessible terrain. Therefore, it is difficult to acquire reliable and representative data to assess this endangered species’ actual distribution area accurately. On the other hand, MaxEnt has the inherent trait of evaluating only niche-based species presence data. The model predicts the species’ fundamental niche rather than the realized niche, which may result in overestimation [56,57]. Furthermore, predicting species distribution requires the consideration of physiological constraints, response to external factors and drivers, and competition in ecological communities. As it takes time for such factors to take effect, species’ actual geographical distribution may lag behind climate change [58].
Notwithstanding, this species has thus far failed to colonize a larger extent of its suitable areas. This inability could be explained by extrinsic factors, including soil quality unsuitability, soil spatial discontinuity, interspecific competition, geographical barriers, human disturbance, and other unfavorable or stressful habitat conditions [59]. The lack of sufficient field data was one of the reasons for conducting this study. The results showed that the suitability area of H. bodinieri was only 7 × 104 km2 (Table 3), mainly distributed in the Guangxi and Guizhou provinces, indicating a geographically confined suitability area.
The narrow spatial distribution is also related to the innate biological characteristics of H. bodinieri fruits. The fruits have a relatively large size that is difficult to spread over a long distance by wind. Most fruits randomly fall on microhabitats around the mother tree after ripening, creating a strong density dependence effect. Moreover, the fruits contain a large amount of oil, commonly consumed by rodents and other wildlife after falling to the ground [31]. In addition, the excessive collection of seeds by humans for oil and food has contributed to population decline [30]. The heavy natural seed predation and human harvesting have jointly curtailed the soil seed bank and seeds for subsequent germination. Such constraints demand more stringent requirements to protect and expand the range of H. bodinieri. Other studies have shown that the constricted spatial spread of rare and endangered plants is usually attributed to dispersal limitations and demographic stochasticity [60,61]. The resulting short-range dispersal distance can bring spatial aggregation [61]. Aggregated distribution may lead to patchy and mosaic distribution on the spatial scale and intensify competition among individuals of the species for limited environmental resources [47]. In sum, the combined consequences of the low reproductive survival rate and deleterious human activities have notably shrunk its distribution area and population size.
Our analysis identified suitable habitats for the ex situ conservation of H. bodinieri, providing a critical basis to protect its potential habitat. The distribution area, mainly the karst mountain region, denotes a unique landform that can provide relatively safe refuges for H. bodinieri to adapt more effectively to or survive the intensifying looming climate change. However, the spatial distribution of plants is influenced by abiotic and biotic environments, such as temperature, precipitation, altitude, presence of predators, human disturbance, geographic barriers, soil and vegetation type, etc. [62]. Especially after prolonged human interference, many suitable habitats have been eliminated, accompanied by altered forest land use. Moreover, past herbarium distribution records are not from the same year, and recent surveys are absent. Therefore, it is necessary to investigate the endangered plant’s distribution status in detail and combine different biotic and abiotic factors to predict climate change’s impacts on its distribution [7].
Some measures can be implemented to sustain and expand the range of H. bodinieri. They entail conducting deeper cognate studies to expand the knowledge base and translate research findings to enhance conservation practice. They include crucial issues such as the potential distribution of pollinators and seed dispersers, expanding existing nature reserves and protecting the surrounding habitat of the existing community, research on the impact of climate change on ecological relationships between this endangered plant and the surrounding associated trees [56], establishing new protected areas in high-risk regions in the context of climate change, protecting existing fruiting mother trees, and increasing viable seed sources to foster the reproduction and production of offspring. The areas predicted to be suitable but currently not occupied by the species offer candidate sites for prioritized conservation and propagation. It is necessary to strengthen habitat protection further and raise public awareness. Protection policies can be reinforced to reduce human impacts and implement ex situ protection. More comprehensive and accurate spatial data can be acquired to refine the H. bodinieri habitat distribution simulation.
5. Conclusions
The suitable areas of H. bodinieri in China were predicted by MaxEnt modeling, generating excellent outcomes with high accuracy. The highly suitable habitats for this plant were found primarily in China’s Sichuan, Guizhou, Yunnan, Guangxi, and Guangdong provinces. The key environmental variables regulating its distribution are the precipitation of the warmest quarter (Bio18) and minimum temperature of the coldest month (Bio6), with the optimal conditions at 2–5 °C and 580–720 mm, respectively. These results help us to pinpoint the specific conditions for the optimal growth of the species and provide the scientific basis to improve the management and conservation of this endangered species.
Author Contributions
Data curation, G.W., Z.G. and H.Y.; methodology, C.X.; software, C.X., L.W. and H.Y.; writing—original draft, G.W.; validation, C.X. and C.J.; formal analysis, G.W.; writing—review and editing, C.J. and C.X.; visualization, L.W. and C.X.; investigation, G.W., Z.G. and H.Y. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Guangxi Natural Science Foundation (No.2023GXNSFAA026422); Scientific Research Foundation of Guangxi Normal University for Nationalities (No.2021BS002); Natural Science Foundation of Hainan Province (No. 423MS061), Education Department of Hainan Province (No. Hnky2023ZD-17); National Natural Science Foundation of China (No. 32360417).
Institutional Review Board Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Bellard, C.; Bertelsmeier, C.; Leadley, P.; Thuiller, W.; Courchamp, F. Impacts of climate change on the future of biodiversity. Ecol. Lett. 2012, 15, 365–377. [Google Scholar] [CrossRef] [PubMed]
- Dawson, T.P.; Jackson, S.T.; House, J.I.; Prentice, I.C.; Mace, G.M. Beyond predictions: Biodiversity conservation in a changing climate. Science 2011, 332, 53–58. [Google Scholar] [CrossRef] [PubMed]
- Shrestha, S.; Asch, F.; Brueck, H.; Giese, M.; Dusserre, J.; Ramanantsoanirina, A. Phenological responses of upland rice grown along an altitudinal gradient. Environ. Exp. Bot. 2013, 89, 1–10. [Google Scholar] [CrossRef]
- Graham, E.M.; Reside, A.E.; Atkinson, I.; Baird, D.; Hodgson, L.; James, C.S.; VanDerWal, J.J. Climate change and biodiversity in Australia: A systematic modelling approach to nationwide species distributions. Australas. J. Environ. Manag. 2019, 26, 112–123. [Google Scholar] [CrossRef]
- Sorte, C.J.B.; Ibáñez, I.; Blumenthal, D.M.; Molinari, N.A.; Miller, L.P.; Grosholz, E.D.; Diez, J.M.; D’Antonio, C.M.; Olden, J.D.; Jones, S.J.; et al. Poised to prosper? A cross-system comparison of climate change effects on native and non-native species performance. Ecol. Lett. 2013, 16, 261–270. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.T.; Gliottone, I.; Pham, M.P. Current and future predicting habitat suitability map of Cunninghamia konishii Hayata using MaxEnt model under climate change in Northern Vietnam. Eur. J. Ecol. 2021, 7, 1–17. [Google Scholar] [CrossRef]
- Blach-Overgaard, A.; Svenning, J.-C.; Dransfield, J.; Greve, M.; Balslev, H. Determinants of palm species distributions across Africa: The relative roles of climate, non-climatic environmental factors, and spatial constraints. Ecography 2010, 33, 380–391. [Google Scholar] [CrossRef]
- Ferrarini, A.; Dai, J.; Bai, Y.; Alatalo, J.M. Redefining the climate niche of plant species: A novel approach for realistic predictions of species distribution under climate change. Sci. Total Environ. 2019, 671, 1086–1093. [Google Scholar] [CrossRef]
- Liu, L.; Guan, L.; Zhao, H.; Huang, Y.; Mou, Q.; Liu, K.; Chen, T.; Wang, X.; Zhang, Y.; Wei, B.; et al. Modeling habitat suitability of Houttuynia cordata Thunb (Ceercao) using MaxEnt under climate change in China. Ecol. Inform. 2021, 63, 101324. [Google Scholar] [CrossRef]
- Selwood, K.E.; McGeoch, M.A.; Mac Nally, R. The effects of climate change and land-use change on demographic rates and population viability. Biol. Rev. 2015, 90, 837–853. [Google Scholar] [CrossRef]
- Román-Palacios, C.; Wiens, J.J. Recent responses to climate change reveal the drivers of species extinction and survival. Proc. Natl. Acad. Sci. USA 2020, 117, 4211–4217. [Google Scholar] [CrossRef] [PubMed]
- Wasowicz, P.; Przedpelska-Wasowicz, E.M.; Kristinsson, H. Alien vascular plants in Iceland: Diversity, spatial patterns, temporal trends, and the impact of climate change. Flora Morphol. Distrib. Funct. Ecol. Plants 2013, 208, 648–673. [Google Scholar] [CrossRef]
- Lippmann, R.; Babben, S.; Menger, A.; Delker, C.; Quint, M. Development of wild and cultivated plants under global warming conditions. Curr. Biol. 2019, 29, 1326–1338. [Google Scholar] [CrossRef]
- Yan, G.; Zhang, G.F. Predicting the potential distribution of endangered Parrotia subaequalis in China. Forests 2022, 13, 1595. [Google Scholar] [CrossRef]
- Abdelaal, M.; Fois, M.; Fenu, G.; Bacchetta, G. Using MaxEnt modeling to predict the potential distribution of the endemic plant Rosa arabica Crép. in Egypt. Ecol. Inform. 2019, 50, 68–75. [Google Scholar] [CrossRef]
- Kamyo, T.; Asanok, L. Modeling habitat suitability of Dipterocarpus alatus (Dipterocarpaceae) using MaxEnt along the Chao Phraya River in Central Thailand. For. Sci. Technol. 2020, 16, 1–7. [Google Scholar] [CrossRef]
- Soilhi, Z.; Sayari, N.; Benalouache, N.; Mekki, M. Predicting current and future distributions of Mentha pulegium L. in Tunisia under climate change conditions, using the MaxEnt model. Ecol. Inform. 2022, 68, 101533. [Google Scholar] [CrossRef]
- Tran, V.D.; Vu, T.T.; Tran, Q.B.; Nguyen, T.H.; Ta, T.N.; Ha, T.M.; Nguyen, H.V. Predicting suitable distribution for an endemic, rare and threatened species (Grey-shanked douc langur, Pygathrix cinerea Nadler, 1997) using MaxEnt model. Appl. Ecol. Environ. Res. 2018, 16, 1275–1291. [Google Scholar] [CrossRef]
- Bentlage, B.; Peterson, A.T.; Barve, N.; Cartwright, P. Plumbing the depths: Extending ecological niche modelling and species distribution modelling in three dimensions. Glob. Ecol. Biogeogr. 2013, 22, 952–961. [Google Scholar] [CrossRef]
- Mousazade, M.; Ghanbarian, G.; Pourghasemi, H.R.; Safaeian, R.; Cerdà, A. Maxent Data Mining Technique and its comparison with a bivariate statistical model for predicting the potential distribution of Astragalus Fasciculifolius Boiss. in Fars, Iran. Sustainability 2019, 11, 3452. [Google Scholar] [CrossRef]
- Çoban, H.O.; Örücü, Ö.K.; Arslan, E.S. MaxEnt modeling for predicting the current and future potential geographical distribution of Quercus libani Olivier. Sustainability 2020, 12, 2671. [Google Scholar] [CrossRef]
- Morales, N.S.; Fernández, I.C.; Baca-González, V. MaxEnt’s parameter configuration and small samples: Are we paying attention to recommendations? A systematic review. PeerJ 2017, 5, e3093. [Google Scholar] [CrossRef] [PubMed]
- Pearson, R.G.; Raxworthy, C.J.; Nakamura, M.; Townsend Peterson, A. ORIGINAL ARTICLE: Predicting species distributions from small numbers of occurrence records: A test case using cryptic geckos in Madagascar. J. Biogeogr. 2007, 34, 102–117. [Google Scholar] [CrossRef]
- Anand, V.; Oinam, B.; Singh, I.H. Predicting the current and future potential spatial distribution of endangered Rucervus eldii eldii (Sangai) using MaxEnt model. Environ. Monit. Assess. 2021, 193, 147. [Google Scholar] [CrossRef] [PubMed]
- Kramer-Schadt, S.; Niedballa, J.; Pilgrim, J.D.; Schröder, B.; Lindenborn, J.; Reinfelder, V.; Stillfried, M.; Heckmann, I.; Scharf, A.K.; Augeri, D.M.; et al. The importance of correcting for sampling bias in MaxEnt species distribution models. Divers. Distrib. 2013, 19, 1366–1379. [Google Scholar] [CrossRef]
- Warren, D.L.; Seifert, S.N. Ecological niche modeling in Maxent: The importance of model complexity and the performance of model selection criteria. Ecol. Appl. 2011, 21, 335–342. [Google Scholar] [CrossRef]
- Cotrina-Sánchez, A.; Rojas Briceño, N.B.; Bandopadhyay, S.; Ghosh, S.; Torres Guzmán, C.; Oliva, M.; Guzman, B.K.; Salas López, R. Biogeographic distribution of Cedrela spp. genus in Peru using MaxEnt modeling: A conservation and restoration approach. Diversity 2021, 13, 261. [Google Scholar] [CrossRef]
- Dad, J.M.; Rashid, I. Differential responses of Kashmir Himalayan threatened medicinal plants to anticipated climate change. Environ. Conserv. 2022, 49, 33–41. [Google Scholar] [CrossRef]
- Cao, L.M.; Xia, N.H.; Deng, Y.F. Embryology of Handeliodendron bodinieri (Sapindaceae) and its systematic value: Development of male and female gametophytes. Plant Syst. Evol. 2008, 274, 17–23. [Google Scholar] [CrossRef]
- He, R.; Wang, J.; Huang, H. Long-distance gene dispersal inferred from spatial genetic structure in Handeliodendron bodinieri, an endangered tree from karst forest in southwest China. Biochem. Syst. Ecol. 2012, 44, 295–302. [Google Scholar] [CrossRef]
- Xiong, Z.; Ran, J.; Tan, C.; Yu, P.; Qin, H.; Wei, J. The seed ecological characteristics of endangered Handeliodendron bod inierei. Acta Ecol. Sin. 2003, 23, 820–825. [Google Scholar]
- Huang, S.; Luo, W. Biological behaviors and conservation of Handeliodendron Bodinieri. J. Ecol. Rural Environ. 2001, 17, 21–22. [Google Scholar] [CrossRef]
- Xie, C.; Li, X.; Guo, S.; Jiang, M.; Li, Z. Research trends of the rare and endangered plant Handeliodendron bodinierei. Mol. Plant Breed. 2020, 18, 1725–1730. [Google Scholar] [CrossRef]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Puchałka, R.; Dyderski, M.K.; Vítková, M.; Sádlo, J.; Klisz, M.; Netsvetov, M.; Prokopuk, Y.; Matisons, R.; Mionskowski, M.; Wojda, T.; et al. Black locust (Robinia pseudoacacia L.) range contraction and expansion in Europe under changing climate. Glob. Chang. Biol. 2021, 27, 1587–1600. [Google Scholar] [CrossRef]
- Liu, D.W.; Xie, C.P.; Jim, C.Y.; Liu, Y.J.; Hou, S.L. Predicting the Potential Distribution of the alien invasive alligator gar Atractosteus spatula in China. Sustainability 2023, 15, 6419. [Google Scholar] [CrossRef]
- Xu, D.P.; Zhuo, Z.H.; Wang, R.L.; Ye, M.; Pu, B. Modeling the distribution of Zanthoxylum armatum in China with MaxEnt modeling. Glob. Ecol. Conserv. 2019, 19, e00691. [Google Scholar] [CrossRef]
- Xie, C.P.; Huang, B.Y.; Jim, C.Y.; Han, W.D.; Liu, D.W. Predicting differential habitat suitability of Rhodomyrtus tomentosa under current and future climate scenarios in China. For. Ecol. Manag. 2021, 501, 119696. [Google Scholar] [CrossRef]
- Muscarella, R.; Galante, P.J.; Soley-Guardia, M.; Boria, R.A.; Kass, J.M.; Uriarte, M.; Anderson, R.P. ENMeval: An R package for conducting spatially independent evaluations and estimating optimal model complexity for MaxEnt ecological niche models. Methods Ecol. Evol. 2014, 11, 1198–1205. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef]
- Sarma, K.; Roy, S.J.; Kalita, B.; Baruah, P.S.; Bawri, A.; Nath, M.J.; Baruah, U.D.; Sahariah, D.; Saikia, A.; Tanti, B. Habitat suitability of Gymnocladus assamicus-A critically endangered plant of Arunachal Pradesh, India using machine learning and statistical modeling. Acta Ecol. Sin. 2022, 42, 398–406. [Google Scholar] [CrossRef]
- Beale, C.M.; Lennon, J.J. Incorporating uncertainty in predictive species distribution modelling. Philos. Trans. R. Soc. B Biol. Sci. 2012, 367, 247–258. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Li, L.; Wu, G.; Yuan, L.; Meng, C.; Guo, H.; Zhou, Y.; Ma, C.; Gao, Y.; Xue, Y.; et al. Habitat suitability assessment of Panthera uncia in Qilian Mountains of Qinghai based on MaxEnt modeling. Acta Ecol. Sin. 2023, 43, 2202–2209. [Google Scholar] [CrossRef]
- Gao, B.; Wei, H.; Guo, Y.; Gu, W. Using GIS and MaxEnt to analyze the potential distribution of Abies chensiensis. Chin. J. Ecol. 2015, 34, 843–852. [Google Scholar] [CrossRef]
- Remya, K.; Ramachandran, A.; Jayakumar, S. Predicting the current and future suitable habitat distribution of Myristica dactyloides Gaertn. using MaxEnt model in the Eastern Ghats, India. Ecol. Eng. 2015, 82, 184–188. [Google Scholar] [CrossRef]
- Veloz, S.D.; Williams, J.W.; Blois, J.L.; He, F.; Otto-Bliesner, B.; Liu, Z. No-analog climates and shifting realized niches during the late quaternary: Implications for 21st-century predictions by species distribution models. Glob. Change Biol. 2012, 18, 1698–1713. [Google Scholar] [CrossRef]
- Wang, G.H.; Pan, Y.; Qin, G.L.; Tan, W.N.; Lu, C.H. Population structure and spatial distribution pattern of Kmeria septentrionalis an endangered species in karst habitat. For. Res. 2021, 34, 81–87. [Google Scholar] [CrossRef]
- Gao, J.W.; Xie, G.W.; Lin, Z.G.; He, J.L.; Wang, Y.L.; Qiu, C.M.; Huang, J.J. Population dynamics of rare and endangered plant Handeliodendron bodinieri (Levl.) Rehd in Leye County of Guangxi region. Guangdong Agric. Sci. 2015, 18, 43–48. [Google Scholar] [CrossRef]
- Lan, X.; Wang, J.; Fu, C.; Li, L.; Yuan, M.; Tan, T.; Du, F. Prediction of suitable distribution area of Magnolia sieboldii in China based on the optimized MaxEnt model. J. Northwest For. Univ. 2022, 37, 100–106. [Google Scholar] [CrossRef]
- Leng, X.H.; Xue, L.; Wang, J.; Li, S.; Yang, Z.L.; Ren, H.D.; Yao, X.H.; Wu, Z.Y.; Li, J.Y. Physiological Responses of Handeliodendron bodinieri(Levl.) Rehd. to Exogenous Calcium Supply under Drought Stress. Forests 2020, 11, 69. [Google Scholar] [CrossRef]
- Khanum, R.; Mumtaz, A.S.; Kumar, S. Predicting impacts of climate change on medicinal asclepiads of Pakistan using Maxent modeling. Acta Oecologica 2013, 49, 23–31. [Google Scholar] [CrossRef]
- Wang, D.; Zhao, W.; Cheng, B.; Zhang, J. Potential Suitable Areas of Crataegus pinnatifida in China based on MaxEnt Modeling. Sci. Silvae Sin. 2022, 58, 43–50. [Google Scholar] [CrossRef]
- Khodorova, N.V.; Boitel-Conti, M. The Role of Temperature in the Growth and Flowering of Geophytes. Plants 2013, 2, 699–711. [Google Scholar] [CrossRef]
- Atkinson, C.J.; Brennan, R.M.; Jones, H.G. Declining chilling and its impact on temperate perennial crops. Environ. Exp. Bot. 2013, 91, 48–62. [Google Scholar] [CrossRef]
- Rupa, D.; Saikat, B. Influence of abiotic stresses on seed production and quality. In Seed Biology Updates; Jimenez-Lopez, J.C., Ed.; IntechOpen: Rijeka, Croatia, 2022. [Google Scholar] [CrossRef]
- Zhou, Y.R.; Lu, X.; Zhang, G.F. Potentially differential impacts on niche overlap between Chinese endangered Zelkova schneideriana and its associated tree species under climate change. Front. Ecol. Evol. 2023, 11, 1218149. [Google Scholar] [CrossRef]
- Yang, X.Q.; Kushwaha, S.P.S.; Saran, S.; Xu, J.C. Maxent modeling for predicting the potential distribution of medicinal plant, Justicia adhatoda L. in Lesser Himalayan foothills. Ecol. Eng. 2013, 51, 83–87. [Google Scholar] [CrossRef]
- Zhang, L.; Jing, Z.N.; Li, Z.Y.; Liu, Y.; Fang, S.Z. Predictive modeling of suitable habitats for Cinnamomum camphora (L.) presl using maxent model under climate change in China. Int. J. Environ. Res. Public Health 2019, 16, 3185. [Google Scholar] [CrossRef]
- Wang, L.; Yilihamu, Y. Prediction of potential suitable areas for a Chinese endemic shrub Sophora davidii using the MaxEnt Model. Chin. J. Ecol. 2021, 40, 3114–3124. [Google Scholar] [CrossRef]
- Aiba, M.; Takafumi, H.; Hiura, T. Interspecific differences in determinants of plant species distribution and the relationships with functional traits. J. Ecol. 2012, 100, 950–957. [Google Scholar] [CrossRef]
- Clark, A.T.; Detto, M.; Muller-Landau, H.C.; Schnitzer, S.A.; Wright, S.J.; Condit, R.; Hubbell, S.P. Functional traits of tropical trees and lianas explain spatial structure across multiple scales. J. Ecol. 2018, 106, 795–806. [Google Scholar] [CrossRef]
- Asanok, L.; Kamyo, T.; Marod, D. Maximum entropy modeling for the conservation of Hopea odorata in riparian forests, central Thailand. Biodiversitas 2020, 21, 4663–4670. [Google Scholar] [CrossRef]
Figure 1.
The images of H. bodinieri and its actual distribution area (white circles) in two provinces of Southwest China. The pictures on the left side show the flowers, fruits, and complete plants of the H. bodinieri from top to bottom.
Figure 1.
The images of H. bodinieri and its actual distribution area (white circles) in two provinces of Southwest China. The pictures on the left side show the flowers, fruits, and complete plants of the H. bodinieri from top to bottom.

Figure 2.
The validation of the model predicting H. bodinieri distribution: (a) omission rate; and (b) ROC curve.
Figure 2.
The validation of the model predicting H. bodinieri distribution: (a) omission rate; and (b) ROC curve.

Figure 3.
The relative predictive powers of the eight environmental variables based on the jackknife test of regularized training gain, test gain, and area under the curve (AUC) in MaxEnt.
Figure 3.
The relative predictive powers of the eight environmental variables based on the jackknife test of regularized training gain, test gain, and area under the curve (AUC) in MaxEnt.

Figure 4.
Response curves of the environmental variables to distribution probability.

Figure 5.
The predicted H. bodinieri distribution range obtained by MaxEnt modeling.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
List of environmental variables considered in the course of model development. The eight variables with the code in bold font were chosen for the MaxEnt modeling study.
Table 1.
List of environmental variables considered in the course of model development. The eight variables with the code in bold font were chosen for the MaxEnt modeling study.
| Code | Environmental Variable | Unit |
|---|---|---|
| Bio1 | Annual mean temperature | °C |
| Bio2 | Mean diurnal range (mean of monthly (maximum temp-minimum temp)) | °C |
| Bio3 | Isothermality (Bio2/Bio7) (×100) | - |
| Bio4 | Temperature seasonality (standard deviation×100) | °C |
| Bio5 | Maximum temperature of the warmest month | °C |
| Bio6 | Minimum temperature of the coldest month | °C |
| Bio7 | Temperature annual range (Bio5-Bio6) | °C |
| Bio8 | Mean temperature of the wettest quarter | °C |
| Bio9 | Mean temperature of the driest quarter | °C |
| Bio10 | Mean temperature of the warmest quarter | °C |
| Bio11 | Mean temperature of the coldest quarter | °C |
| Bio12 | Annual precipitation | mm |
| Bio13 | Precipitation of the wettest month | mm |
| Bio14 | Precipitation of the driest month | mm |
| Bio15 | Precipitation seasonality (coefficient of variation) | - |
| Bio16 | Precipitation of the wettest quarter | mm |
| Bio17 | Precipitation of the driest quarter | mm |
| Bio18 | Precipitation of the warmest quarter | mm |
| Bio19 | Precipitation of the coldest quarter | mm |
Table 2.
Percent contribution and permutation importance levels of the eight environmental variables included in the MaxEnt, ranked by percentage contribution.
Table 2.
Percent contribution and permutation importance levels of the eight environmental variables included in the MaxEnt, ranked by percentage contribution.
| Code | Bioclimatic Variable | Percent Contribution | Permutation Importance |
|---|---|---|---|
| Bio18 | Precipitation of the warmest quarter | 32.7 | 6.9 |
| Bio6 | Minimum temperature of the coldest month | 26.3 | 75.7 |
| Bio2 | Mean diurnal range | 20.5 | 13 |
| Bio15 | Precipitation seasonality | 8.4 | 2.1 |
| Bio4 | Temperature seasonality | 7.7 | 0.3 |
| Bio14 | Precipitation of the driest month | 3.1 | 1.3 |
| Bio13 | Precipitation of the wettest month | 0.9 | 0.4 |
| Bio10 | Mean temperature of the warmest quarter | 0.4 | 0.2 |
Table 3.
Predicted suitable areas for H. bodinieri under the current climate in various provinces or autonomous regions (104 km2). The ratio denotes the predicted suitable area divided by the total land area of the respective province or autonomous.
Table 3.
Predicted suitable areas for H. bodinieri under the current climate in various provinces or autonomous regions (104 km2). The ratio denotes the predicted suitable area divided by the total land area of the respective province or autonomous.
| Province or Autonomous Region | Predicted Suitable Area Ratio | ||||
|---|---|---|---|---|---|
| Fail | Poor | Fair | Good | Excellent | |
| Guangxi | 15.87 | 2.10 | 1.15 | 0.50 | 1.19 |
| Guizhou | 10.99 | 1.92 | 1.11 | 1.22 | 0.71 |
| Guangdong | 14.72 | 0.64 | 0.06 | 0.05 | 0.03 |
| Sichuan | 42.42 | 2.29 | 0.63 | 0.06 | 0.00 |
| Yunnan | 32.40 | 1.40 | 0.27 | 0.02 | 0.00 |
| Total | 116.4 | 8.35 | 3.22 | 1.85 | 1.93 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Wang, G.; Xie, C.; Wei, L.; Gao, Z.; Yang, H.; Jim, C. Predicting Suitable Habitats for China’s Endangered Plant Handeliodendron bodinieri (H. Lév.) Rehder. Diversity 2023, 15, 1033. https://doi.org/10.3390/d15101033
AMA Style
Wang G, Xie C, Wei L, Gao Z, Yang H, Jim C. Predicting Suitable Habitats for China’s Endangered Plant Handeliodendron bodinieri (H. Lév.) Rehder. Diversity. 2023; 15(10):1033. https://doi.org/10.3390/d15101033
Chicago/Turabian StyleWang, Guohai, Chunping Xie, Lijuan Wei, Zequn Gao, Honglan Yang, and Chiyung Jim. 2023. "Predicting Suitable Habitats for China’s Endangered Plant Handeliodendron bodinieri (H. Lév.) Rehder" Diversity 15, no. 10: 1033. https://doi.org/10.3390/d15101033
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.