
Mercury Biogeochemistry and Biomagnification in the Mediterranean Sea: Current Knowledge and Future Prospects in the Context of Climate Change

Department of Physics, Earth and Environmental Sciences, University of Siena, Via P. A. Mattioli, 4, IT-53100 Siena, Italy
*
Author to whom correspondence should be addressed.
Coasts 2024, 4(1), 89-107; https://doi.org/10.3390/coasts4010006
Submission received: 12 December 2023
/
Revised: 17 January 2024
/
Accepted: 1 February 2024
/
Published: 4 February 2024
Abstract
:In the 1970s, the discovery of much higher mercury (Hg) concentrations in Mediterranean fish than in related species of the same size from the Atlantic Ocean raised serious concerns about the possible health effects of neurotoxic monomethylmercury (MMHg) on end consumers. After 50 years, the cycling and fluxes of the different chemical forms of the metal between air, land, and marine environments are still not well defined. However, current knowledge indicates that the anomalous Hg accumulation in Mediterranean organisms is mainly due to the re-mineralization of organic material, which favors the activity of methylating microorganisms and increases MMHg concentrations in low-oxygen waters. The compound is efficiently bio-concentrated by very small phytoplankton cells, which develop in Mediterranean oligotrophic and phosphorous-limited waters and are then transferred to grazing zooplankton. The enhanced bioavailability of MMHg together with the slow growth of organisms and more complex and longer Mediterranean food webs could be responsible for its anomalous accumulation in tuna and other long-lived predatory species. The Mediterranean Sea is a “hotspot” of climate change and has a rich biodiversity, and the increasing temperature, salinity, acidification, and stratification of seawater will likely reduce primary production and change the composition of plankton communities. These changes will likely affect the accumulation of MMHg at lower trophic levels and the biomagnification of its concentrations along the food web; however, changes are difficult to predict. The increased evasion of gaseous elemental mercury (Hg°) from warming surface waters and lower primary productivity could decrease the Hg availability for biotic (and possibly abiotic) methylation processes, but lower oxygen concentrations in deep waters, more complex food webs, and the reduced growth of top predators could increase their MMHg content. Despite uncertainties, in Mediterranean regions historically affected by Hg inputs from anthropogenic and geogenic sources, such as those in the northwestern Mediterranean and the northern Adriatic Sea, rising seawater levels, river flooding, and storms will likely favor the mobilization of Hg and organic matter and will likely maintain high Hg bioaccumulation rates for a long time. Long-term studies will, therefore, be necessary to evaluate the impact of climate change on continental Hg inputs in the Mediterranean basin, on air–sea exchanges, on possible changes in the composition of biotic communities, and on MMHg formation and its biomagnification along food webs. In this context, to safeguard the health of heavy consumers of local seafood, it appears necessary to develop information campaigns, promote initiatives for the consumption of marine organisms at lower trophic levels, and organize large-scale surveys of Hg accumulation in the hair or urine of the most exposed population groups.
1. Introduction
The poisoning incidents that occurred in Minamata (Japan) in the late 1950s highlighted how the consumption of seafood containing high concentrations of the potent neurotoxic monomethylmercury (MMHg) released by industrial plants constituted a serious threat to human health. Global awareness of environmental and health risks increased with the discovery [1] that, in all aquatic ecosystems, inorganic Hg can be transformed via natural processes into the easily bioaccumulative MMHg. It soon emerged that total Hg concentrations in Mediterranean fish were much higher than those found in specimens of the same species and size caught in the Atlantic Ocean [2,3,4,5]. Methylmercury has a remarkably long biological half-life (a few months instead of a few days or weeks for inorganic Hg) and is, therefore, biomagnified in the long-lived carnivorous species of marine food webs. Thus, in subsequent years, it was discovered that MMHg accumulation in Mediterranean fish could be a potential health risk for apex consumers such as seabirds, marine mammals, and some groups of fishermen and their families (e.g., [6,7,8,9]).
To address the accelerating degradation of the world’s oceans and coastal areas, in 1972, the United Nations Conference on the Human Environment launched the Regional Seas Programme, and in 1976, Mediterranean governments and the European Community adopted the Barcelona Convention for the Protection of the Mediterranean Sea Against Pollution. The Mediterranean Action Plan (MAP) was the first in a series of regional initiatives aimed at controlling environmental pollution caused by Hg and persistent organic compounds and promoting the sustainable use of marine and coastal resources [10]. In addition, in 2017, the Minamata Convention on Mercury [11] entered into force, giving further impetus to global Hg monitoring and policy efforts for the protection of human health and the environment. Nonetheless, recent investigations on the distribution, biogeochemical cycle, and possible biological effects of Hg in the Mediterranean Sea (e.g., [12,13,14,15]) attest, overall, that Hg concentrations in several predatory fish still exceed European Union regulatory thresholds, and in coastal human populations, they are higher than in people living elsewhere in Europe or on other continents [16]. In western Mediterranean birds, such as the European shag, northern gannet, and great cormorant, Hg concentrations are still close to or higher than those that cause reproductive alterations in other bird species [17].
Despite significant scientific advances in this metal’s global biogeochemistry (e.g., [18,19,20]), knowledge gaps remain about Hg fluxes between air, land, the anthroposphere, and the marine environment, and the causes of its anomalous bioaccumulation in Mediterranean organisms are not yet fully understood. Moreover, it seems difficult to foresee the future because the physicochemical and bioecological processes underlying the cycling of Hg and its bioaccumulation are influenced by climate change.
Total Hg concentrations in aquatic ecosystems are largely controlled by air–water exchanges, and declines in anthropogenic emissions coupled with the increased evasion of elemental Hg (Hg°) from warming surface waters will likely reduce the metal bioavailability for marine organisms. In the Atlantic bluefin tuna, for instance, Hg bioaccumulation was found to decrease between 2004 and 2012, consistent with reductions in anthropogenic Hg emissions in North America and atmospheric Hg° concentrations in the North Atlantic in the same period [21]. However, decreasing trends in atmospheric Hg concentrations are not always reflected by corresponding trends in Hg concentrations in aquatic organisms (e.g., [22,23]), and in some regions, local aquatic and terrestrial processes may be more important than atmospheric deposition as drivers of Hg bioaccumulation [24]. In the Mediterranean Sea, for instance, Hg bioavailability can be supported by geogenic inputs such as land-based Hg-mineralizations or submarine volcanoes, hydrothermal vents, and cold seeps or by the mobilization of legacy metal released by past human activities. In hydrothermal vents near the island of Panarea (Aeolian Arc), Bagnato et al. [25] measured total Hg concentrations three orders of magnitude higher than in the overlaying seawater column, and Andaloro et al. [26] attributed the metal accumulation in sediments and organisms of the southern Tyrrhenian Sea (the deepest region of the western Mediterranean basin) to submarine volcanic emissions. In addition, the Mediterranean region is crossed by the Iberian Hg belt, which extends from Almaden (Spain) to Monte Amiata (central Italy), Idrija (Slovenia), Karaburun and other locations of Aegean Turkey, and the Numidian belt (Algeria). Past mining and smelting activities and the natural weathering processes of mineralized areas and mine dumps have mobilized large amounts of Hg, which are now buried in riverine and marine sediments of the northern Tyrrhenian and Adriatic seas [27,28,29] and the Aegean shelf [30].
The Mediterranean is a semi-enclosed basin with a low rate of water renewal, and many coastal marine ecosystems are also polluted by past and current anthropogenic Hg inputs via industrial, agricultural, and urban wastewater (e.g., [27,31,32,33]). Moreover, given the recent warming (20% faster than the global average) and changing patterns of atmospheric precipitation (a decrease in mean precipitation and an increase in precipitation variability), the Mediterranean region is considered a climate change hotspot, that is, one of the most responsive regions to global change ([34] and refs therein). Although the effects of such changes, as well as those of water salinity and pH, on marine biogeochemistry are poorly understood, it is likely that they are modifying the cycling of Hg [20]. The rise in sea levels (3.6 ± 0.3 mm year−1 in the period 2000–2018; [35]), together with the subsiding of various coastal environments such as the Po Delta and the growing intensity of waves and storms, is making the flooding and erosion of sandy coasts and river estuaries more severe, favoring the mobilization of formerly deposited Hg. The acidification of oligotrophic Mediterranean waters could enhance the already high rate of Hg methylation and promote the growth of small phytoplankton species that accumulate Hg2+ and MMHg more efficiently than larger species [36]. In the Mediterranean Sea, water temperatures have risen by about 1.5 °C since the end of the 19th century compared with the global average of 1.1 °C [37], and further warming will favor the entry of new alien species from the Red Sea and the geographic expansion of those already established in the basin [38]. Human activities such as overfishing, the accidental or deliberate introduction of exotic species, and waste dumping will likely contribute to changing the composition of biotic communities and the length of food webs with possible significant effects on the biomagnification of MMHg.
This review summarizes recent progress in understanding the geochemical, biological, and ecological processes responsible for the so-called “Mediterranean Hg anomaly”. Peer-reviewed articles investigating Hg and the impact of climate change on the marine environment were selected from those listed in Scopus (https://www.scopus.com/) and other databases, referring mainly to the most recent ones dealing with the Mediterranean Sea region. In discussing the possible impacts of climate change on the biogeochemical cycle of Hg in the Mediterranean and its enhanced biomagnification in marine food webs, the potential health risks for the most exposed population groups will also be taken into consideration. Critical themes and unresolved questions will be highlighted to guide future research with particular reference to the following:
- Hg inputs into the Mediterranean from widespread geochemical, volcanic, geothermal, and anthropogenic sources and an evaluation of air–sea exchanges;
- The impact of the warming and acidification of seawater on the biogeochemical cycle of Hg;
- Possible changes in the composition of biotic communities and food webs and in bioecological processes involved in MMHg biomagnification.
2. The Unique Features of the Mediterranean Sea
The increased bioaccumulation of Hg in Mediterranean organisms is inextricably linked to the specific physicochemical and bioecological characteristics of this semi-enclosed basin, which has been impacted by human activities since ancient times.
Given its complex geological history and past connections with adjacent cold and/or warm seas, the Mediterranean Sea harbors a rich biodiversity with over 17,000 species, approximately one-fifth of which are endemic, with both temperate and subtropical elements [39]. The establishment of species from other seas has been favored by the variety of marine habitats resulting from the strong thermal gradient, wind-driven currents, atmospheric precipitations, and the terrestrial nutrient supply from around the northwestern and the southeastern basins. However, over the few last decades, the accidental and intentional introductions of many exotic species have endangered local endemic communities and become a serious threat to the integrity and functioning of Mediterranean ecosystems.
For many decades after the opening of the Suez Canal in 1869, the hypersaline waters of the Bitter Lakes prevented the migration of organisms from the Red Sea into the Mediterranean, but over time, the salinity of the lakes decreased and no longer constituted a barrier. Moreover, the construction of the Aswan High Dam on the Nile River in the 1960s reduced the inflow of freshwater and nutrients and increased the salinity and oligotrophy of the eastern Mediterranean, generating environmental conditions similar to those of the Red Sea. In recent decades, biological invasions have been further facilitated by the rapid warming of the Mediterranean Sea, the deepening and widening of the Suez Canal, aquaculture facilities, the aquarium trade, ballast waters, and fouling organisms from the numerous vessels operating in the basin. Currently, around 1000 alien species have been identified in the Mediterranean Sea [38].
On a global scale, coastal waters occupy 0.2% of the ocean volume; nonetheless, they receive about one-third of the entire Hg input into the oceans [40], and in the Mediterranean basin, this ratio is probably still greater. Although the Mediterranean Sea constitutes less than 1% of the world’s oceans, more than 50% of global cinnabar (HgS) deposits are concentrated in its region, many of which have been intensely exploited in the past. Moreover, a number of potential submarine sources of Hg have been reported in volcanic-associated ecosystems in the Aegean and Aeolian Arcs and in the Gulf of Naples. According to a recent database [41], there are at least 156 such ecosystems in 21 different regions of the Mediterranean basin, from 1 to 3800 m in depth. Carbonate formations and karst phenomena are widespread along the Mediterranean coasts, and many aquifers are directly connected to the sea, sometimes at great depths [42]. These aquifers, although little or not studied at all, are likely to represent additional sources of bioavailable Hg for marine organisms.
Being almost landlocked, the Mediterranean Sea has very limited tides and, especially in the eastern basin, water evaporation far exceeds the contributions derived from atmospheric precipitation and river inputs. The basin receives surface waters from the Atlantic Ocean, which, as they move eastward, become warmer and saltier and sink to intermediate depths, generating the Levantine Intermediate Water. The latter moves westward and, after a residence time of about 100 years, returns to the Atlantic, exporting nutrients and contributing to the Mediterranean oligotrophy. Moreover, Mediterranean waters have a higher nitrogen/phosphorous (N/P) ratio than the global ocean, and the P shortage limits primary productivity [43]. In nutrient-poor environments, such as the ultra-oligotrophic waters of the Levantine Basin, most chlorophyll is concentrated in rather deep waters, and autotrophic communities are dominated by cyanobacteria and very small algal cells (pico-phytoplankton) [44].
Given the shallow depth of the Strait of Gibraltar, cold deep waters from the Atlantic cannot enter the Mediterranean Sea, and the seawater temperature is always >13 °C, even at depths >5000 m. This thermal “anomaly” strongly influences thermohaline circulation and the physicochemical and bioecological processes involved in the biogeochemical cycle of Hg. The possible effects of climate change on the primary productivity of the Mediterranean Sea have been little studied and are poorly understood [34]. Surface temperature trends indicate general warming during the last few decades and, together with the likely decrease in atmospheric precipitation, will probably make seawater warmer, saltier, and more acidic, further reducing vertical mixing and the availability of oxygen and nutrients for organisms [45]. These changes will push several endemic species beyond their tolerance thresholds and, together with the establishment of invasive alien species, could modify the composition of biotic communities.
3. Sources and Cycling of Hg in the Mediterranean Sea
Mercury released from anthropogenic sources such as incinerators, fossil fuel combustion, or cement production enters seawater primarily through atmospheric precipitation and the dry deposition of gaseous and particulate Hg aerosols [46]. Although still poorly constrained, in the Mediterranean region, significant amounts of atmospheric Hg are released by forest fires, active volcanoes, fumaroles, solfataras, and geothermal fields. The contribution of these natural sources in southern and central Italy has been highlighted since the 1990s (e.g., [47,48,49,50]). Depending on the season and meteorological conditions, a significant proportion of atmospheric Hg also comes from distant sources [15]. Estimated modeled fluxes of dry and wet Hg depositions are about 19.6 and 18.1 Mg year−1, respectively, and anthropogenic emissions likely constitute 20–25% [51,52]. Interestingly, although the AMAP/UNEP Hg emission inventories for 2005 and 2010 reported a 33% decrease in anthropogenic Hg emissions, in the abovementioned studies, the simulated deposition of Hg in the Mediterranean Sea decreased over the same period by only 12%.
Total Hg concentrations in the open waters of the Mediterranean are in the same range or slightly higher than those measured in the Atlantic and Pacific Oceans, while in samples from the continental shelf of the northwestern basin and the northern Adriatic Sea, the values are significantly higher (Table 1). Hg contamination of coastal ecosystems is due to local anthropogenic sources and, in the northern Tyrrhenian and Adriatic seas, also to the inputs from rivers draining cinnabar-mineralized areas and abandoned mining and smelting dumps (e.g., [28,30,53,54,55]). In the Gulf of Trieste, for example, the inputs of particulate and dissolved Hg from the Soča/Isonzo River were estimated to be equal to 1500 and 8.6 kg year−1, respectively [56]. More than 2000 tons of metal were deposited in the Gulf during the mining and smelting history of Idrjia [57], and a few hundred tons of Hg also accumulated in the western (Italian) and eastern (Croatian) coasts of the Adriatic from the discharges of chlor-alkali plants and other anthropogenic sources [54].
In the Mediterranean Sea, the total Hg input by rivers has been estimated to be around 6 Mg year−1, and although most of the metal associated with riverine particles is probably deposited on shelves, sediments may also be transported by turbidite currents in the abyssal plain [15,61]. According to Amos et al. [62], 11 to 50% of particulate Hg discharged by rivers can enter the open sea. While most global models attribute all of the ocean’s Hg load to atmospheric deposition, Liu et al. [40], through a high-spatial-resolution dataset, estimated that, in coastal ocean areas, rivers are the main source of Hg (about three times greater than atmospheric deposition), and under climate change and the increasing frequency of extreme flow events, riverine Hg inputs are likely to increase.
Mercury enters the marine environment from atmospheric or anthropogenic and natural terrestrial sources, mainly as dissolved or particulate inorganic forms, and in seawater, it has a strong affinity for chloride, organic matter, and inorganic sulfides. Metal ions forming particulates can settle in estuarine and shelf sediments, which constitute a steady and long-term source of Hg [63]. In the water column, biological and photochemical reactions can reduce Hg2+ to Hg° (often with consequent oversaturation and escape from surface waters), and through biotic and abiotic pathways, Hg2+ can be converted into MMHg. The latter, in turn, can either be accumulated by organisms, or transformed into gaseous dimethylmercury (DMHg), or degraded back into Hg2+ via other biotic and abiotic pathways [63]. Since methylation and demethylation occur simultaneously, the difference in speed between the two processes dictates the bioaccumulation of the metal.
The formation of MMHg is primarily due to biotic processes, and while Hg bound to soluble organic matter and neutral sulfide (HgS°) is directly bioavailable to methylating microorganisms, redox changes in the sedimentary environment can release Hg2+ from deposited organic matter and Mn-Fe oxides into the pore water [58,64]. Solid HgS particles are insoluble in water and are generally believed to play a minimal role in the biogeochemical cycling of Hg. However, in the vertical sediment profile of the Hg-contaminated Augusta Bay (Sicily), Oliveri et al. [65] found that sulfur-oxidizing chemolithoautotrophic bacteria promote Hg mobilization in sedimentary pore waters in the Fe-Mn reduction zone enriched with SO42−. In addition, dissolved organic matter can adsorb onto the surface of HgS nanoparticles, increasing the dissolution of the metal and providing a food source for sulfate-reducing methylating bacteria [66], and extracellular polymeric substances continuously secreted by methylating anaerobic bacteria can enhance the dissolution of cinnabar [67].
Despite a number of recent studies conducted with novel approaches, such as the use of stable Hg isotopes or dynamic flux chambers (e.g., [18]), our knowledge of Hg cycling in the marine environment is still insufficient. The gaps in knowledge are particularly relevant in the Mediterranean Sea, where, according to Cossa et al. [15], there is insufficient information on temporal variations in air–sea exchanges; point source inputs; submarine hydrothermal and groundwater discharges; and exchanges between coastal and open sea environments. In particular, it appears necessary to acquire a better knowledge of the relative bioavailability of Hg in atmospheric deposition and seawater and to assess the role of sediments as a secondary source of Hg2+ and MMHg, especially in coastal environments impacted by natural sources and by past and ongoing human activities. To this end, the accurate quantification of the different Hg chemical species and their fluxes between biotic and abiotic compartments is necessary. However, as discussed by Bowman et al. [19], the results of international intercalibration exercises show that improvements are needed even for the analytical determination of MMHg in seawater.
4. Methylation and Demethylation of Mercury
The bioavailability and methylation of Hg are influenced by many environmental parameters such as temperature, pH, redox potential, the presence of organic matter, and S- and Fe-organic compounds. Essentially, the rate of Hg methylation depends on the availability of ionic mercury (Hg2+) and its uptake by methylating microorganisms and their metabolic activity. However, the mechanisms governing Hg2+ transport into microbial cells, its methylation, and the release of MMHg are not yet well understood [68]. In anaerobic estuarine and coastal environments, hgcAB gene pairs that encode proteins for Hg methylation have been identified in sulfate- and Fe-reducing bacteria and methanogens (e.g., [69]). Since coastal marine sediments are usually characterized by higher concentrations of organic matter and Hg bioavailability, they should play a major role in MMHg production compared with offshore sediments. However, the amount of bioavailable MMHg in the marine environment also depends on its demethylation and the interconversion between monomethyl- (MMHg) and dimethylmercury (DMHg), usually collectively referred to as MeHg. In general, in surface sediments (0–20 cm), methylation exceeds demethylation, whereas the latter process increases with increasing depth [70]. The bio-demethylation of Hg occurs through reductive and oxidative processes: the former are carried out by aerobic microorganisms equipped with the mer operon, and the transcription of the merB gene produces an enzyme capable of splitting the C-Hg bond, generating CH4 and Hg°; the latter are due to anaerobic microorganisms lacking the mer operon and generating Hg2+, CO2, and CH4. In the open sea, and especially in surface waters, photochemical degradation is the dominant pathway in MMHg decomposition [71].
Among cultured microorganisms, only anaerobic bacteria and archaea are known to produce MMHg; nonetheless, tuna and other pelagic predatory fish that have no connection to the sedimentary environment and benthic fauna usually have very high MMHg concentrations. Over the last decade, several studies have shown that sediments are not the main source of MMHg in the marine environment. Mason et al. [72] found that, in the global oceans, most of the MMHg accumulated by marine organisms is produced in upper waters (<1000 m), and its maximum concentrations are usually found in low-oxygen thermocline waters (e.g., [19]). Moreover, recent oceanographic expeditions (e.g., [73]) show that Hg can also be methylated in oxic subsurface waters, and this finding is supported by the results of studies on the Hg stable isotopic signatures of marine particulates, zooplankton, and pelagic fish [74,75]. In Antarctic Sea ice, Gionfriddo et al. [76] found a potential Hg-methylating microorganism with HgcA-like proteins: a microaerophilic and nitrite-oxidizing marine bacterium belonging to the genus Nitrospina. Applying recent methods for hgcAB gene identification in metagenomic and metatranscriptomic datasets, Villar et al. [73] revealed that the genetic potential for Hg methylation is widespread in the oxic subsurface waters of all oceans. Nitrospina bacteria have also been identified as the predominant and widespread microorganisms expressing the hgcAB gene cluster in the surface waters of the Mediterranean Sea.
Despite analytical difficulties and the fast interconversion between MMHg and DMHg, concentrations of MeHg species in low-oxygen intermediate Mediterranean waters appear to indicate a higher methylation capacity than that in the oceans (Table 1). The activity of methylating microorganisms is favored in the low-oxygen zone of the stratified water column, where slowly sinking organic materials are re-mineralized [58]. While in the Arctic, Atlantic, and Pacific waters, MeHg is usually <26% of total Hg concentrations [19], in Mediterranean waters, it can constitute up to 86% of total Hg, and the basin is a source of organic Hg for the North Atlantic [15]. Direct measurements and modeled zonal and vertical distributions of MeHg indicate a halving of concentrations from the western to the eastern Mediterranean Sea, with the lowest values occurring in the least productive areas (Table 1). Moreover, in the eastern basin, the widespread ultra-oligotrophic conditions and very low primary productivity, together with higher irradiance, water temperature, and transparency, likely favor the biological and photochemical degradation of MeHg rather than its formation (Figure 1).
5. Methylmercury Biomagnification in Mediterranean Food Webs and Potential Risks for Human Health
Ever since it was discovered that Hg concentrations in certain Mediterranean fish were much higher than those found in specimens of the same species and size in the Atlantic Ocean, this so-called “Mediterranean Hg anomaly” has been a rather enigmatic issue [6]. The causes were to be found in the numerous factors involved in the production and bioaccumulation of MMHg, such as the unique geological, physicochemical, and hydrodynamic characteristics of the basin; its oligotrophy; and trophic networks made up of organisms that are rather small and grow slower than related species in other seas. After 50 years, many aspects of the Mediterranean Hg cycle are still poorly understood; however, available data indicate that the increased production and bioavailability of MMHg occur in the oxygen minimum zone during maximum nano- and picoplankton cell abundance [58]. Nitrospina bacteria expressing the hgcAB gene cluster could also contribute to MMHg formation in oxic Mediterranean subsurface waters. Through passive transport mechanisms, phytoplankton cells absorb and accumulate MMHg in their cytoplasm at concentrations >100,000 times higher than those in seawater [77].
In the oligotrophic and phosphorous-limited waters of the Mediterranean Sea, most primary production is due to very small autotrophic organisms with a very high surface-to-volume ratio [44]. The slow sinking of the organic remains of phytoplankton, zooplankton, and fecal pellets promotes their re-mineralization, oxygen consumption, and the availability of Hg for methylating microorganisms [58]. Their metabolism could also be favored by the warm water of the Mediterranean Sea [78]. MMHg bio-concentrated by nano- and pico-phytoplankton cells and in the microbial food web is ingested by grazing zooplankton and begins its biomagnification at the earliest levels of marine food webs (Figure 2).
The collection and chemical analysis of plankton of different sizes are quite difficult because, especially in the small size fraction (<100 µm), the samples contain autotrophic and heterotrophic organisms, as well as organic and inorganic particles. Therefore, most of the literature data on Hg concentrations refer to wide ranges of planktonic cell sizes (e.g., [81]). Recently, Tesán-Onrubia et al. [79] measured total Hg and MMHg concentrations in different size fractions of Mediterranean phyto- and zooplankton. They found a decrease in total Hg and MeHg concentrations with increasing phytoplankton cell sizes (Figure 2). In seawater, they measured low concentrations of dissolved Hg (94 ± 32 pg L−1) and MeHg (13 ± 7 pg L−1), and consequently, their bioconcentration in phytoplankton cells was very high: 106 and 105 times, respectively. As shown in Figure 2, the total Hg content decreased in zooplankton organisms, which have longer life cycles than phytoplankton and can at least partially eliminate it. By contrast, MeHg content increased by about one order of magnitude from phytoplankton to larger macrozooplankton organisms that feed on micro- and mesozooplankton (Figure 2). To better highlight MeHg biomagnification at the lower levels of trophic food webs, Figure 2 also reports the concentrations measured by Buckman et al. [80] in unfiltered water samples and the most abundant macrozooplankton taxa from different regions of the western and eastern Mediterranean basins, including some sites with anthropogenic and natural sources of Hg. In agreement with the data reported above on the spatial distribution of MMHg in Mediterranean waters, the highest concentrations were found in samples from the western Mediterranean and, in particular, from the Tyrrhenian and Balearic Seas.
As a consequence, when compared with specimens of the same species, weight, and length from mesotrophic seas, fish from the Mediterranean Sea grow more slowly, ingest prey with higher MMHg concentrations for longer, and have less possibility of “diluting” the metal body burden. Furthermore, the minute size of many primary producers and consumers likely contributes to lengthening Mediterranean food webs and increasing MMHg biomagnification.
In general, fish from the northwestern basin, the Adriatic Sea, and coastal areas most impacted by natural and anthropogenic sources of the metal have higher Hg concentrations than related species from the Aegean or Ionian Sea (Table 2). The mean Hg concentrations in some orders of fish from polluted and unpolluted marine areas of the world [24] are also reported in Table 2 for comparison. However, in the same fishing ground, Hg bioaccumulation in each individual of a species reflects its life history, and those living in or migrating to deeper waters often have higher Hg concentrations. Girolametti et al. [82], for instance, found that the total Hg content in red mullet from Adriatic coastal waters (0.11 ± 0.08 µg g−1 ww) was three times lower than in samples collected in the open sea (0.35 ± 0.22). Large pelagic fish can prey on epipelagic and mesopelagic organisms, and individuals feeding in deeper waters usually accumulate more Hg (e.g., [83]). Tuna, for instance, are long-lived opportunists that feed across vast marine areas and can dive to depths of several hundred meters. They can accumulate very high MMHg concentrations in the edible muscle and, being of considerable commercial interest, contribute to a significant proportion of the global population’s Hg intake. Tseng et al. [84] found that MMHg concentrations in tuna correlated with those in waters (from the surface to 1000 m depth) and in phyto- and zooplankton. Furthermore, by measuring stable Hg isotopes in juvenile Pacific bluefin tuna and their prey, Madigan et al. [85] found that increased mesopelagic foraging reduced tuna growth and increased their MMHg content.
Owing to the notable biodiversity and the number of edible marine species, people living in the Mediterranean coastal zone have always been strong consumers of seafood caught or farmed in the basin. The estimated mean annual per capita consumption is about 25 kg, with up to five courses per week for people living in some islands and coastal areas [86]. Fish and seafood contribute over 93% of the total Hg adsorbed by the general population, and dietary exposure to MMHg in heavy and frequent consumers can damage the kidneys, lungs, and cardiovascular and nervous systems. Overexposure in pregnant women through seafood consumption can have adverse effects on the brain development of embryos and a potential reduction in intelligence quotient (IQ) in children [91].
As shown by the large database on Hg concentrations in Mediterranean organisms analyzed between 1969 and 2015 [12], Hg concentrations in the muscles of several benthic and pelagic species of carnivorous fish are above the upper safe limits established for human consumption by the European Commission [92] (in general, equal to 0.5 µg g−1 ww or, for certain species, 1 µg g−1 ww). Therefore, especially in the most polluted areas of the Mediterranean Sea, it is necessary to evaluate the potential risks of exposure, at least for the most sensitive population segment. Capodiferro et al. [86] analyzed 58 species of wild fish and seafood sold on the markets of Spain, France, and Italy and discovered that, among all analyzed specimens of different lengths and weights, only thirteen species of pelagic or demersal fish, such as sardines, anchovies, seabream, and brown meagre, had Hg concentrations lower than the EC threshold limit. Combining the average Hg concentrations in all organisms analyzed and the mean fish consumption in the three countries, they estimated a weekly Hg intake of about 1.1 µg kg−1 body wt compared with the provisional tolerable weekly intake (TWI) of 1.6 µg Hg kg−1 body wt for adults recommended by the Joint FAO/WHO Expert Committee on Food Additives [93]. Many other studies have estimated Hg ingestion through fish consumption in several Mediterranean countries, but uncertainties still remain regarding the reliability of actual human exposure. The TWI recommended by FAO/WHO, for example, is expressed as total Hg, which is easier and less expensive to determine than MMHg. However, in the muscles of predatory fish, more than 90% of total Hg is usually MMHg; in planktivorous fish and most crustaceans and mollusks, this percentage is significantly lower (e.g., [94]). Therefore, the US-EPA and Health Canada also express tolerable intake limits for Hg as MMHg.
In addition to seafood, by far the main source of Hg in our diet, humans are also exposed to other food sources of this metal such as rice and mushrooms (e.g., [95]) or to Hg released from dental fillings, skin-lightening creams, or through the inhalation of Hg°. Although fish can be consumed raw, most people cook seafood, and during commonly used cooking procedures, foods lose moisture and MMHg, and total Hg concentrations usually tend to increase (e.g., [96]). The availability of Hg from ingested seafood is influenced by many factors, such as the type of consumed organisms, the cooking method, and the co-occurrence of macro- and micronutrients in the diet. Most estimates of the Hg absorption rate (usually referred to as bio-accessibility) assume that about 100% of total Hg in fish muscles is MMHg (which is only true for carnivorous species), and after reviewing the available literature, Bradley et al. [97] concluded that the absorption of MMHg by humans ranges from 12% to 79% and that of Hg2+ from 49% to 69%. For a reliable assessment of health risks, it is also necessary to evaluate the effectiveness of selenium (Se) in mitigating Hg toxicity. High-protein foods such as fish are major sources of Se, and since the 1960s, when it was discovered that Hg can be sequestered in Hg-Se complexes in mammals, many studies have investigated the Se/Hg molar ratio. Unfortunately, molecular interactions between the two elements are not yet fully understood, and the protection of Se against long-term exposure to Hg remains to be definitely proven [98]. For this reason, in a very recent review of the available data on Hg in seafood and the potential risks to consumer health, Zampetti and Barndt [99] concluded that approaches to consumer safety based on the 1:1 (Se:Hg) molar ratio are likely premature.
The consumption of fish brings important health benefits, as it provides proteins, vitamins, minerals, and unsaturated essential fatty acids such as omega-3, and a large part of the fish and shellfish consumed by the coastal populations of the Mediterranean come from local capture or aquaculture. It, therefore, seems necessary to identify, in different areas, marine species with excessive MMHg concentrations and to encourage the consumption of organisms at lower trophic levels with lower Hg concentrations. It also seems appropriate to extend the monitoring of Hg levels in chronically exposed population groups using hair and urine samples since such a non-invasive collection of samples and their preservation require very simple procedures, and tests for the determination of total Hg concentrations are cheap and widespread [100].
6. Climate Change and the Biogeochemical Cycle of Hg in the Mediterranean
The concluding statement of the Global Hg Assessment 2018 [16], “The presence of legacy Hg and the potential for climate change to influence its remobilization have complicated our ability to assess potential future changes”, fits very well with the situation of the Mediterranean Sea. In this semi-enclosed basin, which constitutes a hotspot of climate change and has received large amounts of Hg from geogenic and anthropogenic sources for centuries, it seems difficult to predict possible changes in the bioavailability of Hg and the responses of biotic communities. Models of Hg biogeochemical cycling in the marine environment are often spatially limited, rely on small sample sizes, or are not adequately validated. However, new coupled atmosphere–land–ocean models have recently been developed (e.g., [36,101,102]) that take into account the most recent findings on the redox chemistry of atmospheric Hg and the Hg isotopic composition of seawater, as well as the higher global estimates of riverine Hg inputs. Overall, these models predict that climate change and rising seawater temperatures will reduce the solubility of atmospheric Hg° and increase its escape from the oceans, with fluxes higher than previously estimated [101]. However, total Hg concentrations in surface seawater also depend on wind speed near the surface, which promotes air–sea exchanges. In ocean regions where wind speeds will decrease, total Hg concentrations in surface waters will increase by about the same magnitude as Hg° evasion [102]. While warming polar seawater will likely increase organic carbon mineralization and MMHg production (e.g., [76]), in mid- and low-latitude oceans, rising temperatures are expected to reduce MMHg production [101]. The Mediterranean Sea is very sensitive to climate change, and warming waters will likely increase vertical stratification, further reducing nutrient availability, primary production, and oxygen availability in deep waters. Microplastics are widespread in the Mediterranean Sea, and according to a recent study [103], zooplankton grazing on very small plastic debris increase the production of organic particles, and their remineralization during sinking contributes to a further loss of oxygen.
Increasing atmospheric CO2 concentrations acidify seawater and should favor the development of small phytoplankton cells [36], which already contribute to the notable bioconcentration of Hg and MMHg in Mediterranean autotrophs. Although oxygen depletion in deep thermocline waters may increase MMHg production, the lower productivity of phytoplankton may decrease the scavenging of Hg from the water column by organic materials and its availability to methylating microorganisms. Increased metabolic demand due to warmer and less dense seawater and the spread of smaller autotrophic organisms will likely change the size and composition of zooplankton communities, and global models (e.g., [101]) predict an increase in small-sized zooplankton that prefer to feed on smaller algal cells and a significant decrease in larger species. These changes could give rise to more complex and longer food webs, which could enhance the biomagnification of MMHg. However, despite large regional variations in response to climate change, overall global models predict that smaller zooplankton organisms should reduce MMHg biomagnification [101].
Hotter, saltier, and more acidic Mediterranean waters; reduced primary productivity; and changes in zooplankton communities [104] will likely influence the growth and composition of communities at higher trophic levels. Mediterranean pelagic and benthic food webs could also be affected by the spread of invasive alien species. Although it seems very difficult to predict the potential impact of these changes, we cannot exclude an increase in the biomagnification of MMHg by carnivorous species as a result of longer food chains and a likely reduction in their growth rate and adult size. Furthermore, as shown by [105] for Atlantic cod, overfishing Mediterranean predatory species such as tuna and swordfish may result in dietary shifts and enhanced MMHg accumulation.
Climate change and human activities are modifying the composition and amounts of river inputs, and in a semi-enclosed basin like the Mediterranean Sea, there will be a significant impact on the structure and functioning of coastal ecosystems. Although the possible effects of these changes are poorly understood, using a coupled physico-biogeochemical model, Pagés et al. [106], for instance, found a 60% decrease in river phosphate inputs in the eastern Mediterranean basin since the late 1980s. This change probably contributed to the decrease in the abundance and size of copepods, followed by a reduction in the size of anchovies and sardines.
The only real sink of Hg is burial in soils and sediments, but climate change is enhancing metal mobilization through the increasing occurrence of extreme weather events and exceptional river flows. In the last few decades, the magnitude of climate change and its impacts in the Mediterranean region have exceeded global averages, and changes in the hydrological cycle have become evident with torrential rain events triggering flash floods [107]. Fornasaro et al. [108], for instance, investigated morphological changes that occurred over the last 200 years along the Paglia River (central Italy) and their effects on Hg distribution in the riverbed and floodplain. This river drains areas with numerous cinnabar deposits on the eastern slopes of Mount Amiata, which was intensively exploited by Hg mining and metallurgical activities until a few decades ago. The study found that the morphological changes and the transport, deposition, and re-suspension processes do not allow for a Hg clean-up of the river, and the variability of its flow and, especially, exceptional flood events are increasing estimated Hg inputs (about 40 kg year−1) into the Tyrrhenian Sea. Furthermore, the subsidence of several coastal areas, the increasing frequency and intensity of waves and storms, tidal flooding, and permanent flooding are also increasing the mobilization of Hg and organic matter from estuaries, coastal plains, and lagoons. Sea level rise contributes to the loss of sandy shores and coastal ecosystems. In the Mediterranean, the process is not uniform in space and time; however, Calafat et al. [35] estimated that, after a decrease (an average rate of −0.3 ± 0.5 mm year−1) between 1960 and 1989, the Mediterranean level started to rise with an average rate of 3.6 ± 0.3 mm year−1 in the period 2000–2018, and this faster increase was detected in the Adriatic, Aegean, and Levantine Seas. A number of studies at regional and local scales report the potential impact of the future Mediterranean Sea level rise on alluvial coastal plains, shoreline retreats, and permanent submergence processes (e.g., [109]). Among the main consequences of these processes is the mobilization of dissolved organic matter (DOM) and organic carbon (DOC), which could significantly influence the biogeochemical cycle of Hg in coastal marine ecosystems. The increased inputs of DOM and DOC enhance those of Hg and MMHg (e.g., [110]), contribute to the dissolution of HgS particles, and provide a food source for methylating bacteria [66]. Positive relationships between DOM concentrations, bacterial activity, and MMHg formation have often been reported in marine waters and sediments. Moreover, experiments under controlled conditions in oxic and brackish coastal waters by Rodríguez et al. [96] showed that increased MMHg production under higher DOM inputs is due to an increased bacterial production rate and an abundance of Hg2+ methylating genes rather than changes in bacterial taxonomic composition or Hg2+ methylation genes. Therefore, although it is difficult to predict the possible effects of climate change on the biogeochemical cycle of Hg in the Mediterranean Sea, the “natural” biomagnification of MMHg along food chains will likely persist for a long time, and any reduction in anthropogenic emissions will have limited effects.
7. Future Directions and Research Needs
Despite the number of studies and scientific advances reported above, there are still large uncertainties about the inputs, distribution, and cycling of Hg in the Mediterranean Sea; furthermore, the impact of climate change is making it increasingly difficult to fill data and knowledge gaps. In any case, despite the progressive warming of marine waters and changes in atmospheric deposition patterns, it is necessary to obtain consistent field and laboratory data to define the Hg budget in the Mediterranean. Spatial and temporal changes in the deposition and evasion of different chemical forms of Hg at the air–sea interface need to be quantified through monitoring networks and suitable model studies. Similarly, it is necessary to assess the continental inputs of Hg and DOC from rivers, point sources, and coastal areas. The acquisition of these data will be of particular significance given the increasing frequency of extreme weather events, rising sea levels, shoreline retreats, and the submergence of alluvial coastal plains. Moreover, very few data are available on the transfer of this metal from coastal areas to the open sea and the abyssal plain, and data on Hg contributions to Mediterranean waters from submarine hydrothermal vents, groundwater discharges, and cold seeps are almost entirely lacking. Improvements are needed for more accurate quantification in waters of different chemical species of Hg, rates of Hg2+ methylation, MMHg demethylation, and interconversion between MMHg and DMHg. Although to date most studies have focused on Hg biomethylation, it would seem necessary to also investigate the possible presence of methyl donors in the marine environment and the possible formation of MMHg through abiotic processes. Contrary to what was believed in the past, recent studies show that, in aquatic ecosystems, sediment is not the main source of MMHg, which can form, to a greater extent, in the water column. This finding represents a great achievement in understanding the biogeochemical cycle of Hg, but at the same time, it raises important questions that need to be addressed. For instance, do the same or different processes determine MMHg availability and bioaccumulation in benthic and pelagic organisms? Furthermore, biotic communities in the water column are likely to be more directly affected than benthic ones by changes expected in the Mediterranean basin, such as decreasing nutrient availability, water warming, and stratification. Although this is a rather difficult task, it seems advisable to examine the responses of pelagic and benthic communities to the impacts of climate and environmental changes, including the possible spread of invasive alien species and possible changes in food webs. In this context, for a better understanding of the possible changes in the biogeochemical cycle of Hg, it would be necessary to monitor, above all, the possible spatiotemporal changes in the communities of microbial autotrophs, small prokaryotic and eukaryotic heterotrophs, and the grazing zooplankton (i.e., the organisms that are mostly responsible for the enhanced production, bioconcentration, and bioaccumulation of MMHg in Mediterranean food webs). Finally, as regards the potential health risks for consumers of Mediterranean seafood, it is necessary to carefully verify the actual bio-accessibility of Hg and the protective role of Se.
8. Conclusions
Since the 1970s, it has been known that Mediterranean organisms accumulate higher Hg concentrations than related species of the same size caught in the Atlantic Ocean, and it was hypothesized that the Hg “anomaly” was due to the impact of natural and anthropogenic sources of the metal in this semi-enclosed and oligotrophic basin and to specific eco-physiological characteristics of Mediterranean biota. Although the inputs and fluxes of the different chemical forms of Hg among the different biogeochemical compartments are not yet well defined, studies conducted in recent decades substantially confirm the initial hypotheses: the basin receives relatively high fluxes of Hg from atmospheric, riverine, and geogenic sources, and the metal accumulates especially in coastal sediments and organisms of the northwestern Mediterranean and northern Adriatic. The biotic and abiotic processes leading to the formation of MMHg, its demethylation, and the interconversion between MMHg and DMHg in marine waters have not been fully elucidated. However, the “anomalous” bioaccumulation of MMHg in Mediterranean pelagic organisms is probably due to the greater bioavailability of MMHg in the water column and its efficient uptake by small nano- and pico-phytoplankton cells that develop in the oligotrophic and phosphorous-limited waters of the basin. In this environment, grazing invertebrates and their consumers grow slowly, and their small size likely contributes to longer food webs, further enhancing MMHg biomagnifications in long-lived predatory species.
The Mediterranean Sea is a hotspot of climate change but is also a region of high biodiversity. Possible changes in water salinity, temperature, pH, and water stratification, together with the spread of invasive alien species, make it difficult to predict future changes in food webs and MMHg biomagnification. Recent atmosphere–land–ocean models predict that warming seawater will increase the emission of gaseous Hg° and water stratification with a consequent reduction in bioavailable nutrients in the photic zone and oxygen in deeper waters. However, a possible decrease in wind speed could reduce the Hg° evasion from surface waters. The acidification of Mediterranean seawater and increasing oligotrophy will probably favor the further development of small phyto- and zooplankton cells, while oxygen depletion could increase MMHg production, and possibly longer food webs could enhance its biomagnification. Changes in food webs will likely cause dietary shifts and lower growth rates in organisms at higher trophic levels, which could lead to enhanced MMHg in carnivorous fish and potential risks for the health of terminal consumers such as seabirds, marine mammals, and humans. In the open sea, a decrease in autotroph biomass could decrease the scavenging of Hg and MMHg from the water column and the production of MMHg by methylating microorganisms. However, for pelagic carnivorous species such as tuna, the possible reduction in MMHg availability could be “compensated” for by changes in their diet and a decreasing growth rate. In coastal ecosystems historically affected by Hg inputs from anthropogenic and geogenic sources, climate change through enhanced river floods and the inundation of coastal plains will likely increase the mobilization of Hg, DOM, and DOC and the bioaccumulation of MMHg.
Thus, future research should aim to gain better knowledge of changes in Hg fluxes at the air–sea interface and riverine Hg inputs, as well as those relating to submarine hydrothermal vents, groundwaters, shoreline retreats, the submersion of alluvial coastal plains, and the possible transfer of Hg and organic matter to the open sea and the abyssal plain. Until now, most studies have focused on Hg biomethylation, but it would seem necessary to also investigate the possible contribution of MMHg from abiotic methylation processes. Although a rather difficult task, it seems advisable to assess how changes in nutrient availability, water temperature, acidity, and stratification—together with the spread of invasive alien species—will affect the structure and composition of Mediterranean pelagic and benthic food webs. In this context, possible spatiotemporal changes in autotroph communities and those of small prokaryotic and eukaryotic heterotrophs will be very useful for a better understanding of anomalous Hg bioaccumulation in Mediterranean organisms.
In general, significant decreases in the MMHg bioaccumulation in key commercial species of the Mediterranean Sea are not foreseeable in the near future; it, therefore, appears necessary to identify fishing areas and edible species with excessive Hg concentrations, develop information campaigns, encourage the consumption of species at lower trophic levels, and monitor Hg with hair or urine of consumers exposed to an excessive dietary intake of the metal.
Author Contributions
Conceptualization, writing—original draft preparation, R.B.; writing—review and editing, visualization, E.R.; supervision, R.B. and E.R. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Jensen, S.; Jernelöv, A. Biological methylation of mercury in aquatic organisms. Nature 1969, 223, 753–754. [Google Scholar] [CrossRef]
- Ciusa, W.; Giaccio, M. Il contenuto in rame, zinco, mercurio e piombo di alcune specie ittiche dell’Adriatico. Quad. Merceol. 1971, 11, 1–12. [Google Scholar]
- Thibaud, Y. Tenure en mercure dans quelques poissons de consommation courante. Sci. Peche 1971, 209, 1–10. [Google Scholar]
- Bernhard, M.; Renzoni, A. Mercury concentrations in Mediterranean marine organisms and their environment: Natural or anthropogenic origin. Thalass. Jugosl. 1977, 13, 265–300. [Google Scholar]
- Stoeppler, M.; Bernhard, M.; Backhaus, F.; Schulte, E.K. Comparative studies on trace metal levels ion marine biota. I. Mercury in marine organisms from western Italian coast, the Strait of Gibraltar, and the North Sea. Sci. Total Environ. 1979, 13, 209–223. [Google Scholar] [CrossRef]
- Aston, S.R.; Fowler, S.W. Mercury in the open Mediterranean: Evidence of contamination? Sci. Total Environ. 1985, 43, 13–26. [Google Scholar] [CrossRef] [PubMed]
- Renzoni, A.; Focardi, S.; Fossi, C.; Leonzio, C.; Mayol, J. Comparison between concentrations of mercury and other contaminants in eggs and tissues of Cory’s shearwater Calonectris diomedea collected on Atlantic and Mediterranean islands. Environ. Pollut. 1986, 40, 17–35. [Google Scholar] [CrossRef]
- Renzoni, A.; Zino, F.; Franchi, E. Mercury levels along the food chain and risk for exposed populations. Environ. Res. 1998, 77, 68–72. [Google Scholar] [CrossRef] [PubMed]
- Leonzio, C.; Focardi, S.; Fossi, M.C. Heavy metals and selenium in stranded dolphins of the northern Tyrrhenian (NW Mediterranean). Sci. Total Environ. 1992, 119, 77–84. [Google Scholar] [CrossRef]
- UNEP. Mediterranean Action Plan (MAP); UN Environment Programme, Chemicals and Health Branch: Geneva, Switzerland, 2019; Available online: https://www.unep.org/unepmap/resources/publications (accessed on 2 October 2023).
- Minamata Convention on Mercury. Available online: https://www.mercuryconvention.org (accessed on 10 October 2023).
- Cinnirella, S.; Bruno, D.E.; Pirrone, N.; Horvat, M.; Zivkovic, I.; Evers, D.C.; Johnson, S.; Sunderland, E.M. Mercury concentrations in biota in the Mediterranean Sea, a compilation of 40 years of surveys. Sci. Data 2019, 6, 205. [Google Scholar] [CrossRef]
- Costantini, D.; Bustamante, P.; Bault-Favrou, M.; Dell’Olmo, G. Pattern of mercury exposure and relationships with isotopes and markers of oxidative status in chicks of a Mediterranean seabird. Environ. Pollut. 2020, 260, 114095. [Google Scholar] [CrossRef]
- Rosati, G.; Canu, D.; Lazzari, P.; Solidoro, C. Assessing the spatial and temporal variability of methylmecury biogeochemistry and bioaccumulation in the Mediterranean Sea with a coupled 3D model. Biogeosciences 2022, 19, 3663–3682. [Google Scholar] [CrossRef]
- Cossa, D.; Knoery, J.; Bănaru, D.; Harmelin-Vivien, M.; Sonke, J.E.; Hedgecock, I.M.; Bravo, A.G.; Rosati, G.; Canu, D.; Horvat, M.; et al. Mediterranean mercury assessment 2022: An updated budget, health consequences, and research perspectives. Environ. Sci. Technol. 2022, 56, 3840–3862. [Google Scholar] [CrossRef] [PubMed]
- UNEP. Global Mercury Assessment 2018; UN Environment Programme, Chemicals and Health Branch: Geneva, Switzerland, 2019; Available online: https://www.unep.org/resources/publication/global-mercury-assessment-2018 (accessed on 2 October 2023).
- Albertos, S.; Brerenguer, N.I.; Sánchez-Virosta, P.; Gómez-Ramírez, P.; Jiménez, P.; Torres-Chaparro, M.Y.; Valverde, I.; Navas, I.; María-Mojica, P.; Garcíá-Fernández, A.; et al. Mercury exposure in birds linked to marine ecosystems in the Western Mediterranean. Arch. Environ. Contam. Toxicol. 2020, 79, 435–453. [Google Scholar] [CrossRef]
- Gustin, M.S.; Bowman, K.; Branfireum, B.; Chetelat, J.M.; Eckley, C.S.; Hammerschmidt, C.R.; Lamborg, C.; Lyman, S.; Martinez-Cortizas, A.; Sommar, J.; et al. Mercury biogeochemical cycling: A synthesis of recent scientific advances. Sci. Total Environ. 2020, 737, 139619. [Google Scholar] [CrossRef]
- Bowman, K.L.; Lamborg, C.H.; Agather, A.M. A global perspective on mercury cycling in the ocean. Sci. Total Environ. 2020, 710, 136166. [Google Scholar] [CrossRef] [PubMed]
- Sonke, J.E.; Angot, H.; Zhang, Y.; Poulain, A.; Björn, E.; Schartup, A. Global change effects on biogeochemical mercury cycling. Ambio 2023, 52, 853–876. [Google Scholar] [CrossRef]
- Lee, C.S.; Lutcavage, M.; Chandler, E.; Madigan, D.J.; Cerrato, R.M.; Fisher, N.S. Declining mercury concentrations in bluefin tuna reflect reduced emissions to the North Atlantic Ocean. Environ. Sci. Technol. 2016, 50, 12825–12830. [Google Scholar] [CrossRef]
- Wang, F.; Outridge, P.M.; Feng, X.; Meng, B.; Heimbürger-Boavid, L.-E.; Mason, R.P. How closely do mercury trends in fish and other aquatic wildlife track those in the atmosphere? Implications for evaluating the effectiveness of the Minamata Convention. Sci. Total Environ. 2019, 674, 58–70. [Google Scholar] [CrossRef]
- Morris, A.D.; Wilson, S.J.; Fryer, R.J.; Thomas, P.J.; Hudelson, K.; Andreasen, B.; Blévin, P.; Bustamante, P.; Chastel, O.; Christensen, G.; et al. Temporal trends of mercury in Arctic biota: 10 more years of progress in Arctic monitoring. Sci. Total Environ. 2022, 839, 155803. [Google Scholar] [CrossRef]
- AMAP/UNEP. Technical Background Report for the Global Mercury Assessment 2018; Arctic Monitoring and Assessment Programme: Oslo, Norway; UN Environment Programme, Chemicals and Health Branch: Geneva, Switzerland, 2019; 426p. [Google Scholar]
- Bagnato, E.; Oliveri, E.; Acquavita, A.; Covelli, S.; Petranich, E.; Barra, M.; Italiano, F.; Parello, F.; Sprovieri, M. Hydrochemical mercury and air-sea exchange over the submarine hydrothermal vents off-shore Panarea Island (Aeolian Arc, Tyrrhenian Sea). Mar. Chem. 2017, 194, 63–78. [Google Scholar] [CrossRef]
- Andaloro, F.; Romeo, T.; Renzi, M.; Guerranti, C.; Perra, G.; Consoli, P.; Perzia, P.; Focardi, S.E. Alteration of potential harmful elements levels in sediments and biota from the central Mediterranean Sea (Aeolian Archipelago) following an episode of intense volcanic activity. Environ. Monit. Assess. 2012, 184, 4035–4047. [Google Scholar] [CrossRef]
- Baldi, F.; Bargagli, R. Mercury pollution in marine sediments near a chlor-alkali plant: Distribution and availability of the metal. Sci. Total Environ. 1984, 39, 15–26. [Google Scholar] [CrossRef]
- Covelli, S.; Faganelli, J.; Horvat, M.; Brambati, A. Mercury contamination of coastal sediments as the result of long-term cinnabar mining activity (Gulf of Trieste, northern Adriatic Sea). Appl. Geochem. 2001, 16, 541–558. [Google Scholar] [CrossRef]
- Rimondi, V.; Costagliola, P.; Lattanzi, P.; Morelli, G.; Cara, G.; Cencetti, C.; Fagotti, C.; Fredduzzi, A.; Marchetti, G.; Sconocchia, A.; et al. A 200 km-long mercury contamination of the Paglia and Tiber floodplain: Monitoring results and implications for environmental management. Environ. Pollut. 2019, 255, 113191. [Google Scholar] [CrossRef]
- Ozkan, E.Y.; Buyukisik, H.B.; Kontas, A.; Turkdogan, M. A survey of metal concentrations in marine sediment cores in the vicinity of an old mercury-mining area in Karaburun, Aegean Sea. Environ. Sci. Pollut. Res. 2017, 24, 13823–13836. [Google Scholar] [CrossRef]
- Degetto, S.; Schintu, M.; Contu, A.; Sbrignadello, G. Santa Gilla Lagoon (Italy): A mercury sediment pollution case study. Contamination assessment and restoration of the site. Sci. Total Environ. 1997, 204, 49–56. [Google Scholar] [CrossRef]
- Tessier, E.; Garnier, C.; Mullot, J.-U.; Lenoble, V.; Arnaud, M.; Raynaud, M.; Mounier, S. Study of the spatial and historical distribution of sediment inorganic contamination in the Toulon Bay (France). Mar. Pollut. Bull. 2011, 62, 2075–2086. [Google Scholar] [CrossRef] [PubMed]
- Rosati, G.; Solidoro, C.; Canu, D. Mercury dynamics in a changing coastal area over industrial and post-industrial phases: Lessons from the Venice Lagoon. Sci. Total Environ. 2020, 743, 140586. [Google Scholar] [CrossRef] [PubMed]
- Richon, C.; Dutay, J.-C.; Bopp, L.; Le Vu, B.; Orr, J.C.; Somot, S.; Dulac, F. Biogeochemical response of the Mediterranean Sea to the transient SRES-A2 climate change scenario. Biogeosciences 2019, 16, 135–165. [Google Scholar] [CrossRef]
- Calafat, F.M.; Frederiske, T.; Horsburgh, K. The sources of sea-level changes in the Mediterranean Sea since 1960. J. Geophys. Res. 2022, 127, e2022JC019061. [Google Scholar] [CrossRef]
- Zhang, Y.; Dutkiewicz, S.; Sunderland, E.M. Impact of climate change on methylmercury formations and bioaccumulation in the 21th century ocean. One Earth 2021, 4, 279–288. [Google Scholar] [CrossRef]
- MedECC. Climate and Environmental Change in the Mediterranean Basin—Current Situation and Risks for the Future: First Mediterranean Assessment Report; Cramer, W., Guiot, J., Marini, K., Eds.; Union for the Mediterranean, Plan Bleu, UNEP/MAP: Marseille, France, 2020; 632p. [Google Scholar] [CrossRef]
- Zenetos, A.; Galanidi, M. Mediterranean non indigenous species at the start of the 2020s: Recent changes. Mar. Biodivers. Rec. 2020, 13, 10. [Google Scholar] [CrossRef]
- Coll, M.; Piroddi, C.; Steenbeek, J.; Kaschner, K.; Ben Rais Lasram, F.; Aguzzi, J.; Ballesteros, E.; Nike Bianchi, C.; Corbera, J.; Dailianis, T.; et al. The biodiversity of the Mediterranean Sea: Estimates, patterns and threats. PLoS ONE 2010, 5, e11842. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Zhang, Q.; Maavara, T.; Liu, S.; Wang, X.; Raymond, P.A. Rivers as the largest source of mercury to coastal oceans worldwide. Nat. Geosci. 2021, 14, 672–677. [Google Scholar] [CrossRef]
- Costa, V.; Sciuretti, V.; Consoli, P.; Manea, E.; Menini, E.; Andaloro, F.; Romeo, T.; Danovaro, R. Volcanic-associated ecosystems of the Mediterranean Sea: A systematic map and an interactive tool to support their conservation. PeerJ 2023, 11, e15162. [Google Scholar] [CrossRef]
- Bakalowicz, M. Coastal karst groundwater in the Mediterranean: A resource to be preferably exploited onshore, not from karst submarine springs. Geosciences 2018, 8, 258. [Google Scholar] [CrossRef]
- Lazzari, P.; Solidoro, C.; Salon, S.; Bolzon, G. Spatial variability of phosphate and nitrate in the Mediterranean Sea: A modeling approach. Deep Sea Res. 2016, 108, 39–52. [Google Scholar] [CrossRef]
- Sammartino, M.; Di Cicco, A.; Marullo, S.; Santoleri, R. Spatio-temporal variability of micro-, nano- and pico-phytoplankton in the Mediterranean Sea from satellite ocean colour data of SeaWiFS. Ocean Sci. 2015, 11, 759–778. [Google Scholar] [CrossRef]
- Adloff, F.; Somot, S.; Sevault, F.; Jordà, G.; Aznar, R.; Dèqué, M.; Herrmann, M.; Marcos, M.; Dubois, C.; Padorno, E.; et al. Mediterranean Sea response to climate change in ensemble of twenty first century scenarios. Clim. Dyn. 2015, 45, 2775–2802. [Google Scholar] [CrossRef]
- Baumann, Z.; Jonsson, S.; Mason, R.P. Geochemistry of mercury in the marine environment. Encycl. Ocean. Sci. 2019, 6, 301–308. [Google Scholar] [CrossRef]
- Bargagli, R.; Barghigiani, C. Lichen biomonitoring of mercury emission and deposition in mining, geothermal and volcanic areas of Italy. Environ. Monit. Assess. 1991, 16, 265–275. [Google Scholar] [CrossRef]
- Edner, H.; Ragnarson, P.; Svanberg, S.; Wallinder, E.; Deliso, A.; Ferrara, R.; Maserti, B.E. Differential absorption lidar mapping of atmospheric atomic mercury in Italian geothermal fields. J. Geophys. Res. 1992, 97, 3779–3786. [Google Scholar] [CrossRef]
- Bargagli, R.; Cateni, D.; Nelli, L.; Olmastroni, S.; Zagarese, B. Environmental impact of trace element emissions from geothermal power plants. Arch. Environ. Contam. Toxicol. 1997, 33, 172–181. [Google Scholar] [CrossRef] [PubMed]
- Ferrara, R.; Mazzolai, B.; Lanzillotta, E.; Nucaro, E.; Pirrone, N. Volcanoes as emission sources of atmospheric mercury in the Mediterranean basin. Sci. Total Environ. 2000, 259, 115–121. [Google Scholar] [CrossRef]
- Gencarelli, C.N.; De Simone, F.; Hedgecock, I.M.; Sprovieri, F.; Yang, X.; Pirrone, N. European and Mediterranean mercury modelling: Local and long-range contribution to the deposition flux. Atmos. Environ. 2015, 117, 162–168. [Google Scholar] [CrossRef]
- De Simone, F.; Gencarelli, C.N.; Hedgecock, I.M.; Pirrone, N. A modeling comparison of mercury deposition from current anthropogenic mercury emission inventories. Environ. Sci. Technol. 2016, 50, 5154–5162. [Google Scholar] [CrossRef]
- Baldi, F.; Bargagli, R. Chemical leaching and specific surface measurements of marine sediments in the evaluation of mercury contamination near cinnabar deposits. Mar. Environ. Res. 1982, 6, 69–82. [Google Scholar] [CrossRef]
- Kotnik, J.; Horvat, M.; Ogrinc, N.; Fajon, V.; Žagar, D.; Cossa, D.; Sprovieri, F.; Pirrone, N. Mercury speciation in the Adriatic Sea. Mar. Pollut. Bull. 2015, 96, 136–148. [Google Scholar] [CrossRef]
- Cossa, D.; Durrieu de Madron, X.; Schäfer, J.; Guédron, S.; Mrusczak, S.; Castelle, S.; Naudin, J.-J. Sources and exchange of mercury in the waters of the Northwestern Mediterranean margin. Progr. Oceanogr. 2018, 163, 172–183. [Google Scholar] [CrossRef]
- Širca, A.; Rajar, R.; Harris, R.C.; Horvat, M. Mercury transport and fate in the Gulf of Trieste (Northern Adriatic)—A two dimensional modelling approach. Environ. Model. Softw. 1999, 14, 645–655. [Google Scholar] [CrossRef]
- Žagar, D.; Knap, A.; Warwick, J.; Rajar, R.; Horvat, M.; Četina, M. Modelling of mercury transport and transformation processes in the Idrijca and Soča river system. Sci. Total Environ. 2006, 368, 149–163. [Google Scholar] [CrossRef]
- Heimbürger, L.-H.; Cossa, D.; Marty, J.-C.; Migon, C.; Averty, B.; Dufour, A.; Ras, J. Methyl mercury distributions in relation to the presence of nano- and picoplankton in an oceanic water column (Ligurian Sea, North-western Mediterranean). Geochim. Cosmochim. Acta 2010, 74, 5549–5559. [Google Scholar] [CrossRef]
- Jiskra, M.; Heimbürger-Boavida, L.-E.; Desgranges, M.-M.; Petrova, M.V.; Dufour, A.; Ferreira-Araujo, B.; Masbou, J.; Chmeleff, J.; Melilotus, T.; Point, D.; et al. Mercury stable isotopes constrain atmospheric sources to the ocean. Nature 2021, 597, 678–682. [Google Scholar] [CrossRef] [PubMed]
- Munson, K.M.; Lamborg, C.H.; Swarr, G.J.; Saito, M.A. Mercury species concentrations and fluxes in the Central Tropical Pacific Ocean. Glob. Biogeochem. Cycles 2015, 29, 656–676. [Google Scholar] [CrossRef]
- Palanques, A.; Guillen, G.; Puig, P.; Durrieu de Madron, X. Storm-driven shelf-to-canyon suspended sediment transport at the southwestern Gulf of Lions. Cont. Shelf Res. 2008, 28, 1947–1956. [Google Scholar] [CrossRef]
- Amos, H.M.; Jacob, D.J.; Kocman, D.; Horowitz, H.M.; Zhang, Y.; Dutkiewicz, S.; Horvat, M.; Corbitt, E.S.; Krabbenhoft, D.P.; Sunderland, E.M. Global biogeochemical implications of mercury discharges from rivers and sediment burial. Environ. Sci. Technol. 2014, 48, 9514–9522. [Google Scholar] [CrossRef]
- Hsu-Kim, H.; Kucharzyk, K.H.; Zhang, T.; Deshusses, A. Mechanisms regulating mercury bioavailability for methylating microorganisms in the aquatic environment: A critical review. Environ. Sci. Technol. 2013, 47, 2441–2456. [Google Scholar] [CrossRef]
- Cossa, D.; Gobeil, C. Mercury speciation in the Lower St. Lawrence Estuary. Can. J. Fish. Aquat. Sci. 2000, 57, 138–147. [Google Scholar] [CrossRef]
- Oliveri, E.; Salvagio Manta, D.; Bonsignore, M.; Cappello, S.; Tranchida, G.; Bagnato, E.; Sabatino, N.; Santisi, S.; Sprovieri, M. Mobility of mercury in contaminated marine sediments: Biogeochemical pathways. Mar. Chem. 2016, 186, 1–10. [Google Scholar] [CrossRef]
- Thian, L.; Guan, W.; Ji, Y.; He, X.; Chen, W.; Alvarez, P.J.J.; Zhang, T. Microbial methylation potential of mercury sulphide particles dictated by surface structure. Nat. Geosci. 2021, 14, 409–416. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhao, Z.; Fang, Q.; Qiao, R.; Zhang, T. Extracellular polymeric substances enhance dissolution and microbial methylation of mercury sulphide minerals. Environ. Sci. Process. Impacts 2023, 25, 44–55. [Google Scholar] [CrossRef] [PubMed]
- Luo, H.; Cheng, Q.; He, D.; Sun, J.; Li, J.; Pan, X. Recent advances in microbial mercury methylation: A review on methylation habitat, methylator, mechanism, and influencing factors. Proc. Saf. Environ. Prot. 2023, 170, 286–296. [Google Scholar] [CrossRef]
- Regnell, O.; Watras, C.J. Microbial mercury methylation in aquatic environments: A critical review of published field and laboratory studies. Environ. Sci. Technol. 2019, 53, 4–19. [Google Scholar] [CrossRef] [PubMed]
- Drott, A.; Lambertsson, L.; Björn, E.; Skyllberg, U. Importance of dissolved neutral mercury sulphides for methyl mercury production in contaminated sediments. Environ. Sci. Technol. 2007, 41, 2270–2276. [Google Scholar] [CrossRef]
- Hammerschmidt, C.R.; Fitzgerald, W.F. Iron-mediated photochemical decomposition of methylmercury in the Arctic Alaskan lake. Environ. Sci. Technol. 2010, 44, 6138–6143. [Google Scholar] [CrossRef]
- Mason, R.P.; Choi, A.L.; Fitzgerald, W.F.; Hammerschmidt, C.R.; Lamborg, C.H.; Soerense, A.L.; Sunderland, E.M. Mercury biogeochemical cycling in the ocean and policy implications. Environ. Res. 2012, 119, 101–117. [Google Scholar] [CrossRef]
- Villar, E.; Cabrol, L.; Heimbürger-Boavida, L.-E. Widespread microbial mercury methylation genes in the global ocean. Environ. Microbiol. Rep. 2020, 12, 277–287. [Google Scholar] [CrossRef]
- Blum, J.D. Marine mercury breakdown. Nat. Geosci. 2011, 4, 139–140. [Google Scholar] [CrossRef]
- Motta, L.C.; Blum, J.D.; Johnson, M.W.; Umhau, B.P.; Popp, B.N.; Washburn, S.J.; Drazen, J.C.; Benitez-Nelson, C.R.; Hannides, C.C.S.; Close, H.G.; et al. Mercury cycling in the North Pacific subtropical gyre as revealed by mercury stable isotope ratios. Glob. Biogeochem. Cycles 2019, 33, 777–794. [Google Scholar] [CrossRef]
- Gionfriddo, C.M.; Tate, M.T.; Wick, R.R.; Schultz, M.B.; Zemia, A.; Thelen, M.P.; Schofield, R.; Krabbenhoft, D.P.; Holt, K.E.; Moreau, J.W. Microbial mercury methylation in Antarctic sea ice. Nat. Microbiol. 2016, 1, 16127. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.S.; Fisher, N.S. Methylmercury uptake by diverse marine phytoplankton. Limnol. Oceanog. 2016, 61, 1626–1639. [Google Scholar] [CrossRef]
- Bacci, E. Mercury in the Mediterranean. Mar. Pollut. Bull. 1989, 20, 59–63. [Google Scholar] [CrossRef]
- Tesán-Onrubia, J.A.; Heimbürger-Boavida, L.-E.; Dufour, A.; Harmelin-Vivien, M.; García-Arévalo, I.; Knoery, J.; Thomas, B.; Carlotti, F.; Tedetti, M.; Bănaru, D. Bioconcentration, bioaccumulation and biomagnification of mercury in plankton of the Mediterranean Sea. Mar. Pollut. Bull. 2023, 194, 115439. [Google Scholar] [CrossRef]
- Buckman, K.L.; Lane, O.; Kotnik, J.; Bratkic, A.; Sprovieri, F.; Horvat, M.; Pirrone, N.; Evers, D.C.; Chen, C.Y. Spatial and taxonomic variation of mercury concentration in low trophic level fauna from Mediterranean Sea. Ecotoxicology 2018, 27, 1341–1352. [Google Scholar] [CrossRef]
- Bargagli, R.; Monaci, F.; Sanchez-Hernandez, J.C.; Cateni, D. Biomagnification of mercury in an Antarctic marine coastal food web. Mar. Ecol. Prog. Ser. 1998, 169, 65–76. [Google Scholar] [CrossRef]
- Girolametti, F.; Frapiccini, E.; Annibaldi, A.; Illuminati, S.; Panfili, M.; Marini, M.; Santojanni, A.; Truzzi, C. Total mercury (THg) content in red mullet (Mullus barbatus) from Adriatic Sea (central Mediterranean Sea): Relation to biological parameters, sampling area and human health risk assessment. Appl. Sci. 2022, 12, 10083. [Google Scholar] [CrossRef]
- Choy, C.A.; Popp, B.N.; Kaneko, J.J.; Drazen, J.C. The influence of depth on mercury levels in pelagic fishes and their prey. Proc. Natl. Acad. Sci. USA 2009, 106, 13865–13869. [Google Scholar] [CrossRef]
- Tseng, C.-M.; Ang, S.-J.; Chen, Y.-S.; Shiao, J.-S.; Lamborg, C.H.; He, X.; Reinferder, J.R. Bluefin tuna reveal global patterns of mercury pollution and bioavailability in the world’s oceans. Proc. Natl. Acad. Sci. USA 2021, 118, e2111205118. [Google Scholar] [CrossRef]
- Madigan, D.J.; Li, M.; Yin, R.; Baumann, H.; Snodgrass, O.E.; Dewar, H.; Krabbenhoft, D.P.; Baumann, Z.; Fisher, N.S.; Balcom, P.; et al. Mercury stable isotopes reveal influence of foraging depth on mercury concentrations and growth in Pacific bluefin tuna. Environ. Sci. Technol. 2018, 52, 6256–6264. [Google Scholar] [CrossRef]
- Capodiferro, M.; Marco, E.; Grimalt, J.O. Wild fish and seafood species in the western Mediterranean Sea with low safe mercury concentrations. Environ. Pollut. 2022, 314, 120274. [Google Scholar] [CrossRef] [PubMed]
- Aissioui, S.; Poirier, L.; Amara, R.; Ramdane, Z. Concentrations of lead, cadmium and mercury in sardines, Sardina pilchardus (Walbaum, 1972) from the Algerian coast and health risks. J. Food Compos. Anal. 2022, 109, 104490. [Google Scholar] [CrossRef]
- Aissioui, S.; Poirier, L.; Amara, R.; Ramdane, Z. Concentrations of lead, cadmium, and mercury in Mullus barbatus from the Algerian coast and health risk associated to its consumption. Reg. Stud. Mar. Sci. 2021, 47, 101959. [Google Scholar] [CrossRef]
- El-Sherbiny, M.M.; Sallam, K.I. Residual content and health risk assessment of mercury, lead and cadmium in sardine and mackerel from Mediterranean Sea coast, Egypt. J. Food Compos. Anal. 2021, 2021, 103749. [Google Scholar] [CrossRef]
- Renieri, E.A.; Safenkova, I.V.; Alegakis, A.K.; Slutskaya, E.S.; Kokaraki, V.; Kentouri, M.; Dzantiev, B.B.; Tsatsakis, A.M. Cadmium, lead and mercury in muscle tissue of gilthead seabream and seabass: Risk evaluation for consumers. Food Chem. Toxicol. 2019, 124, 439–449. [Google Scholar] [CrossRef] [PubMed]
- Abbass, K.; Huusko, A.; Knutsen, H.K.; Nieminen, P.; Myllynen, P.; Meltzer, H.M.; Vahakangas, K.; Rautio, A. Quantitative estimation of mercury intake by toxicokinetic modelling based on total mercury levels in humans. Environ. Int. 2018, 114, 1–11. [Google Scholar] [CrossRef] [PubMed]
- European Commission. Directive 2013/39/EU of the European Parliament and of the Council of 12 August 2013. Off. J. Eur. Union 2013, L. 226, 1–17. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/HTML/?uri=CELEX:32013L0039 (accessed on 1 October 2023).
- World Health Organization; Food and Agriculture Organization. Evaluation of certain contaminants in food. Seventy-second Report of the Joint FAO/WHO Expert Committee on Food Additives; WHO-Technical report series 959. In Proceedings of the Seventy-Second Meeting of the Joint FAO/WHO Expert Committee on Food Additives, Rome, Italy, 16–25 February 2010; Available online: https://www.who.int/publications/i/item/9789241209595 (accessed on 1 October 2023).
- Rodríguez, J.; Andersson, A.; Björn, E.; Timonen, S.; Brugel, S.; Skrobonja, A.; Rowe, O. Inputs of dissolved organic matter enhance bacterial production and methylmercury formation in oxic coastal waters. Front. Microbiol. 2022, 13, 809166. [Google Scholar] [CrossRef]
- Bargagli, R.; Baldi, F. Mercury and methylmercury in higher fungi and their relation with the substrata in a cinnabar mining area. Chemosphere 1984, 13, 1059–1071. [Google Scholar] [CrossRef]
- Rodrigues, E.T.; Coelho, J.P.; Pereira, E.; Pardal, M.A. Are mercury in fishery products appropriate to ensure low risk to high fish-consumption populations? Mar. Pollut. Bull. 2023, 186, 114464. [Google Scholar] [CrossRef]
- Bradley, M.A.; Barst, B.D.; Basu, N. A review of mercury bioavailability in humans and fish. Int. J. Environ. Res. Public Health 2017, 14, 169. [Google Scholar] [CrossRef] [PubMed]
- Tinggi, U.; Perkins, A.V. Selenium status: Its interactions with dietary mercury exposure and implications in human health. Nutrients 2022, 14, 5308. [Google Scholar] [CrossRef]
- Zampetti, C.J.; Barndt, J.E. Co-considering selenium concentrations alters mercury-based fish and seafood consumption advice: A data compilation and critical assessment. Environ. Sci. Technol. Lett. 2023, 10, 179–185. [Google Scholar] [CrossRef]
- Wang, Y.; Li, L.; Yao, C.; Tian, X.; Wu, Y.; Xie, Q.; Wang, D. Mercury in human hair and its implications for health investigation. Curr. Opin. Environ. Sci. Health 2021, 22, 100271. [Google Scholar] [CrossRef]
- Wang, Y.; Wu, P.; Zhang, Y. Climate-driven changes of global marine mercury cycles in 2100. Proc. Natl. Acad. Sci. USA 2023, 120, e2202488120. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhang, P.; Song, Z.; Huang, S.; Yuan, T.; Wu, P.; Shah, V.; Liu, M.; Chen, L.; Wang, X.; et al. An updated global mercury budget from a coupled atmosphere-land-ocean model: 40% more re-emissions buffer the effect of primary emission reductions. One Earth 2023, 6, 316–325. [Google Scholar] [CrossRef]
- Kvale, K.; Oschlies, A. Recovery from microplastic-induced marine deoxygenation make take centuries. Nat. Geosci. 2023, 16, 10–12. [Google Scholar] [CrossRef]
- Reale, M.; Cossarini, G.; Lazzari, P.; Lovato, T.; Bolzon, G.; Masina, S.; Solidoro, C.; Salon, S. Acidification, deoxygenation, and nutrient and biomass declines in a warming Mediterranean Sea. Biogeosciences 2022, 19, 4035–4065. [Google Scholar] [CrossRef]
- Schartup, A.T.; Thackray, C.P.; Qureshi, A.; Dassuncao, C.; Gillespie, K.; Hanke, A.; Sunderland, E.M. Climate change and overfishing increase neurotoxicant in marine predators. Nature 2019, 572, 648–650. [Google Scholar] [CrossRef]
- Pagés, R.; Baklouti, M.; Barrier, N.; Richon, C.; Dutay, J.-C.; Motin, T. Changes in rivers inputs during the last decades significantly impacted the biogeochemistry of the eastern Mediterranean basin: A modelling study. Progr. Oceanog. 2020, 181, 102242. [Google Scholar] [CrossRef]
- Zittis, G.; Almazroui, M.; Alpert, P.; Ciais, P.; Cramer, W.; Dahdal, Y.; Fnais, M.; Francis, D.; Hadjinicolaou, P.; Howari, F.; et al. Climate change and weather extremes in the Eastern Mediterranean and Middle East. Rev. Geophys. 2022, 60, e2021RG000762. [Google Scholar] [CrossRef]
- Fornasaro, S.; Morelli, G.; Rimondi, V.; Fagotti, C.; Friani, R.; Lattanzi, P.; Costagliola, P. The extensive mercury contamination in soil and legacy sediments of the Paglia River basin (Tuscany, Italy): Interplay between Hg-mining waste discharges along rivers, 1960s economic boom, and ongoing climate change. J. Soils Sediments 2022, 22, 656–671. [Google Scholar] [CrossRef]
- Antonioli, F.; De Falco, G.; Lo Presti, V.; Moretti, L.; Scardino, G.; Anzidei, M.; Mastronuzzi, G. Relative sea-level rise and potential submersion risk for 2100 on 16 coastal plains of the Mediterranean Sea. Waters 2020, 12, 2173. [Google Scholar] [CrossRef]
- Krabbenhoft, D.P.; Sunderland, E.M. Global change and mercury. Science 2013, 341, 1457–1458. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Sources and cycling of mercury in the Mediterranean Sea.

Figure 2.
Total Hg accumulation and MeHg biomagnification at the lower levels of the Mediterranean food web. Concentrations (ng g−1 dw) in phyto- and zooplankton of different sizes from [79]; those in different macrozooplankton taxa are from [80].


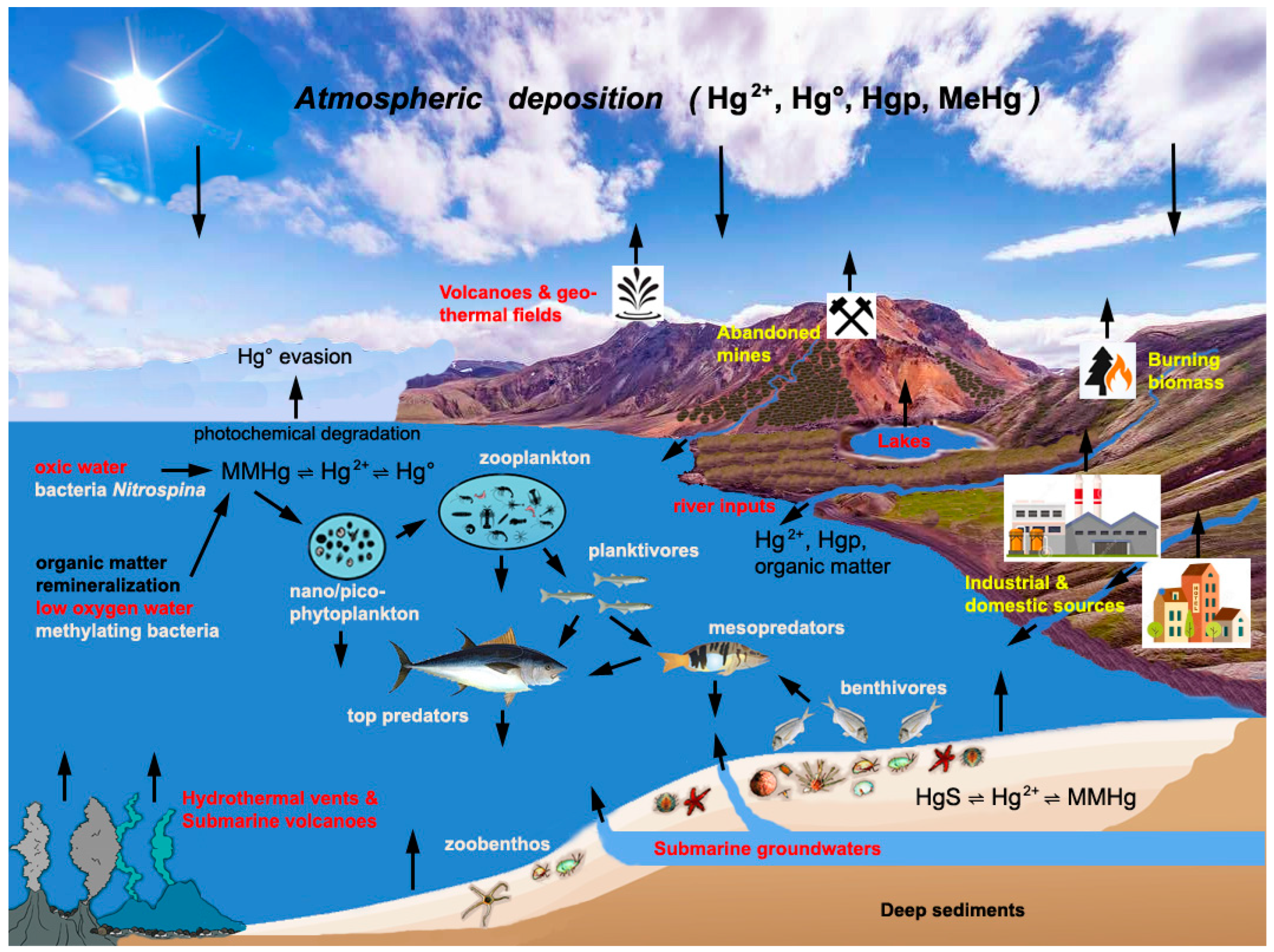{kind=link}
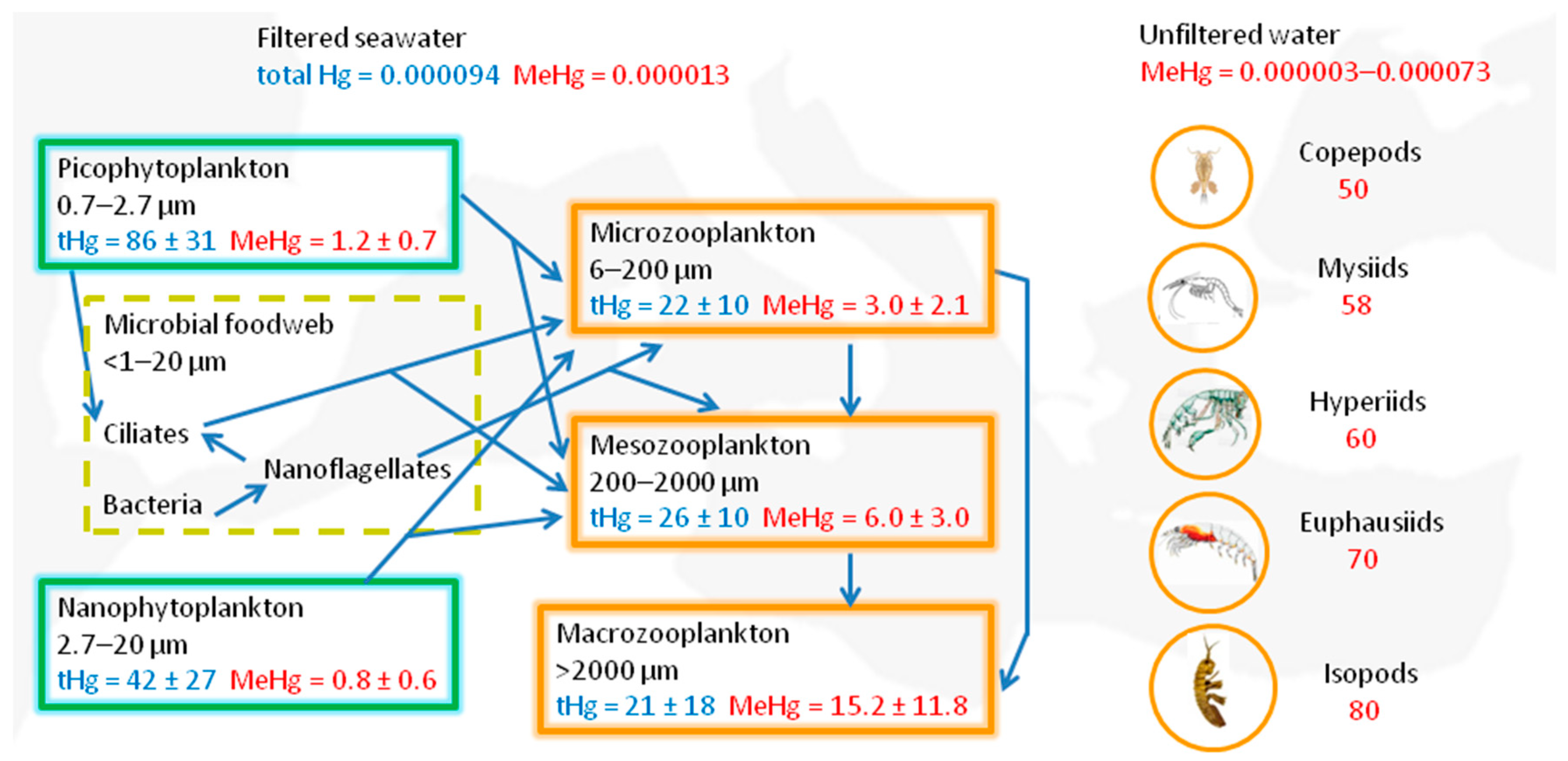{kind=link}
Table 1.
Total Hg and MeHg concentrations (mean ± SD or range, pmol L−1) in filtered and unfiltered water samples from the Mediterranean Sea, Atlantic Ocean, and Pacific Ocean. NA, not analyzed.
Table 1.
Total Hg and MeHg concentrations (mean ± SD or range, pmol L−1) in filtered and unfiltered water samples from the Mediterranean Sea, Atlantic Ocean, and Pacific Ocean. NA, not analyzed.
| Location | Depth | Total Hg | MeHg | References |
|---|---|---|---|---|
| Mediterranean Sea | ||||
| Ligurian Sea | From surface to 2200 m | NA | 0.30 ± 0.17 | [58] |
| Adriatic Sea | From surface to 1000 m | 0.78–6.97 | 0.12–2.48 | [54] |
| Marseille (Endoume Pier) | surface | 4.9–6.7 | NA | [59] |
| 20 km off-shore | From surface to 1400 m | 0.8–1.1 | 0.04–0.40 | |
| Gulf of Lion | Inner shelf | 1.52 ± 1.00 | NA | [15] |
| Continental slope | 1.09 ± 0.15 | NA | ||
| W Mediterranean | From 500 to 3000 m | 1.02 ± 0.12 | <0.02–0.71 | |
| E Mediterranean | Intermediate waters | 1.02 ± 0.12 | <0.02–0.23 | |
| Atlantic Ocean | ||||
| North Atlantic | From surface to 4500 m | 0.46–0.83 | 0.04–0.4 | [59] |
| North Atlantic | <150 m | 0.69 ± 0.39 | 0.13 ± 0.12 | [19] |
| 150–1000 m | 0.96 ± 0.34 | 0.22 ± 0. 05 | ||
| Gulf of Cadiz | From surface to 4000 m | 0.86 ± 0.27 | NA | [15] |
| Pacific Ocean | ||||
| Central North | <150 m | 0.47 ± 0.36 | 0.07 ± 0.03 | [60] |
| >150–1000 m | 1.1 ± 0.29 | 0.21 ± 0.02 | ||
| Equatorial | <150 m | 0.45 ± 0.31 | 0.08 ± 0.02 | |
| >150–1000 m | 0.93 ± 0.23 | 0.14 ± 0.01 |
Table 2.
Total mercury concentrations (µg g−1 ww) in the muscles of commercial fish species from different regions of the Mediterranean Sea compared with global mean values for some fish orders [24].
Table 2.
Total mercury concentrations (µg g−1 ww) in the muscles of commercial fish species from different regions of the Mediterranean Sea compared with global mean values for some fish orders [24].
| Fishing Area | Species | Length (cm) Mean or Range | Total Hg Mean ± SD | References |
|---|---|---|---|---|
| NW Mediterranean Spain, France, Italy | Engraulis encrasicolus | 14 | 0.077 ± 0.05 | [86] |
| Sardina pilchardus | 16 | 0.084 ± 0.04 | ||
| Mullus surmuletus | 20 | 0.18 ± 0.12 | ||
| Sparus aurata | 36 | 0.17 ± 0.13 | ||
| Solea solea | - | 0.10 (median) | ||
| Scomber scombrus | - | 0.41 (median) | ||
| Trachurus trachurus | - | 0.35 (median) | ||
| Conger conger | - | 0.38 (median) | ||
| Dentex dentex | - | 1.00 (median) | ||
| Adriatic Sea | Mullus barbatus | 10–17 | 0.20 ± 0.15 | [82] |
| Algerian coast | Sardina pilchardus | 12–15 | 0.10 ± 0.03 | [87] |
| Mullus barbatus | 13–16 | 0.18 ± 0.07 | [88] | |
| Egypt coast | Sardina pilchardus | 17.6 ± 8.9 | 0.27 ± 0.02 | [89] |
| Aegean Sea | Sparus aurata | 29.5 ± 0.8 | 0.037 ± 0.022 | [90] |
| Dicentrarchus labrax | 34.4 ± 1.4 | 0.034 ± 0.016 | ||
| Global | Order | No. locations | Total Hg | [24] |
| Clupeiformes | 142 | 0.06 ± 0.18 | ||
| Perciformes | 1430 | 0.37 ± 0.48 | ||
| Pleuronectiformes | 112 | 0.26 ± 0.28 | ||
| Gadiformes | 85 | 0.12 ± 0.14 | ||
| Mugiliformes | 91 | 0.08 ± 0.15 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Bargagli, R.; Rota, E. Mercury Biogeochemistry and Biomagnification in the Mediterranean Sea: Current Knowledge and Future Prospects in the Context of Climate Change. Coasts 2024, 4, 89-107. https://doi.org/10.3390/coasts4010006
AMA Style
Bargagli R, Rota E. Mercury Biogeochemistry and Biomagnification in the Mediterranean Sea: Current Knowledge and Future Prospects in the Context of Climate Change. Coasts. 2024; 4(1):89-107. https://doi.org/10.3390/coasts4010006
Chicago/Turabian StyleBargagli, Roberto, and Emilia Rota. 2024. "Mercury Biogeochemistry and Biomagnification in the Mediterranean Sea: Current Knowledge and Future Prospects in the Context of Climate Change" Coasts 4, no. 1: 89-107. https://doi.org/10.3390/coasts4010006