A Review of Mercury Bioavailability in Humans and Fish

Abstract
:1. Introduction
(“mercury” OR “* Hg” OR “methylmercury” OR “* MeHg” OR “CH3Hg”) AND (“assimil * efficiency” OR “intestine * transfer” OR “gut” OR “uptake pathway”) AND “fish”.
“203Hg” AND (“accumulation” OR “diet”) AND “fish”.
(“mercury” OR “* Hg” OR “methylmercury” OR “* MeHg” OR “CH3Hg”) AND (“bioavailab *” OR “bioaccess *” OR “uptake” OR “transport” OR “assimilate *” OR “absor *”) AND “human”.
(“mercury” OR “* Hg” OR “methylmercury” OR “* MeHg” OR “CH3Hg”) AND (“bioavailab *” OR “bioaccess *” OR “uptake” OR “transport” OR “assimilate *” OR “absor *”) AND “human”.
(“mercury” OR “Methylmercury”) AND (“bioaccessibility” OR “bioavailability” OR “Caco-2”) AND (“fish” OR “seafood” OR “shellfish”).
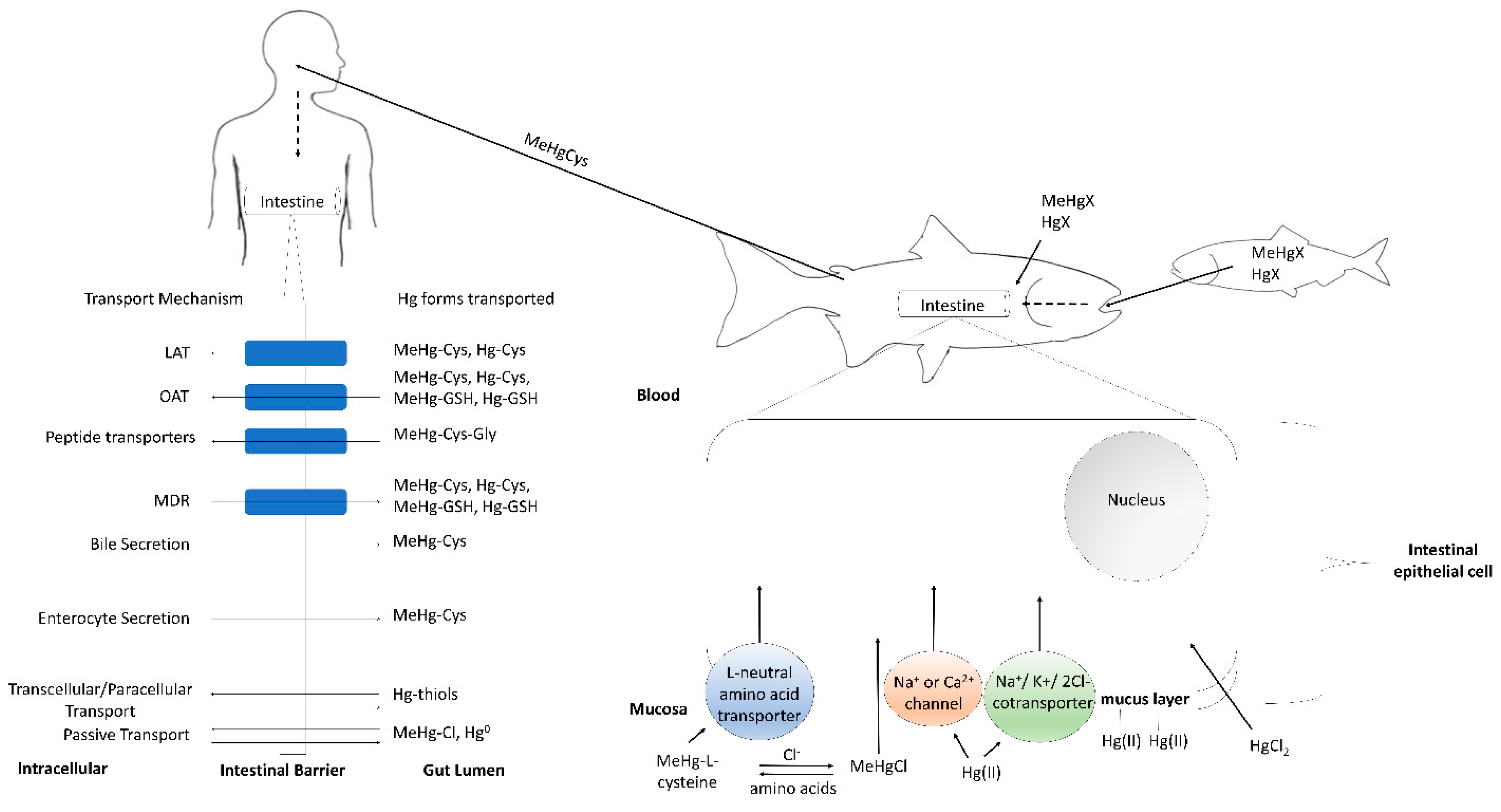
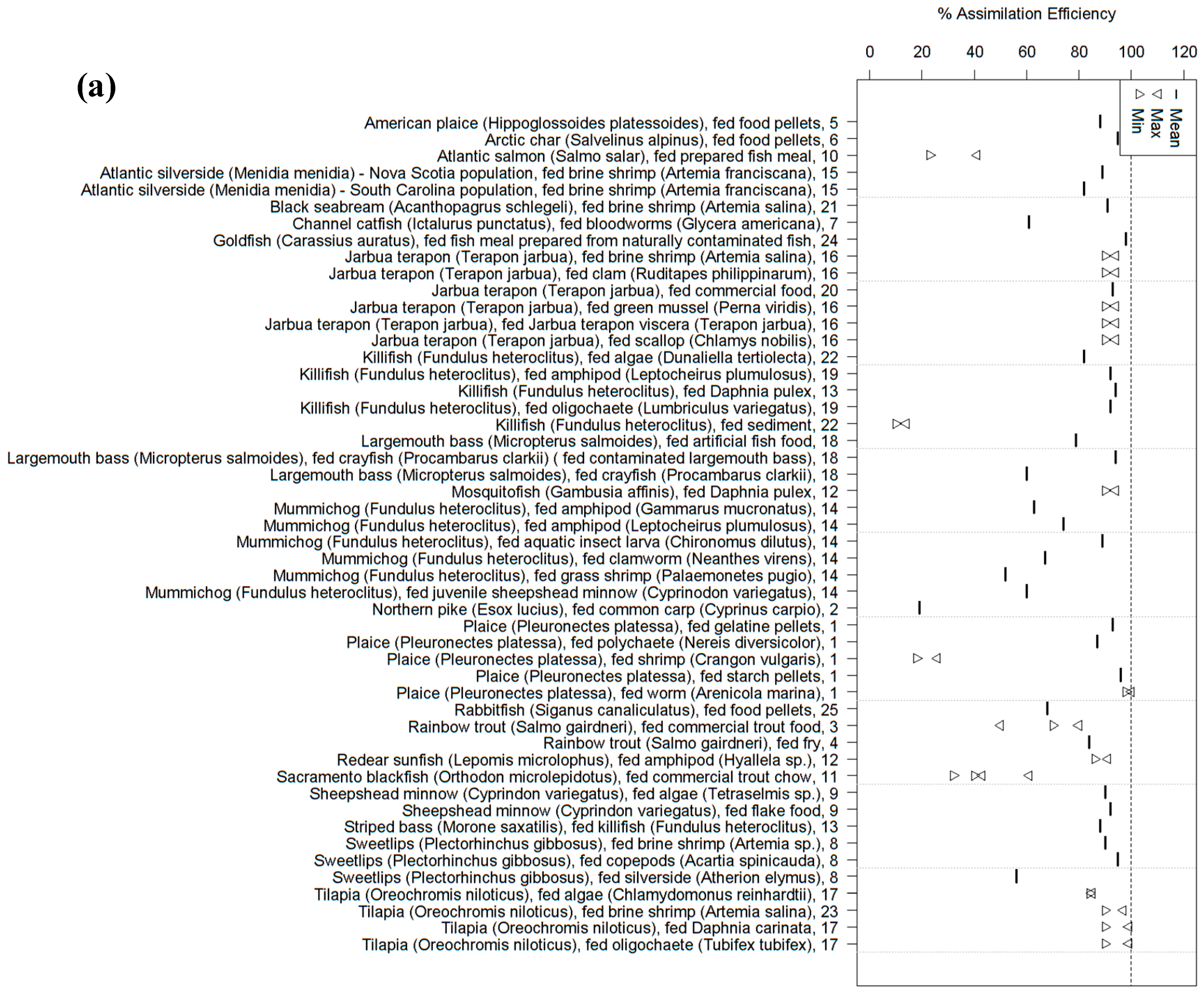
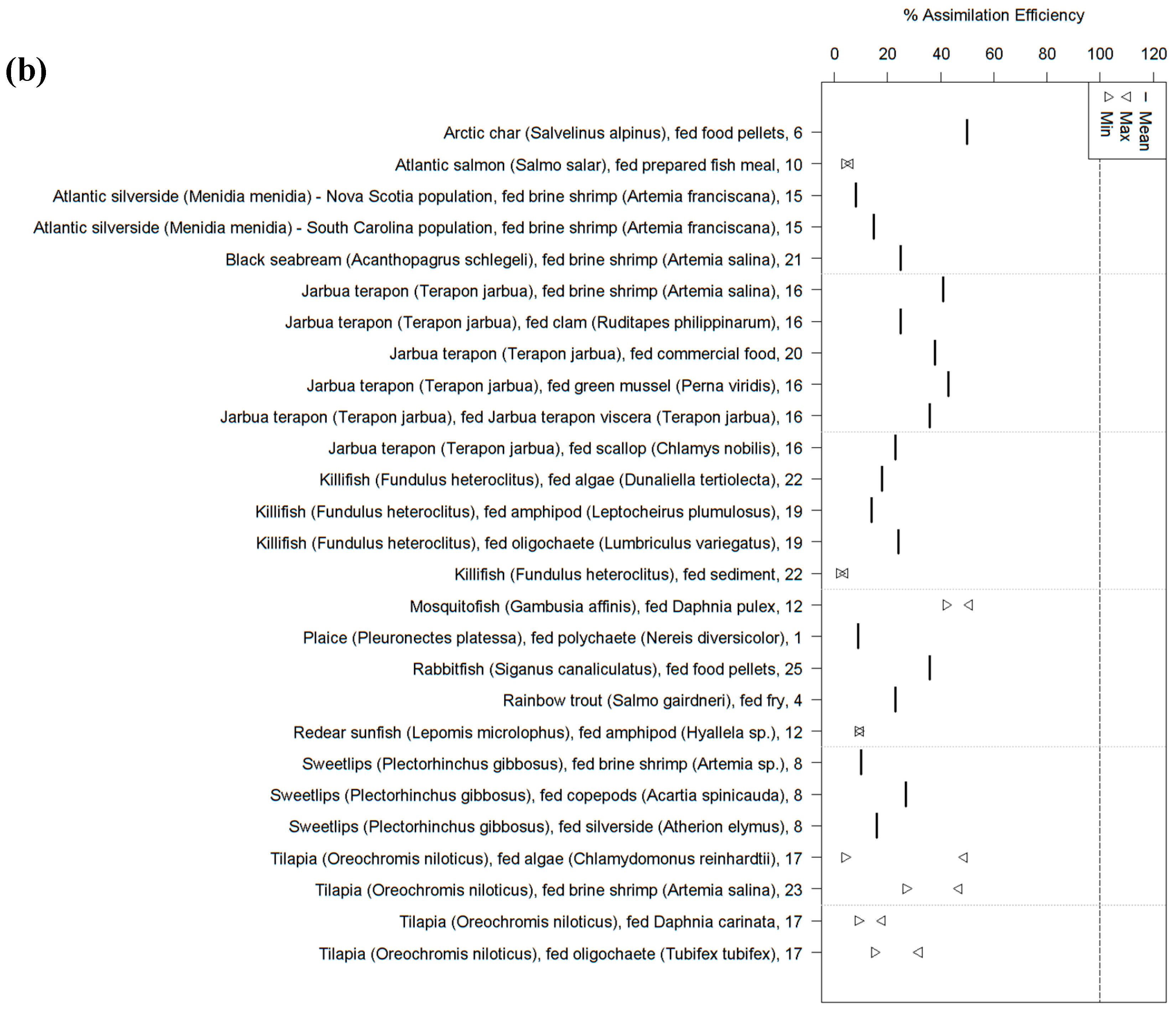
2. Fish
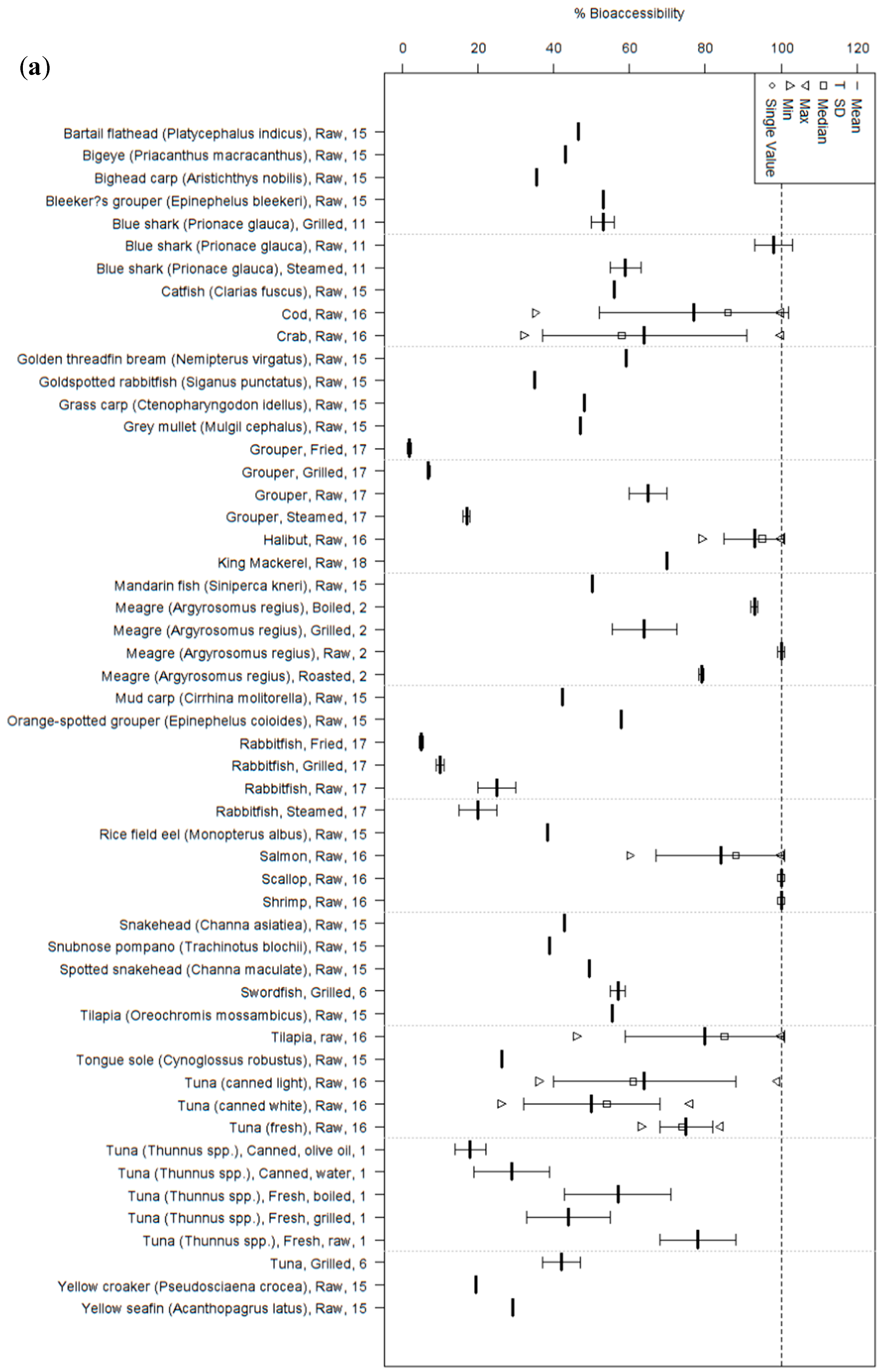
3. Humans
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Ha, E.; Basu, N.; Bose-O’Reilly, S.; Dorea, J.G.; McSorley, E.; Sakamoto, M.; Chan, H.M. Current progress on understanding the impact of mercury on human health. Environ. Res. 2016, 152, 419–433. [Google Scholar] [CrossRef] [PubMed]
- Scheuhammer, A.M.; Basu, N.; Evers, D.C.; Heinz, G.H.; Sandheinrich, M.; Bank, M.S. Toxicology of mercury in fish and wildlife: Recent advances. In Mercury in the Environment: Pattern and Process; Bank, M.S., Ed.; University of California Press: Berkeley, CA, USA, 2012. [Google Scholar]
- Grandjean, P.; Weihe, P.; Jørgensen, P.; Clarkson, T.; Cernichiari, E.; Viderø, T. Impact of maternal seafood diet on fetal exposure to mercury, selenium, and lead. Arch. Environ. Health Int. J. 1992, 47, 185–195. [Google Scholar] [CrossRef] [PubMed]
- Julshamn, K.; Andersen, A.; Ringdal, O.; Mørkøre, J. Trace elements intake in the Faroe Islands I. Element levels in edible parts of pilot whales (Globicephalus meleanus). Sci. Total Environ. 1987, 65, 53–62. [Google Scholar] [CrossRef]
- Zhang, H.; Feng, X.; Larssen, T.; Qiu, G.; Vogt, R.D. In inland China, rice, rather than fish, is the major pathway for methylmercury exposure. Environ. Health Perspect. 2010, 118, 1183–1188. [Google Scholar] [CrossRef] [PubMed]
- Mergler, D.; Anderson, H.A.; Chan, L.H.M.; Mahaffey, K.R.; Murray, M.; Sakamoto, M.; Stern, A.H. Methylmercury exposure and health effects in humans: A worldwide concern. J. Hum. Environ. 2007, 36, 3–11. [Google Scholar] [CrossRef]
- Sheehan, M.C.; Burke, T.A.; Navas-Acien, A.; Breysse, P.N.; McGready, J.; Fox, M.A. Global methylmercury exposure from seafood consumption and risk of developmental neurotoxicity: A systematic review. Bull. World Health Organ. 2014, 92, 254–269. [Google Scholar] [CrossRef] [PubMed]
- Karagas, M.R.; Choi, A.L.; Oken, E.; Horvat, M.; Schoeny, R.; Kamai, E.; Cowell, W.; Grandjean, P.; Korrick, S. Evidence on the human health effects of low-level methylmercury exposure. Environ. Health Perspect. 2012, 120, 799–806. [Google Scholar] [CrossRef] [Green Version]
- Mozaffarian, D.; Rimm, E.B. Fish intake, contaminants, and human health: Evaluating the risks and the benefits. JAMA 2006, 296, 1885–1899. [Google Scholar] [CrossRef] [PubMed]
- Swain, E.B.; Jakus, P.M.; Rice, G.; Lupi, F.; Maxson, P.A.; Pacyna, J.M.; Penn, A.; Spiegel, S.J.; Veiga, M.M. Socioeconomic consequences of mercury use and pollution. J. Hum. Environ. 2007, 36, 45–61. [Google Scholar] [CrossRef]
- Mahaffey, K.R.; Rice, G.E.; Swartout, J. Mercury Study Report to Congress Volume IV: An Assessment of Exposure to Mercury in the United States. Available online: http://www.epa.gov/mercury/report.htm (accessed on 29 November 2016).
- Di Giulio, R.T.; Hinton, D.E. The Toxicology of Fishes; CRC Press: Boca Raton, FL, USA, 2008. [Google Scholar]
- Versantvoort, C.H.M.; van de Kamp, E.; Rompelberg, C.J.M. Development and Applicability of an In Vitro Digestion Model in Assessing the Bioaccessibility of Contaminants from Food. Available online: http://rivm.openrepository.com/rivm/bitstream/10029/18885/10021/320102002.pdf (accessed on 29 November 2016).
- Lemes, M.; Wang, F. Methylmercury speciation in fish muscle by HPLC-ICP-MS following enzymatic hydrolysis. J. Anal. At. Spectrom. 2009, 24, 663–668. [Google Scholar] [CrossRef]
- Leaner, J.J.; Mason, R.P. Methylmercury accumulation and fluxes across the intestine of channel catfish, Ictalurus punctatus. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2002, 132, 247–259. [Google Scholar] [CrossRef]
- Hoyle, I.; Handy, R. Dose-dependent inorganic mercury absorption by isolated perfused intestine of rainbow trout, Oncorhynchus mykiss, involves both amiloride-sensitive and energy-dependent pathways. Aquat. Toxicol. 2005, 72, 147–159. [Google Scholar] [CrossRef] [PubMed]
- Pärt, P.; Lock, R. Diffusion of calcium, cadmium and mercury in a mucous solution from rainbow trout. Comp. Biochem. Physiol. C Comp. Pharmacol. 1983, 76, 259–263. [Google Scholar] [CrossRef]
- Bridges, C.C.; Zalups, R.K. Transport of inorganic mercury and methylmercury in target tissues and organs. J. Toxicol. Environ. Health 2010, 13, 385–410. [Google Scholar] [CrossRef] [PubMed]
- Hall, B.; Bodaly, R.; Fudge, R.; Rudd, J.; Rosenberg, D. Food as the dominant pathway of methylmercury uptake by fish. Water, Air Soil Pollut. 1997, 100, 13–24. [Google Scholar]
- Hrenchuk, L.E.; Blanchfield, P.J.; Paterson, M.J.; Hintelmann, H.H. Dietary and waterborne mercury accumulation by yellow perch: A field experiment. Environ. Sci. Technol. 2011, 46, 509–516. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Wong, M.-H.; Wang, W.-X. Mercury exposure in the freshwater tilapia Oreochromis niloticus. Environ. Pollut. 2010, 158, 2694–2701. [Google Scholar] [CrossRef] [PubMed]
- Pickhardt, P.C.; Stepanova, M.; Fisher, N.S. Contrasting uptake routes and tissue distributions of inorganic and methylmercury in mosquitofish (Gambusia affinis) and redear sunfish (Lepomis microlophus). Environ. Toxicol. Chem. 2006, 25, 2132–2142. [Google Scholar] [CrossRef]
- Wang, R.; Wang, W.-X. Importance of speciation in understanding mercury bioaccumulation in tilapia controlled by salinity and dissolved organic matter. Environ. Sci. Technol. 2010, 44, 7964–7969. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Lemes, M.; Khan, M. Metallomics of mercury: Role of thiol-and selenol-containing biomolecules. In Environmental Chemistry and Toxicology of Mercury; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2012; pp. 517–544. [Google Scholar]
- Kidd, K.; Batchelar, K. 5—Mercury. In Fish Physiology; Wood, C.M., Farrell, A.P., Colin, J.B., Eds.; Academic Press: Cambridge, MA, USA, 2011; Volume 31, pp. 237–295. [Google Scholar]
- Boudou, A.; Ribeyre, F. Experimental study of trophic contamination of Salmo gairdneri by two mercury compounds—HgCl2 and CH3HgCl—Analysis at the organism and organ levels. Water Air Soil Pollut. 1985, 26, 137–148. [Google Scholar] [CrossRef]
- Wang, W.-X.; Wong, R.S. Bioaccumulation kinetics and exposure pathways of inorganic mercury and methylmercury in a marine fish, the sweetlips Plectorhinchus gibbosus. Mar. Ecol. Prog. Ser. 2003, 261, 257–268. [Google Scholar] [CrossRef]
- Goto, D.; Wallace, W.G. Influences of prey-and predator-dependent processes on cadmium and methylmercury trophic transfer to mummichogs (Fundulus heteroclitus). Can. J. Fish. Aquat. Sci. 2009, 66, 836–846. [Google Scholar]
- Dutton, J.; Fisher, N.S. Bioavailability of sediment-bound and algal metals to killifish Fundulus heteroclitus. Aquat. Biol. 2012, 16, 85–96. [Google Scholar] [CrossRef]
- Houck, A.; Cech, J.J. Effects of dietary methylmercury on juvenile Sacramento blackfish bioenergetics. Aquat. Toxicol. 2004, 69, 107–123. [Google Scholar] [CrossRef] [PubMed]
- Rodgers, D.; Beamish, F. Dynamics of dietary methylmercury in rainbow trout, Salmo gairdneri. Aquat. Toxicol. 1982, 2, 271–290. [Google Scholar] [CrossRef]
- Li, J.; Drouillard, K.G.; Branfireun, B.; Haffner, G.D. Comparison of the toxicokinetics and bioaccumulation potential of mercury and polychlorinated biphenyls in goldfish (Carassius auratus). Environ. Sci. Technol. 2015, 49, 11019–11027. [Google Scholar] [CrossRef] [PubMed]
- Phillips, G.; Gregory, R. Assimilation efficiency of dietary methylmercury by northern pike (Esox lucius). J. Fish. Board Can. 1979, 36, 1516–1519. [Google Scholar] [CrossRef]
- Oliveira Ribeiro, C.; Rouleau, C.; Pelletier, E.; Audet, C.; Tjälve, H. Distribution kinetics of dietary methylmercury in the arctic charr (Salvelinus alpinus). Environ. Sci. Technol. 1999, 33, 902–907. [Google Scholar] [CrossRef]
- Leaner, J.J.; Mason, R.P. Methylmercury uptake and distribution kinetics in sheepshead minnows, Cyprinodon variegatus, after exposure to CH3Hg-spiked food. Environ. Toxicol. Chem. 2004, 23, 2138–2146. [Google Scholar] [CrossRef]
- Giblin, F.; Massaro, E.J. The erythrocyte transport and transfer of methylmercury to the tissues of the rainbow trout (Salmo gairdneri). Toxicology 1975, 5, 243–254. [Google Scholar] [CrossRef]
- Berntssen, M.; Hylland, K.; Julshamn, K.; Lundebye, A.K.; Waagbø, R. Maximum limits of organic and inorganic mercury in fish feed. Aquac. Nutr. 2004, 10, 83–97. [Google Scholar] [CrossRef]
- Sandheinrich, M.; Wiener, J. Methylmercury in freshwater fish: Recent advances in assessing toxicity of environmentally relevant exposures. Environ. Contam. Biota Interpret. Tissue Conc. 2011, 2, 169–190. [Google Scholar]
- Bloom, N.S. On the chemical form of mercury in edible fish and marine invertebrate tissue. Can. J. Fish. Aquat. Sci. 1992, 49, 1010–1017. [Google Scholar] [CrossRef]
- Harris, H.H.; Pickering, I.J.; George, G.N. The chemical form of mercury in fish. Science 2003, 301, 1203. [Google Scholar] [CrossRef] [PubMed]
- Trudel, M.; Rasmussen, J.B. Modeling the elimination of mercury by fish. Environ. Sci. Technol. 1997, 31, 1716–1722. [Google Scholar] [CrossRef]
- Van Walleghem, J.L.; Blanchfield, P.J.; Hintelmann, H. Elimination of mercury by yellow perch in the wild. Environ. Sci. Technol. 2007, 41, 5895–5901. [Google Scholar] [CrossRef] [PubMed]
- Van Walleghem, J.L.; Blanchfield, P.J.; Hrenchuk, L.E.; Hintelmann, H. Mercury elimination by a top predator, Esox lucius. Environ. Sci. Technol. 2013, 47, 4147–4154. [Google Scholar] [CrossRef] [PubMed]
- Aberg, B.; Ekman, L.; Falk, R.; Greitz, U.; Persson, G.; Snihs, J. Metabolism of methyl mercury (203Hg) compounds in man. Arch. Environ. Health 1969, 19, 478–484. [Google Scholar] [CrossRef] [PubMed]
- Miettinen, J.K.; Rahola, T.; Hattula, T.; Rissanen, K.; Tillander, M. Elimination of 203Hg-methylmercury in man. Ann. Clin. Res. 1971, 3, 116–122. [Google Scholar] [PubMed]
- Government of Canada. What Is A Food Guide Serving of Meat and Alternatives?—Canada’s Food Guide. Available online: http://www.hc-sc.gc.ca/fn-an/food-guide-aliment/choose-choix/meat-viande/serving-portion-eng.php (accessed on 29 November 2016).
- U.S. Food and Drug Administration. CFR—Code of Federal Regulations Title 21. Available online: https://www.accessdata.fda.gov/scripts/cdrh/cfdocs/cfcfr/cfrsearch.cfm?fr=101.12 (accessed on 29 November 2016).
- Laparra, J.M.; Velez, D.; Montoro, R.; Barbera, R.; Farre, R. Estimation of arsenic bioavailability in edible seaweed by an in vitro digestion method. J. Agric. Food Chem. 2003, 51, 6080–6085. [Google Scholar] [CrossRef] [PubMed]
- Costa, S.; Afonso, C.; Cardoso, C.; Batista, I.; Chaveiro, N.; Nunes, M.L.; Bandarra, N.M. Fatty acids, mercury, and methylmercury bioaccessibility in salmon (Salmo salar) using an in vitro model: Effect of culinary treatment. Food Chem. 2015, 185, 268–276. [Google Scholar] [CrossRef] [PubMed]
- Calatayud, M.; Devesa, V.; Virseda, J.R.; Barbera, R.; Montoro, R.; Velez, D. Mercury and selenium in fish and shellfish: Occurrence, bioaccessibility and uptake by Caco-2 cells. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2012, 50, 2696–2702. [Google Scholar] [CrossRef] [PubMed]
- Siedlikowski, M.; Bradley, M.; Kubow, S.; Goodrich, J.M.; Franzblau, A.; Basu, N. Bioaccessibility and bioavailability of methylmercury from seafood commonly consumed in North America: In vitro and epidemiological studies. Environ. Res. 2016, 149, 266–273. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.S.; Xu, W.F.; Chen, Z.J.; Cheng, Z.; Ge, L.C.; Man, Y.B.; Giesy, J.P.; Du, J.; Wong, C.K.; Wong, M.H. In vitro estimation of exposure of Hong Kong residents to mercury and methylmercury via consumption of market fishes. J. Hazard. Mater. 2013, 248–249, 387–393. [Google Scholar] [CrossRef] [PubMed]
- He, M.; Wang, W.X. Factors affecting the bioaccessibility of methylmercury in several marine fish species. J. Agric. Food Chem. 2011, 59, 7155–7162. [Google Scholar] [CrossRef] [PubMed]
- Ouedraogo, O.; Amyot, M. Effects of various cooking methods and food components on bioaccessibility of mercury from fish. Environ. Res. 2011, 111, 1064–1069. [Google Scholar] [CrossRef] [PubMed]
- Costa, S.; Afonso, C.; Bandarra, N.M.; Gueifao, S.; Castanheira, I.; Carvalho, M.L.; Cardoso, C.; Nunes, M.L. The emerging farmed fish species meagre (Argyrosomus regius): How culinary treatment affects nutrients and contaminants concentration and associated benefit-risk balance. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2013, 60, 277–285. [Google Scholar] [CrossRef] [PubMed]
- Burger, J.; Dixon, C.; Boring, S.; Gochfeld, M. Effect of deep-frying fish on risk from mercury. J. Toxicol. Environ. Health 2003, 66, 817–828. [Google Scholar] [CrossRef] [PubMed]
- Torres-Escribano, S.; Ruiz, A.; Barrios, L.; Vélez, D.; Montoro, R. Influence of mercury bioaccessibility on exposure assessment associated with consumption of cooked predatory fish in Spain. J. Sci. Food Agric. 2011, 91, 981–986. [Google Scholar] [CrossRef] [PubMed]
- Perelló, G.; Martí-Cid, R.; Llobet, J.M.; Domingo, J.L. Effects of various cooking processes on the concentrations of arsenic, cadmium, mercury, and lead in foods. J. Agric. Food Chem. 2008, 56, 11262–11269. [Google Scholar] [CrossRef] [PubMed]
- Armbruster, G.; Gutenmann, W.H.; Lisk, D.J. The effects of six methods of cooking on residues of mercury in striped bass. Nutr. Rep. Int. 1988, 37, 123–126. [Google Scholar]
- Gutenmann, W.H.; Lisk, D.J. Higher average mercury concentrations in fish fillets after skinning and fat removal. J. Food Saf. 1991, 11, 99–103. [Google Scholar] [CrossRef]
- Blanco, S.L.; Gonzalez, J.C.; Vieites, J.M. Mercury, cadmium and lead levels in samples of the main traded fish and shellfish species in Galicia, Spain. Food Addit. Contam. 2008, 1, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.H.; Teng, P.Y.; Chen, C.Y.; Hsu, C.C. Organic and total mercury levels in bigeye tuna, Thunnus obesus, harvested by Taiwanese fishing vessels in the Atlantic and Indian Oceans. Food Addit. Contam. 2011, 4, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Groth, E. Ranking the contributions of commercial fish and shellfish varieties to mercury exposure in the United States: Implications for risk communication. Environ. Res. 2010, 110, 226–236. [Google Scholar] [CrossRef] [PubMed]
- Moon, H.B.; Kim, S.J.; Park, H.; Jung, Y.S.; Lee, S.; Kim, Y.H.; Choi, M. Exposure assessment for methyl and total mercury from seafood consumption in Korea, 2005 to 2008. J. Environ. Monit. 2011, 13, 2400–2405. [Google Scholar] [CrossRef] [PubMed]
- Tran, N.L.; Barraj, L.M.; Bi, X.; Schuda, L.C.; Moya, J. Estimated long-term fish and shellfish intake—National Health and Nutrition Examination Survey. J. Expo. Sci. Environ. Epidemiol. 2013, 23, 128–136. [Google Scholar] [CrossRef] [PubMed]
- Afonso, C.; Costa, S.; Cardoso, C.; Oliveira, R.; Lourenco, H.M.; Viula, A.; Batista, I.; Coelho, I.; Nunes, M.L. Benefits and risks associated with consumption of raw, cooked, and canned tuna (Thunnus spp.) based on the bioaccessibility of selenium and methylmercury. Environ. Res. 2015, 143, 130–137. [Google Scholar] [CrossRef] [PubMed]
- George, G.N.; Singh, S.P.; Prince, R.C.; Pickering, I.J. Chemical forms of mercury and selenium in fish following digestion with simulated gastric fluid. Chem. Res. Toxicol. 2008, 21, 2106–2110. [Google Scholar] [CrossRef] [PubMed]
- Dunn, J.D.; Clarkson, T.W.; Magos, L. Ethanol reveals novel mercury detoxification step in tissues. Science 1981, 213, 1123–1125. [Google Scholar] [CrossRef] [PubMed]
- Chapman, L.; Chan, H.M. The influence of nutrition on methyl mercury intoxication. Environ. Health Perspect. 2000, 108, 29–56. [Google Scholar] [CrossRef] [PubMed]
- Shim, S.M.; Ferruzzi, M.G.; Kim, Y.C.; Janle, E.M.; Santerre, C.R. Impact of phytochemical-rich foods on bioaccessibility of mercury from fish. Food Chem. 2009, 112, 46–50. [Google Scholar] [CrossRef]
- Passos, C.J.; Mergler, D.; Gaspar, E.; Morais, S.; Lucotte, M.; Larribe, F.; Davidson, R.; de Grosbois, S. Eating tropical fruit reduces mercury exposure from fish consumption in the Brazilian Amazon. Environ. Res. 2003, 93, 123–130. [Google Scholar] [CrossRef]
- Laird, B.D.; Shade, C.; Gantner, N.; Chan, H.M.; Siciliano, S.D. Bioaccessibility of mercury from traditional northern country foods measured using an in vitro gastrointestinal model is independent of mercury concentration. Sci. Total Environ. 2009, 407, 6003–6008. [Google Scholar] [CrossRef] [PubMed]
- Laird, B.D.; Chan, H.M. Bioaccessibility of metals in fish, shellfish, wild game, and seaweed harvested in British Columbia, Canada. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2013, 58, 381–387. [Google Scholar] [CrossRef] [PubMed]
- Aduayom, I.; Campbell, P.G.C.; Denizeau, F.; Jumarie, C. Different transport mechanisms for cadmium and mercury in Caco-2 cells: Inhibition of Cd uptake by Hg without evidence for reciprocal effects. Toxicol. Appl. Pharm. 2003, 189, 56–67. [Google Scholar] [CrossRef]
- Rostami-Hodjegan, A. Physiologically based pharmacokinetics joined with in vitro-in vivo extrapolation of ADME: A marriage under the arch of systems pharmacology. Clin. Pharmacol. Ther. 2012, 92, 50–61. [Google Scholar] [CrossRef] [PubMed]
- Berntssen, M.H.; Hylland, K.; Lundebye, A.K.; Julshamn, K. Higher faecal excretion and lower tissue accumulation of mercury in Wistar rats from contaminated fish than from methylmercury chloride added to fish. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2004, 42, 1359–1366. [Google Scholar] [CrossRef] [PubMed]
- Sherman, L.S.; Blum, J.D.; Franzblau, A.; Basu, N. New insight into biomarkers of human mercury exposure using naturally occurring mercury stable isotopes. Environ. Sci. Technol. 2013, 47, 3403–3409. [Google Scholar] [CrossRef] [PubMed]
- Hur, S.J.; Lim, B.O.; Decker, E.A.; McClements, D.J. In vitro human digestion models for food applications. Food Chem. 2011, 125, 1–12. [Google Scholar] [CrossRef]
- Laird, B.D.; Weiseth, B.; Packull-McCormick, S.R.; Peak, D.; Dodd, M.; Siciliano, S.D. Solid-liquid separation method governs the in vitro bioaccessibility of metals in contaminated soil-like test materials. Chemosphere 2015, 134, 544–549. [Google Scholar] [CrossRef] [PubMed]
- Rowland, I.R.; Davies, M.J.; Evans, J.G. Tissue content of mercury in rats given methylmercuric chloride orally: Influence of intestinal flora. Arch. Environ. Health 1980, 35, 155–160. [Google Scholar] [CrossRef] [PubMed]
- Rowland, I.R.; Robinson, R.D.; Doherty, R.A. Effects of diet on mercury metabolism and excretion in mice given methylmercury: Role of gut flora. Arch. Environ. Health 1984, 39, 401–408. [Google Scholar] [CrossRef] [PubMed]
- Basu, N.; Goodrich, J.M.; Head, J. Ecogenetics of mercury: From genetic polymorphisms and epigenetics to risk assessment and decision-making. Environ. Toxicol. Chem. 2014, 33, 1248–1258. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
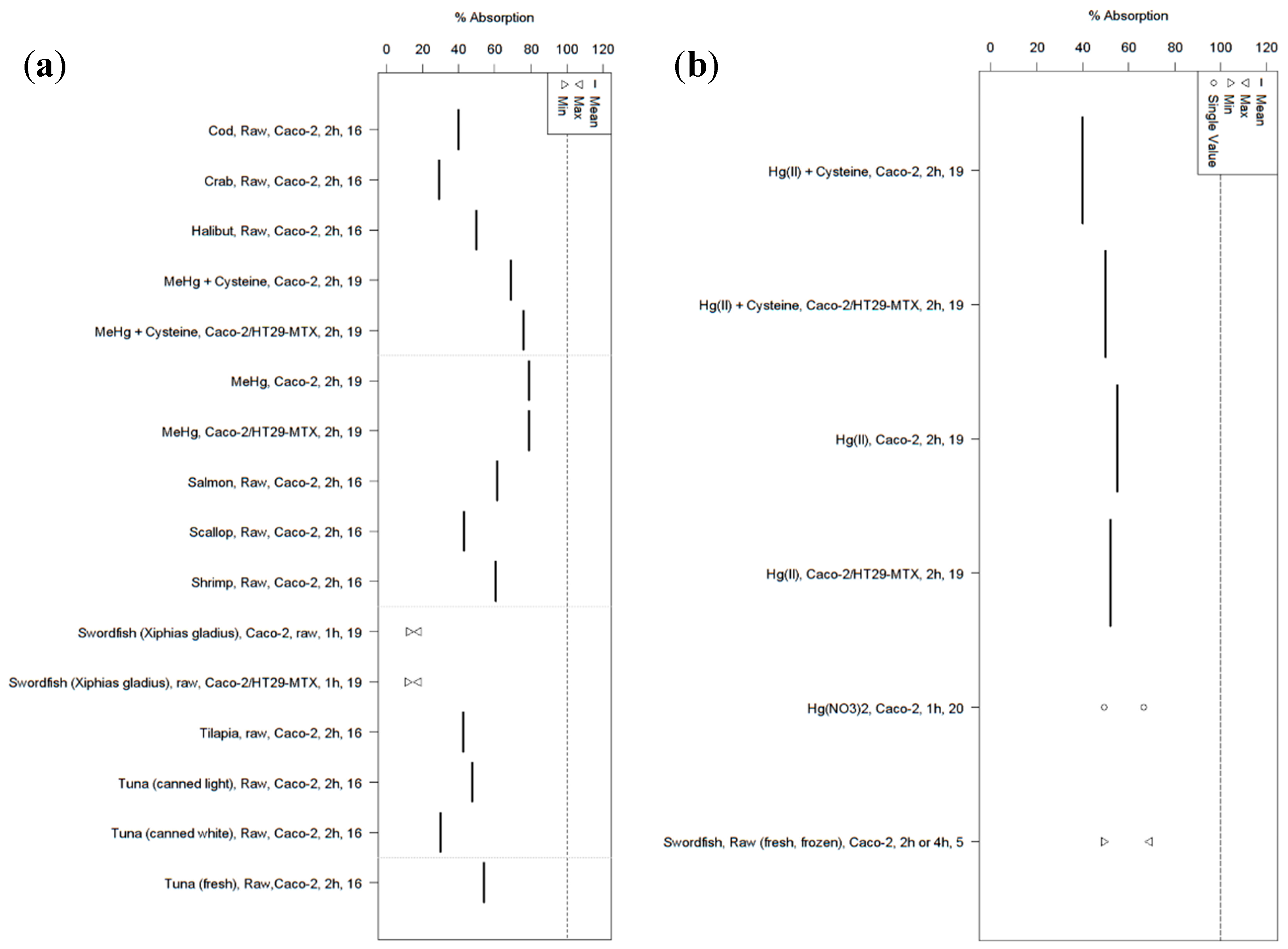{kind=link}
| Fish Species | Cooking Treatment | Hg Concentration Effect | Weight Measurement | Hg Bioaccessibility Effect | Reference |
|---|---|---|---|---|---|
| Meagre (farmed; Argyrosomus regius) | Grilled | increased total Hg 33%; increased MeHg 25% | unclear; probably wet weight | Not studied | [55] |
| Boiled | no change | unclear; probably wet weight | Not studied | ||
| Roasted | increased total Hg 19%; increased MeHg 19% | unclear; probably wet weight | Not studied | ||
| Largemouth bass (Micropterus salmoides) | Deep fried (with breading) | increased total Hg 45% | wet weight | Not studied | [56] |
| Deep fried (without breading) | increased total Hg 75% | wet weight | Not studied | ||
| Spanish mackerel (Scomberomorus macalatus), cat shark (Scyliorhinus sp.), red tuna (Thunnus thynnus) | Fried | no change | dry weight | total Hg: 65% lower; MeHg: 85% lower | [54] |
| Boiled | no change | dry weight | total Hg: 38% lower; MeHg: 54% lower | ||
| Fresh swordfish (Xiphias gladius), frozen tope shark (Galeorhinus galeus), frozen bonito (Sarda sp.), fresh tuna (Thunnus sp.) | Hot plate/griddle | total Hg increased: swordfish (43%), tuna (32%), tope shark (22%), bonito (20%) | wet weight | total Hg in raw fish: 42% (13%–87%); total Hg in cooked fish: 26% (6%–49%) | [57] |
| Hot plate/griddle | no change | dry weight | Not studied | ||
| Sardine, hake, tuna | Fried | no change | wet weight | Not studied | [58] |
| Grilled | no change | wet weight | Not studied | ||
| Hake | Roasted | no change | wet weight | Not studied | |
| Boiled | no change | wet weight | Not studied | ||
| Striped bass | Baked | no change | dry weight | Not studied | [59] |
| Broiled | no change | dry weight | Not studied | ||
| Fried | no change | dry weight | Not studied | ||
| Microwaved | no change | dry weight | Not studied | ||
| Poached | no change | dry weight | Not studied | ||
| Steamed | no change | dry weight | Not studied | ||
| Brown trout (Salmo trutta) | Skinned, trimmed fat | increased total Hg 25% (male fish), 32% (female fish) | dry weight | Not studied | [60] |
| Food or Nutrient | Hg Exposure | Model | Description of Interaction | Reference |
|---|---|---|---|---|
| Tea, coffee | 1–4 μg/g dry weight Hg in fish (tuna, shark, mackerel) | in vitro digestion—bioaccessibility | Tea and coffee reduced total Hg bioaccessibility 10%–60%, depending on species | [54] |
| Corn starch | 1–4 μg/g dry weight Hg in fish (tuna, shark, mackerel) | in vitro digestion—bioaccessibility | Corn starch reduced total Hg bioaccessibility by 20% (tuna only) | |
| Tropical Fruits | Hg in fish meals | Human, prospective study | Consumption of 1 fruit/day was associated with lower hair Hg than consumption of <1 fruit/day | [71] |
| Green tea extract | fish tissue | in vitro digestion—bioaccessibility | reduced Hg bioaccessibility by 82%–92% | [70] |
| Black tea extract | fish tissue | in vitro digestion—bioaccessibility | reduced Hg bioaccessibility by 88%–91% | |
| Soy protein | fish tissue | in vitro digestion—bioaccessibility | reduced Hg bioaccessibility by 44%–87% | |
| Grapefruit juice | fish tissue | in vitro digestion—bioaccessibility | no reduction of bioaccessible Hg | |
| Wheat bran | fish tissue | in vitro digestion—bioaccessibility | reduced Hg bioaccessibility by 84% | |
| OAT bran | fish tissue | in vitro digestion—bioaccessibility | reduced Hg bioaccessibility by 59%–85% | |
| Psyllium | fish tissue | in vitro digestion—bioaccessibility | reduced Hg bioaccessibility by 15%–31% |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bradley, M.A.; Barst, B.D.; Basu, N. A Review of Mercury Bioavailability in Humans and Fish. Int. J. Environ. Res. Public Health 2017, 14, 169. https://doi.org/10.3390/ijerph14020169
Bradley MA, Barst BD, Basu N. A Review of Mercury Bioavailability in Humans and Fish. International Journal of Environmental Research and Public Health. 2017; 14(2):169. https://doi.org/10.3390/ijerph14020169
Chicago/Turabian StyleBradley, Mark A., Benjamin D. Barst, and Niladri Basu. 2017. "A Review of Mercury Bioavailability in Humans and Fish" International Journal of Environmental Research and Public Health 14, no. 2: 169. https://doi.org/10.3390/ijerph14020169