

Insights into the Function of Aquaporins in Gastrointestinal Fluid Absorption and Secretion in Health and Disease


1
Department of Biosciences, Biotechnologies and Environment, University of Bari Aldo Moro, 70125 Bari, Italy
2
Laboratory of Pathophysiological and Nutritional Biochemistry, Faculty of Medicine, Université Libre de Bruxelles, 1070 Brussels, Belgium
*
Author to whom correspondence should be addressed.
Cells 2023, 12(17), 2170; https://doi.org/10.3390/cells12172170
Submission received: 1 July 2023
/
Revised: 16 August 2023
/
Accepted: 17 August 2023
/
Published: 29 August 2023
(This article belongs to the Special Issue Advances in Aquaporins II)
Abstract
:Aquaporins (AQPs), transmembrane proteins permeable to water, are involved in gastrointestinal secretion. The secretory products of the glands are delivered either to some organ cavities for exocrine glands or to the bloodstream for endocrine glands. The main secretory glands being part of the gastrointestinal system are salivary glands, gastric glands, duodenal Brunner’s gland, liver, bile ducts, gallbladder, intestinal goblet cells, exocrine and endocrine pancreas. Due to their expression in gastrointestinal exocrine and endocrine glands, AQPs fulfill important roles in the secretion of various fluids involved in food handling. This review summarizes the contribution of AQPs in physiological and pathophysiological stages related to gastrointestinal secretion.
1. Introduction
Aquaporins (AQPs) represent a family of proteins expressed in all living organisms that contain six transmembrane domains with both N- and C-terminal ends located intracellularly [1]. AQPs have a tetrameric organization conferring their functionality [1,2]. The mammalian family of AQPs is made of thirteen members (AQP0 to AQP12) [1]. Some AQPs are mostly permeable to water (AQP0, AQP1, AQP2, AQP4, AQP5, AQP6, AQP8), while others are also permeable to glycerol (AQP3, AQP7, AQP9, AQP10, AQP11) [1,3] or urea (AQP3, AQP6, AQP7, AQP9, AQP10) [4]. The permeability of one AQP still remains to be specified (AQP12) [4]. Some AQPs also ensure the transport of gas, such as carbon dioxide (AQP0, AQP1, AQP4, AQP5, AQP6, AQP9), nitric oxide (AQP1, AQP4), ammonia (AQP1, AQP3, AQP6, AQP7, AQP8, AQP9) and oxygen (AQP1) [4]. In addition, some AQPs can facilitate the movement of hydrogen peroxide (peroxiporins: AQP1, AQP3, AQP5, AQP8, AQP11), some ions (AQP0, AQP1, AQP6), silicon (AQP3, AQP7 AQP9, AQP10), antimonite and arsenite (AQP7, AQP9) [4].
AQPs are distributed throughout the entire body and fulfill a wide range of physiological functions [1,5]. AQPs are expressed in exocrine and endocrine glands to ensure glandular secretion [6]. This review summarizes the involvement of AQPs in the physiology of glandular secretion ensured by the main exocrine and endocrine glands distributed along the gastrointestinal tract and being involved in food handling. Hence, a particular focus is put on the role of AQPs in glandular secretion ensured by salivary glands, stomach glands, duodenal Brunner’s glands, intestinal Goblet cells, bile-secreting glands and pancreas. Moreover, for each gland, we address the role of relevant AQPs in association with diseases and, when relevant, the new therapeutic options and perspectives to modulate AQPs function in some of the secretory glands.
2. Salivary Glands
Saliva secretion is ensured by major and minor salivary glands [7,8]. Major salivary glands, present in pairs, include parotid glands located in front of and below each ear, submandibular glands located below the jaw, and sublingual glands located under the tongue. Minor salivary glands line the upper aerodigestive tract mucosa and most of the oral mucosa. The salivary glands are organized into lobes made of lobules. Each lobule consists of clusters of acinar cells, forming the acini, and interconnected ductal cells, forming the ducts. The acini and some ducts are surrounded by myoepithelial cells facilitating saliva secretion upon contraction [9]. The serous, mucous and seromucous acini (made respectively of serous, mucous or a mixture of both serous and mucous acinar cells) contribute to the secretion of a watery and mucus-rich secretion [9]. The ductal system is made of intralobular (intercalated and striated), interlobular and interlobar (excretory) ducts.
2.1. Role of AQPs in Saliva Secretion
Saliva formation involves both acinar and ductal cells. At first, acini produce an isosmotic fluid rich in NaCl and water. Then, ducts ensure the reuptake of some of the NaCl and the bicarbonate secretion. These combined processes lead to the secretion of hypotonic saliva into the oral cavity [10,11]. AQP5, located at the apical membrane of acinar cells, ensures water movement to the lumen of the acini following sodium and chloride secretion creating an osmotic gradient, playing a major role in saliva secretion [12,13]. Indeed, Aqp5 knockout mice presented a 60% decrease in pilocarpine-stimulated saliva secretion, as well as more viscous and hypertonic saliva [12,13]. Furthermore, acinar cells from parotid and submandibular glands from these mice displayed decreased water permeability (65% and 77%, respectively) [13]. From these studies and others, it was inferred that AQP5 is responsible for transmembrane water movement across the apical plasma membrane of acinar cells [10,11,14,15]. It was suggested that molecular mechanism involving AQP5 allows transcellular and paracellular water movement in a collective manner [16,17]. Under physiological conditions, muscarinic stimulation of acinar cells leads to intracellular calcium increase triggering AQP5 translocation to the plasma membrane [18,19,20]. Concomitantly, serous acini from all human salivary glands [21,22] and from rat [22,23,24,25] and mouse [18,26,27] submandibular and parotid glands mainly express AQP5 at their apical plasma membrane. Figure 1 summarizes the role of AQP5 in saliva formation in acinar cells. AQP5 expression was also reported in rat and mouse ductal cells [18,25,28,29]. However, physiologically, this localization is difficult to explain since ductal cells are water impermeable [30]. Rats with an AQP5 point mutation displayed reduced AQP5 expression and saliva secretion [31]. However, to the best of our knowledge, no human AQP5 mutation has so far been linked to salivary gland dysfunction.
In addition to AQP5, other AQPs are present in salivary glands [32,33]. Indeed, AQP1 is expressed in mouse [34] and human [21,22,35,36,37] endothelial and myoepithelial cells and in rat endothelial cells [29,38,39,40,41]. In human salivary glands, AQP3 was localized at the basolateral membrane of serous and mucous acinar cells but not in ductal cells [21,36,37], and Aqp4, Aqp6 and Aqp7 mRNAs but not their proteins have also been detected [21,37]. In rats, the expression of both AQP3 and AQP4 remains a subject of debate [28,29,42,43]. In mice, AQP3, AQP4 and AQP8 are expressed at the basolateral membrane of acinar and ductal cells [34]. Rat parotid glands expressed AQP6 in secretory granule membrane [44]. In mouse salivary glands, AQP7 was detected in the endothelial cells [27]. AQP8 is expressed in salivary gland myoepithelial cells [45,46,47]. In mouse salivary glands, AQP9 distribution remains to be fully assessed [26,27,33] and AQP11 was detected in ductal cells [26,27]. Moreover, different profiles of AQPs expression have been documented during the development of salivary glands in mice, rats and humans [29,48,49,50,51]. The employment of Aqp1, Aqp4 and Aqp8 knockout mice has ruled out their physiological involvement in saliva secretion [12,52,53].
2.2. Role of AQPs in Diseases Affecting Salivary Gland Function
Abnormal AQP5 expression and/or distribution has been associated with some pathophysiological conditions affecting salivary glands including Sjögren’s syndrome, head and neck cancer treated with ionizing radiation therapy, and diabetes.
2.2.1. Sjögren’s Syndrome
Sjögren’s syndrome is an autoimmune disease characterized by lymphocytic infiltration of exocrine glands, in particular salivary and lacrimal glands, often leading to xerostomia (dry mouth) and keratoconjunctivitis sicca [54]. Due to the frequent xerostomia observed in patients with Sjögren’s syndrome, it was hypothesized that altered AQP5 expression and/or localization may account for this observation [22]. Impaired AQP5 localization was detected [22,55] or not [56,57,58] in the salivary glands of patients with Sjögren’s syndrome. The use of distinct patient subsets and/or antibodies may account for these apparent divergent data. In several mouse models of Sjögren’s syndrome, modified AQP5 distribution has indisputably been documented [59,60,61,62,63,64]. Several parameters may account for the altered AQP5 localization [65] such as the presence of inflammatory infiltrates [61], cytokines [66,67,68,69], autoantibodies against muscarinic M3 receptors [70,71], alteration of the machinery regulating intracellular calcium levels [65,72,73,74]. Very recently, AQP5 interacting protein partners have been suggested to regulate AQP5 trafficking in salivary glands from patients with Sjögren’s syndrome [75,76]. Indeed, both ezrin and prolactin-inducible protein, two newly identified protein partners of AQP5 in salivary glands, presented altered expression and/or localization that may account for abnormal AQP5 localization [75,76]. Both prolactin-inducible protein and ezrin interacted with the C-terminus end of human AQP5 [75,76]. Anti-AQP5 antibodies, recently discovered in blood samples of patients with Sjögren’s syndrome, may also take part in salivary gland dysfunction [77]. So far, patients- and methodologies-related variabilities may account for the heterogeneity in the detection of anti-AQP5 antibodies among patients suffering from Sjögren’s syndrome [78] and their association with decreased unstimulated saliva flow [77,79]. Therefore, further studies will be necessary to assess whether anti-AQP5 antibodies lead to salivary gland dysfunction. Rituximab, a B-cell depleting agent, reverted the decreased AQP5 expression detected in the salivary gland and the reduced saliva flow [80]. The beneficial effect of Rituximab on unstimulated saliva flow rate was confirmed in a larger cohort of patients with Sjögren’s syndrome [81], but it remains to assess if this effect may be due to restored AQP5 expression. Overall, abnormal AQP5 localization in salivary glands from patients with Sjögren’s syndrome resulting from multifactorial events may occur in subsets of patients and participate in salivary gland dysfunction.
Modified expression of other AQPs has been found in the salivary glands of patients with Sjögren’s syndrome. Indeed, lower AQP1 [36] and AQP4 [82] expression in myoepithelial cells have been documented. Treatment of a patient suffering from Sjögren’s syndrome with Rituximab reverted the decreased AQP1 expression detected in the salivary gland [80]. However, no association has been found between AQP1 autoantibodies detected in patients with Sjögren’s syndrome [78,83] and reduced saliva flow rate [83]. Besides, saliva secretion was not modified in Aqp1 and Aqp4 knockout mice [12,52]. Overall, these data suggest no involvement of AQP1 and AQP4 in saliva secretion.
2.2.2. Other Diseases
Reduced or loss of AQP5 expression [84,85] and/or altered AQP5 translocation [86] may participate to xerostomia in patients with head and neck cancer treated with ionizing radiation therapy. In addition, decreased AQP5 expression and saliva secretion were also detected following irradiation of the neck area in mice and rats [87,88,89,90,91] and could be reverted in rats by administration of pilocarpine, a muscarinic receptor agonist [92].
2.3. New Therapeutic Options to Modulate AQPs in Diseases Affecting Salivary Glands
To alleviate xerostomia, AQPs have been targeted or modulated by pharmaceutical approaches and have also been used in gene therapy. In vitro, DNA demethylation agents increased AQP5 expression in salivary gland cell lines [68,96]. In vivo, the use of cevimeline, a pharmacological muscarinic agonist agent, restored AQP5 trafficking in salivary glands of different rat models characterized by impaired AQP5 localization [97,98,99]. In addition, the cystic fibrosis transmembrane regulator (CFTR) corrector and potentiator restored AQP5 expression and salivary gland function in a mouse model of Sjögren’s syndrome [100]. Very recently, in a mouse model of Sjögren’s syndrome, anticeramide treatment using myriocin was reported to attenuate inflammation, increase AQP5 expression, and to re-establish salivary gland function [101]. Gene therapy using recombinant adenoviral or adeno-associated viral vectors coding for AQP1 (AdhAQP1) were capable to restore saliva flow to irradiated salivary glands of animals and human with Sjögren’s syndrome [102,103,104,105,106,107], as well as to resolve inflammation [108]. In the clinic, patients with radiotherapy for head and neck cancer or with Sjögren’s syndrome represent ideal target populations for AQP1 gene therapy. However, the advance of AQP1 gene therapy relies on the development of new viral vectors promoting more efficient and persistent gene expression, as well as less immune reactivity. The substitution of the endogenous AQP1 gene promoter with the cytomegalovirus (CMV) promoter using CRISPR-CAS9 gene editing increased AQP1 expression in vitro [109]. Overall, if limitations related to gene therapy can be overcome later on, gene therapy may be clinically beneficial to patients suffering from xerostomia consequent to head and neck cancer treated with ionizing radiation therapy or Sjögren’s syndrome.
3. Gastric Glands
Mammalian gastric glands are sitting in invaginations formed by the epithelium of the gastric mucosa (gastric pits). According to their localization, they are classified into cardiac (in the cardia of the stomach), fundic (in the body and fundus of the stomach) and pyloric glands (in the antrum of the pylorus connecting the stomach to the duodenum). Gastric glands are composed of a variety of cells including several types of mucous cells producing mucous, progenitor cells replenishing the gastric epithelium, parietal cells secreting gastric acid and bicarbonate ions, chief cells releasing pepsinogen, D cells secreting somatostatin, G cells secreting gastrin, X cells secreting ghrelin, enterochromaffin cells releasing serotonin and enterochromaffin-like cells releasing histamine [110].
3.1. Function of AQPs in Gastric Secretion
The stomach expresses many AQPs. Aqp1, Aqp3, Aqp4, Aqp5, Aqp7, Aqp8, Aqp10 and Aqp11 mRNAs have been detected in the fundus, while Aqp1, Aqp2, Aqp 3, Aqp 5, Aqp 7, Aqp 11 mRNAs have been detected in the antrum of the pylorus [111,112,113]. However, the presence of only a few AQPs has been confirmed at the protein level, including AQP3, AQP4, and AQP5 as described in more detail hereafter.
Upon histamine stimulation, AQP4 located at the basolateral plasma membrane of chief and parietal cells [43,114,115,116,117] undergo phosphorylation and internalization in the vesicle-recycling compartment to escape the degradative pathway [118]. Internalized AQP4 can recycle back to the plasma membrane following histamine wash out. Therefore, AQP4 can dynamically be regulated by endocytosis and recycling. It was proposed that the simultaneous decrease in histamine response and localization of AQP4 at the cell surface account for the down-regulation of gastric secretion [118]. However, the hypothesized role of AQP4 in water movement accompanying gastric secretion has been ruled out using Aqp4 knockout mice [119], thereby suggesting a compensatory role of other AQPs in such a process. Additional studies are necessary to assess if AQP4 may still play a role in the maintenance of the volume of gastric cells. The function of AQP5, located at the apical and lateral membranes of pyloric glands [120], remains to be determined in the stomach.
3.2. Pathophysiological Involvement of AQPs in Gastric Diseases
Quite a few AQPs have been involved in pathogenic mechanisms of diseases affecting the stomach, in particular, gastric cancer and chronic gastritis [113,121,122,123,124,125,126,127,128].
AQP3 and AQP5 ensure significant functions in gastric cancer [129]. They induce the invasion and metastasis of gastric cancer cells by regulating epithelial-mesenchymal transition [123,130]. Diminished levels of miR-877, miR874, and miR370 increased AQP3 expression and epithelial-mesenchymal transition [131,132,133]. The levels of long noncoding RNA LINC00659, acting as a molecular sponge of miR-370, were upregulated in gastric cancers and associated with tumor stage, metastasis and poorer prognosis [133]. LINC00659 knockdown suppressed the proliferation, metastasis and epithelial-mesenchymal transition in gastric cancer cells in vitro [133]. Mechanistically LINC00659 competes with miR-370 to increase AQP3 expression in gastric cancer [133]. Recent work reported the identification and validation of AQP5 as a potentially specific marker of gastric cancer stem cells, as well as the role of AQP5 in the self-renewal and tumorigenesis of gastric cancer stem cells by complementing the effect of leucine-rich repeat-containing G protein-coupled receptor 5 (LGR5) [134]. Mechanistically, AQP5 regulated the autophagy and stemness of gastric cancer stem cells by recruiting E3 ligase Tripartite Motif Family Like 1 (TRIM21) to the key autophagy protein Unc-51 Like Autophagy Activating Kinase 1 (ULK1) and inducing the ubiquitination of ULK1 [134]. In Epstein-Barr virus (EBV)-associated gastric cancer, the EBV-encoded protein latent membrane protein 2A (LMP2A) could down-regulate AQP3 by inhibiting the activation of Mechanistic Target Of Rapamycin Kinase (mTOR) signaling pathway and further inhibit autophagy and migration of gastric cancer cells [135]. Furthermore, AQP3 increased the expression of Eukaryotic Translation Initiation Factor 4E Binding Protein 1 (EIF4ABP1 also named 4E-BP1; a target of mTOR) and its phosphorylation by activating Mitogen-Activated Protein Kinase 1 (MAPK1; also named Extracellular Signal-Regulated Kinase (ERK)) signaling pathway, which induced autophagy and promoted cell proliferation in gastric cancer cells [135]. Helicobacter pylori infection, representing the major cause of chronic gastritis, can promote AQP3 and AQP5 expressions which correlate with the progression of gastric carcinoma [136,137]. Helicobacter pylori induced both AQP3 and AQP5 expression by activating downstream HIF-1a or ERK1/2, MEK, respectively [136,137]. The expression of some AQPs in gastric cancer may be used as predictive prognostic gastric cancer biomarkers considering their association with overall patient survival [127,138].
Low Aqp4 expression may represent a good biomarker for spasmolytic polypeptide-expressing metaplasia (SPEM) (a regenerative lesion of the gastric mucosa that can evolve towards cancer). The lowered gastrin levels in SPEM [139,140] may account for the decreased Aqp4 expression as gastrin can regulate Aqp4 expression [141].
3.3. New Therapeutic Options to Modulate AQPs in Diseases Affecting Stomach
Considering their role in gastric cancers, AQPs may represent additional molecular therapeutic targets [142]. Moreover, targeting specific miRNA and long noncoding RNA involved in the regulation of AQPs may also provide additional therapeutic options in the future.
4. Duodenal Brunner’s Gland and Intestinal Goblet Cells
Mammalian duodenal Brunner’s glands lie in the submucosal layer of the proximal duodenum and are innervated by cholinergic vagal and polypeptidergic nerves releasing acetylcholine and vasoactive intestinal peptide (VIP), respectively. In most mammalian species, they are found in decreasing abundance from the pylorus-duodenum junction to the biliary/pancreatic duct [143]. The glands, made of clusters of serous cells and branching ducts, secrete a fluid rich in mucin glycoproteins containing a limited amount of bicarbonate and a variety of peptides (such as trypsin inhibitors, antimicrobial peptides, and growth factors). Intestinal goblet cells are scattered in the epithelia of the small intestine and secrete a fluid rich in mucins [144]. The role of goblet cells relies on the secretion of fluid serving as an important barrier to prevent pathogens to invade the intestinal mucosa and cause inflammation, and in the delivery of luminal antigens to antigen-presenting cells to initiate adaptative immune response [145].
4.1. Role of AQPs in Fluid Secretion of Duodenal Brunner’s Gland and Intestinal Goblet Cells
So far, the roles of AQPs in duodenal Brunner’s gland and intestinal goblet cells have been poorly studied. In Brunner’s gland cells, the expression of AQP5 has been detected at the apical, lateral and secretory granule membranes [120] and that of AQP1 at the level of the apical and lateral membranes [146]. VIP stimulated the secretion of bicarbonate and proteins and increased the secretion rate of rat Brunner’s gland [147] by mechanisms involving a cAMP-dependent signaling cascade inducing translocation of AQP5, but not of AQP1, from secretory granules to the apical plasma membrane [143,146]. Once located at the apical plasma membrane, AQP5 is anticipated to facilitate water movement and thereof contribute to increased fluid secretion. The co-localization and co-trafficking of CFTR with AQP5 support the existence of a parallel pathway for electrolyte secretion and osmotic water movement [143]. Additional studies using Aqp5 knockout mice will be valuable to corroborate the involvement of AQP5 in Brunner’s gland secretion. Concerning the expression of AQPs in intestinal goblet cells, only Aqp9 mRNA has been documented in a subset of these cells [148]. Overall, further experiments are necessary to obtain more precise data concerning the expression and function of AQPs in both these duodenal and intestinal secretory cells.
4.2. Role of AQPs in Diseases Affecting the Duodenal Brunner’s Glands and Intestinal Goblet Cells
Celiac disease and cystic fibrosis have been characterized by decreased AQP5 expression in Brunner’s glands which may contribute to the altered duodenal secretion occurring in these diseases [143]. However, the participation of AQP5 in the pathogenesis of celiac disease and cystic fibrosis requires further investigation. To the best of our knowledge, no data are available concerning the role of AQPs in diseases affecting intestinal goblet cells. However, if AQPs are involved in mucin secretion, their dysregulation is anticipated to impair intestinal homeostasis and participate in diseases associated with goblet cells dysfunction, such as colitis and infections.
5. Glands Ensuring Bile Secretion
Bile is a complex biologic fluid rich in water (95%) which contains bile acids, cholesterol, phospholipids, bile pigments, proteins, metabolites, hormones, and inorganic ions [149]. Bile ensures the excretion of cholesterol (in the form of unesterified cholesterol or as bile acids, the latter assisting the emulsification and absorption of lipid compounds in the duodenum) and the elimination of various toxins and drugs. In healthy conditions, adult humans produce each day between 0.8 to 1.0 L of hepatic bile daily at a rate of 30–40 mL per hour [150]. Bile fluid formation begins at the canalicular (apical) membrane of hepatocytes. It involves the secretion of ions and osmotically-active compounds-mainly through the bile salt transporter (e.g., the bile salt export pump (BSEP)) and the organic anion transporter (e.g., the multidrug resistance-associated protein 2, (MRP2)) creating the osmotic force that drives the parallel water flow [150]. The composition of human canalicular bile is then modified by the epithelial cells lining the ductal lumen. Bile is stored and concentrated in the gallbladder (cystic bile), and released into the duodenum during fat digestion [151,152]. While bile water is mostly recovered in the proximal portion of the small intestine [153] bile salts are reabsorbed in the distal ileum from where they return to the liver via the enterohepatic circulation [154,155]. Bile acid-independent bile flow also exists and results from an active secretion of osmotically active inorganic electrolytes and organic anions. Bile formation results from continuous ductal/ductural secretion and canalicular bile flow [150].
In mammals, the epithelial cells lining the hepatobiliary tree express a variety of AQPs with distinct subcellular localizations (Table 1). As in the blood vessels of other body districts, the endothelial cells of the hepatobiliary system express AQP1 [41].
5.1. Role of AQPs in Glands Ensuring Bile Secretion
5.1.1. Liver
Rodent and human hepatocytes express high levels of AQP8 and AQP9 [156,157,158,160,161,169]. Murine hepatocytes [170] and immortalized human hepatocyte cell line Huh7 also express AQP11 [156]. AQP3 and AQP7 have also been detected in the human liver [156]. The redundancy in AQP expression may be due to their distinctive subcellular localization and molecular selectivity [159,171]. In hepatocytes, AQP8, AQP9 and AQP11 fulfill significant functions, whereas the roles of AQP3 and AQP7 in hepatocytes remain unclear.
In hepatocytes, AQP8 shows multiple subcellular localizations that range from the canalicular membrane and subapical vesicles to organelles such as the mitochondria and the smooth endoplasmic reticulum [157,169]. Hepatocyte AQP8 has been suggested to mediate the secretion of canalicular bile water [172], preserve the cytoplasm osmolarity during glycogen synthesis or degradation [169], facilitate the ammonia movement in mitochondrial ammonium detoxification and ureagenesis [173,174,175,176] and mediate the efflux of hydrogen peroxide out of mitochondria during the oxidative stress [177,178]. AQP8 may exert a role in the cholesterol biosynthesis modulated by the sterol regulatory element-binding protein (SREBP) [179,180,181]. Based on its peroxiporin activity, AQP8 has recently been implicated in the differential regulation of metabolic signaling by α1- and β-adrenoceptors (ARs) leading to the induction of Ca2+ ion mobilization [182]. Extensive work has been carried out in investigating the role of AQP8 in canalicular bile secretion [159]. Choleretic agonists such as dibutyryl cAMP or glucagon induced the translocation of subapical vesicles incorporating AQP8 to the canalicular plasma membrane through a phosphatidylinositol-3-kinase (PI3K)-dependent microtubule-associated pathway [183]. This was accompanied by an increase in the water permeability of the canalicular plasma membrane with consequent osmotic movement of water into the bile canaliculus [172,184,185] (Figure 2). Glucagon also increased the AQP8 expression by decreasing its degradation via a mechanism involving cAMP-PKA and PI3K signal pathways in rat primary hepatocytes [186].
The pleiotropic functional relevance of AQP9 results from its permeability to various neutral solutes (i.e., glycerol and other polyols, H2O2, urea, carbamides, nucleosides, monocarboxylates, purines, pyrimidines and metalloid arsenic) in addition to water [187]. In rodent hepatocytes, AQP9 is localized at the sinusoidal domain of the basolateral plasma membrane [161] and ensures the principal entry pathway for blood glycerol into hepatocytes during fasting [188,189,190,191]. Imported glycerol is rapidly converted into glycerol-3-phosphate (G3P) acting as a major substrate for gluconeogenesis during early starvation. AQP9 has also relevance in lipid homeostasis since G3P is needed for triacylglycerols (TAGs) synthesis [192]. Hepatocyte AQP9 has been involved in rodent bile formation and in extruding catabolic urea by facilitating the entry of water and the exit of urea from and to portal blood, respectively [193,194]. In rodents, the negative regulation of Aqp9 transcription by insulin [195] may account for enhanced hepatic AQP9 expression in insulin resistance [196,197]. The functional relevance of AQP9 in metabolic homeostasis and energy balance was also revealed by the diminished liver glycerol permeability and the enhanced concentrations of plasma glycerol and TAGs detected in AQP9-depleted knockout mice [193,198]. Reduction in AQP9 levels and liver glycerol permeability were seen in hepatocytes of murine models of obesity and subjects with obesity with type 2 diabetes [199,200]. The hepatic expression of AQP9 is also controlled by leptin [192,201]. However, the modulation exerted by these hormones on AQP9 appears to be different between humans and rodents [197]. Gender-related dimorphism of liver AQP9 expression is seen both in rodents and humans in line with the distinctions between the two sexes in handling glycerol for metabolic purposes [201]. In rat hepatoma cell lines, AQP9 played a role in the lipid-lowering activity of silybin, a nutraceutical phytocompound, through modulation of the autophagic process and lipid droplet composition [202]. Liver AQP9 seems also to have immune relevance considering its involvement in the early acute phase of the inflammatory response triggered by TLR4 ligands in mice [203]. Furthermore, the inhibition of AQP9 with the selective and potent blocker RG100204 abolished the LPS-induced increase in NO and O2− levels in FaO hepatoma cells [204].
Mouse liver AQP11 was involved in rough endoplasmic reticulum homeostasis and liver regeneration through a mechanism that remains unclear [156,170]. The recent functional identification of AQP11 as a peroxiporin suggests its potential implication in the regulation of intracellular H2O2 balance to avoid ER stress [205]. Additional studies will be valuable to determine the exact role of this AQP in the liver.
5.1.2. Bile Ducts
In cholangiocytes, secretin induces bile secretion through a cascade of events including the activation of the cAMP signaling pathway [206], a Cl− efflux via CFTR driving the extrusion of HCO3− into the lumen via apical AE2 (chloride/bicarbonate exchanger) and an osmotically-driven water movement through AQP1 located at the apical membrane [206]. In human and rodent cholangiocytes, AQP1 mediates the apical secretion of water during both basal- and secretin-induced ductal bile formation [41,207,208]. AQP1 is also found in subapical membrane vesicles [162], co-expressed with AE2 and CFTR [209], undergoing exocytic insertion into the apical membrane of cholangiocytes upon secretin stimulation [162,209]. In the new notion of functional bile secretory unit, AQP4 and AQP1 located at the basolateral and apical plasma membrane of cholangiocytes [162,210] are allowing water movement to ensure the maintenance of the relative iso-osmolar status of the cell during ductal bile formation. This novel concept is in line with the physical association that exists between the cholangiocyte basolateral membrane and the peribiliary vascular plexus that surrounds the bile ducts and from which bile water derives [159,211] (Figure 3).
However, the water permeability of cholangiocytes isolated from AQP1-depleted knockout mice was not diminished [212]. Compensatory upregulation of other mouse cholangiocytes AQPs (i.e., AQP8) has been invoked to explain this observation [213,214]. Intrahepatic bile ducts not only secrete but also absorb water as demonstrated in isolated rodent intrahepatic bile duct units [151]. It was hypothesized that osmotic absorption of water is triggered by the active uptake of sodium-coupled glucose and bile salts through the sodium/glucose transporter 1 (SGLT1) and apical sodium-dependent bile acid transporter (ASBT), respectively [206]. Somatostatin, gastrin and insulin may decrease ductal bile secretion by lowering the intracellular levels of cAMP and the subsequent translocation of AQP1, CFTR and AE2 induced by secretin. This mechanism may explain why somatostatin reduces ductal secretion while stimulating the net absorption of ductal water.
5.1.3. Gallbladder
The mammalian gallbladder stores bile produced through hepatobiliary secretion and plays key roles in lipid digestion and maintenance of metabolic homeostasis. The movement of water across the gallbladder epithelium is led by osmotic forces generated from active salt absorption and secretion [159,215]. Both human and murine gallbladder epithelial cells express AQP1 and AQP8. AQP1 is present both at the apical and at the basolateral plasma membrane of the epithelial cells that line the neck of the organ [163,216]. AQP1 is also localized at the corpus portion in subapical vesicles and plasma membrane [164]. In the murine gallbladder, AQP1 was slightly upregulated by leptin [217]. AQP8 is present at the plasma membrane and, to a lower extent, in intracellular vesicles of the gallbladder epithelium of various species [41,157]. In gallbladder cholangiocytes, AQP1, AQP8, and CFTR were up-regulated by the liver X receptor β (LXRβ), an oxysterol-activated transcription factor highly present in the gallbladder epithelium [218]. A molecular partnership between CFTR and AQPs has also been found in mouse Sertoli cells [219]. In mouse gallbladder epithelium, AQP1 accounted for high water permeability [165]. The osmotic water permeability was independent of cAMP and osmotic gradient size and direction. As in bile duct cholangiocytes, subapical AQP1 is translocated to the apical membrane to ensure water movement across the gallbladder epithelium. Based on its subcellular pattern of localization, gallbladder AQP8 is likely involved in the absorption of water and, to a lesser extent, in the secretion of water into the lumen [157] (Figure 4). However, the exact relevance of AQP1 and AQP8 in gallbladder function remains a debated argument due to contradictory reported results. Indeed, similar bile salt concentrations were found from gallbladders of wild-type and AQP1-ablated mice, with no apparent functional substitution of AQP1 by AQP8 [165]. However, this observation was not supported by the temporal association observed between diminished gallbladder concentrating function and decreased AQP1 or AQP8 levels [164], and the effects leptin replacement in leptin-deleted mice altering the gallbladder volume acting on the AQP-mediated absorption/secretion of water [220]. Additional studies will be worth to clarify the question.
5.2. Role of AQPs in Diseases Affecting the Glands Secreting Bile
Various diseases that affect the hepatobiliary tree have been linked to perturbed bile fluid secretion and resulting cholestasis [159,221]. Dysregulated hepatobiliary AQPs and altered bile secretion have been observed in various experimental models of cholestasis. Studies with cellular and murine models of gallstone disease suggest an association between altered AQP expression/localization in cholangiocytes and gallbladder concentrating function [159].
5.2.1. Liver and Bile Ducts
Dysregulated expression of AQP8 at the canalicular side of hepatocytes contributes to the development of cholestasis in several experimental models of cholestasis including extrahepatic obstructive cholestasis [222], estrogen-induced cholestasis [223] and sepsis-induced cholestasis [224]. The association between downregulation of canalicular AQP8 and decreased canalicular osmotic water permeability suggests relevance for AQP8 in cholestasis [207,223]. The conjoint occurrence of reduced solute transport and impaired water permeability may account for cholestasis [225]. Interestingly, adenoviral transfer of the human AQP1 gene to rat liver improved bile flow in estrogen-induced cholestasis with potential therapeutic implications for cholestatic diseases [226]. New considerations have been made regarding the occurrence of cholestasis and its progression [227,228] taking into account that a reduction in the paracellular or transcellular canalicular water flow had no significant effect on bile acid excretion [229]. Basolateral AQP9, facilitating the movement of water from the sinusoidal blood into the hepatocyte, was downregulated at a post-transcriptional level in a rodent model of extrahepatic cholestasis [194]. Hepatic cystogenesis accompanied by altered expression and subcellular distribution of AQP1 (together with CFTR and AE2) was seen in a rat model of autosomal recessive polycystic kidney disease. Liver cysts likely resulted from the increased fluid accumulation triggered by the overexpression and ectopic localization of AQP1, CFTR, and AE2 in cystic cholangiocytes [230]. Disruption of the Aqp11 gene in mice leads to intracellular vacuolization of periportal hepatocytes and a severe form of polycystic kidney disease (PKD) with uremic death before weaning due to renal failure [170]. Since the life span of Aqp11−/− mice was limited by kidney disease, the liver phenotype could be premature. Polycystic livers are expectable in Aqp11 knockout mice since cysts are often seen in the biliary epithelia of patients and mice with PKD [231]. Further studies are needed to assess whether the PKD provoked by the depletion of Aqp11 in mice triggers the same liver cysts induced by the autosomal recessive form of PKD, a well-known form of PKD caused by homologous congenital polycystic kidney.
5.2.2. Gallbladder
Gallstone disease (cholecystolithiasis) is characterized by a high prevalence (10–15% in adulthood) and cost in Western countries [232,233]. The majority of gallstones are cholesterol gallstones, while the minority are pigment stones that contain less than 30% cholesterol [234]. The prevalence of gallstones augments with age, while being associated with multiple risk factors [232,235]. Cholesterol gallstone disease results from altered cholesterol homeostasis inducing crystallization of bile cholesterol and reduced gallbladder contractility [236]. Cholesterol gallstone disease is currently considered as the hepatobiliary expression of the metabolic syndrome, being frequently associated with obesity, type 2 diabetes, dyslipidemia, and hyperinsulinemia. Reduced expression of the gallbladder AQP1 and AQP8 was associated with a decrement of the gallbladder concentrating ability in mice fed a lithogenic diet [164]. Dysregulated Aqp1 and Aqp4 mRNA levels were found in the gallbladder of leptin-deficient obese [Lep(ob)] mice undergoing leptin replacement [217]. Besides showing the characteristic obesity, Lep(ob) mice show enhanced gallbladder volumes and diminished gallbladder contractility, the latter reflecting gallbladder stasis [220]. However, in humans, no significant relationship has been found between AQP1 and AQP8 expression in the gallbladder epithelium and the occurrence of gallbladder stones [163]. A recent study has reported that sex-specific expression and localization of hepatobiliary AQPs, associated with reduced biliary water movement, may account for lower cholesterol gallstone prevalence in female thyroid hormone (TH) deficient mice [237]. Additional experiments will be valuable to better understand the function of AQPs in gallstone disease.
5.3. New Therapeutic Options Using AQPs in Diseases Affecting Bile Secretion
Downregulated canalicular expression of BSEP and MRP2 and AQP8 plays a key role in the development of hepatocellular cholestasis [150,238]. There are still no studies aimed at modulating the expression of endogenous AQP to counteract clinical conditions with reduced bile secretion. However, compelling experimental evidence indicates that hepatic AQP gene transfer may improve hepatocellular cholestasis [239]. Recent studies using rodent models of hepatocellular cholestasis showed that the hepatic transfer of an adenoviral vector encoding for human AQP1 (AdhAQP1) improves bile secretion and re-establishes high serum levels of bile salts [226,240,241,242]. Interestingly, the AdhAQP1-transduced hepatocytes showed that the heterologous expression of hAQP1 at the canalicular membrane led to an increase in the osmotic membrane water permeability with an induction of the transport activities of BSEP and MRP2 by their redistribution in canalicular lipid rafts likely through the interaction with the cholesterol-binding protein caveolin-1.
6. Pancreas
6.1. Exocrine Pancreas
Histologically, the exocrine pancreas represents a major part of the pancreas (±90%). The glandular structure of the exocrine pancreas is very similar to that of salivary glands but presents few particularities. Indeed, the exocrine pancreas is made of only serous acinar cells, contains centroacinar cells (extension of intercalated ducts into each acinus) and a main collecting duct ensures pancreatic fluid drainage.
The pancreatic fluid, controlled by neurotransmitters (i.e., acetylcholine, cholecystokinin and secretin), neutralizes the stomach acid and ensures food digestion [243].
6.1.1. Role of AQPs in Exocrine Pancreatic Secretion
The distribution of AQPs in the exocrine pancreas varies according to species. In the human exocrine pancreas, despite the detection of several Aqps mRNA (i.e., Aqp1, Aqp3, Aqp4, Aqp5, Aqp8 and Aqp12), the protein expression of only a few of these AQPs has been confirmed (i.e., AQP1, AQP5 and AQP8) [244,245]. AQP1 was located in endothelial cells, centroacinar cells (apical membrane), intercalated ductal cells [244] and pancreatic zymogen granules [246,247]. AQP5 and AQP8 were, respectively, located in the apical membrane of both intercalated ductal cells and acinar cells, respectively [244].
In rat exocrine pancreas, Aqp1, Aqp4, Aqp5, Aqp8 but no Aqp12 mRNA have been detected [244,245,248]. AQP1 was localized to intralobular (apical and basolateral membrane) and interlobular ductal cells (apical and basolateral membranes and caveolae and vesicle-like structures) [249,250], acinar zymogen granules [246] and endothelial cells [248]. AQP5 was detected in centroacinar and intercalated ductal cells (apical membrane) [251]. AQP8 was located at the apical plasma membrane of acinar cells [245].
The murine exocrine pancreas expresses AQP1 and AQP5 in interlobular ductal cells (apical membrane), AQP5 in intercalated and intralobular ductal cells (apical membrane) [251], and AQP12 in acinar cells (intracellularly) [252].
Pancreatic juice output originates from the secretion of a small isotonic fluid by acinar cells. In acinar cells, water movement is ensured through AQP8 and AQP1 (located at the apical membrane). The content of the initial pancreatic fluid is then modified by the ductal cells which secrete ions and ensure the majority of the water movement to the ductal lumen via AQP1 (located at the apical and basolateral membranes) and AQP5 (located at the apical membrane) [10,251,253]. In rats, AQP8 is responsible for most water permeability (90%) in acinar cells [248]. However, Aqp8 knockout mice show normal exocrine pancreatic function, possibly due to the major contribution of ductal cells to transcellular water transport [53]. In rats, AQP1 contributes to basal and GTP-mediated vesicle water movement and swelling of pancreatic acinar zymogen granules [246,247]. In rat interlobular ductal cells, AQP1 is responsible for most of the pancreatic juice secretion induced by secretin [250]. However, both Aqp1 and Aqp5 knockout mice show no modification in exocrine pancreatic function [244], suggesting their low contribution to pancreatic fluid secretion and/or the presence of compensatory mechanisms. Therefore, the use of double Aqp1 and Aqp5 knockout mice are anticipated to precise the overall contribution of these AQPs to the pancreatic juice formation. A study using Aqp12 knockout mice has suggested AQP12 may participate in the mechanisms that control the exocrine pancreatic secretory function in response to following rapid and intense stimulation [254]. Very recently, the contribution of AQP12 to pancreatic juice secretion has also been revealed in patients presenting a nonsense mutation in the Aqp12B gene, one of the two homologous Aqp12 genes (Aqp12A and Aqp12B) [255]. Overall, additional studies are still necessary to evaluate the specific contribution of each AQP to pancreatic fluid secretion.
6.1.2. Role of AQPs in Diseases Affecting Exocrine Pancreatic Secretion
Some AQPs have been incriminated in the pathogenesis of pancreatitis, cystic fibrosis and cancer [129,256,257,258]. Indeed, AQP1 expression was decreased in a rat model of acute necrotizing pancreatitis [259] and in a mouse model of pancreatic exocrine insufficiency [260]. However, the role of AQP1 in pancreatitis remains to be further explored. Beyond affecting the pancreatic function, pancreatitis can affect the lung and colon characterized by impaired AQPs expression [261,262,263]. AQP5 expression was decreased in a mouse model of autoimmune pancreatitis [100]. In that model, the CFTR corrector C18 and the CFTR potentiator VX770 rescued CFTR expression, corrected AQP5 expression and pancreatic fluid secretion and alleviated tissue inflammation [100]. However, the effects of these molecules on the AQP1 expression remain to be assessed. Nevertheless, CFTR correctors may represent strong candidates for the treatment of pancreatitis. Considering gene therapeutic approaches have been considered in pancreatic diseases [264], it may also be interesting to evaluate the effects of viral vectors encoding AQP1 or AQP5 [102,103,107,265].
CFTR, located in ductal cells, participates in pancreatic fluid secretion [266]. In cystic fibrosis, a genetic disease resulting from autosomal mutations in the CFTR gene, pancreatic juice secretion is altered [267]. In guinea-pig and mouse models of cystic fibrosis, the underlying mechanisms causing decreased AQP1 expression and reduced pancreatic fluid secretion remain to be elucidated [268,269].
In various cancers, AQPs have been involved in cell proliferation, migration, adhesion, invasion, metastasis, drug resistance, angiogenesis and epithelial-mesenchymal transition [270,271,272,273,274,275,276,277]. AQPs have been implicated in the pathogenesis of pancreatic ductal adenocarcinoma (PDAC), the most prevalent, aggressive and lethal pancreatic cancer [256,278]. The role of AQPs in cell proliferation, migration, and apoptosis have been studied using PDAC cell lines [256,278,279]. In addition, AQP3 and AQP5 have been implicated not only in cell migration but also in cell-cell-adhesion by modulation of cell biomechanical properties in a PDAC cell line [280]. Using a panel of PDAC cell lines, miR-874 was shown to regulate Aqp3 mRNA expression which promoted cell proliferation and survival (via increased expression and activity of mTOR and downstream targets) [281]. However, the role of AQPs in PDAC tissues has been less investigated. The expression of Aqp1, Aqp3, Aqp4, Aqp5 and Aqp8 mRNAs were detected in PDAC [244]. Moreover, Aqp1 and Aqp3 mRNA were upregulated in PDAC as compared to healthy pancreatic tissue [282]. The expression of AQP1 and AQP3 protein were increased in PDAC, predictive markers of poor prognosis, but associated with patients’ survival [258,281,282]. However, a study reported decreased Aqp3 and Aqp8 mRNA expression in PDAC [283]. These discrepant data may result from distinct patient cohort characteristics and analytic methodologies. Regarding AQP5 protein in PDAC, its localization was altered, and its expression was increased (more in moderately differentiated than in poorly differentiated PDAC) and correlated with tumor differentiation status and aggressiveness [258]. Further studies will be warranted to better understand the involvement of AQPs in PDAC, their usefulness as predictive or prognostic markers and their contribution to chemo-resistance.
6.2. Endocrine Pancreas
As opposed to the exocrine pancreas, the endocrine pancreas only accounts for a minor portion of the entire pancreatic tissue (±10%). Despite its minor representation, the endocrine pancreas fulfills important endocrinological functions by ensuring the secretion of two major metabolic hormones regulating glucose metabolism such as insulin and glucagon as well as other functionally relevant peptide hormones. The endocrine pancreas consists of islets of Langerhans scattered among the pancreatic tissue. Islets of Langerhans contain insulin-producing ß-cells surrounded by glucagon-producing α-cells, somatostatin-producing δ-cells, and pancreatic polypeptide-producing PP cells [284]. The primary role of the B-cells is to release insulin during the postprandial state to decrease glycemia [285,286]. Insulin secretion by B-cells results from the succession of several events. The first event consists of the entrance of glucose through the glucose transporter type 2 (GLUT2). Then the metabolization of glucose induces an increase in the intracellular ATP content which inhibits ATP-sensitive K+ channels that leads to membrane depolarization, and subsequent opening of voltage-dependent Ca2+ channels. The resulting intracellular calcium elevation triggers the exocytosis of insulin-containing granules into the bloodstream [285]. Furthermore, glucose induces b-cell swelling [287] which causes the activation of volume-regulated anion channel (VRAC) with consequent cell membrane depolarization and in turn the activation of voltage-dependent Ca2+ channels. The latter allows calcium entry which triggers insulin secretion [288,289].
6.2.1. Role of AQPs in Endocrine Pancreatic Secretion
Very recently, the presence of Aqp7 mRNA has been reported in a-, b- and d-cells from human islets of Langerhans [290]. Rat β-cells express AQP7 [291,292,293] and mouse β-cells express AQP5, AQP7, AQP8 [292] and AQP12 [294]. Further studies are still required to obtain a comprehensive map of AQPs distribution in human and rodent islets of Langerhans.
While data have ruled out the involvement of AQP12 in insulin secretion [294], several studies have pointed out the role of AQP7 in the regulation of intracellular glycerol content, insulin production and secretion in b-cells. Indeed, Aqp7 knockout mice were characterized by reduced β-cell size and mass, insulin content and cAMP-driven glycerol release [293,295]. Furthermore, the mice displayed higher basal and glucose-stimulated insulin secretion rates, glycerol and TAG contents and glycerol kinase activity [293]. The mice’s genetic background are likely influencing the phenotype of the Aqp7 knockout mice, i.e., the presence of hyperinsulinemia [293,295] combined with the presence [295] or absence [293] of hyperglycemia, or the presence of normoglycemia with undetermined insulin levels [296].
In vitro, extracellular isosmotic glycerol-induced sequential cell swelling, volume-regulated anion channel (VRAC) activation, membrane depolarization, calcium entry, and insulin secretion in both rat β-cells and pancreatic b-cell line BRIN-BD [291,297,298]. These effects are likely mediated by the entrance of glycerol and its subsequent metabolization [291]. Ex vivo, β-cells from Aqp7 knockout mice displayed lower insulin secretion in response to D-glucose, extracellular hypotonicity or extracellular isosmotic addition of glycerol [292]. By allowing the passage of glycerol, AQP7 may regulate insulin secretion by acting at a distal downstream site in the insulin exocytosis pathway [292]. A recent analysis of AQP7 by cryo-electron microscopy and gas chromatography/mass spectrometry have suggested that the central pore formed by the tetrameric structure of AQP7 is permeable to glycerol-3-phosphate, while each individual pore formed by monomeric AQP7 is permeable to glycerol [290]. Furthermore, crystal structure revealed that the AQP7 dimer of tetramers of opposite lipidic layers interacts to form octamer [290]. It was hypothesized AQP7 octamer may serve as a junction protein and promote cell-cell adhesion between β-cells as well as in the rosette-like structures of b-cells around blood capillaries (also expressing AQP7) [290]. Hence, AQP7 could be considered a junctional protein. However, functional evidence for the role of AQP7 octamer in cell-cell adhesion is still lacking and further studies will be required to address this question. Therefore, AQP7 may facilitate intercellular movement and distribution of glycerol as well as other metabolites such as glycerol-3-phosphate. Considering adherens junction have been shown to be important for insulin secretion [299,300], AQP7 may therefore be involved in the control of insulin secretion through its function of glycerol and glycerol-3-phosphate channel and its feature to act as a junctional protein. Figure 5A recapitulates the role of AQP7 in β-cell physiology.
6.2.2. Role of AQPs in Diseases Related to Altered Endocrine Pancreatic Secretion
Inflammation that has been involved in diabetes [301] can alter AQPs expression. Indeed, tumor necrosis factor α (TNF-α) reduced AQP7 and insulin secretion, but increased AQP12 levels, in rat pancreatic β-cell line RIN-m5F [302]. In the same cells, lipopolysaccharides raised both AQP7 and AQP12 levels but lowered insulin release [302]. Furthermore, in RIN-m5F cells incubated with (TNF-α) or lipopolysaccharides, overexpression and silencing of AQP7 or AQP12 showed the role of AQP7 in insulin release and of AQP12 in inflammation [302]. AQP8, expressed at the mitochondrial and plasma membranes of rat RIN-m5F β -cells, reduced cytokine-induced cell toxicity [303].
The human Aqp7 gene is present in a chromosomal region that has been associated with type 2 diabetes [304] and metabolic syndrome [305]. In addition, single nucleotide polymorphisms in the Aqp7 gene have been linked to obesity and/or type 2 diabetes in Caucasians [306,307] and with type 2 diabetes in the Chinese Han population [308]. Very recent data showed decreased Aqp7 mRNA in islets from type 2 diabetic patients, a negative correlation between Aqp7 mRNA expression in islets and body mass index, a strong association between a genetic variant of the Aqp7 gene and random glucose and fasting glucose adjusted for body mass index [290]. Other variants of Aqp7 gene were significantly associated with type 2 diabetes or glycated hemoglobin [290]. Altogether, data suggest that some genetic variants of Aqp7 gene can be associated with metabolic features playing a role in type 2 diabetes. Still, the direct link between Aqp7 gene genetic variants and protein dysfunction remains to be proven.
In obese patients with type 2 diabetes, sleeve gastrectomy induced diabetes remission possibly via an increase in glucagon-like peptide 1 (GLP-1) levels and a decrease in ghrelin levels [309,310]. Following sleeve gastrectomy in rats, it has been shown that increased GLP-1 levels and decreased ghrelin levels were responsible for AQP7 upregulation in b-cells which contributed to the improvement of insulin secretion and pancreatic steatosis [294]. Aqp7 knockout mice displayed adult-onset obesity and hyperglycemia [293,295,311]. Altogether, these data suggest that dysregulated AQP7 expression is linked to dysregulated insulin secretion, thereof suggesting AQP7 as a potential therapeutic target for human obesity-associated type 2 diabetes. Interestingly, Metformin, a hypoglycemic drug used to treat diabetic patients, rescued AQP7 expression to induce insulin secretion in the rat in vitro and in vivo models of type 2 diabetes by suppressing the Mitogen-Activated Protein Kinase p38 and c-Jun N-terminal kinase (JNK) pathway [312]. Figure 5B,C recapitulate the role of AQP7 in β-cell during type 2 diabetes untreated or treated with Metformin.
While DNA methylation, an epigenetic modification, of the Aqp7 gene promoter has been correlated with the decreased expression in human white adipose tissue and adiposity [313,314], further studies will be necessary to determine if such epigenetic modification can affect Aqp7 gene expression and insulin secretion in β-cells.
Further studies are needed to pursue deciphering the role of AQPs in the pathogenesis of type 2 diabetes and associated inflammation.
7. Conclusions
Studies on the expression and role of AQPs throughout the gastrointestinal system have been highly instructive to provide useful insights into complex molecular mechanisms by which gastrointestinal exocrine and endocrine glands secrete and reabsorb water in concert with organic and inorganic compounds. In addition, important acquisitions have been made about the pathophysiological implication of AQPs in multiple gastrointestinal diseases accompanied by fluid imbalance.
In salivary glands, AQP5 is the main AQP playing a key role in saliva secretion. In addition, deregulation of its expression may participate in the xerostomia manifesting in patients suffering from Sjögren’s syndrome, head and neck cancer treated with ionizing radiation therapy, or diabetes. Several therapeutic options have therefore been tested to modulate and/or restore the expression of AQP5 to alleviate xerostomia. In gastric glands, the role of AQP4 in gastric secretion remains a source of debate and a better understanding of its role in gastric cell volume maintenance will require further investigations. In gastric cancers, AQP3 and AQP5 may represent additional therapeutic targets for therapeutic intervention considering they have been shown to participate in the epithelial-mesenchymal transition of the gastric epithelium, to be regulated by miRNA and long coding RNA, and to represent interesting predictive prognostic biomarkers. So far, studies have solely investigated the expression of AQPs, but not their roles, in duodenal and intestinal secretory glands. So far, AQP5 and AQP1 have been hypothesized to participate in duodenal Brunner’s gland secretion, but additional studies will be worth to validate such a hypothesis. The role of AQP9 expression in intestinal goblet cells also remains to be studied further. In glands ensuring bile secretion, the expression and role of AQPs appear to be relatively specific to the different cell types involved including hepatocytes (AQP8 and AQP9), bile ducts (AQP1, AQP4), and gallbladder (AQP1 and AQP8). Deregulation of AQPs expression in cells involved in bile secretion has been linked to pathologies such as cholestasis and gallstone disease suggesting relevance as new therapeutic targets. Interestingly, hepatic gene transfer of human AQP1 has been reported to ameliorate the bile secretory failure in hepatocellular cholestasis by enhancing both biliary output and choleretic efficiency of key osmotic solutes such as bile salts and glutathione. Nevertheless, further studies will be valuable to deepen the current understanding of the complex mechanisms involved in bile secretion under physiological and pathological conditions. In the pancreas, the relative contribution of AQP1, AQP5, AQP8 and AQP12 to the exocrine secretion remains poorly understood. Some of these AQPs may play a role in cystic fibrosis and cancer and therefore, be considered interesting drug targets. Despite the detection of several AQPs in the endocrine pancreas, only AQP7 has been shown to date to participate in insulin secretion, and to undergo deregulation in type 2 diabetes that can be rescued by hypoglycemic drugs. Still, additional experiments will contribute to further precise the role of AQPs in exocrine and endocrine pancreatic secretions in healthy and pathological conditions.
Despite efforts made so far to decipher the regulation and precise function of AQPs in gastrointestinal glands, some questions remain open deserving additional studies. However, while studying the ultimate pathophysiological role of AQPs in several clinical disorders, novel pharmacological strategies might appear soon to treat gastrointestinal secretory/absorptive failures in which gastrointestinal glands epithelia are the target cell.
Author Contributions
Conceptualization, C.D.; writing—original draft preparation, C.D. and G.C.; writing—review and editing, C.D. and G.C.; visualization, C.D. and G.C.; supervision, C.D.; project administration, C.D.; funding acquisition, C.D. and G.C. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Fund Octave Dupont from the Royal Academy of Belgium (C.D.). G.C. was supported by a grant from the Italian Government (“Fondo Integrativo Speciale per la Ricerca 2020”—FISR 2020 CoVAPin, grant #FISR2020IP_04051) and, in part, by a grant from the University of Bari (“Horizon Europe Seeds 2022-2023” NextgenerationEU Programma MUR—Fondo Promozione e Sviluppo-DM 737/2021-Uniba Euroseeds ID #S10) and another grant from the Italian Government (“Programmi di Ricerca Scientifica di Rilevante Interesse Nazionale 2017” PRIN2017 #2017J92TM5).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
This review is in memory of Lucantonio Debellis as a person of great human qualities and as an excellent physiologist and pathophysiologist of the gastrointestinal system. The Figures were partly generated using Servier Medical Art, provided by Servier, licensed under a Creative Commons Attribution 3.0 unported license (https://creativecommons.org/licenses/by/3.0/).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Agre, P. Aquaporin Water Channels (Nobel Lecture). Angew. Chem. Int. Ed. 2004, 43, 4278–4290. [Google Scholar] [CrossRef]
- Verbavatz, J.M.; Brown, D.; Sabolić, I.; Valenti, G.; Ausiello, D.A.; Van Hoek, A.N.; Ma, T.; Verkman, A.S. Tetrameric Assembly of CHIP28 Water Channels in Liposomes and Cell Membranes: A Freeze-Fracture Study. J. Cell Biol. 1993, 123, 605–618. [Google Scholar] [CrossRef] [PubMed]
- Madeira, A.; Fernández-Veledo, S.; Camps, M.; Zorzano, A.; Moura, T.F.; Ceperuelo-Mallafré, V.; Vendrell, J.; Soveral, G. Human Aquaporin-11 Is a Water and Glycerol Channel and Localizes in the Vicinity of Lipid Droplets in Human Adipocytes. Obesity 2014, 22, 2010–2017. [Google Scholar] [CrossRef]
- Geng, X.; Shao, G.; Jiang, T.; Yang, B. Transport Characteristics of Aquaporins. Adv. Exp. Med. Biol. 2023, 1398, 53–64. [Google Scholar] [CrossRef] [PubMed]
- Takata, K.; Matsuzaki, T.; Tajika, Y. Aquaporins: Water Channel Proteins of the Cell Membrane. Prog. Histochem. Cytochem. 2004, 39, 1–83. [Google Scholar] [CrossRef] [PubMed]
- Calamita, G.; Delporte, C. Aquaporins in Glandular Secretion. In Aquaporins; Springer: Singapore, 2023; pp. 225–249. [Google Scholar]
- Amano, O.; Mizobe, K.; Bando, Y.; Sakiyama, K. Anatomy and Histology of Rodent and Human Major Salivary Glands: Overview of the Japan Salivary Gland Society-Sponsored Workshop. Acta Histochem. Cytochem. 2012, 45, 241–250. [Google Scholar] [CrossRef]
- Young, J.A.; Van Lennep, E.W. The Morphology of Salivary Glands; Academic Press: London, UK; New York, NY, USA, 1978; ISBN 978-0-12-772950-3. [Google Scholar]
- Redman, R. Development of Salivary Glands. In The Salivary System; Sreebny, L.M., Ed.; CRC Press: Boca Raton, FL, USA, 1987; pp. 1–20. ISBN 13978-0849366895. [Google Scholar]
- Lee, M.G.; Ohana, E.; Park, H.W.; Yang, D.; Muallem, S. Molecular Mechanism of Pancreatic and Salivary Gland Fluid and HCO3 Secretion. Physiol. Rev. 2012, 92, 39–74. [Google Scholar] [CrossRef]
- Melvin, J.E.; Yule, D.; Shuttleworth, T.; Begenisich, T. Regulation of Fluid and Electrolyte Secretion in Salivary Gland Acinar Cells. Annu. Rev. Physiol. 2005, 67, 445–469. [Google Scholar] [CrossRef]
- Ma, T.; Song, Y.; Gillespie, A.; Carlson, E.J.; Epstein, C.J.; Verkman, A.S. Defective Secretion of Saliva in Transgenic Mice Lacking Aquaporin-5 Water Channels. J. Biol. Chem. 1999, 274, 20071–20074. [Google Scholar]
- Krane, C.M.; Melvin, J.E.; Nguyen, H.V.; Richardson, L.; Towne, J.E.; Doetschman, T.; Menon, A.G. Salivary Acinar Cells from Aquaporin 5-Deficient Mice Have Decreased Membrane Water Permeability and Altered Cell Volume Regulation. J. Biol. Chem. 2001, 276, 23413–23420. [Google Scholar] [CrossRef] [PubMed]
- Maclaren, O.J.; Sneyd, J.; Crampin, E.J. What Do Aquaporin Knockout Studies Tell Us about Fluid Transport in Epithelia? J. Membr. Biol. 2013, 246, 297–305. [Google Scholar] [CrossRef] [PubMed]
- Sneyd, J.; Crampin, E.; Yule, D. Multiscale Modelling of Saliva Secretion. Math. Biosci. 2014, 257, 69–79. [Google Scholar] [CrossRef] [PubMed]
- Hill, A.E.; Shachar-Hill, B. A New Approach to Epithelial Isotonic Fluid Transport: An Osmosensor Feedback Model. J. Membr. Biol. 2006, 210, 77–90. [Google Scholar] [CrossRef] [PubMed]
- Kawedia, J.D.; Nieman, M.L.; Boivin, G.P.; Melvin, J.E.; Kikuchi, K.-I.; Hand, A.R.; Lorenz, J.N.; Menon, A.G. Interaction between Transcellular and Paracellular Water Transport Pathways through Aquaporin 5 and the Tight Junction Complex. Proc. Natl. Acad. Sci. USA 2007, 104, 3621–3626. [Google Scholar] [CrossRef]
- Matsuzaki, T.; Suzuki, T.; Koyama, H.; Tanaka, S.; Takata, K. Aquaporin-5 (AQP5), a Water Channel Protein, in the Rat Salivary and Lacrimal Glands: Immunolocalization and Effect of Secretory Stimulation. Cell Tissue Res. 1999, 295, 513–521. [Google Scholar] [PubMed]
- Ishikawa, Y.; Eguchi, T.; Skowronski, M.T.; Ishida, H. Acetylcholine Acts on M3 Muscarinic Receptors and Induces the Translocation of Aquaporin5 Water Channel via Cytosolic Ca2+ Elevation in Rat Parotid Glands. Biochem. Biophys. Res. Commun. 1998, 245, 835–840. [Google Scholar] [CrossRef]
- Cho, G.; Bragiel, A.M.; Wang, D.; Pieczonka, T.D.; Skowronski, M.T.; Shono, M.; Nielsen, S.; Ishikawa, Y. Activation of Muscarinic Receptors in Rat Parotid Acinar Cells Induces AQP5 Trafficking to Nuclei and Apical Plasma Membrane. Biochim. Biophys. Acta 2015, 1850, 784–793. [Google Scholar] [CrossRef]
- Wang, W.; Hart, P.S.; Piesco, N.P.; Lu, X.; Gorry, M.C.; Hart, T.C. Aquaporin Expression in Developing Human Teeth and Selected Orofacial Tissues. Calcif. Tissue Int. 2003, 72, 222–227. [Google Scholar] [CrossRef]
- Steinfeld, S.; Cogan, E.; King, L.S.; Agre, P.; Kiss, R.; Delporte, C. Abnormal Distribution of Aquaporin-5 Water Channel Protein in Salivary Glands from Sjögren’s Syndrome Patients. Lab. Investig. 2001, 81, 143–148. [Google Scholar] [CrossRef]
- Raina, S.; Preston, G.M.; Guggino, W.B.; Agre, P. Molecular Cloning and Characterization of an Aquaporin CDNA from Salivary, Lacrimal, and Respiratory Tissues. J. Biol. Chem. 1995, 270, 1908–1912. [Google Scholar] [CrossRef] [PubMed]
- Funaki, H.; Yamamoto, T.; Koyama, Y.; Kondo, D.; Yaoita, E.; Kawasaki, K.; Kobayashi, H.; Sawaguchi, S.; Abe, H.; Kihara, I. Localization and Expression of AQP5 in Cornea, Serous Salivary Glands, and Pulmonary Epithelial Cells. Am. J. Physiol. 1998, 275, C1151–C1157. [Google Scholar] [CrossRef] [PubMed]
- Murdiastuti, K.; Miki, O.; Yao, C.; Parvin, M.N.; Kosugi-Tanaka, C.; Akamatsu, T.; Kanamori, N.; Hosoi, K. Divergent Expression and Localization of Aquaporin 5, an Exocrine-Type Water Channel, in the Submandibular Gland of Sprague-Dawley Rats. Pflug. Arch. 2002, 445, 405–412. [Google Scholar] [CrossRef]
- Larsen, H.S.; Aure, M.H.; Peters, S.B.; Larsen, M.; Messelt, E.B.; Kanli Galtung, H. Localization of AQP5 during Development of the Mouse Submandibular Salivary Gland. J. Mol. Histol. 2011, 42, 71–81. [Google Scholar] [CrossRef] [PubMed]
- Aure, M.H.; Ruus, A.-K.; Galtung, H.K. Aquaporins in the Adult Mouse Submandibular and Sublingual Salivary Glands. J. Mol. Histol. 2014, 45, 69–80. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, S.; King, L.S.; Christensen, B.M.; Agre, P. Aquaporins in Complex Tissues. II. Subcellular Distribution in Respiratory and Glandular Tissues of Rat. Am. J. Physiol. 1997, 273, C1549–C1561. [Google Scholar] [CrossRef]
- Akamatsu, T.; Parvin, M.N.; Murdiastuti, K.; Kosugi-Tanaka, C.; Yao, C.; Miki, O.; Kanamori, N.; Hosoi, K. Expression and Localization of Aquaporins, Members of the Water Channel Family, during Development of the Rat Submandibular Gland. Pflug. Arch. 2003, 446, 641–651. [Google Scholar] [CrossRef]
- Mangos, J.A.; McSherry, N.R. Micropuncture Study of Urea Excretion in Parotid Saliva of the Rat. Am. J. Physiol. 1970, 218, 1329–1332. [Google Scholar] [CrossRef]
- Murdiastuti, K.; Purwanti, N.; Karabasil, M.R.; Li, X.; Yao, C.; Akamatsu, T.; Kanamori, N.; Hosoi, K. A Naturally Occurring Point Mutation in the Rat Aquaporin 5 Gene, Influencing Its Protein Production by and Secretion of Water from Salivary Glands. Am. J. Physiol. Gastrointest. Liver Physiol. 2006, 291, G1081–G1088. [Google Scholar] [CrossRef]
- D’Agostino, C.; Elkashty, O.A.; Chivasso, C.; Perret, J.; Tran, S.D.; Delporte, C. Insight into Salivary Gland Aquaporins. Cells 2020, 9, E1547. [Google Scholar] [CrossRef]
- Delporte, C. Aquaporins in Salivary Glands and Pancreas. Biochim. Biophys. Acta 2014, 1840, 1524–1532. [Google Scholar] [CrossRef]
- Nakamura, M.; Saga, T.; Watanabe, K.; Takahashi, N.; Tabira, Y.; Kusukawa, J.; Yamaki, K.-I. An Immunohistochemistry-Based Study on Aquaporin (AQP)-1, 3, 4, 5 and 8 in the Parotid Glands, Submandibular Glands and Sublingual Glands of Sjögren’s Syndrome Mouse Models Chronically Administered Cevimeline. Kurume Med. J. 2013, 60, 7–19. [Google Scholar] [CrossRef] [PubMed]
- Mobasheri, A.; Marples, D. Expression of the AQP-1 Water Channel in Normal Human Tissues: A Semiquantitative Study Using Tissue Microarray Technology. Am. J. Physiol. Cell Physiol. 2004, 286, C529–C537. [Google Scholar] [CrossRef] [PubMed]
- Beroukas, D.; Hiscock, J.; Gannon, B.J.; Jonsson, R.; Gordon, T.P.; Waterman, S.A. Selective Down-Regulation of Aquaporin-1 in Salivary Glands in Primary Sjögren’s Syndrome. Lab. Investig. 2002, 82, 1547–1552. [Google Scholar] [CrossRef] [PubMed]
- Gresz, V.; Burghardt, B.; Ferguson, C.J.; Hurley, P.T.; Takács, M.; Nielsen, S.; Varga, G.; Zelles, T.; Case, R.M.; Steward, M.C. Expression of Aquaporin 1 (AQP1) Water Channels in Human Labial Salivary Glands. Arch. Oral Biol. 1999, 44 (Suppl. S1), S53–S57. [Google Scholar] [CrossRef] [PubMed]
- Agre, P.; Preston, G.M.; Smith, B.L.; Jung, J.S.; Raina, S.; Moon, C.; Guggino, W.B.; Nielsen, S. Aquaporin CHIP: The Archetypal Molecular Water Channel. Am. J. Physiol. 1993, 265, F463–F476. [Google Scholar] [CrossRef]
- He, X.; Tse, C.M.; Donowitz, M.; Alper, S.L.; Gabriel, S.E.; Baum, B.J. Polarized Distribution of Key Membrane Transport Proteins in the Rat Submandibular Gland. Pflug. Arch. 1997, 433, 260–268. [Google Scholar] [CrossRef]
- Li, J.; Nielsen, S.; Dai, Y.; Lazowski, K.W.; Christensen, E.I.; Tabak, L.A.; Baum, B.J. Examination of Rat Salivary Glands for the Presence of the Aquaporin CHIP. Pflug. Arch. 1994, 428, 455–460. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, S.; Smith, B.L.; Christensen, E.I.; Agre, P. Distribution of the Aquaporin CHIP in Secretory and Resorptive Epithelia and Capillary Endothelia. Proc. Natl. Acad. Sci. USA 1993, 90, 7275–7279. [Google Scholar] [CrossRef]
- King, L.S.; Nielsen, S.; Agre, P. Aquaporins in Complex Tissues. I. Developmental Patterns in Respiratory and Glandular Tissues of Rat. Am. J. Physiol. 1997, 273, C1541–C1548. [Google Scholar] [CrossRef]
- Frigeri, A.; Gropper, M.A.; Turck, C.W.; Verkman, A.S. Immunolocalization of the Mercurial-Insensitive Water Channel and Glycerol Intrinsic Protein in Epithelial Cell Plasma Membranes. Proc. Natl. Acad. Sci. USA 1995, 92, 4328–4331. [Google Scholar] [CrossRef] [PubMed]
- Matsuki-Fukushima, M.; Fujita-Yoshigaki, J.; Murakami, M.; Katsumata-Kato, O.; Yokoyama, M.; Sugiya, H. Involvement of AQP6 in the Mercury-Sensitive Osmotic Lysis of Rat Parotid Secretory Granules. J. Membr. Biol. 2013, 246, 209–214. [Google Scholar] [CrossRef]
- Wellner, R.B.; Redman, R.S.; Swaim, W.D.; Baum, B.J. Further Evidence for AQP8 Expression in the Myoepithelium of Rat Submandibular and Parotid Glands. Pflug. Arch. Eur. J. Physiol. 2006, 451, 642–645. [Google Scholar] [CrossRef] [PubMed]
- Koyama, Y.; Yamamoto, T.; Kondo, D.; Funaki, H.; Yaoita, E.; Kawasaki, K.; Sato, N.; Hatakeyama, K.; Kihara, I. Molecular Cloning of a New Aquaporin from Rat Pancreas and Liver. J. Biol. Chem. 1997, 272, 30329–30333. [Google Scholar] [CrossRef] [PubMed]
- Elkjær, M.-L.; Nejsum, L.N.; Gresz, V.; Kwon, T.-H.; Jensen, U.B.; Frøkiær, J.; Nielsen, S. Immunolocalization of Aquaporin-8 in Rat Kidney, Gastrointestinal Tract, Testis, and Airways. Am. J. Physiol. Ren. Physiol. 2001, 281, F1047–F1057. [Google Scholar] [CrossRef] [PubMed]
- Larsen, H.S.; Ruus, A.-K.; Galtung, H.K. Aquaporin Expression Patterns in the Developing Mouse Salivary Gland. Eur. J. Oral Sci. 2009, 117, 655–662. [Google Scholar] [CrossRef] [PubMed]
- de Paula, F.; Tucker, A.S.; Teshima, T.H.N.; de Souza, M.M.; Coutinho-Camillo, C.M.; Nico, M.M.S.; Lourenço, S.V. Characteristics of Aquaporin 1, 3, and 5 Expression during Early Murine Salivary Gland Development. J. Anat. 2021, 238, 794–806. [Google Scholar] [CrossRef]
- De Paula, F.; Teshima, T.H.N.; Hsieh, R.; Souza, M.M.; Coutinho-Camillo, C.M.; Nico, M.M.S.; Lourenco, S.V. The Expression of Water Channel Proteins during Human Salivary Gland Development: A Topographic Study of Aquaporins 1, 3 and 5. J. Mol. Histol. 2017, 48, 329–336. [Google Scholar] [CrossRef]
- Hosoi, K.; Yao, C.; Hasegawa, T.; Yoshimura, H.; Akamatsu, T. Dynamics of Salivary Gland AQP5 under Normal and Pathologic Conditions. Int. J. Mol. Sci. 2020, 21, 1182. [Google Scholar] [CrossRef]
- Verkman, A.S.; Yang, B.; Song, Y.; Manley, G.T.; Ma, T. Role of Water Channels in Fluid Transport Studied by Phenotype Analysis of Aquaporin Knockout Mice. Exp. Physiol. 2000, 85, 233s–241s. [Google Scholar] [CrossRef]
- Yang, B.; Song, Y.; Zhao, D.; Verkman, A.S. Phenotype Analysis of Aquaporin-8 Null Mice. Am. J. Physiol. Cell Physiol. 2005, 288, C1161–C1170. [Google Scholar] [CrossRef]
- Parisis, D.; Chivasso, C.; Perret, J.; Soyfoo, M.S.; Delporte, C. Current State of Knowledge on Primary Sjögren’s Syndrome, an Autoimmune Exocrinopathy. J. Clin. Med. 2020, 9, E2299. [Google Scholar] [CrossRef] [PubMed]
- Yoshimura, S.; Nakamura, H.; Horai, Y.; Nakajima, H.; Shiraishi, H.; Hayashi, T.; Takahashi, T.; Kawakami, A. Abnormal Distribution of AQP5 in Labial Salivary Glands Is Associated with Poor Saliva Secretion in Patients with Sjögren’s Syndrome Including Neuromyelitis Optica Complicated Patients. Mod. Rheumatol. 2016, 26, 384–390. [Google Scholar] [CrossRef]
- Gresz, V.; Horvath, A.; Gera, I.; Nielsen, S.; Zelles, T. Immunolocalization of AQP5 in Resting and Stimulated Normal Labial Glands and in Sjögren’s Syndrome. Oral Dis. 2015, 21, e114–e120. [Google Scholar] [CrossRef] [PubMed]
- Beroukas, D.; Hiscock, J.; Jonsson, R.; Waterman, S.A.; Gordon, T.P. Subcellular Distribution of Aquaporin 5 in Salivary Glands in Primary Sjögren’s Syndrome. Lancet 2001, 358, 1875–1876. [Google Scholar] [CrossRef]
- Teos, L.Y.; Zhang, Y.; Cotrim, A.P.; Swaim, W.; Won, J.H.; Ambrus, J.; Shen, L.; Bebris, L.; Grisius, M.; Jang, S.-I.; et al. IP3R Deficit Underlies Loss of Salivary Fluid Secretion in Sjögren’s Syndrome. Sci. Rep. 2015, 5, 13953. [Google Scholar] [CrossRef]
- Konttinen, Y.T.; Tensing, E.-K.; Laine, M.; Porola, P.; Törnwall, J.; Hukkanen, M. Abnormal Distribution of Aquaporin-5 in Salivary Glands in the NOD Mouse Model for Sjögren’s Syndrome. J. Rheumatol. 2005, 32, 1071–1075. [Google Scholar]
- Soyfoo, M.S.; De Vriese, C.; Debaix, H.; Martin-Martinez, M.D.; Mathieu, C.; Devuyst, O.; Steinfeld, S.D.; Delporte, C. Modified Aquaporin 5 Expression and Distribution in Submandibular Glands from NOD Mice Displaying Autoimmune Exocrinopathy. Arthritis Rheum. 2007, 56, 2566–2574. [Google Scholar] [CrossRef]
- Soyfoo, M.S.; Konno, A.; Bolaky, N.; Oak, J.S.; Fruman, D.; Nicaise, C.; Takiguchi, M.; Delporte, C. Link between Inflammation and Aquaporin-5 Distribution in Submandibular Gland in Sjögren’s Syndrome? Oral Dis. 2012, 18, 568–574. [Google Scholar] [CrossRef] [PubMed]
- Satoh, K.; Narita, T.; Matsuki-Fukushima, M.; Okabayashi, K.; Ito, T.; Senpuku, H.; Sugiya, H. E2f1-Deficient NOD/SCID Mice Have Dry Mouth Due to a Change of Acinar/Duct Structure and the down-Regulation of AQP5 in the Salivary Gland. Pflug. Arch. 2013, 465, 271–281. [Google Scholar] [CrossRef]
- Saito, K.; Mori, S.; Kodama, T. McH-Lpr/Lpr-RA1 Mice: A Novel Spontaneous Mouse Model of Autoimmune Sialadenitis. Immunol. Lett. 2021, 237, 3–10. [Google Scholar] [CrossRef]
- Lin, X.; Song, J.; Shaw, P.-C.; Ng, T.-B.; Wong, R.N.-S.; Sze, S.C.-W.; Tong, Y.; Lee, K.-F.; Zhang, K.Y. An Autoimmunized Mouse Model Recapitulates Key Features in the Pathogenesis of Sjögren’s Syndrome. Int. Immunol. 2011, 23, 613–624. [Google Scholar] [CrossRef] [PubMed]
- Chivasso, C.; D’Agostino, C.; Parisis, D.; Soyfoo, M.S.; Delporte, C. Involvement of Aquaporin 5 in Sjögren’s Syndrome. Autoimmun. Rev. 2023, 22, 103268. [Google Scholar] [CrossRef] [PubMed]
- Limaye, A.; Hall, B.E.; Zhang, L.; Cho, A.; Prochazkova, M.; Zheng, C.; Walker, M.; Adewusi, F.; Burbelo, P.D.; Sun, Z.J.; et al. Targeted TNF-α Overexpression Drives Salivary Gland Inflammation. J. Dent. Res. 2019, 98, 713–719. [Google Scholar] [CrossRef]
- Miyagi, Y.; Kondo, Y.; Kusuda, Y.; Hori, Y.; Yamazaki, S.; Munemasa, T.; Mukaibo, T.; Masaki, C.; Hosokawa, R. Submandibular Gland-Specific Inflammaging-Induced Hyposalivation in the Male Senescence-Accelerated Mouse Prone -1 Line (SAM-P1). Biogerontology 2019, 20, 421–432. [Google Scholar] [CrossRef]
- Yamamura, Y.; Motegi, K.; Kani, K.; Takano, H.; Momota, Y.; Aota, K.; Yamanoi, T.; Azuma, M. TNF-α Inhibits Aquaporin 5 Expression in Human Salivary Gland Acinar Cells via Suppression of Histone H4 Acetylation. J. Cell Mol. Med. 2012, 16, 1766–1775. [Google Scholar] [CrossRef]
- Yao, C.; Purwanti, N.; Karabasil, M.R.; Azlina, A.; Javkhlan, P.; Hasegawa, T.; Akamatsu, T.; Hosoi, T.; Ozawa, K.; Hosoi, K. Potential Down-Regulation of Salivary Gland AQP5 by LPS via Cross-Coupling of NF-KappaB and p-c-Jun/c-Fos. Am. J. Pathol. 2010, 177, 724–734. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.H.; Gauna, A.E.; Perez, G.; Park, Y.; Pauley, K.M.; Kawai, T.; Cha, S. Autoantibodies against Muscarinic Type 3 Receptor in Sjögren’s Syndrome Inhibit Aquaporin 5 Trafficking. PLoS ONE 2013, 8, e53113. [Google Scholar] [CrossRef]
- Li, J.; Ha, Y.-M.; Kü, N.-Y.; Choi, S.-Y.; Lee, S.J.; Oh, S.B.; Kim, J.-S.; Lee, J.-H.; Lee, E.B.; Song, Y.W.; et al. Inhibitory Effects of Autoantibodies on the Muscarinic Receptors in Sjögren’s Syndrome. Lab. Investig. 2004, 84, 1430–1438. [Google Scholar] [CrossRef] [PubMed]
- Ambudkar, I. Calcium Signaling Defects Underlying Salivary Gland Dysfunction. Biochim. Biophys. Acta Mol. Cell Res. 2018, 1865, 1771–1777. [Google Scholar] [CrossRef]
- Jang, S.-I.; Tandon, M.; Teos, L.; Zheng, C.; Warner, B.M.; Alevizos, I. Dual Function of MiR-1248 Links Interferon Induction and Calcium Signaling Defects in Sjögren’s Syndrome. EBioMedicine 2019, 48, 526–538. [Google Scholar] [CrossRef]
- Xiao, F.; Du, W.; Zhu, X.; Tang, Y.; Liu, L.; Huang, E.; Deng, C.; Luo, C.; Han, M.; Chen, P.; et al. IL-17 Drives Salivary Gland Dysfunction via Inhibiting TRPC1-Mediated Calcium Movement in Sjögren’s Syndrome. Clin. Transl. Immunol. 2021, 10, e1277. [Google Scholar] [CrossRef]
- Chivasso, C.; Nesverova, V.; Järvå, M.; Blanchard, A.; Rose, K.L.; Öberg, F.K.; Wang, Z.; Martin, M.; Lhotellerie, F.; Zindy, E.; et al. Unraveling Human AQP5-PIP Molecular Interaction and Effect on AQP5 Salivary Glands Localization in SS Patients. Cells 2021, 10, 2108. [Google Scholar] [CrossRef]
- Chivasso, C.; Hagströmer, C.J.; Rose, K.L.; Lhotellerie, F.; Leblanc, L.; Wang, Z.; Moscato, S.; Chevalier, C.; Zindy, E.; Martin, M.; et al. Ezrin Is a Novel Protein Partner of Aquaporin-5 in Human Salivary Glands and Shows Altered Expression and Cellular Localization in Sjögren’s Syndrome. Int. J. Mol. Sci. 2021, 22, 9213. [Google Scholar] [CrossRef] [PubMed]
- Alam, J.; Koh, J.H.; Kim, N.; Kwok, S.-K.; Park, S.-H.; Song, Y.W.; Park, K.; Choi, Y. Detection of Autoantibodies against Aquaporin-5 in the Sera of Patients with Primary Sjögren’s Syndrome. Immunol. Res. 2016, 64, 848–856. [Google Scholar] [CrossRef]
- Tzartos, J.S.; Stergiou, C.; Daoussis, D.; Zisimopoulou, P.; Andonopoulos, A.P.; Zolota, V.; Tzartos, S.J. Antibodies to Aquaporins Are Frequent in Patients with Primary Sjögren’s Syndrome. Rheumatology 2017, 56, 2114–2122. [Google Scholar] [CrossRef] [PubMed]
- Jeon, S.; Lee, J.; Park, S.-H.; Kim, H.-D.; Choi, Y. Associations of Anti-Aquaporin 5 Autoantibodies with Serologic and Histopathological Features of Sjögren’s Syndrome. J. Clin. Med. 2019, 8, 1863. [Google Scholar] [CrossRef] [PubMed]
- Ring, T.; Kallenbach, M.; Praetorius, J.; Nielsen, S.; Melgaard, B. Successful Treatment of a Patient with Primary Sjögren’s Syndrome with Rituximab. Clin. Rheumatol. 2006, 25, 891–894. [Google Scholar] [CrossRef]
- Carubbi, F.; Cipriani, P.; Marrelli, A.; Benedetto, P.D.; Ruscitti, P.; Berardicurti, O.; Pantano, I.; Liakouli, V.; Alvaro, S.; Alunno, A.; et al. Efficacy and Safety of Rituximab Treatment in Early Primary Sjögren’s Syndrome: A Prospective, Multi-Center, Follow-up Study. Arthritis Res. Ther. 2013, 15, R172. [Google Scholar] [CrossRef]
- Sisto, M.; Lorusso, L.; Ingravallo, G.; Nico, B.; Ribatti, D.; Ruggieri, S.; Lofrumento, D.D.; Lisi, S. Abnormal Distribution of AQP4 in Minor Salivary Glands of Primary Sjögren’s Syndrome Patients. Autoimmunity 2017, 50, 202–210. [Google Scholar] [CrossRef]
- Alam, J.; Choi, Y.S.; Koh, J.H.; Kwok, S.-K.; Park, S.-H.; Song, Y.W.; Park, K.; Choi, Y. Detection of Autoantibodies against Aquaporin-1 in the Sera of Patients with Primary Sjögren’s Syndrome. Immune Netw. 2017, 17, 103–109. [Google Scholar] [CrossRef]
- Takagi, K.; Yamaguchi, K.; Sakurai, T.; Asari, T.; Hashimoto, K.; Terakawa, S. Secretion of Saliva in X-Irradiated Rat Submandibular Glands. Radiat. Res. 2003, 159, 351–360. [Google Scholar] [CrossRef]
- Choi, J.H.; Wu, H.-G.; Jung, K.C.; Lee, S.H.; Kwon, E.K. Apoptosis and Expression of AQP5 and TGF-Beta in the Irradiated Rat Submandibular Gland. Cancer Res. Treat. 2009, 41, 145–154. [Google Scholar] [CrossRef]
- Asari, T.; Maruyama, K.; Kusama, H. Salivation Triggered by Pilocarpine Involves Aquaporin-5 in Normal Rats but Not in Irradiated Rats. Clin. Exp. Pharmacol. Physiol. 2009, 36, 531–538. [Google Scholar] [CrossRef]
- Kim, J.H.; Jeong, B.K.; Jang, S.J.; Yun, J.W.; Jung, M.H.; Kang, K.M.; Kim, T.G.; Woo, S.H. Alpha-Lipoic Acid Ameliorates Radiation-Induced Salivary Gland Injury by Preserving Parasympathetic Innervation in Rats. Int. J. Mol. Sci. 2020, 21, 2260. [Google Scholar] [CrossRef] [PubMed]
- Araujo, M.V.T.; Spadella, M.A.; Chies, A.B.; Arruda, G.V.; Santos, T.d.M.; Cavariani, M.M.; Domeniconi, R.F. Effect of Low Radiation Dose on the Expression and Location of Aquaporins in Rat Submandibular Gland. Tissue Cell 2018, 53, 104–110. [Google Scholar] [CrossRef] [PubMed]
- Takakura, K.; Takaki, S.; Takeda, I.; Hanaue, N.; Kizu, Y.; Tonogi, M.; Yamane, G. Effect of Cevimeline on Radiation-Induced Salivary Gland Dysfunction and AQP5 in Submandibular Gland in Mice. Bull. Tokyo Dent. Coll. 2007, 48, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Zhao, D.; Gong, B.; Xu, Y.; Sun, H.; Yang, B.; Zhao, X. Decreased Saliva Secretion and Down-Regulation of AQP5 in Submandibular Gland in Irradiated Rats. Radiat. Res. 2006, 165, 678–687. [Google Scholar] [CrossRef]
- Wu, Y.-H.; Xu, H.; Yao, Q.-T.; Liu, S.-H.; Yakupu, A.; Lu, L.-D.; Shi, L. Effect of Ionizing Radiation on the Secretion of the Paracellular Pathway in Rat Submandibular Glands. Hua Xi Kou Qiang Yi Xue Za Zhi 2021, 39, 267–273. [Google Scholar] [CrossRef]
- Yao, Q.-T.; Wu, Y.-H.; Liu, S.-H.; Song, X.-B.; Xu, H.; Li, J.; Shi, L. Pilocarpine Improves Submandibular Gland Dysfunction in Irradiated Rats by Downregulating the Tight Junction Protein Claudin-4. Oral Dis. 2021, 28, 1528–1538. [Google Scholar] [CrossRef]
- Biswas, R.; Ahn, J.C.; Moon, J.H.; Kim, J.; Choi, Y.-H.; Park, S.Y.; Chung, P.-S. Low-Level Laser Therapy with 850 nm Recovers Salivary Function via Membrane Redistribution of Aquaporin 5 by Reducing Intracellular Ca2+ Overload and ER Stress during Hyperglycemia. Biochim. Biophys. Acta Gen. Subj. 2018, 1862, 1770–1780. [Google Scholar] [CrossRef]
- Soyfoo, M.S.; Bolaky, N.; Depoortere, I.; Delporte, C. Relationship between Aquaporin-5 Expression and Saliva Flow in Streptozotocin-Induced Diabetic Mice? Oral Dis. 2012, 18, 501–505. [Google Scholar] [CrossRef]
- Cui, F.; Hu, M.; Li, R.; Li, B.; Huang, D.; Ma, W.; Jia, X.; Lv, Z. Insulin on Changes in Expressions of Aquaporin-1, Aquaporin-5, and Aquaporin-8 in Submandibular Salivary Glands of Rats with Streptozotocin-Induced Diabetes. Int. J. Clin. Exp. Pathol. 2021, 14, 221–229. [Google Scholar]
- Wu, F.; Wang, J.; Sun, J.; Shen, L.; Liu, M.; Zhao, E. Procaine Stimulates Aquaporin-5 Expression in Human Salivary Gland Ductal Cells via the Suppression of DNA Methyltransferase-1. Mol. Med. Rep. 2018, 17, 7996–8002. [Google Scholar] [CrossRef]
- Wang, D.; Yuan, Z.; Inoue, N.; Cho, G.; Shono, M.; Ishikawa, Y. Abnormal Subcellular Localization of AQP5 and Downregulated AQP5 Protein in Parotid Glands of Streptozotocin-Induced Diabetic Rats. Biochim. Biophys. Acta 2011, 1810, 543–554. [Google Scholar] [CrossRef]
- Inoue, N.; Iida, H.; Yuan, Z.; Ishikawa, Y.; Ishida, H. Age-Related Decreases in the Response of Aquaporin-5 to Acetylcholine in Rat Parotid Glands. J. Dent. Res. 2003, 82, 476–480. [Google Scholar] [CrossRef]
- Ishikawa, Y.; Iida, H.; Ishida, H. The Muscarinic Acetylcholine Receptor-Stimulated Increase in Aquaporin-5 Levels in the Apical Plasma Membrane in Rat Parotid Acinar Cells Is Coupled with Activation of Nitric Oxide/CGMP Signal Transduction. Mol. Pharm. 2002, 61, 1423–1434. [Google Scholar] [CrossRef] [PubMed]
- Zeng, M.; Szymczak, M.; Ahuja, M.; Zheng, C.; Yin, H.; Swaim, W.; Chiorini, J.A.; Bridges, R.J.; Muallem, S. Restoration of CFTR Activity in Ducts Rescues Acinar Cell Function and Reduces Inflammation in Pancreatic and Salivary Glands of Mice. Gastroenterology 2017, 153, 1148–1159. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Wu, L.; Chen, Y.; Yan, Z.; Fu, J.; Luo, Z.; Du, J.; Guo, L.; Xu, J.; Liu, Y. Anticeramide Improves Sjögren’s Syndrome by Blocking BMP6-Induced Th1. J. Dent. Res. 2023, 102, 93–102. [Google Scholar] [CrossRef]
- Delporte, C.; O’Connell, B.C.; He, X.; Lancaster, H.E.; O’Connell, A.C.; Agre, P.; Baum, B.J. Increased Fluid Secretion after Adenoviral-Mediated Transfer of the Aquaporin-1 CDNA to Irradiated Rat Salivary Glands. Proc. Natl. Acad. Sci. USA 1997, 94, 3268–3273. [Google Scholar] [CrossRef] [PubMed]
- Baum, B.J.; Zheng, C.; Cotrim, A.P.; Goldsmith, C.M.; Atkinson, J.C.; Brahim, J.S.; Chiorini, J.A.; Voutetakis, A.; Leakan, R.A.; Van Waes, C.; et al. Transfer of the AQP1 CDNA for the Correction of Radiation-Induced Salivary Hypofunction. Biochim. Biophys. Acta 2006, 1758, 1071–1077. [Google Scholar] [CrossRef] [PubMed]
- Shan, Z.; Li, J.; Zheng, C.; Liu, X.; Fan, Z.; Zhang, C.; Goldsmith, C.M.; Wellner, R.B.; Baum, B.J.; Wang, S. Increased Fluid Secretion after Adenoviral-Mediated Transfer of the Human Aquaporin-1 CDNA to Irradiated Miniature Pig Parotid Glands. Mol. Ther. 2005, 11, 444–451. [Google Scholar] [CrossRef]
- O’Connell, A.C.; Baccaglini, L.; Fox, P.C.; O’Connell, B.C.; Kenshalo, D.; Oweisy, H.; Hoque, A.T.; Sun, D.; Herscher, L.L.; Braddon, V.R.; et al. Safety and Efficacy of Adenovirus-Mediated Transfer of the Human Aquaporin-1 CDNA to Irradiated Parotid Glands of Non-Human Primates. Cancer Gene Ther. 1999, 6, 505–513. [Google Scholar] [CrossRef] [PubMed]
- Baum, B.J.; Alevizos, I.; Zheng, C.; Cotrim, A.P.; Liu, S.; McCullagh, L.; Goldsmith, C.M.; Burbelo, P.D.; Citrin, D.E.; Mitchell, J.B.; et al. Early Responses to Adenoviral-Mediated Transfer of the Aquaporin-1 CDNA for Radiation-Induced Salivary Hypofunction. Proc. Natl. Acad. Sci. USA 2012, 109, 19403–19407. [Google Scholar] [CrossRef]
- Nakamura, H.; Tanaka, T.; Zheng, C.; Afione, S.A.; Warner, B.M.; Noguchi, M.; Atsumi, T.; Chiorini, J.A. Correction of LAMP3-Associated Salivary Gland Hypofunction by Aquaporin Gene Therapy. Sci. Rep. 2022, 12, 18570. [Google Scholar] [CrossRef]
- Lai, Z.; Yin, H.; Cabrera-Pérez, J.; Guimaro, M.C.; Afione, S.; Michael, D.G.; Glenton, P.; Patel, A.; Swaim, W.D.; Zheng, C.; et al. Aquaporin Gene Therapy Corrects Sjögren’s Syndrome Phenotype in Mice. Proc. Natl. Acad. Sci. USA 2016, 113, 5694–5699. [Google Scholar] [CrossRef]
- Wang, Z.; Wang, Y.; Wang, S.; Zhang, L.-R.; Zhang, N.; Cheng, Z.; Liu, Q.; Shields, K.J.; Hu, B.; Passineau, M.J. CRISPR-Cas9 HDR System Enhances AQP1 Gene Expression. Oncotarget 2017, 8, 111683–111696. [Google Scholar] [CrossRef]
- Engevik, A.C.; Kaji, I.; Goldenring, J.R. The Physiology of the Gastric Parietal Cell. Physiol. Rev. 2020, 100, 573–602. [Google Scholar] [CrossRef]
- Zhu, C.; Chen, Z.; Jiang, Z. Expression, Distribution and Role of Aquaporin Water Channels in Human and Animal Stomach and Intestines. Int. J. Mol. Sci. 2016, 17, E1399. [Google Scholar] [CrossRef] [PubMed]
- Mobasheri, A.; Wray, S.; Marples, D. Distribution of AQP2 and AQP3 Water Channels in Human Tissue Microarrays. J. Mol. Histol. 2005, 36, 1–14. [Google Scholar] [CrossRef]
- Laforenza, U. Water Channel Proteins in the Gastrointestinal Tract. Mol. Asp. Med. 2012, 33, 642–650. [Google Scholar] [CrossRef]
- Koyama, Y.; Yamamoto, T.; Tani, T.; Nihei, K.; Kondo, D.; Funaki, H.; Yaoita, E.; Kawasaki, K.; Sato, N.; Hatakeyama, K.; et al. Expression and Localization of Aquaporins in Rat Gastrointestinal Tract. Am. J. Physiol. 1999, 276, C621–C627. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Tola, V.B.; Fang, P.; Soybel, D.I.; Van Hoek, A.N. Partitioning of Aquaporin-4 Water Channel MRNA and Protein in Gastric Glands. Dig. Dis. Sci. 2003, 48, 2027–2036. [Google Scholar] [CrossRef] [PubMed]
- Fujita, A.; Horio, Y.; Nielsen, S.; Nagelhus, E.A.; Hata, F.; Ottersen, O.P.; Kurachi, Y. High-Resolution Immunogold Cytochemistry Indicates That AQP4 Is Concentrated along the Basal Membrane of Parietal Cell in Rat Stomach. FEBS Lett. 1999, 459, 305–309. [Google Scholar] [CrossRef] [PubMed]
- Misaka, T.; Abe, K.; Iwabuchi, K.; Kusakabe, Y.; Ichinose, M.; Miki, K.; Emori, Y.; Arai, S. A Water Channel Closely Related to Rat Brain Aquaporin 4 Is Expressed in Acid- and Pepsinogen-Secretory Cells of Human Stomach. FEBS Lett. 1996, 381, 208–212. [Google Scholar] [CrossRef] [PubMed]
- Carmosino, M.; Procino, G.; Tamma, G.; Mannucci, R.; Svelto, M.; Valenti, G. Trafficking and Phosphorylation Dynamics of AQP4 in Histamine-Treated Human Gastric Cells. Biol. Cell 2007, 99, 25–36. [Google Scholar] [CrossRef]
- Wang, K.S.; Komar, A.R.; Ma, T.; Filiz, F.; McLeroy, J.; Hoda, K.; Verkman, A.S.; Bastidas, J.A. Gastric Acid Secretion in Aquaporin-4 Knockout Mice. Am. J. Physiol. Gastrointest. Liver Physiol. 2000, 279, G448–G453. [Google Scholar] [CrossRef]
- Parvin, M.N.; Tsumura, K.; Akamatsu, T.; Kanamori, N.; Hosoi, K. Expression and Localization of AQP5 in the Stomach and Duodenum of the Rat. Biochim. Biophys. Acta 2002, 1542, 116–124. [Google Scholar] [CrossRef]
- Huang, Y.-H.; Zhou, X.-Y.; Wang, H.-M.; Xu, H.; Chen, J.; Lv, N.-H. Aquaporin 5 Promotes the Proliferation and Migration of Human Gastric Carcinoma Cells. Tumour. Biol. 2013, 34, 1743–1751. [Google Scholar] [CrossRef]
- Shen, L.; Zhu, Z.; Huang, Y.; Shu, Y.; Sun, M.; Xu, H.; Zhang, G.; Guo, R.; Wei, W.; Wu, W. Expression Profile of Multiple Aquaporins in Human Gastric Carcinoma and Its Clinical Significance. Biomed. Pharm. 2010, 64, 313–318. [Google Scholar] [CrossRef]
- Chen, J.; Wang, T.; Zhou, Y.-C.; Gao, F.; Zhang, Z.-H.; Xu, H.; Wang, S.-L.; Shen, L.-Z. Aquaporin 3 Promotes Epithelial-Mesenchymal Transition in Gastric Cancer. J. Exp. Clin. Cancer Res. 2014, 33, 38. [Google Scholar] [CrossRef] [PubMed]
- Lastraioli, E.; Iorio, J.; Arcangeli, A. Ion Channel Expression as Promising Cancer Biomarker. Biochim. Biophys. Acta 2015, 1848, 2685–2702. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Wang, Y.; Wang, S.; Shen, L. Hyperglycemia Promotes Human Gastric Carcinoma Progression via Aquaporin 3. Dig. Dis. Sci. 2015, 60, 2338–2345. [Google Scholar] [CrossRef] [PubMed]
- Thapa, S.; Chetry, M.; Huang, K.; Peng, Y.; Wang, J.; Wang, J.; Zhou, Y.; Shen, Y.; Xue, Y.; Ji, K. Significance of Aquaporins’ Expression in the Prognosis of Gastric Cancer. Biosci. Rep. 2018, 38, BSR20171687. [Google Scholar] [CrossRef] [PubMed]
- Shiozaki, A.; Marunaka, Y.; Otsuji, E. Roles of Ion and Water Channels in the Cell Death and Survival of Upper Gastrointestinal Tract Cancers. Front. Cell Dev. Biol. 2021, 9, 616933. [Google Scholar] [CrossRef]
- Wang, Z.; Wang, Y.; He, Y.; Zhang, N.; Chang, W.; Niu, Y. Aquaporin-1 Facilitates Proliferation and Invasion of Gastric Cancer Cells via GRB7-Mediated ERK and Ras Activation. Anim. Cells Syst. 2020, 24, 253–259. [Google Scholar] [CrossRef]
- Moosavi, M.-S.; Elham, Y. Aquaporins 1, 3 and 5 in Different Tumors, Their Expression, Prognosis Value and Role as New Therapeutic Targets. Pathol. Oncol. Res. 2019, 26, 615–625. [Google Scholar] [CrossRef]
- Ying, W.; Zheng, K.; Wu, Y.; Wang, O. Pannexin 1 Mediates Gastric Cancer Cell Epithelial-Mesenchymal Transition via Aquaporin 5. Biol. Pharm. Bull. 2021, 44, 1111–1119. [Google Scholar] [CrossRef]
- Jiang, B.; Li, Z.; Zhang, W.; Wang, H.; Zhi, X.; Feng, J.; Chen, Z.; Zhu, Y.; Yang, L.; Xu, H.; et al. MiR-874 Inhibits Cell Proliferation, Migration and Invasion through Targeting Aquaporin-3 in Gastric Cancer. J. Gastroenterol. 2014, 49, 1011–1025. [Google Scholar] [CrossRef]
- Zhu, H.; Wu, Y.; Kang, M.; Zhang, B. MiR-877 Suppresses Gastric Cancer Progression by Downregulating AQP3. J. Int. Med. Res. 2020, 48, 300060520903661. [Google Scholar] [CrossRef]
- Wang, Y.; Guo, Y.; Zhuang, T.; Xu, T.; Ji, M. SP1-Induced Upregulation of LncRNA LINC00659 Promotes Tumour Progression in Gastric Cancer by Regulating MiR-370/AQP3 Axis. Front. Endocrinol. 2022, 13, 936037. [Google Scholar] [CrossRef] [PubMed]
- Zhao, R.; He, B.; Bie, Q.; Cao, J.; Lu, H.; Zhang, Z.; Liang, J.; Wei, L.; Xiong, H.; Zhang, B. AQP5 Complements LGR5 to Determine the Fates of Gastric Cancer Stem Cells through Regulating ULK1 Ubiquitination. J. Exp. Clin. Cancer Res. 2022, 41, 322. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.; Zhang, X.; Wang, J.; Song, H.; Liu, W.; Luo, B. Molecular Mechanism of Aquaporin 3 (AQP3) Regulating by LMP2A and Its Crosstalk with 4E-BP1 via ERK Signaling Pathway in EBV-Associated Gastric Cancer. Virus Res. 2022, 322, 198947. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Xu, X.; Yang, H.; Wang, H.; Ouyang, Y.; Zhou, Y.; Peng, C.; Yuan, Z.; He, C.; Zeng, C.; et al. Activation of Aquaporin 5 by Carcinogenic Helicobacter Pylori Infection Promotes Epithelial-Mesenchymal Transition via the MEK/ERK Pathway. Helicobacter 2021, 26, e12842. [Google Scholar] [CrossRef] [PubMed]
- Wen, J.; Wang, Y.; Gao, C.; Zhang, G.; You, Q.; Zhang, W.; Zhang, Z.; Wang, S.; Peng, G.; Shen, L. Helicobacter Pylori Infection Promotes Aquaporin 3 Expression via the ROS-HIF-1α-AQP3-ROS Loop in Stomach Mucosa: A Potential Novel Mechanism for Cancer Pathogenesis. Oncogene 2018, 37, 3549–3561. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Wang, Z.; Li, X.; Liu, R.; Li, B.; Huang, L.; Chen, Y.; Zhang, C.; Zhang, H.; Li, Y.; et al. Total Glucosides of Paeony (TGP) Alleviates Constipation and Intestinal Inflammation in Mice Induced by Sjögren’s Syndrome. J. Ethnopharmacol. 2020, 260, 113056. [Google Scholar] [CrossRef]
- Zhao, Y.; Deng, Z.; Ma, Z.; Zhang, M.; Wang, H.; Tuo, B.; Li, T.; Liu, X. Expression Alteration and Dysfunction of Ion Channels/Transporters in the Parietal Cells Induces Gastric Diffused Mucosal Injury. Biomed. Pharm. 2022, 148, 112660. [Google Scholar] [CrossRef]
- Fukuhara, S.; Matsuzaki, J.; Tsugawa, H.; Masaoka, T.; Miyoshi, S.; Mori, H.; Fukushima, Y.; Yasui, M.; Kanai, T.; Suzuki, H. Mucosal Expression of Aquaporin-4 in the Stomach of Histamine Type 2 Receptor Knockout Mice and Helicobacter Pylori-Infected Mice. J. Gastroenterol. Hepatol. 2014, 29 (Suppl. S4), 53–59. [Google Scholar] [CrossRef]
- Jain, R.N.; Brunkan, C.S.; Chew, C.S.; Samuelson, L.C. Gene Expression Profiling of Gastrin Target Genes in Parietal Cells. Physiol. Genom. 2006, 24, 124–132. [Google Scholar] [CrossRef] [PubMed]
- Xia, J.; Wang, H.; Li, S.; Wu, Q.; Sun, L.; Huang, H.; Zeng, M. Ion Channels or Aquaporins as Novel Molecular Targets in Gastric Cancer. Mol. Cancer 2017, 16, 54. [Google Scholar] [CrossRef]
- Collaco, A.M.; Jakab, R.L.; Hoekstra, N.E.; Mitchell, K.A.; Brooks, A.; Ameen, N.A. Regulated Traffic of Anion Transporters in Mammalian Brunner’s Glands: A Role for Water and Fluid Transport. Am. J. Physiol. Gastrointest. Liver Physiol. 2013, 305, G258–G275. [Google Scholar] [CrossRef] [PubMed]
- Gustafsson, J.K.; Johansson, M.E.V. The Role of Goblet Cells and Mucus in Intestinal Homeostasis. Nat. Rev. Gastroenterol. Hepatol. 2022, 19, 785–803. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Yu, M. Role of Goblet Cells in Intestinal Barrier and Mucosal Immunity. J. Inflamm. Res. 2021, 14, 3171–3183. [Google Scholar] [CrossRef]
- Parvin, M.N.; Kurabuchi, S.; Murdiastuti, K.; Yao, C.; Kosugi-Tanaka, C.; Akamatsu, T.; Kanamori, N.; Hosoi, K. Subcellular Redistribution of AQP5 by Vasoactive Intestinal Polypeptide in the Brunner’s Gland of the Rat Duodenum. Am. J. Physiol. Gastrointest. Liver Physiol. 2005, 288, G1283–G1291. [Google Scholar] [CrossRef] [PubMed]
- Kirkegaard, P.; Lundberg, J.M.; Poulsen, S.S.; Olsen, P.S.; Fahrenkrug, J.; Hökfelt, T.; Christiansen, J. Vasoactive Intestinal Polypeptidergic Nerves and Brunner’s Gland Secretion in the Rat. Gastroenterology 1981, 81, 872–878. [Google Scholar] [CrossRef]
- Okada, S.; Misaka, T.; Matsumoto, I.; Watanabe, H.; Abe, K. Aquaporin-9 Is Expressed in a Mucus-Secreting Goblet Cell Subset in the Small Intestine. FEBS Lett. 2003, 540, 157–162. [Google Scholar] [CrossRef]
- Mahmoud, F.; Mullen, A.; Sainsbury, C.; Rushworth, G.F.; Yasin, H.; Abutheraa, N.; Mueller, T.; Kurdi, A. Meta-Analysis of Factors Associated with Antidiabetic Drug Prescribing for Type 2 Diabetes Mellitus. Eur. J. Clin. Investig. 2023, 53, e13997. [Google Scholar] [CrossRef]
- Boyer, J.L. Bile Formation and Secretion. Compr. Physiol. 2013, 3, 1035–1078. [Google Scholar] [CrossRef]
- Masyuk, A.I.; Masyuk, T.V.; Tietz, P.S.; Splinter, P.L.; LaRusso, N.F. Intrahepatic Bile Ducts Transport Water in Response to Absorbed Glucose. Am. J. Physiol. Cell Physiol. 2002, 283, C785–C791. [Google Scholar] [CrossRef]
- Müller, M.; Jansen, P.L. Molecular Aspects of Hepatobiliary Transport. Am. J. Physiol. 1997, 272, G1285–G1303. [Google Scholar] [CrossRef]
- Ma, T.; Verkman, A.S. Aquaporin Water Channels in Gastrointestinal Physiology. J. Physiol. 1999, 517 Pt 2, 317–326. [Google Scholar] [CrossRef] [PubMed]
- Vlahcevic, Z.R.; Jairath, S.K.; Heuman, D.M.; Stravitz, R.T.; Hylemon, P.B.; Avadhani, N.G.; Pandak, W.M. Transcriptional Regulation of Hepatic Sterol 27-Hydroxylase by Bile Acids. Am. J. Physiol. 1996, 270, G646–G652. [Google Scholar] [CrossRef]
- Sherlock, S.; Dooley, J. Diseases of the Liver and Biliary System, 11th ed.; Blackwell Science: Oxford, UK, 2002; ISBN 978-0-632-05582-1. [Google Scholar]
- Gregoire, F.; Lucidi, V.; Zerrad-Saadi, A.; Virreira, M.; Bolaky, N.; Delforge, V.; Lemmers, A.; Donckier, V.; Devière, J.; Demetter, P.; et al. Analysis of Aquaporin Expression in Liver with a Focus on Hepatocytes. Histochem. Cell Biol. 2015, 144, 347–363. [Google Scholar] [CrossRef]
- Calamita, G.; Ferri, D.; Bazzini, C.; Mazzone, A.; Bottà, G.; Liquori, G.E.; Paulmichl, M.; Portincasa, P.; Meyer, G.; Svelto, M. Expression and Subcellular Localization of the AQP8 and AQP1 Water Channels in the Mouse Gall-Bladder Epithelium. Biol. Cell 2005, 97, 415–423. [Google Scholar] [CrossRef] [PubMed]
- Calamita, G.; Mazzone, A.; Bizzoca, A.; Cavalier, A.; Cassano, G.; Thomas, D.; Svelto, M. Expression and Immunolocalization of the Aquaporin-8 Water Channel in Rat Gastrointestinal Tract. Eur. J. Cell Biol. 2001, 80, 711–719. [Google Scholar] [CrossRef]
- Portincasa, P.; Calamita, G. Water Channel Proteins in Bile Formation and Flow in Health and Disease: When Immiscible Becomes Miscible. Mol. Asp. Med. 2012, 33, 651–664. [Google Scholar] [CrossRef] [PubMed]
- Nihei, K.; Koyama, Y.; Tani, T.; Yaoita, E.; Ohshiro, K.; Adhikary, L.P.; Kurosaki, I.; Shirai, Y.; Hatakeyama, K.; Yamamoto, T. Immunolocalization of Aquaporin-9 in Rat Hepatocytes and Leydig Cells. Arch. Histol. Cytol. 2001, 64, 81–88. [Google Scholar] [CrossRef]
- Elkjaer, M.; Vajda, Z.; Nejsum, L.N.; Kwon, T.; Jensen, U.B.; Amiry-Moghaddam, M.; Frøkiaer, J.; Nielsen, S. Immunolocalization of AQP9 in Liver, Epididymis, Testis, Spleen, and Brain. Biochem. Biophys. Res. Commun. 2000, 276, 1118–1128. [Google Scholar] [CrossRef]
- Marinelli, R.A.; Tietz, P.S.; Pham, L.D.; Rueckert, L.; Agre, P.; LaRusso, N.F. Secretin Induces the Apical Insertion of Aquaporin-1 Water Channels in Rat Cholangiocytes. Am. J. Physiol. 1999, 276, G280–G286. [Google Scholar] [CrossRef] [PubMed]
- Ambe, P.C.; Gödde, D.; Zirngibl, H.; Störkel, S. Aquaporin-1 and 8 Expression in the Gallbladder Mucosa Might Not Be Associated with the Development of Gallbladder Stones in Humans. Eur. J. Clin. Investig. 2016, 46, 227–233. [Google Scholar] [CrossRef]
- Van Erpecum, K.J.; Wang, D.Q.-H.; Moschetta, A.; Ferri, D.; Svelto, M.; Portincasa, P.; Hendrickx, J.-J.; Schipper, M.; Calamita, G. Gallbladder Histopathology during Murine Gallstone Formation: Relation to Motility and Concentrating Function. J. Lipid Res. 2006, 47, 32–41. [Google Scholar] [CrossRef]
- Li, L.; Zhang, H.; Ma, T.; Verkman, A.S. Very High Aquaporin-1 Facilitated Water Permeability in Mouse Gallbladder. Am. J. Physiol. Gastrointest. Liver Physiol. 2009, 296, G816–G822. [Google Scholar] [CrossRef]
- Hung, K.-C.; Hsieh, P.-M.; Hsu, C.-Y.; Lin, C.-W.; Feng, G.-M.; Chen, Y.-S.; Hung, C.-H. Expression of Aquaporins in Rat Liver Regeneration. Scand. J. Gastroenterol. 2012, 47, 676–685. [Google Scholar] [CrossRef]
- Tardelli, M.; Moreno-Viedma, V.; Zeyda, M.; Itariu, B.K.; Langer, F.B.; Prager, G.; Stulnig, T.M. Adiponectin Regulates Aquaglyceroporin Expression in Hepatic Stellate Cells Altering Their Functional State. J. Gastroenterol. Hepatol. 2017, 32, 253–260. [Google Scholar] [CrossRef] [PubMed]
- Lee, P.J.; Park, H.-J.; Cho, N.; Kim, H.P. Aquaporin 11-Dependent Inhibition of Proliferation by Deuterium Oxide in Activated Hepatic Stellate Cells. Molecules 2018, 23, E3209. [Google Scholar] [CrossRef]
- Ferri, D.; Mazzone, A.; Liquori, G.E.; Cassano, G.; Svelto, M.; Calamita, G. Ontogeny, Distribution, and Possible Functional Implications of an Unusual Aquaporin, AQP8, in Mouse Liver. Hepatology 2003, 38, 947–957. [Google Scholar] [CrossRef] [PubMed]
- Morishita, Y.; Matsuzaki, T.; Hara-chikuma, M.; Andoo, A.; Shimono, M.; Matsuki, A.; Kobayashi, K.; Ikeda, M.; Yamamoto, T.; Verkman, A.; et al. Disruption of Aquaporin-11 Produces Polycystic Kidneys Following Vacuolization of the Proximal Tubule. Mol. Cell Biol. 2005, 25, 7770–7779. [Google Scholar] [CrossRef] [PubMed]
- Portincasa, P.; Moschetta, A.; Mazzone, A.; Palasciano, G.; Svelto, M.; Calamita, G. Water Handling and Aquaporins in Bile Formation: Recent Advances and Research Trends. J. Hepatol. 2003, 39, 864–874. [Google Scholar] [CrossRef] [PubMed]
- Huebert, R.C.; Splinter, P.L.; Garcia, F.; Marinelli, R.A.; LaRusso, N.F. Expression and Localization of Aquaporin Water Channels in Rat Hepatocytes. Evidence for a Role in Canalicular Bile Secretion. J. Biol. Chem. 2002, 277, 22710–22717. [Google Scholar] [CrossRef]
- Soria, L.R.; Fanelli, E.; Altamura, N.; Svelto, M.; Marinelli, R.A.; Calamita, G. Aquaporin-8-Facilitated Mitochondrial Ammonia Transport. Biochem. Biophys. Res. Commun. 2010, 393, 217–221. [Google Scholar] [CrossRef]
- Capiglioni, A.M.; Müller, G.L.; Marrone, J.; Alvarez, M.L.; Marinelli, R.A. Enhanced Ammonia Detoxification to Urea in Hepatocytes Transduced with Human Aquaporin-8 Gene. Biotechnol. Bioeng. 2021, 118, 4331–4337. [Google Scholar] [CrossRef]
- Calamita, G.; Gena, P.; Meleleo, D.; Ferri, D.; Svelto, M. Water Permeability of Rat Liver Mitochondria: A Biophysical Study. Biochim. Biophys. Acta 2006, 1758, 1018–1024. [Google Scholar] [CrossRef] [PubMed]
- Capiglioni, A.M.; Capitani, M.C.; Marrone, J.; Marinelli, R.A. Adenoviral Transfer of Human Aquaporin-8 Gene to Mouse Liver Improves Ammonia-Derived Ureagenesis. Cells 2023, 12, 1535. [Google Scholar] [CrossRef]
- Marinelli, R.A.; Marchissio, M.J. Mitochondrial Aquaporin-8: A Functional Peroxiporin? Antioxid. Redox Signal. 2013, 19, 896. [Google Scholar] [CrossRef] [PubMed]
- Marchissio, M.J.; Francés, D.E.A.; Carnovale, C.E.; Marinelli, R.A. Mitochondrial Aquaporin-8 Knockdown in Human Hepatoma HepG2 Cells Causes ROS-Induced Mitochondrial Depolarization and Loss of Viability. Toxicol. Appl. Pharm. 2012, 264, 246–254. [Google Scholar] [CrossRef] [PubMed]
- Danielli, M.; Capiglioni, A.M.; Marrone, J.; Marinelli, R.A. Further Evidence for the Involvement of Mitochondrial Aquaporin-8 in Hepatocyte Lipid Synthesis. Biochimie 2021, 188, 16–19. [Google Scholar] [CrossRef] [PubMed]
- Danielli, M.; Marrone, J.; Capiglioni, A.M.; Marinelli, R.A. Mitochondrial Aquaporin-8 Is Involved in SREBP-Controlled Hepatocyte Cholesterol Biosynthesis. Free Radic. Biol. Med. 2019, 131, 370–375. [Google Scholar] [CrossRef]
- Danielli, M.; Capiglioni, A.M.; Marrone, J.; Calamita, G.; Marinelli, R.A. Cholesterol Can Modulate Mitochondrial Aquaporin-8 Expression in Human Hepatic Cells. IUBMB Life 2017, 69, 341–346. [Google Scholar] [CrossRef]
- Vilchis-Landeros, M.; Guinzberg, R.; Riveros-Rosas, H.; Villalobos-Molina, R.; Piña, E. Aquaporin 8 Is Involved in H2 O2 -Mediated Differential Regulation of Metabolic Signaling by A1- and β-Adrenoceptors in Hepatocytes. FEBS Lett. 2020, 594, 1564–1576. [Google Scholar] [CrossRef]
- Gradilone, S.A.; Carreras, F.I.; Lehmann, G.L.; Marinelli, R.A. Phosphoinositide 3-Kinase Is Involved in the Glucagon-Induced Translocation of Aquaporin-8 to Hepatocyte Plasma Membrane. Biol. Cell 2005, 97, 831–836. [Google Scholar] [CrossRef]
- Gradilone, S.A.; García, F.; Huebert, R.C.; Tietz, P.S.; Larocca, M.C.; Kierbel, A.; Carreras, F.I.; Larusso, N.F.; Marinelli, R.A. Glucagon Induces the Plasma Membrane Insertion of Functional Aquaporin-8 Water Channels in Isolated Rat Hepatocytes. Hepatology 2003, 37, 1435–1441. [Google Scholar] [CrossRef]
- García, F.; Kierbel, A.; Larocca, M.C.; Gradilone, S.A.; Splinter, P.; LaRusso, N.F.; Marinelli, R.A. The Water Channel Aquaporin-8 Is Mainly Intracellular in Rat Hepatocytes, and Its Plasma Membrane Insertion Is Stimulated by Cyclic AMP. J. Biol. Chem. 2001, 276, 12147–12152. [Google Scholar] [CrossRef] [PubMed]
- Soria, L.R.; Gradilone, S.A.; Larocca, M.C.; Marinelli, R.A. Glucagon Induces the Gene Expression of Aquaporin-8 but Not That of Aquaporin-9 Water Channels in the Rat Hepatocyte. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2009, 296, R1274–R1281. [Google Scholar] [CrossRef] [PubMed]
- da Silva, I.V.; Garra, S.; Calamita, G.; Soveral, G. The Multifaceted Role of Aquaporin-9 in Health and Its Potential as a Clinical Biomarker. Biomolecules 2022, 12, 897. [Google Scholar] [CrossRef]
- Florio, M.; Engfors, A.; Gena, P.; Larsson, J.; Massaro, A.; Timpka, S.; Reimer, M.K.; Kjellbom, P.; Beitz, E.; Johanson, U.; et al. Characterization of the Aquaporin-9 Inhibitor RG100204 In Vitro and in Db/Db Mice. Cells 2022, 11, 3118. [Google Scholar] [CrossRef] [PubMed]
- Gena, P.; Buono, N.D.; D’Abbicco, M.; Mastrodonato, M.; Berardi, M.; Svelto, M.; Lopez, L.; Calamita, G. Dynamical Modeling of Liver Aquaporin-9 Expression and Glycerol Permeability in Hepatic Glucose Metabolism. Eur. J. Cell Biol. 2017, 96, 61–69. [Google Scholar] [CrossRef] [PubMed]
- Calamita, G.; Gena, P.; Ferri, D.; Rosito, A.; Rojek, A.; Nielsen, S.; Marinelli, R.A.; Frühbeck, G.; Svelto, M. Biophysical Assessment of Aquaporin-9 as Principal Facilitative Pathway in Mouse Liver Import of Glucogenetic Glycerol. Biol. Cell 2012, 104, 342–351. [Google Scholar] [CrossRef] [PubMed]
- Jelen, S.; Wacker, S.; Aponte-Santamaría, C.; Skott, M.; Rojek, A.; Johanson, U.; Kjellbom, P.; Nielsen, S.; de Groot, B.L.; Rützler, M. Aquaporin-9 Protein Is the Primary Route of Hepatocyte Glycerol Uptake for Glycerol Gluconeogenesis in Mice. J. Biol. Chem. 2011, 286, 44319–44325. [Google Scholar] [CrossRef]
- Rodríguez, A.; Catalán, V.; Gómez-Ambrosi, J.; García-Navarro, S.; Rotellar, F.; Valentí, V.; Silva, C.; Gil, M.J.; Salvador, J.; Burrell, M.A.; et al. Insulin- and Leptin-Mediated Control of Aquaglyceroporins in Human Adipocytes and Hepatocytes Is Mediated via the PI3K/Akt/MTOR Signaling Cascade. J. Clin. Endocrinol. Metab. 2011, 96, E586–E597. [Google Scholar] [CrossRef]
- Jelen, S.; Gena, P.; Lebeck, J.; Rojek, A.; Praetorius, J.; Frøkiaer, J.; Fenton, R.A.; Nielsen, S.; Calamita, G.; Rützler, M. Aquaporin-9 and Urea Transporter-A Gene Deletions Affect Urea Transmembrane Passage in Murine Hepatocytes. Am. J. Physiol. Gastrointest. Liver Physiol. 2012, 303, G1279–G1287. [Google Scholar] [CrossRef]
- Calamita, G.; Ferri, D.; Gena, P.; Carreras, F.I.; Liquori, G.E.; Portincasa, P.; Marinelli, R.A.; Svelto, M. Altered Expression and Distribution of Aquaporin-9 in the Liver of Rat with Obstructive Extrahepatic Cholestasis. Am. J. Physiol. Gastrointest. Liver Physiol. 2008, 295, G682–G690. [Google Scholar] [CrossRef] [PubMed]
- Kuriyama, H.; Shimomura, I.; Kishida, K.; Kondo, H.; Furuyama, N.; Nishizawa, H.; Maeda, N.; Matsuda, M.; Nagaretani, H.; Kihara, S.; et al. Coordinated Regulation of Fat-Specific and Liver-Specific Glycerol Channels, Aquaporin Adipose and Aquaporin 9. Diabetes 2002, 51, 2915–2921. [Google Scholar] [CrossRef]
- Carbrey, J.M.; Gorelick-Feldman, D.A.; Kozono, D.; Praetorius, J.; Nielsen, S.; Agre, P. Aquaglyceroporin AQP9: Solute Permeation and Metabolic Control of Expression in Liver. Proc. Natl. Acad. Sci. USA 2003, 100, 2945–2950. [Google Scholar] [CrossRef]
- Bernardino, R.L.; Marinelli, R.A.; Maggio, A.; Gena, P.; Cataldo, I.; Alves, M.G.; Svelto, M.; Oliveira, P.F.; Calamita, G. Hepatocyte and Sertoli cell Aquaporins, recent advances and research trends. Int. J. Mol. Sci. 2016, 17, 1096. [Google Scholar] [CrossRef] [PubMed]
- Rojek, A.M.; Skowronski, M.T.; Füchtbauer, E.-M.; Füchtbauer, A.C.; Fenton, R.A.; Agre, P.; Frøkiaer, J.; Nielsen, S. Defective Glycerol Metabolism in Aquaporin 9 (AQP9) Knockout Mice. Proc. Natl. Acad. Sci. USA 2007, 104, 3609–3614. [Google Scholar] [CrossRef]
- Rodríguez, A.; Gena, P.; Méndez-Giménez, L.; Rosito, A.; Valentí, V.; Rotellar, F.; Sola, I.; Moncada, R.; Silva, C.; Svelto, M.; et al. Reduced Hepatic Aquaporin-9 and Glycerol Permeability Are Related to Insulin Resistance in Non-Alcoholic Fatty Liver Disease. Int. J. Obes. 2014, 38, 1213–1220. [Google Scholar] [CrossRef] [PubMed]
- Gena, P.; Mastrodonato, M.; Portincasa, P.; Fanelli, E.; Mentino, D.; Rodríguez, A.; Marinelli, R.A.; Brenner, C.; Frühbeck, G.; Svelto, M.; et al. Liver Glycerol Permeability and Aquaporin-9 Are Dysregulated in a Murine Model of Non-Alcoholic Fatty Liver Disease. PLoS ONE 2013, 8, e78139. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, A.; Ezquerro, S.; Méndez-Giménez, L.; Becerril, S.; Frühbeck, G. Revisiting the Adipocyte: A Model for Integration of Cytokine Signaling in the Regulation of Energy Metabolism. Am. J. Physiol. Endocrinol. Metab. 2015, 309, E691–E714. [Google Scholar] [CrossRef]
- Baldini, F.; Portincasa, P.; Grasselli, E.; Damonte, G.; Salis, A.; Bonomo, M.; Florio, M.; Serale, N.; Voci, A.; Gena, P.; et al. Aquaporin-9 Is Involved in the Lipid-Lowering Activity of the Nutraceutical Silybin on Hepatocytes through Modulation of Autophagy and Lipid Droplets Composition. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2020, 1865, 158586. [Google Scholar] [CrossRef]
- Tesse, A.; Gena, P.; Rützler, M.; Calamita, G. Ablation of Aquaporin-9 Ameliorates the Systemic Inflammatory Response of LPS-Induced Endotoxic Shock in Mouse. Cells 2021, 10, 435. [Google Scholar] [CrossRef]
- Mohammad, S.; O’Riordan, C.E.; Verra, C.; Aimaretti, E.; Alves, G.F.; Dreisch, K.; Evenäs, J.; Gena, P.; Tesse, A.; Rützler, M.; et al. RG100204, A Novel Aquaporin-9 Inhibitor, Reduces Septic Cardiomyopathy and Multiple Organ Failure in Murine Sepsis. Front. Immunol. 2022, 13, 900906. [Google Scholar] [CrossRef] [PubMed]
- Ishibashi, K.; Tanaka, Y.; Morishita, Y. The Role of Mammalian Superaquaporins inside the Cell: An Update. Biochim. Biophys. Acta Biomembr. 2021, 1863, 183617. [Google Scholar] [CrossRef]
- Masyuk, T.; LaRusso, N. Polycystic Liver Disease: New Insights into Disease Pathogenesis. Hepatology 2006, 43, 906–908. [Google Scholar] [CrossRef]
- Marinelli, R.A.; Lehmann, G.L.; Soria, L.R.; Marchissio, M.J. Hepatocyte Aquaporins in Bile Formation and Cholestasis. Front. Biosci. 2011, 16, 2642–2652. [Google Scholar] [CrossRef] [PubMed]
- Marinelli, R.A.; LaRusso, N.F. Aquaporin Water Channels in Liver: Their Significance in Bile Formation. Hepatology 1997, 26, 1081–1084. [Google Scholar] [CrossRef]
- Tietz, P.S.; Marinelli, R.A.; Chen, X.-M.; Huang, B.; Cohn, J.; Kole, J.; McNiven, M.A.; Alper, S.; LaRusso, N.F. Agonist-Induced Coordinated Trafficking of Functionally Related Transport Proteins for Water and Ions in Cholangiocytes. J. Biol. Chem. 2003, 278, 20413–20419. [Google Scholar] [CrossRef] [PubMed]
- Marinelli, R.A.; Pham, L.D.; Tietz, P.S.; LaRusso, N.F. Expression of Aquaporin-4 Water Channels in Rat Cholangiocytes. Hepatology 2000, 31, 1313–1317. [Google Scholar] [CrossRef]
- Calamita, G.; Ferri, D.; Gena, P.; Liquori, G.E.; Marinelli, R.A.; Meyer, G.; Portincasa, P.; Svelto, M. Water Transport into Bile and Role in Bile Formation. Curr. Drug. Targets Immune Endocr. Metab. Disord. 2005, 5, 137–142. [Google Scholar] [CrossRef]
- Mennone, A.; Verkman, A.S.; Boyer, J.L. Unimpaired Osmotic Water Permeability and Fluid Secretion in Bile Duct Epithelia of AQP1 Null Mice. Am. J. Physiol. Gastrointest. Liver Physiol. 2002, 283, G739–G746. [Google Scholar] [CrossRef]
- Poling, H.M.; Mohanty, S.K.; Tiao, G.M.; Huppert, S.S. A Comprehensive Analysis of Aquaporin and Secretory Related Gene Expression in Neonate and Adult Cholangiocytes. Gene Expr. Patterns 2014, 15, 96–103. [Google Scholar] [CrossRef]
- Ueno, Y.; Alpini, G.; Yahagi, K.; Kanno, N.; Moritoki, Y.; Fukushima, K.; Glaser, S.; LeSage, G.; Shimosegawa, T. Evaluation of Differential Gene Expression by Microarray Analysis in Small and Large Cholangiocytes Isolated from Normal Mice. Liver Int. 2003, 23, 449–459. [Google Scholar] [CrossRef]
- Reuss, L.; Segal, Y.; Altenberg, G. Regulation of Ion Transport across Gallbladder Epithelium. Annu. Rev. Physiol. 1991, 53, 361–373. [Google Scholar] [CrossRef]
- Calamita, G.; Ferri, D.; Gena, P.; Liquori, G.E.; Cavalier, A.; Thomas, D.; Svelto, M. The Inner Mitochondrial Membrane Has Aquaporin-8 Water Channels and Is Highly Permeable to Water. J. Biol. Chem. 2005, 280, 17149–17153. [Google Scholar] [CrossRef] [PubMed]
- Swartz-Basile, D.A.; Lu, D.; Basile, D.P.; Graewin, S.J.; Al-Azzawi, H.; Kiely, J.M.; Mathur, A.; Yancey, K.; Pitt, H.A. Leptin Regulates Gallbladder Genes Related to Absorption and Secretion. Am. J. Physiol. Gastrointest. Liver Physiol. 2007, 293, G84–G90. [Google Scholar] [CrossRef] [PubMed]
- Sweed, N.; Kim, H.-J.; Hultenby, K.; Barros, R.; Parini, P.; Sancisi, V.; Strandvik, B.; Gabbi, C. Liver X Receptor β Regulates Bile Volume and the Expression of Aquaporins and Cystic Fibrosis Transmembrane Conductance Regulator in the Gallbladder. Am. J. Physiol. Gastrointest. Liver Physiol. 2021, 321, G243–G251. [Google Scholar] [CrossRef]
- Ribeiro, J.C.; Bernardino, R.L.; Carrageta, D.F.; Soveral, G.; Calamita, G.; Alves, M.G.; Oliveira, P.F. CFTR Modulates Aquaporin-Mediated Glycerol Permeability in Mouse Sertoli Cells. Cell Mol. Life Sci. 2022, 79, 592. [Google Scholar] [CrossRef]
- Goldblatt, M.I.; Swartz-Basile, D.A.; Svatek, C.L.; Nakeeb, A.; Pitt, H.A. Decreased Gallbladder Response in Leptin-Deficient Obese Mice. J. Gastrointest. Surg. 2002, 6, 438–442, discussion 443–444. [Google Scholar] [CrossRef] [PubMed]
- Zollner, G.; Trauner, M. Mechanisms of Cholestasis. Clin. Liver Dis. 2008, 12, 1–26. [Google Scholar] [CrossRef] [PubMed]
- Carreras, F.I.; Gradilone, S.A.; Mazzone, A.; García, F.; Huang, B.Q.; Ochoa, J.E.; Tietz, P.S.; Larusso, N.F.; Calamita, G.; Marinelli, R.A. Rat Hepatocyte Aquaporin-8 Water Channels Are down-Regulated in Extrahepatic Cholestasis. Hepatology 2003, 37, 1026–1033. [Google Scholar] [CrossRef] [PubMed]
- Carreras, F.I.; Lehmann, G.L.; Ferri, D.; Tioni, M.F.; Calamita, G.; Marinelli, R.A. Defective Hepatocyte Aquaporin-8 Expression and Reduced Canalicular Membrane Water Permeability in Estrogen-Induced Cholestasis. Am. J. Physiol. Gastrointest. Liver Physiol. 2007, 292, G905–G912. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, G.L.; Marinelli, R.A. Peritoneal Sepsis Downregulates Liver Expression of Aquaporin-8: A Water Channel Involved in Bile Secretion. Liver Int. 2009, 29, 317–318. [Google Scholar] [CrossRef]
- Lehmann, G.-L.; Larocca, M.-C.; Soria, L.-R.; Marinelli, R.-A. Aquaporins: Their Role in Cholestatic Liver Disease. World J. Gastroenterol. 2008, 14, 7059–7067. [Google Scholar] [CrossRef]
- Marrone, J.; Lehmann, G.L.; Soria, L.R.; Pellegrino, J.M.; Molinas, S.; Marinelli, R.A. Adenoviral Transfer of Human Aquaporin-1 Gene to Rat Liver Improves Bile Flow in Estrogen-Induced Cholestasis. Gene Ther. 2014, 21, 1058–1064. [Google Scholar] [CrossRef] [PubMed]
- Javitt, N.B. Hepatic Bile Formation: Bile Acid Transport and Water Flow into the Canalicular Conduit. Am. J. Physiol. Gastrointest. Liver Physiol. 2020, 319, G609–G618. [Google Scholar] [CrossRef] [PubMed]
- Marinelli, R.A.; Vore, M.; Javitt, N.B. Hepatic Bile Formation: Canalicular Osmolarity and Paracellular and Transcellular Water Flow. J. Pharm. Exp. Ther. 2019, 371, 713–717. [Google Scholar] [CrossRef]
- Matsumoto, K.; Imasato, M.; Yamazaki, Y.; Tanaka, H.; Watanabe, M.; Eguchi, H.; Nagano, H.; Hikita, H.; Tatsumi, T.; Takehara, T.; et al. Claudin 2 Deficiency Reduces Bile Flow and Increases Susceptibility to Cholesterol Gallstone Disease in Mice. Gastroenterology 2014, 147, 1134–1145.e10. [Google Scholar] [CrossRef] [PubMed]
- Banales, J.M.; Masyuk, T.V.; Bogert, P.S.; Huang, B.Q.; Gradilone, S.A.; Lee, S.-O.; Stroope, A.J.; Masyuk, A.I.; Medina, J.F.; LaRusso, N.F. Hepatic Cystogenesis Is Associated with Abnormal Expression and Location of Ion Transporters and Water Channels in an Animal Model of Autosomal Recessive Polycystic Kidney Disease. Am. J. Pathol. 2008, 173, 1637–1646. [Google Scholar] [CrossRef]
- Chen, X.-M.; O’Hara, S.P.; Huang, B.Q.; Splinter, P.L.; Nelson, J.B.; LaRusso, N.F. Localized Glucose and Water Influx Facilitates Cryptosporidium Parvum Cellular Invasion by Means of Modulation of Host-Cell Membrane Protrusion. Proc. Natl. Acad. Sci. USA 2005, 102, 6338–6343. [Google Scholar] [CrossRef]
- Portincasa, P.; Moschetta, A.; Palasciano, G. Cholesterol Gallstone Disease. Lancet 2006, 368, 230–239. [Google Scholar] [CrossRef]
- Sandler, R.S.; Everhart, J.E.; Donowitz, M.; Adams, E.; Cronin, K.; Goodman, C.; Gemmen, E.; Shah, S.; Avdic, A.; Rubin, R. The Burden of Selected Digestive Diseases in the United States. Gastroenterology 2002, 122, 1500–1511. [Google Scholar] [CrossRef]
- Wang, D.Q.-H.; Cohen, D.E.; Carey, M.C. Biliary Lipids and Cholesterol Gallstone Disease. J. Lipid Res. 2009, 50 (Suppl. 1), S406–S411. [Google Scholar] [CrossRef] [PubMed]
- Katsika, D.; Grjibovski, A.; Einarsson, C.; Lammert, F.; Lichtenstein, P.; Marschall, H.-U. Genetic and Environmental Influences on Symptomatic Gallstone Disease: A Swedish Study of 43,141 Twin Pairs. Hepatology 2005, 41, 1138–1143. [Google Scholar] [CrossRef] [PubMed]
- Portincasa, P.; Di Ciaula, A.; Wang, H.H.; Palasciano, G.; van Erpecum, K.J.; Moschetta, A.; Wang, D.Q.-H. Coordinate Regulation of Gallbladder Motor Function in the Gut-Liver Axis. Hepatology 2008, 47, 2112–2126. [Google Scholar] [CrossRef] [PubMed]
- Kube, I.; Kowalczyk, M.; Hofmann, U.; Ghallab, A.; Hengstler, J.G.; Führer, D.; Zwanziger, D. Hepatobiliary Thyroid Hormone Deficiency Impacts Bile Acid Hydrophilicity and Aquaporins in Cholestatic C57BL/6J Mice. Int. J. Mol. Sci. 2022, 23, 12355. [Google Scholar] [CrossRef]
- Calamita, G.; Delporte, C.; Marinelli, R.A. Hepatobiliary, salivary glands and pancreatic aquaporins in health and disease. In Aquaporins in Health and Disease: New Molecular Targets for Drug Discovery; Soveral, G., Nielsen, S., Casini, A., Eds.; CRC Press: Boca Raton, FL, USA, 2016; p. 183.e205. ISBN 9781138894013. [Google Scholar]
- Marrone, J.; Danielli, M.; Gaspari, C.I.; Capiglioni, A.M.; Marinelli, R.A. Aquaporin Gene Transfer for Hepatocellular Cholestasis. Biochimie 2021, 188, 12–15. [Google Scholar] [CrossRef]
- Marrone, J.; Soria, L.R.; Danielli, M.; Lehmann, G.L.; Larocca, M.C.; Marinelli, R.A. Hepatic Gene Transfer of Human Aquaporin-1 Improves Bile Salt Secretory Failure in Rats with Estrogen-Induced Cholestasis. Hepatology 2016, 64, 535–548. [Google Scholar] [CrossRef]
- Marrone, J.; Danielli, M.; Gaspari, C.I.; Marinelli, R.A. Adenovirus-Mediated Human Aquaporin-1 Expression in Hepatocytes Improves Lipopolysaccharide-Induced Cholestasis. IUBMB Life 2017, 69, 978–984. [Google Scholar] [CrossRef]
- Marrone, J.; Tocchetti, G.N.; Danielli, M.; Mottino, A.D.; Marinelli, R.A. Improved Hepatic MRP2/ABCC2 Transport Activity in LPS-Induced Cholestasis by Aquaporin-1 Gene Transfer. Biochimie 2019, 165, 179–182. [Google Scholar] [CrossRef]
- Konturek, S.J.; Zabielski, R.; Konturek, J.W.; Czarnecki, J. Neuroendocrinology of the Pancreas; Role of Brain-Gut Axis in Pancreatic Secretion. Eur. J. Pharmacol. 2003, 481, 1–14. [Google Scholar] [CrossRef]
- Burghardt, B.; Elkaer, M.-L.; Kwon, T.-H.; Rácz, G.Z.; Varga, G.; Steward, M.C.; Nielsen, S. Distribution of Aquaporin Water Channels AQP1 and AQP5 in the Ductal System of the Human Pancreas. Gut 2003, 52, 1008–1016. [Google Scholar] [CrossRef]
- Isokpehi, R.D.; Rajnarayanan, R.V.; Jeffries, C.D.; Oyeleye, T.O.; Cohly, H.H.P. Integrative Sequence and Tissue Expression Profiling of Chicken and Mammalian Aquaporins. BMC Genom. 2009, 10 (Suppl. S2), S7. [Google Scholar] [CrossRef]
- Cho, S.-J.; Sattar, A.K.M.A.; Jeong, E.-H.; Satchi, M.; Cho, J.A.; Dash, S.; Mayes, M.S.; Stromer, M.H.; Jena, B.P. Aquaporin 1 Regulates GTP-Induced Rapid Gating of Water in Secretory Vesicles. Proc. Natl. Acad. Sci. USA 2002, 99, 4720–4724. [Google Scholar] [CrossRef]
- Abu-Hamdah, R.; Cho, W.-J.; Cho, S.-J.; Jeremic, A.; Kelly, M.; Ilie, A.E.; Jena, B.P. Regulation of the Water Channel Aquaporin-1: Isolation and Reconstitution of the Regulatory Complex. Cell Biol. Int. 2004, 28, 7–17. [Google Scholar] [CrossRef]
- Hurley, P.T.; Ferguson, C.J.; Kwon, T.H.; Andersen, M.L.; Norman, A.G.; Steward, M.C.; Nielsen, S.; Case, R.M. Expression and Immunolocalization of Aquaporin Water Channels in Rat Exocrine Pancreas. Am. J. Physiol. Gastrointest. Liver Physiol. 2001, 280, G701–G709. [Google Scholar] [CrossRef] [PubMed]
- Furuya, S.; Naruse, S.; Ko, S.B.H.; Ishiguro, H.; Yoshikawa, T.; Hayakawa, T. Distribution of Aquaporin 1 in the Rat Pancreatic Duct System Examined with Light- and Electron-Microscopic Immunohistochemistry. Cell Tissue Res. 2002, 308, 75–86. [Google Scholar] [CrossRef] [PubMed]
- Ko, S.B.H.; Naruse, S.; Kitagawa, M.; Ishiguro, H.; Furuya, S.; Mizuno, N.; Wang, Y.; Yoshikawa, T.; Suzuki, A.; Shimano, S.; et al. Aquaporins in Rat Pancreatic Interlobular Ducts. Am. J. Physiol. Gastrointest. Liver Physiol. 2002, 282, G324–G331. [Google Scholar] [CrossRef]
- Burghardt, B.; Nielsen, S.; Steward, M.C. The Role of Aquaporin Water Channels in Fluid Secretion by the Exocrine Pancreas. J. Membr. Biol. 2006, 210, 143–153. [Google Scholar] [CrossRef]
- Itoh, T.; Rai, T.; Kuwahara, M.; Ko, S.B.H.; Uchida, S.; Sasaki, S.; Ishibashi, K. Identification of a Novel Aquaporin, AQP12, Expressed in Pancreatic Acinar Cells. Biochem. Biophys. Res. Commun. 2005, 330, 832–838. [Google Scholar] [CrossRef] [PubMed]
- McManaman, J.L.; Reyland, M.E.; Thrower, E.C. Secretion and Fluid Transport Mechanisms in the Mammary Gland: Comparisons with the Exocrine Pancreas and the Salivary Gland. J. Mammary Gland. Biol. Neoplasia 2006, 11, 249–268. [Google Scholar] [CrossRef]
- Ohta, E.; Itoh, T.; Nemoto, T.; Kumagai, J.; Ko, S.B.H.; Ishibashi, K.; Ohno, M.; Uchida, K.; Ohta, A.; Sohara, E.; et al. Pancreas-Specific Aquaporin 12 Null Mice Showed Increased Susceptibility to Caerulein-Induced Acute Pancreatitis. Am. J. Physiol. Cell Physiol. 2009, 297, C1368–C1378. [Google Scholar] [CrossRef]
- Eiseler, K.; Dropmann, L.M.; Bugert, P.; Ewers, M.; Witt, H. Genetic Analysis of the Aquaporin Water Channels AQP12A and AQP12B in Patients with Chronic Pancreatitis. Pancreatology 2022, 22, 1079–1083. [Google Scholar] [CrossRef]
- Arsenijevic, T.; Perret, J.; Van Laethem, J.-L.; Delporte, C. Aquaporins Involvement in Pancreas Physiology and in Pancreatic Diseases. Int. J. Mol. Sci. 2019, 20, 5052. [Google Scholar] [CrossRef]
- Nagaraju, G.P.; Basha, R.; Rajitha, B.; Alese, O.B.; Alam, A.; Pattnaik, S.; El-Rayes, B. Aquaporins: Their Role in Gastrointestinal Malignancies. Cancer Lett. 2016, 373, 12–18. [Google Scholar] [CrossRef]
- Direito, I.; Paulino, J.; Vigia, E.; Brito, M.A.; Soveral, G. Differential Expression of Aquaporin-3 and Aquaporin-5 in Pancreatic Ductal Adenocarcinoma. J. Surg. Oncol. 2017, 115, 980–996. [Google Scholar] [CrossRef]
- Feng, D.; Peng, W.; Chen, Y.; Chen, T.; Tian, J.; Si, H.; Cai, J.; Rao, Y.; Han, F.; Zhao, R. Down-Regulation of Aquaporin 1 in Rats with Experimental Acute Necrotizing Pancreatitis. Pancreas 2012, 41, 1092–1098. [Google Scholar] [CrossRef]
- Gabbi, C.; Kim, H.-J.; Hultenby, K.; Bouton, D.; Toresson, G.; Warner, M.; Gustafsson, J.-A. Pancreatic Exocrine Insufficiency in LXRbeta-/- Mice Is Associated with a Reduction in Aquaporin-1 Expression. Proc. Natl. Acad. Sci. USA 2008, 105, 15052–15057. [Google Scholar] [CrossRef]
- Xu, J.; Huang, B.; Wang, Y.; Tong, C.; Xie, P.; Fan, R.; Gao, Z. Emodin Ameliorates Acute Lung Injury Induced by Severe Acute Pancreatitis through the Up-Regulated Expressions of AQP1 and AQP5 in Lung. Clin. Exp. Pharmacol. Physiol. 2016, 43, 1071–1079. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.; Zhang, B.; Chen, Q.; Zhang, J.; Lei, B.; Li, B.; Wei, Y.; Zhai, R.; Liang, Z.; He, S.; et al. The Mechanism Underlying Alpinetin-Mediated Alleviation of Pancreatitis-Associated Lung Injury through Upregulating Aquaporin-1. Drug. Des. Devel. Ther. 2016, 10, 841–850. [Google Scholar] [CrossRef] [PubMed]
- Xie, R.; Wang, J.; Yao, Y.; Qi, M.; Huang, S.; Zhao, Z.; Chen, Y.; Yang, Z.; Sheng, H.; Fei, J.; et al. Fluid Resuscitation via the Rectum Ameliorates Hemodynamic Disorders through Adjusting Aquaporin Expression in an Experimental Severe Acute Pancreatitis Model. Exp. Ther. Med. 2019, 17, 437–443. [Google Scholar] [CrossRef] [PubMed]
- Kamimura, K.; Yokoo, T.; Terai, S. Gene Therapy for Pancreatic Diseases: Current Status. Int. J. Mol. Sci. 2018, 19, 3415. [Google Scholar] [CrossRef]
- Delporte, C.; O’Connell, B.C.; He, X.; Ambudkar, I.S.; Agre, P.; Baum, B.J. Adenovirus-Mediated Expression of Aquaporin-5 in Epithelial Cells. J. Biol. Chem. 1996, 271, 22070–22075. [Google Scholar] [CrossRef] [PubMed]
- Saint-Criq, V.; Gray, M.A. Role of CFTR in Epithelial Physiology. Cell Mol. Life Sci. 2017, 74, 93–115. [Google Scholar] [CrossRef] [PubMed]
- Hegyi, P.; Seidler, U.; Kunzelmann, K. CFTR-beyond the Airways: Recent Findings on the Role of the CFTR Channel in the Pancreas, the Intestine and the Kidneys. J. Cyst. Fibros. 2023, 22 (Suppl. S1), S17–S22. [Google Scholar] [CrossRef]
- Venglovecz, V.; Pallagi, P.; Kemény, L.V.; Balázs, A.; Balla, Z.; Becskeházi, E.; Gál, E.; Tóth, E.; Zvara, Á.; Puskás, L.G.; et al. The Importance of Aquaporin 1 in Pancreatitis and Its Relation to the CFTR Cl- Channel. Front. Physiol. 2018, 9, 854. [Google Scholar] [CrossRef] [PubMed]
- Ko, S.B.H.; Yamamoto, A.; Azuma, S.; Song, H.; Kamimura, K.; Nakakuki, M.; Gray, M.A.; Becq, F.; Ishiguro, H.; Goto, H. Effects of CFTR Gene Silencing by SiRNA or the Luminal Application of a CFTR Activator on Fluid Secretion from Guinea-Pig Pancreatic Duct Cells. Biochem. Biophys. Res. Commun. 2011, 410, 904–909. [Google Scholar] [CrossRef]
- Papadopoulos, M.C.; Saadoun, S. Key Roles of Aquaporins in Tumor Biology. Biochim. Biophys. Acta 2015, 1848, 2576–2583. [Google Scholar] [CrossRef]
- Verkman, A.S.; Hara-Chikuma, M.; Papadopoulos, M.C. Aquaporins—New Players in Cancer Biology. J. Mol. Med. 2008, 86, 523–529. [Google Scholar] [CrossRef]
- Tomita, Y.; Dorward, H.; Yool, A.J.; Smith, E.; Townsend, A.R.; Price, T.J.; Hardingham, J.E. Role of Aquaporin 1 Signalling in Cancer Development and Progression. Int. J. Mol. Sci. 2017, 18, 299. [Google Scholar] [CrossRef]
- Dajani, S.; Saripalli, A.; Sharma-Walia, N. Water Transport Proteins-Aquaporins (AQPs) in Cancer Biology. Oncotarget 2018, 9, 36392–36405. [Google Scholar] [CrossRef]
- Morishita, K.; Watanabe, K.; Ichijo, H. Cell Volume Regulation in Cancer Cell Migration Driven by Osmotic Water Flow. Cancer Sci. 2019, 110, 2337–2347. [Google Scholar] [CrossRef]
- Wang, L.; Zhang, Y.; Wu, X.; Yu, G. Aquaporins: New Targets for Cancer Therapy. Technol. Cancer Res. Treat. 2016, 15, 821–828. [Google Scholar] [CrossRef] [PubMed]
- Moon, C.S.; Moon, D.; Kang, S.K. Aquaporins in Cancer Biology. Front. Oncol. 2022, 12, 782829. [Google Scholar] [CrossRef]
- Smith, I.M.; Banerjee, S.; Moses, A.K.; Stroka, K.M. Aquaporin-Mediated Dysregulation of Cell Migration in Disease States. Cell Mol. Life Sci. 2023, 80, 48. [Google Scholar] [CrossRef] [PubMed]
- Bruun-Sørensen, A.S.; Edamana, S.; Login, F.H.; Borgquist, S.; Nejsum, L.N. Aquaporins in Pancreatic Ductal Adenocarcinoma. APMIS 2021, 129, 700–705. [Google Scholar] [CrossRef]
- Schnipper, J.; Dhennin-Duthille, I.; Ahidouch, A.; Ouadid-Ahidouch, H. Ion Channel Signature in Healthy Pancreas and Pancreatic Ductal Adenocarcinoma. Front. Pharmacol. 2020, 11, 568993. [Google Scholar] [CrossRef] [PubMed]
- Silva, P.M.; da Silva, I.V.; Sarmento, M.J.; Silva, Í.C.; Carvalho, F.A.; Soveral, G.; Santos, N.C. Aquaporin-3 and Aquaporin-5 Facilitate Migration and Cell-Cell Adhesion in Pancreatic Cancer by Modulating Cell Biomechanical Properties. Cells 2022, 11, 1308. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Huang, L.; Shao, M. Aquaporin 3 Facilitates Tumor Growth in Pancreatic Cancer by Modulating MTOR Signaling. Biochem. Biophys. Res. Commun. 2017, 486, 1097–1102. [Google Scholar] [CrossRef]
- Zou, W.; Yang, Z.; Li, D.; Liu, Z.; Zou, Q.; Yuan, Y. AQP1 and AQP3 Expression Are Associated with Severe Symptoms and Poor-Prognosis of the Pancreatic Ductal Adenocarcinoma. Appl. Immunohistochem. Mol. Morphol. 2019, 27, 40–47. [Google Scholar] [CrossRef]
- Zaccagnino, A.; Pilarsky, C.; Tawfik, D.; Sebens, S.; Trauzold, A.; Novak, I.; Schwab, A.; Kalthoff, H. In Silico Analysis of the Transportome in Human Pancreatic Ductal Adenocarcinoma. Eur. Biophys. J. 2016, 45, 749–763. [Google Scholar] [CrossRef]
- Langerhans, P. Beiträge zur Mikroscopischen Anatomie der Bauchspeichel Druse. In Inaugural-Dissertation; Gustav Lange, Inc.: Berlin, Germany, 1869. [Google Scholar]
- Henquin, J.C. Regulation of Insulin Secretion: A Matter of Phase Control and Amplitude Modulation. Diabetologia 2009, 52, 739–751. [Google Scholar] [CrossRef]
- Sener, A.; Malaisse, W.J. The Stimulus-Secretion Coupling of Amino Acid-Induced Insulin Release. Insulinotropic Action of L-Alanine. Biochim. Biophys. Acta 2002, 1573, 100–104. [Google Scholar] [CrossRef] [PubMed]
- Miley, H.E.; Sheader, E.A.; Brown, P.D.; Best, L. Glucose-Induced Swelling in Rat Pancreatic Beta-Cells. J. Physiol. 1997, 504 Pt 1, 191–198. [Google Scholar] [CrossRef]
- Best, L.; Miley, H.E.; Yates, A.P. Activation of an Anion Conductance and Beta-Cell Depolarization during Hypotonically Induced Insulin Release. Exp. Physiol. 1996, 81, 927–933. [Google Scholar] [CrossRef] [PubMed]
- Drews, G.; Krippeit-Drews, P.; Düfer, M. Electrophysiology of Islet Cells. Adv. Exp. Med. Biol. 2010, 654, 115–163. [Google Scholar] [CrossRef]
- Huang, P.; Venskutonytė, R.; Prasad, R.B.; Ardalani, H.; de Maré, S.W.; Fan, X.; Li, P.; Spégel, P.; Yan, N.; Gourdon, P.; et al. Cryo-EM Structure Supports a Role of AQP7 as a Junction Protein. Nat. Commun. 2023, 14, 600. [Google Scholar] [CrossRef] [PubMed]
- Best, L.; Brown, P.D.; Yates, A.P.; Perret, J.; Virreira, M.; Beauwens, R.; Malaisse, W.J.; Sener, A.; Delporte, C. Contrasting Effects of Glycerol and Urea Transport on Rat Pancreatic Beta-Cell Function. Cell Physiol. Biochem. 2009, 23, 255–264. [Google Scholar] [CrossRef]
- Louchami, K.; Best, L.; Brown, P.; Virreira, M.; Hupkens, E.; Perret, J.; Devuyst, O.; Uchida, S.; Delporte, C.; Malaisse, W.J.; et al. A New Role for Aquaporin 7 in Insulin Secretion. Cell Physiol. Biochem. 2012, 29, 65–74. [Google Scholar] [CrossRef]
- Matsumura, K.; Chang, B.H.-J.; Fujimiya, M.; Chen, W.; Kulkarni, R.N.; Eguchi, Y.; Kimura, H.; Kojima, H.; Chan, L. Aquaporin 7 Is a Beta-Cell Protein and Regulator of Intraislet Glycerol Content and Glycerol Kinase Activity, Beta-Cell Mass, and Insulin Production and Secretion. Mol. Cell Biol. 2007, 27, 6026–6037. [Google Scholar] [CrossRef]
- Méndez-Giménez, L.; Becerril, S.; Camões, S.P.; da Silva, I.V.; Rodrigues, C.; Moncada, R.; Valentí, V.; Catalán, V.; Gómez-Ambrosi, J.; Miranda, J.P.; et al. Role of Aquaporin-7 in Ghrelin- and GLP-1-Induced Improvement of Pancreatic β-Cell Function after Sleeve Gastrectomy in Obese Rats. Int. J. Obes. 2017, 41, 1394–1402. [Google Scholar] [CrossRef]
- Hibuse, T.; Maeda, N.; Funahashi, T.; Yamamoto, K.; Nagasawa, A.; Mizunoya, W.; Kishida, K.; Inoue, K.; Kuriyama, H.; Nakamura, T.; et al. Aquaporin 7 Deficiency Is Associated with Development of Obesity through Activation of Adipose Glycerol Kinase. Proc. Natl. Acad. Sci. USA 2005, 102, 10993–10998. [Google Scholar] [CrossRef]
- Skowronski, M.T.; Lebeck, J.; Rojek, A.; Praetorius, J.; Füchtbauer, E.-M.; Frøkiaer, J.; Nielsen, S. AQP7 Is Localized in Capillaries of Adipose Tissue, Cardiac and Striated Muscle: Implications in Glycerol Metabolism. Am. J. Physiol. Ren. Physiol. 2007, 292, F956–F965. [Google Scholar] [CrossRef]
- Delporte, C.; Virreira, M.; Crutzen, R.; Louchami, K.; Sener, A.; Malaisse, W.J.; Beauwens, R. Functional Role of Aquaglyceroporin 7 Expression in the Pancreatic Beta-Cell Line BRIN-BD11. J. Cell Physiol. 2009, 221, 424–429. [Google Scholar] [CrossRef] [PubMed]
- Virreira, M.; Perret, J.; Delporte, C. Pancreatic Beta-Cells: Role of Glycerol and Aquaglyceroporin 7. Int. J. Biochem. Cell Biol. 2011, 43, 10–13. [Google Scholar] [CrossRef]
- Dissanayake, W.C.; Sorrenson, B.; Shepherd, P.R. The Role of Adherens Junction Proteins in the Regulation of Insulin Secretion. Biosci. Rep. 2018, 38, BSR20170989. [Google Scholar] [CrossRef] [PubMed]
- Cottle, L.; Gan, W.J.; Gilroy, I.; Samra, J.S.; Gill, A.J.; Loudovaris, T.; Thomas, H.E.; Hawthorne, W.J.; Kebede, M.A.; Thorn, P. Structural and Functional Polarisation of Human Pancreatic Beta Cells in Islets from Organ Donors with and without Type 2 Diabetes. Diabetologia 2021, 64, 618–629. [Google Scholar] [CrossRef] [PubMed]
- Tsalamandris, S.; Antonopoulos, A.S.; Oikonomou, E.; Papamikroulis, G.-A.; Vogiatzi, G.; Papaioannou, S.; Deftereos, S.; Tousoulis, D. The Role of Inflammation in Diabetes: Current Concepts and Future Perspectives. Eur. Cardiol. 2019, 14, 50–59. [Google Scholar] [CrossRef]
- da Silva, I.V.; Cardoso, C.; Méndez-Giménez, L.; Camoes, S.P.; Frühbeck, G.; Rodríguez, A.; Miranda, J.P.; Soveral, G. Aquaporin-7 and Aquaporin-12 Modulate the Inflammatory Phenotype of Endocrine Pancreatic Beta-Cells. Arch. Biochem. Biophys. 2020, 691, 108481. [Google Scholar] [CrossRef]
- Krüger, C.; Jörns, A.; Kaynert, J.; Waldeck-Weiermair, M.; Michel, T.; Elsner, M.; Lenzen, S. The Importance of Aquaporin-8 for Cytokine-Mediated Toxicity in Rat Insulin-Producing Cells. Free Radic. Biol. Med. 2021, 174, 135–143. [Google Scholar] [CrossRef]
- Lindgren, C.M.; Mahtani, M.M.; Widén, E.; McCarthy, M.I.; Daly, M.J.; Kirby, A.; Reeve, M.P.; Kruglyak, L.; Parker, A.; Meyer, J.; et al. Genomewide Search for Type 2 Diabetes Mellitus Susceptibility Loci in Finnish Families: The Botnia Study. Am. J. Hum. Genet. 2002, 70, 509–516. [Google Scholar] [CrossRef]
- Loos, R.J.F.; Katzmarzyk, P.T.; Rao, D.C.; Rice, T.; Leon, A.S.; Skinner, J.S.; Wilmore, J.H.; Rankinen, T.; Bouchard, C. Genome-Wide Linkage Scan for the Metabolic Syndrome in the HERITAGE Family Study. J. Clin. Endocrinol. Metab. 2003, 88, 5935–5943. [Google Scholar] [CrossRef] [PubMed]
- Ceperuelo-Mallafré, V.; Miranda, M.; Chacón, M.R.; Vilarrasa, N.; Megia, A.; Gutiérrez, C.; Fernández-Real, J.M.; Gómez, J.M.; Caubet, E.; Frühbeck, G.; et al. Adipose Tissue Expression of the Glycerol Channel Aquaporin-7 Gene Is Altered in Severe Obesity but Not in Type 2 Diabetes. J. Clin. Endocrinol. Metab. 2007, 92, 3640–3645. [Google Scholar] [CrossRef]
- Prudente, S.; Flex, E.; Morini, E.; Turchi, F.; Capponi, D.; De Cosmo, S.; Tassi, V.; Guida, V.; Avogaro, A.; Folli, F.; et al. A Functional Variant of the Adipocyte Glycerol Channel Aquaporin 7 Gene Is Associated with Obesity and Related Metabolic Abnormalities. Diabetes 2007, 56, 1468–1474. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Chen, G.; Tu, Q.; Wu, J.; Qin, Y.; Zhu, Z.; Shen, Y.; Yan, L.; Han, A.; Xiang, Q.; et al. Associations between Aquaglyceroporin Gene Polymorphisms and Risk of Type 2 Diabetes Mellitus. Biomed. Res. Int. 2018, 2018, 8167538. [Google Scholar] [CrossRef] [PubMed]
- Nannipieri, M.; Baldi, S.; Mari, A.; Colligiani, D.; Guarino, D.; Camastra, S.; Barsotti, E.; Berta, R.; Moriconi, D.; Bellini, R.; et al. Roux-En-Y Gastric Bypass and Sleeve Gastrectomy: Mechanisms of Diabetes Remission and Role of Gut Hormones. J. Clin. Endocrinol. Metab. 2013, 98, 4391–4399. [Google Scholar] [CrossRef]
- Vigneshwaran, B.; Wahal, A.; Aggarwal, S.; Priyadarshini, P.; Bhattacharjee, H.; Khadgawat, R.; Yadav, R. Impact of Sleeve Gastrectomy on Type 2 Diabetes Mellitus, Gastric Emptying Time, Glucagon-Like Peptide 1 (GLP-1), Ghrelin and Leptin in Non-Morbidly Obese Subjects with BMI 30-35.0 Kg/M2: A Prospective Study. Obes. Surg. 2016, 26, 2817–2823. [Google Scholar] [CrossRef]
- Maeda, N.; Funahashi, T.; Hibuse, T.; Nagasawa, A.; Kishida, K.; Kuriyama, H.; Nakamura, T.; Kihara, S.; Shimomura, I.; Matsuzawa, Y. Adaptation to Fasting by Glycerol Transport through Aquaporin 7 in Adipose Tissue. Proc. Natl. Acad. Sci. USA 2004, 101, 17801–17806. [Google Scholar] [CrossRef]
- He, X.; Gao, F.; Hou, J.; Li, T.; Tan, J.; Wang, C.; Liu, X.; Wang, M.; Liu, H.; Chen, Y.; et al. Metformin Inhibits MAPK Signaling and Rescues Pancreatic Aquaporin 7 Expression to Induce Insulin Secretion in Type 2 Diabetes Mellitus. J. Biol. Chem. 2021, 297, 101002. [Google Scholar] [CrossRef]
- Kerr, A.G.; Sinha, I.; Dadvar, S.; Arner, P.; Dahlman, I. Epigenetic Regulation of Diabetogenic Adipose Morphology. Mol. Metab. 2019, 25, 159–167. [Google Scholar] [CrossRef]
- Agha, G.; Houseman, E.A.; Kelsey, K.T.; Eaton, C.B.; Buka, S.L.; Loucks, E.B. Adiposity Is Associated with DNA Methylation Profile in Adipose Tissue. Int. J. Epidemiol. 2015, 44, 1277–1287. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Role of AQP5 in saliva production in acinar cells. Upon neuronal excitation, acetylcholine, adrenaline, noradrenaline and vasoactive intestinal peptide bind to their receptor and induce either the activation of phospholipase C or adenylyl cyclase resulting in an increase in the intracellular calcium or cyclic adenosine monophosphate (cAMP) concentration. This leads to the trafficking of intracellular vesicles containing AQP5 to the apical plasma membrane and the secretion of sodium and chloride. The latter creates an osmotic gradient allowing the passage of water to the acini lumen. AQP5 is also suggested to play the role of osmosensor involved in the control of paracellular flow as well. A: adrenalin; Ac: acetylcholine; AC: adenylyl cyclase; Ca++: calcium ions; cAMP: cyclic adenosine monophosphate; Cl−: chloride ions; NA: noradrenaline; Na+: sodium ions; PLC: phospholipase C; VIP: vasoactive intestinal peptide. Figure was adapted from [6], reproduced with permission from SNCSC, the Licensed Material is not part of the governing OA license but has been reproduced with permission).
Figure 1.
Role of AQP5 in saliva production in acinar cells. Upon neuronal excitation, acetylcholine, adrenaline, noradrenaline and vasoactive intestinal peptide bind to their receptor and induce either the activation of phospholipase C or adenylyl cyclase resulting in an increase in the intracellular calcium or cyclic adenosine monophosphate (cAMP) concentration. This leads to the trafficking of intracellular vesicles containing AQP5 to the apical plasma membrane and the secretion of sodium and chloride. The latter creates an osmotic gradient allowing the passage of water to the acini lumen. AQP5 is also suggested to play the role of osmosensor involved in the control of paracellular flow as well. A: adrenalin; Ac: acetylcholine; AC: adenylyl cyclase; Ca++: calcium ions; cAMP: cyclic adenosine monophosphate; Cl−: chloride ions; NA: noradrenaline; Na+: sodium ions; PLC: phospholipase C; VIP: vasoactive intestinal peptide. Figure was adapted from [6], reproduced with permission from SNCSC, the Licensed Material is not part of the governing OA license but has been reproduced with permission).

Figure 2.
Role of AQP8 and AQP9 in canalicular bile formation and secretion in hepatocytes. AQP8 mediates the osmotically-driven transport of water into the bile canaliculus across the canalicular membrane, whereas AQP9 ensures the diffusion of water from the sinusoidal blood into the hepatocytes. An AQP-independent movement of water also occurs through the paracellular route passing across the tight junctions to enter the bile canaliculus (blue arrows). Glucagon triggers the microtubule-dependent canalicular targeting of AQP8-containing subapical vesicles. AQP8, present in the mitochondria and the smooth endoplasmic reticulum, may ensure additional roles beyond facilitating the canalicular secretion of bile water. AQP9 ensures the main pathway for the import of lipolytic glycerol from sinusoidal blood (see Table 1). cAMP: cyclic adenosine monophosphate; BC: bile canaliculus; GR: glucagon receptor; PKA: protein kinase A; SAV: subapical vesicles; ST: salt transporters. Figure was adapted from [6], reproduced with permission from SNCSC, the Licensed Material is not part of the governing OA license but has been reproduced with permission).
Figure 2.
Role of AQP8 and AQP9 in canalicular bile formation and secretion in hepatocytes. AQP8 mediates the osmotically-driven transport of water into the bile canaliculus across the canalicular membrane, whereas AQP9 ensures the diffusion of water from the sinusoidal blood into the hepatocytes. An AQP-independent movement of water also occurs through the paracellular route passing across the tight junctions to enter the bile canaliculus (blue arrows). Glucagon triggers the microtubule-dependent canalicular targeting of AQP8-containing subapical vesicles. AQP8, present in the mitochondria and the smooth endoplasmic reticulum, may ensure additional roles beyond facilitating the canalicular secretion of bile water. AQP9 ensures the main pathway for the import of lipolytic glycerol from sinusoidal blood (see Table 1). cAMP: cyclic adenosine monophosphate; BC: bile canaliculus; GR: glucagon receptor; PKA: protein kinase A; SAV: subapical vesicles; ST: salt transporters. Figure was adapted from [6], reproduced with permission from SNCSC, the Licensed Material is not part of the governing OA license but has been reproduced with permission).

Figure 3.
Role of AQP1 and AQP4 in bile secretion in intrahepatic duct cholangiocytes. Secretin, through cAMP, induces the microtubule-dependent apical targeting and translocation of subapical vesicles containing AQP1, CFTR (Cl− channels) and AE2 (Cl−/HCO3− exchangers) to the apical plasma membrane. The exit of Cl− via CFTR drives the extrusion of HCO3− into the ductal lumen through AE2. The presence of HCO3− and Cl− ions generate an osmotic gradient allowing the movement of water from blood plasma (via AQP4 located at the basolateral membrane) to the biliary lumen (via AQP1 located at the apical membrane). AE2: anion exchanges isoform 2; CFTR: cystic fibrosis transmembrane conductance regulator; SAV: subapical vesicles; SR: secretin receptor. Figure was adapted from [6], reproduced with permission from SNCSC, the Licensed Material is not part of the governing OA license but has been reproduced with permission).
Figure 3.
Role of AQP1 and AQP4 in bile secretion in intrahepatic duct cholangiocytes. Secretin, through cAMP, induces the microtubule-dependent apical targeting and translocation of subapical vesicles containing AQP1, CFTR (Cl− channels) and AE2 (Cl−/HCO3− exchangers) to the apical plasma membrane. The exit of Cl− via CFTR drives the extrusion of HCO3− into the ductal lumen through AE2. The presence of HCO3− and Cl− ions generate an osmotic gradient allowing the movement of water from blood plasma (via AQP4 located at the basolateral membrane) to the biliary lumen (via AQP1 located at the apical membrane). AE2: anion exchanges isoform 2; CFTR: cystic fibrosis transmembrane conductance regulator; SAV: subapical vesicles; SR: secretin receptor. Figure was adapted from [6], reproduced with permission from SNCSC, the Licensed Material is not part of the governing OA license but has been reproduced with permission).

Figure 4.
Role of AQP1 and AQP8 in gallbladder bile absorption/secretion. Apical AQP8 and AQP1 promote the osmotic absorption and secretion of water into/from the gallbladder lumen, respectively. Basolateral AQP1 allows the entry/extrusion of water into/out of the epithelial cells. AQP-independent movement of water likely occurs through the tight junctions (paracellular route; blue arrow). The nuclear receptor LXRβ has been suggested to regulate bile volume and the expression of AQP1 and AQP8 together with that of CFTR in the gallbladder of male mice [218]. Thus, the flow of water through the gallbladder epithelium would occur by means of multiple mechanisms involving AQPs and CFTR. CFTR: cystic fibrosis transmembrane conductance regulator. LXRβ: liver X receptor β. SAV: subapical vesicle.
Figure 4.
Role of AQP1 and AQP8 in gallbladder bile absorption/secretion. Apical AQP8 and AQP1 promote the osmotic absorption and secretion of water into/from the gallbladder lumen, respectively. Basolateral AQP1 allows the entry/extrusion of water into/out of the epithelial cells. AQP-independent movement of water likely occurs through the tight junctions (paracellular route; blue arrow). The nuclear receptor LXRβ has been suggested to regulate bile volume and the expression of AQP1 and AQP8 together with that of CFTR in the gallbladder of male mice [218]. Thus, the flow of water through the gallbladder epithelium would occur by means of multiple mechanisms involving AQPs and CFTR. CFTR: cystic fibrosis transmembrane conductance regulator. LXRβ: liver X receptor β. SAV: subapical vesicle.

Figure 5.
Role of AQP7 in β-cell. (A) In healthy β-cells, glycerol entry via AQP7 leads to successive cell swelling, VRAC activation, membrane depolarization, calcium entry, and insulin secretion. In addition, it is hypothesized that AQP7 may act as a junction protein by participating in β-cell rosette formation and intercellular movement and distribution of glycerol as well as other metabolites such as glycerol-3-phosphate. (B) During type 2 diabetes, increased phosphorylated p38 and JNK decreased AQP7 expression which may lead to decreased insulin secretion. (C) Metformin was capable to inhibit p38 and JNK phosphorylated and promoting AQP7 expression and insulin secretion. ATP: adenosine triphosphate; Ca++: calcium ions; DHAP: dihydroxyacetone phosphate GK: glycerol kinase; Glu: glucose; Gro: glycerol; Gro3P: glycerol-3-phosphate; JNK: c-Jun N-terminal kinase; K+: potassium ions; KATP channels: ATP-sensitive potassium channels; NAD+: nicotinamide adenine dinucleotide; NADH: reduced nicotinamide adenine dinucleotide; P: phosphate; p38: Mitogen-Activated Protein Kinase p38; Pyr: pyruvate VRAC: volume-regulated anion channel.
Figure 5.
Role of AQP7 in β-cell. (A) In healthy β-cells, glycerol entry via AQP7 leads to successive cell swelling, VRAC activation, membrane depolarization, calcium entry, and insulin secretion. In addition, it is hypothesized that AQP7 may act as a junction protein by participating in β-cell rosette formation and intercellular movement and distribution of glycerol as well as other metabolites such as glycerol-3-phosphate. (B) During type 2 diabetes, increased phosphorylated p38 and JNK decreased AQP7 expression which may lead to decreased insulin secretion. (C) Metformin was capable to inhibit p38 and JNK phosphorylated and promoting AQP7 expression and insulin secretion. ATP: adenosine triphosphate; Ca++: calcium ions; DHAP: dihydroxyacetone phosphate GK: glycerol kinase; Glu: glucose; Gro: glycerol; Gro3P: glycerol-3-phosphate; JNK: c-Jun N-terminal kinase; K+: potassium ions; KATP channels: ATP-sensitive potassium channels; NAD+: nicotinamide adenine dinucleotide; NADH: reduced nicotinamide adenine dinucleotide; P: phosphate; p38: Mitogen-Activated Protein Kinase p38; Pyr: pyruvate VRAC: volume-regulated anion channel.



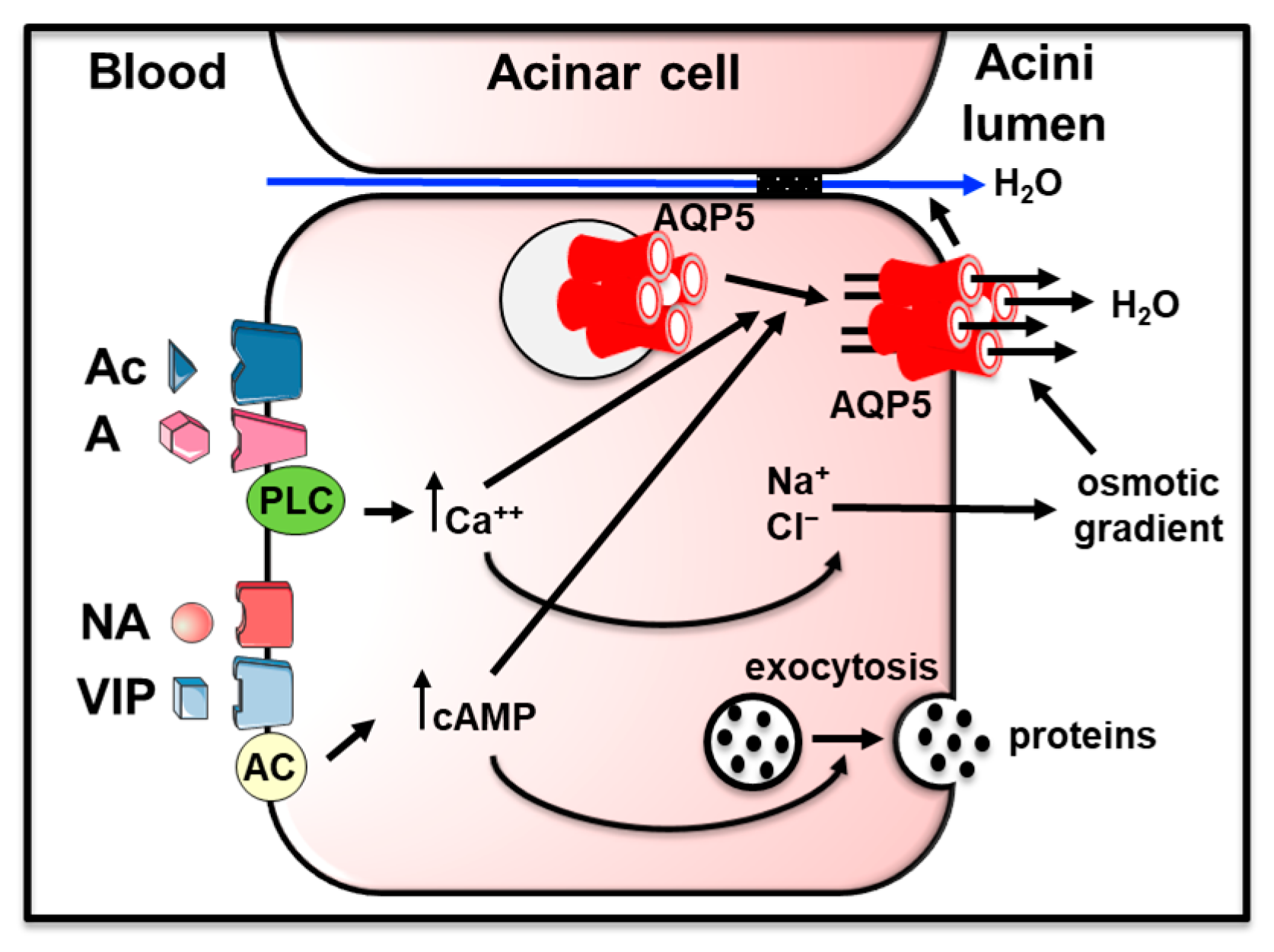{kind=link}
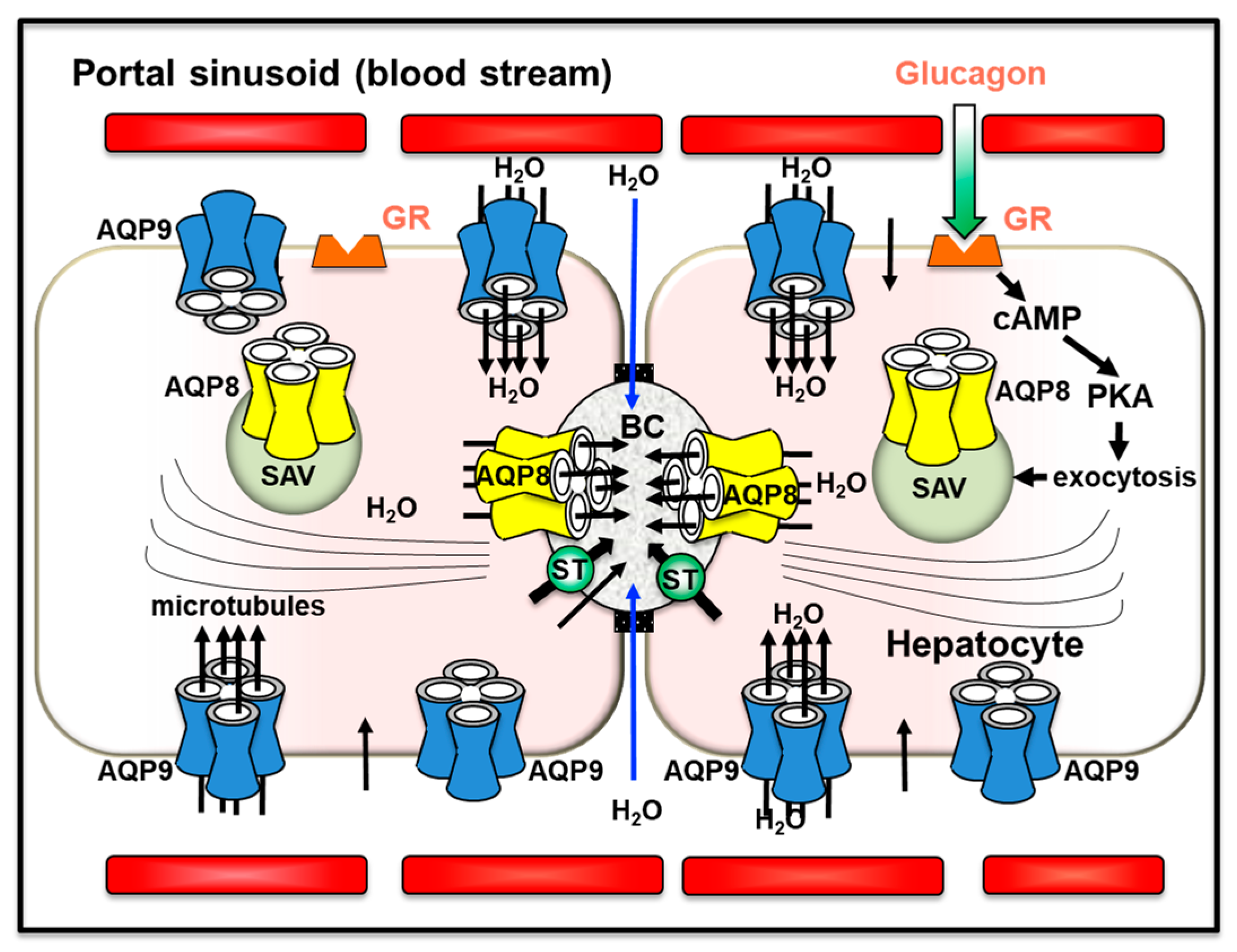{kind=link}
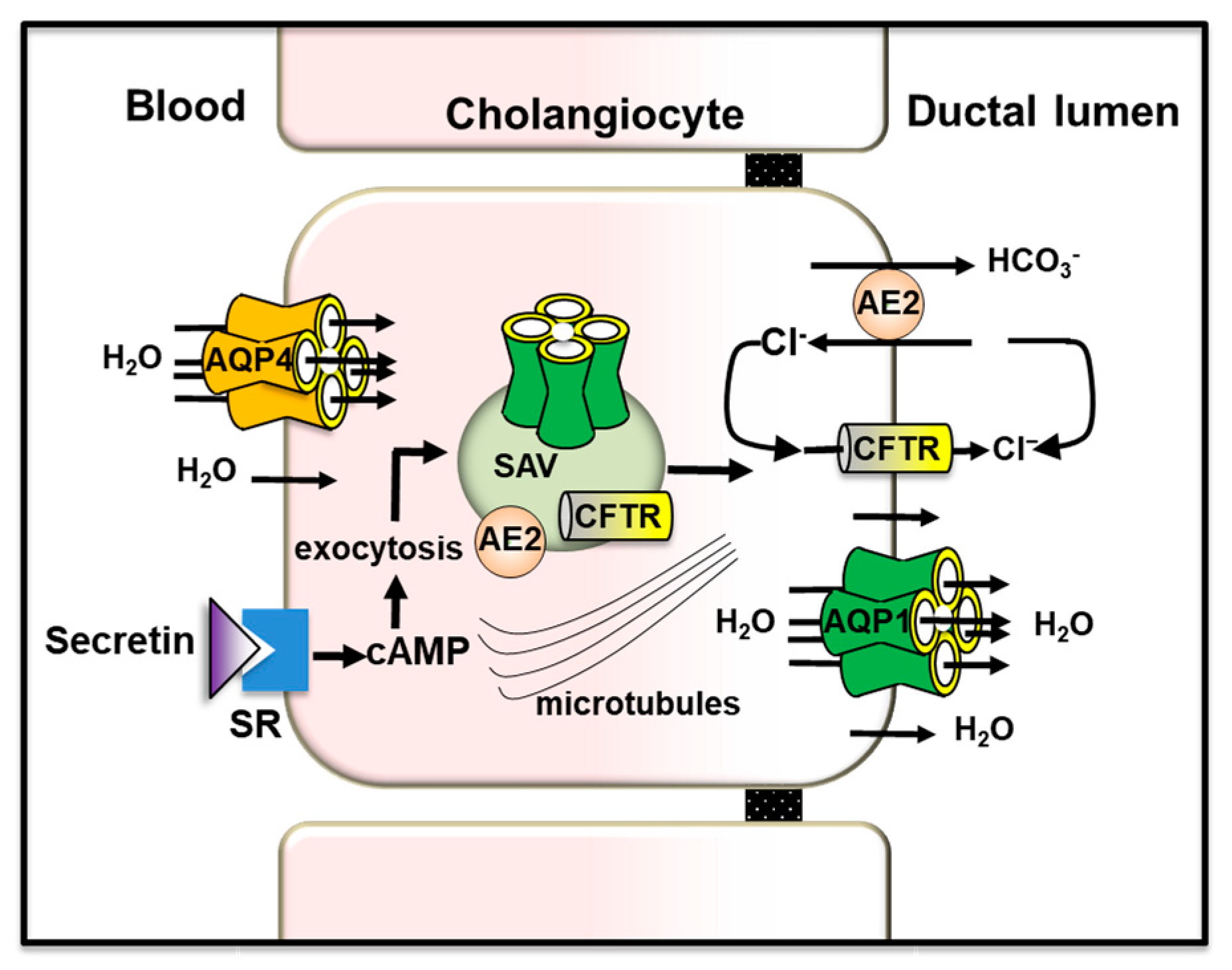{kind=link}
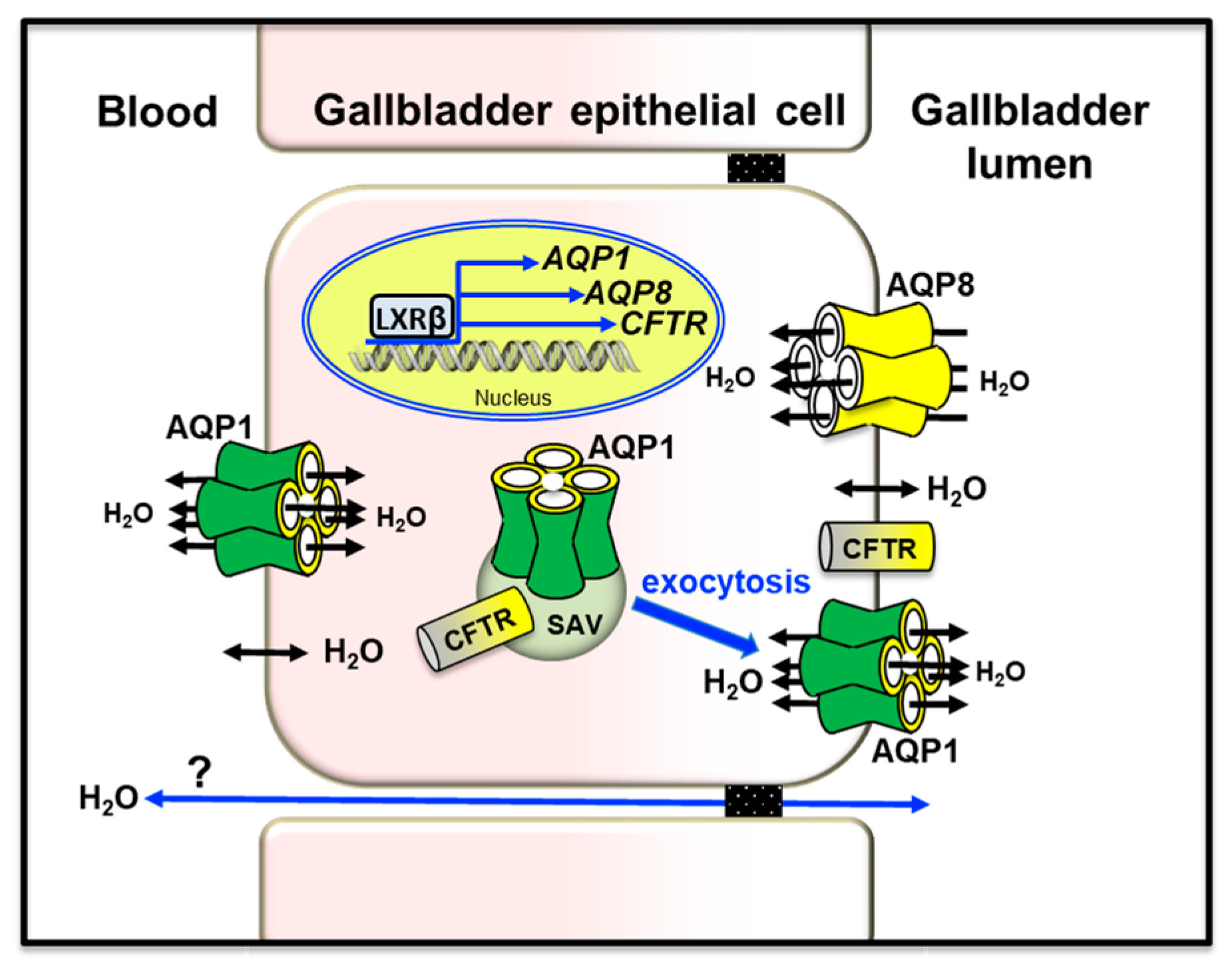{kind=link}
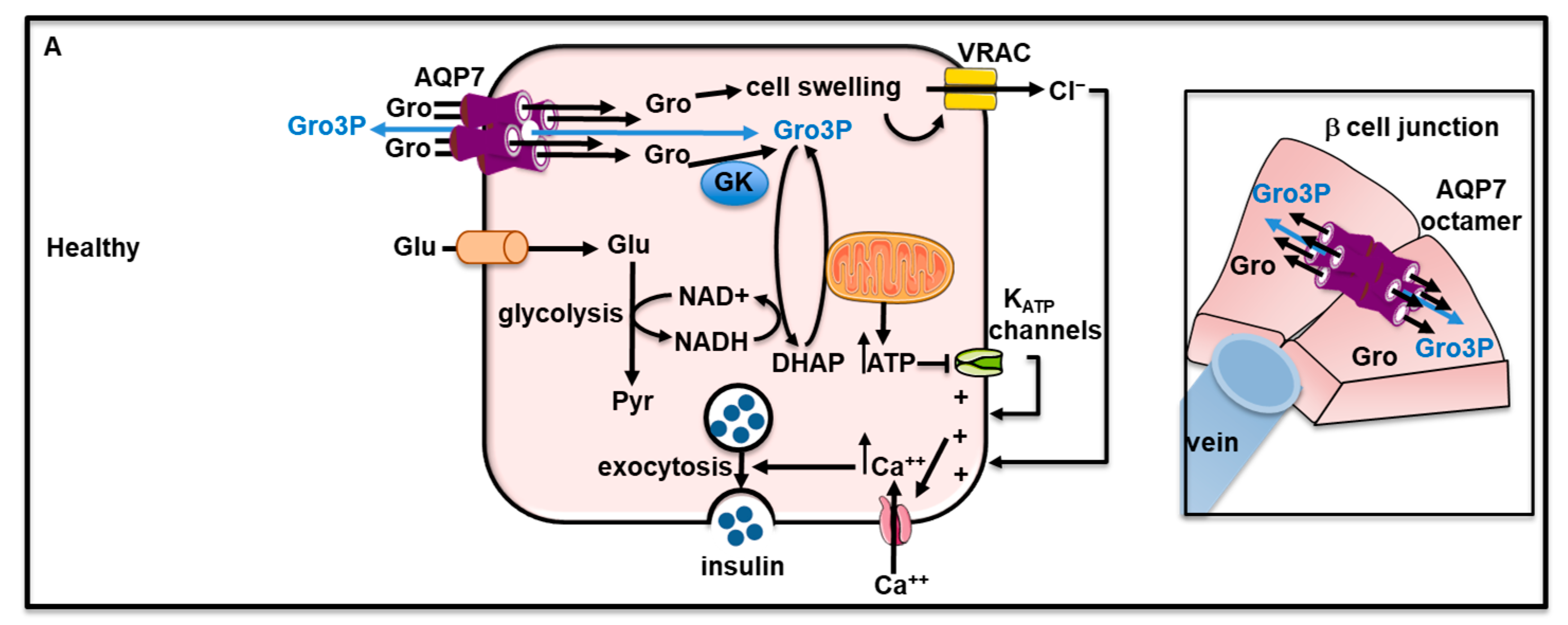{kind=link}
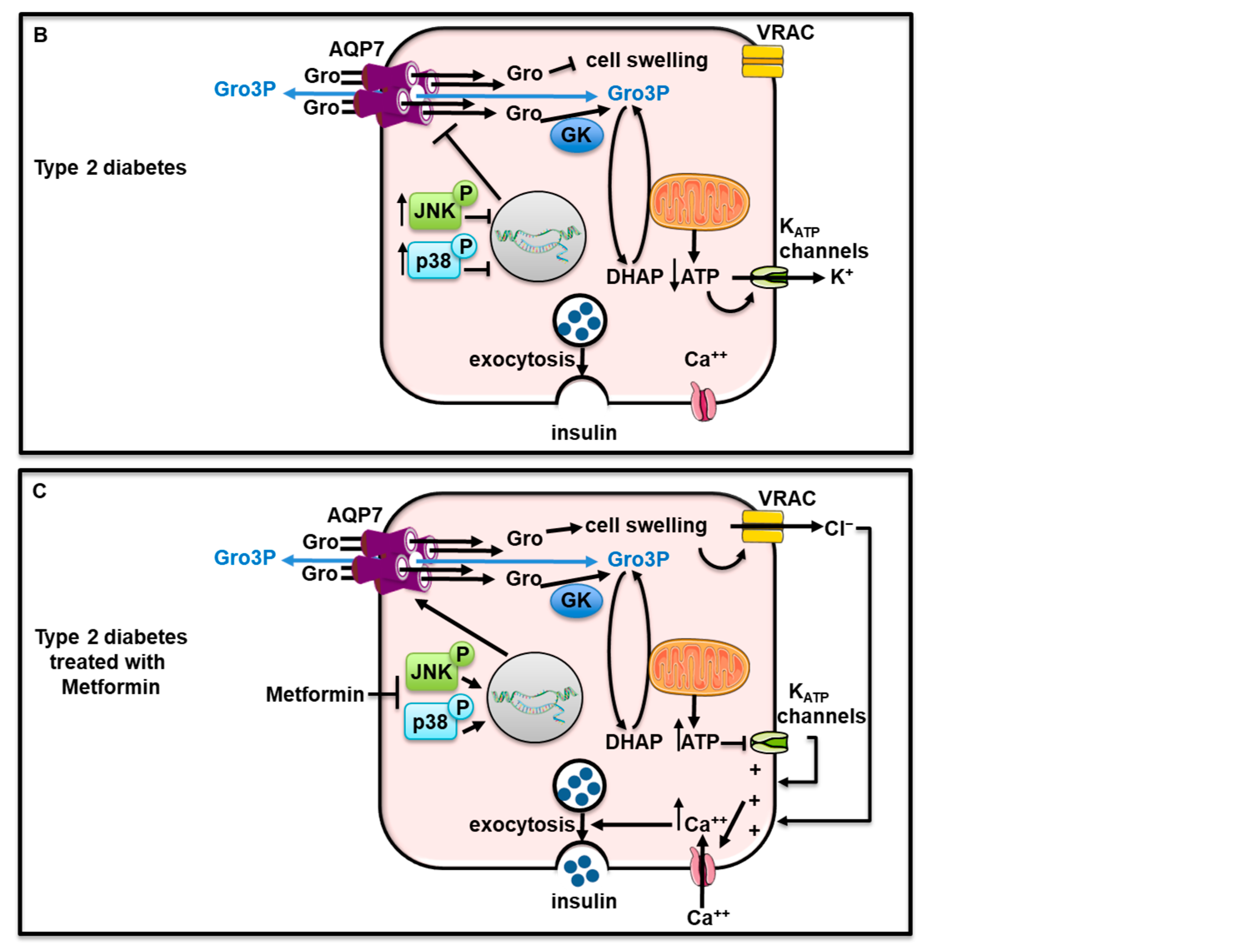{kind=link}
Table 1.
Location and roles of hepatobiliary aquaporins of functional relevance.
| Hepatobiliary Section | AQP | Cellular Location and Species | Subcellular Location | Suggested Functional Involvement | References |
|---|---|---|---|---|---|
| Liver parenchyma | AQP3 | Hep (h) | N.D. | Unclear | |
| AQP7 | Hep (h) | N.D. | Unclear | ||
| AQP8 | Hep (r, m, h) | APM, SAV, IMM, SER | Canalicular bile secretion; cytoplasmic osmotic homeostasis; mitochondrial ammonia detoxification and ureagenesis; mitochondrial H2O2 release; cholesterol biosynthesis; regulation of metabolic signaling | [156,157,158,159] | |
| AQP9 | Hep (r, m, h) | BLM | Glycerol uptake during starvation; lipid homeostasis; water movement from sinusoidal blood; catabolic urea secretion; | [156,160,161] | |
| AQP11 | Hep (m) | RER | RER homeostasis; liver regeneration | ||
| Intrahepatic bile ducts | AQP1 | Chol (m, r, h) | APM, SAV, BLM | Secretion and absorption of ductal bile water | [41] |
| AQP4 | Chol (m, r) | BLM | Secretion and absorption of ductal bile water | [162] | |
| Gallbladder | AQP1 | EpC (m, h) | APM, BLM, SAV | Cystic bile absorption/secretion | [41,163,164,165] |
| AQP8 | EpC (m, h) | APM, SAV | Cystic bile absorption (?) | [163,164] | |
| PS; PVP; BV | AQP1 | EC (h) | APM, BLM | Bile formation and flow | [41] |
| Other hepatic cell types | AQP3 | KC (h) | PM | Cell migration; proinflammatory cytokines secretion (?) | [156] |
| AQP8 | KC (r) | PM | Repopulation of Kupffer cells during liver regeneration (?) | [166] | |
| AQP3 | SC (h) | PM | Adiponectin-mediated inhibition of SC cells activation | [167] | |
| AQP11 | SC (r) | N.D. | Control of activated SC proliferation | [168] |
AM, apical plasma membrane; BLM, basolateral plasma membrane; BV, blood vessels; EC: endothelial cells; EpC: epithelial cells; h: human; Hep: hepatocytes; IMM, inner mitochondrial membrane; KC: Kupffer cells; m: mouse; ND: not determined; PM, plasma membrane; PS: portal sinusoids; PVP, peribiliary vascular plexus; r; rat; RER, rough endoplasmic reticulum; SAV, subapical membrane vesicles; SC: stellate cells; SER, smooth endoplasmic reticulum.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Calamita, G.; Delporte, C. Insights into the Function of Aquaporins in Gastrointestinal Fluid Absorption and Secretion in Health and Disease. Cells 2023, 12, 2170. https://doi.org/10.3390/cells12172170
AMA Style
Calamita G, Delporte C. Insights into the Function of Aquaporins in Gastrointestinal Fluid Absorption and Secretion in Health and Disease. Cells. 2023; 12(17):2170. https://doi.org/10.3390/cells12172170
Chicago/Turabian StyleCalamita, Giuseppe, and Christine Delporte. 2023. "Insights into the Function of Aquaporins in Gastrointestinal Fluid Absorption and Secretion in Health and Disease" Cells 12, no. 17: 2170. https://doi.org/10.3390/cells12172170
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.