
Real-Time Visualization of Cytosolic and Mitochondrial ATP Dynamics in Response to Metabolic Stress in Cultured Cells

 ,
,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Design and Validation of an In Vitro Dual-ATP Indicator (smacATPi)
2.2. Generation and Validation of neg-smacATPi
2.3. Cell Culture
2.4. HEK293T Cell Transfection
2.5. Mouse Embryonic Fibroblast (MEF) Isolation
2.6. Electroporation of MEF Cells
2.7. Isolation and Culture of Adult Rat Cardiomyocytes (ARCM)
2.8. Generation of Adenovirus for smacATPi
2.9. Drug Selection
2.10. Evaluation of Mitochondrial Membrane Potential
2.11. Evaluation of Mitochondrial pH Change
2.12. Image Acquisition
2.13. Hypoxic Conditions for HEK293T Cells
2.14. Statistical Analysis
3. Results
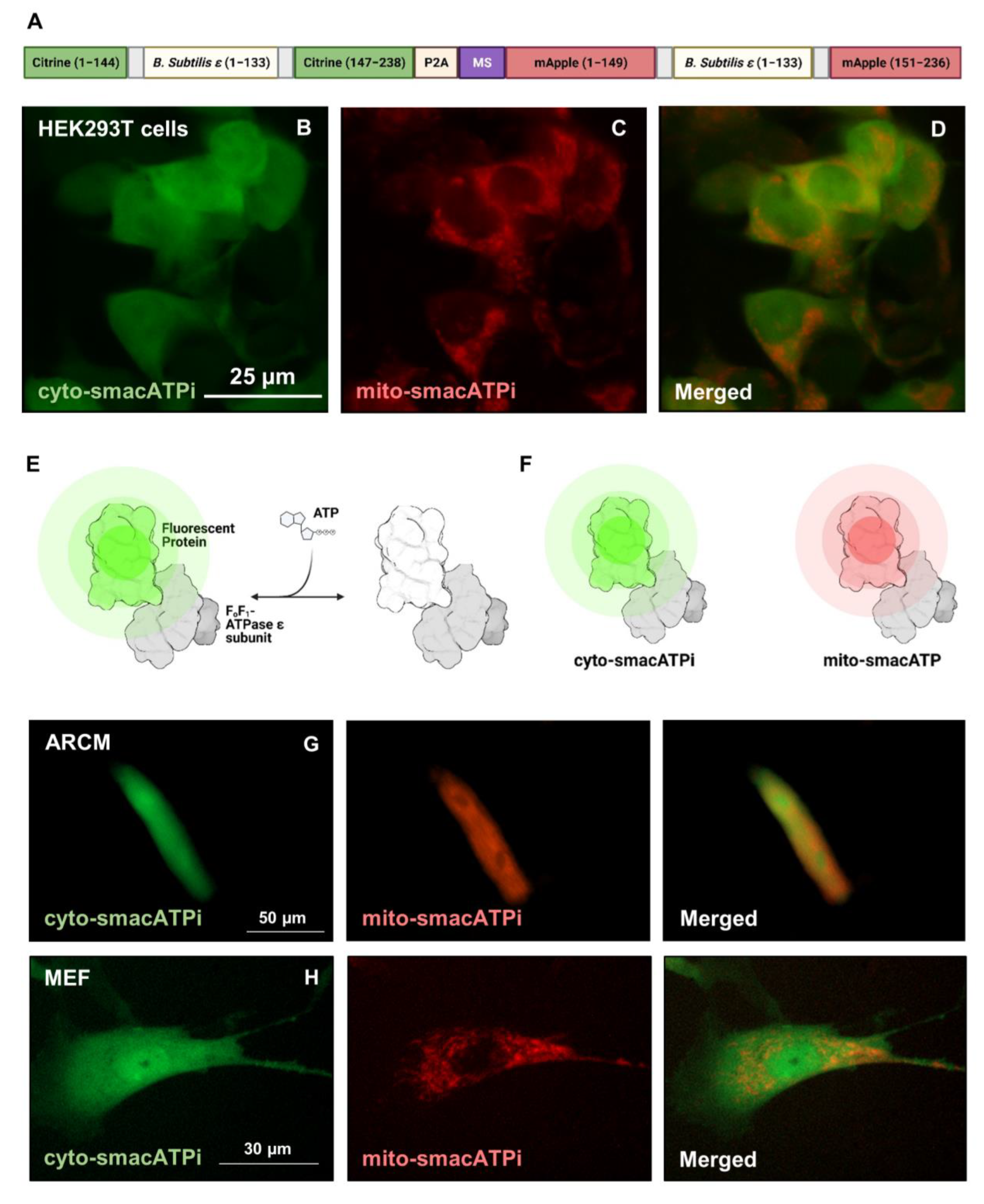
3.1. Design, Generation, and Validation of smacATPi (Simultaneous Mitochondrial and Cytosolic ATP Indicator)
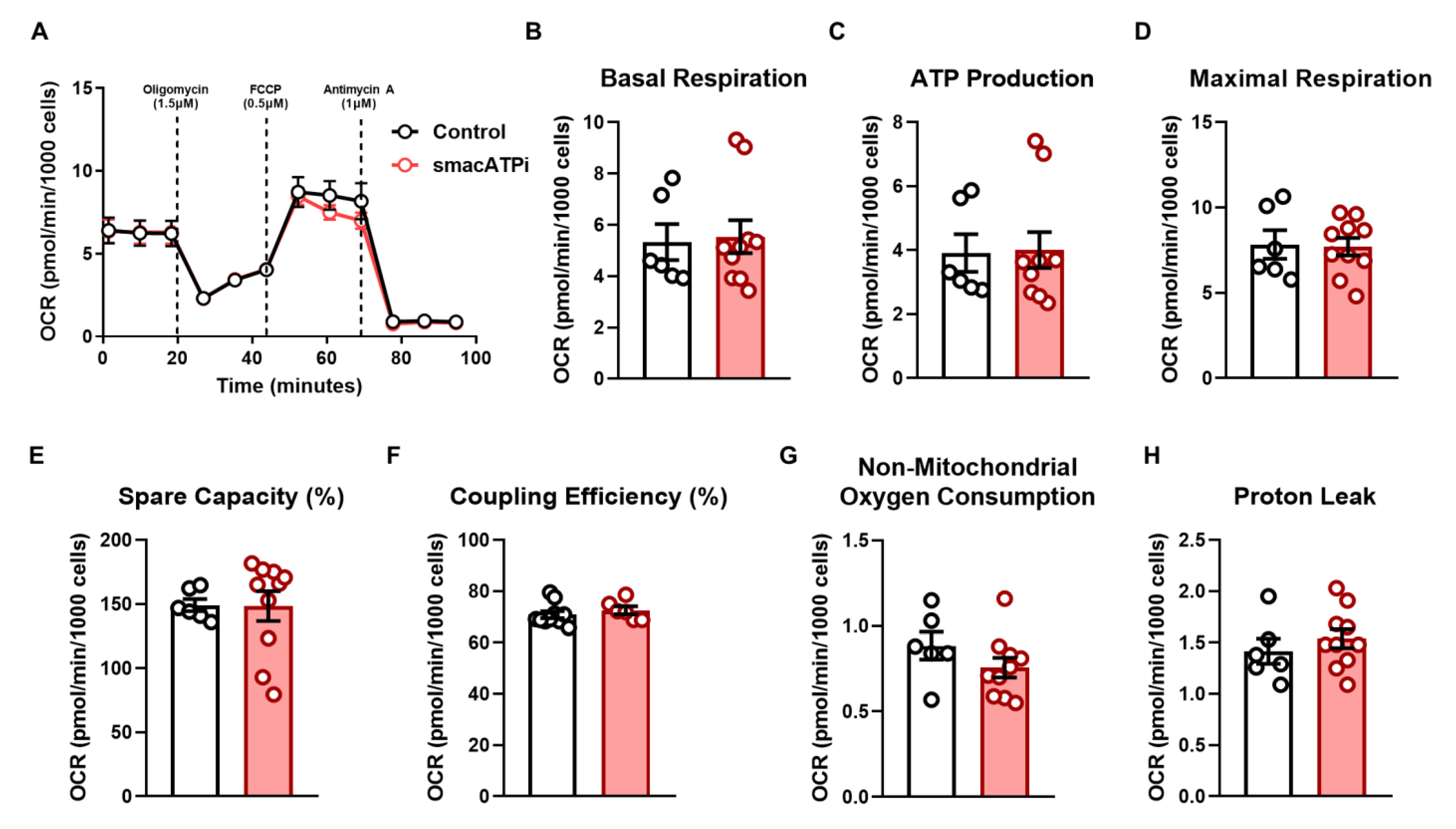
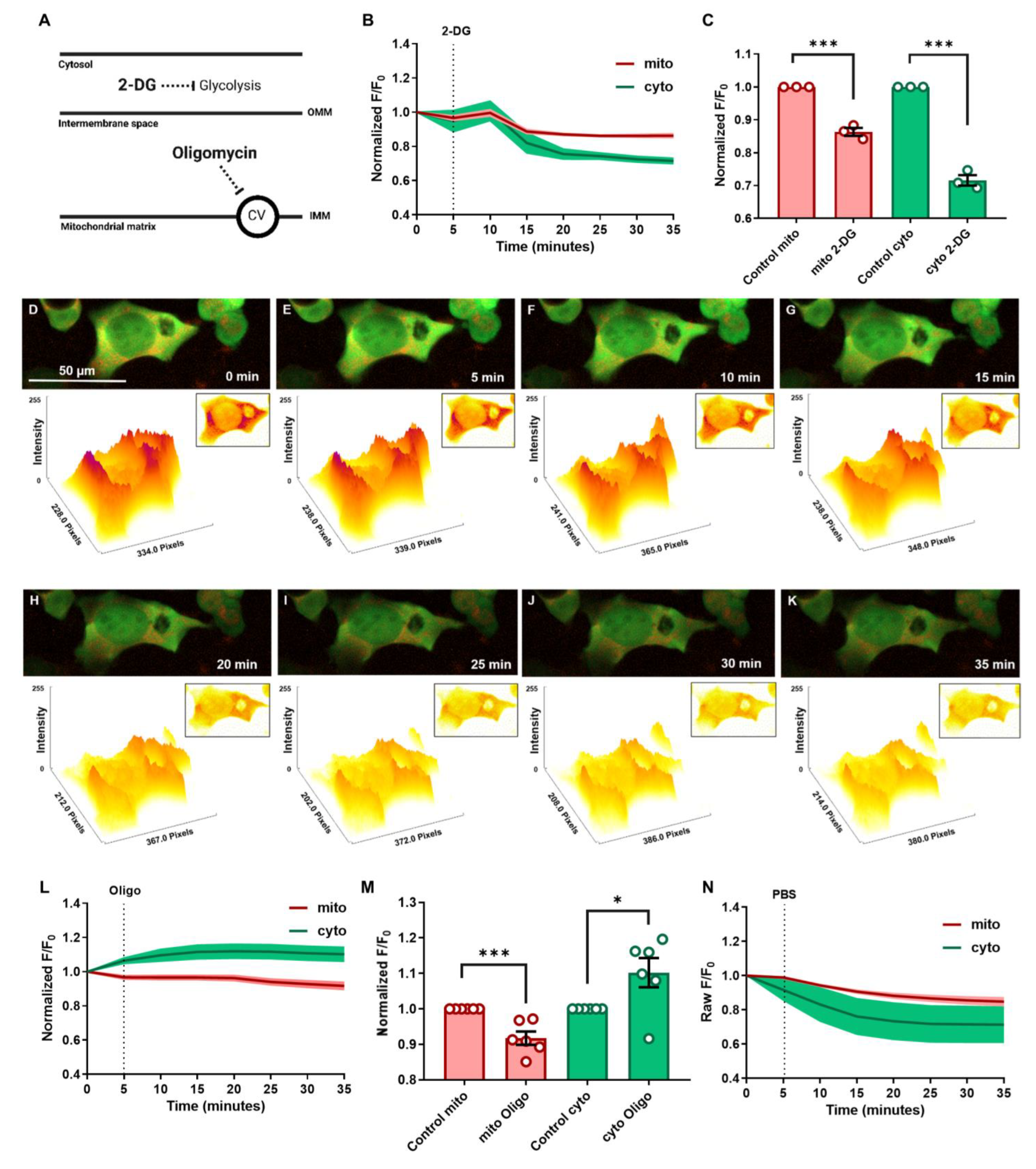
3.2. Cytosolic and Mitochondrial ATP Dynamics and Trafficking in Response to Glycolytic and Mitochondrial Inhibitions under Normoxic Conditions
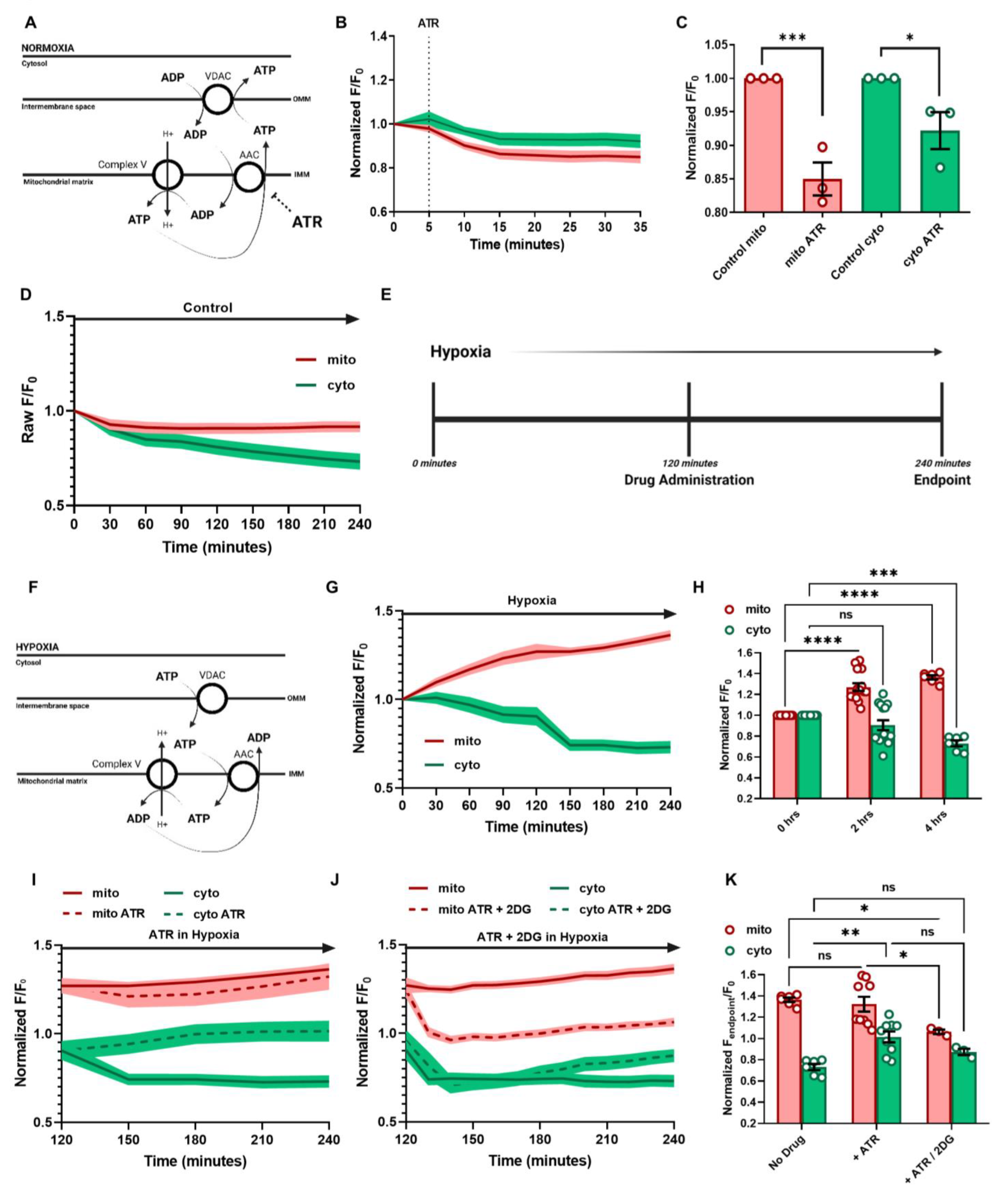
3.3. ATP Regulation and Cellular Response in Hypoxia
4. Discussion
4.1. Feasibility of the Dual ATP Indicator for Real-Time ATP Visualization within the Cytosol and Mitochondria of Cultured Cells
4.2. Real-Time Spatiotemporal Cellular ATP Dynamics in the Presence of Mitochondrial and Metabolic Inhibitors
4.3. ATP Regulation under Hypoxic Conditions
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| 2-DG | 2-Deoxy-D-glucose |
| AA | Antimycin A |
| ADP | Adenosine diphosphate |
| ATP | Adenosine triphosphate |
| ATR | Atractyloside |
| FBS | Fetal bovine serum |
| IMM | Inner mitochondrial membrane |
| OCR | Oxygen consumption rate |
| OMM | Outer mitochondrial membrane |
| OXPHOS | Oxidative phosphorylation |
| PBS | Phosphate buffered saline |
| smacATPi | simultaneous mitochondrial and cytosolic ATP indicator |
| VDAC1 | voltage-dependent anion channel 1 |
References
- Johnson, T.A.; Jinnah, H.A.; Kamatani, N. Shortage of Cellular ATP as a Cause of Diseases and Strategies to Enhance ATP. Front. Pharmacol. 2019, 10, 98. [Google Scholar] [CrossRef] [Green Version]
- Zhang, F.; Huang, P.-J.J.; Liu, J. Sensing Adenosine and ATP by Aptamers and Gold Nanoparticles: Opposite Trends of Color Change from Domination of Target Adsorption Instead of Aptamer Binding. ACS Sens. 2020, 5, 2885–2893. [Google Scholar] [CrossRef]
- Imamura, H.; Nhat, K.P.H.; Togawa, H.; Saito, K.; Iino, R.; Kato-Yamada, Y.; Nagai, T.; Noji, H. Visualization of ATP levels inside single living cells with fluorescence resonance energy transfer-based genetically encoded indicators. Proc. Natl. Acad. Sci. USA 2009, 106, 15651–15656. [Google Scholar] [CrossRef] [Green Version]
- Depaoli, M.R.; Karsten, F.; Madreiter-Sokolowski, C.T.; Klec, C.; Gottschalk, B.; Bischof, H.; Eroglu, E.; Waldeck-Weiermair, M.; Simmen, T.; Graier, W.F.; et al. Faculty Opinions recommendation of Real-Time Imaging of Mitochondrial ATP Dynamics Reveals the Metabolic Setting of Single Cells. Cell Rep. 2019, 25, 501–512.e3. [Google Scholar] [CrossRef] [Green Version]
- Conley, J.M.; Radhakrishnan, S.; Valentino, S.A.; Tantama, M. Imaging extracellular ATP with a genetically-encoded, ratiometric fluorescent sensor. PLoS ONE 2017, 12, e0187481. [Google Scholar] [CrossRef] [Green Version]
- Rajendran, M.; Dane, E.; Conley, J.; Tantama, M. Imaging Adenosine Triphosphate (ATP). Biol. Bull. 2016, 231, 73–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, D.; Yang, Q. Genetically Encoded ATP Biosensors for Direct Monitoring of Cellular ATP Dynamics. Cells 2022, 11, 1920. [Google Scholar] [CrossRef] [PubMed]
- Arai, S.; Kriszt, R.; Harada, K.; Looi, L.-S.; Matsuda, S.; Wongso, D.; Suo, S.; Ishiura, S.; Tseng, Y.-H.; Raghunath, M.; et al. RGB-Color Intensiometric Indicators to Visualize Spatiotemporal Dynamics of ATP in Single Cells. Angew. Chem. Int. Ed. 2018, 57, 10873–10878. [Google Scholar] [CrossRef]
- Klingenberg, M. The ADP and ATP transport in mitochondria and its carrier. Biochim. Biophys. Acta (BBA) Biomembr. 2008, 1778, 1978–2021. [Google Scholar] [CrossRef] [Green Version]
- Aprille, J.R. Mechanism and regulation of the mitochondrial ATP-Mg/Pi carrier. J. Bioenerg. Biomembr. 1993, 25, 473–481. [Google Scholar] [CrossRef]
- Hara, K.Y.; Kondo, A. ATP regulation in bioproduction. Microb. Cell Factories 2015, 14, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chinopoulos, C.; Gerencser, A.A.; Mandi, M.; Mathe, K.; Töröcsik, B.; Doczi, J.; Turiak, L.; Kiss, G.; Konrad, C.; Vajda, S.; et al. Forward operation of adenine nucleotide translocase during F0F1-ATPase reversal: Critical role of matrix substrate-level phosphorylation. FASEB J. 2010, 24, 2405–2416. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Han, X.; Lin, Y. Dissecting the regulation and function of ATP at the single-cell level. PLoS Biol. 2018, 16, e3000095. [Google Scholar] [CrossRef]
- Area-Gomez, E.; Guardia-Laguarta, C.; Schon, E.A.; Przedborski, S. Mitochondria, OxPhos, and neurodegeneration: Cells are not just running out of gas. J. Clin. Investig. 2019, 129, 34–45. [Google Scholar] [CrossRef] [Green Version]
- Fernandez-Vizarra, E.; Zeviani, M. Mitochondrial disorders of the OXPHOS system. FEBS Lett. 2020, 595, 1062–1106. [Google Scholar] [CrossRef] [PubMed]
- García-Aguilar, A.; Cuezva, J.M. A Review of the Inhibition of the Mitochondrial ATP Synthase by IF1 in vivo: Reprogramming Energy Metabolism and Inducing Mitohormesis. Front. Physiol. 2018, 9, 1322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farhat, E.; Cheng, H.; Romestaing, C.; Pamenter, M.; Weber, J.-M. Goldfish Response to Chronic Hypoxia: Mitochondrial Respiration, Fuel Preference and Energy Metabolism. Metabolites 2021, 11, 187. [Google Scholar] [CrossRef]
- St-Pierre, J.; Brand, M.D.; Boutilier, R.G. Mitochondria as ATP consumers: Cellular treason in anoxia. Proc. Natl. Acad. Sci. USA 2000, 97, 8670–8674. [Google Scholar] [CrossRef] [Green Version]
- Belisario, D.C.; Kopecka, J.; Pasino, M.; Akman, M.; De Smaele, E.; Donadelli, M.; Riganti, C. Hypoxia Dictates Metabolic Rewiring of Tumors: Implications for Chemoresistance. Cells 2020, 9, 2598. [Google Scholar] [CrossRef]
- Fontaine, E.; Leverve, X. Basics in clinical nutrition: Metabolic response to hypoxia. e-SPEN Eur. e-J. Clin. Nutr. Metab. 2008, 3, e285–e288. [Google Scholar] [CrossRef] [Green Version]
- Hellwig-Bürgel, T.; Stiehl, D.P.; Wagner, A.E.; Metzen, E.; Jelkmann, W. Review: Hypoxia-Inducible Factor-1 (HIF-1): A Novel Transcription Factor in Immune Reactions. J. Interf. Cytokine Res. 2005, 25, 297–310. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.W.; Ko, J.; Ju, C.; Eltzschig, H.K. Hypoxia signaling in human diseases and therapeutic targets. Exp. Mol. Med. 2019, 51, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solaini, G.; Baracca, A.; Lenaz, G.; Sgarbi, G. Hypoxia and mitochondrial oxidative metabolism. Biochim. Biophys. Acta (BBA) Bioenerg. 2010, 1797, 1171–1177. [Google Scholar] [CrossRef] [Green Version]
- Denko, N.C. Hypoxia, HIF1 and glucose metabolism in the solid tumour. Nat. Rev. Cancer 2008, 8, 705–713. [Google Scholar] [CrossRef] [PubMed]
- Stewart, M.J.; Steenkamp, V. The Biochemistry and Toxicity of Atractyloside: A Review. Ther. Drug Monit. 2000, 22, 641–649. [Google Scholar] [CrossRef]
- Wheaton, W.W.; Chandel, N.S.; Chihanga, T.; Ruby, H.N.; Ma, Q.; Bashir, S.; Devarajan, P.; Kennedy, M.A.; Bernard, O.; Jeny, F.; et al. Hypoxia. 2. Hypoxia regulates cellular metabolism. Am. J. Physiol. Physiol. 2011, 300, C385–C393. [Google Scholar] [CrossRef] [Green Version]
- DeLuca, M.; McElroy, W.D. Kinetics of the firefly luciferase catalyzed reactions. Biochemistry 1974, 13, 921–925. [Google Scholar] [CrossRef]
- Morciano, G.; Sarti, A.C.; Marchi, S.; Missiroli, S.; Falzoni, S.; Raffaghello, L.; Pistoia, V.; Giorgi, C.; Di Virgilio, F.; Pinton, P. Use of luciferase probes to measure ATP in living cells and animals. Nat. Protoc. 2017, 12, 1542–1562. [Google Scholar] [CrossRef]
- Marsin, A.-S.; Bouzin, C.; Bertrand, L.; Hue, L. The Stimulation of Glycolysis by Hypoxia in Activated Monocytes Is Mediated by AMP-activated Protein Kinase and Inducible 6-Phosphofructo-2-kinase. J. Biol. Chem. 2002, 277, 30778–30783. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Gu, Y.; Yu, X. Assessing tissue metabolism by phosphorous-31 magnetic resonance spectroscopy and imaging: A methodology review. Quant. Imaging Med. Surg. 2017, 7, 707–716. [Google Scholar] [CrossRef] [Green Version]
- Chinopoulos, C.; Adam-Vizi, V. Mitochondria as ATP consumers in cellular pathology. Biochim. Biophys. Acta (BBA) Mol. Basis Dis. 2010, 1802, 221–227. [Google Scholar] [CrossRef] [Green Version]
- Leyssens, A.; Nowicky, A.V.; Patterson, L.; Crompton, M.; Duchen, M.R. The relationship between mitochondrial state, ATP hydrolysis, [Mg2+]i and [Ca2+]i studied in isolated rat cardiomyocytes. J. Physiol. 1996, 496, 111–128. [Google Scholar] [CrossRef] [PubMed]
- Obatomi, D.K.; Bach, P.H. Inhibition of mitochondrial respiration and oxygen uptake in isolated rat renal tubular fragments by atractyloside. Toxicol. Lett. 1996, 89, 155–161. [Google Scholar] [CrossRef]
- Ruprecht, J.J.; King, M.S.; Zögg, T.; Aleksandrova, A.A.; Pardon, E.; Crichton, P.G.; Steyaert, J.; Kunji, E.R. The Molecular Mechanism of Transport by the Mitochondrial ADP/ATP Carrier. Cell 2019, 176, 435–447.e15. [Google Scholar] [CrossRef] [Green Version]
- Chinopoulos, C.; Kiss, G.; Kawamata, H.; Starkov, A.A. Measurement of ADP–ATP Exchange in Relation to Mitochondrial Transmembrane Potential and Oxygen Consumption. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 2014; Volume 542, pp. 333–348. [Google Scholar] [CrossRef] [Green Version]
- Kawamata, H.; Starkov, A.A.; Manfredi, G.; Chinopoulos, C. A kinetic assay of mitochondrial ADP–ATP exchange rate in permeabilized cells. Anal. Biochem. 2010, 407, 52–57. [Google Scholar] [CrossRef] [Green Version]
- Santi, R. Pharmacological properties and mechanism of action of atractyloside. J. Pharm. Pharmacol. 1964, 16, 437–438. [Google Scholar] [CrossRef]
- Kunji, E.R.S.; Aleksandrova, A.; King, M.S.; Majd, H.; Ashton, V.L.; Cerson, E.; Springett, R.; Kibalchenko, M.; Tavoulari, S.; Crichton, P.G.; et al. The transport mechanism of the mitochondrial ADP/ATP carrier. Biochim. Biophys. Acta (BBA) Mol. Cell Res. 2016, 1863, 2379–2393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ley-Ngardigal, S.; Bertolin, G. Approaches to monitor ATP levels in living cells: Where do we stand? FEBS J. 2022, 289, 7940–7969. [Google Scholar] [CrossRef]
- Tsuyama, T.; Kishikawa, J.-I.; Han, Y.-W.; Harada, Y.; Tsubouchi, A.; Noji, H.; Kakizuka, A.; Yokoyama, K.; Uemura, T.; Imamura, H. In Vivo Fluorescent Adenosine 5′-Triphosphate (ATP) Imaging of Drosophila melanogaster and Caenorhabditis elegans by Using a Genetically Encoded Fluorescent ATP Biosensor Optimized for Low Temperatures. Anal. Chem. 2013, 85, 7889–7896. [Google Scholar] [CrossRef]
- Maldonado, E.N.; DeHart, D.N.; Patnaik, J.; Klatt, S.C.; Gooz, M.B.; Lemasters, J.J. ATP/ADP Turnover and Import of Glycolytic ATP into Mitochondria in Cancer Cells Is Independent of the Adenine Nucleotide Translocator. J. Biol. Chem. 2016, 291, 19642–19650. [Google Scholar] [CrossRef] [Green Version]
- Manfredi, G. Measurements of ATP in mammalian cells. Methods 2002, 26, 317–326. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Wen, J.; Li, H.; Sun, S.; Xu, Y. Fluorescent probes for recognition of ATP. Chin. Chem. Lett. 2017, 28, 1916–1924. [Google Scholar] [CrossRef]
- Lobas, M.A.; Tao, R.; Nagai, J.; Kronschläger, M.T.; Borden, P.M.; Marvin, J.S.; Looger, L.L.; Khakh, B.S. A genetically encoded single-wavelength sensor for imaging cytosolic and cell surface ATP. Nat. Commun. 2019, 10, 711. [Google Scholar] [CrossRef] [PubMed] [Green Version]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
White, D., III; Lauterboeck, L.; Mobasheran, P.; Kitaguchi, T.; Chaanine, A.H.; Yang, Q. Real-Time Visualization of Cytosolic and Mitochondrial ATP Dynamics in Response to Metabolic Stress in Cultured Cells. Cells 2023, 12, 695. https://doi.org/10.3390/cells12050695
White D III, Lauterboeck L, Mobasheran P, Kitaguchi T, Chaanine AH, Yang Q. Real-Time Visualization of Cytosolic and Mitochondrial ATP Dynamics in Response to Metabolic Stress in Cultured Cells. Cells. 2023; 12(5):695. https://doi.org/10.3390/cells12050695
Chicago/Turabian StyleWhite, Donnell, III, Lothar Lauterboeck, Parnia Mobasheran, Tetsuya Kitaguchi, Antoine H. Chaanine, and Qinglin Yang. 2023. "Real-Time Visualization of Cytosolic and Mitochondrial ATP Dynamics in Response to Metabolic Stress in Cultured Cells" Cells 12, no. 5: 695. https://doi.org/10.3390/cells12050695