
Deciphering Molecular Mechanisms and Intervening in Physiological and Pathophysiological Processes of Ca2+ Signaling Mechanisms Using Optogenetic Tools


Abstract
:1. Introduction
1.1. Ca2+—The Versatile Signaling Protein
1.2. Ca2+ Entry Pathways
1.2.1. Voltage-Gated Ca2+ Ion Channels (VGCCs)
1.2.2. Store-Dependent Ca2+ Ion Channels
1.2.3. Store-Independent Ca2+ Ion Channels
1.2.4. Transient Receptor Potential Ion Channels
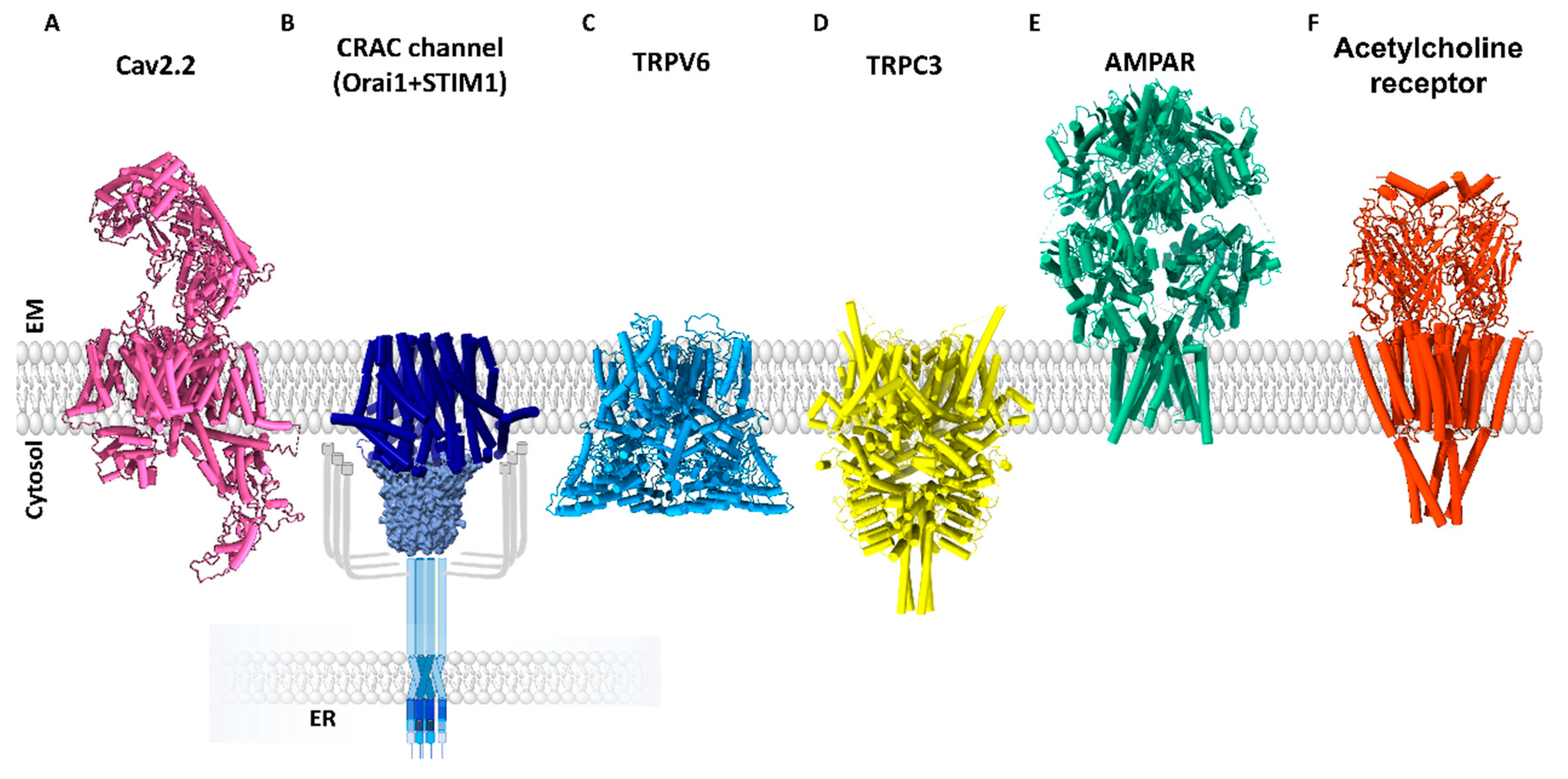
1.2.5. Receptor Ca2+ Ion Channels
2. Optogenetics to Unravel the Mechanism of Ca2+ Ion Channels
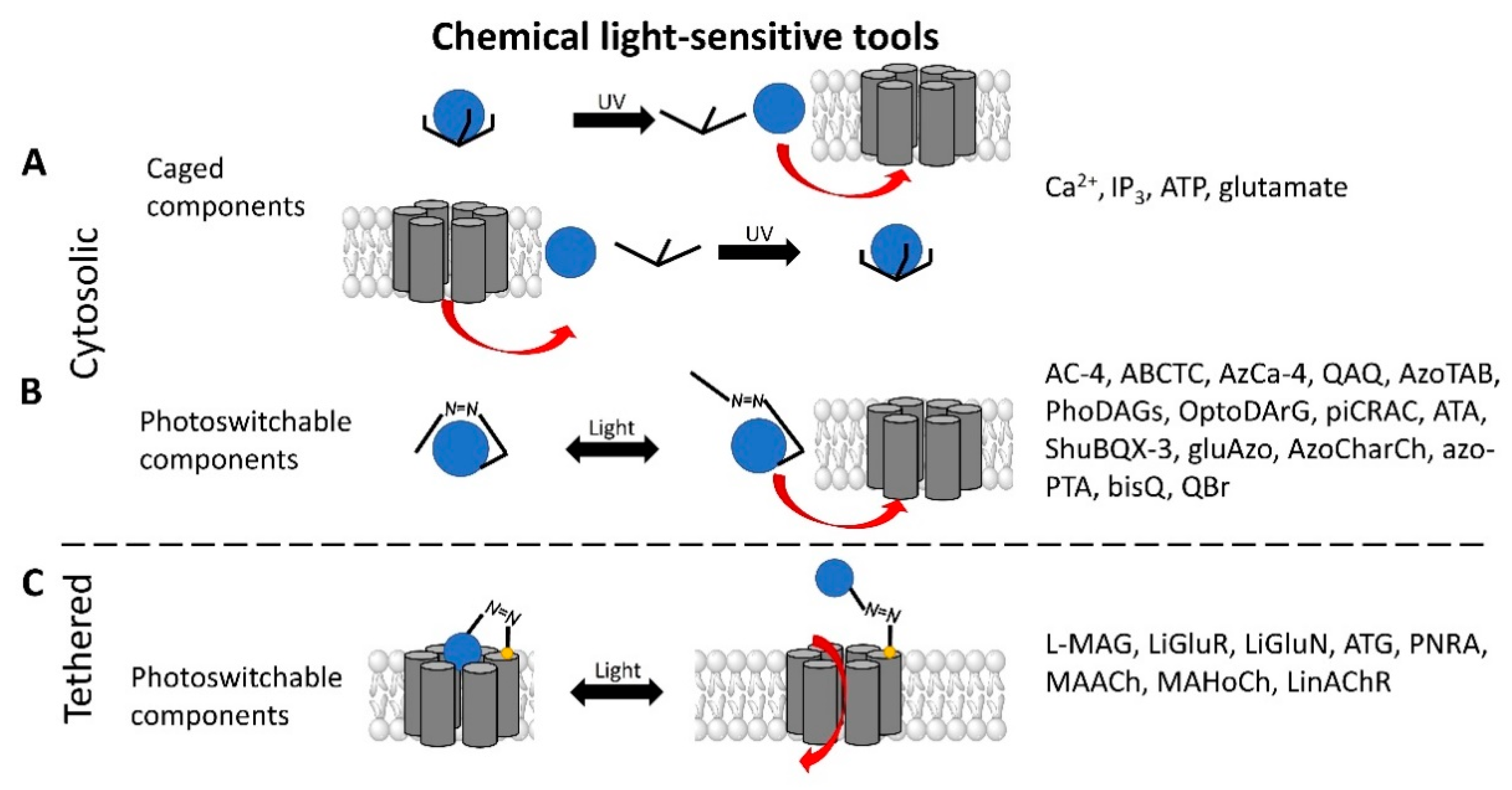
2.1. Light Sensitive Chemical Tools
2.1.1. Light-Sensitive Chemical Tools That Modulate Cytosolic Ca2+ Levels
2.1.2. Light-Sensitive Chemical Tools Interfering with Ca2+ Signaling Cascades
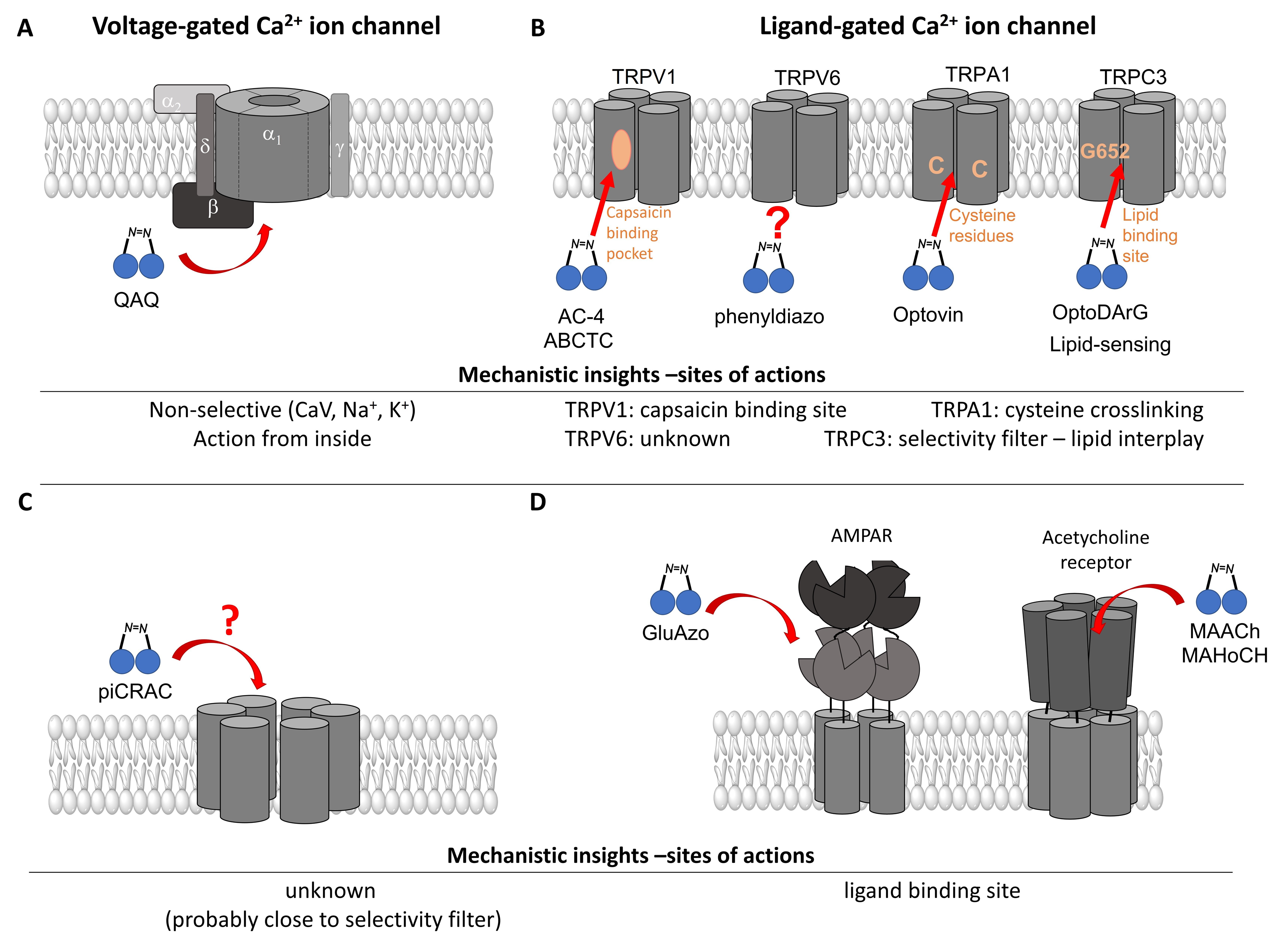
2.1.3. Light Sensitive Chemical Tools That Interfere with Ca2+ Permeable Ion Channels
- Photochromic Ligands Acting on Voltage-Gated Ca2+ Ion Channels
- Photosensitive Ligands Acting on TRP and CRAC Ion Channels
- Photochromic Ligands Acting on Ca2+ Permeable Receptor Ion Channels
2.1.4. Advantages and Disadvantages of Photochromic Ligands
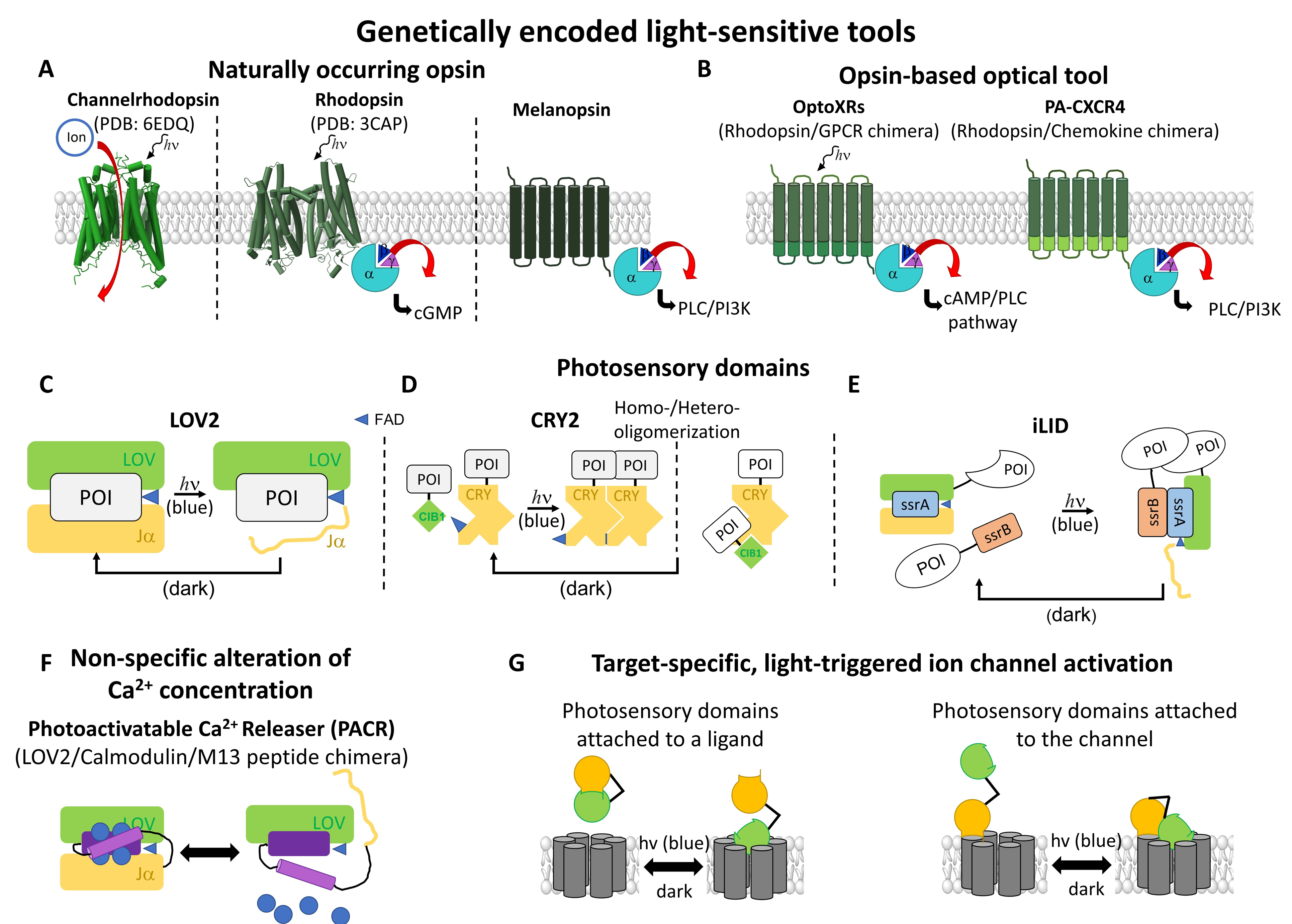
2.2. Light-Sensitive Genetically Encoded Tools
2.2.1. Opsins in Ca2+ Signaling

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Classification | Light-Sensitive Tool | Photosensitive Module | Wave-Length | t1/2 ON | t1/2 OFF | Organism/ Cell Type | Ref. |
|---|---|---|---|---|---|---|---|
| Opsin in Ca2+ signaling | Opto-XRs a | rhodopsin/GPCR chimeras | 504 ± 6 nm/dark | Optoß2AR = 2,1s, Optoa1AR = 125 ms | Optoß2AR = 3 s, Optoa1AR = 533 ms | HEK293 mice | [211] |
| PA-CXCR4 b | chimeric, photoactivatable rhodopsin-chemokine receptor | 505 nm | 3–20 min | HEK293 mice | [214] | ||
| PACR c | LOV2, AsLOV2 | 470 nm/dark | C. elegans HeLa | [215] | |||
| Photosensory proteins | opto-FGFR1 d | AsCryptochrome 2 (CRY2PHR) | 488 nm/dark | ~15 sec | ~40 sec | HeLa HUVECs | [216] |
| opto-RGK e | LOV2-SsrA/SspB | 470 nm/dark | HEK293 C2C12 HL-1 cardiomyocytes | [39] | |||
| OptoCRAC f | LOV2(404–46) | 470 nm/dark | 23.4 ± 4.2 sec | 24.9 ± 4.8 sec | HeLa | [105] | |
| Opto-STIM1 g | CRY2(1–498) | 470 nm/dark | 64.5 ± 4.8 sec | 274 ± 23.7 sec | HeLa | [104] | |
| hBACCS1 h | LOV2 (404–538) | 470 nm/dark | 4.5-fold in [Ca2+] after 30 s | HEK293T | [103] | ||
| hBACCS2 h | LOV2 (404–538) | 470 nm/dark | ~30 sec | ~180 sec | COS-7 HEK293 HUVEC hESCs astrocyte zebrafish embryo mouse hippocampus | ||
| dmBACCS2 i | LOV2 (404–538) | 470 nm/dark | ~30 sec | ~150 sec | |||
| dmBACCS2 NS i | LOV2(404–538) (N425S) | 470 nm/dark | ~30 sec | ~30 sec | |||
| dmBACCS2 i VL | LOV2(404–538) (V416L) | 470 nm/dark | ~30 sec | ~480 sec | |||
| LOCa3 j | AsLOV2 (404–546) | 470 nm/dark | 48.69 ± 4.53 s;~75 sec | 56.84 ± 3.79 sec | HEK293 | [217] |
2.2.2. Photosensory Proteins
- Genetically-Encoded Tools that Modulate Cytosolic Ca2+ Levels
- Light-Triggered Control of Receptor Activity
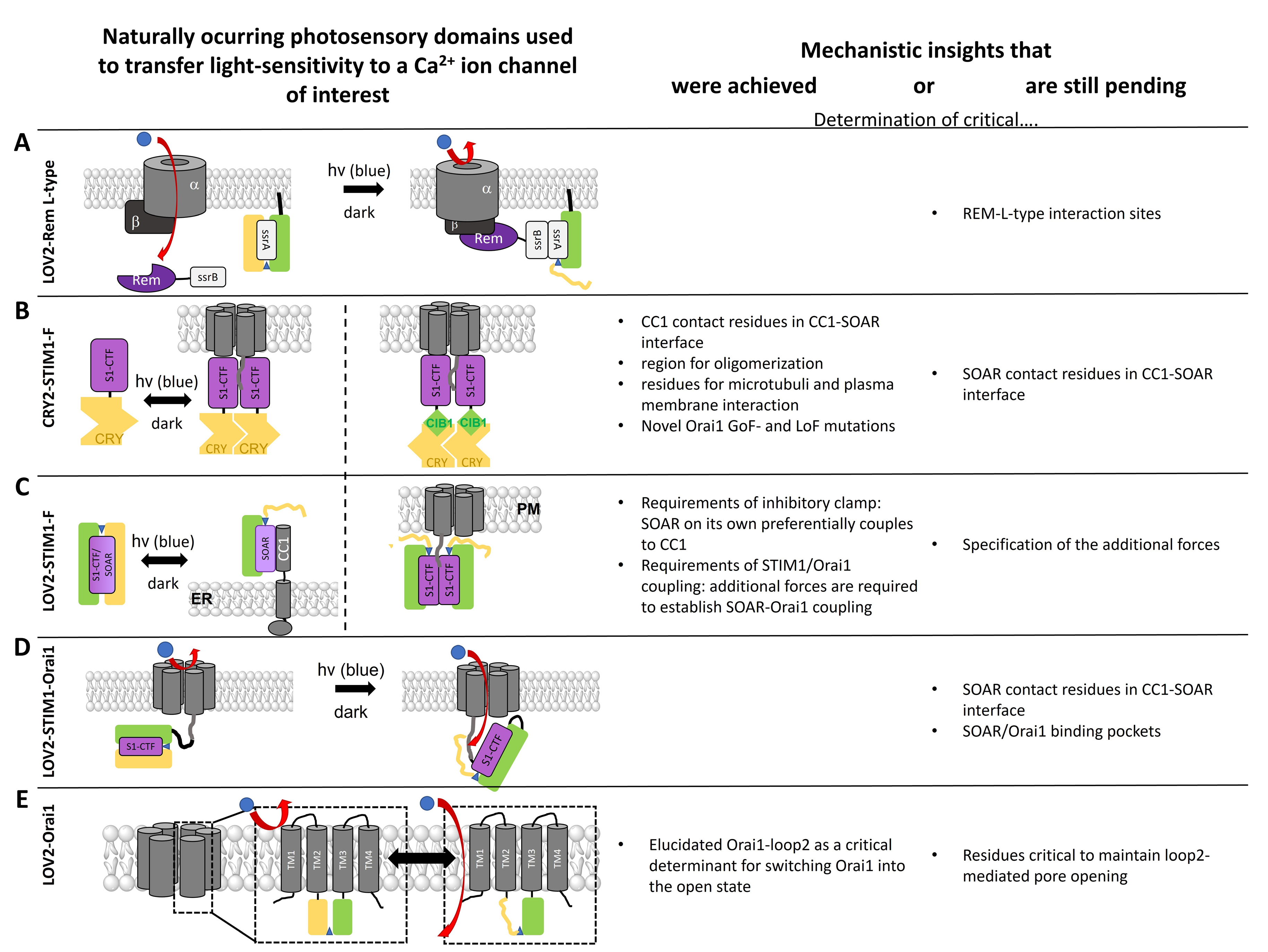
- Photosensory Domains to Transfer Light Sensitivity to Ca2+ Ion Channels
- LOV2 to Modulate L-Type Function
- Light-Sensitive STIM1 Variants Based on Light-Induced Oligomerization
- Light-Sensitive STIM1 Variants Based on Light-Induced Uncaging
- Light-Sensitive Orai Variants
2.2.3. Advantages and Disadvantages of Naturally Occurring Photosensory Proteins
3. Physiological and Pathophysiological Relevance of Novel Optical Tools in the Field of Ca2+ Permeable Ion Channels, Both In Vitro and In Vivo
3.1. Light Sensitive Chemical Tools in Downstream Signaling and Physiology
3.1.1. Caged Ligands
- Caged Ca2+
- Caged Glutamate
- Photolabile EGTA, photocaged IP3 and photocaged lipids
3.1.2. Photosensory Ligands
- In Vitro Applications of the Photosensitive Ligands to Unravel Ca2+-Dependent Processes
- In Vivo Applications of the Photosensitive Ligands to Unravel Ca2+-Dependent Processes
3.2. Light Sensitive Genetically Encoded Tools in Downstream Signaling and Physiology
3.2.1. Opsins
3.2.2. Photosensory Domains
- Light-Sensitive Receptor Opto-FGFR1
- Light-Sensitive CRAC Channel Tools
3.3. Conclusion on In Vitro and In Vivo Application of Optogenetic
4. Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AA | arachidonic acid |
| ACh | acetylincholine |
| AMPA | α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid |
| ARC | arachidonate-regulated calcium current |
| ATP | adenosine-triphosphate |
| BACCS | Blue light-activated Ca2+ channel switch |
| CAD | CRAC activation domain |
| CaM | Calmodulin |
| CIB1 | cryptochrome-interacting basic helix-loop-helix 1 |
| CRAC | Ca2+ relase-activated Ca2+ channel |
| CRY2 | Cryptochrome 2 |
| DAG | diacylglycerol |
| eOS1 | enhanced OptoSTIM1 |
| ER | endoplasmic reticulum |
| ESC | embryonic stem cell |
| FGFR1 | fibroblast growth factor receptor 1 |
| FMN | Flavin mononucleotide |
| FRET | Förster resonance energy transfer |
| GECA | genetically encoded actuator |
| GFP | green flourescent protein |
| HUVEC | human umbilical vein endothelial cell |
| iGluR | ionotropic glutamate receptor |
| iLID | improved light-induced dimer |
| IP3 | inositol 1,4,5-triphosphate |
| LOCa | light-operated Ca2+ channel |
| LOV | Love-oxygen-voltage |
| LRC | LTC4-regulated calcium current |
| LTC4 | leukotriene C4 |
| monSTIM1 | monster-OptoSTIM1 |
| nAChR | nicotinic acetylcholine receptor |
| NFAT | Nuclear factor of activated T-cells |
| NMDR | N-Methyl-D-aspartate receptor |
| PACR | photoactivatable Ca2+ releaser |
| PAS | Period-ARNT-Singleminded |
| PCB | phytocyanobilin |
| PCL | photochromic ligands |
| PHR | photolyase homology region |
| PL | photoswitchable ligand |
| PLC | Phospholipase C |
| PM | plasma membrane |
| PTL | photoswitchable tethered ligand |
| RGC | retinal ganglion cell |
| RTK | receptor tyrosine kinase |
| SICE | store independent calcium entry |
| SOAR | STIM1 Orai activating region |
| SOCE | store-operated calcium entry |
| STIM1 | stromal interaction molecule |
| TM | transmembrane |
| TRP | transient receptor potential -A ankyrin; -C canonical; -M melastatin; -ML mucolipin; -PP polycystic; -V vanilloid |
| UCNP | upconversion nanoparticle |
| UVR8 | UV resistance locus 8 |
| VGCC | Voltage-gated ion channels |
References
- Berridge, M.J.; Bootman, M.D.; Roderick, H.L. Calcium signalling: Dynamics, homeostasis and remodelling. Nat. Rev. Mol. Cell Biol. 2003, 4, 517–529. [Google Scholar] [CrossRef] [Green Version]
- Di Capite, J.L.; Bates, G.J.; Parekh, A.B. Mast cell CRAC channel as a novel therapeutic target in allergy. Curr. Opin. Allergy Clin. Immunol. 2011, 11, 33–38. [Google Scholar] [CrossRef]
- Ruhle, B.; Trebak, M. Emerging roles for native Orai Ca2+ channels in cardiovascular disease. Curr. Top. Membr. 2013, 71, 209–235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eder, P. Cardiac Remodeling and Disease: SOCE and TRPC Signaling in Cardiac Pathology. Adv. Exp. Med. Biol. 2017, 993, 505–521. [Google Scholar] [CrossRef]
- Kiselyov, K.I.; Shin, D.M.; Wang, Y.; Pessah, I.N.; Allen, P.D.; Muallem, S. Gating of store-operated channels by conformational coupling to ryanodine receptors. Mol. Cell 2000, 6, 421–431. [Google Scholar] [CrossRef] [Green Version]
- Ong, H.L.; Subedi, K.P.; Son, G.Y.; Liu, X.; Ambudkar, I.S. Tuning store-operated calcium entry to modulate Ca(2+)-dependent physiological processes. Biochim. Et Biophys. Acta. Mol. Cell Res. 2019, 1866, 1037–1045. [Google Scholar] [CrossRef] [PubMed]
- Berridge, M.J. Calcium microdomains: Organization and function. Cell Calcium 2006, 40, 405–412. [Google Scholar] [CrossRef]
- Berridge, M.J.; Bootman, M.D.; Lipp, P. Calcium—A life and death signal. Nature 1998, 395, 645–648. [Google Scholar] [CrossRef] [PubMed]
- Berridge, M.J.; Lipp, P.; Bootman, M.D. The versatility and universality of calcium signalling. Nat. Rev. Mol. Cell Biol. 2000, 1, 11–21. [Google Scholar] [CrossRef]
- Vaeth, M.; Feske, S. Ion channelopathies of the immune system. Curr. Opin. Immunol. 2018, 52, 39–50. [Google Scholar] [CrossRef]
- Parys, J.B.; Bultynck, G. Calcium signaling in health, disease and therapy. Biochim. Biophys. Acta (BBA)-Mol. Cell Res. 2018, 1865, 1657–1659. [Google Scholar] [CrossRef]
- Lorenzon, N.M.; Beam, K.G. Disease causing mutations of calcium channels. Channels 2008, 2, 163–179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lorenzon, N.M.; Beam, K.G. Calcium channelopathies. Kidney Int. 2000, 57, 794–802. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S. Pharmacological Inhibition of Voltage-gated Ca(2+) Channels for Chronic Pain Relief. Curr. Neuropharmacol. 2013, 11, 606–620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azimi, I.; Stevenson, R.J.; Zhang, X.; Meizoso-Huesca, A.; Xin, P.; Johnson, M.; Flanagan, J.U.; Chalmers, S.B.; Yoast, R.E.; Kapure, J.S.; et al. A new selective pharmacological enhancer of the Orai1 Ca(2+) channel reveals roles for Orai1 in smooth and skeletal muscle functions. ACS Pharmacol. Transl. Sci. 2020, 3, 135–147. [Google Scholar] [CrossRef] [Green Version]
- McDonough, S.I. Calcium ion channels: Challenges and successes in drug discovery. Wiley Interdiscip. Rev. Membr. Transp. Signal. 2013, 2, 85–104. [Google Scholar] [CrossRef]
- Bird, G.S.; Putney, J.W., Jr. Pharmacology of Store-Operated Calcium Entry Channels. In Calcium Entry Channels in Non-Excitable Cells; Kozak, J.A., Putney, J.W., Jr., Eds.; CRC Press: Boca Raton, FL, USA, 2018; pp. 311–324. [Google Scholar]
- Jairaman, A.; Prakriya, M. Molecular pharmacology of store-operated CRAC channels. Channels 2013, 7, 402–414. [Google Scholar] [CrossRef] [Green Version]
- Elliott, A.C. Recent developments in non-excitable cell calcium entry. Cell Calcium 2001, 30, 73–93. [Google Scholar] [CrossRef]
- Catterall, W.A. Voltage-gated calcium channels. Cold Spring Harb. Perspect. Biol. 2011, 3, a003947. [Google Scholar] [CrossRef]
- Prevarskaya, N.; Skryma, R.; Shuba, Y. Ion Channels in Cancer: Are Cancer Hallmarks Oncochannelopathies? Physiol Rev. 2018, 98, 559–621. [Google Scholar] [CrossRef] [Green Version]
- Pitt, G.S.; Matsui, M.; Cao, C. Voltage-Gated Calcium Channels in Nonexcitable Tissues. Annu Rev. Physiol 2021, 83, 183–203. [Google Scholar] [CrossRef] [PubMed]
- Kaestner, L.; Wang, X.; Hertz, L.; Bernhardt, I. Voltage-Activated Ion Channels in Non-excitable Cells-A Viewpoint Regarding Their Physiological Justification. Front. Physiol. 2018, 9, 450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Putney, J.W.; Steinckwich-Besancon, N.; Numaga-Tomita, T.; Davis, F.M.; Desai, P.N.; D’Agostin, D.M.; Wu, S.; Bird, G.S. The functions of store-operated calcium channels. Biochim. Biophys. Acta Mol. Cell Res. 2017, 1864, 900–906. [Google Scholar] [CrossRef] [PubMed]
- Putney, J.W. Store-Operated Calcium Entry: An Historical Overview. Adv. Exp. Med. Biol 2017, 981, 205–214. [Google Scholar] [CrossRef] [PubMed]
- Putney, J.W. The physiological function of store-operated calcium entry. Neurochem. Res. 2011, 36, 1157–1165. [Google Scholar] [CrossRef] [Green Version]
- Feng, M.Y.; Rao, R. New insights into store-independent Ca(2+) entry: Secretory pathway calcium ATPase 2 in normal physiology and cancer. Int. J. Oral Sci. 2013, 5, 71–74. [Google Scholar] [CrossRef]
- Cantonero, C.; Sanchez-Collado, J.; Gonzalez-Nunez, M.A.; Salido, G.M.; Lopez, J.J.; Jardin, I.; Rosado, J.A. Store-independent Orai1-mediated Ca(2+) entry and cancer. Cell Calcium 2019, 80, 1–7. [Google Scholar] [CrossRef]
- Kaczmarek-Hajek, K.; Lorinczi, E.; Hausmann, R.; Nicke, A. Molecular and functional properties of P2X receptors--recent progress and persisting challenges. Purinergic Signal. 2012, 8, 375–417. [Google Scholar] [CrossRef] [Green Version]
- North, R.A. Molecular physiology of P2X receptors. Physiol. Rev. 2002, 82, 1013–1067. [Google Scholar] [CrossRef]
- Petrovic, M.; Horak, M.; Sedlacek, M.; Vyklicky, L., Jr. Physiology and pathology of NMDA receptors. Prague Med. Rep. 2005, 106, 113–136. [Google Scholar]
- Catterall, W.A.; Perez-Reyes, E.; Snutch, T.P.; Striessnig, J. International Union of Pharmacology. XLVIII. Nomenclature and structure-function relationships of voltage-gated calcium channels. Pharmacol. Rev. 2005, 57, 411–425. [Google Scholar] [CrossRef]
- Dolphin, A.C. The alpha2delta subunits of voltage-gated calcium channels. Biochim. Biophys. Acta 2013, 1828, 1541–1549. [Google Scholar] [CrossRef] [Green Version]
- Dolphin, A.C. Voltage-gated calcium channels and their auxiliary subunits: Physiology and pathophysiology and pharmacology. J. Physiol. 2016, 594, 5369–5390. [Google Scholar] [CrossRef] [Green Version]
- Striessnig, J. Voltage-gated calcium channels—From basic mechanisms to disease. J. Physiol. 2016, 594, 5817–5821. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.Y.; Sun, J.; Huang, M.Y.; Wang, Y.S.; Hou, M.F.; Sun, Y.; He, H.; Krishna, N.; Chiu, S.J.; Lin, S.; et al. STIM1 overexpression promotes colorectal cancer progression, cell motility and COX-2 expression. Oncogene 2015, 34, 4358–4367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buchanan, P.J.; McCloskey, K.D. Ca(V) channels and cancer: Canonical functions indicate benefits of repurposed drugs as cancer therapeutics. Eur. Biophys. J. 2016, 45, 621–633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gueguinou, M.; Gambade, A.; Felix, R.; Chantome, A.; Fourbon, Y.; Bougnoux, P.; Weber, G.; Potier-Cartereau, M.; Vandier, C. Lipid rafts, KCa/ClCa/Ca2+ channel complexes and EGFR signaling: Novel targets to reduce tumor development by lipids? Biochim. Biophys. Acta 2015, 1848, 2603–2620. [Google Scholar] [CrossRef] [Green Version]
- Ma, G.; Liu, J.; Ke, Y.; Liu, X.; Li, M.; Wang, F.; Han, G.; Huang, Y.; Wang, Y.; Zhou, Y. Optogenetic Control of Voltage-Gated Calcium Channels. Angew. Chem. 2018, 57, 7019–7022. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.L.; Yu, Y.; Roos, J.; Kozak, J.A.; Deerinck, T.J.; Ellisman, M.H.; Stauderman, K.A.; Cahalan, M.D. STIM1 is a Ca2+ sensor that activates CRAC channels and migrates from the Ca2+ store to the plasma membrane. Nature 2005, 437, 902–905. [Google Scholar] [CrossRef]
- Liou, J.; Kim, M.L.; Heo, W.D.; Jones, J.T.; Myers, J.W.; Ferrell, J.E., Jr.; Meyer, T. STIM is a Ca2+ sensor essential for Ca2+-store-depletion-triggered Ca2+ influx. Curr. Biol. 2005, 15, 1235–1241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feske, S.; Gwack, Y.; Prakriya, M.; Srikanth, S.; Puppel, S.H.; Tanasa, B.; Hogan, P.G.; Lewis, R.S.; Daly, M.; Rao, A. A mutation in Orai1 causes immune deficiency by abrogating CRAC channel function. Nature 2006, 441, 179–185. [Google Scholar] [CrossRef]
- Prakriya, M.; Feske, S.; Gwack, Y.; Srikanth, S.; Rao, A.; Hogan, P.G. Orai1 is an essential pore subunit of the CRAC channel. Nature 2006, 443, 230–233. [Google Scholar] [CrossRef]
- Wu, M.M.; Buchanan, J.; Luik, R.M.; Lewis, R.S. Ca2+ store depletion causes STIM1 to accumulate in ER regions closely associated with the plasma membrane. J. Cell Biol. 2006, 174, 803–813. [Google Scholar] [CrossRef]
- Prakriya, M.; Lewis, R.S. Store-Operated Calcium Channels. Physiol. Rev. 2015, 95, 1383–1436. [Google Scholar] [CrossRef] [Green Version]
- Fahrner, M.; Grabmayr, H.; Romanin, C. Mechanism of STIM activation. Curr. Opin. Physiol. 2020, 17, 74–79. [Google Scholar] [CrossRef] [PubMed]
- Grabmayr, H.; Romanin, C.; Fahrner, M. STIM Proteins: An Ever-Expanding Family. Int. J. Mol. Sci. 2020, 22, 378. [Google Scholar] [CrossRef]
- Butorac, C.; Krizova, A.; Derler, I. Review: Structure and Activation Mechanisms of CRAC Channels. Adv. Exp. Med. Biol. 2020, 1131, 547–604. [Google Scholar] [CrossRef] [PubMed]
- Hou, X.; Outhwaite, I.R.; Pedi, L.; Long, S.B. Cryo-EM structure of the calcium release-activated calcium channel Orai in an open conformation. eLife 2020, 9, e62772. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Wu, G.; Yu, Y.; Chen, X.; Ji, R.; Lu, J.; Li, X.; Zhang, X.; Yang, X.; Shen, Y. Molecular understanding of calcium permeation through the open Orai channel. PLoS Biol. 2019, 17, e3000096. [Google Scholar] [CrossRef]
- Hou, X.; Burstein, S.R.; Long, S.B. Structures reveal opening of the store-operated calcium channel Orai. eLife 2018, 7, e36758. [Google Scholar] [CrossRef]
- Hou, X.; Pedi, L.; Diver, M.M.; Long, S.B. Crystal structure of the calcium release-activated calcium channel Orai. Science 2012, 338, 1308–1313. [Google Scholar] [CrossRef] [Green Version]
- Schindl, R.; Frischauf, I.; Bergsmann, J.; Muik, M.; Derler, I.; Lackner, B.; Groschner, K.; Romanin, C. Plasticity in Ca2+ selectivity of Orai1/Orai3 heteromeric channel. Proc. Natl. Acad. Sci. USA 2009, 106, 19623–19628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gwack, Y.; Srikanth, S.; Feske, S.; Cruz-Guilloty, F.; Oh-hora, M.; Neems, D.S.; Hogan, P.G.; Rao, A. Biochemical and functional characterization of Orai proteins. J. Biol. Chem. 2007, 282, 16232–16243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lis, A.; Peinelt, C.; Beck, A.; Parvez, S.; Monteilh-Zoller, M.; Fleig, A.; Penner, R. CRACM1, CRACM2, and CRACM3 are store-operated Ca2+ channels with distinct functional properties. Curr. Biol. 2007, 17, 794–800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; Meraner, P.; Kwon, H.T.; Machnes, D.; Oh-hora, M.; Zimmer, J.; Huang, Y.; Stura, A.; Rao, A.; Hogan, P.G. STIM1 gates the store-operated calcium channel ORAI1 in vitro. Nat. Struct. Mol. Biol. 2010, 17, 112–116. [Google Scholar] [CrossRef] [Green Version]
- Yoast, R.E.; Emrich, S.M.; Trebak, M. The anatomy of native CRAC channel(s). Curr. Opin. Physiol. 2020, 17, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Tiffner, A.; Maltan, L.; Weiss, S.; Derler, I. The Orai Pore Opening Mechanism. Int. J. Mol. Sci. 2021, 22, 533. [Google Scholar] [CrossRef] [PubMed]
- Tiffner, A.; Schober, R.; Hoeglinger, C.; Bonhenry, D.; Pandey, S.; Lunz, V.; Sallinger, M.; Frischauf, I.; Fahrner, M.; Lindinger, S.; et al. CRAC channel opening is determined by a series of Orai1 gating checkpoints in the transmembrane and cytosolic regions. J. Biol. Chem. 2020. [Google Scholar] [CrossRef]
- Zhang, X.; Gueguinou, M.; Trebak, M. Store-Independent Orai Channels Regulated by STIM. In Calcium Entry Channels in Non-Excitable Cells; Kozak, J.A., Putney, J.W., Jr., Eds.; CRC Press: Boca Raton, FL, USA; Taylor & Francis: Abingdon, UK, 2018; pp. 197–214. [Google Scholar]
- Tiffner, A.; Derler, I. Isoform-Specific Properties of Orai Homologues in Activation, Downstream Signaling, Physiology and Pathophysiology. Int. J. Mol. Sci. 2021, 22, 8020. [Google Scholar] [CrossRef]
- Motiani, R.K.; Stolwijk, J.A.; Newton, R.L.; Zhang, X.; Trebak, M. Emerging roles of Orai3 in pathophysiology. Channels 2013, 7, 392–401. [Google Scholar] [CrossRef] [Green Version]
- Feng, M.; Grice, D.M.; Faddy, H.M.; Nguyen, N.; Leitch, S.; Wang, Y.; Muend, S.; Kenny, P.A.; Sukumar, S.; Roberts-Thomson, S.J.; et al. Store-independent activation of Orai1 by SPCA2 in mammary tumors. Cell 2010, 143, 84–98. [Google Scholar] [CrossRef] [Green Version]
- Gueguinou, M.; Harnois, T.; Crottes, D.; Uguen, A.; Deliot, N.; Gambade, A.; Chantome, A.; Haelters, J.P.; Jaffres, P.A.; Jourdan, M.L.; et al. SK3/TRPC1/Orai1 complex regulates SOCE-dependent colon cancer cell migration: A novel opportunity to modulate anti-EGFR mAb action by the alkyl-lipid Ohmline. Oncotarget 2016, 7, 36168–36184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clarysse, L.; Gueguinou, M.; Potier-Cartereau, M.; Vandecasteele, G.; Bougnoux, P.; Chevalier, S.; Chantome, A.; Vandier, C. cAMP-PKA inhibition of SK3 channel reduced both Ca2+ entry and cancer cell migration by regulation of SK3-Orai1 complex. Pflug. Arch. 2014, 466, 1921–1932. [Google Scholar] [CrossRef] [PubMed]
- Chantome, A.; Potier-Cartereau, M.; Clarysse, L.; Fromont, G.; Marionneau-Lambot, S.; Gueguinou, M.; Pages, J.C.; Collin, C.; Oullier, T.; Girault, A.; et al. Pivotal role of the lipid Raft SK3-Orai1 complex in human cancer cell migration and bone metastases. Cancer Res. 2013, 73, 4852–4861. [Google Scholar] [CrossRef] [Green Version]
- Gueguinou, M.; Crottes, D.; Chantome, A.; Rapetti-Mauss, R.; Potier-Cartereau, M.; Clarysse, L.; Girault, A.; Fourbon, Y.; Jezequel, P.; Guerin-Charbonnel, C.; et al. The SigmaR1 chaperone drives breast and colorectal cancer cell migration by tuning SK3-dependent Ca(2+) homeostasis. Oncogene 2017, 36, 3640–3647. [Google Scholar] [CrossRef]
- Peretti, M.; Badaoui, M.; Girault, A.; Van Gulick, L.; Mabille, M.P.; Tebbakha, R.; Sevestre, H.; Morjani, H.; Ouadid-Ahidouch, H. Original association of ion transporters mediates the ECM-induced breast cancer cell survival: Kv10.1-Orai1-SPCA2 partnership. Sci. Rep. 2019, 9, 1175. [Google Scholar] [CrossRef]
- Montell, C.; Rubin, G.M. Molecular characterization of the Drosophila trp locus: A putative integral membrane protein required for phototransduction. Neuron 1989, 2, 1313–1323. [Google Scholar] [CrossRef]
- Nilius, B.; Owsianik, G.; Voets, T.; Peters, J.A. Transient receptor potential cation channels in disease. Physiol. Rev. 2007, 87, 165–217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gees, M.; Colsoul, B.; Nilius, B. The role of transient receptor potential cation channels in Ca2+ signaling. Cold Spring Harb. Perspect. Biol. 2010, 2, a003962. [Google Scholar] [CrossRef] [Green Version]
- Benemei, S.; Patacchini, R.; Trevisani, M.; Geppetti, P. TRP channels. Curr. Opin. Pharmacol. 2015, 22, 18–23. [Google Scholar] [CrossRef]
- Cao, E. Structural mechanisms of transient receptor potential ion channels. J. Gen. Physiol. 2020, 152. [Google Scholar] [CrossRef]
- Brauchi, S.; Orio, P. Voltage sensing in thermo-TRP channels. Adv. Exp. Med. Biol. 2011, 704, 517–530. [Google Scholar] [CrossRef]
- Zheng, J. Molecular mechanism of TRP channels. Compr. Physiol. 2013, 3, 221–242. [Google Scholar] [CrossRef] [Green Version]
- Ong, H.L.; de Souza, L.B.; Ambudkar, I.S. Role of TRPC Channels in Store-Operated Calcium Entry. Adv. Exp. Med. Biol. 2016, 898, 87–109. [Google Scholar] [CrossRef]
- Cheng, K.T.; Ong, H.L.; Liu, X.; Ambudkar, I.S. Contribution and regulation of TRPC channels in store-operated Ca2+ entry. Curr. Top. Membr. 2013, 71, 149–179. [Google Scholar] [CrossRef] [Green Version]
- Salido, G.M.; Sage, S.O.; Rosado, J.A. TRPC channels and store-operated Ca(2+) entry. Biochim. Biophys. Acta 2009, 1793, 223–230. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Trebak, M. Transient receptor potential canonical 7: A diacylglycerol-activated non-selective cation channel. Handb. Exp. Pharmacol. 2014, 222, 189–204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dryer, S.E.; Kim, E.Y. Permeation and Rectification in Canonical Transient Receptor Potential-6 (TRPC6) Channels. Front. Physiol. 2018, 9, 1055. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.P.; Zeng, W.; Huang, G.N.; Worley, P.F.; Muallem, S. STIM1 heteromultimerizes TRPC channels to determine their function as store-operated channels. Nat. Cell Biol. 2007, 9, 636–645. [Google Scholar] [CrossRef]
- Worley, P.F.; Zeng, W.; Huang, G.N.; Yuan, J.P.; Kim, J.Y.; Lee, M.G.; Muallem, S. TRPC channels as STIM1-regulated store-operated channels. Cell Calcium 2007, 42, 205–211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meents, J.E.; Ciotu, C.I.; Fischer, M.J.M. TRPA1: A molecular view. J. Neurophysiol. 2019, 121, 427–443. [Google Scholar] [CrossRef]
- Prevarskaya, N.; Zhang, L.; Barritt, G. TRP channels in cancer. Biochim. Biophys. Acta 2007, 1772, 937–946. [Google Scholar] [CrossRef] [Green Version]
- Canales, J.; Morales, D.; Blanco, C.; Rivas, J.; Díaz, N.; Angelopoulos, I.; Cerda, O. A TR(i)P to Cell Migration: New Roles of TRP Channels in Mechanotransduction and Cancer. Front. Physiol. 2019, 10, 757. [Google Scholar] [CrossRef]
- Shapovalov, G.; Ritaine, A.; Skryma, R.; Prevarskaya, N. Role of TRP ion channels in cancer and tumorigenesis. Semin. ImmunoPathol. 2016, 38, 357–369. [Google Scholar] [CrossRef]
- Curcic, S.; Tiapko, O.; Groschner, K. Photopharmacology and opto-chemogenetics of TRPC channels-some therapeutic visions. Pharmacol. Ther. 2019, 200, 13–26. [Google Scholar] [CrossRef]
- Pankratov, Y.; Lalo, U. Calcium permeability of ligand-gated Ca2+ channels. Eur. J. Pharmacol. 2014, 739, 60–73. [Google Scholar] [CrossRef] [PubMed]
- Twomey, E.C.; Sobolevsky, A.I. Structural Mechanisms of Gating in Ionotropic Glutamate Receptors. Biochemistry 2018, 57, 267–276. [Google Scholar] [CrossRef]
- Wollmuth, L.P. Ion permeation in ionotropic glutamate receptors: Still dynamic after all these years. Curr. Opin. Physiol. 2018, 2, 36–41. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Sheng, M. NMDA receptors in nervous system diseases. Neuropharmacology 2013, 74, 69–75. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.; Goodman, L.; Fourie, C.; Schenk, S.; Leitch, B.; Montgomery, J.M. AMPA Receptors as Therapeutic Targets for Neurological Disorders. Adv. Protein Chem. Struct. Biol. 2016, 103, 203–261. [Google Scholar] [CrossRef] [PubMed]
- Yi, H.; Talmon, G.; Wang, J. Glutamate in cancers: From metabolism to signaling. J. Biomed. Res. 2019, 34, 260–270. [Google Scholar] [CrossRef] [PubMed]
- Burnstock, G.; Kennedy, C. P2X receptors in health and disease. Adv. Pharmacol. 2011, 61, 333–372. [Google Scholar] [CrossRef] [PubMed]
- Di Virgilio, F.; Vultaggio-Poma, V.; Sarti, A.C. P2X receptors in cancer growth and progression. Biochem. Pharm. 2021, 187, 114350. [Google Scholar] [CrossRef]
- Thompson, A.J.; Lester, H.A.; Lummis, S.C. The structural basis of function in Cys-loop receptors. Q. Rev. Biophys. 2010, 43, 449–499. [Google Scholar] [CrossRef] [Green Version]
- Lindstrom, J. Nicotinic acetylcholine receptors in health and disease. Mol. Neurobiol. 1997, 15, 193–222. [Google Scholar] [CrossRef]
- Chen, J.; Cheuk, I.W.Y.; Shin, V.Y.; Kwong, A. Acetylcholine receptors: Key players in cancer development. Surg. Oncol. 2019, 31, 46–53. [Google Scholar] [CrossRef] [PubMed]
- Traynelis, S.F.; Wollmuth, L.P.; McBain, C.J.; Menniti, F.S.; Vance, K.M.; Ogden, K.K.; Hansen, K.B.; Yuan, H.; Myers, S.J.; Dingledine, R. Glutamate receptor ion channels: Structure, regulation, and function. Pharmacol. Rev. 2010, 62, 405–496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wonnacott, S.; Bermudez, I.; Millar, N.S.; Tzartos, S.J. Nicotinic acetylcholine receptors. Br. J. Pharm. 2018, 175, 1785–1788. [Google Scholar] [CrossRef] [Green Version]
- Guru, A.; Post, R.J.; Ho, Y.Y.; Warden, M.R. Making Sense of Optogenetics. Int. J. Neuropsychopharmacol. 2015, 18, pyv079. [Google Scholar] [CrossRef] [PubMed]
- Boyden, E.S.; Zhang, F.; Bamberg, E.; Nagel, G.; Deisseroth, K. Millisecond-timescale, genetically targeted optical control of neural activity. Nat. Neurosci. 2005, 8, 1263–1268. [Google Scholar] [CrossRef]
- Ishii, T.; Sato, K.; Kakumoto, T.; Miura, S.; Touhara, K.; Takeuchi, S.; Nakata, T. Light generation of intracellular Ca(2+) signals by a genetically encoded protein BACCS. Nat. Commun. 2015, 6, 8021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kyung, T.; Lee, S.; Kim, J.E.; Cho, T.; Park, H.; Jeong, Y.M.; Kim, D.; Shin, A.; Kim, S.; Baek, J.; et al. Optogenetic control of endogenous Ca(2+) channels in vivo. Nat. Biotechnol. 2015, 33, 1092–1096. [Google Scholar] [CrossRef]
- He, L.; Zhang, Y.; Ma, G.; Tan, P.; Li, Z.; Zang, S.; Wu, X.; Jing, J.; Fang, S.; Zhou, L.; et al. Near-infrared photoactivatable control of Ca(2+) signaling and optogenetic immunomodulation. eLife 2015, 4, e10024. [Google Scholar] [CrossRef]
- Paoletti, P.; Ellis-Davies, G.C.R.; Mourot, A. Optical control of neuronal ion channels and receptors. Nat. Rev. Neurosci. 2019, 20, 514–532. [Google Scholar] [CrossRef]
- Berlin, S.; Isacoff, E.Y. Synapses in the spotlight with synthetic optogenetics. EMBO Rep. 2017, 18, 677–692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ellis-Davies, G.C. Caged compounds: Photorelease technology for control of cellular chemistry and physiology. Nat. Methods 2007, 4, 619–628. [Google Scholar] [CrossRef] [Green Version]
- Ellis-Davies, G.C.; Kaplan, J.H. Nitrophenyl-EGTA, a photolabile chelator that selectively binds Ca2+ with high affinity and releases it rapidly upon photolysis. Proc. Natl. Acad. Sci. USA 1994, 91, 187–191. [Google Scholar] [CrossRef] [Green Version]
- Zimmermann, B.; Somlyo, A.V.; Ellis-Davies, G.C.R.; Kaplan, J.H.; Somlyo, A.P. Kinetics of Prephosphorylation Reactions and Myosin Light Chain Phosphorylation in Smooth Muscle: Flash Photolysis studies with caged calcium and caged ATP (∗). J. Biol. Chem. 1995, 270, 23966–23974. [Google Scholar] [CrossRef] [Green Version]
- Ellis-Davies, G.C.R.; Barsotti, R.J. Tuning caged calcium: Photolabile analogues of EGTA with improved optical and chelation properties. Cell Calcium 2006, 39, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Tsien, R.Y.; Zucker, R.S. Control of cytoplasmic calcium with photolabile tetracarboxylate 2-nitrobenzhydrol chelators. Biophys. J. 1986, 50, 843–853. [Google Scholar] [CrossRef] [Green Version]
- Morad, M.; Davies, N.W.; Kaplan, J.H.; Lux, H.D. Inactivation and block of calcium channels by photo-released Ca2+ in dorsal root ganglion neurons. Science 1988, 241, 842–844. [Google Scholar] [CrossRef]
- Harootunian, A.T.; Kao, J.P.; Tsien, R.Y. Agonist-induced calcium oscillations in depolarized fibroblasts and their manipulation by photoreleased Ins(1,4,5)P3, Ca++, and Ca++ buffer. Cold Spring Harb Symp Quant. Biol 1988, 53 Pt 2, 935–943. [Google Scholar] [CrossRef]
- Mulligan, I.P.; Ashley, C.C. Rapid relaxation of single frog skeletal muscle fibres following laser flash photolysis of the caged calcium chelator, diazo-2. FEBS Lett. 1989, 255, 196–200. [Google Scholar] [CrossRef] [Green Version]
- Lancaster, B.; Zucker, R.S. Photolytic manipulation of Ca2+ and the time course of slow, Ca(2+)-activated K+ current in rat hippocampal neurones. J. Physiol. 1994, 475, 229–239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palmer, R.E.; Mulligan, I.P.; Nunn, C.; Ashley, C.C. Striated scallop muscle relaxation: Fast force transients produced by photolysis of Diazo-2. Biochem. Biophys. Res. Commun. 1990, 168, 295–300. [Google Scholar] [CrossRef]
- Fryer, M.W.; Zucker, R.S. Ca(2+)-dependent inactivation of Ca2+ current in Aplysia neurons: Kinetic studies using photolabile Ca2+ chelators. J. Physiol. 1993, 464, 501–528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sah, P.; Clements, J.D. Photolytic manipulation of [Ca2+]i reveals slow kinetics of potassium channels underlying the afterhyperpolarization in hippocampal pyramidal neurons. J. Neurosci. Off. J. Soc. Neurosci. 1999, 19, 3657–3664. [Google Scholar] [CrossRef] [Green Version]
- Walker, J.W.; Somlyo, A.V.; Goldman, Y.E.; Somlyo, A.P.; Trentham, D.R. Kinetics of smooth and skeletal muscle activation by laser pulse photolysis of caged inositol 1,4,5-trisphosphate. Nature 1987, 327, 249–252. [Google Scholar] [CrossRef]
- Adams, S.R.; Tsien, R.Y. Controlling cell chemistry with caged compounds. Annu. Rev. Physiol. 1993, 55, 755–784. [Google Scholar] [CrossRef]
- Kaplan, J.H.; Somlyo, A.P. Flash photolysis of caged compounds: New tools for cellular physiology. Trends Neurosci. 1989, 12, 54–59. [Google Scholar] [CrossRef]
- Gurney, A.M.; Lester, H.A. Light-flash physiology with synthetic photosensitive compounds. Physiol. Rev. 1987, 67, 583–617. [Google Scholar] [CrossRef]
- Tan, P.; He, L.; Han, G.; Zhou, Y. Optogenetic Immunomodulation: Shedding Light on Antitumor Immunity. Trends Biotechnol. 2017, 35, 215–226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zucker, R.S.; Haydon, P.G. Membrane potential has no direct role in evoking neurotransmitter release. Nature 1988, 335, 360–362. [Google Scholar] [CrossRef]
- Kaplan, J.H.; Ellis-Davies, G.C. Photolabile chelators for the rapid photorelease of divalent cations. Proc. Natl. Acad. Sci. USA 1988, 85, 6571–6575. [Google Scholar] [CrossRef] [Green Version]
- Goldman, Y.E.; Kaplan, J.H. Activation of skeletal muscle fibers by photolysis of DM-nitrophen, a new caged Ca2+. Biophys. J. 1988, 53, 25a. [Google Scholar]
- Dantzig, J.A.; Walker, J.W.; Trentham, D.R.; Goldman, Y.E. Relaxation of muscle fibers with adenosine 5’-[gamma-thio]triphosphate (ATP[gamma S]) and by laser photolysis of caged ATP[gamma S]: Evidence for Ca2+-dependent affinity of rapidly detaching zero-force cross-bridges. Proc. Natl. Acad. Sci. USA 1988, 85, 6716–6720. [Google Scholar] [CrossRef] [Green Version]
- Losonczy, A.; Magee, J.C. Integrative properties of radial oblique dendrites in hippocampal CA1 pyramidal neurons. Neuron 2006, 50, 291–307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Somlyo, A.P.; Walker, J.W.; Goldman, Y.E.; Trentham, D.R.; Kobayashi, S.; Kitazawa, T.; Somlyo, A.V. Inositol trisphosphate, calcium and muscle contraction. Philos. Trans. R. Soc. London. Ser. B Biol. Sci. 1988, 320, 399–414. [Google Scholar] [CrossRef]
- Wang, Y.; Deshpande, M.; Payne, R. 2-Aminoethoxydiphenyl borate inhibits phototransduction and blocks voltage-gated potassium channels in Limulus ventral photoreceptors. Cell Calcium 2002, 32, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Li, W.-h.; Llopis, J.; Whitney, M.; Zlokarnik, G.; Tsien, R.Y. Cell-permeant caged InsP3 ester shows that Ca2+ spike frequency can optimize gene expression. Nature 1998, 392, 936–941. [Google Scholar] [CrossRef] [PubMed]
- Miyamoto, A.; Mikoshiba, K. Probes for manipulating and monitoring IP3. Cell Calcium 2017, 64, 57–64. [Google Scholar] [CrossRef]
- Berridge, M.J. The Inositol Trisphosphate/Calcium Signaling Pathway in Health and Disease. Physiol. Rev. 2016, 96, 1261–1296. [Google Scholar] [CrossRef] [Green Version]
- Horne, J.H.; Meyer, T. Elementary calcium-release units induced by inositol trisphosphate. Science 1997, 276, 1690–1693. [Google Scholar] [CrossRef] [PubMed]
- Hartung, K.; Grell, E.; Hasselbach, W.; Bamberg, E. Electrical pump currents generated by the Ca2+-ATPase of sarcoplasmic reticulum vesicles adsorbed on black lipid membranes. Biochim. Biophys. Acta 1987, 900, 209–220. [Google Scholar] [CrossRef]
- Wieboldt, R.; Gee, K.R.; Niu, L.; Ramesh, D.; Carpenter, B.K.; Hess, G.P. Photolabile precursors of glutamate: Synthesis, photochemical properties, and activation of glutamate receptors on a microsecond time scale. Proc. Natl. Acad. Sci. USA 1994, 91, 8752–8756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuzaki, M.; Ellis-Davies, G.C.; Nemoto, T.; Miyashita, Y.; Iino, M.; Kasai, H. Dendritic spine geometry is critical for AMPA receptor expression in hippocampal CA1 pyramidal neurons. Nat. Neurosci. 2001, 4, 1086–1092. [Google Scholar] [CrossRef] [Green Version]
- Matsuzaki, M.; Honkura, N.; Ellis-Davies, G.C.; Kasai, H. Structural basis of long-term potentiation in single dendritic spines. Nature 2004, 429, 761–766. [Google Scholar] [CrossRef] [PubMed]
- Brini, M.; Carafoli, E. Calcium pumps in health and disease. Physiol. Rev. 2009, 89, 1341–1378. [Google Scholar] [CrossRef] [Green Version]
- Walker, J.W.; McCray, J.A.; Hess, G.P. Photolabile protecting groups for an acetylcholine receptor ligand. Synthesis and photochemistry of a new class of o-nitrobenzyl derivatives and their effects on receptor function. Biochemistry 1986, 25, 1799–1805. [Google Scholar] [CrossRef]
- Milburn, T.; Matsubara, N.; Billington, A.P.; Udgaonkar, J.B.; Walker, J.W.; Carpenter, B.K.; Webb, W.W.; Marque, J.; Denk, W.; McCray, J.A.; et al. Synthesis, photochemistry, and biological activity of a caged photolabile acetylcholine receptor ligand. Biochemistry 1989, 28, 49–55. [Google Scholar] [CrossRef]
- Banala, S.; Arvin, M.C.; Bannon, N.M.; Jin, X.T.; Macklin, J.J.; Wang, Y.; Peng, C.; Zhao, G.; Marshall, J.J.; Gee, K.R.; et al. Photoactivatable drugs for nicotinic optopharmacology. Nat. Methods 2018, 15, 347–350. [Google Scholar] [CrossRef] [Green Version]
- Passlick, S.; Thapaliya, E.R.; Chen, Z.; Richers, M.T.; Ellis-Davies, G.C.R. Optical probing of acetylcholine receptors on neurons in the medial habenula with a novel caged nicotine drug analogue. J. Physiol. 2018, 596, 5307–5318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niu, L.; Wieboldt, R.; Ramesh, D.; Carpenter, B.K.; Hess, G.P. Synthesis and characterization of a caged receptor ligand suitable for chemical kinetic investigations of the glycine receptor in the 3-microseconds time domain. Biochemistry 1996, 35, 8136–8142. [Google Scholar] [CrossRef] [PubMed]
- Palma-Cerda, F.; Auger, C.; Crawford, D.J.; Hodgson, A.C.; Reynolds, S.J.; Cowell, J.K.; Swift, K.A.; Cais, O.; Vyklicky, L.; Corrie, J.E.; et al. New caged neurotransmitter analogs selective for glutamate receptor sub-types based on methoxynitroindoline and nitrophenylethoxycarbonyl caging groups. Neuropharmacology 2012, 63, 624–634. [Google Scholar] [CrossRef] [PubMed]
- Gee, K.R.; Niu, L.; Schaper, K.; Jayaraman, V.; Hess, G.P. Synthesis and photochemistry of a photolabile precursor of N-methyl-D-aspartate (NMDA) that is photolyzed in the microsecond time region and is suitable for chemical kinetic investigations of the NMDA receptor. Biochemistry 1999, 38, 3140–3147. [Google Scholar] [CrossRef]
- Hartung, K.; Froehlich, J.P.; Fendler, K. Time-resolved charge translocation by the Ca-ATPase from sarcoplasmic reticulum after an ATP concentration jump. Biophys J. 1997, 72, 2503–2514. [Google Scholar] [CrossRef] [Green Version]
- Mourot, A.; Fehrentz, T.; Le Feuvre, Y.; Smith, C.M.; Herold, C.; Dalkara, D.; Nagy, F.; Trauner, D.; Kramer, R.H. Rapid optical control of nociception with an ion-channel photoswitch. Nat. Methods 2012, 9, 396–402. [Google Scholar] [CrossRef]
- Fehrentz, T.; Kuttruff, C.A.; Huber, F.M.; Kienzler, M.A.; Mayer, P.; Trauner, D. Exploring the pharmacology and action spectra of photochromic open-channel blockers. Chembiochem A Eur. J. Chem. Biol. 2012, 13, 1746–1749. [Google Scholar] [CrossRef]
- Banghart, M.R.; Mourot, A.; Fortin, D.L.; Yao, J.Z.; Kramer, R.H.; Trauner, D. Photochromic blockers of voltage-gated potassium channels. Angew. Chem. 2009, 48, 9097–9101. [Google Scholar] [CrossRef] [Green Version]
- Frolova, S.R.; Gaiko, O.; Tsvelaya, V.A.; Pimenov, O.Y.; Agladze, K.I. Photocontrol of Voltage-Gated Ion Channel Activity by Azobenzene Trimethylammonium Bromide in Neonatal Rat Cardiomyocytes. PLoS ONE 2016, 11, e0152018. [Google Scholar] [CrossRef]
- Frank, J.A.; Moroni, M.; Moshourab, R.; Sumser, M.; Lewin, G.R.; Trauner, D. Photoswitchable fatty acids enable optical control of TRPV1. Nat. Commun 2015, 6, 7118. [Google Scholar] [CrossRef] [Green Version]
- Stein, M.; Breit, A.; Fehrentz, T.; Gudermann, T.; Trauner, D. Optical control of TRPV1 channels. Angew. Chem. 2013, 52, 9845–9848. [Google Scholar] [CrossRef] [PubMed]
- Kokel, D.; Cheung, C.Y.; Mills, R.; Coutinho-Budd, J.; Huang, L.; Setola, V.; Sprague, J.; Jin, S.; Jin, Y.N.; Huang, X.P.; et al. Photochemical activation of TRPA1 channels in neurons and animals. Nat. Chem. Biol. 2013, 9, 257–263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lam, P.Y.; Mendu, S.K.; Mills, R.W.; Zheng, B.; Padilla, H.; Milan, D.J.; Desai, B.N.; Peterson, R.T. A high-conductance chemo-optogenetic system based on the vertebrate channel Trpa1b. Sci. Rep. 2017, 7, 11839. [Google Scholar] [CrossRef]
- Leinders-Zufall, T.; Storch, U.; Bleymehl, K.; Mederos, Y.S.M.; Frank, J.A.; Konrad, D.B.; Trauner, D.; Gudermann, T.; Zufall, F. PhoDAGs Enable Optical Control of Diacylglycerol-Sensitive Transient Receptor Potential Channels. Cell Chem. Biol. 2018, 25, 215–223.e213. [Google Scholar] [CrossRef] [Green Version]
- Lichtenegger, M.; Tiapko, O.; Svobodova, B.; Stockner, T.; Glasnov, T.N.; Schreibmayer, W.; Platzer, D.; de la Cruz, G.G.; Krenn, S.; Schober, R.; et al. An optically controlled probe identifies lipid-gating fenestrations within the TRPC3 channel. Nat. Chem. Biol. 2018, 14, 396–404. [Google Scholar] [CrossRef]
- Yang, X.; Ma, G.; Zheng, S.; Qin, X.; Li, X.; Du, L.; Wang, Y.; Zhou, Y.; Li, M. Optical Control of CRAC Channels Using Photoswitchable Azopyrazoles. J. Am. Chem. Soc. 2020, 142, 9460–9470. [Google Scholar] [CrossRef] [PubMed]
- Stawski, P.; Sumser, M.; Trauner, D. A photochromic agonist of AMPA receptors. Angew. Chem. 2012, 51, 5748–5751. [Google Scholar] [CrossRef]
- Wolter, T.; Steinbrecher, T.; Trauner, D.; Elstner, M. Ligand photo-isomerization triggers conformational changes in iGluR2 ligand binding domain. PLoS ONE 2014, 9, e92716. [Google Scholar] [CrossRef]
- Barber, D.M.; Liu, S.A.; Gottschling, K.; Sumser, M.; Hollmann, M.; Trauner, D. Optical control of AMPA receptors using a photoswitchable quinoxaline-2,3-dione antagonist. Chem. Sci. 2017, 8, 611–615. [Google Scholar] [CrossRef] [Green Version]
- Volgraf, M.; Gorostiza, P.; Szobota, S.; Helix, M.R.; Isacoff, E.Y.; Trauner, D. Reversibly caged glutamate: A photochromic agonist of ionotropic glutamate receptors. J. Am. Chem. Soc. 2007, 129, 260–261. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Wolter, T.; Kubar, T.; Sumser, M.; Trauner, D.; Elstner, M. Molecular Dynamics Investigation of gluazo, a Photo-Switchable Ligand for the Glutamate Receptor GluK2. PLoS ONE 2015, 10, e0135399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abrams, Z.R.; Warrier, A.; Trauner, D.; Zhang, X. A Signal Processing Analysis of Purkinje Cells in vitro. Front. Neural Circuits 2010, 4, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, D.; Herault, K.; Isacoff, E.Y.; Oheim, M.; Ropert, N. Optogenetic activation of LiGluR-expressing astrocytes evokes anion channel-mediated glutamate release. J. Physiol. 2012, 590, 855–873. [Google Scholar] [CrossRef]
- Levitz, J.; Popescu, A.T.; Reiner, A.; Isacoff, E.Y. A Toolkit for Orthogonal and in vivo Optical Manipulation of Ionotropic Glutamate Receptors. Front. Mol. Neurosci. 2016, 9, 2. [Google Scholar] [CrossRef] [Green Version]
- Izquierdo-Serra, M.; Trauner, D.; Llobet, A.; Gorostiza, P. Optical modulation of neurotransmission using calcium photocurrents through the ion channel LiGluR. Front. Mol. Neurosci. 2013, 6, 3. [Google Scholar] [CrossRef] [Green Version]
- Levitz, J.; Pantoja, C.; Gaub, B.; Janovjak, H.; Reiner, A.; Hoagland, A.; Schoppik, D.; Kane, B.; Stawski, P.; Schier, A.F.; et al. Optical control of metabotropic glutamate receptors. Nat. Neurosci. 2013, 16, 507–516. [Google Scholar] [CrossRef]
- Berlin, S.; Szobota, S.; Reiner, A.; Carroll, E.C.; Kienzler, M.A.; Guyon, A.; Xiao, T.; Trauner, D.; Isacoff, E.Y. A family of photoswitchable NMDA receptors. eLife 2016, 5, e12040. [Google Scholar] [CrossRef]
- Deal, W.J.; Erlanger, B.F.; Nachmansohn, D. Photoregulation of biological activity of photochromic reagents, III. Photoregulation of bioelectricity by acetylcholine receptor inhibitors. Proc. Natl. Acad. Sci. USA 1969, 64, 1230–1234. [Google Scholar] [CrossRef] [Green Version]
- Bartels, E.; Wassermann, N.H.; Erlanger, B.F. Photochromic activators of the acetylcholine receptor. Proc. Natl. Acad. Sci. USA 1971, 68, 1820–1823. [Google Scholar] [CrossRef] [Green Version]
- Tochitsky, I.; Banghart, M.R.; Mourot, A.; Yao, J.Z.; Gaub, B.; Kramer, R.H.; Trauner, D. Optochemical control of genetically engineered neuronal nicotinic acetylcholine receptors. Nat. Chem. 2012, 4, 105–111. [Google Scholar] [CrossRef]
- Magome, N.; Kanaporis, G.; Moisan, N.; Tanaka, K.; Agladze, K. Photo-control of excitation waves in cardiomyocyte tissue culture. Tissue Eng. Part A 2011, 17, 2703–2711. [Google Scholar] [CrossRef] [PubMed]
- Cao, E.; Liao, M.; Cheng, Y.; Julius, D. TRPV1 structures in distinct conformations reveal activation mechanisms. Nature 2013, 504, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Gavva, N.R.; Tamir, R.; Klionsky, L.; Norman, M.H.; Louis, J.C.; Wild, K.D.; Treanor, J.J. Proton activation does not alter antagonist interaction with the capsaicin-binding pocket of TRPV1. Mol. Pharm. 2005, 68, 1524–1533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cunha, M.R.; Bhardwaj, R.; Lindinger, S.; Butorac, C.; Romanin, C.; Hediger, M.A.; Reymond, J.L. Photoswitchable Inhibitor of the Calcium Channel TRPV6. ACS Med. Chem. Lett. 2019, 10, 1341–1345. [Google Scholar] [CrossRef]
- Frank, J.A.; Yushchenko, D.A.; Hodson, D.J.; Lipstein, N.; Nagpal, J.; Rutter, G.A.; Rhee, J.S.; Gottschalk, A.; Brose, N.; Schultz, C.; et al. Photoswitchable diacylglycerols enable optical control of protein kinase C. Nat. Chem. Biol. 2016, 12, 755–762. [Google Scholar] [CrossRef] [Green Version]
- Ashmole, I.; Duffy, S.M.; Leyland, M.L.; Morrison, V.S.; Begg, M.; Bradding, P. CRACM/Orai ion channel expression and function in human lung mast cells. J. Allergy Clin. Immunol. 2012, 129, 1628–1635.e1622. [Google Scholar] [CrossRef] [Green Version]
- Derler, I.; Schindl, R.; Fritsch, R.; Heftberger, P.; Riedl, M.C.; Begg, M.; House, D.; Romanin, C. The action of selective CRAC channel blockers is affected by the Orai pore geometry. Cell Calcium 2013, 53, 139–151. [Google Scholar] [CrossRef]
- van Kruchten, R.; Braun, A.; Feijge, M.A.; Kuijpers, M.J.; Rivera-Galdos, R.; Kraft, P.; Stoll, G.; Kleinschnitz, C.; Bevers, E.M.; Nieswandt, B.; et al. Antithrombotic potential of blockers of store-operated calcium channels in platelets. Arterioscler. Thromb. Vasc. Biol. 2012, 32, 1717–1723. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Xin, P.; Yoast, R.E.; Emrich, S.M.; Johnson, M.T.; Pathak, T.; Benson, J.C.; Azimi, I.; Gill, D.L.; Monteith, G.R.; et al. Distinct pharmacological profiles of ORAI1, ORAI2, and ORAI3 channels. Cell Calcium 2020, 91, 102281. [Google Scholar] [CrossRef]
- Waldherr, L.; Tiffner, A.; Mishra, D.; Sallinger, M.; Schober, R.; Frischauf, I.; Schmidt, T.; Handl, V.; Sagmeister, P.; Kockinger, M.; et al. Blockage of Store-Operated Ca(2+) Influx by Synta66 is Mediated by Direct Inhibition of the Ca(2+) Selective Orai1 Pore. Cancers 2020, 12, 2876. [Google Scholar] [CrossRef]
- Reiter, A.; Skerra, A.; Trauner, D.; Schiefner, A. A photoswitchable neurotransmitter analogue bound to its receptor. Biochemistry 2013, 52, 8972–8974. [Google Scholar] [CrossRef] [PubMed]
- Volgraf, M.; Gorostiza, P.; Numano, R.; Kramer, R.H.; Isacoff, E.Y.; Trauner, D. Allosteric control of an ionotropic glutamate receptor with an optical switch. Nat. Chem. Biol. 2006, 2, 47–52. [Google Scholar] [CrossRef]
- Szobota, S.; Gorostiza, P.; Del Bene, F.; Wyart, C.; Fortin, D.L.; Kolstad, K.D.; Tulyathan, O.; Volgraf, M.; Numano, R.; Aaron, H.L.; et al. Remote control of neuronal activity with a light-gated glutamate receptor. Neuron 2007, 54, 535–545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gorostiza, P.; Volgraf, M.; Numano, R.; Szobota, S.; Trauner, D.; Isacoff, E.Y. Mechanisms of photoswitch conjugation and light activation of an ionotropic glutamate receptor. Proc. Natl. Acad. Sci. USA 2007, 104, 10865–10870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Numano, R.; Szobota, S.; Lau, A.Y.; Gorostiza, P.; Volgraf, M.; Roux, B.; Trauner, D.; Isacoff, E.Y. Nanosculpting reversed wavelength sensitivity into a photoswitchable iGluR. Proc. Natl. Acad. Sci. USA 2009, 106, 6814–6819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kienzler, M.A.; Reiner, A.; Trautman, E.; Yoo, S.; Trauner, D.; Isacoff, E.Y. A red-shifted, fast-relaxing azobenzene photoswitch for visible light control of an ionotropic glutamate receptor. J. Am. Chem. Soc. 2013, 135, 17683–17686. [Google Scholar] [CrossRef] [Green Version]
- Rullo, A.; Reiner, A.; Reiter, A.; Trauner, D.; Isacoff, E.Y.; Woolley, G.A. Long wavelength optical control of glutamate receptor ion channels using a tetra-ortho-substituted azobenzene derivative. Chem. Commun. 2014, 50, 14613–14615. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Szobota, S.; Wang, Y.; Volgraf, M.; Liu, Z.; Sun, C.; Trauner, D.; Isacoff, E.Y.; Zhang, X. All optical interface for parallel, remote, and spatiotemporal control of neuronal activity. Nano. Lett. 2007, 7, 3859–3863. [Google Scholar] [CrossRef]
- Carroll, E.C.; Berlin, S.; Levitz, J.; Kienzler, M.A.; Yuan, Z.; Madsen, D.; Larsen, D.S.; Isacoff, E.Y. Two-photon brightness of azobenzene photoswitches designed for glutamate receptor optogenetics. Proc. Natl. Acad. Sci. USA 2015, 112, E776–E785. [Google Scholar] [CrossRef] [Green Version]
- Izquierdo-Serra, M.; Gascon-Moya, M.; Hirtz, J.J.; Pittolo, S.; Poskanzer, K.E.; Ferrer, E.; Alibes, R.; Busque, F.; Yuste, R.; Hernando, J.; et al. Two-photon neuronal and astrocytic stimulation with azobenzene-based photoswitches. J. Am. Chem. Soc. 2014, 136, 8693–8701. [Google Scholar] [CrossRef]
- Reiner, A.; Isacoff, E.Y. Tethered ligands reveal glutamate receptor desensitization depends on subunit occupancy. Nat. Chem. Biol. 2014, 10, 273–280. [Google Scholar] [CrossRef] [Green Version]
- Hartrampf, F.W.W.; Barber, D.M.; Gottschling, K.; Leippe, P.; Hollmann, M.; Trauner, D. Development of a photoswitchable antagonist of NMDA receptors. Tetrahedron 2017, 73, 4905–4912. [Google Scholar] [CrossRef]
- Damijonaitis, A.; Broichhagen, J.; Urushima, T.; Hull, K.; Nagpal, J.; Laprell, L.; Schonberger, M.; Woodmansee, D.H.; Rafiq, A.; Sumser, M.P.; et al. AzoCholine Enables Optical Control of Alpha 7 Nicotinic Acetylcholine Receptors in Neural Networks. ACS Chem. Neurosci. 2015, 6, 701–707. [Google Scholar] [CrossRef]
- Klippenstein, V.; Hoppmann, C.; Ye, S.; Wang, L.; Paoletti, P. Optocontrol of glutamate receptor activity by single side-chain photoisomerization. eLife 2017, 6, e25808. [Google Scholar] [CrossRef]
- Poulsen, M.H.; Poshtiban, A.; Klippenstein, V.; Ghisi, V.; Plested, A.J.R. Gating modules of the AMPA receptor pore domain revealed by unnatural amino acid mutagenesis. Proc. Natl. Acad. Sci. USA 2019, 116, 13358–13367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klippenstein, V.; Ghisi, V.; Wietstruk, M.; Plested, A.J. Photoinactivation of glutamate receptors by genetically encoded unnatural amino acids. J. Neurosci. Off. J. Soc. Neurosci. 2014, 34, 980–991. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, Y.; Yamamura, H.; Ohya, S.; Imaizumi, Y. Caveolin-1 facilitates the direct coupling between large conductance Ca2+-activated K+ (BKCa) and Cav1.2 Ca2+ channels and their clustering to regulate membrane excitability in vascular myocytes. J. Biol. Chem. 2013, 288, 36750–36761. [Google Scholar] [CrossRef] [Green Version]
- Kramer, R.H.; Mourot, A.; Adesnik, H. Optogenetic pharmacology for control of native neuronal signaling proteins. Nat. Neurosci. 2013, 16, 816–823. [Google Scholar] [CrossRef]
- Furuta, T. Designing Caged Compounds for Spatiotemporal Control of Cellular Chemistry. J. Synth. Org. Chem. Jpn. 2012, 70, 1164–1169. [Google Scholar] [CrossRef] [Green Version]
- Ellis-Davies, G.C. Development and application of caged calcium. Methods Enzymol. 2003, 360, 226–238. [Google Scholar] [CrossRef] [PubMed]
- Hull, K.; Morstein, J.; Trauner, D. In Vivo Photopharmacology. Chem. Rev. 2018, 118, 10710–10747. [Google Scholar] [CrossRef] [PubMed]
- Izquierdo-Serra, M.; Bautista-Barrufet, A.; Trapero, A.; Garrido-Charles, A.; Díaz-Tahoces, A.; Camarero, N.; Pittolo, S.; Valbuena, S.; Pérez-Jiménez, A.; Gay, M.; et al. Optical control of endogenous receptors and cellular excitability using targeted covalent photoswitches. Nat. Commun. 2016, 7, 12221. [Google Scholar] [CrossRef] [PubMed]
- Allen, B.D.; Singer, A.C.; Boyden, E.S. Principles of designing interpretable optogenetic behavior experiments. Learn Mem. 2015, 22, 232–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, F.; Vierock, J.; Yizhar, O.; Fenno, L.E.; Tsunoda, S.; Kianianmomeni, A.; Prigge, M.; Berndt, A.; Cushman, J.; Polle, J.; et al. The microbial opsin family of optogenetic tools. Cell 2011, 147, 1446–1457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deisseroth, K. Optogenetics: 10 years of microbial opsins in neuroscience. Nat. Neurosci. 2015, 18, 1213–1225. [Google Scholar] [CrossRef] [Green Version]
- Fenno, L.; Yizhar, O.; Deisseroth, K. The development and application of optogenetics. Annu. Rev. Neurosci. 2011, 34, 389–412. [Google Scholar] [CrossRef]
- Spangler, S.M.; Bruchas, M.R. Optogenetic approaches for dissecting neuromodulation and GPCR signaling in neural circuits. Curr. Opin. Pharmacol. 2017, 32, 56–70. [Google Scholar] [CrossRef] [Green Version]
- Airan, R.D.; Thompson, K.R.; Fenno, L.E.; Bernstein, H.; Deisseroth, K. Temporally precise in vivo control of intracellular signalling. Nature 2009, 458, 1025–1029. [Google Scholar] [CrossRef]
- Ernst, O.P.; Lodowski, D.T.; Elstner, M.; Hegemann, P.; Brown, L.S.; Kandori, H. Microbial and animal rhodopsins: Structures, functions, and molecular mechanisms. Chem. Rev. 2014, 114, 126–163. [Google Scholar] [CrossRef]
- Nagel, G.; Szellas, T.; Huhn, W.; Kateriya, S.; Adeishvili, N.; Berthold, P.; Ollig, D.; Hegemann, P.; Bamberg, E. Channelrhodopsin-2, a directly light-gated cation-selective membrane channel. Proc. Natl. Acad. Sci. USA 2003, 100, 13940–13945. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Hyun, Y.M.; Lim, K.; Lee, H.; Cummings, R.J.; Gerber, S.A.; Bae, S.; Cho, T.Y.; Lord, E.M.; Kim, M. Optogenetic control of chemokine receptor signal and T-cell migration. Proc. Natl Acad. Sci. USA 2014, 111, 6371–6376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fukuda, N.; Matsuda, T.; Nagai, T. Optical control of the Ca2+ concentration in a live specimen with a genetically encoded Ca2+-releasing molecular tool. ACS Chem. Biol. 2014, 9, 1197–1203. [Google Scholar] [CrossRef] [PubMed]
- Kim, N.; Kim, J.M.; Lee, M.; Kim, C.Y.; Chang, K.Y.; Heo, W.D. Spatiotemporal control of fibroblast growth factor receptor signals by blue light. Chem. Biol. 2014, 21, 903–912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, L.; Wang, L.; Zeng, H.; Tan, P.; Ma, G.; Zheng, S.; Li, Y.; Sun, L.; Dou, F.; Siwko, S.; et al. Engineering of a bona fide light-operated calcium channel. Nat. Commun. 2021, 12, 164. [Google Scholar] [CrossRef]
- van Wyk, M.; Pielecka-Fortuna, J.; Lowel, S.; Kleinlogel, S. Restoring the ON Switch in Blind Retinas: Opto-mGluR6, a Next-Generation, Cell-Tailored Optogenetic Tool. PLoS Biol. 2015, 13, e1002143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hughes, C.E.; Nibbs, R.J.B. A guide to chemokines and their receptors. FEBS J. 2018, 285, 2944–2971. [Google Scholar] [CrossRef]
- Lu, X.; Shen, Y.; Campbell, R.E. Engineering Photosensory Modules of Non-Opsin-Based Optogenetic Actuators. Int. J. Mol. Sci. 2020, 21, 6522. [Google Scholar] [CrossRef] [PubMed]
- Sancar, A. Structure and function of DNA photolyase and cryptochrome blue-light photoreceptors. Chem. Rev. 2003, 103, 2203–2237. [Google Scholar] [CrossRef] [PubMed]
- Cashmore, A.R.; Jarillo, J.A.; Wu, Y.J.; Liu, D. Cryptochromes: Blue light receptors for plants and animals. Science 1999, 284, 760–765. [Google Scholar] [CrossRef]
- Lin, C.; Shalitin, D. Cryptochrome structure and signal transduction. Annu. Rev. Plant Biol. 2003, 54, 469–496. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Liu, B.; Zhao, C.; Pepper, M.; Lin, C. The action mechanisms of plant cryptochromes. Trends Plant Sci. 2011, 16, 684–691. [Google Scholar] [CrossRef] [Green Version]
- Kennedy, M.J.; Hughes, R.M.; Peteya, L.A.; Schwartz, J.W.; Ehlers, M.D.; Tucker, C.L. Rapid blue-light-mediated induction of protein interactions in living cells. Nat. Methods 2010, 7, 973–975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Che, D.L.; Duan, L.; Zhang, K.; Cui, B. The Dual Characteristics of Light-Induced Cryptochrome 2, Homo-oligomerization and Heterodimerization, for Optogenetic Manipulation in Mammalian Cells. ACS Synth. Biol. 2015, 4, 1124–1135. [Google Scholar] [CrossRef] [Green Version]
- Zemelman, B.V.; Nesnas, N.; Lee, G.A.; Miesenböck, G. Photochemical gating of heterologous ion channels: Remote control over genetically designated populations of neurons. Proc. Natl. Acad. Sci. USA 2003, 100, 1352–1357. [Google Scholar] [CrossRef] [Green Version]
- Levskaya, A.; Weiner, O.D.; Lim, W.A.; Voigt, C.A. Spatiotemporal control of cell signalling using a light-switchable protein interaction. Nature 2009, 461, 997–1001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krauss, U.; Minh, B.Q.; Losi, A.; Gartner, W.; Eggert, T.; von Haeseler, A.; Jaeger, K.E. Distribution and phylogeny of light-oxygen-voltage-blue-light-signaling proteins in the three kingdoms of life. J. Bacteriol. 2009, 191, 7234–7242. [Google Scholar] [CrossRef] [Green Version]
- Harper, S.M.; Neil, L.C.; Gardner, K.H. Structural basis of a phototropin light switch. Science 2003, 301, 1541–1544. [Google Scholar] [CrossRef] [PubMed]
- Renicke, C.; Schuster, D.; Usherenko, S.; Essen, L.O.; Taxis, C. A LOV2 domain-based optogenetic tool to control protein degradation and cellular function. Chem. Biol. 2013, 20, 619–626. [Google Scholar] [CrossRef] [Green Version]
- Okajima, K.; Aihara, Y.; Takayama, Y.; Nakajima, M.; Kashojiya, S.; Hikima, T.; Oroguchi, T.; Kobayashi, A.; Sekiguchi, Y.; Yamamoto, M.; et al. Light-induced conformational changes of LOV1 (light oxygen voltage-sensing domain 1) and LOV2 relative to the kinase domain and regulation of kinase activity in Chlamydomonas phototropin. J. Biol. Chem. 2014, 289, 413–422. [Google Scholar] [CrossRef] [Green Version]
- Halavaty, A.S.; Moffat, K. N- and C-terminal flanking regions modulate light-induced signal transduction in the LOV2 domain of the blue light sensor phototropin 1 from Avena sativa. Biochemistry 2007, 46, 14001–14009. [Google Scholar] [CrossRef] [PubMed]
- Ma, G.; Wen, S.; Huang, Y.; Zhou, Y. The STIM-Orai Pathway: Light-Operated Ca(2+) Entry Through Engineered CRAC Channels. Adv. Exp. Med. Biol 2017, 993, 117–138. [Google Scholar] [CrossRef] [PubMed]
- Pudasaini, A.; El-Arab, K.K.; Zoltowski, B.D. LOV-based optogenetic devices: Light-driven modules to impart photoregulated control of cellular signaling. Front. Mol. Biosci. 2015, 2, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Motta-Mena, L.B.; Reade, A.; Mallory, M.J.; Glantz, S.; Weiner, O.D.; Lynch, K.W.; Gardner, K.H. An optogenetic gene expression system with rapid activation and deactivation kinetics. Nat. Chem. Biol. 2014, 10, 196–202. [Google Scholar] [CrossRef] [PubMed]
- Nihongaki, Y.; Suzuki, H.; Kawano, F.; Sato, M. Genetically engineered photoinducible homodimerization system with improved dimer-forming efficiency. ACS Chem. Biol. 2014, 9, 617–621. [Google Scholar] [CrossRef]
- Guntas, G.; Hallett, R.A.; Zimmerman, S.P.; Williams, T.; Yumerefendi, H.; Bear, J.E.; Kuhlman, B. Engineering an improved light-induced dimer (iLID) for controlling the localization and activity of signaling proteins. Proc. Natl. Acad. Sci. USA 2015, 112, 112–117. [Google Scholar] [CrossRef] [Green Version]
- Shcherbakova, D.M.; Shemetov, A.A.; Kaberniuk, A.A.; Verkhusha, V.V. Natural photoreceptors as a source of fluorescent proteins, biosensors, and optogenetic tools. Annu. Rev. Biochem. 2015, 84, 519–550. [Google Scholar] [CrossRef] [Green Version]
- Kim, N.; Kim, J.M.; Heo, W.D. Optogenetic Control of Fibroblast Growth Factor Receptor Signaling. Methods Mol. Biol. 2016, 1408, 345–362. [Google Scholar] [CrossRef]
- Béguin, P.; Nagashima, K.; Gonoi, T.; Shibasaki, T.; Takahashi, K.; Kashima, Y.; Ozaki, N.; Geering, K.; Iwanaga, T.; Seino, S. Regulation of Ca2+ channel expression at the cell surface by the small G-protein kir/Gem. Nature 2001, 411, 701–706. [Google Scholar] [CrossRef]
- Finlin, B.S.; Crump, S.M.; Satin, J.; Andres, D.A. Regulation of voltage-gated calcium channel activity by the Rem and Rad GTPases. Proc. Natl. Acad. Sci. USA 2003, 100, 14469–14474. [Google Scholar] [CrossRef] [Green Version]
- Colecraft, H.M. Designer genetically encoded voltage-dependent calcium channel inhibitors inspired by RGK GTPases. J. Physiol. 2020, 598, 1683–1693. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.N.; Berggren, P.O. Beta-cell CaV channel regulation in physiology and pathophysiology. Am. J. Physiology. Endocrinol. Metab. 2005, 288, E16–E28. [Google Scholar] [CrossRef]
- Park, C.Y.; Hoover, P.J.; Mullins, F.M.; Bachhawat, P.; Covington, E.D.; Raunser, S.; Walz, T.; Garcia, K.C.; Dolmetsch, R.E.; Lewis, R.S. STIM1 clusters and activates CRAC channels via direct binding of a cytosolic domain to Orai1. Cell 2009, 136, 876–890. [Google Scholar] [CrossRef] [Green Version]
- Yuan, J.P.; Zeng, W.; Dorwart, M.R.; Choi, Y.J.; Worley, P.F.; Muallem, S. SOAR and the polybasic STIM1 domains gate and regulate Orai channels. Nat. Cell Biol. 2009, 11, 337–343. [Google Scholar] [CrossRef]
- Korzeniowski, M.K.; Manjarres, I.M.; Varnai, P.; Balla, T. Activation of STIM1-Orai1 involves an intramolecular switching mechanism. Sci. Signal. 2010, 3, ra82. [Google Scholar] [CrossRef] [Green Version]
- Luik, R.M.; Wang, B.; Prakriya, M.; Wu, M.M.; Lewis, R.S. Oligomerization of STIM1 couples ER calcium depletion to CRAC channel activation. Nature 2008, 454, 538–542. [Google Scholar] [CrossRef] [PubMed]
- Fegan, A.; White, B.; Carlson, J.C.; Wagner, C.R. Chemically controlled protein assembly: Techniques and applications. Chem. Rev. 2010, 110, 3315–3336. [Google Scholar] [CrossRef]
- Ma, G.; He, L.; Liu, S.; Xie, J.; Huang, Z.; Jing, J.; Lee, Y.T.; Wang, R.; Luo, H.; Han, W.; et al. Optogenetic engineering to probe the molecular choreography of STIM1-mediated cell signaling. Nat. Commun. 2020, 11, 1039. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Srinivasan, P.; Razavi, S.; Seymour, S.; Meraner, P.; Gudlur, A.; Stathopulos, P.B.; Ikura, M.; Rao, A.; Hogan, P.G. Initial activation of STIM1, the regulator of store-operated calcium entry. Nat. Struct. Mol. Biol. 2013, 20, 973–981. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bugaj, L.J.; Choksi, A.T.; Mesuda, C.K.; Kane, R.S.; Schaffer, D.V. Optogenetic protein clustering and signaling activation in mammalian cells. Nat. Methods 2013, 10, 249–252. [Google Scholar] [CrossRef]
- Taslimi, A.; Vrana, J.D.; Chen, D.; Borinskaya, S.; Mayer, B.J.; Kennedy, M.J.; Tucker, C.L. An optimized optogenetic clustering tool for probing protein interaction and function. Nat. Commun. 2014, 5, 4925. [Google Scholar] [CrossRef]
- He, L.; Zhang, Q.; Zhou, Y.; Huang, Y. Optogenetic Approaches to Control Calcium Entry in Non-Excitable Cells. In Calcium Entry Channels in Non-Excitable Cells; Kozak, J.A., Putney, J.W., Jr., Eds.; CRC Press: Boca Raton, FL, USA, 2018; pp. 145–160. [Google Scholar]
- Ma, G.; Wen, S.; He, L.; Huang, Y.; Wang, Y.; Zhou, Y. Optogenetic toolkit for precise control of calcium signaling. Cell Calcium 2017, 64, 36–46. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Kyung, T.; Chung, J.H.; Kim, N.; Keum, S.; Lee, J.; Park, H.; Kim, H.M.; Lee, S.; Shin, H.S.; et al. Non-invasive optical control of endogenous Ca(2+) channels in awake mice. Nat. Commun. 2020, 11, 210. [Google Scholar] [CrossRef] [Green Version]
- Bohineust, A.; Garcia, Z.; Corre, B.; Lemaitre, F.; Bousso, P. Optogenetic manipulation of calcium signals in single T cells in vivo. Nat. Commun. 2020, 11, 1143. [Google Scholar] [CrossRef] [PubMed]
- Park, H.; Kim, N.Y.; Lee, S.; Kim, N.; Kim, J.; Heo, W.D. Optogenetic protein clustering through fluorescent protein tagging and extension of CRY2. Nat. Commun. 2017, 8, 30. [Google Scholar] [CrossRef] [PubMed]
- Fahrner, M.; Muik, M.; Schindl, R.; Butorac, C.; Stathopulos, P.; Zheng, L.; Jardin, I.; Ikura, M.; Romanin, C. A coiled-coil clamp controls both conformation and clustering of stromal interaction molecule 1 (STIM1). J. Biol. Chem. 2014, 289, 33231–33244. [Google Scholar] [CrossRef] [Green Version]
- Ma, G.; Wei, M.; He, L.; Liu, C.; Wu, B.; Zhang, S.L.; Jing, J.; Liang, X.; Senes, A.; Tan, P.; et al. Inside-out Ca(2+) signalling prompted by STIM1 conformational switch. Nat. Commun. 2015, 6, 7826. [Google Scholar] [CrossRef] [Green Version]
- He, L.; Jing, J.; Zhu, L.; Tan, P.; Ma, G.; Zhang, Q.; Nguyen, N.T.; Wang, J.; Zhou, Y.; Huang, Y. Optical control of membrane tethering and interorganellar communication at nanoscales. Chem. Sci. 2017, 8, 5275–5281. [Google Scholar] [CrossRef] [Green Version]
- Krizova, A.; Maltan, L.; Derler, I. Critical parameters maintaining authentic CRAC channel hallmarks. Eur. Biophys. J. 2019, 48, 425–445. [Google Scholar] [CrossRef] [Green Version]
- Fahrner, M.; Pandey, S.K.; Muik, M.; Traxler, L.; Butorac, C.; Stadlbauer, M.; Zayats, V.; Krizova, A.; Plenk, P.; Frischauf, I.; et al. Communication between N terminus and loop2 tunes Orai activation. J. Biol. Chem. 2018, 293, 1271–1285. [Google Scholar] [CrossRef] [Green Version]
- Butorac, C.; Muik, M.; Derler, I.; Stadlbauer, M.; Lunz, V.; Krizova, A.; Lindinger, S.; Schober, R.; Frischauf, I.; Bhardwaj, R.; et al. A novel STIM1-Orai1 gating interface essential for CRAC channel activation. Cell Calcium 2019, 79, 57–67. [Google Scholar] [CrossRef] [PubMed]
- Srikanth, S.; Jung, H.J.; Ribalet, B.; Gwack, Y. The intracellular loop of Orai1 plays a central role in fast inactivation of Ca2+ release-activated Ca2+ channels. J. Biol. Chem. 2010, 285, 5066–5075. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yizhar, O.; Fenno, L.E.; Prigge, M.; Schneider, F.; Davidson, T.J.; O’Shea, D.J.; Sohal, V.S.; Goshen, I.; Finkelstein, J.; Paz, J.T.; et al. Neocortical excitation/inhibition balance in information processing and social dysfunction. Nature 2011, 477, 171–178. [Google Scholar] [CrossRef]
- Goto, Y.; Kondo, Y.; Aoki, K. Visualization and Manipulation of Intracellular Signaling. Adv. Exp. Med. Biol. 2021, 1293, 225–234. [Google Scholar] [CrossRef] [PubMed]
- Boyle, P.M.; Karathanos, T.V.; Trayanova, N.A. “Beauty is a light in the heart”: The transformative potential of optogenetics for clinical applications in cardiovascular medicine. Trends Cardiovasc. Med. 2015, 25, 73–81. [Google Scholar] [CrossRef] [Green Version]
- Housley, G.D.; Raybould, N.P.; Thorne, P.R. Fluorescence imaging of Na+ influx via P2X receptors in cochlear hair cells. Hear. Res. 1998, 119, 1–13. [Google Scholar] [CrossRef]
- Kotter, R.; Schubert, D.; Dyhrfjeld-Johnsen, J.; Luhmann, H.J.; Staiger, J.F. Optical release of caged glutamate for stimulation of neurons in the in vitro slice preparation. J. Biomed. Opt. 2005, 10, 11003. [Google Scholar] [CrossRef] [PubMed]
- Rial Verde, E.M.; Zayat, L.; Etchenique, R.; Yuste, R. Photorelease of GABA with Visible Light Using an Inorganic Caging Group. Front. Neural Circuits 2008, 2, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, F.; Smith, N.A.; Xu, Q.; Goldman, S.; Peng, W.; Huang, J.H.; Takano, T.; Nedergaard, M. Photolysis of caged Ca2+ but not receptor-mediated Ca2+ signaling triggers astrocytic glutamate release. J. Neurosci. Off. J. Soc. Neurosci. 2013, 33, 17404–17412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adams, S.R.; Lev-Ram, V.; Tsien, R.Y. A new caged Ca2+, azid-1, is far more photosensitive than nitrobenzyl-based chelators. Chem. Biol. 1997, 4, 867–878. [Google Scholar] [CrossRef] [Green Version]
- Gurney, A.M.; Tsien, R.Y.; Lester, H.A. Activation of a potassium current by rapid photochemically generated step increases of intracellular calcium in rat sympathetic neurons. Proc. Natl. Acad. Sci. USA 1987, 84, 3496–3500. [Google Scholar] [CrossRef] [Green Version]
- Noguchi, J.; Matsuzaki, M.; Ellis-Davies, G.C.; Kasai, H. Spine-neck geometry determines NMDA receptor-dependent Ca2+ signaling in dendrites. Neuron 2005, 46, 609–622. [Google Scholar] [CrossRef] [Green Version]
- Branco, T.; Clark, B.A.; Hausser, M. Dendritic discrimination of temporal input sequences in cortical neurons. Science 2010, 329, 1671–1675. [Google Scholar] [CrossRef]
- Denk, W. Two-photon scanning photochemical microscopy: Mapping ligand-gated ion channel distributions. Proc. Natl. Acad. Sci. USA 1994, 91, 6629–6633. [Google Scholar] [CrossRef] [Green Version]
- Huang, X.P.; Sreekumar, R.; Patel, J.R.; Walker, J.W. Response of cardiac myocytes to a ramp increase of diacylglycerol generated by photolysis of a novel caged diacylglycerol. Biophys. J. 1996, 70, 2448–2457. [Google Scholar] [CrossRef] [Green Version]
- McDonald, T.V.; Premack, B.A.; Gardner, P. Flash photolysis of caged inositol 1,4,5-trisphosphate activates plasma membrane calcium current in human T cells. J. Biol. Chem. 1993, 268, 3889–3896. [Google Scholar] [CrossRef]
- Tomida, T.; Hirose, K.; Takizawa, A.; Shibasaki, F.; Iino, M. NFAT functions as a working memory of Ca2+ signals in decoding Ca2+ oscillation. EMBO J. 2003, 22, 3825–3832. [Google Scholar] [CrossRef] [Green Version]
- Quann, E.J.; Merino, E.; Furuta, T.; Huse, M. Localized diacylglycerol drives the polarization of the microtubule-organizing center in T cells. Nat. Immunol. 2009, 10, 627–635. [Google Scholar] [CrossRef] [PubMed]
- Tiapko, O.; Shrestha, N.; Lindinger, S.; Guedes de la Cruz, G.; Graziani, A.; Klec, C.; Butorac, C.; Graier, W.F.; Kubista, H.; Freichel, M.; et al. Lipid-independent control of endothelial and neuronal TRPC3 channels by light. Chem. Sci. 2019, 10, 2837–2842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Izquierdo-Serra, M.; Trauner, D.; Llobet, A.; Gorostiza, P. Optical control of calcium-regulated exocytosis. Biochim. Biophys.Acta 2013, 1830, 2853–2860. [Google Scholar] [CrossRef] [PubMed]
- Kauwe, G.; Isacoff, E.Y. Rapid feedback regulation of synaptic efficacy during high-frequency activity at the Drosophila larval neuromuscular junction. Proc. Natl. Acad. Sci. USA 2013, 110, 9142–9147. [Google Scholar] [CrossRef] [Green Version]
- Adesnik, H.; Nicoll, R.A.; England, P.M. Photoinactivation of native AMPA receptors reveals their real-time trafficking. Neuron 2005, 48, 977–985. [Google Scholar] [CrossRef] [Green Version]
- Patterson, M.A.; Szatmari, E.M.; Yasuda, R. AMPA receptors are exocytosed in stimulated spines and adjacent dendrites in a Ras-ERK-dependent manner during long-term potentiation. Proc. Natl. Acad. Sci. USA 2010, 107, 15951–15956. [Google Scholar] [CrossRef] [Green Version]
- Khiroug, L.; Giniatullin, R.; Klein, R.C.; Fayuk, D.; Yakel, J.L. Functional mapping and Ca2+ regulation of nicotinic acetylcholine receptor channels in rat hippocampal CA1 neurons. J. Neurosci. Off. J. Soc. Neurosci. 2003, 23, 9024–9031. [Google Scholar] [CrossRef] [Green Version]
- Pittolo, S.; Gomez-Santacana, X.; Eckelt, K.; Rovira, X.; Dalton, J.; Goudet, C.; Pin, J.P.; Llobet, A.; Giraldo, J.; Llebaria, A.; et al. An allosteric modulator to control endogenous G protein-coupled receptors with light. Nat. Chem. Biol. 2014, 10, 813–815. [Google Scholar] [CrossRef]
- Rovira, X.; Trapero, A.; Pittolo, S.; Zussy, C.; Faucherre, A.; Jopling, C.; Giraldo, J.; Pin, J.P.; Gorostiza, P.; Goudet, C.; et al. OptoGluNAM4.1, a Photoswitchable Allosteric Antagonist for Real-Time Control of mGlu4 Receptor Activity. Cell Chem. Biol. 2016, 23, 929–934. [Google Scholar] [CrossRef] [Green Version]
- Gomez-Santacana, X.; Pittolo, S.; Rovira, X.; Lopez, M.; Zussy, C.; Dalton, J.A.; Faucherre, A.; Jopling, C.; Pin, J.P.; Ciruela, F.; et al. Illuminating Phenylazopyridines To Photoswitch Metabotropic Glutamate Receptors: From the Flask to the Animals. ACS Cent. Sci. 2017, 3, 81–91. [Google Scholar] [CrossRef]
- Wyart, C.; Del Bene, F.; Warp, E.; Scott, E.K.; Trauner, D.; Baier, H.; Isacoff, E.Y. Optogenetic dissection of a behavioural module in the vertebrate spinal cord. Nature 2009, 461, 407–410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janovjak, H.; Szobota, S.; Wyart, C.; Trauner, D.; Isacoff, E.Y. A light-gated, potassium-selective glutamate receptor for the optical inhibition of neuronal firing. Nat. Neurosci. 2010, 13, 1027–1032. [Google Scholar] [CrossRef]
- Zussy, C.; Gomez-Santacana, X.; Rovira, X.; De Bundel, D.; Ferrazzo, S.; Bosch, D.; Asede, D.; Malhaire, F.; Acher, F.; Giraldo, J.; et al. Dynamic modulation of inflammatory pain-related affective and sensory symptoms by optical control of amygdala metabotropic glutamate receptor 4. Mol. Psychiatry 2018, 23, 509–520. [Google Scholar] [CrossRef] [PubMed]
- Font, J.; Lopez-Cano, M.; Notartomaso, S.; Scarselli, P.; Di Pietro, P.; Bresoli-Obach, R.; Battaglia, G.; Malhaire, F.; Rovira, X.; Catena, J.; et al. Optical control of pain in vivo with a photoactive mGlu5 receptor negative allosteric modulator. eLife 2017, 6, e23545. [Google Scholar] [CrossRef] [Green Version]
- Durand-de Cuttoli, R.; Mondoloni, S.; Marti, F.; Lemoine, D.; Nguyen, C.; Naude, J.; d’Izarny-Gargas, T.; Pons, S.; Maskos, U.; Trauner, D.; et al. Manipulating midbrain dopamine neurons and reward-related behaviors with light-controllable nicotinic acetylcholine receptors. eLife 2018, 7, e37487. [Google Scholar] [CrossRef]
- Caporale, N.; Kolstad, K.D.; Lee, T.; Tochitsky, I.; Dalkara, D.; Trauner, D.; Kramer, R.; Dan, Y.; Isacoff, E.Y.; Flannery, J.G. LiGluR restores visual responses in rodent models of inherited blindness. Mol. Ther. J. Am. Soc. Gene Ther. 2011, 19, 1212–1219. [Google Scholar] [CrossRef] [PubMed]
- Gaub, B.M.; Berry, M.H.; Holt, A.E.; Reiner, A.; Kienzler, M.A.; Dolgova, N.; Nikonov, S.; Aguirre, G.D.; Beltran, W.A.; Flannery, J.G.; et al. Restoration of visual function by expression of a light-gated mammalian ion channel in retinal ganglion cells or ON-bipolar cells. Proc. Natl. Acad. Sci. USA 2014, 111, E5574–E5583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berry, M.H.; Holt, A.; Levitz, J.; Broichhagen, J.; Gaub, B.M.; Visel, M.; Stanley, C.; Aghi, K.; Kim, Y.J.; Cao, K.; et al. Restoration of patterned vision with an engineered photoactivatable G protein-coupled receptor. Nat. Commun. 2017, 8, 1862. [Google Scholar] [CrossRef] [PubMed]
- Tochitsky, I.; Kienzler, M.A.; Isacoff, E.; Kramer, R.H. Restoring Vision to the Blind with Chemical Photoswitches. Chem. Rev. 2018, 118, 10748–10773. [Google Scholar] [CrossRef]
- Deisseroth, K. Optogenetics. Nat. Methods 2011, 8, 26–29. [Google Scholar] [CrossRef]
- Gradinaru, V.; Mogri, M.; Thompson, K.R.; Henderson, J.M.; Deisseroth, K. Optical deconstruction of parkinsonian neural circuitry. Science 2009, 324, 354–359. [Google Scholar] [CrossRef] [Green Version]
- Reinbothe, T.M.; Safi, F.; Axelsson, A.S.; Mollet, I.G.; Rosengren, A.H. Optogenetic control of insulin secretion in intact pancreatic islets with beta-cell-specific expression of Channelrhodopsin-2. Islets 2014, 6, e28095. [Google Scholar] [CrossRef] [Green Version]
- Deisseroth, K.; Feng, G.; Majewska, A.K.; Miesenbock, G.; Ting, A.; Schnitzer, M.J. Next-generation optical technologies for illuminating genetically targeted brain circuits. J. Neurosci. Off. J. Soc. Neurosci. 2006, 26, 10380–10386. [Google Scholar] [CrossRef] [Green Version]
- Aravanis, A.M.; Wang, L.P.; Zhang, F.; Meltzer, L.A.; Mogri, M.Z.; Schneider, M.B.; Deisseroth, K. An optical neural interface: In vivo control of rodent motor cortex with integrated fiberoptic and optogenetic technology. J. Neural Eng. 2007, 4, S143–S156. [Google Scholar] [CrossRef] [PubMed]
- Ye, H.; Daoud-El Baba, M.; Peng, R.W.; Fussenegger, M. A synthetic optogenetic transcription device enhances blood-glucose homeostasis in mice. Science 2011, 332, 1565–1568. [Google Scholar] [CrossRef] [PubMed]
- Holz, G.G.t.; Kuhtreiber, W.M.; Habener, J.F. Pancreatic beta-cells are rendered glucose-competent by the insulinotropic hormone glucagon-like peptide-1(7-37). Nature 1993, 361, 362–365. [Google Scholar] [CrossRef] [Green Version]
- Parsons, G.B.; Souza, D.W.; Wu, H.; Yu, D.; Wadsworth, S.G.; Gregory, R.J.; Armentano, D. Ectopic expression of glucagon-like peptide 1 for gene therapy of type II diabetes. Gene Ther. 2007, 14, 38–48. [Google Scholar] [CrossRef] [Green Version]
- Hannanta-Anan, P.; Chow, B.Y. Optogenetic Control of Calcium Oscillation Waveform Defines NFAT as an Integrator of Calcium Load. Cell Syst. 2016, 2, 283–288. [Google Scholar] [CrossRef] [Green Version]
- Beiert, T.; Bruegmann, T.; Sasse, P. Optogenetic activation of Gq signalling modulates pacemaker activity of cardiomyocytes. Cardiovasc. Res. 2014, 102, 507–516. [Google Scholar] [CrossRef] [Green Version]
- Viard, P.; Butcher, A.J.; Halet, G.; Davies, A.; Nurnberg, B.; Heblich, F.; Dolphin, A.C. PI3K promotes voltage-dependent calcium channel trafficking to the plasma membrane. Nat. Neurosci. 2004, 7, 939–946. [Google Scholar] [CrossRef]
- Kim, J.H.; Lkhagvadorj, S.; Lee, M.R.; Hwang, K.H.; Chung, H.C.; Jung, J.H.; Cha, S.K.; Eom, M. Orai1 and STIM1 are critical for cell migration and proliferation of clear cell renal cell carcinoma. Biochem. Biophys. Res. Commun. 2014, 448, 76–82. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, L.A.; Larson, M.H.; Morsut, L.; Liu, Z.; Brar, G.A.; Torres, S.E.; Stern-Ginossar, N.; Brandman, O.; Whitehead, E.H.; Doudna, J.A.; et al. CRISPR-mediated modular RNA-guided regulation of transcription in eukaryotes. Cell 2013, 154, 442–451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ran, F.A.; Hsu, P.D.; Wright, J.; Agarwala, V.; Scott, D.A.; Zhang, F. Genome engineering using the CRISPR-Cas9 system. Nat. Protoc. 2013, 8, 2281–2308. [Google Scholar] [CrossRef] [Green Version]
- Hsu, P.D.; Lander, E.S.; Zhang, F. Development and applications of CRISPR-Cas9 for genome engineering. Cell 2014, 157, 1262–1278. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, N.T.; He, L.; Martinez-Moczygemba, M.; Huang, Y.; Zhou, Y. Rewiring Calcium Signaling for Precise Transcriptional Reprogramming. ACS Synth. Biol. 2018, 7, 814–821. [Google Scholar] [CrossRef]
- Sato, M.; Asano, T.; Hosomichi, J.; Ono, T.; Nakata, T. Optogenetic manipulation of intracellular calcium by BACCS promotes differentiation of MC3T3-E1 cells. Biochem. Biophys. Res. Commun. 2018, 506, 716–722. [Google Scholar] [CrossRef]
- Sun, L.; Blair, H.C.; Peng, Y.; Zaidi, N.; Adebanjo, O.A.; Wu, X.B.; Wu, X.Y.; Iqbal, J.; Epstein, S.; Abe, E.; et al. Calcineurin regulates bone formation by the osteoblast. Proc. Natl. Acad. Sci. USA 2005, 102, 17130–17135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, S.; Liu, Y.; Lipsky, S.; Cho, M. Physical manipulation of calcium oscillations facilitates osteodifferentiation of human mesenchymal stem cells. FASEB J. 2007, 21, 1472–1480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winslow, M.M.; Pan, M.; Starbuck, M.; Gallo, E.M.; Deng, L.; Karsenty, G.; Crabtree, G.R. Calcineurin/NFAT signaling in osteoblasts regulates bone mass. Dev. Cell 2006, 10, 771–782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zayzafoon, M. Calcium/calmodulin signaling controls osteoblast growth and differentiation. J. Cell. Biochem. 2006, 97, 56–70. [Google Scholar] [CrossRef]
- Ramamoorth, M.; Narvekar, A. Non viral vectors in gene therapy- an overview. J. Clin. Diagn. Res. JCDR 2015, 9, GE01–GE06. [Google Scholar] [CrossRef]
- Joshi, J.; Rubart, M.; Zhu, W. Optogenetics: Background, Methodological Advances and Potential Applications for Cardiovascular Research and Medicine. Front. Bioeng. Biotechnol. 2020, 7, 466. [Google Scholar] [CrossRef] [Green Version]
- Yizhar, O.; Fenno, L.E.; Davidson, T.J.; Mogri, M.; Deisseroth, K. Optogenetics in neural systems. Neuron 2011, 71, 9–34. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Lu, L.; Ye, S. Genetic Code Expansion and Optoproteomics. Yale J. Biol. Med. 2017, 90, 599–610. [Google Scholar] [PubMed]
- Chin, J.W. Expanding and reprogramming the genetic code. Nature 2017, 550, 53–60. [Google Scholar] [CrossRef] [PubMed]





| Cages | Photosensitive Tool | Chemical Structure | Cell Line/ Organism | Ref. |
|---|---|---|---|---|
| Ca2+ | Diazo-2 |  | fibroblasts | [114] |
| Diazo-4 |  | rat hippocampal pyramidal cells Aplysia neurons | [118,119] | |
| Nitr chelators |  | Cell bodies of cultured neurons of the fresh water snail Helisoma trivolvis | [125] | |
| DM-nitrophen |  | skinned rabbit psoas fibers | [126,127] | |
| ATP | O-caged ATP-[γS] |  | Hippocampal CA1 Pyramidal Neurons | [128] |
| glutamate | gluEPSPs |  | Hippocampal CA1 Pyramidal Neurons | [129] |
| IP3 |  | frog skeletal muscle | [130] |
| Photosensitive Tool | Chemical Structure | Target | wavelength | Mechanism | Cell Line/ Organism | Ref. |
|---|---|---|---|---|---|---|
| QAQ |  | blocks voltage-gated K+, Na+ and Ca2+channels of TRPV1-expressing neurons | 380 nm/ 500 nm light | light-dependent blocker of the Nav and Kv channel in its trans configuration and acts only at the inner side, but not at the outer side of the ion channel. | Rat hippocampal neuron, HEK-293T cells, mouse DRG neurons, spinal cord slices | [149] |
| AzoTAB |  | blocks voltage-gated Ca2+ ion channels | 365 nm/ 490 nm | Its trans-isomer blocks NaV and CaV channels, resulting in the suppression of the spontaneous electric excitability in the cardiomyocytes. | Neonatal Rat Cardiomyocytes | [152] |
| AC-4 |  | TRPV1 | 360 nm/ 440 nm | AC-4 is able to inhibit the TRPV1 channel. It acts as a trans-antagonist on the voltage-activated channel and as a cis-antagonist in experiments with the agonist capsaicin (TRPV1 agonist) | dorsal root ganglion neurons and C-fibre nociceptors, HEK293T cell | [153] |
| ABDTC |  | TRPV1 | 370 nm/ 470 nm | Cis-antagonist on voltage-activated TRPV1 | HEK293T cells | [154] |
| optovin |  | TRPA1 | 405 nm/ dark | Control TRPA1 activity by a light-activated intermediate state that reversibly forms covalent thioether with redox-sensitive key cysteines in the channel. | HEK293T cells, dorsal root ganglion sensory neurons, zebrafish, TRPA1-KO mice, DRG neurons, human cardiomyocytes | [155,156] |
| PhoDAGs |  | TRPC2/3/6 | 365 nm/ 470 nm | The photoswitchable diacylglycerols have photoswitchable fatty acids that enable the optical manipulation of TRPC2/3/6 activity. | HEK293 cells, mammalian tissue slices, mouse vomeronasal sensory neurons (VSNs), murine vomeronasal organ tissue slices | [157] |
| OptoDArG |  | TRPC3 | 365 nm/ 430 nm | Photochromic ligand for TRPC3 channel that assists the activation of TRPC3 mutant, containing the G652A substitution near the selectivity filter. | HEK293 cells | [158] |
| piCRAC-1 |  | CRAC channel | 365 nm/ 415 nm | The amide group of the CRAC channel inhibitors (e.g: GSK, Synta 66) is substituted by azo-group to convert it into photoswitchable derivatives. | HEK293 cells, zebrafish embryos | [159] |
| ATA-3 |  | AMPA receptors | 440 nm-480 nm/ dark | Its trans-isomer binds to the ligand-binding domains within AMPA receptors and allows for full blockage of the receptor, whereas the cis-isomer breaks up quickly. | HEK293T cells, mouse cortical slices, hippocampal neurons, TKO mouse retina | [160,161] |
| ShuBQX-3 |  | AMPA receptors | 460 nm/ 600 nm | ShuBQX-3 is supposed to attach to the glutamate receptor ligand-binding domain | HEK293T cells, Xenopus oocytes, hippocampal neurons | [162] |
| gluAzo |  | Kainate receptor | 380 nm/ 500 nm | Because the binding pocket of the glutamate moiety of GluAzo photoswitch is analogous to that of soluble glutamate, it can activate and inhibit the kainate receptors reversibly. | rat hippocampal neurons, HEK293 cells, purkinje cells | [163,164,165] |
| LiGluR |  | Kainate receptor | 460 nm or 840 nm (two-photon)/ dark | It can induce the pore opening of the kainate receptor variants in an allosteric manner because it is located in close proximity to the ligand-binding domain | Bovine chromaffin cells, astrocytes, visual cortex of awake mice, zebrafish, hippocampal neurons, HEK293T cell | [166,167,168] |
| LimGluRs |  | Kainate receptor | 380 nm or 820 (two-photon)/ 500 nm | It can induce the pore opening of the kainate receptor variants in an allosteric manner because it is localized in close proximity to the ligand-binding domain | HEK293T cell, shippocampal neurons, zebrafish larvae, astrocytes, chromaffin cells, TKO mice | [169] |
| LiGluN |  | Activates or inhibit NMDA receptors (GluN1, GluN2A, GluN2B) | 360 nm-405 nm (two-photon)/ 460 nm-560 nm | It can trigger the pore opening of the kainate receptor variants in an allosteric manner because it is localized in close proximity to the ligand-binding domain | HEK293T cells, hippocampal slices, larval zebrafish, GluN2A-knockout neonate mice | [170] |
| AzoCharCh |  | nAChRs | UV | AzoCharCh is a derivative of the carbamylcholine (potent depolarizing agent) that can reversibly inhibit acetylcholinesterase activity and function at low concentrations as receptor inhibitors block the depolarization generated by CharCh. | Electrophorous Electroplaques | [171] |
| Azo-PTA |  | nAChRs | UV | Azo-PTA is a derivative of the phenyltrimethylammonium (potent depolarizing agent) that can reversibly inhibit acetylcholinesterase activity and function at low concentrations as receptor inhibitors block the depolarization generated by CharCh. | Electrophorous Electroplaques | [171] |
| bisQ |  | nAChRs | UV | Its trans-isomer is a potent activator of Electrophorus electroplaques, in which it can depolarize the electrogenic membrane of the electroplaque. It functions as inhibitor of acetylcholinesterase. | Electrophorous Electroplaques | [172] |
| QBr |  | nAChRs | UV | trans-QBr can covalently bind to the electroplaques membrane and depolarize the membrane. It functions as an inhibitor of acetylcholinesterase. | Electrophorous Electroplaques | [172] |
| MAACh |  | nAChRs | 380 nm/ 500 nm or dark | The MAACh is used as a photoswitchable agonist for the nAChR receptor. | Xenopus oocytes | [173] |
| MAHoCh |  | nAChRs | 380 nm/ 500 nm or dark | The MAHoCh is a light-activated antagonist for the nAChR receptor. | Xenopus oocytes | [173] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maltan, L.; Najjar, H.; Tiffner, A.; Derler, I. Deciphering Molecular Mechanisms and Intervening in Physiological and Pathophysiological Processes of Ca2+ Signaling Mechanisms Using Optogenetic Tools. Cells 2021, 10, 3340. https://doi.org/10.3390/cells10123340
Maltan L, Najjar H, Tiffner A, Derler I. Deciphering Molecular Mechanisms and Intervening in Physiological and Pathophysiological Processes of Ca2+ Signaling Mechanisms Using Optogenetic Tools. Cells. 2021; 10(12):3340. https://doi.org/10.3390/cells10123340
Chicago/Turabian StyleMaltan, Lena, Hadil Najjar, Adéla Tiffner, and Isabella Derler. 2021. "Deciphering Molecular Mechanisms and Intervening in Physiological and Pathophysiological Processes of Ca2+ Signaling Mechanisms Using Optogenetic Tools" Cells 10, no. 12: 3340. https://doi.org/10.3390/cells10123340