
Working Memory Maintenance of Visual and Auditory Spatial Information Relies on Supramodal Neural Codes in the Dorsal Frontoparietal Cortex

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
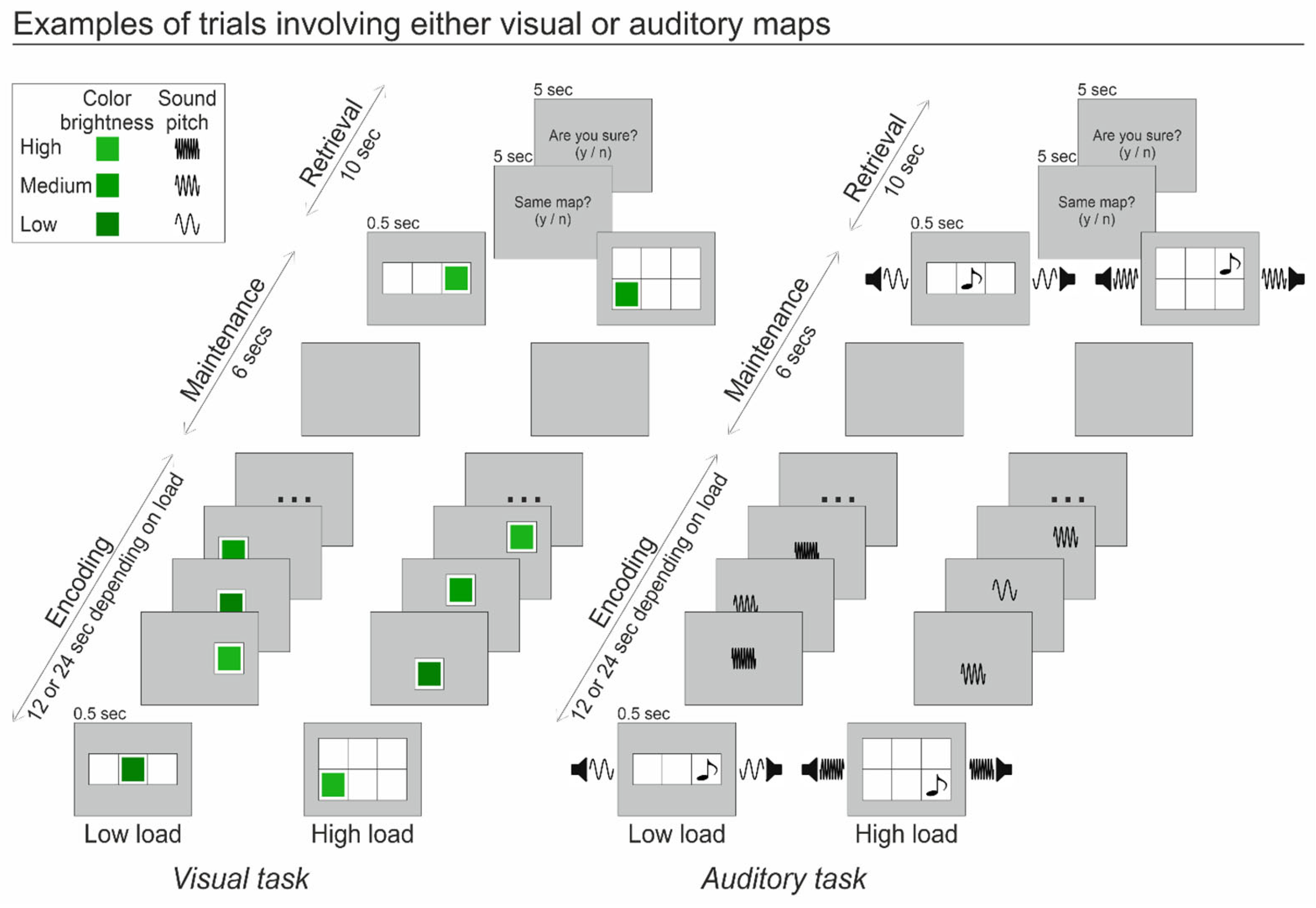
2.2. Stimuli and Procedure
2.3. Magnetic Resonance Imaging
2.4. fMRI Data Analysis
2.5. Multi-Voxel Pattern Analysis (MVPA)
2.6. Statistical Approach
3. Results
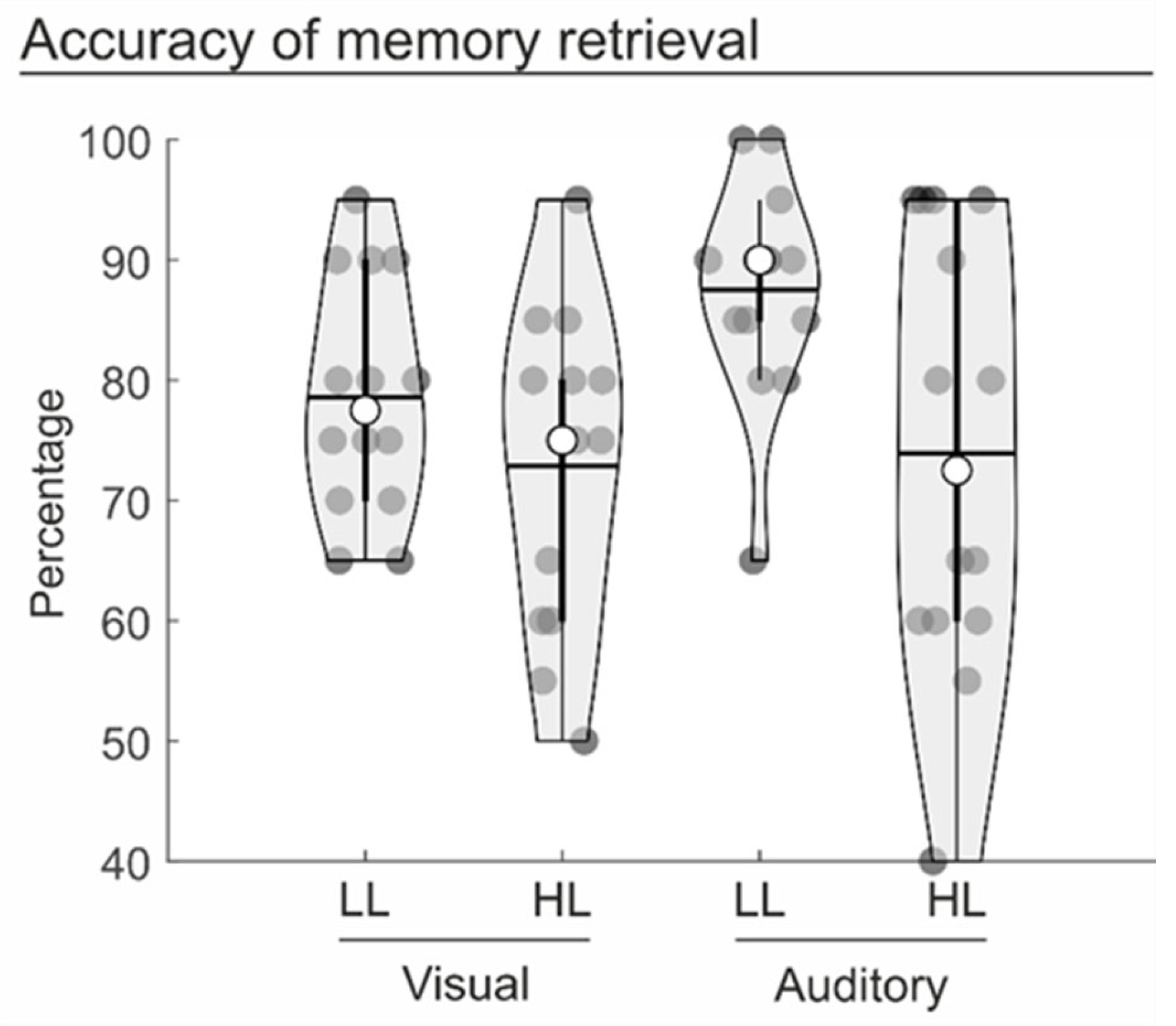
3.1. Behavioural Data
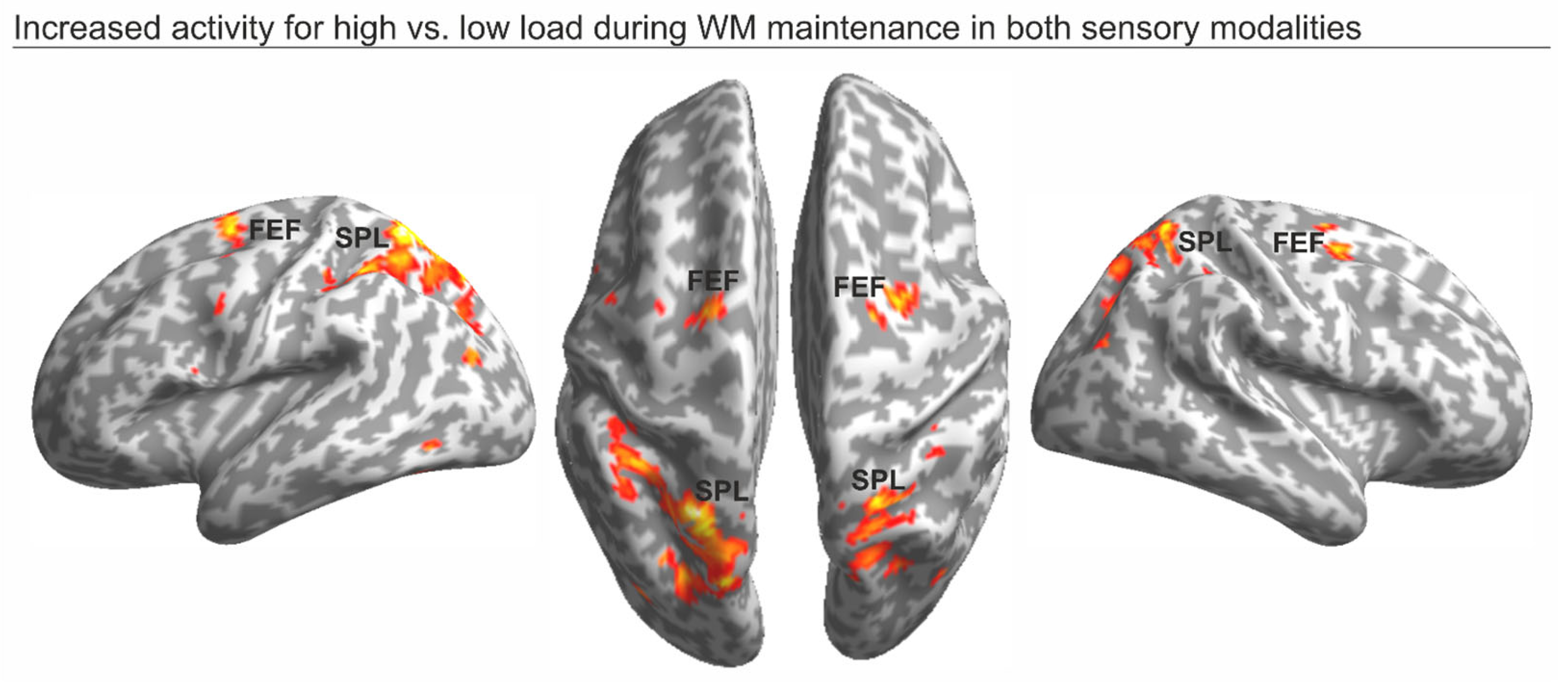
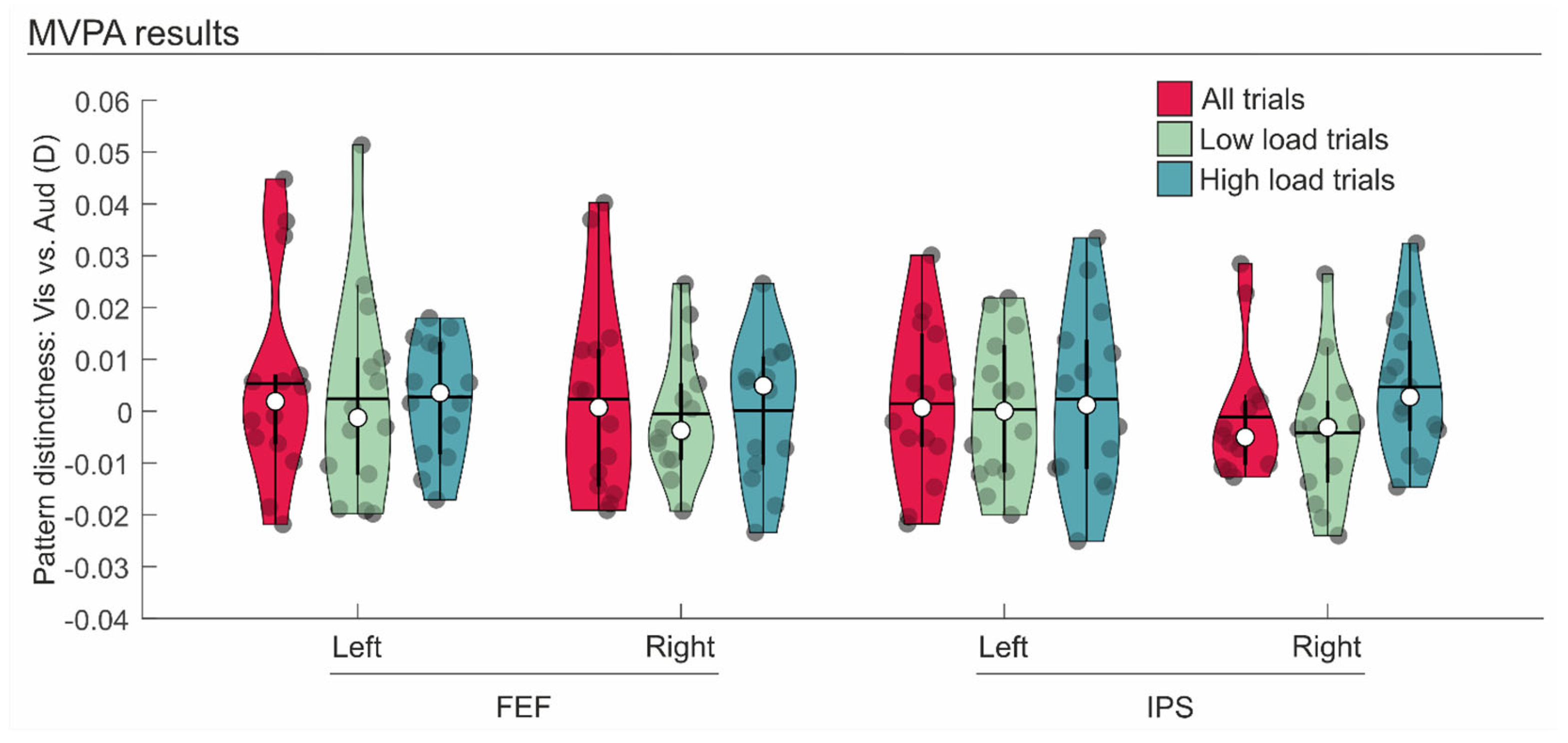
3.2. fMRI Data
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Baddeley, A. Working memory. Science 1992, 255, 556–559. [Google Scholar] [CrossRef]
- Cowan, N. Working Memory Capacity; Psychology Press: New York, NY, USA, 2005. [Google Scholar]
- Sternberg, S. High-speed scanning in human memory. Science 1966, 153, 652–654. [Google Scholar] [CrossRef]
- Gazzaley, A.; Nobre, A.C. Top-down modulation: Bridging selective attention and working memory. Trends Cogn. Sci. 2012, 16, 129–135. [Google Scholar] [CrossRef] [PubMed]
- Corbetta, M.; Shulman, G.L. Control of goal-directed and stimulus-driven attention in the brain. Nat. Rev. Neurosci. 2002, 3, 201–215. [Google Scholar] [CrossRef]
- Linden, D.E.; Bittner, R.A.; Muckli, L.; Waltz, J.A.; Kriegeskorte, N.; Goebel, R.; Singer, W.; Munk, M.H. Cortical capacity constraints for visual working memory: Dissociation of fMRI load effects in a fronto-parietal network. Neuroimage 2003, 20, 1518–1530. [Google Scholar] [CrossRef] [PubMed]
- McAfoose, J.; Baune, B.T. Exploring visual–spatial working memory: A critical review of concepts and models. Neuropsychol. Rev. 2009, 19, 130–142. [Google Scholar] [CrossRef] [PubMed]
- Todd, J.J.; Marois, R. Capacity limit of visual short-term memory in human posterior parietal cortex. Nature 2004, 428, 751–754. [Google Scholar] [CrossRef]
- Xu, Y.; Chun, M.M. Dissociable neural mechanisms supporting visual short-term memory for objects. Nature 2006, 440, 91–95. [Google Scholar] [CrossRef] [PubMed]
- Awh, E.; Vogel, E.K.; Oh, S.H. Interactions between attention and working memory. Neuroscience 2006, 139, 201–208. [Google Scholar] [CrossRef]
- Bledowski, C.; Kaiser, J.; Rahm, B. Basic operations in working memory: Contributions from functional imaging studies. Behav. Brain Res. 2010, 214, 172–179. [Google Scholar] [CrossRef]
- Ikkai, A.; Curtis, C.E. Common neural mechanisms supporting spatial working memory, attention and motor intention. Neuropsychologia 2011, 49, 1428–1434. [Google Scholar] [CrossRef] [PubMed]
- Linden, D.E. The working memory networks of the human brain. Neuroscientist 2007, 13, 257–267. [Google Scholar] [CrossRef]
- Postle, B.R. Distraction-spanning sustained activity during delayed recognition of locations. Neuroimage 2006, 30, 950–962. [Google Scholar] [CrossRef]
- Tamber-Rosenau, B.J.; Esterman, M.; Chiu, Y.C.; Yantis, S. Cortical mechanisms of cognitive control for shifting attention in vision and working memory. J. Cogn. Neurosci. 2011, 23, 2905–2919. [Google Scholar] [CrossRef] [PubMed]
- Van der Lubbe, R.H.; Bundt, C.; Abrahamse, E.L. Internal and external spatial attention examined with lateralized EEG power spectra. Brain Res. 2014, 1583, 179–192. [Google Scholar] [CrossRef] [PubMed]
- Van der Lubbe, R.H.J.; Panek, B.; Jahangier, I.; Asanowicz, D. Lateralized connectivity in the alpha band between parietal and occipital sources when spatial attention is externally and internally directed. Front. Cognit. 2023, 2, 1145854. [Google Scholar] [CrossRef]
- Emrich, S.M.; Riggall, A.C.; LaRocque, J.J.; Postle, B.R. Distributed patterns of activity in sensory cortex reflect the precision of multiple items maintained in visual short-term memory. J. Neurosci. 2013, 33, 6516–6523. [Google Scholar] [CrossRef]
- Li, D.; Christ, S.E.; Cowan, N. Domain-general and domain-specific functional networks in working memory. Neuroimage 2014, 102, 646–656. [Google Scholar] [CrossRef]
- Majerus, S.; Cowan, N.; Péters, F.; Van Calster, L.; Phillips, C.; Schrouff, J. Cross-modal decoding of neural patterns associated with working memory: Evidence for attention-based accounts of working memory. Cereb. Cortex 2016, 26, 166–179. [Google Scholar] [CrossRef]
- Riggall, A.C.; Postle, B.R. The relationship between working memory storage and elevated activity as measured with functional magnetic resonance imaging. J. Neurosci. 2012, 32, 12990–12998. [Google Scholar] [CrossRef]
- Naghavi, H.R.; Nyberg, L. Common fronto-parietal activity in attention, memory, and consciousness: Shared demands on integration? Conscious. Cogn. 2005, 14, 390–425. [Google Scholar] [CrossRef]
- Santangelo, V.; Macaluso, E. Spatial attention and audiovisual processing. In The New Handbook of Multisensory Processing; Stein, B.E., Ed.; MIT Press: Cambridge, MA, USA, 2012; pp. 359–370. [Google Scholar]
- Eger, E.; Sterzer, P.; Russ, M.O.; Giraud, A.L.; Kleinschmidt, A. A supramodal number representation in human intraparietal cortex. Neuron 2003, 37, 719–726. [Google Scholar] [CrossRef] [PubMed]
- Farah, M.J.; Wong, A.B.; Monheit, M.A.; Morrow, L.A. Parietal lobe mechanisms of spatial attention: Modality-specific or supramodal? Neuropsychologia 1989, 27, 461–470. [Google Scholar] [CrossRef] [PubMed]
- Macaluso, E.; Frith, C.D.; Driver, J. Supramodal effects of covert spatial orienting triggered by visual or tactile events. J. Cogn. Neurosci. 2002, 14, 389–401. [Google Scholar] [CrossRef] [PubMed]
- Spitzer, B.; Blankenburg, F. Supramodal parametric working memory processing in humans. J. Neurosci. 2012, 32, 3287–3295. [Google Scholar] [CrossRef] [PubMed]
- Spitzer, B.; Gloel, M.; Schmidt, T.T.; Blankenburg, F. Working memory coding of analog stimulus properties in the human prefrontal cortex. Cereb. Cortex 2014, 24, 2229–2236. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; Chen, C.; Spagna, A.; Wu, X.; Mackie, M.A.; Russell-Giller, S.; Xu, P.; Luo, Y.J.; Liu, X.; Hof, P.R.; et al. The functional anatomy of cognitive control: A domain-general brain network for uncertainty processing. J. Comp. Neurol. 2020, 528, 1265–1292. [Google Scholar] [CrossRef] [PubMed]
- Chambers, C.D.; Stokes, M.G.; Mattingley, J.B. Modality-specific control of strategic spatial attention in parietal cortex. Neuron 2004, 44, 925–930. [Google Scholar] [CrossRef]
- Dade, L.A.; Zatorre, R.J.; Evans, A.C.; Jones-Gotman, M. Working memory in another dimension: Functional imaging of human olfactory working memory. Neuroimage 2001, 14, 650–660. [Google Scholar] [CrossRef]
- Matsumoto, M.; Sakurada, T.; Yamamoto, S.I. Distinct bilateral prefrontal activity patterns associated with the qualitative aspect of working memory characterized by individual sensory modality dominance. PLoS ONE 2020, 15, e0238235. [Google Scholar] [CrossRef]
- Ruchkin, D.S.; Berndt, R.S.; Johnson, R., Jr.; Ritter, W.; Grafman, J.; Canoune, H.L. Modality-specific processing streams in verbal working memory: Evidence from spatio-temporal patterns of brain activity. Cogn. Brain Res. 1997, 6, 95–113. [Google Scholar] [CrossRef] [PubMed]
- Crottaz-Herbette, S.; Anagnoson, R.T.; Menon, V. Modality effects in verbal working memory: Differential prefrontal and parietal responses to auditory and visual stimuli. Neuroimage 2004, 21, 340–351. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Jimenez, R.; Avila, C.; Garcia-Navarro, C.; Bagney, A.; Aragon, A.M.; Ventura-Campos, N.; Martinez-Gras, I.; Forn, C.; Ponce, G.; Rubio, G.; et al. Differential dorsolateral prefrontal cortex activation during a verbal n-back task according to sensory modality. Behav. Brain Res. 2009, 205, 299–302. [Google Scholar] [CrossRef] [PubMed]
- Schumacher, E.H.; Lauber, E.; Awh, E.; Jonides, J.; Smith, E.E.; Koeppe, R.A. PET evidence for an amodal verbal working memory system. Neuroimage 1996, 3, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Mastroberardino, S.; Santangelo, V.; Botta, F.; Marucci, F.S.; Olivetti Belardinelli, M. How the bimodal format of presentation affects working memory: An overview. Cogn. Process. 2008, 9, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Kravitz, D.J.; Saleem, K.S.; Baker, C.I.; Mishkin, M. A new neural framework for visuospatial processing. Nat. Rev. Neurosci. 2011, 12, 217–230. [Google Scholar] [CrossRef] [PubMed]
- Friston, K.J.; Glaser, D.E.; Henson, R.N.; Kiebel, S.; Phillips, C.; Ashburner, J. Classical and Bayesian inference in neuroimaging: Applications. Neuroimage 2002, 16, 484–512. [Google Scholar] [CrossRef]
- Santangelo, V.; Bordier, C. Large-scale brain networks underlying successful and unsuccessful encoding, maintenance, and retrieval of everyday scenes in visuospatial working memory. Front. Psychol. 2019, 10, 233. [Google Scholar] [CrossRef]
- Santangelo, V.; Macaluso, E. Visual saliency improve spatial working memory via enhanced parieto-temporal functional connectivity. J. Neurosci. 2013, 33, 4110–4117. [Google Scholar] [CrossRef]
- Santangelo, V.; Di Francesco, S.A.; Mastroberardino, S.; Macaluso, E. Parietal cortex integrates contextual and saliency signals during the encoding of natural scenes in working memory. Hum. Brain Mapp. 2015, 36, 5003–5017. [Google Scholar] [CrossRef]
- Allefeld, C.; Haynes, J.D. Searchlight-based multi-voxel pattern analysis of fMRI by cross-validated MANOVA. Neuroimage 2014, 89, 345–357. [Google Scholar] [CrossRef] [PubMed]
- Grootswagers, T.; Wardle, S.G.; Carlson, T.A. Decoding dynamic brain patterns from evoked responses: A tutorial on multivariate pattern analysis applied to time series neuroimaging data. J. Cogn. Neurosci. 2017, 29, 677–697. [Google Scholar] [CrossRef] [PubMed]
- Haxby, J.V.; Connolly, A.C.; Guntupalli, J.S. Decoding neural representational spaces using multivariate pattern analysis. Annu. Rev. Neurosci. 2014, 37, 435–456. [Google Scholar] [CrossRef] [PubMed]
- Christophel, T.B.; Iamshchinina, P.; Yan, C.; Allefeld, C.; Haynes, J.D. Cortical specialization for attended versus unattended working memory. Nat. Neurosci. 2018, 21, 494–496. [Google Scholar] [CrossRef] [PubMed]
- Gardumi, A.; Ivanov, D.; Hausfeld, L.; Valente, G.; Formisano, E.; Uludağ, K. The effect of spatial resolution on decoding accuracy in fMRI multivariate pattern analysis. Neuroimage 2016, 132, 32–42. [Google Scholar] [CrossRef]
- Keysers, C.; Gazzola, V.; Wagenmakers, E.J. Using Bayes factor hypothesis testing in neuroscience to establish evidence of absence. Nat. Neurosci. 2020, 23, 788–799. [Google Scholar] [CrossRef]
- Corbetta, M.; Patel, G.; Shulman, G.L. The reorienting system of the human brain: From environment to theory of mind. Neuron 2008, 58, 306–324. [Google Scholar] [CrossRef]
- Nyberg, L.; Dahlin, E.; Stigsdotter Neely, A.; Bäckman, L. Neural correlates of variable working memory load across adult age and skill: Dissociative patterns within the fronto-parietal network. Scand. J. Psychol. 2009, 50, 41–46. [Google Scholar] [CrossRef]
- Santangelo, V.; Macaluso, E. The contribution of working memory to divided attention. Hum. Brain Mapp. 2013, 34, 158–175. [Google Scholar] [CrossRef]
- Lee, S.H.; Kravitz, D.J.; Baker, C.I. Goal-dependent dissociation of visual and prefrontal cortices during working memory. Nat. Neurosci. 2013, 16, 997–999. [Google Scholar] [CrossRef]
- Christophel, T.B.; Klink, P.C.; Spitzer, B.; Roelfsema, P.R.; Haynes, J.D. The distributed nature of working memory. Trends Cogn. Sci. 2017, 21, 111–124. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Baker, C.I. Multi-voxel decoding and the topography of maintained information during visual working memory. Front. Syst. Neurosci. 2016, 10, 2. [Google Scholar] [CrossRef]
- Serences, J.T.; Ester, E.F.; Vogel, E.K.; Awh, E. Stimulus-specific delay activity in human primary visual cortex. Psychol. Sci. 2009, 20, 207–214. [Google Scholar] [CrossRef] [PubMed]
- Sprague, T.C.; Ester, E.F.; Serences, J.T. Reconstructions of information in visual spatial working memory degrade with memory load. Curr. Biol. 2014, 24, 2174–2180. [Google Scholar] [CrossRef] [PubMed]
- Xing, Y.; Ledgeway, T.; McGraw, P.V.; Schluppeck, D. Decoding working memory of stimulus contrast in early visual cortex. J. Neurosci. 2013, 33, 10301–10311. [Google Scholar] [CrossRef] [PubMed]
- Christophel, T.B.; Hebart, M.N.; Haynes, J.D. Decoding the contents of visual short-term memory from human visual and parietal cortex. J. Neurosci. 2012, 32, 12983–12989. [Google Scholar] [CrossRef] [PubMed]
- Christophel, T.B.; Cichy, R.M.; Hebart, M.N.; Haynes, J.D. Parietal and early visual cortices encode working memory content across mental transformations. Neuroimage 2015, 106, 198–206. [Google Scholar] [CrossRef]
- Jerde, T.A.; Merriam, E.P.; Riggall, A.C.; Hedges, J.H.; Curtis, C.E. Prioritized maps of space in human frontoparietal cortex. J. Neurosci. 2012, 32, 17382–17390. [Google Scholar] [CrossRef]
- Christophel, T.B.; Haynes, J.D. Decoding complex flow-field patterns in visual working memory. Neuroimage 2014, 91, 43–51. [Google Scholar] [CrossRef]
- Naughtin, C.K.; Mattingley, J.B.; Dux, P.E. Distributed and overlapping neural substrates for object individuation and identification in visual short-term memory. Cereb. Cortex 2016, 26, 566–575. [Google Scholar] [CrossRef]
- Ester, E.F.; Sprague, T.C.; Serences, J.T. Parietal and frontal cortex encode stimulus-specific mnemonic representations during visual working memory. Neuron 2015, 87, 893–905. [Google Scholar] [CrossRef] [PubMed]
- Kowialiewski, B.; Krasnoff, J.; Mizrak, E.; Oberauer, K. Verbal working memory encodes phonological and semantic information differently. Cognition 2023, 233, 105364. [Google Scholar] [CrossRef] [PubMed]
- Santangelo, V.; Spence, C. Assessing the effect of verbal working memory load on visuo-spatial exogenous orienting. Neurosci. Lett. 2007, 413, 105–109. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Brain Region | Cluster p-FWE-Corr | Cluster Size (k) | Peak t-Value | Peak Coordinates X Y Z |
|---|---|---|---|---|
| Left FEF | 0.018 | 192 | 7.22 | −22 −4 58 |
| Right FEF | 0.016 | 198 | 6.32 | 26 −2 48 |
| Left SPL | <0.001 | 1979 | 10.77 | −22 −62 62 |
| Right SPL | <0.001 | 595 | 8.69 | 22 −62 64 |
| Contrast | Brain Region | t(13) | p-Value | Cohen’s d | BF10 |
|---|---|---|---|---|---|
| Visual vs. Auditory: all trials | Left FEF | 0.991 | 0.170 | 0.265 | 0.668 |
| Right FEF | 0.458 | 0.327 | 0.122 | 0.392 | |
| Left SPL | 0.350 | 0.366 | 0.094 | 0.357 | |
| Right SPL | −0.325 | 0.625 | −0.087 | 0.216 | |
| Visual vs. Auditory: only low load trials | Left FEF | 0.453 | 0.329 | 0.121 | 0.391 |
| Right FEF | −0.165 | 0.564 | −0.044 | 0.240 | |
| Left SPL | 0.097 | 0.462 | 0.026 | 0.290 | |
| Right SPL | −1.168 | 0.868 | −0.312 | 0.141 | |
| Visual vs. Auditory: only high load trials | Left FEF | 0.898 | 0.193 | 0.240 | 0.604 |
| Right FEF | 0.035 | 0.486 | 0.009 | 0.277 | |
| Left SPL | 0.501 | 0.312 | 0.134 | 0.408 | |
| Right SPL | 1.326 | 0.104 | 0.354 | 0.986 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rizza, A.; Pedale, T.; Mastroberardino, S.; Olivetti Belardinelli, M.; Van der Lubbe, R.H.J.; Spence, C.; Santangelo, V. Working Memory Maintenance of Visual and Auditory Spatial Information Relies on Supramodal Neural Codes in the Dorsal Frontoparietal Cortex. Brain Sci. 2024, 14, 123. https://doi.org/10.3390/brainsci14020123
Rizza A, Pedale T, Mastroberardino S, Olivetti Belardinelli M, Van der Lubbe RHJ, Spence C, Santangelo V. Working Memory Maintenance of Visual and Auditory Spatial Information Relies on Supramodal Neural Codes in the Dorsal Frontoparietal Cortex. Brain Sciences. 2024; 14(2):123. https://doi.org/10.3390/brainsci14020123
Chicago/Turabian StyleRizza, Aurora, Tiziana Pedale, Serena Mastroberardino, Marta Olivetti Belardinelli, Rob H. J. Van der Lubbe, Charles Spence, and Valerio Santangelo. 2024. "Working Memory Maintenance of Visual and Auditory Spatial Information Relies on Supramodal Neural Codes in the Dorsal Frontoparietal Cortex" Brain Sciences 14, no. 2: 123. https://doi.org/10.3390/brainsci14020123