
fNIRS-Based Differences in Cortical Activation during Tool Use, Pantomimed Actions, and Meaningless Actions between Children with and without Autism Spectrum Disorder (ASD)

Abstract
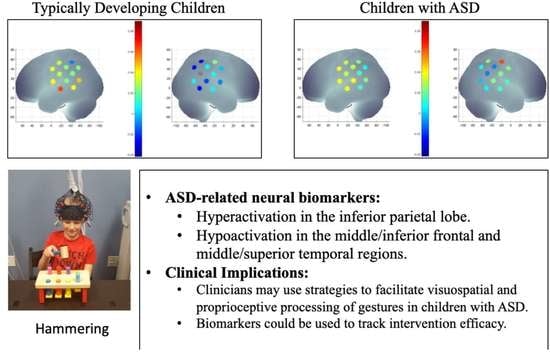
:
1. Introduction
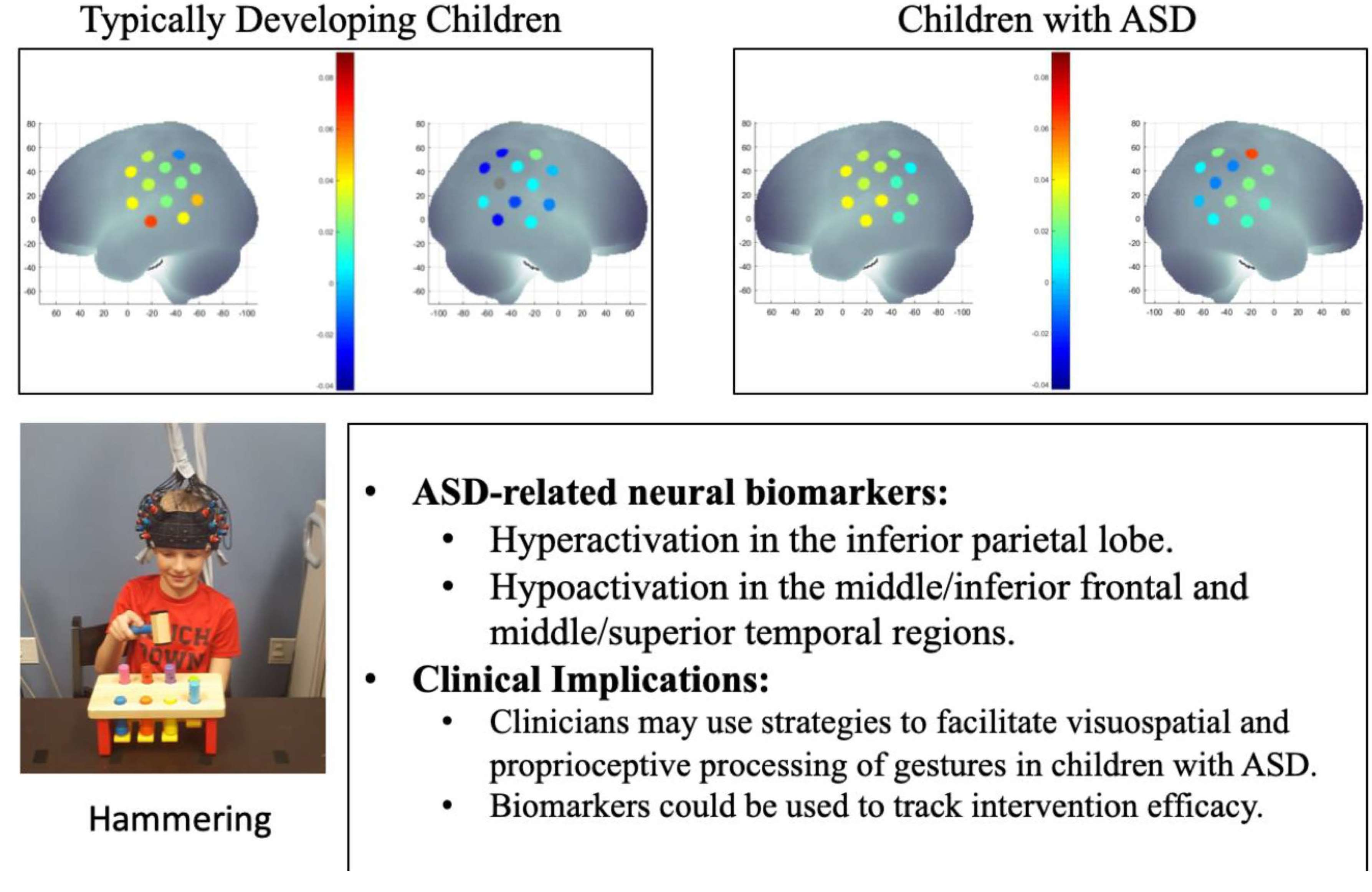
1.1. Behavioral Framework for Tool Use
1.2. Neural Mechanisms Underlying Tool Use
1.3. Dyspraxia in Children with ASD
1.4. ASD-Related Atypical Cortical Activation during Tool Use and Pantomimed Tool Use
1.5. Aims and Hypotheses of This Study
2. Materials and Methods
2.1. Participants
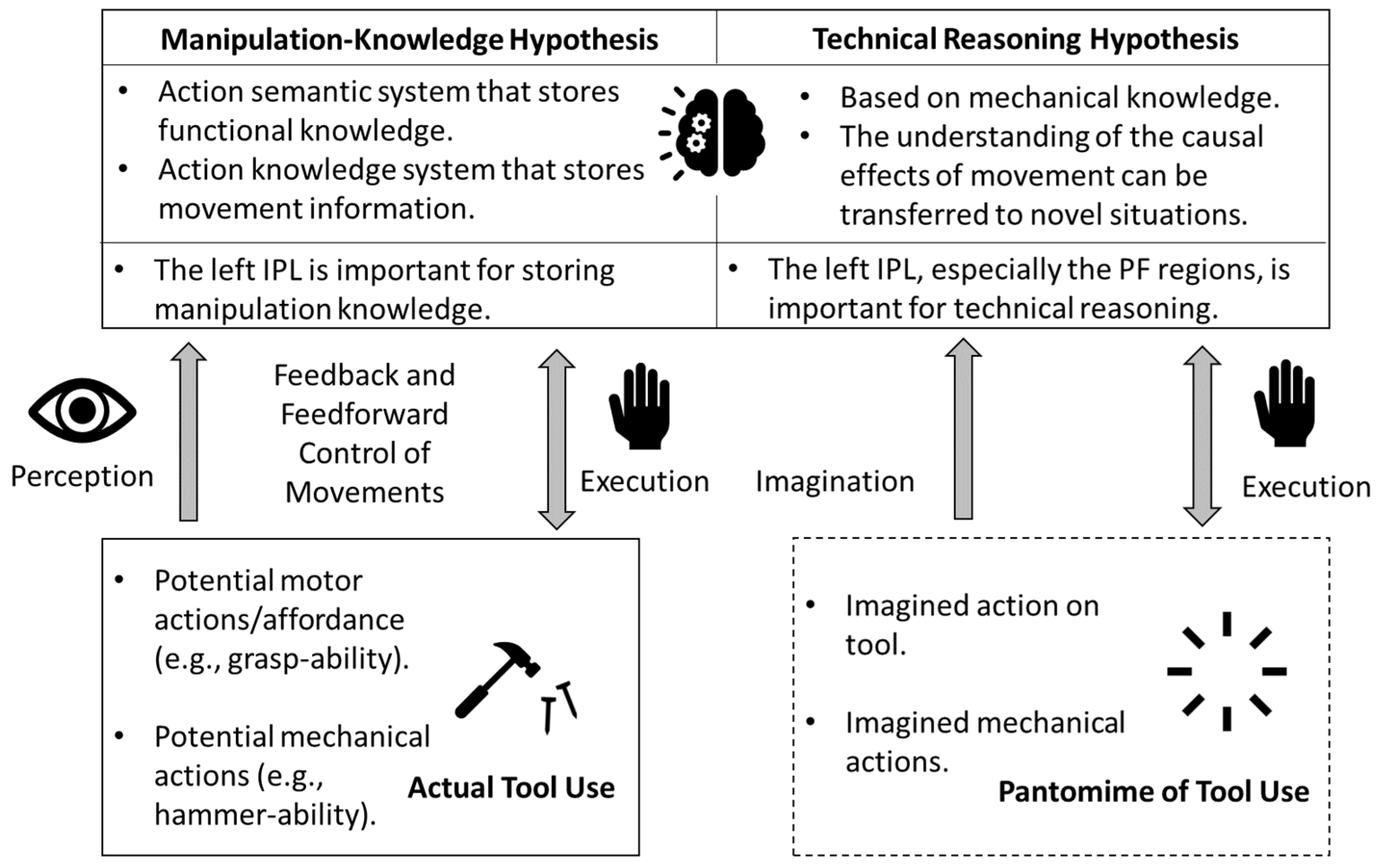
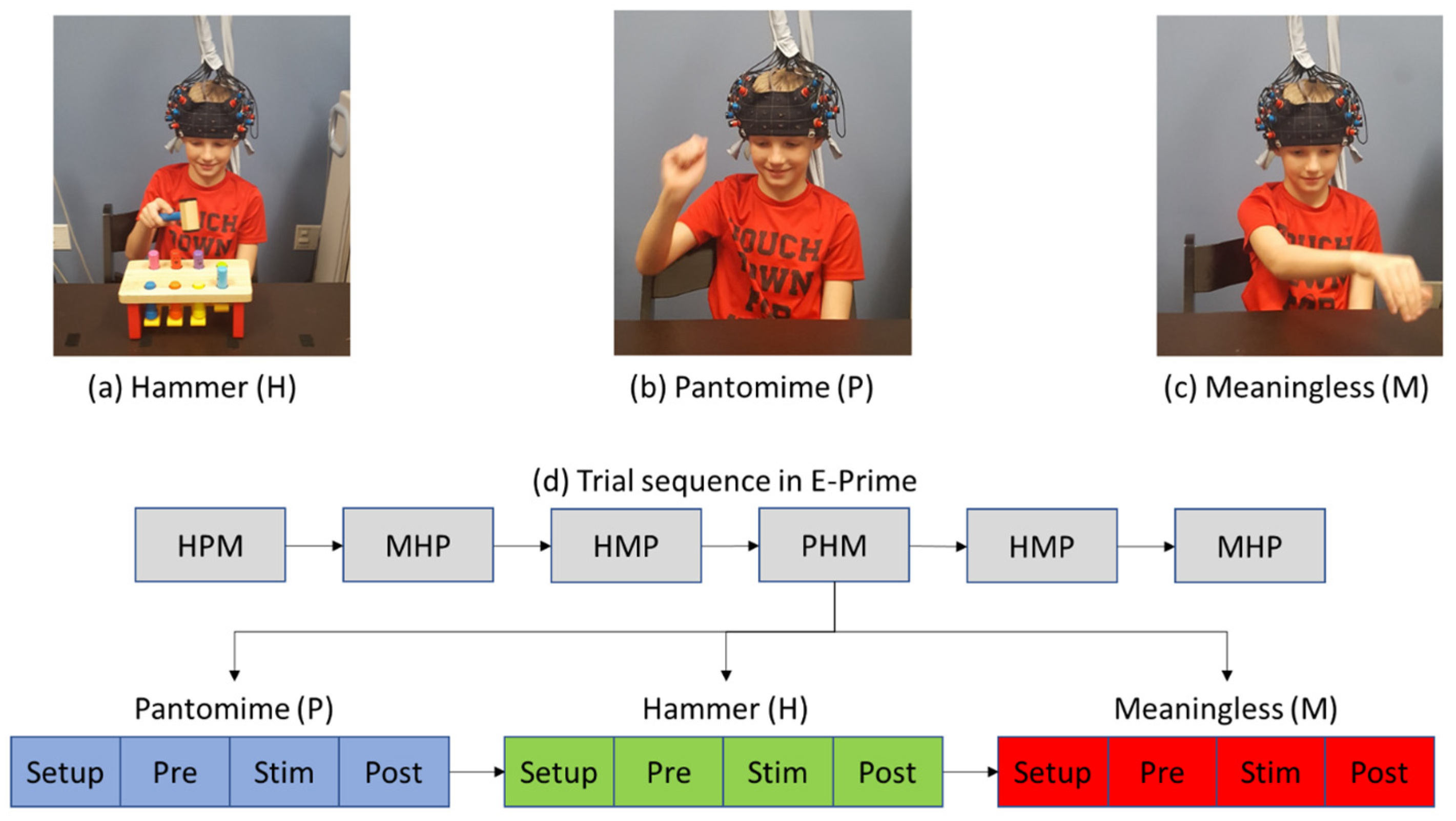
2.2. Study Procedures
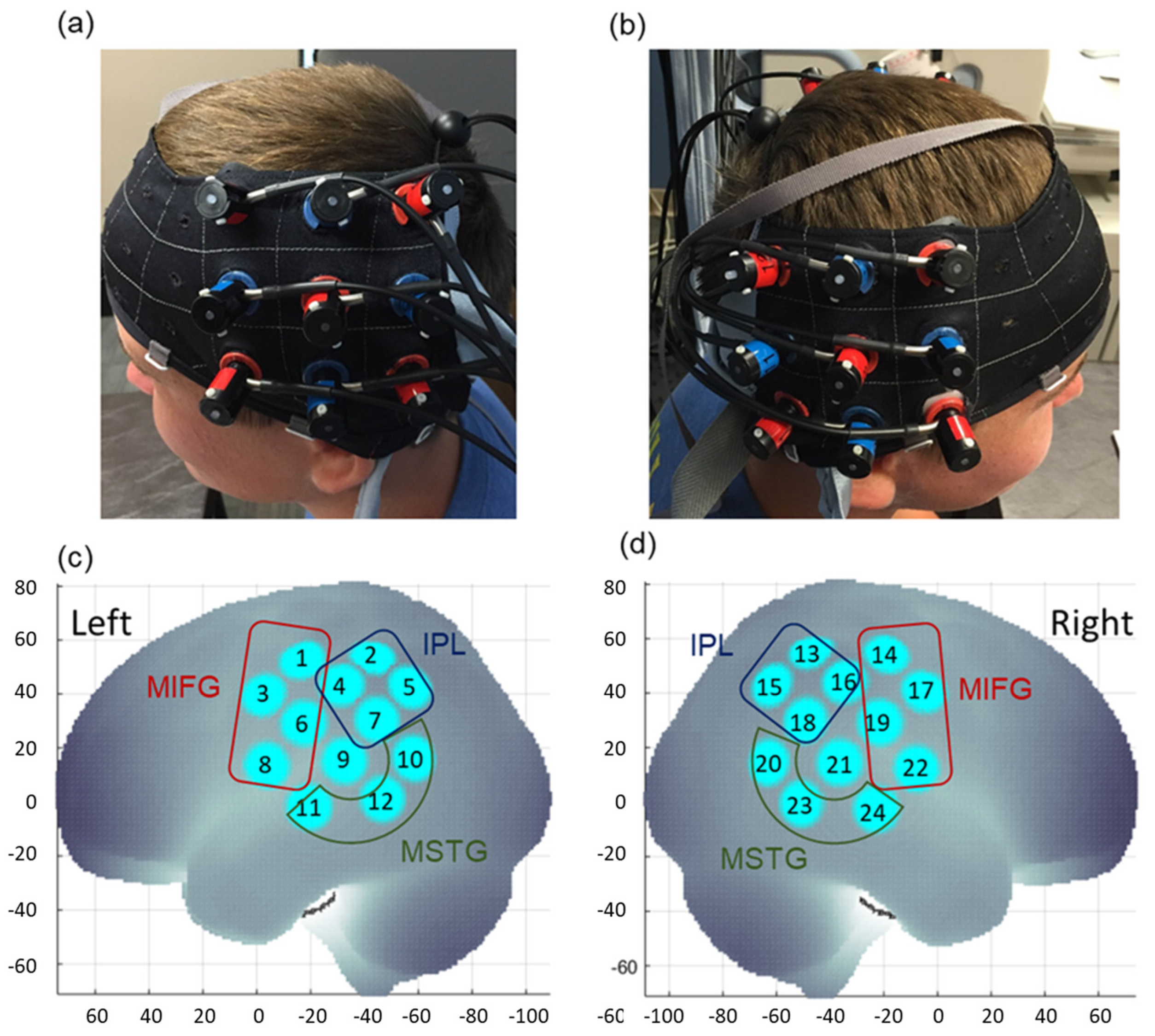
2.3. fNIRS Data Collection
2.4. Spatial Registration Approach
2.5. Video Coding for SIPT-PP and SIPT-BMC
2.6. Processing Cortical-Activation Data
2.7. Data Exclusion for Cortical Activation Data
2.8. Statistical Analyses
3. Results
3.1. Praxis Performance
3.2. Cortical Activation
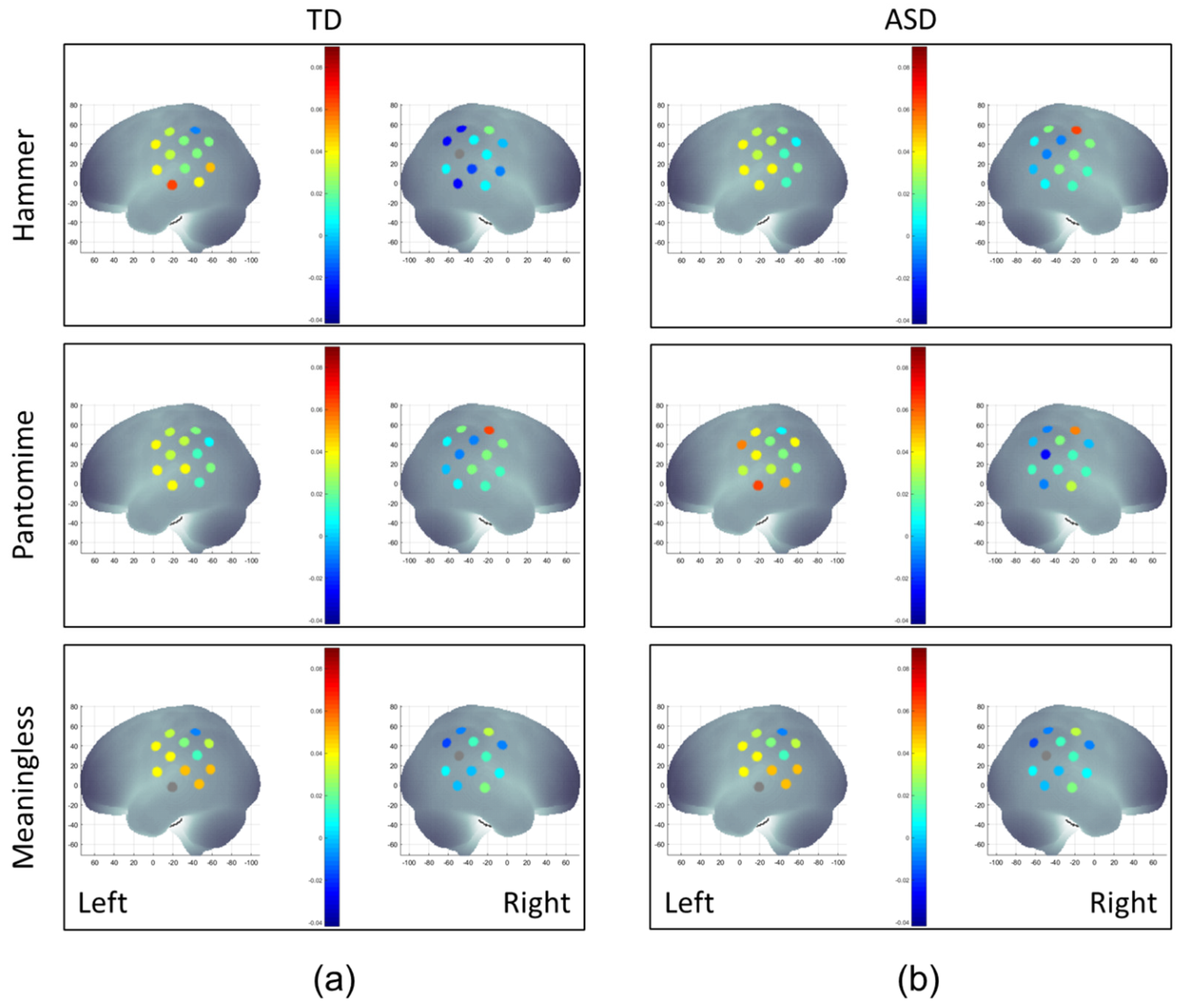
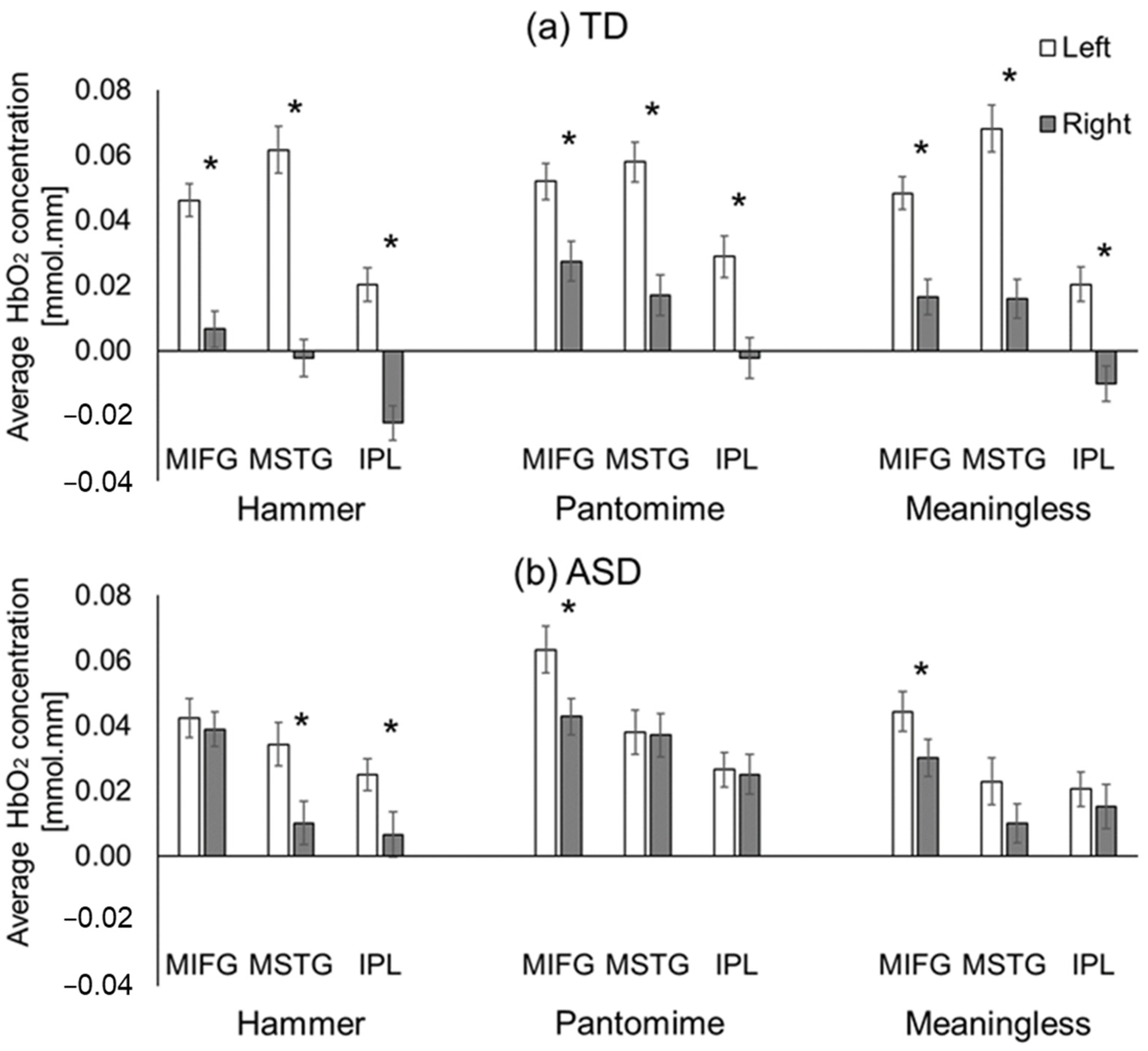
3.2.1. Hemispheric Differences
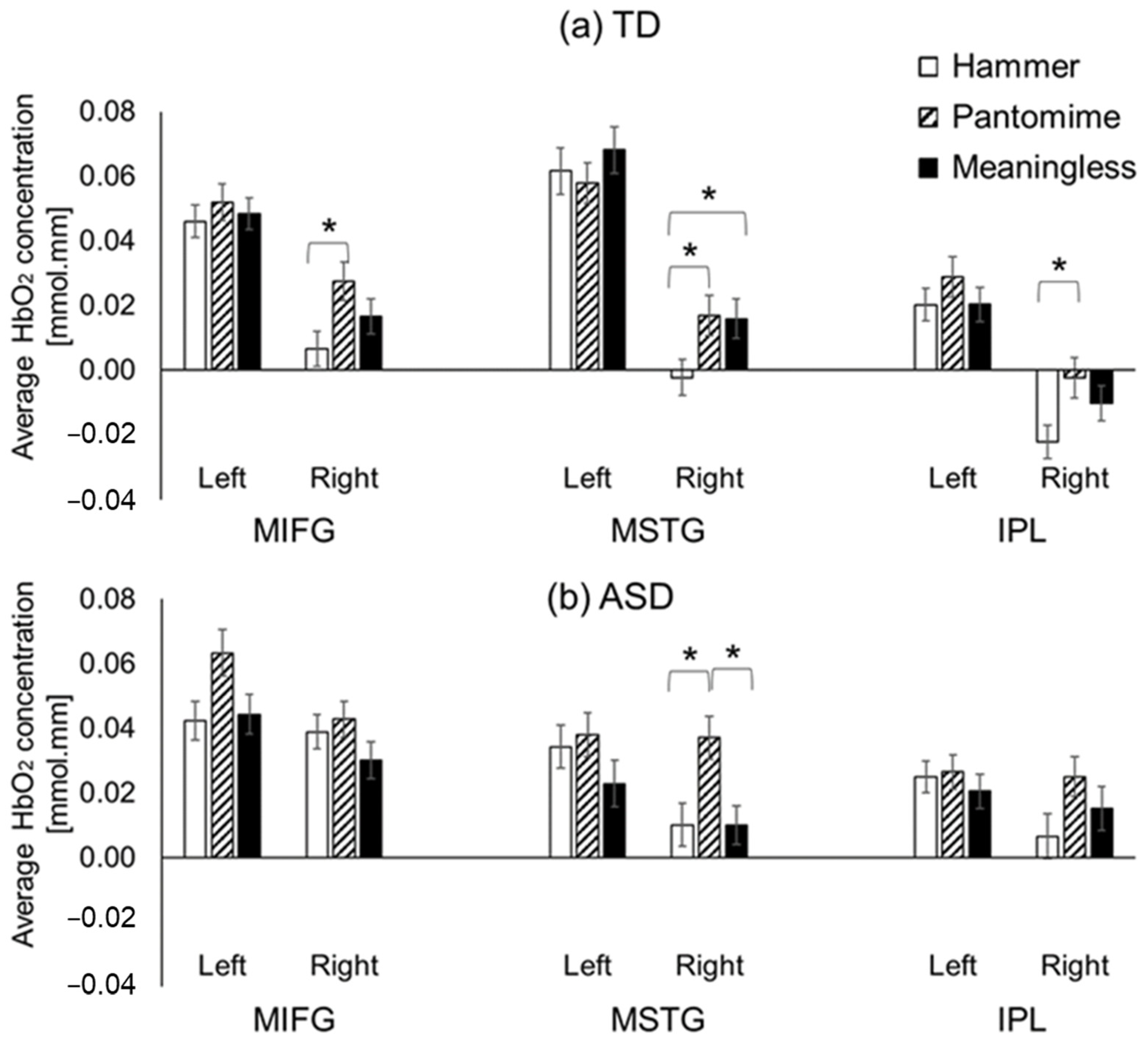
3.2.2. Condition-Related Differences
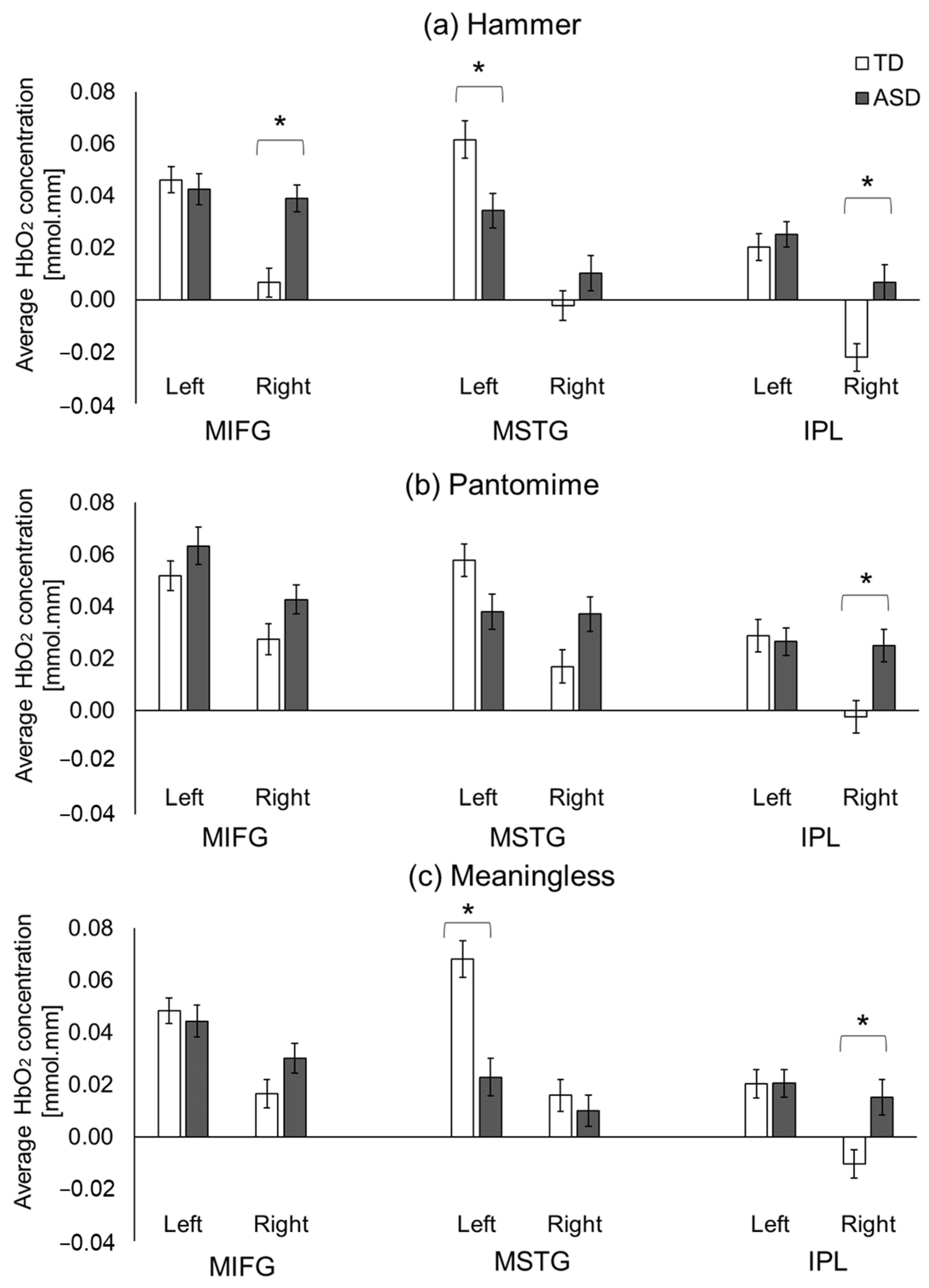
3.2.3. ASD-Related/Group Differences
3.3. Correlations between Cortical Activation and Praxis Performance
3.4. Correlations between Cortical Activation and ADOS Scores in Children with ASD
3.5. Correlations between Cortical Activation and VABS Scores
4. Discussion
4.1. Impaired Praxis Performance in Children with ASD
4.2. Left-Lateralized Activation in Children without ASD and More Bilateral Activation in Children with ASD during Gestural Performance
4.3. Greater Right-Hemispheric Activation during Pantomimed Actions in Children with and without ASD
4.4. Atypical Activation of the Parietal, Frontal, and Temporal Networks during Actual and Pantomimed Tool Use in Children with ASD
4.4.1. Intact Left IPL but Hyperactivation of Right IPL in Children with ASD
4.4.2. ASD-Related Hyperactivation of Right MIFG Region during Actual Tool Use
4.4.3. ASD-Related Hypoactivation of Left MSTG When Performing Tool Use and Meaningless Gestures
4.5. Limitations and Future Directions
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders, 5th ed.; American Psychiatric Publishing: Washington, DC, USA, 2013. [Google Scholar]
- Bhat, A.N.; Landa, R.J.; Galloway, J.C. Current perspectives on motor functioning in infants, children, and adults with autism spectrum disorders. Phys. Ther. 2011, 91, 1116–1129. [Google Scholar] [CrossRef] [Green Version]
- Kaur, M.; Srinivasan, S.; Bhat, A. Comparing motor performance, praxis, coordination, and interpersonal synchrony between children with and without Autism Spectrum Disorder (ASD). Res. Dev. Disabil. 2018, 72, 79–95. [Google Scholar] [CrossRef]
- Bhat, A. Is motor impairment in ASD distinct from developmental coordination disorder (DCD)? A Report from the SPARK study. Phys. Ther. 2020, 100, 633–644. [Google Scholar] [CrossRef]
- Bhat, A. Motor impairment increases in children with ASD as a function of social communication, cognitive, and functional impairments, repetitive behavior severity, and comorbid diagnoses: A SPARK study report. Autism Res. 2021, 14, 202–219. [Google Scholar] [CrossRef]
- Bhat, A.; Boulton, A.; Tulsky, D. A further study of relations between motor, social communication/language, cognitive, and functional impairments as well as repetitive behavior severity in children with Autism Spectrum Disorder (ASD): A SPARK dataset analysis. Autism Res. 2022, 15, 1156–1178. [Google Scholar] [CrossRef]
- Mostofsky, S.H.; Dubey, P.; Jerath, V.K.; Jansiewicz, E.M.; Goldberg, M.C.; Denckla, M.B. Developmental dyspraxia is not limited to imitation in children with autism spectrum disorders. J. Int. Neuropsychol. Soc. 2006, 12, 314–326. [Google Scholar] [CrossRef] [Green Version]
- Gizzonio, V.; Avanzini, P.; Campi, C.; Orivoli, S.; Piccolo, B.; Cantalupo, G.; Tassinari, C.A.; Rizzolatti, G.; Fabbri-Destro, M. Failure in pantomime action execution correlates with the severity of social behavior deficits in children with autism: A praxis study. J. Autism Dev. Disord. 2015, 45, 3085–3097. [Google Scholar] [CrossRef]
- Bhat, A.; Srinivasan, S.; Woxholdt, C.; Shield, A. Differences in praxis performance and receptive language during fingerspelling between deaf children with and without autism spectrum disorder. Autism 2018, 22, 271–282. [Google Scholar] [CrossRef]
- Dewey, D.; Cantell, M.; Crawford, S.G. Motor and gestural performance in children with autism spectrum disorders, developmental coordination disorder, and/or attention deficit hyperactivity disorder. J. Int. Neuropsychol. Soc. 2007, 13, 246–256. [Google Scholar] [CrossRef]
- Dziuk, M.A.; Gidley Larson, J.C.; Apostu, A.; Mahone, E.M.; Denckla, M.B.; Mostofsky, S.H. Dyspraxia in autism: Association with motor, social, and communicative deficits. Dev. Med. Child. Neurol. 2007, 49, 734–739. [Google Scholar] [CrossRef]
- Dowell, L.R.; Mahone, E.M.; Mostofsky, S.H. Associations of postural knowledge and basic motor skill with dyspraxia in autism: Implication for abnormalities in distributed connectivity and motor learning. Neuropsychology 2009, 23, 563–570. [Google Scholar] [CrossRef] [Green Version]
- Shield, A.; Knape, K.; Henry, M.; Srinivasan, S.; Bhat, A. Impaired praxis in gesture imitation by deaf children with autism spectrum disorder. Autism Dev. Lang. Impair. 2017, 2. [Google Scholar] [CrossRef]
- Ambrose, S.H. Paleolithic technology and human evolution. Science 2001, 291, 1748–1753. [Google Scholar] [CrossRef]
- Bartolo, A.; Ham, H.S. A cognitive overview of limb apraxia. Curr. Neurol. Neurosci. Rep. 2016, 16, 75. [Google Scholar] [CrossRef]
- Canzano, L.; Scandola, M.; Gobbetto, V.; Moretto, G.; D’Imperio, D.; Moro, V. The representation of objects in apraxia: From action execution to error awareness. Front. Hum. Neurosci. 2016, 10, 39. [Google Scholar] [CrossRef]
- Ellis, R.; Tucker, M. Micro-affordance: The potentiation of components of action by seen objects. Br. J. Psychol. 2000, 91, 451–471. [Google Scholar] [CrossRef] [Green Version]
- Osiurak, F.; Federico, G. Four ways of (mis-)conceiving embodiment in tool use. Synthese 2020, 199, 3853–3879. [Google Scholar] [CrossRef]
- Foundas, A.L.; Duncan, E.S. Limb apraxia: A disorder of learned skilled movement. Curr. Neurol. Neurosci. Rep. 2019, 19, 82. [Google Scholar] [CrossRef]
- Osiurak, F.; Claidière, N.; Bluet, A.; Brogniart, J.; Lasserre, S.; Bonhoure, T.; Di Rollo, L.; Gorry, N.; Polette, Y.; Saude, A.; et al. Technical reasoning bolsters cumulative technological culture through convergent transformations. Sci. Adv. 2022, 8, eabl7446. [Google Scholar] [CrossRef]
- Osiurak, F.; Claidière, N.; Federico, G. Bringing cumulative technological culture beyond copying versus reasoning. Trends Cogn. Sci. 2023, 27, 30–42. [Google Scholar] [CrossRef]
- Mostofsky, S.H.; Ewen, J.B. Altered connectivity and action model formation in autism is autism. Neuroscientist 2011, 174, 437–448. [Google Scholar] [CrossRef] [Green Version]
- Wymbs, N.F.; Grafton, S.T. The human motor system supports sequence-specific representations over multiple training-dependent timescales. Cereb. Cortex 2015, 2511, 4213–4225. [Google Scholar] [CrossRef] [Green Version]
- Buxbaum, L.J.; Kyle, K.M.; Menon, R. On beyond mirror neurons: Internal representations subserving imitation and recognition of skilled object-related actions in humans. Brain Res. Cogn. Brain Res. 2005, 25, 226–239. [Google Scholar] [CrossRef]
- Raymer, A.M.; Maher, L.M.; Foundas, A.L.; Heilman, K.M.; Rothi, L.J. The significance of body part as tool errors in limb apraxia. Brain Cogn. 1997, 34, 287–292. [Google Scholar] [CrossRef] [Green Version]
- Poole, J.L.; Gallagher, J.; Janosky, J.; Qualls, C. The mechanisms for adult-onset apraxia and developmental dyspraxia: An examination and comparison of error patterns. Am. J. Occup. Ther. 1997, 51, 339–346. [Google Scholar] [CrossRef] [Green Version]
- Donkervoort, M.; Dekker, J.; van den Ende, E.; Stehmann-Saris, J.C.; Deelman, B.G. Prevalence of apraxia among patients with a first left hemisphere stroke in rehabilitation centres and nursing homes. Clin. Rehabil. 2000, 14, 130–136. [Google Scholar] [CrossRef] [Green Version]
- Ogawa, K.; Imai, F. Hand-independent representation of tool-use pantomimes in the left anterior intraparietal cortex. Exp. Brain Res. 2016, 234, 3677–3687. [Google Scholar] [CrossRef] [Green Version]
- Brandi, M.L.; Wohlschläger, A.; Sorg, C.; Hermsdörfer, J. The neural correlates of planning and executing actual tool use. J. Neurosci. 2014, 34, 13183–13194. [Google Scholar] [CrossRef] [Green Version]
- Ishibashi, R.; Pobric, G.; Saito, S.; Lambon Ralph, M.A. The neural network for tool-related cognition: An activation likelihood estimation meta-analysis of 70 neuroimaging contrasts. Cogn. Neuropsychol. 2016, 33, 241–256. [Google Scholar] [CrossRef] [Green Version]
- Ohgami, Y.; Matsuo, K.; Uchida, N.; Nakai, T. An fMRI study of tool-use gestures: Body part as object and pantomime. Neuroreport 2004, 15, 1903–1906. [Google Scholar] [CrossRef]
- Johnson-Frey, S.H.; Newman-Norlund, R.; Grafton, S.T. A distributed left hemisphere network active during planning of everyday tool use skills. Cereb. Cortex 2005, 15, 681–695. [Google Scholar] [CrossRef] [Green Version]
- Kaltenbach, M.L.; Brett, M.; Patterson, K. Actions speak louder than functions: The importance of manipulability and action in tool representation. J. Cogn. Neurosci. 2003, 15, 30–46. [Google Scholar]
- Binkofski, F.; Buccino, G.; Stephan, K.M.; Rizzolatti, G.; Seitz, R.J.; Freund, H.J. A parieto-premotor network for object manipulation: Evidence from neuroimaging. Exp. Brain Res. 1991, 128, 210–213. [Google Scholar] [CrossRef]
- Buchwald, M.; Przybylski, Ł.; Króliczak, G. Decoding brain states for planning functional grasps of tools: A functional magnetic resonance imaging multivoxel pattern analysis study. J. Int. Neuropsychol. Soc. 2018, 24, 1013–1025. [Google Scholar] [CrossRef]
- Fischer, J.; Mikhael, J.G.; Tenenbaum, J.B.; Kanwisher, N. Functional neuroanatomy of intuitive physical inference. Proc. Natl. Acad. Sci. USA 2016, 113, E5072–E5081. [Google Scholar] [CrossRef] [Green Version]
- Federico, G.; Reynaud, E.; Navarro, J.; Lesourd, M.; Gaujoux, V.; Lamberton, F.; Ibarrola, D.; Cavaliere, C.; Alfano, V.; Aiello, M.; et al. The cortical thickness of the area PF of the left inferior parietal cortex mediates technical-reasoning skills. Sci. Rep. 2022, 12, 11840. [Google Scholar] [CrossRef]
- Osiurak, F.; Reynaud, E.; Baumard, J.; Rossetti, Y.; Bartolo, A.; Lesourd, M. Pantomime of tool use: Looking beyond apraxia. Brain Commun. 2021, 3, fcab263. [Google Scholar] [CrossRef]
- Tanaka, S.; Inui, T.; Iwaki, S.; Konishi, J.; Nakai, T. Neural substrates involved in imitating finger configurations: An fMRI study. Neuroreport 2001, 12, 1171–1174. [Google Scholar] [CrossRef] [Green Version]
- Hermsdörfer, J.; Goldenberg, G.; Wachsmuth, C.; Conrad, B.; Ceballos-Baumann, A.O.; Bartenstein, P.; Schwaiger, M.; Boecker, H. Cortical correlates of gesture processing: Clues to the cerebral mechanisms underlying apraxia during the imitation of meaningless gestures. NeuroImage 2001, 14, 149–161. [Google Scholar] [CrossRef]
- Imazu, S.; Sugio, T.; Tanaka, S.; Inui, T. Differences between actual and imagined usage of chopsticks: An fMRI study. Cortex 2007, 43, 301–307. [Google Scholar] [CrossRef]
- Lausberg, H.; Kazzer, P.; Heekeren, H.R.; Wartenburger, I. Pantomiming tool use with an imaginary tool in hand as compared to demonstration with tool in hand specifically modulates the left middle and superior temporal gyri. Cortex 2015, 71, 1–14. [Google Scholar] [CrossRef]
- Muth, A.; Hönekopp, J.; Falter, C.M. Visuo-spatial performance in autism: A meta-analysis. J. Autism Dev. Disord. 2014, 44, 3245–3263. [Google Scholar] [CrossRef] [Green Version]
- Caron, M.J.; Mottron, L.; Berthiaume, C.; Dawson, M. Cognitive mechanisms, specificity and neural underpinnings of visuospatial peaks in autism. Brain 2006, 129 Pt 7, 1789–1802. [Google Scholar] [CrossRef] [Green Version]
- Williams, J.H.; Whiten, A.; Suddendorf, T.; Perrett, D.I. Imitation, mirror neurons and autism. Neurosci. Biobehav. Rev. 2001, 25, 287–295. [Google Scholar] [CrossRef] [Green Version]
- Wadsworth, H.M.; Maximo, J.O.; Lemelman, A.R.; Clayton, K.; Sivaraman, S.; Deshpande, H.D.; Ver Hoef, L.; Kana, R.K. The action imitation network and motor imitation in children and adolescents with autism. Neuroscience 2017, 343, 147–156. [Google Scholar] [CrossRef]
- Wymbs, N.F.; Nebel, M.B.; Ewen, J.B.; Mostofsky, S.H. Altered inferior parietal functional connectivity is correlated with praxis and social skill performance in children with autism spectrum disorder. Cereb. Cortex 2020, 31, 2639–2652. [Google Scholar] [CrossRef]
- Fourie, E.; Palser, E.R.; Pokorny, J.J.; Neff, M.; Rivera, S.M. Neural processing and production of gesture in children and adolescents with autism spectrum disorder. Front. Psychol. 2020, 10, 3045. [Google Scholar] [CrossRef]
- Ewen, J.B.; Lakshmanan, B.M.; Pillai, A.S.; McAuliffe, D.; Nettles, C.; Hallett, M.; Crone, N.E.; Mostofsky, S.H. Decreased modulation of EEG oscillations in high-functioning autism during a motor control task. Front. Hum. Neurosci. 2016, 10, 198. [Google Scholar] [CrossRef] [Green Version]
- Lloyd-Fox, S.; Blasi, A.; Elwell, C.E. Illuminating the developing brain: The past, present and future of functional near infrared spectroscopy. Neurosci. Biobehav. Rev. 2010, 34, 269–284. [Google Scholar] [CrossRef]
- McPartland, J.C.; Lerner, M.D.; Bhat, A.; Clarkson, T.; Jack, A.; Koohsari, S.; Matuskey, D.; McQuaid, G.A.; Su, W.C.; Trevisan, D.A. Looking back at the next 40 years of ASD neuroscience research. J. Autism Dev. Disord. 2021, 51, 4333–4353. [Google Scholar] [CrossRef]
- Bhat, A.; Hoffman, M.; Trost, S.; Eilbott, J.; Pelphrey, K. Cortical activation patterns during action observation, action execution, & interpersonal synchrony in adults: A functional near-infrared spectroscopy (fNIRS) study. Front. Hum. Neurosci. 2017, 11, 431. [Google Scholar]
- Su, W.C.; Culotta, M.; Mueller, J.; Tsuzuki, D.; Pelphrey, K.; Bhat, A. Differences in cortical activation patterns during action observation, action execution, and interpersonal synchrony between children with or without autism spectrum disorder (ASD): An fNIRS pilot study. PLoS ONE 2020, 15, e0240301. [Google Scholar] [CrossRef] [PubMed]
- Su, W.C.; Culotta, M.L.; Hoffman, M.D.; Trost, S.L.; Pelphrey, K.A.; Tsuzuki, D.; Bhat, A.N. Developmental differences in cortical activation during action observation, action execution and interpersonal synchrony: An fNIRS study. Front. Hum. Neurosci. 2020, 14, 57. [Google Scholar] [CrossRef] [Green Version]
- Su, W.C.; Culotta, M.; Tsuzuki, D.; Bhat, A. Movement kinematics and cortical activation in children with and without autism spectrum disorder during sway synchrony tasks: An fNIRS study. Sci. Rep. 2021, 11, 15035. [Google Scholar] [CrossRef] [PubMed]
- Su, W.C.; Srinivasan, S.; Cleffi, C.; Bhat, A. Short report on research trends during the COVID-19 pandemic and use of telehealth interventions and remote brain research in children with autism spectrum disorder. Autism 2021, 25, 1816–1822. [Google Scholar] [CrossRef]
- Su, W.C.; Culotta, M.; Tsuzuki, D.; Bhat, A. Cortical activation during cooperative joint actions and competition in children with and without an autism spectrum condition (ASC): An fNIRS study. Sci. Rep. 2022, 12, 5177. [Google Scholar] [CrossRef]
- Su, W.C.; Amonkar, N.; Cleffi, C.; Srinivasan, S.; Bhat, A. Neural effects of physical activity and movement interventions in individuals with developmental disabilities: A systematic review. Front. Psychiatry 2022, 13, 794652. [Google Scholar] [CrossRef]
- Hollingshead, A.B. Four Factor Index of Social Status; Yale University: New Haven, CT, USA, 1975. [Google Scholar]
- Lord, C. Autism Diagnostic Observation Schedule, 2nd ed.; Western Psychological Services: Los Angeles, CA, USA, 2012. [Google Scholar]
- Rutter, M.; Bailey, A.; Lord, C. The Social Communication Questionnaire: Manual; Western Psychological Services: Los Angeles, CA, USA, 2003. [Google Scholar]
- Sparrow, S.S.; Cicchetti, D.V.; Balla, D.A. Vineland Adaptive Behavior Scales, 2nd ed.; NCS Pearson: San Antonio, DE, USA, 2005. [Google Scholar]
- Coren, S. The lateral preference inventory for measurement of handedness, footedness, eyedness, and earedness: Norms for young adults. Bull. Psychon. Soc. 1993, 31, 1–3. [Google Scholar] [CrossRef]
- Ayres, A.J. Sensory Integration and Praxis Tests; Western Psychological Publishers: Torrance, CA, USA, 1989. [Google Scholar]
- Klem, G.H.; Lüders, H.O.; Jasper, H.H.; Elger, C. The ten-twenty electrode system of the international federation. The international federation of clinical neurophysiology. Electroencephalogr. Clin. Neurophysiol. Suppl. 1999, 52, 3–6. [Google Scholar]
- Tsuzuki, D.; Cai, D.S.; Dan, H. Stable and convenient spatial registration of stand-alone NIRS data through anchor-based probabilistic registration. Neurosci. Res. 2012, 72, 163–171. [Google Scholar] [CrossRef]
- Okamoto, M.; Dan, H.; Sakamoto, K.; Takeo, K.; Shimizu, K.; Kohno, S.; Oda, I.; Isobe, S.; Suzuki, T.; Kohyama, K.; et al. Three-dimensional probabilistic anatomical cranio-cerebral correlation via the international 10–20 system oriented for transcranial functional brain mapping. NeuroImage 2004, 21, 99–111. [Google Scholar] [CrossRef]
- Shattuck, D.W.; Mirza, M.; Adisetiyo, V.; Hojatkashani, C.; Salamon, G.; Narr, K.L.; Poldrack, R.A.; Bilder, R.M.; Toga, A.W. Construction of a 3D probabilistic atlas of human cortical structures. NeuroImage 2008, 39, 1064–1080. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sutoko, S.; Sato, H.; Maki, A.; Kiguchi, M.; Hirabayashi, Y.; Atsumori, H.; Obata, A.; Funane, T.; Katura, T. Tutorial on platform for optical topography analysis tools. Neurophotonics 2016, 3, 010801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huppert, T.J.; Diamond, S.G.; Franceschini, M.A.; Boas, D.A. HomER: A review of time-series analysis methods for near-infrared spectroscopy of the brain. Appl. Opt. 2009, 48, 280–298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sato, H.; Fuchino, Y.; Kiguchi, M.; Katura, T.; Maki, A.; Yoro, T.; Koizumi, H. Intersubject variability of near-infrared spectroscopy signals during sensorimotor cortex activation. J. Biomed. Opt. 2005, 10, 44001. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.K.; Dan, I. Exploring the false discovery rate in multichannel NIRS. NeuroImage 2006, 33, 542–549. [Google Scholar] [CrossRef]
- Siaperas, P.; Ring, H.A.; McAllister, C.J.; Henderson, S.; Barnett, A.; Watson, P.; Holland, A.J. Atypical movement performance and sensory integration in Asperger’s syndrome. J. Autism Dev. Disord. 2012, 42, 718–725. [Google Scholar] [CrossRef]
- Salowitz, N.M.; Eccarius, P.; Karst, J.; Carson, A.; Schohl, K.; Stevens, S.; Van Hecke, A.V.; Scheidt, R.A. Brief report: Visuo-spatial guidance of movement during gesture imitation and mirror drawing in children with autism spectrum disorders. J. Autism Dev. Disord. 2013, 43, 985–995. [Google Scholar] [CrossRef] [Green Version]
- Morimoto, C.; Hida, E.; Shima, K.; Okamura, H. Temporal processing instability with millisecond accuracy is a cardinal feature of sensorimotor impairments in autism spectrum disorder: Analysis using the synchronized finger-tapping task. J. Autism Dev. Disord. 2018, 48, 351–360. [Google Scholar] [CrossRef] [Green Version]
- Franich, K.; Wong, H.Y.; Yu, A.; To, C. Temporal coordination and prosodic structure in autism spectrum disorder: Timing across speech and non-speech motor domains. J. Autism Dev. Disord. 2021, 51, 2929–2949. [Google Scholar] [CrossRef]
- Haaland, K.Y.; Harrington, D.L.; Knight, R.T. Neural representations of skilled movement. Brain 2000, 123, 2306–2313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Floris, D.L.; Howells, H. Atypical structural and functional motor networks in autism. Prog. Brain Res. 2018, 238, 207–248. [Google Scholar] [PubMed]
- Limanowski, J.; Sarasso, P.; Blankenburg, F. Different responses of the right superior temporal sulcus to visual movement feedback during self-generated vs. externally generated hand movements. Eur. J. Neurosci. 2018, 47, 314–320. [Google Scholar] [CrossRef]
- Amonkar, N.; Su, W.C.; Bhat, A.N.; Srinivasan, S.M. Effects of creative movement therapies on social communication, behavioral-affective, sensorimotor, cognitive, and functional participation skills of individuals with Autism Spectrum Disorder: A systematic review. Front. Psychiatry 2021, 12, 722874. [Google Scholar] [CrossRef]
- Afif, I.Y.; Manik, A.R.; Munthe, K.; Maula, M.I.; Ammarullah, M.I.; Jamari, J.; Winarni, T.I. Physiological effect of deep pressure in reducing anxiety of children with ASD during traveling: A public transportatiov n setting. Bioengineering 2022, 9, 157. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | ASD Group (N = 14) Mean ± SE | TD Group (N = 17) Mean ± SE |
|---|---|---|
| Age | 11.14 ± 0.95 | 10.82 ± 0.69 |
| Sex | 9M, 5F | 11M, 6F |
| Ethnicity | 10C, 2 A, 1 AC, 1BC | 13 C; 1A; 1AI; 2AC |
| SES-Child | 64.57 ± 5.35 | 69.71 ± 4.43 |
| Coren’s handedness score | Right-handed 33.36 ± 1.59 | Right-handed 33.41 ± 1.78 |
| SCQ score | 23.50 ± 3.27 | - |
| ADOS total Score Social affect Repetitive behavior | 18.17 ± 1.84 13.17 ± 1.42 5.00 ± 0.62 | - |
| VABS-II (SS) | 72.50 ± 3.07 * | 110.41 ± 2.91 |
| IQ | 85.64 ± 7.04 * | 114.18 ± 1.71 |
| BOT-2 manual dexterity | 19.93 ± 2.04 * | 27.71 ± 1.08 |
| R-Values | SIPT-PP Total Error | SIPT-BMC Total Error | ||||
|---|---|---|---|---|---|---|
| H | P | M | H | P | M | |
| TD | ||||||
| Left hemisphere | ||||||
| MIFG | −0.283 ** | −0.296 ** | −0.180 | −0.452 ** | −0.426 ** | −0.329 ** |
| MSTG | 0.002 | −0.157 | 0.311 ** | −0.091 | −0.296 ** | 0.150 |
| IPL | 0.063 | −0.093 | −0.042 | −0.337 ** | −0.336 ** | −0.390 ** |
| Right hemisphere | ||||||
| MIFG | 0.025 | 0.064 | 0.019 | −0.122 | −0.225 * | −0.210 * |
| MSTG | 0.111 | 0.098 | 0.192 * | −0.112 | −0.103 | 0.030 |
| IPL | 0.129 | 0.087 | 0.304 ** | −0.132 | −0.169 | −0.058 |
| ASD | ||||||
| Left hemisphere | ||||||
| MIFG | −0.257 * | 0.222 * | 0.287 ** | −0.072 | −0.104 | −0.010 |
| MSTG | 0.212 * | 0.179 | 0.216 * | −0.032 | 0.317 ** | 0.332 ** |
| IPL | 0.160 | 0.095 | 0.190 | 0.105 | 0.213 * | 0.330 ** |
| Right hemisphere | ||||||
| MIFG | 0.434 ** | 0.288 ** | 0.283 ** | 0.186 | 0.153 | 0.049 |
| MSTG | 0.094 | 0.131 | 0.047 | −0.130 | 0.002 | −0.061 |
| IPL | 0.282 ** | 0.235 * | 0.368 ** | 0.432 ** | 0.180 | 0.543 ** |
| R-Values | ADOS-SA | ADOS-RRB | ADOS-Total | ||||||
|---|---|---|---|---|---|---|---|---|---|
| H | P | M | H | P | M | H | P | M | |
| ASD | |||||||||
| Left hemisphere | |||||||||
| MIFG | 0.055 | 0.001 | 0.096 | −0.027 | −0.170 | 0.085 | 0.033 | −0.056 | 0.103 |
| MSTG | −0.132 | 0.310 ** | 0.209 | −0.004 | 0.360 ** | 0.341 ** | −0.103 | 0.361 ** | 0.277 * |
| IPL | 0.290 * | −0.162 | 0.062 | 0.352 ** | −0.068 | 0.237 * | 0.343 ** | −0.149 | 0.128 |
| Right hemisphere | |||||||||
| MIFG | 0.063 | 0.172 | 0.252 * | 0.311 ** | 0.204 | 0.288 * | 0.153 | 0.202 | 0.293 * |
| MSTG | −0.097 | 0.111 | 0.205 | −0.139 | 0.067 | −0.013 | −0.122 | 0.109 | 0.155 |
| IPL | 0.045 | 0.214 | 0.072 | 0.217 | 0.142 | 0.330 ** | 0.108 | 0.214 | 0.167 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Su, W.-C.; Culotta, M.; Mueller, J.; Tsuzuki, D.; Bhat, A. fNIRS-Based Differences in Cortical Activation during Tool Use, Pantomimed Actions, and Meaningless Actions between Children with and without Autism Spectrum Disorder (ASD). Brain Sci. 2023, 13, 876. https://doi.org/10.3390/brainsci13060876
Su W-C, Culotta M, Mueller J, Tsuzuki D, Bhat A. fNIRS-Based Differences in Cortical Activation during Tool Use, Pantomimed Actions, and Meaningless Actions between Children with and without Autism Spectrum Disorder (ASD). Brain Sciences. 2023; 13(6):876. https://doi.org/10.3390/brainsci13060876
Chicago/Turabian StyleSu, Wan-Chun, McKenzie Culotta, Jessica Mueller, Daisuke Tsuzuki, and Anjana Bhat. 2023. "fNIRS-Based Differences in Cortical Activation during Tool Use, Pantomimed Actions, and Meaningless Actions between Children with and without Autism Spectrum Disorder (ASD)" Brain Sciences 13, no. 6: 876. https://doi.org/10.3390/brainsci13060876