
Fine-Tuning the PI3K/Akt Signaling Pathway Intensity by Sex and Genotype-Load: Sex-Dependent Homozygotic Threshold for Somatic Growth but Feminization of Anxious Phenotype in Middle-Aged PDK1 K465E Knock-In and Heterozygous Mice



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Generation of PDK1K465E/K465E Mice and Genotyping Analysis
2.2. Animals
2.3. Behavioral Assessments
2.4. Statistics
3. Results
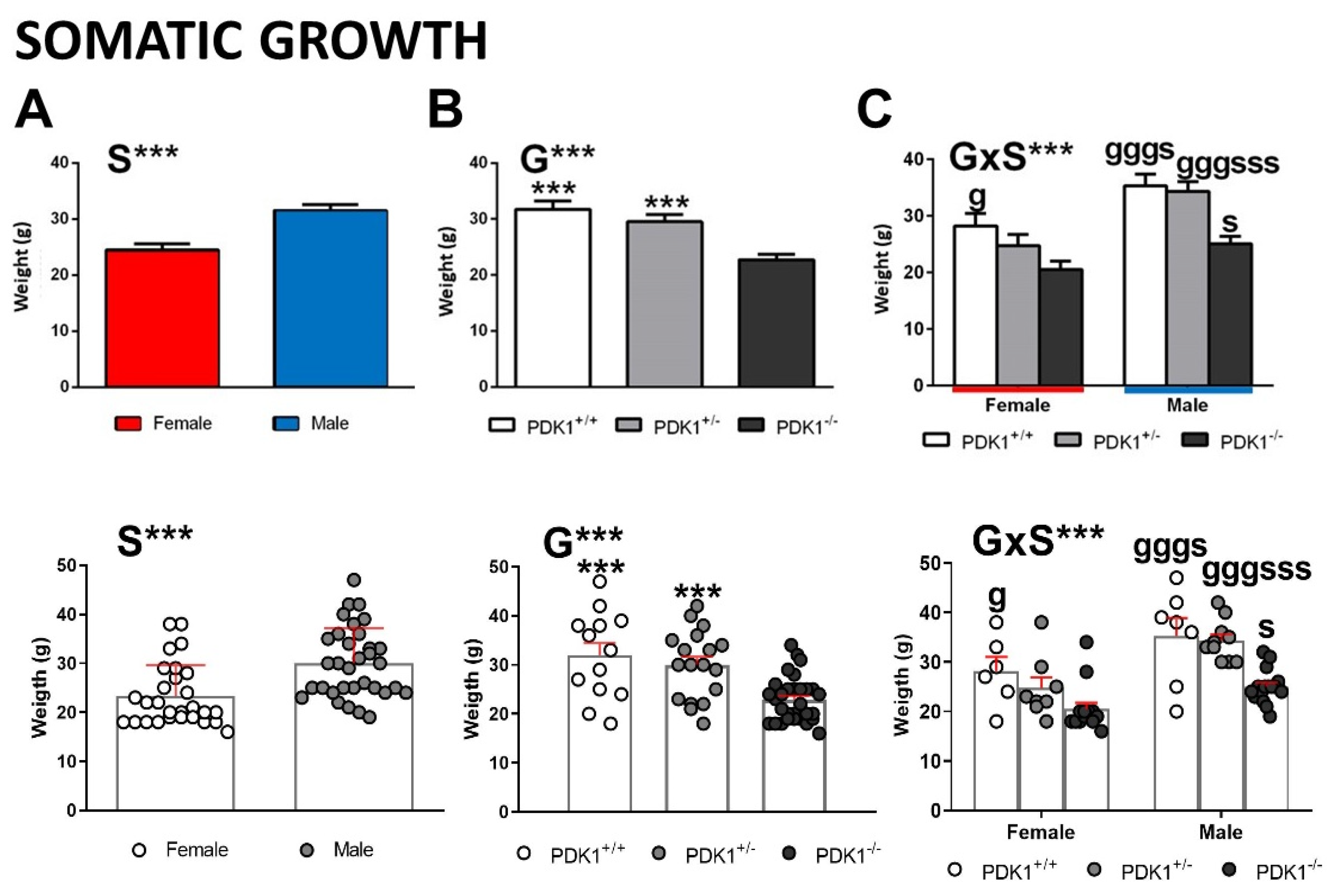
3.1. Somatic Growth/Bodyweight
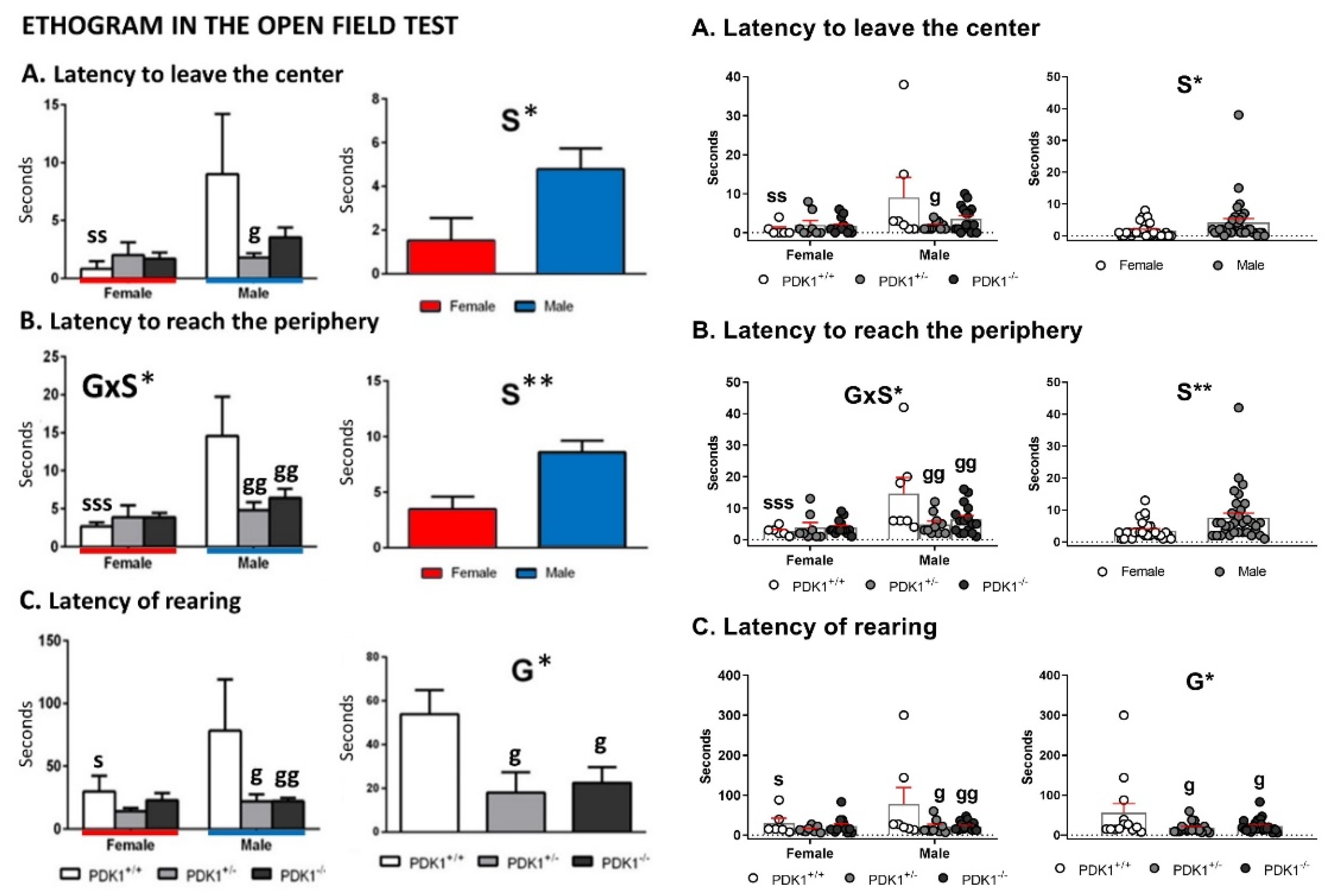
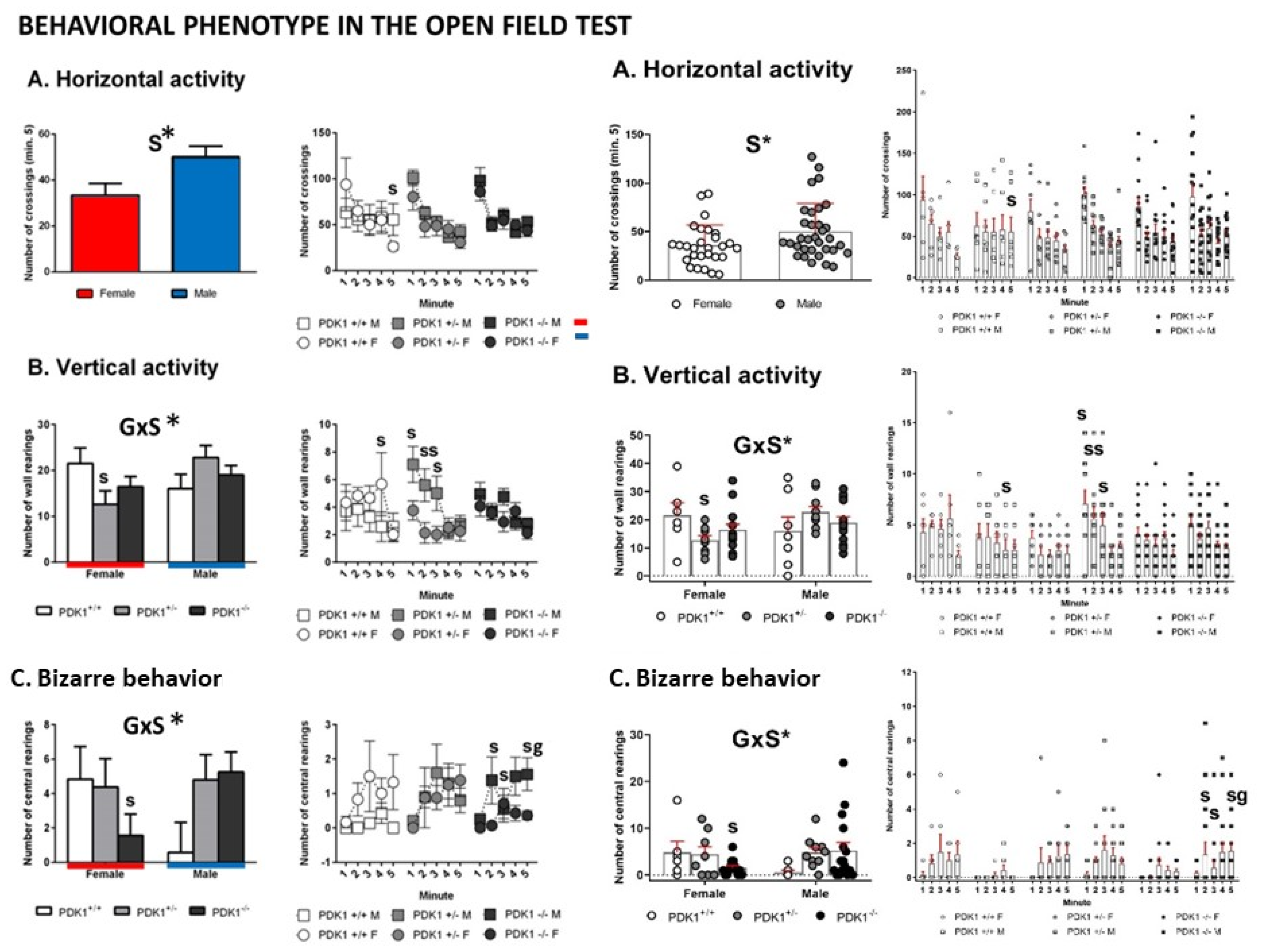
3.2. Open Field Test
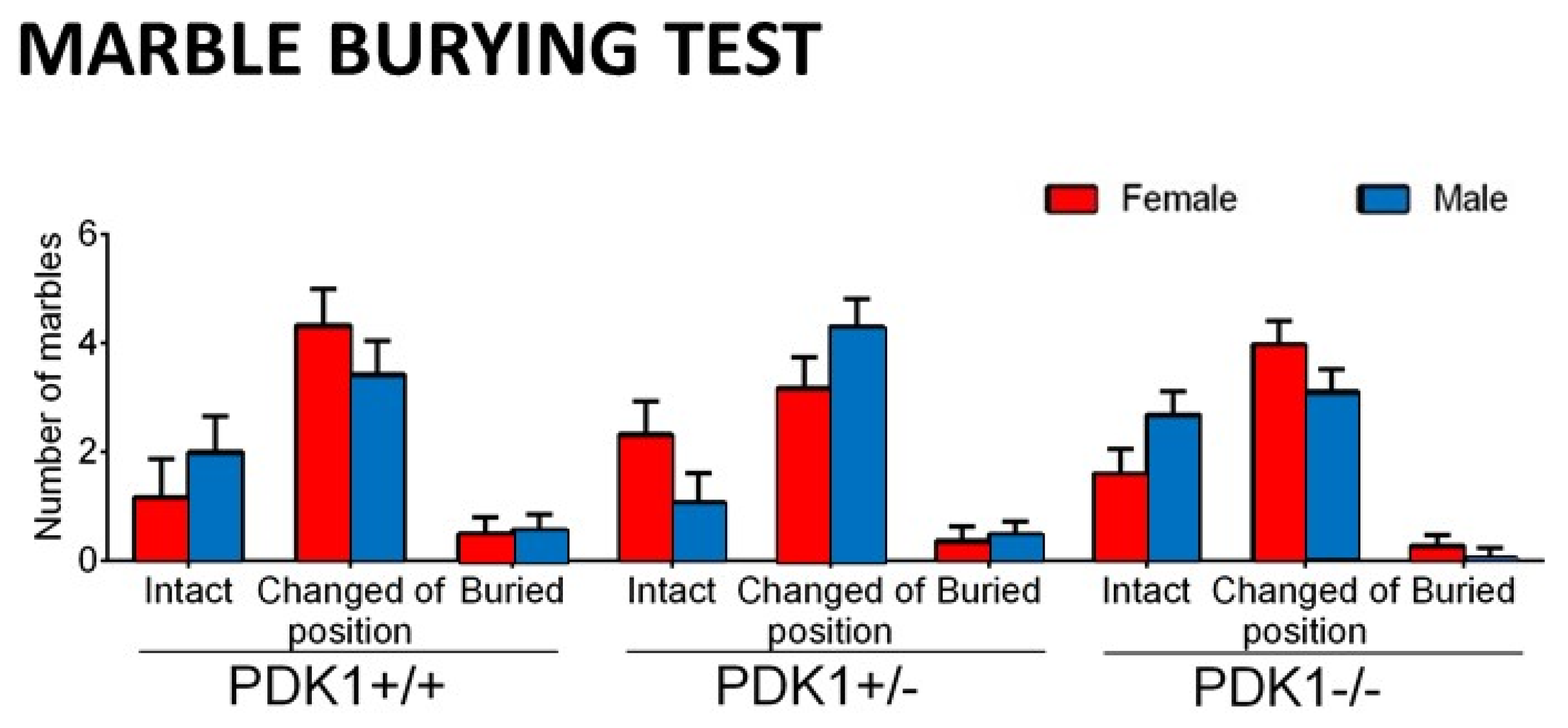
3.3. Marble Burying Test
4. Discussion
4.1. Homozygous-Load of the Mutant Gene Is Needed for the Reduced Somatic Growth Effects Only in Mutant Males
4.2. Similar Neophobia and Locomotion, but Increased Anxiety-Like Phenotype in Mutant Females and Feminized Anxious-Like Phenotype of Mutant Males
4.3. Performance of Mutants Can Resemble Normal under Naturalistic Anxiogenic Conditions
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lenze, E.J.; Wetherell, J.L. A lifespan view of anxiety disorders. Dialogues Clin. Neurosci. 2011, 13, 381–399. [Google Scholar] [CrossRef]
- National Institute of Mental Health. Research Domain Criteria. (Internet) NIHM. Available online: https://www.nimh.nih.gov/research/research-funded-by-nimh/rdoc/index.shtml (accessed on 29 February 2020).
- Asher, J. Genes and Circuitry, Not Just Clinical Observation, to Guide Classification for Research (Internet). NIMH; 2010. Available online: https://www.nimh.nih.gov/news/science-news/2010/genes-and-circuitry-not-just-clinical-observation-to-guide-classification-for-research.shtml (accessed on 29 February 2020).
- Cuthbert, B.N.; Insel, T.R. Toward the future of psychiatric diagnosis: The seven pillars of RDoC. BMC Med. 2013, 11, 126. [Google Scholar] [CrossRef] [Green Version]
- Savage, J.E.; Sawyers, C.; Roberson-Nay, R.; Hettema, J.M. The genetics of anxiety-related negative valence system traits. Am. J. Med. Genet. Part B Neuropsychiatr. Genet. 2017, 174, 156–177. [Google Scholar] [CrossRef] [Green Version]
- Prendergast, B.J.; Onishi, K.G.; Zucker, I. Female mice liberated for inclusion in neuroscience and biomedical research. Neurosci. Biobehav. Rev. 2014, 40, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, S.J.; Scheibye-Knudsen, M.; Longo, D.L.; de Cabo, R. Animal Models of Aging Research: Implications for Human Aging and Age-Related Diseases. Annu. Rev. Anim. Biosci. 2015, 3, 283–303. [Google Scholar] [CrossRef] [Green Version]
- Ackermann, T.F.; Hörtnagl, H.; Wolfer, D.P.; Colacicco, G.; Sohr, R.; Lang, F.; Hellweg, R.; Lang, U.E. Phosphatidylinositide Dependent Kinase Deficiency Increases Anxiety and Decreases GABA and Serotonin Abundance in the Amygdala. Cell. Physiol. Biochem. 2008, 22, 735–744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leibrock, C.; Ackermann, T.F.; Hierlmeier, M.; Lang, F.; Borgwardt, S.; Lang, U.E. Akt2 Deficiency is Associated with Anxiety and Depressive Behavior in Mice. Cell. Physiol. Biochem. 2013, 32, 766–777. [Google Scholar] [CrossRef] [PubMed]
- Cordón-Barris, L.; Pascual-Guiral, S.; Yang, S.; Giménez-Llort, L.; Lope-Piedrafita, S.; Niemeyer, C.; Claro, E.; Lizcano, J.M.; Bayascas, J.R. Mutation of the 3-Phosphoinositide-Dependent Protein Kinase 1 (PDK1) Substrate-Docking Site in the Developing Brain Causes Microcephaly with Abnormal Brain Morphogenesis Independently of Akt, Leading to Impaired Cognition and Disruptive Behaviors. Mol. Cell. Biol. 2016, 36, 2967–2982. [Google Scholar] [CrossRef] [Green Version]
- Bergeron, Y.; Bureau, G.; Laurier-Laurin, M.-É.; Asselin, E.; Massicotte, G.; Cyr, M. Genetic Deletion of Akt3 Induces an Endophenotype Reminiscent of Psychiatric Manifestations in Mice. Front. Mol. Neurosci. 2017, 10, 102. [Google Scholar] [CrossRef] [PubMed]
- Alessi, D.R.; James, S.R.; Downes, C.; Holmes, A.B.; Gaffney, P.R.; Reese, C.B.; Cohen, P. Characterization of a 3-phosphoinositide-dependent protein kinase which phosphorylates and activates protein kinase Bα. Curr. Biol. 1997, 7, 261–269. [Google Scholar] [CrossRef] [Green Version]
- Mora, A.; Komander, D.; van Aalten, D.M.; Alessi, D.R. PDK1, the master regulator of AGC kinase signal transduction. Semin. Cell Dev. Biol. 2004, 15, 161–170. [Google Scholar] [CrossRef] [Green Version]
- Pearce, L.R.; Komander, D.; Alessi, D. The nuts and bolts of AGC protein kinases. Nat. Rev. Mol. Cell Biol. 2010, 11, 9–22. [Google Scholar] [CrossRef] [PubMed]
- Waite, K.; Eickholt, B.J. The Neurodevelopmental Implications of PI3K Signaling. Curr. Top. Microbiol. Immunol. 2010, 346, 245–265. [Google Scholar] [CrossRef]
- Bayascas, J.R. PDK1: The Major Transducer of PI 3-Kinase Actions. Curr. Top. Microbiol. Immunol. 2010, 346, 9–29. [Google Scholar] [CrossRef] [PubMed]
- Jope, R.S.; Roh, R.S. Glycogen Synthase Kinase-3 (GSK3) in Psychiatric Diseases and Therapeutic Interventions. Curr. Drug Targets 2006, 7, 1421–1434. [Google Scholar] [CrossRef]
- Beaulieu, J.-M.; Gainetdinov, R.; Caron, M.G. Akt/GSK3 Signaling in the Action of Psychotropic Drugs. Annu. Rev. Pharmacol. Toxicol. 2009, 49, 327–347. [Google Scholar] [CrossRef] [Green Version]
- Freyberg, Z.; Ferrando, S.J.; Javitch, J.A. Roles of the Akt/GSK-3 and Wnt Signaling Pathways in Schizophrenia and Antipsychotic Drug Action. Am. J. Psychiatry 2010, 167, 388–396. [Google Scholar] [CrossRef] [Green Version]
- Jope, R.S. Glycogen Synthase Kinase-3 in the Etiology and Treatment of Mood Disorders. Front. Mol. Neurosci. 2011, 4, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sachs, B.D.; Rodriguiz, R.M.; Siesser, W.B.; Kenan, A.; Royer, E.L.; Jacobsen, J.P.R.; Wetsel, W.C.; Caron, M.G. The effects of brain serotonin deficiency on behavioural disinhibition and anxiety-like behaviour following mild early life stress. Int. J. Neuropsychopharmacol. 2013, 16, 2081–2094. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, K. An emerging role for Wnt and GSK3 signaling pathways in schizophrenia. Clin. Genet. 2013, 83, 511–517. [Google Scholar] [CrossRef]
- Beurel, E.; Grieco, S.F.; Jope, R.S. Glycogen synthase kinase-3 (GSK3): Regulation, actions, and diseases. Pharmacol. Ther. 2015, 148, 114–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, H.-S.; Zhu, W.-L.; Liu, J.; Luo, Y.-X.; Si, J.-J.; Wang, S.-J.; Xue, Y.-X.; Ding, Z.-B.; Shi, J.; Lu, L. PI3K/Akt Signaling Pathway in the Basolateral Amygdala Mediates the Rapid Antidepressant-like Effects of Trefoil Factor 3. Neuropsychopharmacology 2012, 37, 2671–2683. [Google Scholar] [CrossRef]
- Wang, W.; Lu, Y.; Xue, Z.; Li, C.; Wang, C.; Zhao, X.; Zhang, J.; Wei, X.; Chen, X.; Cui, W.; et al. Rapid-acting antidepressant-like effects of acetyl-l-carnitine mediated by PI3K/AKT/BDNF/VGF signaling pathway in mice. Neuroscience 2015, 285, 281–291. [Google Scholar] [CrossRef] [PubMed]
- Tao, W.; Dong, Y.; Su, Q.; Wang, H.; Chen, Y.; Xue, W.; Chen, C.; Xia, B.; Duan, J.; Chen, G. Liquiritigenin reverses depression-like behavior in unpredictable chronic mild stress-induced mice by regulating PI3K/Akt/mTOR mediated BDNF/TrkB pathway. Behav. Brain Res. 2016, 308, 177–186. [Google Scholar] [CrossRef] [PubMed]
- Dwivedi, Y.; Rizavi, H.S.; Zhang, H.; Roberts, R.C.; Conley, R.R.; Pandey, G.N. Modulation in Activation and Expression of Phosphatase and Tensin Homolog on Chromosome Ten, Akt1, and 3-Phosphoinositide-Dependent Kinase 1: Further Evidence Demonstrating Altered Phosphoinositide 3-Kinase Signaling in Postmortem Brain of Suicide Subjects. Biol. Psychiatry 2010, 67, 1017–1025. [Google Scholar] [CrossRef] [Green Version]
- Neasta, J.; Ben Hamida, S.; Yowell, Q.V.; Carnicella, S.; Ron, D. AKT Signaling Pathway in the Nucleus Accumbens Mediates Excessive Alcohol Drinking Behaviors. Biol. Psychiatry 2011, 70, 575–582. [Google Scholar] [CrossRef] [Green Version]
- Dahlhoff, M.; Siegmund, A.; Golub, Y.; Wolf, E.; Holsboer, F.; Wotjak, C. AKT/GSK-3β/β-catenin signalling within hippocampus and amygdala reflects genetically determined differences in posttraumatic stress disorder like symptoms. Neuroscience 2010, 169, 1216–1226. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Cui, K.; Xing, F.; Liu, X. Akt dependent adult hippocampal neurogenesis regulates the behavioral improvement of treadmill running to mice model of post-traumatic stress disorder. Behav. Brain Res. 2020, 379, 112375. [Google Scholar] [CrossRef]
- Huang, H.; Wang, Q.; Guan, X.; Zhang, X.; Zhang, Y.; Cao, J.; Li, X. Effects of enriched environment on depression and anxiety-like behavior induced by early life stress: A comparison between different periods. Behav. Brain Res. 2021, 411, 113389. [Google Scholar] [CrossRef]
- Komander, D.; Fairservice, A.; Deak, M.; Kular, G.S.; Prescott, A.; Downes, C.P.; Safrany, S.; Alessi, D.; Van Aalten, D.M.F. Structural insights into the regulation of PDK1 by phosphoinositides and inositol phosphates. EMBO J. 2004, 23, 3918–3928. [Google Scholar] [CrossRef] [Green Version]
- Bayascas, J.R.; Wullschleger, S.; Sakamoto, K.; García-Martínez, J.M.; Clacher, C.; Komander, D.; van Aalten, D.M.F.; Boini, K.M.; Lang, F.; Lipina, C.; et al. Mutation of the PDK1 PH Domain Inhibits Protein Kinase B/Akt, Leading to Small Size and Insulin Resistance. Mol. Cell. Biol. 2008, 28, 3258–3272. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.; Pascual-Guiral, S.; Ponce, R.; Giménez-Llort, L.; Baltrons, M.A.; Arancio, O.; Palacio, J.R.; Clos, V.M.; Yuste, V.J.; Bayascas, J.R. Reducing the Levels of Akt Activation by PDK1 Knock-in Mutation Protects Neuronal Cultures against Synthetic Amyloid-Beta Peptides. Front. Aging Neurosci. 2018, 9, 435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zurashvili, T.; Cordón-Barris, L.; Ruiz-Babot, G.; Zhou, X.; Lizcano, J.M.; Gómez, N.; Giménez-Llort, L.; Bayascas, J.R. Interaction of PDK1 with Phosphoinositides Is Essential for Neuronal Differentiation but Dispensable for Neuronal Survival. Mol. Cell. Biol. 2013, 33, 1027–1040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giménez-Llort, L.; Santana-Santana, M.; Bayascas, J.R. The Impact of the PI3K/Akt Signaling Pathway in Anxiety and Working Memory in Young and Middle-Aged PDK1 K465E Knock-In Mice. Front. Behav. Neurosci. 2020, 14, 61. [Google Scholar] [CrossRef] [PubMed]
- McLean, C.P.; Asnaani, A.; Litz, B.T.; Hofmann, S.G. Gender differences in anxiety disorders: Prevalence, course of illness, comorbidity and burden of illness. J. Psychiatr. Res. 2011, 45, 1027–1035. [Google Scholar] [CrossRef] [Green Version]
- Zucker, I.; Beery, A. Males still dominate animal studies. Nat. Cell Biol. 2010, 465, 690. [Google Scholar] [CrossRef]
- Beery, A.; Zucker, I. Sex bias in neuroscience and biomedical research. Neurosci. Biobehav. Rev. 2011, 35, 565–572. [Google Scholar] [CrossRef] [Green Version]
- Wong, H.; Levenga, J.; Laplante, L.; Keller, B.; Cooper-Sansone, A.; Borski, C.; Milstead, R.; Ehringer, M.; Hoeffer, C. Isoform-specific roles for AKT in affective behavior, spatial memory, and extinction related to psychiatric disorders. eLife 2020, 9, 56630. [Google Scholar] [CrossRef]
- Kilkenny, C.; Browne, W.J.; Cuthill, I.C.; Emerson, M.; Altman, D.G. Improving Bioscience Research Reporting: The ARRIVE Guidelines for Reporting Animal Research. PLoS Biol. 2010, 8, e1000412. [Google Scholar] [CrossRef]
- Hall, C.S.; Ballachey, E.L. A study of the rat’s behavior in a field: A contribution to method in comparative psychology. Univ. Calif. Publ. Psychol. 1932, 6, 1–12. [Google Scholar]
- Deacon, R.M.J. Digging and marble burying in mice: Simple methods for in vivo identification of biological impacts. Nat. Protoc. 2006, 1, 122–124. [Google Scholar] [CrossRef]
- Baeta-Corral, R.; Giménez-Llort, L. Bizarre behaviors and risk assessment in 3xTg-AD mice at early stages of the disease. Behav. Brain Res. 2014, 258, 97–105. [Google Scholar] [CrossRef] [PubMed]
- Torres-Lista, V.; López-Pousa, S.; Giménez-Llort, L. Marble-burying is enhanced in 3xTg-AD mice, can be reversed by risperidone and it is modulable by handling. Behav. Process. 2015, 116, 69–74. [Google Scholar] [CrossRef] [PubMed]
- Castillo-Mariqueo, L.; Giménez-Llort, L. Translational Modeling of Psychomotor Function in Normal and AD-Pathological Aging With Special Concerns on the Effects of Social Isolation. Front. Aging 2021, 2, 5. [Google Scholar] [CrossRef]
- Gimenez-Llort, L.; Alveal-Mellado, D. Digging Signatures in 13-Month-Old 3xTg-AD Mice for Alzheimer’s Disease and Its Disruption by Isolation Despite Social Life Since They Were Born. Front. Behav. Neurosci. 2021, 14, 611384. [Google Scholar] [CrossRef]
- Muntsant, A.; Giménez-Llort, L. Impact of Social Isolation on the Behavioral, Functional Profiles, and Hippocampal Atrophy Asymmetry in Dementia in Times of Coronavirus Pandemic (COVID-19): A Translational Neuroscience Approach. Front. Psychiatry 2020, 11, 572583. [Google Scholar] [CrossRef]
- Pinel, J.P.; Treit, D. Burying as a defensive response in rats. J. Comp. Physiol. Psychol. 1978, 92, 708–712. [Google Scholar] [CrossRef] [Green Version]
- Gyertyán, I. Analysis of the marble burying response: Marbles serve to measure digging rather than evoke burying. Behav. Pharmacol. 1995, 6, 24–31. [Google Scholar]
- De Brouwer, G.; Fick, A.; Harvey, B.H.; Wolmarans, D.W. A critical inquiry into marble-burying as a preclinical screening paradigm of relevance for anxiety and obsessive–compulsive disorder: Mapping the way forward. Cogn. Affect. Behav. Neurosci. 2019, 19, 1–39. [Google Scholar] [CrossRef] [Green Version]
- Mahmood, H.M.; Aldhalaan, H.M.; Alshammari, T.K.; Alqasem, M.A.; Alshammari, M.A.; Albekairi, N.A.; AlSharari, S.D. The Role of Nicotinic Receptors in the Attenuation of Autism-Related Behaviors in a Murine BTBR T + tf/J Autistic Model. Autism Res. 2020, 13, 1311–1334. [Google Scholar] [CrossRef]
- Broekkamp, C.L.; Rijk, H.W.; Joly-Gelouin, D.; Lloyd, K.L. Major tranquillizers can be distinguished from minor tranquillizers on the basis of effects on marble burying and swim-induced grooming in mice. Eur. J. Pharmacol. 1986, 126, 223–229. [Google Scholar] [CrossRef]
- Deacon, R.M.; Bannerman, D.; Kirby, B.; Croucher, A.; Rawlins, J.P. Effects of cytotoxic hippocampal lesions in mice on a cognitive test battery. Behav. Brain Res. 2002, 133, 57–68. [Google Scholar] [CrossRef]
- Greene-Schloesser, D.M.; Van Der Zee, E.A.; Sheppard, D.K.; Castillo, M.R.; Gregg, K.A.; Burrow, T.; Foltz, H.; Slater, M.; Bult-Ito, A. Predictive validity of a non-induced mouse model of compulsive-like behavior. Behav. Brain Res. 2011, 221, 55–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitra, S.; Bastos, C.; Chesworth, S.; Frye, C.; Bult-Ito, A. Strain and sex based characterization of behavioral expressions in non-induced compulsive-like mice. Physiol. Behav. 2017, 168, 103–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dixit, P.V.; Sahu, R.; Mishra, D.K. Marble-burying behavior test as a murine model of compulsive-like behavior. J. Pharmacol. Toxicol. Methods 2020, 102, 106676. [Google Scholar] [CrossRef] [PubMed]
- Jimenez-Gomez, C.; Osentoski, A.; Woods, J.H. Pharmacological evaluation of the adequacy of marble burying as an animal model of compulsion and/or anxiety. Behav. Pharmacol. 2011, 22, 711–713. [Google Scholar] [CrossRef] [PubMed] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santana-Santana, M.; Bayascas, J.-R.; Giménez-Llort, L. Fine-Tuning the PI3K/Akt Signaling Pathway Intensity by Sex and Genotype-Load: Sex-Dependent Homozygotic Threshold for Somatic Growth but Feminization of Anxious Phenotype in Middle-Aged PDK1 K465E Knock-In and Heterozygous Mice. Biomedicines 2021, 9, 747. https://doi.org/10.3390/biomedicines9070747
Santana-Santana M, Bayascas J-R, Giménez-Llort L. Fine-Tuning the PI3K/Akt Signaling Pathway Intensity by Sex and Genotype-Load: Sex-Dependent Homozygotic Threshold for Somatic Growth but Feminization of Anxious Phenotype in Middle-Aged PDK1 K465E Knock-In and Heterozygous Mice. Biomedicines. 2021; 9(7):747. https://doi.org/10.3390/biomedicines9070747
Chicago/Turabian StyleSantana-Santana, Mikel, José-Ramón Bayascas, and Lydia Giménez-Llort. 2021. "Fine-Tuning the PI3K/Akt Signaling Pathway Intensity by Sex and Genotype-Load: Sex-Dependent Homozygotic Threshold for Somatic Growth but Feminization of Anxious Phenotype in Middle-Aged PDK1 K465E Knock-In and Heterozygous Mice" Biomedicines 9, no. 7: 747. https://doi.org/10.3390/biomedicines9070747