
Impacts of Reducing Protein Content in Milk Replacer on Growth Performance and Health of Young Calves

 , , ,
, , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals, Housing, and Zootechnical Measures
2.2. Diets, Feeding Concept, and Experimental Design
2.3. Measurement and Sampling Procedure
2.4. Analytical Methods
2.5. Statistical Analysis
3. Results
3.1. Feed Intake
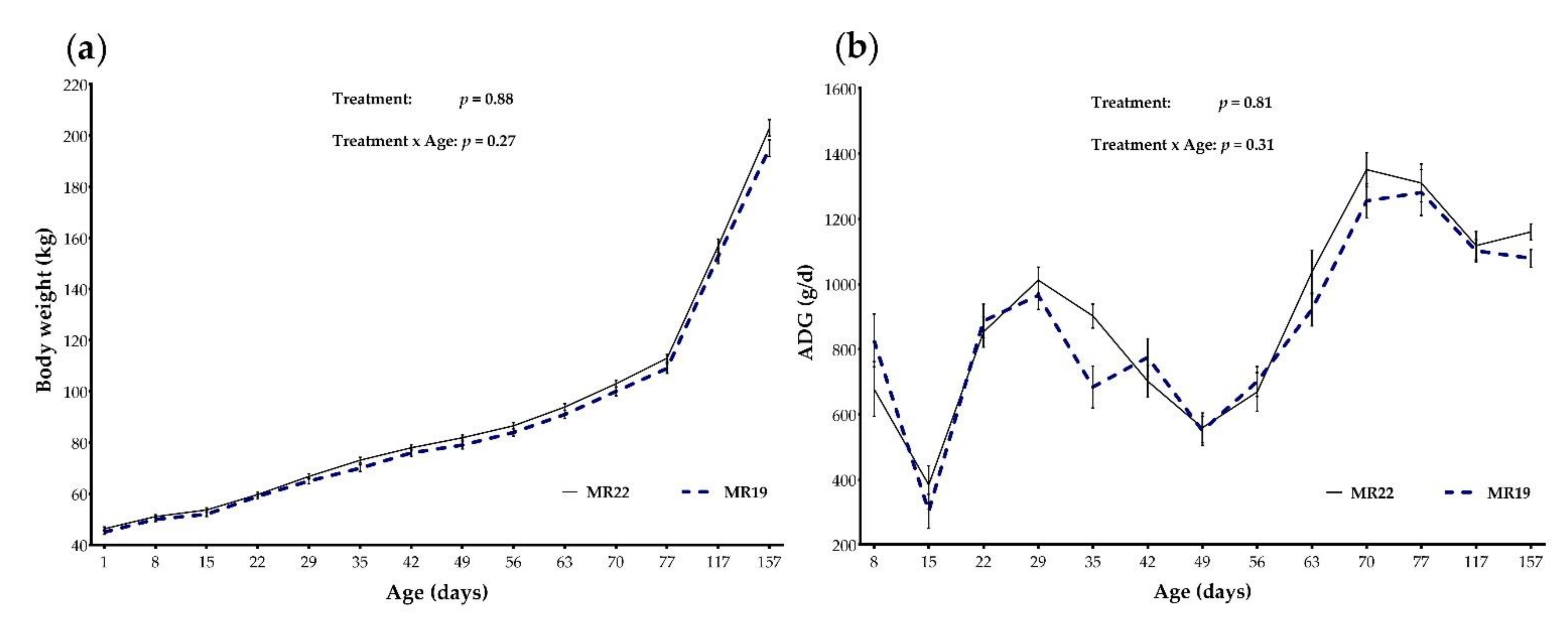
3.2. Body Weight Gain and Sacral Height
3.3. Rectal Temperature
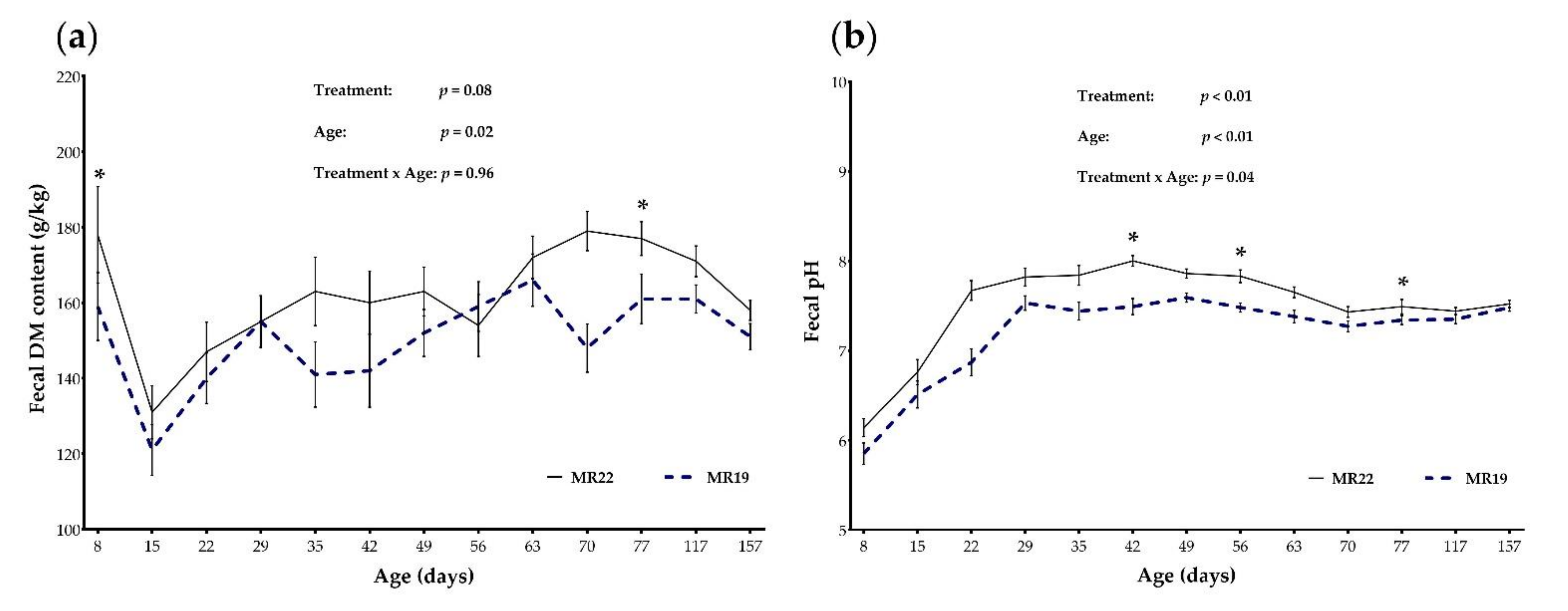
3.4. Fecal Parameters
3.4.1. Characteristics of Fecal Samples
3.4.2. DM Content and pH of Fecal Samples
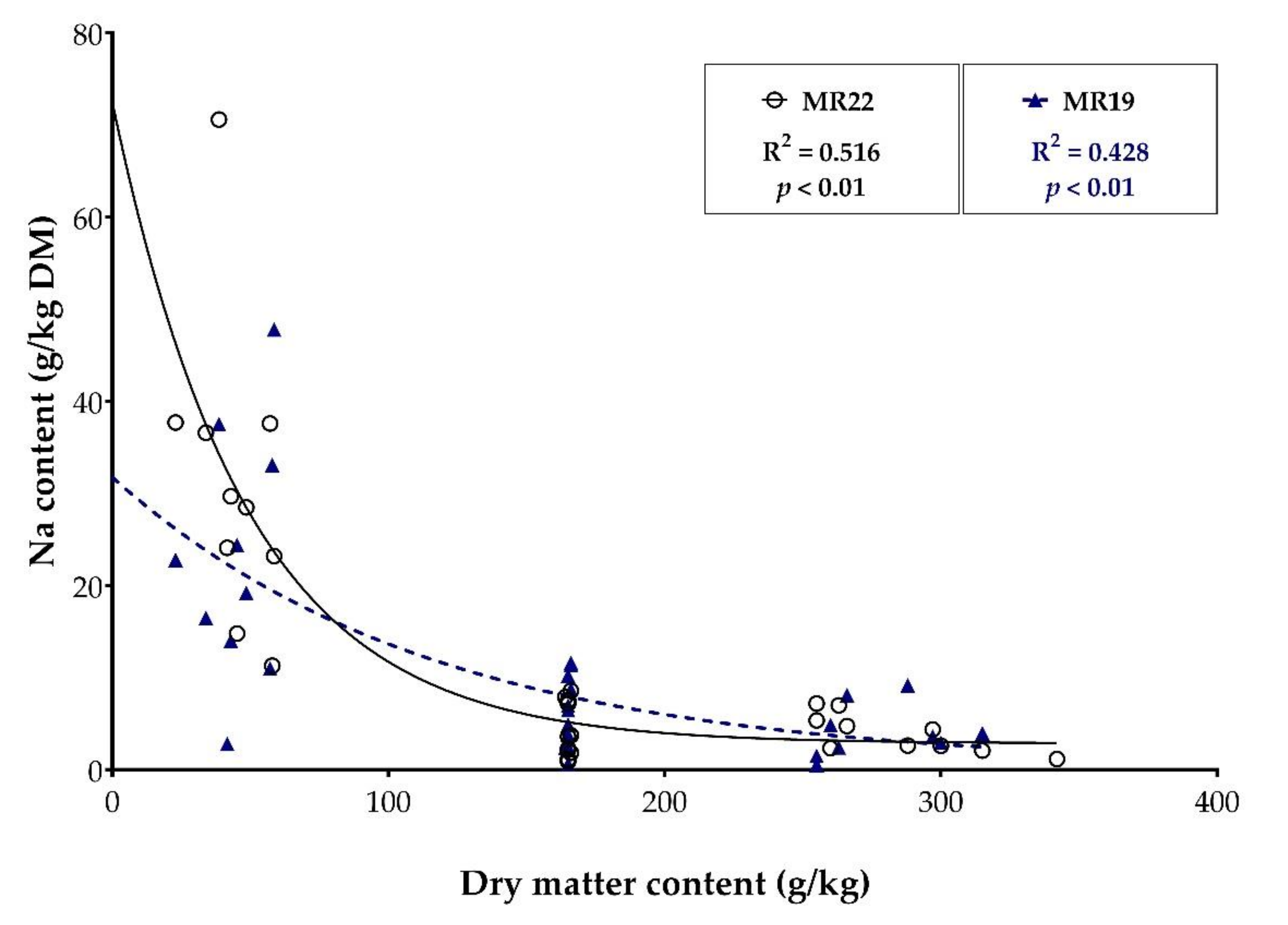
3.4.3. Sodium Content of Fecal Samples
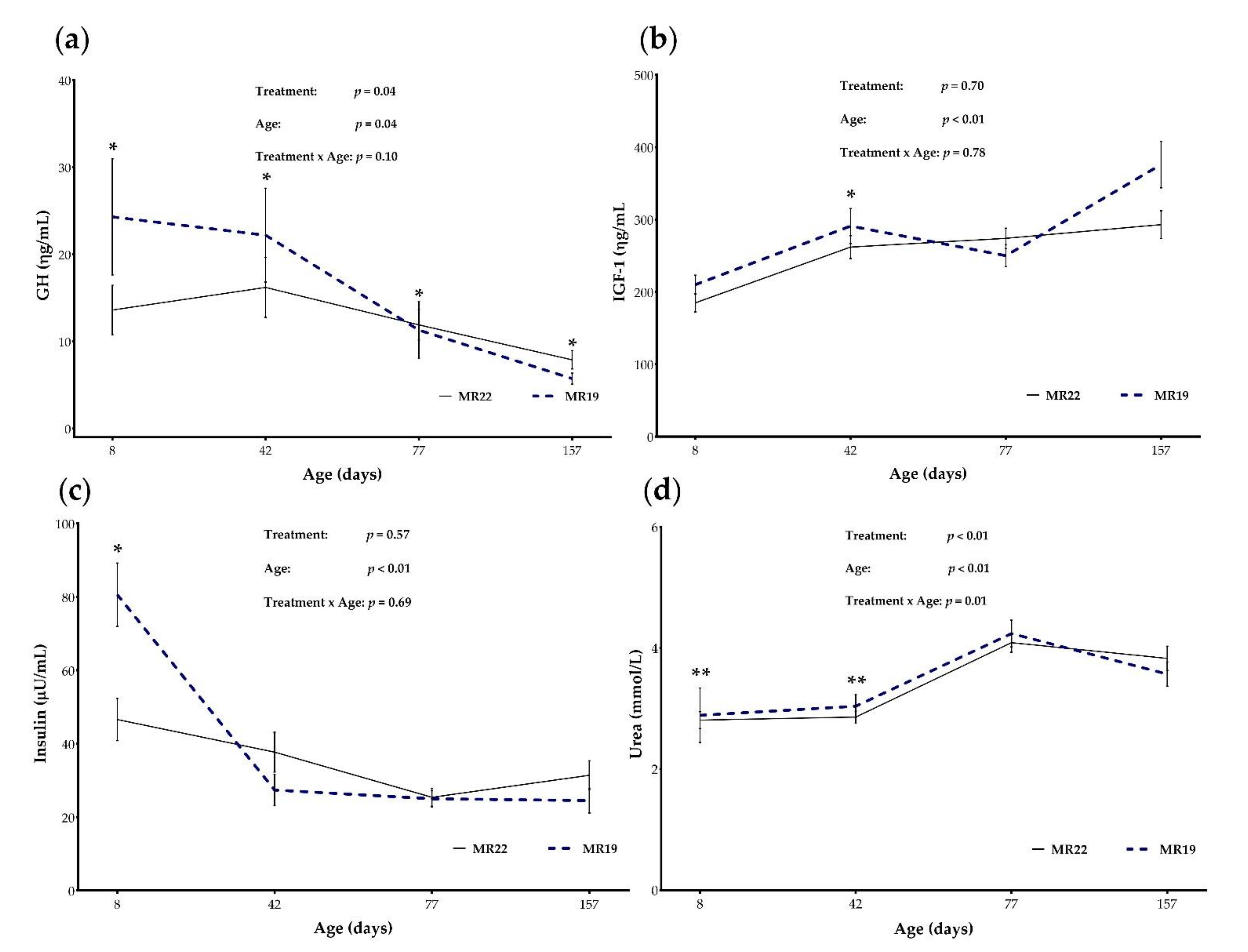
3.5. Blood Parameters
3.5.1. GH and IGF-1
3.5.2. Insulin and Glucose
4. Discussion
4.1. Performance of Calves
4.2. Fecal Quality
4.3. Metabolic Traits
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Conneely, M.; Berry, D.; Murphy, J.; Lorenz, I.; Doherty, M.; Kennedy, E. Effects of milk feeding volume and frequency on body weight and health of dairy heifer calves. Livest. Sci. 2014, 161, 90–94. [Google Scholar] [CrossRef]
- Hulbert, L.E.; Moisá, S.J. Stress, immunity, and the management of calves. J. Dairy Sci. 2016, 99, 3199–3216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carter, H.S.; Renaud, D.L.; Steele, M.A.; Fischer-Tlustos, A.J.; Costa, J.H. A Narrative Review on the Unexplored Potential of Colostrum as a Preventative Treatment and Therapy for Diarrhea in Neonatal Dairy Calves. Animals 2021, 11, 2221. [Google Scholar] [CrossRef]
- Bartlett, K.; McKeith, F.; VandeHaar, M.; Dahl, G.; Drackley, J. Growth and body composition of dairy calves fed milk replacers containing different amounts of protein at two feeding rates. J. Anim. Sci. 2006, 84, 1454–1467. [Google Scholar] [CrossRef]
- Khan, M.; Weary, D.; Von Keyserlingk, M. Invited review: Effects of milk ration on solid feed intake, weaning, and performance in dairy heifers. J. Dairy Sci. 2011, 94, 1071–1081. [Google Scholar] [CrossRef] [PubMed]
- Geiger, A.; Parsons, C.; James, R.; Akers, R. Growth, intake, and health of Holstein heifer calves fed an enhanced preweaning diet with or without postweaning exogenous estrogen. J. Dairy Sci. 2016, 99, 3995–4004. [Google Scholar] [CrossRef]
- Bleach, E.; Gould, M.; Blackie, N.; Beever, D. Growth Performance of Holstein-Friesian Heifer Calves Reared Using Three Milk Replacer Feeding Regimes. In Recent Advances in Animal Nutrition 2005; Garnsworthy, P.C., Wiseman, J., Eds.; Nottingham University Press: Nottingham, UK, 2006; Volume 39, pp. 347–357. [Google Scholar]
- Palczynski, L.J.; Bleach, E.C.; Brennan, M.L.; Robinson, P.A. Appropriate dairy calf feeding from birth to weaning:“it’s an investment for the future”. Animals 2020, 10, 116. [Google Scholar] [CrossRef] [Green Version]
- Albright, J.L.; Arave, C.W. The Behaviour of Cattle; CAB International: Wallingford, UK, 1997. [Google Scholar]
- Diaz, M.; Van Amburgh, M.; Smith, J.; Kelsey, J.; Hutten, E. Composition of growth of Holstein calves fed milk replacer from birth to 105-kilogram body weight. J. Dairy Sci. 2001, 84, 830–842. [Google Scholar] [CrossRef]
- Jasper, J.; Weary, D. Effects of ad libitum milk intake on dairy calves. J. Dairy Sci. 2002, 85, 3054–3058. [Google Scholar] [CrossRef]
- De Paula, M.; Oltramari, C.; Silva, J.d.; Gallo, M.d.C.; Mourão, G.; Bittar, C. Intensive liquid feeding of dairy calves with a medium crude protein milk replacer: Effects on performance, rumen, and blood parameters. J. Dairy Sci. 2017, 100, 4448–4456. [Google Scholar] [CrossRef] [Green Version]
- Bascom, S.; James, R.; McGilliard, M.; Van Amburgh, M. Influence of dietary fat and protein on body composition of Jersey bull calves. J. Dairy Sci. 2007, 90, 5600–5609. [Google Scholar] [CrossRef] [PubMed]
- Hill, T.; Bateman Ii, H.; Aldrich, J.; Schlotterbeck, R. Effect of milk replacer program on digestion of nutrients in dairy calves. J. Dairy Sci. 2010, 93, 1105–1115. [Google Scholar] [CrossRef] [PubMed]
- Johnson, K.F.; Nair, R.V.; Wathes, D. Comparison of the effects of high and low milk-replacer feeding regimens on health and growth of crossbred dairy heifers. Anim. Prod. Sci. 2019, 59, 1648–1659. [Google Scholar] [CrossRef]
- Hammon, H.; Zanker, I.; Blum, J. Delayed colostrum feeding affects IGF-I and insulin plasma concentrations in neonatal calves. J. Dairy Sci. 2000, 83, 85–92. [Google Scholar] [CrossRef]
- Sauter, S.N.; Ontsouka, E.; Roffler, B.; Zbinden, Y.; Philipona, C.; Pfaffl, M.; Breier, B.H.; Blum, J.W.; Hammon, H.M. Effects of dexamethasone and colostrum intake on the somatotropic axis in neonatal calves. Am. J. Physiol. Endocrinol. Metab. 2003, 285, E252–E261. [Google Scholar] [CrossRef] [Green Version]
- Breier, B.; Oliver, M.; Gallaher, B. Regulation of Growth and Metabolism during Postnatal Development. In Ruminant Physiology: Digestion, Metabolism, Growth and Reproduction; Cronje, P.B., Ed.; CABI Publishing: Wallingford, UK, 2000; pp. 187–204. [Google Scholar]
- Akers, R. Major advances associated with hormone and growth factor regulation of mammary growth and lactation in dairy cows. J. Dairy Sci. 2006, 89, 1222–1234. [Google Scholar] [CrossRef]
- Schäff, C.T.; Gruse, J.; Maciej, J.; Mielenz, M.; Wirthgen, E.; Hoeflich, A.; Schmicke, M.; Pfuhl, R.; Jawor, P.; Stefaniak, T. Effects of feeding milk replacer ad libitum or in restricted amounts for the first five weeks of life on the growth, metabolic adaptation, and immune status of newborn calves. PLoS ONE 2016, 11, e0168974. [Google Scholar]
- Haisan, J.; Oba, M.; Ambrose, D.; Steele, M. The effects of offering a high or low plane of milk preweaning on insulin-like growth factor and insulin-like growth factor binding proteins in dairy heifer calves. J. Dairy Sci. 2018, 101, 11441–11446. [Google Scholar] [CrossRef] [Green Version]
- Renaville, R.; Hammadi, M.; Portetelle, D. Role of the somatotropic axis in the mammalian metabolism. Domest. Anim. Endocrinol. 2002, 23, 351–360. [Google Scholar] [CrossRef]
- Hammon, H.; Steinhoff-Wagner, J.; Schönhusen, U.; Metges, C.; Blum, J. Energy metabolism in the newborn farm animal with emphasis on the calf: Endocrine changes and responses to milk-born and systemic hormones. Domest. Anim. Endocrinol. 2012, 43, 171–185. [Google Scholar] [CrossRef]
- Maccari, P.; Wiedemann, S.; Kunz, H.J.; Piechotta, M.; Sanftleben, P.; Kaske, M. Effects of two different rearing protocols for Holstein bull calves in the first 3 weeks of life on health status, metabolism and subsequent performance. J. Anim. Physiol. Anim. Nutr. 2015, 99, 737–746. [Google Scholar] [CrossRef] [PubMed]
- Larson, L.; Owen, F.; Albright, J.; Appleman, R.; Lamb, R.; Muller, L. Guidelines toward more uniformity in measuring and reporting calf experimental data. J. Dairy Sci. 1977, 60, 989–991. [Google Scholar] [CrossRef]
- Chapman, C.E.; Erickson, P.S.; Quigley, J.D.; Hill, T.M.; Bateman, H.G.; Suarez-Mena, F.X.; Schlotterbeck, R.L. Effect of milk replacer program on calf performance and digestion of nutrients with age of the dairy calf. J. Dairy Sci. 2016, 99, 2740–2747. [Google Scholar] [CrossRef] [Green Version]
- Naumann, C.; Bassler, R. Methoden der Landwirtschaftlichen Forschungs-und Untersuchungsanstalt, Biochemische Untersuchung von Futtermitteln. Methodenbuch III (Einschließlich der Achten Ergänzungen); VDLUFA: Darmstadt, Germany, 2012. [Google Scholar]
- Chudy, A. Rostock Feed Evaluation System—An Example of the Transformation of Energy and Nutrient Utilization Models to Practical Application; CABI Publishing: Wallingford, UK, 2006; pp. 366–382. [Google Scholar] [CrossRef]
- Löhnert, H.J.; Ochrimenko, W.I. Untersuchungen zur Verdaulichkeit von Milchaustauschern bei Kalbern; DLG Verlag: Frankfurt, Germany, 1998; p. 75. [Google Scholar]
- Schuhknecht, W.; Schinkel, H. Universalvorschrift fur die Bestimmung von Kalium, Natrium und Lithium nebeneinander. Z. Analyt. Chem. 1963, 194, 176–183. [Google Scholar]
- Roh, S.-G.; Matsunaga, N.; Miyamoto, A.; Hidaka, S.; Hidari, H. Competitive enzyme immunoassay for bovine growth hormone. Endocr. J. 1997, 44, 195–198. [Google Scholar] [CrossRef] [Green Version]
- Kawashima, C.; Sakaguchi, M.; Suzuki, T.; Sasamoto, Y.; Takahashi, Y.; Matsui, M.; Miyamoto, A. Metabolic profiles in ovulatory and anovulatory primiparous dairy cows during the first follicular wave postpartum. J. Reprod. Dev. 2007, 53, 113. [Google Scholar] [CrossRef] [Green Version]
- Akoglu, H. User’s guide to correlation coefficients. Turk. J. Emerg. Med. 2018, 18, 91–93. [Google Scholar] [CrossRef]
- Blome, R.; Drackley, J.; McKeith, F.; Hutjens, M.; McCoy, G. Growth, nutrient utilization, and body composition of dairy calves fed milk replacers containing different amounts of protein. J. Anim. Sci. 2003, 81, 1641–1655. [Google Scholar] [CrossRef]
- Heinrichs, A.J.; Wells, S.J.; Losinger, W.C. A Study of the Use of Milk Replacers for Dairy Calves in the United States. J. Dairy Sci. 1995, 78, 2831–2837. [Google Scholar] [CrossRef]
- Jaeger, B.M.; Ziegler, D.; Schimek, D.; Ziegler, B.; Raeth, M.; Chester-Jones, H.; Casper, D.P. Growth performance of newborn dairy calves fed a milk replacer with 2 protein concentrations at 2 feeding rates. Appl. Anim. Sci. 2020, 36, 48–56. [Google Scholar] [CrossRef]
- Hill, S.; Knowlton, K.; Daniels, K.M.; James, R.; Pearson, R.; Capuco, A.; Akers, R. Effects of milk replacer composition on growth, body composition, and nutrient excretion in preweaned Holstein heifers. J. Dairy Sci. 2008, 91, 3145–3155. [Google Scholar] [CrossRef] [PubMed]
- Chapman, C.E.; Hill, T.; Elder, D.; Erickson, P.S. Nitrogen utilization, preweaning nutrient digestibility, and growth effects of Holstein dairy calves fed 2 amounts of a moderately high protein or conventional milk replacer. J. Dairy Sci. 2017, 100, 279–292. [Google Scholar] [CrossRef] [PubMed]
- Cowles, K.E.; White, R.A.; Whitehouse, N.L.; Erickson, P.S. Growth Characteristics of Calves Fed an Intensified Milk Replacer Regimen with Additional Lactoferrin1. J. Dairy Sci. 2006, 89, 4835–4845. [Google Scholar] [CrossRef]
- NASEM (National Academies of Science, Engineering and Medicine). Nutrient Requirements of Dairy Cattle: Eigth Revised Edition; The National Academies Press: Washington, DC, USA, 2021. [Google Scholar] [CrossRef]
- GfE (Ausschuss für Bedarfnormen der Gesellschaft für Ernährungsphyiologie). Empfehlungen zur Proteinversorgung von Aufzuchtkälbern. Proc. Soc. Nutr. Physiol. 1999, 8, 155–164. [Google Scholar]
- NRC. Nutrient Requirements of Dairy Cattle: 2001; National Academies Press: Washington, DC, USA, 2001. [Google Scholar]
- Heinrichs, A.; Heinrichs, B. A prospective study of calf factors affecting first-lactation and lifetime milk production and age of cows when removed from the herd. J. Dairy Sci. 2011, 94, 336–341. [Google Scholar] [CrossRef] [PubMed]
- Davis, C.L.; Drackley, J.K. The Development, Nutrition, and Management of the Young Calf; Iowa State University Press: Iowa, MO, USA, 1998. [Google Scholar]
- Echeverry-Munera, J.; Leal, L.N.; Wilms, J.N.; Berends, H.; Costa, J.H.; Steele, M.; Martín-Tereso, J. Effect of partial exchange of lactose with fat in milk replacer on ad libitum feed intake and performance in dairy calves. J. Dairy Sci. 2021, 104, 5432–5444. [Google Scholar] [CrossRef]
- Jeffery, H.; Berg, R. Evaluation of milk variables as measures of milk effect on preweaning performance of beef cattle. Can. J. Anim. Sci. 1971, 51, 21–30. [Google Scholar] [CrossRef]
- Lorenz, I.; Fagan, J.; More, S.J. Calf health from birth to weaning. II. Management of diarrhoea in pre-weaned calves. Ir. Vet. J. 2011, 64, 9. [Google Scholar] [CrossRef] [Green Version]
- Cho, Y.-i.; Yoon, K.-J. An overview of calf diarrhea-infectious etiology, diagnosis, and intervention. J. Vet. Sci. 2014, 15, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Dirksen, G. Die Kotuntersuchung. In Die Klinische Untersuchung des Rindes; Rosenberger, G., Ed.; Verlag Paul Parey: Berlin, Germany, 1989; pp. 358–363. [Google Scholar]
- Ballou, M.; Hanson, D.; Cobb, C.; Obeidat, B.; Sellers, M.; Pepper-Yowell, A.; Carroll, J.; Earleywine, T.; Lawhon, S. Plane of nutrition influences the performance, innate leukocyte responses, and resistance to an oral Salmonella enterica serotype Typhimurium challenge in Jersey calves. J. Dairy Sci. 2015, 98, 1972–1982. [Google Scholar] [CrossRef]
- Palmer, A.L.; Beausoleil, N.J.; Boulton, A.C.; Cogger, N. Prevalence of Potential Indicators of Welfare Status in Young Calves at Meat Processing Premises in New Zealand. Animals 2021, 11, 2467. [Google Scholar] [CrossRef] [PubMed]
- Dirksen, G. Krankheiten des Darmes. In Innere Medizin und Chirurgie des Rindes; Dirksen, G., Gründer, H.-D., Stöber, M., Eds.; Verlag Parey in MVS Medizinverlage Stuttgart GmbH & Co. KG: Stuttgart, Germany, 2006; pp. 514–597. [Google Scholar]
- Renaud, D.; Buss, L.; Wilms, J.; Steele, M. Is fecal consistency scoring an accurate measure of fecal dry matter in dairy calves? J. Dairy Sci. 2020, 103, 10709–10714. [Google Scholar] [CrossRef] [PubMed]
- Ireland-Perry, R.; Stallings, C. Fecal consistency as related to dietary composition in lactating Holstein cows. J. Dairy Sci. 1993, 76, 1074–1082. [Google Scholar] [CrossRef]
- Ferreira, J.; Noller, C.; Keyser, R.; Stewart, T. Influence of dietary calcium and protein on fecal pH, consistency, and rate of passage in dairy cattle. J. Dairy Sci. 1980, 63, 1091–1097. [Google Scholar] [CrossRef]
- Erfle, J.; Boila, R.; Teather, R.; Mahadevan, S.; Sauer, F. Effect of pH on fermentation characteristics and protein degradation by rumen microorganisms in vitro. J. Dairy Sci. 1982, 65, 1457–1464. [Google Scholar] [CrossRef]
- Haaland, G.L.; Tyrrell, H.F.; Moe, P.W.; Wheeler, W.E. Effect of Crude Protein Level and Limestone Buffer in Diets Fed at Two Levels of Intake on Rumen pH, Ammonia-Nitrogen, Buffering Capacity and Volatile Fatty Acid Concentration of Cattle. J. Anim. Sci. 1982, 55, 943–950. [Google Scholar] [CrossRef]
- Veira, D.M.; Macleod, G.K.; Burton, J.H.; Stone, J.B. Nutrition of the Weaned Holstein Calf. II. Effect of Dietary Protein Level on Nitrogen Balance, Digestibility and Feed Intake. J. Anim. Sci. 1980, 50, 945–951. [Google Scholar] [CrossRef]
- Wheeler, W.; Noller, C. Gastrointestinal tract pH and starch in feces of ruminants. J. Anim. Sci. 1977, 44, 131–135. [Google Scholar] [CrossRef]
- Schönbrodt, F.D.; Perugini, M. At what sample size do correlations stabilize? J. Res. Pers. 2013, 47, 609–612. [Google Scholar] [CrossRef] [Green Version]
- Fisher, E.; De la Fuente, G. Water and electrolyte studies in newborn calves with particular reference to the effects of diarrhoea. Res. Vet. Sci. 1972, 13, 315–323. [Google Scholar] [CrossRef]
- Hammon, H.; Blum, J.W. The somatotropic axis in neonatal calves can be modulated by nutrition, growth hormone, and Long-R3-IGF-I. Am. J. Physiol. Endocrinol. Metab. 1997, 273, E130–E138. [Google Scholar] [CrossRef]
- Smith, J.; Van Amburgh, M.; Diaz, M.; Lucy, M.; Bauman, D. Effect of nutrient intake on the development of the somatotropic axis and its responsiveness to GH in Holstein bull calves. J. Anim. Sci. 2002, 80, 1528–1537. [Google Scholar] [CrossRef] [PubMed]
- Metges, C.C.; Hammon, H.M. Metabolische Programmierung bei Nutztieren. Übers. Tierernährung 2008, 36, 1–29. [Google Scholar]
- Bassett, J.; Weston, R.; Hogan, J. Dietary regulation of plasma insulin and growth hormone concentrations in sheep. Aust. J. Biol. Sci. 1971, 24, 321–330. [Google Scholar] [CrossRef] [Green Version]
- Hammon, H.; Schiessler, G.; Nussbaum, A.; Blum, J. Feed intake patterns, growth performance, and metabolic and endocrine traits in calves fed unlimited amounts of colostrum and milk by automate, starting in the neonatal period. J. Dairy Sci. 2002, 85, 3352–3362. [Google Scholar] [CrossRef]
- Egli, C.P.; Blum, J.W. Clinical, Haematological, Metabolic and Endocrine Traits During the First Three Months of Life of Suckling Simmentaler Calves Held in a Cow-Calf Operation1. J. Vet. Med. 1998, 45, 99–118. [Google Scholar] [CrossRef] [PubMed]
- Gerrits, W.J.; Decuypere, E.; Verstegen, M.W.; Karabinas, V. Effect of protein and protein-free energy intake on plasma concentrations of insulin-like growth factor I and thyroid hormones in preruminant calves. J. Anim. Sci. 1998, 76, 1356–1363. [Google Scholar] [CrossRef]
- Breier, B.; Gluckman, P.; Bass, J. Plasma concentrations of insulin-like growth factor-I and insulin in the infant calf: Ontogeny and influence of altered nutrition. J. Endocrinol. 1988, 119, 43–50. [Google Scholar] [CrossRef]
- Fürll, M. Spezielle Untersuchungen beim Wiederkäuer. In Klinische Labordiagnostik in der Tiermedizin; Schattauer GmbH: Stuttgart, Germany, 2014; Volume 726–777. [Google Scholar]
- Silper, B.; Lana, A.; Carvalho, A.; Ferreira, C.; Franzoni, A.; Lima, J.; Saturnino, H.; Reis, R.; Coelho, S. Effects of milk replacer feeding strategies on performance, ruminal development, and metabolism of dairy calves. J. Dairy Sci. 2014, 97, 1016–1025. [Google Scholar] [CrossRef]
- Makizadeh, H.; Kazemi-Bonchenari, M.; Mansoori-Yarahmadi, H.; Fakhraei, J.; Khanaki, H.; Drackley, J.; Ghaffari, M. Corn processing and crude protein content in calf starter: Effects on growth performance, ruminal fermentation, and blood metabolites. J. Dairy Sci. 2020, 103, 9037–9053. [Google Scholar] [CrossRef]
- Khan, M.; Lee, H.; Lee, W.; Kim, H.; Ki, K.; Hur, T.; Suh, G.; Kang, S.; Choi, Y. Structural growth, rumen development, and metabolic and immune responses of Holstein male calves fed milk through step-down and conventional methods. J. Dairy Sci. 2007, 90, 3376–3387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vicari, T.; Borne, J.J.G.C.V.D.; Gerrits, W.J.J.; Zbinden, Y.; Blum, J.W. Separation of protein and lactose intake ovel meals dissociates postprandial glucose and insulin concentrations and reduces postprandial glucose and insulin responses in heavy veal calves. Domest. Anim. Endocrinol. 2008, 34, 182–195. [Google Scholar] [CrossRef] [PubMed]
- Hussein, H.A.; Thurmann, J.-P.; Staufenbiel, R. 24-h variations of blood serum metabolites in high yielding dairy cows and calves. BMC Vet. Res. 2020, 16, 327. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item | MR22 1 | MR19 2 |
|---|---|---|
| Ingredient | ||
| Skimmed milk powder | 40.0 | 40.0 |
| Whey powder | 12.5 | 32.5 |
| Whey powder, partially delactosed | 10.0 | - |
| Whey protein powder | 10.0 | - |
| Vegetable fat (coconut/palm) | 18.0 | 18.0 |
| Beer yeast | 0.30 | 0.30 |
| Pregelatinized wheat starch | 2.00 | 2.00 |
| Dextrose | 1.50 | 1.50 |
| Chemical composition | ||
| DM (% of fresh matter) | 96.3 | 96.8 |
| Crude ash | 7.20 | 6.70 |
| Crude protein | 21.8 | 18.5 |
| Ether extract | 19.5 | 18.8 |
| Crude fibre | 0.00 | 0.00 |
| NfE 3 | 51.5 | 56.0 |
| Metabolizable energy (ME), MJ/kg of DM | 17.5 | 17.4 |
| Protein:energy ratio MJ ME/g CP | 1.25 | 1.06 |
| Calcium | 0.87 | 0.79 |
| Phosphorus | 0.71 | 0.65 |
| Sodium | 0.68 | 0.58 |
| Potassium | 1.57 | 1.31 |
| Magnesium | 0.17 | 0.14 |
| Item | Concentrates | Straw | MCF | TMR |
|---|---|---|---|---|
| DM (% of fresh matter) | 87.5 | 89.6 | 87.8 | 46.2 |
| Metabolizable energy (ME), MJ/kg of DM | 13.0 | 6.40 | 12.2 | 10.9 |
| Crude ash | 7.40 | 4.70 | 7.00 | 8.90 |
| Crude protein | 21.1 | 3.00 | 18.7 | 15.3 |
| Ether extract | 4.90 | 1.20 | 4.40 | 3.50 |
| Crude fibre | 6.30 | 46.5 | 11.5 | 18.9 |
| NfE 1 | 60.4 | 44.7 | 58.4 | 53.4 |
| Age (d) | DM intake | MR22 1 | MR19 2 | p-Value |
|---|---|---|---|---|
| 8–42 | MR | 1310 ± 137 | 1300 ± 137 | 0.48 |
| MCF | 91.0 ± 66.0 | 146 ± 120 | <0.01 | |
| Total | 1401 ± 176 | 1446 ± 218 | 0.73 | |
| 43–77 | MR | 555 ± 345 | 560 ± 353 | 0.57 |
| MCF | 1217 ± 664 | 1344 ± 677 | 0.43 | |
| Total | 1772 ± 339 | 1904 ± 317 | 0.06 | |
| 78–157 | TMR | 2627 ± 574 | 2547 ± 564 | 0.81 |
| MCF | 1470 ± 140 | 1532 ± 154 | 0.35 | |
| Total | 4097 ± 893 | 4079 ± 730 | 0.85 |
| Age (d) | MR22 1 | MR19 2 | p-Value | ||
|---|---|---|---|---|---|
| Mean ± SEM | Min-Max | Mean ± SEM | Min-Max | ||
| Initial | 82.3 ± 056 | 76–88 | 81.9 ± 0.57 | 74–88 | 0.46 |
| 8 | 84.4 ± 0.50 | 78–89 | 84.3 ± 0.54 | 79–92 | 0.75 |
| 22 | 88.5 ± 0.43 | 82–93 | 88.8 ± 0.49 | 82–94 | 0.56 |
| 42 | 94.3 ± 0.48 | 87–100 | 94.2 ± 0.46 | 87–99 | 0.74 |
| 77 | 102 ± 0.43 | 95–107 | 103 ± 0.50 | 96–108 | 0.50 |
| 117 | 111 ± 0.49 | 104–117 | 110 ± 0.61 | 105–119 | 0.32 |
| 157 | 118 ± 0.60 | 109–123 | 118 ± 0.64 | 112–124 | 0.68 |
| Characteristic | Pre-Transition 3 and Transition 4 PHASE | p-Value | Postweaning 5 Phase | p-Value | ||
|---|---|---|---|---|---|---|
| MR22 1 (n = 350) | MR19 2 (n = 330) | MR22 1 (n = 102) | MR19 2 (n = 99) | |||
| Color | ||||||
| Brown-green | 60.6% (212) | 54.9% (181) | 0.20 | 99.0% (101) | 100% (99) | 0.32 |
| Brown-orange | 16.9% (59) | 16.1% (53) | 0.79 | 1.00% (1) | 0% (0) | 0.46 |
| Yellow-brown | 22.5% (79) | 28.8% (95) | 0.26 | 0% (0) | 0% (0) | - |
| Whitish-grey | 0% (0) | 0.20% (1) | 0.81 | 0% (0) | 0% (0) | - |
| Consistency | ||||||
| Firm | 0% (0) | 0% (0) | - | 0% (0) | 0% (0) | - |
| Thick Pulpy | 2.00% (7) | 1.82% (6) | 0.67 | 17.7% (18) | 5.05% (5) | 0.02 |
| Medium Pulpy (Pasty) | 63.1% (221) | 57.6% (190) | 0.02 | 68.6% (70) | 75.8% (75) | 0.74 |
| Mushy | 20.0% (70) | 21.5% (71) | 0.76 | 11.8% (12) | 16.2% (16) | 0.58 |
| Loose | 9.70% (34) | 9.70% (32) | 0.64 | 1.96% (2) | 1.01% (1) | 0.70 |
| Watery | 5.14% (18) | 9.39% (31) | 0.25 | 0% (0) | 2.02% (2) | 0.71 |
| Fecal DM Classification | MR22 1 | MR19 2 | ||
|---|---|---|---|---|
| DM Content | Sodium Content | DM Content | Sodium Content | |
| Low | 44.7 ± 11.4 | 31.4 | 51.4 ± 7.48 | 22.9 |
| Medium | 165 ± 0.57 | 4.27 | 153 ± 1.10 | 5.82 |
| High | 284 ± 29.4 | 3.95 | 236 ± 12.7 | 4.85 |
| Item | Treatment | p-Value | |||
|---|---|---|---|---|---|
| MR22 1 | SEM | MR19 2 | SEM | Treatment | |
| GH, ng/mL | |||||
| d 8 | 13.6 b | 2.84 | 24.3 a | 6.67 | <0.01 |
| d 42 3 | 16.2 b | 3.44 | 22.2 a | 5.39 | 0.02 |
| d 77 4 | 11.9 a | 1.75 | 11.3 b | 3.25 | 0.01 |
| d 157 | 7.89 a | 1.05 | 5.73 b | 0.63 | 0.04 |
| IGF-1, ng/mL | |||||
| d 8 | 185 | 12.6 | 210 | 13.2 | 0.92 |
| d 42 3 | 262 b | 15.8 | 291 a | 24.1 | 0.03 |
| d 77 4 | 274 | 14.1 | 250 | 15.2 | 0.75 |
| d 157 | 293 | 19.2 | 376 | 32.1 | 0.07 |
| Insulin, µU/mL | |||||
| d 8 | 46.6 b | 5.76 | 80.6 a | 8.63 | 0.04 |
| d 42 3 | 37.7 | 5.42 | 27.4 | 4.27 | 0.11 |
| d 77 4 | 25.4 | 2.44 | 25.0 | 2.19 | 0.49 |
| d 157 | 31.4 | 3.94 | 24.5 | 3.37 | 0.48 |
| Glucose, mg/dL | |||||
| d 8 | 108 | 7.75 | 132 | 6.67 | 0.43 |
| d 42 3 | 109 | 4.90 | 88.4 | 4.24 | 0.50 |
| d 77 4 | 79.9 | 3.00 | 73.4 | 2.16 | 0.22 |
| d 157 | 78.5 | 1.94 | 90.4 | 2.03 | 0.94 |
| Urea, mmol/L | |||||
| d 8 | 2.81 b | 0.14 | 2.89 a | 0.45 | <0.01 |
| d 42 3 | 2.86 b | 0.10 | 3.04 a | 0.19 | <0.01 |
| d 77 4 | 4.09 | 0.16 | 4.24 | 0.22 | 0.12 |
| d 157 | 3.83 | 0.20 | 3.57 | 0.20 | 0.83 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schubert, D.C.; Chuppava, B.; Hoffmans, S.; Pries, M.; Visscher, C.; Kamphues, J.; Abd El-Wahab, A. Impacts of Reducing Protein Content in Milk Replacer on Growth Performance and Health of Young Calves. Animals 2022, 12, 1756. https://doi.org/10.3390/ani12141756
Schubert DC, Chuppava B, Hoffmans S, Pries M, Visscher C, Kamphues J, Abd El-Wahab A. Impacts of Reducing Protein Content in Milk Replacer on Growth Performance and Health of Young Calves. Animals. 2022; 12(14):1756. https://doi.org/10.3390/ani12141756
Chicago/Turabian StyleSchubert, Dana Carina, Bussarakam Chuppava, Sandra Hoffmans, Martin Pries, Christian Visscher, Josef Kamphues, and Amr Abd El-Wahab. 2022. "Impacts of Reducing Protein Content in Milk Replacer on Growth Performance and Health of Young Calves" Animals 12, no. 14: 1756. https://doi.org/10.3390/ani12141756