Drivers for the artisanal fisheries production in the Magdalena River
 Luz Jiménez-Segura
Luz Jiménez-Segura Juan D. Restrepo-Ángel
Juan D. Restrepo-Ángel Andrés Hernandez-Serna
Andrés Hernandez-Serna- 1Instituto de Biologia, Universidad de Antioquia, Medellín, Colombia
- 2School of Applied Sciences and Engineering, Universidad EAFIT, Medellín, Colombia
- 3Department of Geographical Sciences, University of Maryland, Rockville, MD, United States
We review knowledge on the Magdalena River in Colombia and its fish to identify those drivers that influence the artisanal fisheries production. We identify eight direct drivers (four natural and four anthropogenic) and at least four indirect drivers. Those drivers modify conditions in the fluvial network that promote fish movements, reproduction, and their larvae survivor. Landscape, rains, floods, connectivity of the fluvial net as land cover change, water pollution, hydropower, and alien species are the natural and anthropogenic direct drivers described in this article. The river–lake interaction dynamics in the Magdalena River are determined by two rainy cycles per year. Two seasonal flooding periods induce two cycles in the biological productivity of floodplains because water and sediment inputs. The most visible consequences in these hydrological cycles are the migrations of potamodromous fish and the periodic increase in the artisanal fishery production. Major floodplains are reducing their storage capacity by trapping ∼10%–40% of upstream sediment production. This process induces many research questions about rates of biomass production, carbon fluxes in the basin, impacts of human-induced erosion, and increasing rates of sediment load on floodplain connectivity, but still there is not enough data to answer them. Finally, we make some suggestions toward the sustainability of the Magdalena floodplains. The well-being of the floodplain ecosystems and their connectivity with the main river are the main tools to preserve and manage the ecosystem services of the Magdalena River and its floodplains lakes.
Introduction
Fluvial networks are the hotspots of human activities and one of the most threatened ecosystems in the world (Strayer and Dudgeon, 2010). Thirty one percent of all freshwater fishes and amphibians are included in the Red List of Threatened Species of the International Union for Conservation of Nature (IUCN, 2020). In the last five decades, the understanding of the function, diversity, and complexity of the freshwater ecosystems have demonstrated urgency of action for river conservation and management; the more we know about them, the more we understand what has been changing and the need to promote their protection. Reid et al. (2019) identified 12 threats to freshwater diversity: climate change, e-commerce and invasions, light and noise, expanding hydropower, emerging contaminants, microplastic pollution, declining Calcium, freshwater salinization, harmful algal blooms, infectious diseases, engineered nanomaterials, and cumulative stressors. Recently, Williams-Subiza and Epele (2021) summarized the main drivers modeling the freshwater ecosystems, after a bibliometric analysis of 10 years of published knowledge about them; they identified that scientific research works have been focused mainly in six drivers: climate change, habitat degradation, expanding hydropower, flow change, water pollution, and species invasions.
A driver is any natural or human-induced factor that directly or indirectly causes a change in an ecosystem; there are direct and indirect drivers (Carpenter et al., 2006). The direct drivers are natural or anthropogenic factors that strongly influence the functioning of the ecosystems. Some of them are climate variability and climate change, land conversion, nutrient support to soils for agricultural purposes, natural resources use and exploitation, and biological invasions. Indirect drivers influence more diffusely the direct drivers (i.e., human demography, economic and socio-political trends, scientific advances to improve biomass production, and human culture) (Nelson et al., 2006). Diversity and dynamics of the river biota strongly responds on the combination of indirect and direct drivers. Changes in the artisanal fisheries production can be used as a response variable that explains natural or induced changes in freshwater ecosystems (Hernandez-Barrero et al., 2021). Therefore, freshwater fish can be a good model for describing how these drivers may interact to influence their response in a river in the tropical Andes, exposed to monsoonal climate, high steep relief basins, and high sediment production and because of human-induced activities, posing unprecedented challenges to their natural functioning.
The Magdalena River in Colombia is a unique fluvial system in the tropical region. It is the highest sediment-yielding river (∼710 t km2 y−1) among the rivers that drain South America, transporting a sediment load to the ocean of 184 Mt y−1 (Restrepo et al., 2015). It has a fantastic geologic history because of the uplift of three Andean mountain ranges and the advance of the sea into continental land because of sea level changes during the Pleistocene (Lundberg et al., 1998). It is a morphologically and climatologically diverse basin with large floodplains, high mountains, and episodic local climate events because the interaction of the mountain ranges and the seasonal movement of rains coming from the Pacific Ocean, the Amazonas and Orinoco basins, and the intertropical convergence zone (ITCZ) (Restrepo et al., 2020a). It is born and perishes in the same country and since most of the Colombian population inhabits the basin (∼80%) (The Nature Conservancy, Fundación Alma, Fundación Humedales and Autoridad Nacional de Pesca, 2016), their impacts, including deforestation, water pollution, hydropower, poor soil conservation, mining practices, alien fish species, and increasing rates of urbanization, have accounted for the overall increasing trends in the threats for the rivers conservation. Downstream of 300 m elevation, the lowlands of the Magdalena River cover 10% of the catchment area and are the largest alluvial fans in the northern Andes and the least studied fluvial lakes of major South American rivers. Lakes in these floodplains provide refuge to aquatic biota, controlling floods, and supplying water for human consumption and irrigation, and they are nursery areas for fish species used by artisanal fishery (Jaramillo et al., 2015a). However, the ongoing human activities along the whole basin have dramatically changed these vital ecosystems and fisheries landings (Barletta et al., 2010; Hernandez-Barrero et al., 2021). Because of human activities in the lowlands such as dikes, dams, agriculture, and cattle ranching, more than 30% of the floodplain lakes have lost their natural connections to the main Magdalena River, and many lakes had dried during the last two decades (The Nature Conservancy, Fundación Alma, Fundación Humedales and Autoridad Nacional de Pesca, 2016; Restrepo et al., 2020b).
Freshwater fish richness of the Magdalena River has 233 species (García-Alzate e t al., 2020). There is an important turnover along the elevation gradient with few fish species up the 1,500 m elevation (Jaramillo-Villa et al., 2010; Carvajal-Quintero et al., 2015; Herrera-Pérez et al., 2019). Fish diversity in the lowlands aquatic systems (main river channel, lakes, creeks, and connecting channels) are composed of 173 species; 45% percent are endemic and 36% occur in the lakes. Eighteen of the endemic freshwater species are potamodromous–White fish (following Welcomme et al., 2006) (Usma-Oviedo et al., 2013; López-Casas et al., 2016) and the other ones do not migrate (Usma-Oviedo et al., 2013); although more information is needed to be conclusive about movement strategies. Three endemic species Totumito Abramites eques (Steindachner, 1878), Chipe Hoplosternum magdalenae (Eigenmann, 1913), and Cachegua Trachelyopterus insignis (Steindachner, 1878) are exclusive to floodplain lakes. Fish species distributions in the Magdalena River floodplain lakes are highly nested, and fish diversity is positively related to the lake area (Granado-Lorencio et al., 2012). Forty species of these lowland fish are resource for artisanal fisheries, and nineteen of them are White fish; in the last five decades, fisheries landings had reduced (Valderrama et al., 2020). Because fish production in the Magdalena River strongly depends on the interaction between different drivers in the Magdalena lowlands, our aim in this study is to identify and describe those related with fish and environmental dynamics and sustainability. We use available data on the Magdalena River discharge, sediments, floodplain dynamics, the fish life history and dynamics, and the fisheries landings. Finally, we discuss the science required to understand the importance of interconnectivity toward environmental management and artisanal fisheries sustainability.
Data sources and methods
We focus on direct drivers, and when necessary, we include indirect drivers that may influence a direct driver. Natural direct drivers are: 1) riverine landscape, 2) climate, flow, connectivity, and discharge seasonality, 3) fish dynamics, and 4) flooded area. Antropogenic direct drivers are 5) fisheries, 6) land conversion, 7) water pollution, 7) alien species, and 8) hydropower development. We obtain data for driver’s description from different resources. Information on floodplain geomorphology, tectonic-induced subsidence, and rates of sedimentation were obtained from Tanner (1974), HIMAT (1977), Smith (1986), Latrubesse et al. (2005), Kettner et al. (2010) and Latrubesse (2015). Water level (meters), rains (mm), and Sun bright (hours of light) were obtained from two IDEAM (Instituto de Hidrología, Meteorología y Estudios Ambientales) hydrologic stations in the Puerto Berrio town (Codes 23095010 and 23090110; http://sisaire.ideam.gov.co/ideam-sisaire-web), and total suspended sediments (TSS) (g.ml−1) were measured following the standard operating procedure (gravimetric determination); those data came from the time series between years 2004 and 2005 (Jiménez-Segura 2007; unpublished data).
Water discharge magnitude and variability near major floodplains were derived from satellite-based river discharge images during the 1998–2016 period from the River Watch version 3 system of the Dartmouth Flood Observatory (DFO) at three selected measurement points on the Magdalena and Cauca rivers (Escobar et al., 2016; Restrepo et al., 2020a). Depth of the floodplain lakes are also included (Ramírez and Viña, 1998; Hernández-Atilano et al., 2008; Granado-Lorencio et al., 2012).
Floodplain areas and major physical attributes, including changes in flooding areas and channel avulsion processes are from Patiño and Estupiñán-Suarez (2016). We estimate the total flooded area (km2) of the middle and lower Magdalena basin using Earth observations. The Landsat 5, 7, and 8 satellites when used in combination with one another provide a nominal eight-day interval between observations; though with cloud cover and prioritizations there can be a much longer gap. However, Landsat provides a dataset that can be used to evaluate seasonal and annual fluctuations in open water extent (Restrepo et al., 2020a). We classified every scenario from 1999–2015 into land, water, and bad data (e.g., cloud, shadows, and haze), through the methods described in Potapov et al. (2020). We saved the counts of land and water per month and then aggregated these to obtain the area per season that is inundated for at least 50% of the observations (Pickens et al., 2020). Because the ENSO cycles are definitive in the water discharge in the riverine systems, we explored the relationship between the flooded area in the Magdalena River and the Southern Oscillation Index (SOI) in order to define their association. SOI data came from https://www.ncdc.noaa.gov/teleconnections/enso/indicators/soi/.
Fish dynamics follow environmental guilds (Black non-migratory and White fish migratory) by Welcomme et al. (2006) and reproductive history traits (following Winemiller and Rose 1992: opportunist, equilibrium, and seasonally, with modifications in Jiménez-Segura et al., 2020). Migratory types (White fish) were based on Usma-Oviedo et al. (2013) and López-Casas et al. (2016). Dynamics of water level came from Restrepo et al. (2020a), and fish life strategies in the floodplain lakes came from Jiménez-Segura et al. (2010a). Data about life strategies of potamodromous fish associated with flood pulses was from Jiménez-Segura et al. (2020) and Lasso et al. (2011). Fisheries landings data reported between 2000 and 2014 by the Colombian government were obtained from www.sepec.aunap.gov.co; latest consult was from 12 December 2021) We also constructed a linear model Ln (Catch) = 0.8773*Ln (flooding area (year−1)) + 17.568, which was obtained from the relation between the flooding area and landings data reported in Welcomme (1985) to estimate the expected landings in the Magdalena Basin. We explore the relationship between estimated fisheries landings of the Magdalena River of a year x, with some characteristics of the flooded area, one year before that year x. For each year, we calculate the mean value of the flooded area, their coefficient of variation, the amplitude between the minimum and the maximum value of the flooded area, and the number of days with a value of the flooded area over and under than the mean of the year. As White fish spawns in the Magdalena River are strongly influenced by rainy periods (Jiménez-Segura et al., 2010b), we also explored the relation between ichtyoplankton densities of potamodromous fish with ENSO cycles, the flooded area, and fisheries landings (expected landings) using time series graphs.
Anthropogenic pressures such as water pollution, fragmentation, habitat loss, alien fish species, and fisheries pressure on floodplains were assessed from Galvis and Mojica (2007), Jiménez-Segura et al. (2014), Jaramillo et al. (2015), The Nature Conservancy, Fundación Alma, Fundación Humedales and Autoridad Nacional de Pesca (2016), Angarita et al. (2018), Angarita et al. (2020), Gutiérrez-Moreno and De La Pava (2020), Hernandez-Borrero et al. (2021), López-Casas et al. (2020), Lasso et al. (2020), and Jiménez-Segura et al. (2014) (unpublished data). Hotspots of wetland loss are from Patiño and Estupiñán-Suarez (2016) and The Nature Conservancy, Fundación Alma, Fundación Humedales and Autoridad Nacional de Pesca (2016).
Driver 1: The Magdalena river landscape
The Magdalena River drains an area of 257,438 km2 and is the largest river system in Colombia (Figure 1). Magdalena has two main tributaries, the Cauca River, with a drainage basin area of 66,750 km2 (26% of the Magdalena basin) and the San Jorge River, with a particular monomodal hydrologic pattern (Figure 1). Most tributaries have small drainage areas ranging between 22,000 and 6,000 km2, all of them are responsive to both natural and human-induced change (Restrepo and Syvitski, 2006). The geomorphic setting of Magdalena comprises subsiding foreland areas, an anastomosing river pattern (Figure 2), and tributary systems with high vertical aggradation (Latrubesse et al., 2005).
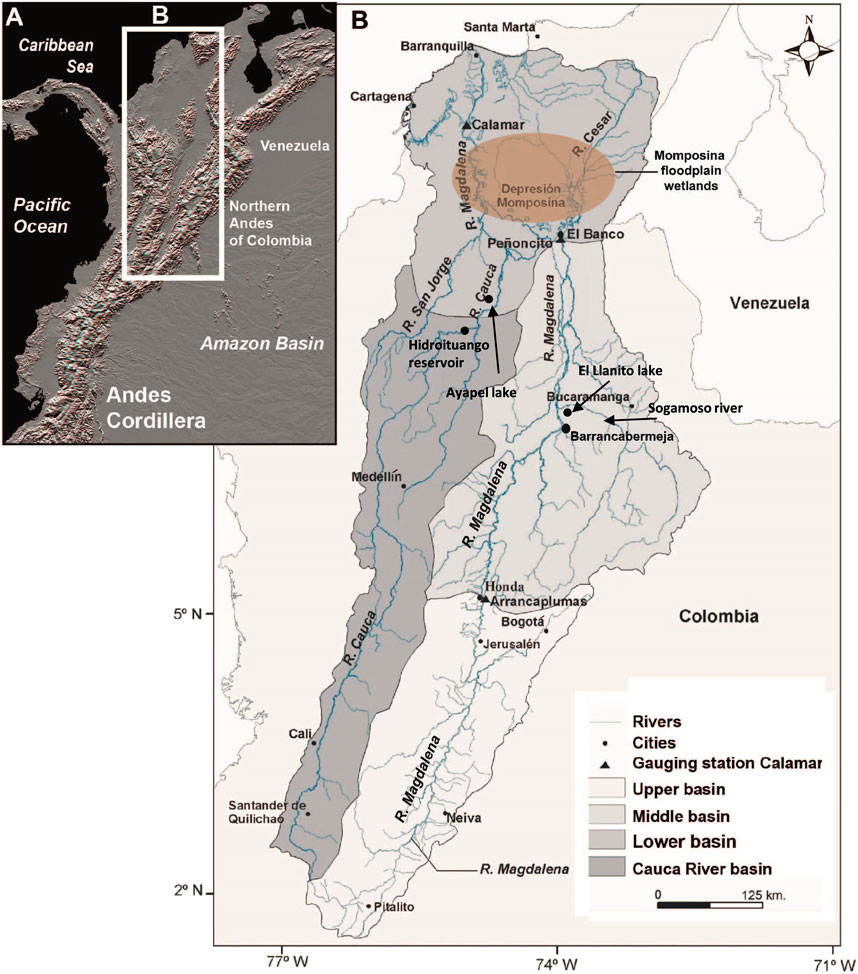
FIGURE 1. (A) Andes of Colombia, showing the location of the Magdalena River basin; (B) upper, middle, and lower sections of the catchment, and the most downstream floodplain wetlands, the Momposina, are also shown.
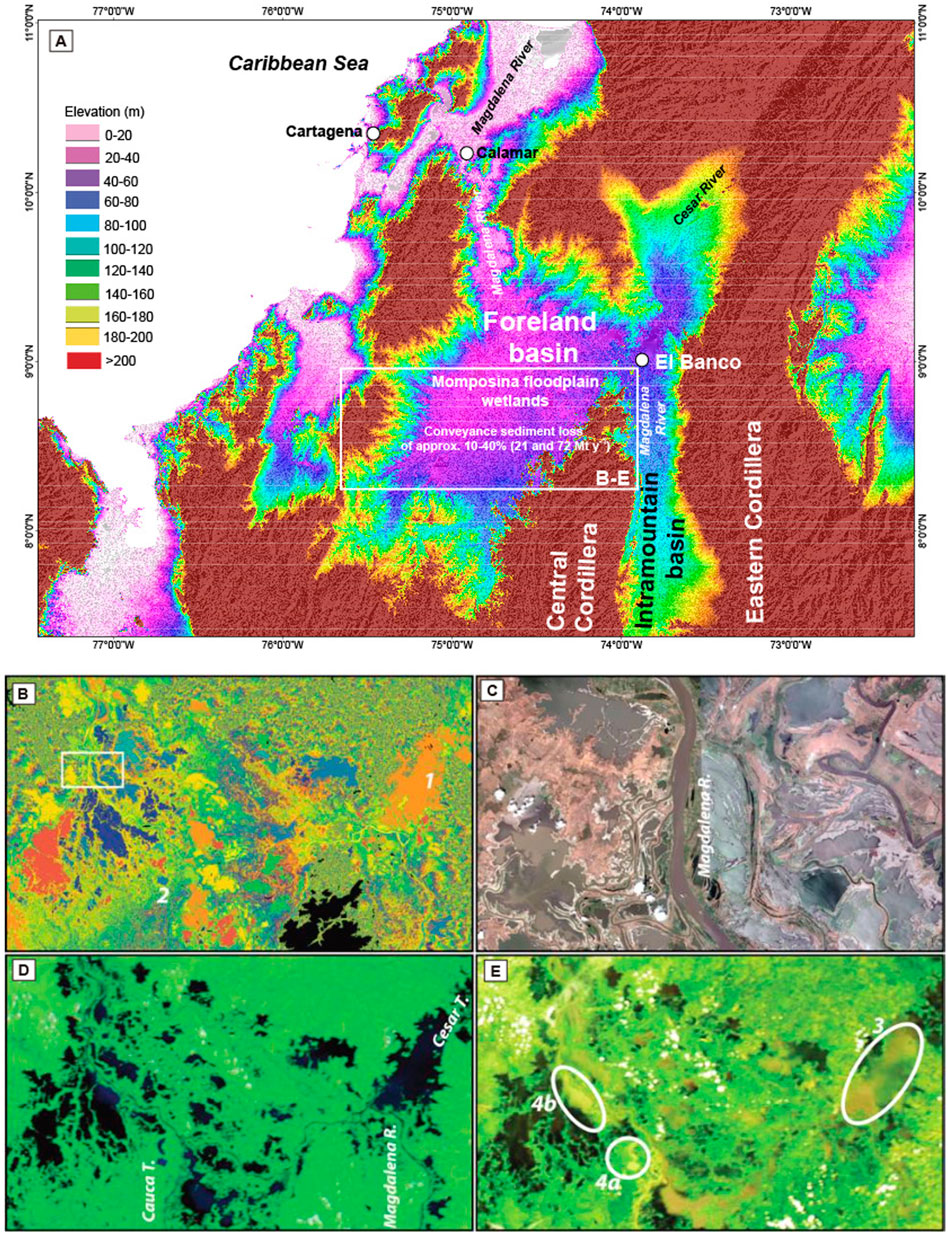
FIGURE 2. (A) Multi-Error-Removed Improved-Terrain DEM of the lower reaches of the Magdalena drainage basin (Data courtesy of Dai Yamazaki, Yamazaki et al., under review). (B,D,E) Representations of the same area and a detailed view (C) of the Momposina floodplain wetlands (modified from Kettner et al., 2010). (B) Display of 1-m vertical intervals of space shuttle radar topography mission data obtained during 11-day mission in February 2000 (i.e., during the dry season). Each color represents a 1-m interval; lakes bordering the Magdalena River as well as the Cesar 1) and the Cauca 2) tributaries are shown. (C) Digital Globe 2009 (Google Earth) detailed view [contents of the white box in (B)] of the lakes created as a result of the tectonic depression. (D) Near-infrared aqua moderate resolution imaging spectroradiometer (MODIS) satellite image during the La Niña event on 10 September 2008. Black shading indicates bodies of water surrounding the Magdalena River and the Cesar and Cauca tributaries. (E) True-color MODIS satellite images [same day and time as image in (D)] clearly showing the sediment plumes in various lakes close to the Magdalena River as well as to the Cesar (3) and the Cauca (4a, 4b) tributaries, indicating that suspended sediment spills into bordering lakes where most of it will be deposited.
Along the river basin there is a high diverse mosaic of freshwater ecosystems (Jaramillo-Villa et al., 2015). The Magdalena River floodplains represent 9.7% of the whole river catchment area. The contributing zones to floodplains are hilly regions with a drainage area of 168,627 km2 (62% of the total basin area). The larger floodplain region with an area of 13,137 km2 is formed in the lower reach of the Magdalena–Cauca rivers system (The Nature Conservancy, Fundación Alma, Fundación Humedales and Autoridad Nacional de Pesca, 2016). The average flooded area in the period between years 2000 and 2014 was 6,532 km2 with fluctuations in the flooded area (range: 1,754–10,474, S.D: 1,634, C.V: 25%). At the lower end of these alluvial plains lies the so-called “Momposina depression wetlands” (Figure 3), the largest avulsive anabranching flood basin with at least 55 m thick of Holocene deposits (Smith, 1986; Latrubbesse et al., 2015). Considering the whole floodplain system, almost 1,900 lakes currently exist in the Magdalena River basin (The Nature Conservancy, Fundación Alma, Fundación Humedales and Autoridad Nacional de Pesca, 2016).
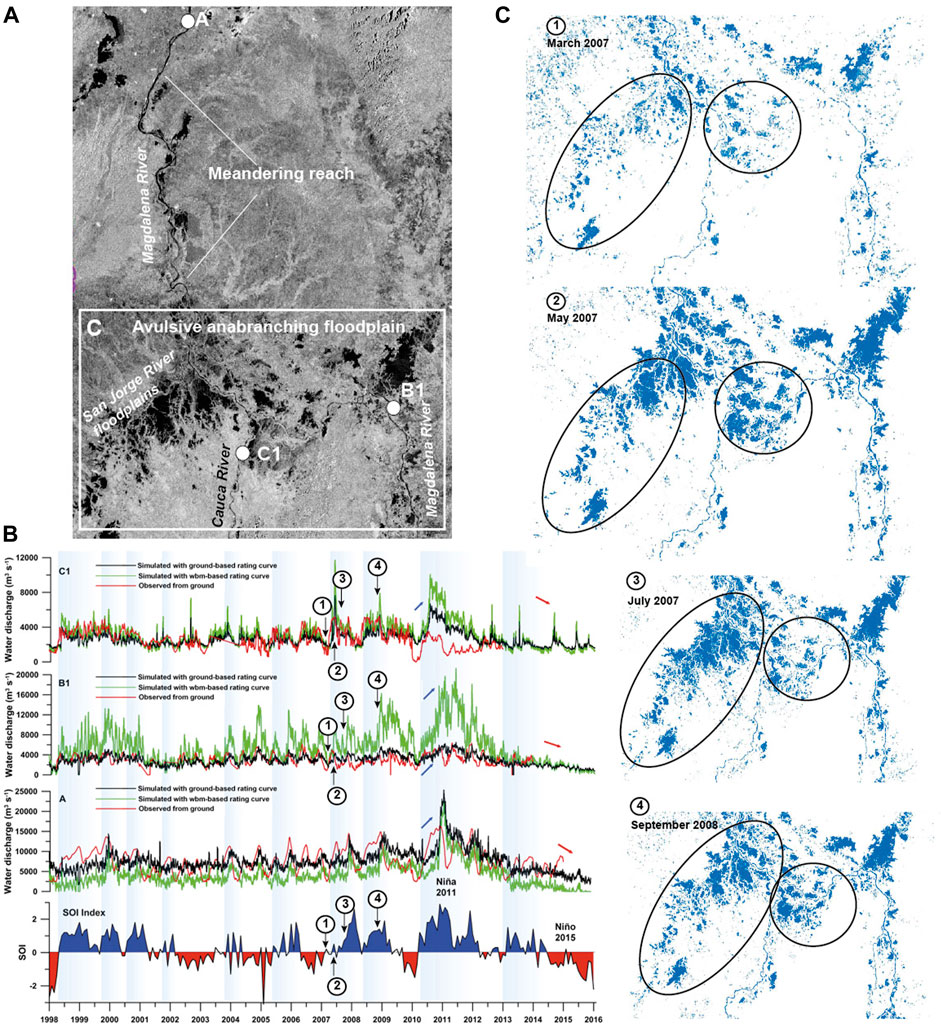
FIGURE 3. (A) Radar image (ALOS-PALSAR) of the lower reaches of the Magdalena River, showing the Momposina depression floodplains and the approximate conveyance sediment loss (images courtesy of The Nature Conservancy, Colombia). (B) Time series of river discharge for the Magdalena River obtained from rating curves using the Water Balance Model (WBM) estimates (green line) and discharge from ground-based gauging stations (black line). We also show ground-based observations of water discharge at each gauging station (red line). The Southern Oscillation Index (SOI) is shown at the bottom (Modified from Escobar et al., 2016). (C) Flooding maps of the Momposina floodplain wetlands during La Niña event 2007–2008 based on ALOS-PALSAR images (images courtesy of The Nature Conservancy, Colombia).
The middle and lower reaches of the Magdalena River occupy two different topographic basins because their sediment deposition rate (Figure 3). Upstream from El Banco, the intramontane basin is bordered by the Eastern and Central Cordilleras. Channels and wetlands extend ∼230 km from the south at Barrancabermeja municipality northward to El Banco municipality in a 25 km wide valley floor. In this middle reach section, Tanner (1974) described these laterally stable channels and associated floodplains as an incomplete floodplain characterized by lakes and marshes and lacking scroll bars and oxbow lakes. Large wetlands, formed by tectonic down warping, indicate that overbank sedimentation was unable to balance tectonic subsidence. At El Banco municipality, Magdalena crosses the partially buried north end of the Central Cordillera before entering the foreland basin of the Momposina depression. Here, the river divides into numerous anastomosing channels. On the southwestern side of the Momposina depression, the Cauca and San Jorge rivers enter from a major valley limited by the Central and Western Cordilleras. The foreland area is oriented parallel with the north–south regional topographic and structural geologic trends. In the northwest part of the foreland basin, all channels converge to form a 100 km long meandering channel reach (Smith, 1986).
Studies on geological evolution of northwestern Colombia indicate that the Magdalena lowlands (Figure 3), the called Momposina depression wetlands, have been moving down or subsiding during the Late Cenozoic. More than 8,000 m of fluvial sediments have accumulated in this sedimentary basin. In addition, during the Andes Cordillera uplift in the Miocene–Paleocene, sedimentary sequences were deposited as a result of tectonism, particularly in the Eastern Cordillera (Campbell, 1974; Irving, 1975; Smith, 1986).
Tanner (1974) suggested that the channels and wetlands were formed by tectonic down warping. According to Smith (1986), basin subsidence is the primary cause of the avulsive anabranching channels and foreland deposits. Thus, ongoing tectonic down warping has produced an actively aggrading depositional environment, consisting of large wetlands or an incomplete floodplain. Based on 14C dates in the Momposina wetlands, HIMAT (1977) found that the rate of sedimentation of 3.8 mm y−1 over 7,500 years was equal to the rate of subsidence, a condition that would favor vertical accretion rather than lateral channel accretion associated with a meandering fluvial model. Overall, wetlands in the Magdalena, covering ∼80%–90% of the foreland depo-basin (Figure 3), consist of open shallow lakes with bottom sediments of clastic mud and marshes with abundant aquatic plants and organic rich mud. Channels, levees, and crevasse-splays account for the remainder.
Lowland creeks, lakes, and connection channels between lakes and the river channel are the main aquatic systems inside the Magdalena River lowlands. Floodplain lakes are between 900–1,000 m and downstream of 250 m elevation. Floodplain lakes are known locally as ciénagas. They are shallow lakes (Z mean = 2.56 m; C.V. = 30%) formed by water accumulation in land depressions. Arias (1985, cited by Welcomme, 1985) defined four ciénaga types according to degree of connectivity to the main river course: type I (a channel connects the lake directly to the main river), type II (a channel connects different lakes with the main river channel), type III (a channel connects the lake to a tributary of the main river channel), and type IV (there is no channel connecting the lake to the main river channel).
Driver 2: Rains, discharge, floods, sediments, and hydrological connectivity
The Magdalena basin is characterized by monsoonal precipitation, with an average rainfall of 2,050 mm y−1. Rainfall regime is bimodal and similar throughout the watershed. There are two wet and two dry seasons; March–May and October–November are high rainfall periods, while December–February and June–September are low rainfall periods. The mean annual water discharge of the Magdalena and Cauca rivers are 7,200 and 2,530 m3 s−1, respectively (Restrepo et al., 2020b). Changes in the sediment concentration discharge and floods are associated with Sun bright and rains (Figure 4). After three or four weeks of the increase in the intensity and frequency of rains (because of the reduction in the Sun bright), water level increases and just in the beginning of the flooding period, there is a sudden increase in the sediments concentration and after the highest flood, they fall.
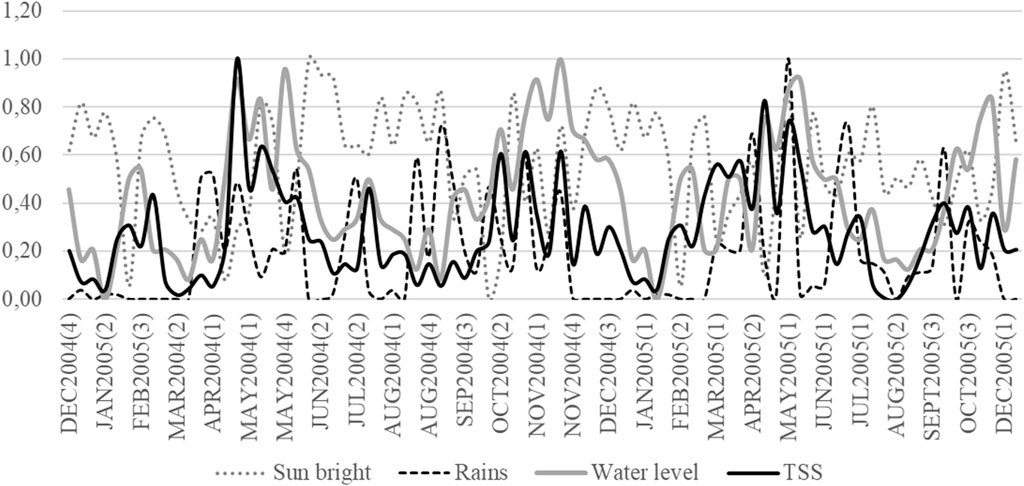
FIGURE 4. Weekly changes in Sun bright, rains, water level, and Total Suspended Solids in the Magdalena River in front of Puerto Berrio city between January 2004 and December 2005. Variables values are movil average for four periods (weeks) and each one was standardized to 0–1 values to let the graphic comparisons. In parenthesis, number of the week in the month.
Drainage basins in the Andes of Colombia (Figure 1) are characterized by high precipitation rates and fast runoff response due to steep slopes (Restrepo et al., 2006a). Regional climate is partly controlled by the occurrence of the El Niño Southern Oscillation (ENSO), which brings dry conditions during the El Niño and wet pulses during La Niña. Inter-annual variability of river discharge shows strong correlations with the ENSO at periods of 3–5 years (Poveda et al., 2001) (Figure 3).
In terms of the hydrologic connectivity between Magdalena River and its floodplain lakes, Restrepo et al. (2020a), based on time series of different connectivity stages using MODIS satellite images, recognized four broad stages of connectivity: 1) river recharge (hydrological connection to floodplain as evidenced by floods), 2) channel-connection between river and floodplain (as evidenced by the change in the water color in the lake), 3) receding flows toward disconnection, and 4) disconnection. During the 2000–2017 the % time of river-floodplain connectivity in the Momposina depression floodplain was 13%. El Niño–Southern Oscillation (ENSO) is a major forcing mechanism of climatic and hydrological anomalies (Glantz, 1997). In tropical South America, particularly Colombia, interannual hydro-climatology is strongly influenced by extreme phases of ENSO (Poveda, 1997). Long-term rainfall analysis confirms that El Niño events are associated with negative rainfall anomalies and La Niña with positive anomalies in northern South America (Poveda and Mesa, 1997). During La Niña years, the positive phase of the ENSO, annual connectivity times increased to 38%. In contrast, the El Niño years witnessed low connectivity time percentages down to 5% (Restrepo et al., 2020a). Further interpretations are based on data and studies focusing on floods, sediment budget, and dispersion.
The lower course of the Magdalena–Cauca Rivers has been strongly affected by floods and associated sedimentary fluxes during the last decades due to human induced erosion in the upper basin. Based on satellite-measured data during the 1998–2016 period at the two most downstream hydrological stations, before the confluence with the Momposina floodplain wetlands (Figure 3), discharge variability shows a robust relation with the ENSO cycle (Figure 3C). Peak water discharges were clearly observed during La Niña events in 2007–2009 and 2010–2011. Thus, high discharges are in close agreement with the observed positive values of the Southern Oscillation Index (Escobar et al., 2016; Restrepo et al., 2020b).
It is worth noting that during La Niña event 2007–2009, the Magdalena floodplains exhibited large variability of hydrological connectivity in terms of flooding areas (Figure 3C). According to Smith (1986), the San Jorge River, characterized by low rates of sediment transport and channel gradient, is a stable channel with prominent levees and minimum avulsion activity. Thus, in many parts of the southern region of the Momposina floodplains, rates of subsidence appear to exceed sedimentation, a condition that causes more lateral connectivity and extensive flooding of wetlands.
Sediment trapping in large river systems is very common. A significant amount of the suspended sediment never reaches the river mouth, as it becomes sequestered in riverbeds, on floodplains, and in reservoirs (Dunne et al., 1998; Goodbred and Kuehl, 1998; Aalto et al., 2002). Once the Magdalena River crosses the floodable area of the Momposina floodplain wetlands (Figure 3), overflow and levee failures cause extensive flooding from April until November, and sediment has been deposited over the last 7,500 yr at a rate of 3–4 mm y−1, in places creating 130-m thick deposits (Smith, 1986; Plazas et al., 1988; Bray, 2006). On the basis of 27 years of observation, Restrepo et al. (2006b) estimate that 14% of the total annual Magdalena sediment loads are trapped in the Momposina floodplains (Figure 2).
In the Momposina wetlands not all lakes are connected with the main river channel even during peak discharge periods. Depending on location and volume, lakes trap sediment as well. Lakes situated in the lower reach of the main stem of a river or tributary tend to capture more sediment than do lakes situated high in the hinterland. The Momposina floodplain lakes form a flooded area for up to 8–9 months of the year. The water emptying process of these lakes is not simultaneous because they have different degrees of river channel–lake connectivity (e.g., lakes types II and III; Figure 3). True-color MODIS satellite images clearly show the sediment plumes in various lakes close to the Magdalena River as well as to the Cesar and the Cauca tributaries, indicating that suspended sediment spilled into bordering lakes during the La Niña event in September 2008 (Figures 3B–E), where most of it will be deposited (Kettner et al., 2010).
Sedimentation rates (Plazas et al., 1988) and an analysis of daily moderate resolution imaging spectroradiometer (MODIS) images (Figure 2) support that these flooded areas trap significant amounts of sediment. Landsat and MODIS images indicate that 25%–33% of the area of the Cesar tributary gets flooded every year. Assuming average deposition rates of 3–4 mm y−1 (Plazas et al., 1988), ∼24–45 Mt y−1 of sediment is trapped on the floodplains of the Cesar tributary given a density of 1,700–1,800 kg m3. Approximately, 17–72 Mt y−1 of sediment are trapped in the lower reaches of the Cauca tributary (Kettner et al., 2010) (Figure 3).
The magnitude of yields for the northern and central regions of the Andes, including the Colombian Cordilleras, is equivalent to rivers draining other orogenic belts in Asia, Insular Asia, and New Zealand, and one order of magnitude larger than yields reported for the Alps (Latrubesse and Restrepo, 2014). Transporting a sediment load to the ocean of 184 Mt y−1 the Magdalena River ranks as the highest sediment-yielding river (∼714 t km−2 y−1) among the large rivers that drain South America (Restrepo et al., 2015).
The abovementioned magnitudes of sediment production in the Andes of Colombia and along the main fluvial system, the Magdalena River, highlight the conditions of sediment trapping occurring in the Magdalena floodplains (Figure 3), which together with the hydrologic variability are the main drivers of floodplain connectivity.
Previous findings estimate that the foreland basin-trapping zone of the Momposina depression may store ca. 21 Mt y−1 of suspended sediment (Restrepo et al., 2006b), while an estimated 53% of the incoming bedload is deposited in the Magdalena foreland (Smith, 1986). The sediment storage in the active sedimentary basin of the lower Magdalena, between 21 and 72 Mt y−1, is of the same magnitude as the sediment trapped into the Pantanal from the Brazilian highlands (ca. 8 Mt y−1 or near 50% of the total sediment entering the region) and sediments deposited in the middle Paraná floodplain in Argentina (17–60 Mt y−1; Latrubesse, 2015).
Natural distribution and deposit of sediment from the highlands to the lowlands have reduced the depth of the main river channel, and this is not a convenient characteristic for the river freight transport because it limits the freighter capacity and size moving along the Magdalena River up from Bocas de Ceniza Port, near Barranquilla city (Caribbean Sea). In different moments in our history, the Colombian government have tried to modify this situation using drags in some sections along the river course, moving from sand from one side to another indefinitely, as a Sisyphus curse. In 2021, the Colombian Congress approved in a first instance a new law (Number 298; Senado de la Republica de Colombia 2020) declaring the Magdalena River channel as a transport highway; the first step for modifying the river as the freight business needs. Now, the transport Colombian authority is promoting a new project to improve the river navigability; the first indirect driver: fluvial transport business. This project is planned to use lateral dikes and drags to redirect the river flow to the river main channel (Instituto Alexander von Humboldt, 2019). Sadly, most of the areas where the sediment accumulates are near to the connection channels with the floodplain lakes. These actions will decrease connectivity between the river channel and these lakes with important changes in nutrient flux and fish larvae drift and inputs in to these nursery areas. Fortunately, the citizen science process involving fishers had improved their opportunities for identifying the negative process that can modify the Magdalena River and the fisheries yield, and by now this project is going to be demanded at the Colombian Congress because the damage it could cause if adequate management strategies are not incorporated.
Driver 2: Fish dynamics in the Magdalena River basin
In the Magdalena River, there is a strong turnover in the fish richness elevation gradient (Jiménez-Segura et al., 2014); the higher diversity is in the lowland creeks and the lower diversity in reservoirs (Jiménez-Segura et al., 2020). Fish assemblage in the lowlands is highly diverse in each of the trophic group, reproductive traits strategy, and swimming/water column position (Marín-Avendaño, 2012; Jiménez-Segura et al., 2020), and migrations of some fish species from the lowlands to the hills during each dry period generate seasonal changes in the fish abundance in the river network and an amazing artisanal fisheries activity, mainly in the lowlands. Jiménez-Segura et al. (2020) identify that there are 22 functional niches in the Magdalena River basin, most of them are in creeks and lakes in the lowlands. Because of their role in the food webs in the aquatic systems (Winemiller et al., 2004; Reis et al., 2020), fish can be considered as a driver in the dynamics in the flux of carbon and energy between habitats along the river basin; even, some fish species can be considered as ecosystems engineers (Flecker 1996). In the Magdalena River, the fish role in the dynamic and structure of the aquatic freshwater ecosystems have been poorly explored.
Flood pulse and fish response in the lakes
Reproduction of fish in the Magdalena River is synchronized with environmental cues. The number of hours of daylight, the flood pulse attributes (magnitude, frequency and duration), and some water characteristics (conductivity and temperature) are cues for activating each reproduction phase of the White fish (Jiménez-Segura, 2007). In the floodplain lakes, black fish ripe during transition from low water to high water conditions (Jiménez-Segura et al., 2010a). Change in the river connectivity influence the water volume in the floodplain lakes due to shifts in discharge, nutrient inputs, and sediment magnitude. Flooding, floods, and dry periods are critical for biological productivity and fisheries production in the floodplain lakes. Along the hydrological cycle, the Ayapel Lake, an aquatic system inside the Momposine depression (Figure 1), expands and reduces its area because riverine inputs; its area changes from 120 km2 in the high waters (mean deep of five m) to ∼50 km2 in the dry period (mean deep of three m) (Restrepo et al., 2006a). Research on the response of some aquatic biota in this lake has demonstrated the influence of the flooding on their densities.
Jiménez-Segura et al. (2010a) described the influence of the hydrological cycle in the trophic guilds of fish fauna in Ayapel Lake. The welfare condition (relationship between fish length and weight, factor k) of omnivore fish (including insect preferences) is observed during high water periods. The higher food supply contained in the root biofilm of these floating macrophytes could explain the importance of the high-water period on the fish condition factor. During the descending water period, Ayapel Lake reduces its area, White fishes move to the river channel, and the floating meadows are flushed out from the lakes. The condition factor of carnivore guild increases during the hydrologic transition from high to low water levels and in the floods, when the zooplankton supply to the pelagic areas is at highest levels (Gallo-Sanchez et al., 2009). Transitional periods along the hydrological cycle have been poorly studied in the Magdalena River. The flooding period is of high importance, it not only contribute with water, nutrients, and sediments to the lateral plain but also fish embryos and larvae, resulting from the previous spawning of White fish species in the main river channel (Ochoa-Orrego et al., 2015).
In the low water period, floating macrophytes trapped in the lake begin to decompose, and coupled with wind stress, sediments from the bottom liberate nutrients into the water column for planktonic organisms; a process that increases planktonic densities (Hernandez-Atilano et al., 2008). Low water periods are characterized by a high concentration of fish in the water column of the Ayapel lagoon and associated tributary creeks (Ríos-Pulgarín et al., 2008) and by the growth of vegetation on subaerial lands. The condition factor of the carnivorous fish increases as densities of black fish increases in the reduced water volume. Without any exception, all fish guilds inside the lake ripen their gonads toward the end of the low water period and during the filling of the lake (rising water period).
During rising waters, water transparency in the Ayapel Lake is reduced due to sediment input from the river and the decomposition of flooded vegetation. Larvae from Black and White fish find plenty food in the lakes as zooplankton densities (Gallo-Sanchez et al., 2009) and phytoplankton bloom increases (Hernández-Atilano et al., 2008). This scenario coupled with the growth of floating macrophytes provides optimal habitat structure for the survival of the new cohort of fish since there is available food of different sizes and protection in the roots of the floating macrophytes.
Granado-Lorencio et al. (2012) found that the distribution of the fish in the Magdalena floodplain lakes is highly nested. Roots of floating macrophytes are habitat for fish larvae, and the flushing out of floating macrophytes from the lake to the river and their drift to lower lands during the descending water period could be an important driving force for this nested pattern in the Magdalena floodplain lakes. Although fish drift with floating macrophytes has already been reported in the Amazon and Paraná rivers (Schiesari et al., 2003; Bulla et al., 2011), little about this is known in Andean rivers.
Granado-Lorencio et al. (2012) found that the similarity in the assemblages of non-migratory fish present in 35 floodplain lakes was not related to the distance in kilometers between them; those distant lakes have an almost identical species composition. We hypothesize that floodplain contraction and expansion of the Magdalena River is the cause of this nested fish assemblage in the floodplain lakes. We base our hypothesis in two evidence: the reproduction of the fish present in the flood lakes occurs during the beginning of the flood of the lateral plane (Jiménez-Segura et al., 2010a), and Jiménez-Segura (2007) found that about 30% of the species that formed the drifting ichthyoplankton were not White fish species. We suggest that the overflow and contraction of the river on the lateral plane may induce the accidental drift of larvae of non-migrating species toward areas downstream of their birth lake, and during periods of low flooding, adults of these non-migrating species may go out into the river and disperse upstream, looking for lagoons with better conditions to survive. River expansion and contraction over its lateral plane and its influence over the fish species composition in floodplain lakes along the river channel is a hypothesis that needs to be validated with research that identify which fish are under the floating macrophytes that are drifting downstream during the flooding period and the species composition in the ichtyoplankton, also drifting in this same hydrological period.
Fish migration cycle
White fish species in the floodplain lakes migrate among four different aquatic systems twice per year in synchrony with rainy periods: floodplain lakes, connection channels between lakes and the main river channel, and the main river channel and tributaries to the main river channel (Figure 5). López-Casas et al. (2016) using tagging and recapture strategies estimated the movement distances between 50 and 1,200 km.
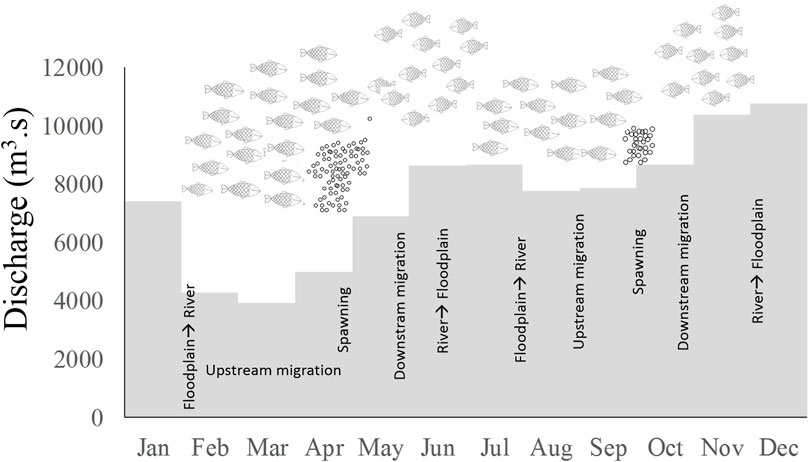
FIGURE 5. Migration cycle of potamodromous fish in the Magdalena River and hydrologic seasonality. Gray bars: Mean of the monthly discharge in the Magdalena River in the period 2003–2014. Adapted from Jiménez-Segura et al. (2020).
As rain diminishes, Sun brightness on the water increases (December), and fish use this as a cue to migrate from lakes to main channels. Upstream migration along the main river (known by the fishers as the subienda migration) is carried out during the first annual low water period (45–60 days, January–February). Migratory fish get into the tributaries and stay there for 30–45 days (López-Casas et al., 2016). Prochilodontid fish such as bocachico Prochilodus magdalenae (Steindachner, 1879) travel up to 1,000 m elevation and Pimelodidae species such as Nicuro Pimelodus yuma (Villa-Navarro, Acero, and Cala, 2017) to 500 m. During the subienda, López-Casas (2015) found that the fishers’ catch behaves in pulses; landings may be the same for a couple of days, and then the following day drops to half the number of fish. This process may suggest that not all the potamodromous fish migrate to the river at the same time as not all floodplain lakes dry at the same time. Migrations from the lakes and upstream to the rivers are carried by groups of fish that leave from each lake at different times during the end of the high water and low water periods (fishers name them punta). To test the hypotheses about pulsing migration from each lake in the Magdalena River more research using telemetry methods is needed in order to describe movements in different moments along the hydrological cycle. Telemetry studies have demonstrated to be effective to describe seasonal movements of migratory fish in South America in order to take management decisions elk dam construction (Hahn, 2012).
With the incoming rains at the start of the rainy season, the river water level increases (March and April) and potamodromous fish spawn Jiménez-Segura et al. (2007). Icththyoplankton densities pulse many times during the flooding period Jiménez-Segura et al. (2010b), Jiménez-Segura et al. (2016); these evidence are probably a consequence that not all migratory fish spawn at the same time because not all fish migrate at the same moment (Punta groups). To improve the knowledge of this interesting behavior, we need more research on long term monitoring of ichthyoplankton densities, integrating telemetry research to follow the migratory routes, and seasons of White fish in the Magdalena basin.
During the peak floods (May and June), embryos and adults drift downstream from the spawning site (fishers name it as Bajanza migration); embryos are incubated during this drift. Adults come back into the floodplain lakes, and fish larvae are passively dispersed into the lake with the floods (Ochoa-Orrego et al., 2015). In July, during the second dry period of the year, a second migration (Mitaca migration) is activated, and migratory and spawning cycles initiate again. Although changes have been found in the ichthyoplankton densities between Bajanza and Mitaca (Jiménez-Segura 2007), the differences in density migration are poorly understood because densities of migratory fishes have not been evaluated. An integrated research using migratory fish abundance and spawning intensity (ichthyoplankton densities) in different areas in the Magdalena basin is needed. Recent analyses about spawning areas in the basin have identified that the higher densities of fish embryo and larvae of migratory fish were observed in those tributaries with a high developed floodplain area and longer stretch of the river channels with a low slope gradient (Moreno et al., 2021).
Driver 4: Flooded area
The flooded area has been associated with biomass production in rivers (Welcomme, 1985) and richness (Reis et al., 2016). The linear model on variations of the flooded area vs. fish landings of the artisanal fishermen proposed by Welcomme (1985) estimates expected landings that are different from those reported by the Colombian government (Figure 6). The Colombian authorities have been monitoring the inland fishery landings during the last 40 years. Colombia is perhaps one of the few countries in South America that maintains monitoring landings despite government changes. In spite of the efforts, the low investment of each Colombian government in the monitoring budget limits the effectiveness in the sampling design to obtain the data of the continental fishing landings in the country and it may affect the accuracy of the collected information.
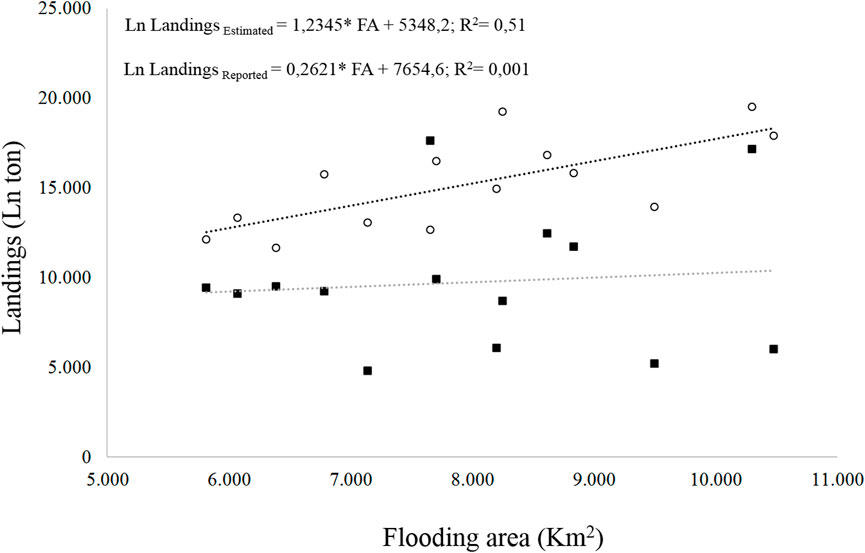
FIGURE 6. Relation between the flooded area and the landings of the freshwater fisheries (estimated empty circle, reported black square).
Our analyses show that the maximum-flooded area and the time of flooding before each yearly fishery period explained 42 and 37% of the fishery landings, respectively; other dispersion variables explain 30% or less (Figure 7). Although these two relations are important, they do not explain more than 50% of the expected landings. To improve the explanation power in the model proposed by Welcomme (1985) to estimate the fisheries landings in the Magdalena River basin could be important to include in the model other information as nutrients and sediment concentration in the water, chlorophyll concentration in the floodplains lakes, and hydrologic connectivity of the fluvial network.
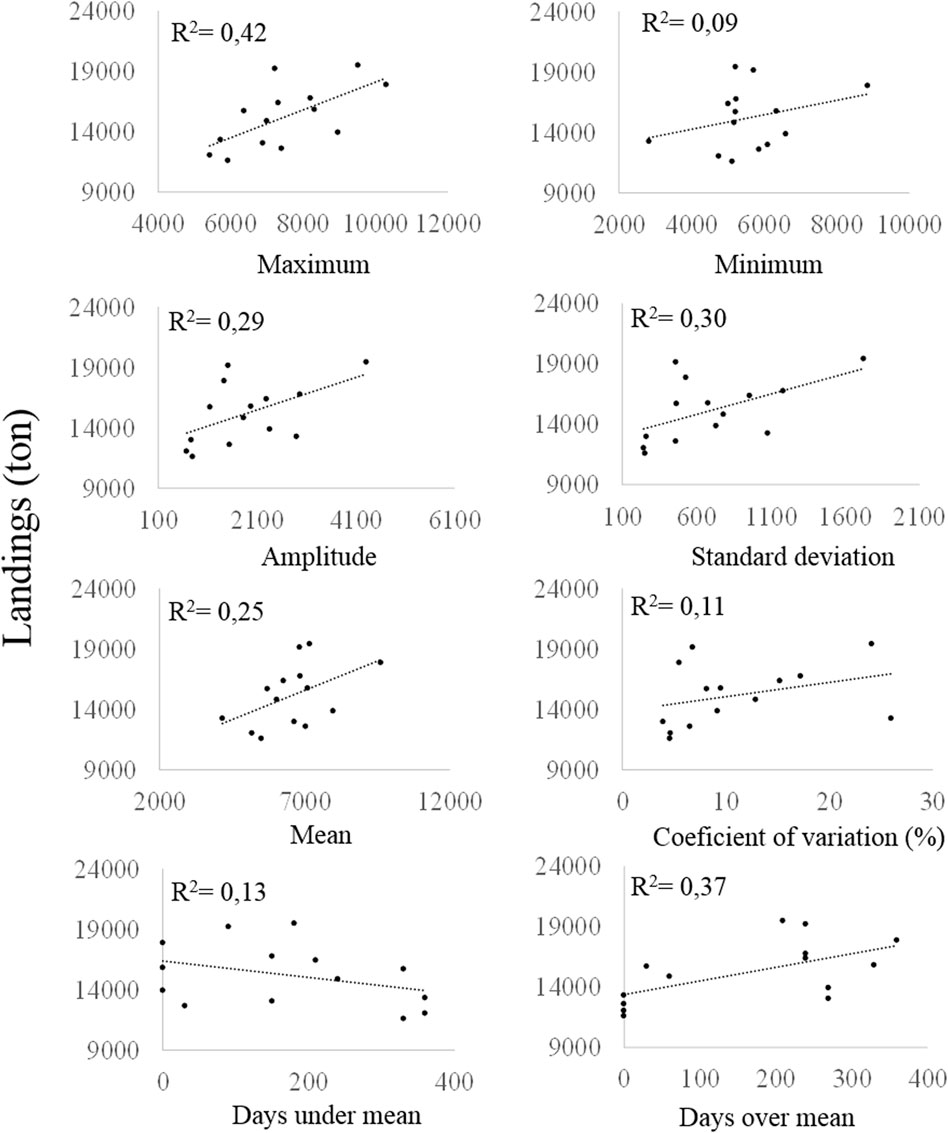
FIGURE 7. Data characteristics of the flooded area and fishery landings (estimated) in the Magdalena River in years 2001–2014.
Water retention for irrigating crops in the lowlands have reduced the discharge of some tributaries (e.g., Cesar River) to the main channel of the Magdalena River and diminishes the flooded area in the lowlands as reservoirs do in those regulated basins with lakes in their lateral plains (Angarita et al., 2020). Those human actions can be considered the second indirect driver influencing not only the flooded area but also the nutrient and carbon inputs in the Magdalena floodplains.
Ichthyoplankton density and expected fishery landings in the Magdalena River were associated to with ENSO cycles and river-flooded area (Figure 8). After peaks in the density of ichthyoplankton and the increase in the flooded area, the landings of the fisher increased two years after. This suggests the high influence of the higher floods on river fisheries productivity. Research on fish larvae in the Parana and Mekong rivers (Baumgartner et al., 2004; Tung et al., 2006) has similarly identified the time and the areas for White fish spawning, but little is known about how they are linked over long-term periods to promote fisheries production.
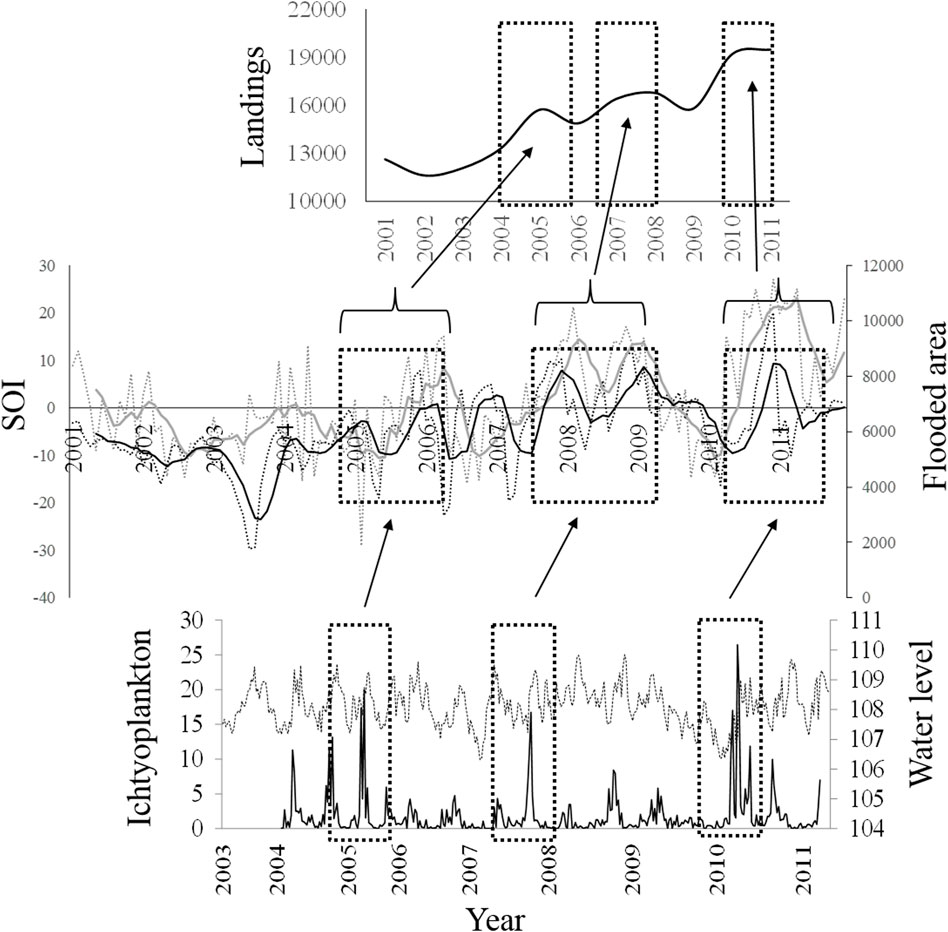
FIGURE 8. Relationship between (A) fisheries landings between years 2000 and 2014, (B) SOI values and flooded area (SOI values: monthly-gray pointed line, moving average for five periods- black pointed line; Floodplain area values: monthly-gray line, moving average for five periods-black line), and (C) ichthyoplankton densities (individuals.m-3, black line) and water level of the Magdalena River (pointed line).
Driver 5: Artisanal fisheries
In 2012, ∼30% of Colombia’s population (47 million people) lived in the floodplain regions, 11% lived in poverty, and 0.3% was fishers (The Nature Conservancy, Fundación Alma, Fundación Humedales and Autoridad Nacional de Pesca, 2016). Fisheries in the floodplains of the Magdalena–Cauca basin are mainly artisanal and for subsistence. Most of the boats used in lakes are wooden and utilize paddles for propulsion, and the most frequent fishing methods are gillnets, cast net, and hooks (AUNAP-UNIMAGDALENA, 2014; Hernandez-Borrero et al., 2021). Fish migrations are extremely important to these riverine human communities, with almost 35,000 fishers and 130,000 people depending on it. They use fish as the main protein source in their diet and for their economic support. In 2010, the economic value of fishery production was ∼24 million dollars (The Nature Conservancy, Fundación Alma, Fundación Humedales and Autoridad Nacional de Pesca, 2016). In addition, there is a strong culture around the fisher life associated with this incredible biological cycle that is reflected in Colombian folklore music, food traditions, and the amphibian way of life (Jaramillo-Villa et al., 2015a).
Fisheries of White fish are implemented in aquatic systems where migrating fish stay: in the floodplain lakes during floods, in the connection channels during the ascending or descending hydro-periods, and in the river channel during the low water period (The Nature Conservancy, Fundación Alma, Fundación Humedales and Autoridad Nacional de Pesca, 2016). Groups of fishermen move along the different aquatic systems, following the puntas. Although there is a synchrony between fishery landings and migration of fish throughout the year, fish catches are permanent in the floodplain lakes. In the Magdalena River, ∼55% of the yearly landings come from the subienda migration, and in this migration, 20% comes from floodplain lakes (Figure 9A; INPA, 1998; INPA, 1999). Based on fisher landings data from the Sogamoso River and its floodplain lake (El Llanito lake) (Figure 1) during 1995 (INGETEC S.A Ingenieros Consultores, 1998), it was estimated that 45% came from the lake; this percentage changed monthly because of the interaction between river discharge and fish migration (Figure 9B).
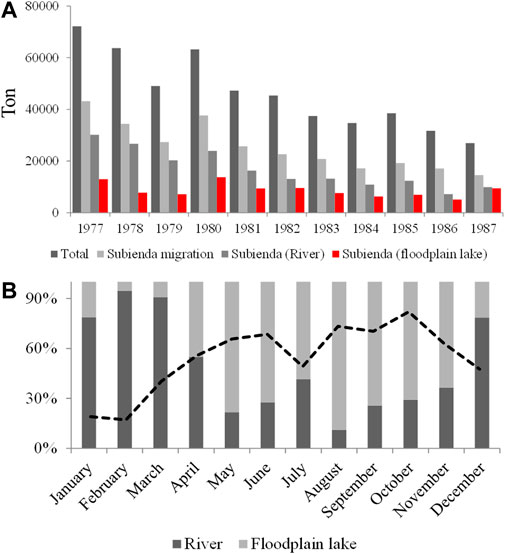
FIGURE 9. Fisheries landings of the Magdalena River. (A) Yearly report from the river channel and floodplain lakes from 1977 to 1987 years; (B) Monthly proportion in the catches obtained by aquatic systems (river, lake) in the year 1995; dotted line: river discharge trend. Data obtained from INPA (1998), INPA (1999).
Forty fish species inhabiting lowlands are recruited by artisanal fisheries (1/4 of the total number used in the basin; Hernandez-Barrero et al., 2021). At present, the bocachico P. magdalenae landings represent between 80 and 95% of the total and other fish species such as Bagre rayado Pseudoplatystoma magdaleniatum (Buitrago-Suárez and Burr, 2007), barbudo Pimelodus yuma (Villa-Navarro and Acero P., 2017), capaz Pimelodus grosskopfii (Steindachner, 1879), blanquillo Sorubim cuspicaudus (Littmann, Burr, and Nass, 2000) and dorada Brycon moorei (Steindachner, 1878), accompany the catches of fishermen (Gutiérrez et al., 2011). Bocachico has provided the highest percent of biomass according to government reports on fishery landings since 1975 and it is recruited along the year (Figure 10). Oreochromis niloticus, an African fish species, has been successfully introduced in the floodplain lakes and reservoirs, and it was reported as the third fish species recruited in the fishery in these aquatic systems after P. yuma (The Nature Conservancy, Fundación Alma, Fundación Humedales and Autoridad Nacional de Pesca, 2016).
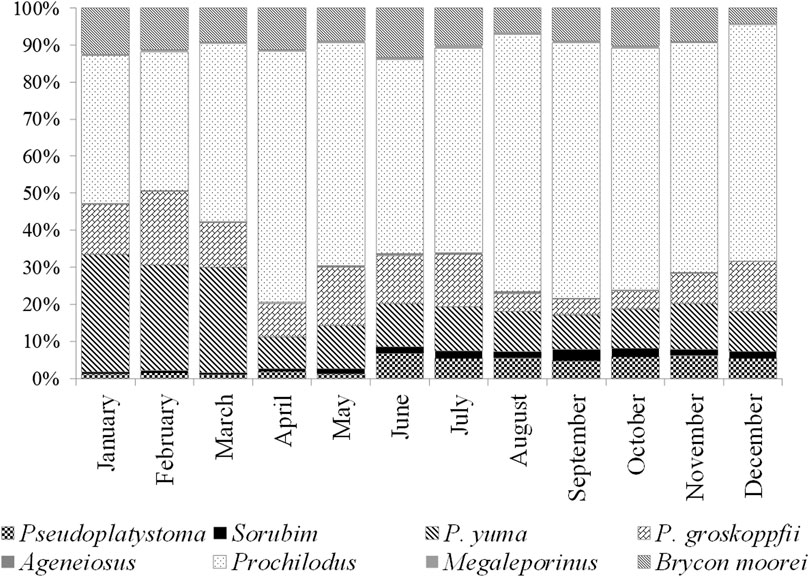
FIGURE 10. Percent of each potamodromous fish species to the monthly landings in the Magdalena River basin from 1993 to 1999. Black line: annual average tons. Data obtained from INPA (1998), INPA (1999).
Floodplain lakes produce ∼50% of the freshwater fishery landings in the Magdalena River. Artisanal fishery landings have been depleted to one tenth in the last four decades due to human-induced floodplain changes and fishery pressure (Jiménez-Segura et al., 2016; Hernandez-Barrero et al., 2021). Some causes have been identified including: overfishing, land use, water pollution, alien species, and hydropower generation. Between 1978 and 2010, the capture by unit of effort has been reduced by half (25,5 to 12,1 kg.day−1. boat−1), the catfish Bagre rayado P. magdaleniatum landings have been halved, and the size of capture have fallen from 890 to 450 mm, the number of species recruited to the fishery has almost doubled, and the new recruited species are mainly black fish (Agujeto Ctenoluccius hujeta, moncholo Hoplias malabaricus, Mojarra azul Andinoacara latifrons, Cucha Hypostomus spp.), with an opportunistic life strategy (low fecundities, parental care, and small body size) (Valderrama et al., 2016). Evidence of fish overexploitation in general and specifically on the most important fish species to fisheries is worrisome. From 1996 to 2007, the size of captured of Bocachico had decreased from 260 to 230 mm, the average age of capture had reduced from 1.6 to 1.3 years, and the mortality by fisheries (F value) has been increased three times (Valderrama et al., 2016). Between 2003 and 2009, the ∼65% of captured individuals were under the legal minimum catch size (230 mm LS), fishery effort was mainly concentrated in young individuals (Valderrama-Barco et al., 2011). If this fish species is the main support for the Magdalena artisanal fisheries, certainly, the food security and the culture of the artisanal fishermen are highly threatened. Fortunately, fisher´s self-regulation has demonstrated good results in the Magdalena river fisheries. Hernandez-Barrero et al. (2021) identified three periods in the fisheries management in the Magdalena River: fishers were regulated by government measures (years 1975–1984), fishers detect the need for changing the control measures because they were failing (years 1985–1992), and the own fishers have been self-regulating their catches (years 1993–2015). In this third period, fishers’ organizations have been promoting the conservation of aquatic ecosystems in order to protect fish biodiversity and their fisheries production. Fisher´s capacities have been favored by activities around citizen science promoted by some academic and research institutions and non-governmental organizations. This change in their minds can be the third indirect drivers: citizen movements for protecting nature.
Driver 6: Land use conversion
Human-derived factors impacting wetlands in Colombia have been shown by Patiño and Estupiñán-Suarez (2016). More than seven million hectares of wetlands have land use classes related to human activities, which correspond to ∼24% of the existing wetland area in the country. The main human induced activities responsible for wetland loss are cattle ranching, agriculture, and deforestation (Galvis and Mojica, 2007), which together account for ∼95% of the wetland loss (Patiño and Estupiñán-Suarez, 2016). The main hotspots of wetland loss in Colombia are in the Magdalena River valley and its main tributary, the Cauca River; a loss of 88% of the wetland area between 1950 and 1980 was reported in the Cauca River floodplains (Beach and Luzzadder-Beach, 2012; Patiño and Estupiñán-Suarez, 2016). There are changes in the flooded area because land use conversion has been suggested as a potential cause on the fall in the fishery landings in the last five decades (Hernández-Barrero et al., 2021). Political interest in the agroindustry (e.g., extensive cattle farms and African palm crops) can be identified as a fourth indirect driver modifying the extension of the flooded area.
Driver 7: Water pollution
The Magdalena basin provides 80% of gross domestic product in Colombia (The Nature Conservancy, Fundación Alma, Fundación Humedales and Autoridad Nacional de Pesca, 2016). Half of towns in the Magdalena River basin put their sewage waters into streams and rivers without any previous treatment and because of the economic development of the basin, the condition of the water is frequently modified due to the discharge of hydrocarbons, organochlorines, organophosphates, heavy metals, organic matter, fecal bacteria, and new contaminants from different sources (oil industry, agriculture, gold mining, and fur and clothes manufacturing) (Gutiérrez-Moreno and De La Pava (2020)). Heavy metals such as lead, cadmium, arsenic, mercury, and chromium have been detected in tissues of the Magdalena River fish (Alvarez et al., 2012; Mancera-Rodríguez and Álvarez-León 2006), with already detected changes in the health of the human population (Vargas-Licona and Marrugo-Negrete, 2019). Recent advances have found metil-parabens in the water of the Magdalena River (Cacua-Ortiz et al., 2020), and some research works on fish metabolism of the Bagre rayado have detected changes with new contaminants (triclosan and ibuprofen). Gallego et al. (2021) found that exposition to them modifies the energy production, the immune response, architecture and cellular physiology, and also liver damage of Bagre rayado.
Driver 8: Alien fish species
Due to the high number of alien species diversity, the Magdalena River can be considered like a Babylonia tower; 43 fish species (e.g., Oreochromis spp., Tilapia nilotica, Trychogaster pectoralis, and Pangasionodon hypophthalmus) have been reported (Lasso et al., 2020). Some of them escape from fish farms, others were translated by sport fishers, and others came from “liberations” of aquarium fish. Stocking has also been considered as an activity that introduces alien fish because of the genome divergence generated by the inbreeding problem in the fishery farms (Lasso et al., 2020). The Aquaculture and Fishery Authority in Colombia have found an important resource to improve freshwater aquaculture in our country along the last five decades in the foreign fish species but their impact in the current fish communities is unknown. Although some research advances had tried to delimitate the impact of some of these alien species as a way to stop their fish culture (Lasso et al., 2020; Castellanos-Mejia et al., 2021), there are strong political’ pressures in the Colombian Congress to legalize the cultivation of some of them (e.g., Pangasiodon hypophthalmus) and others have been declared by law as “domesticated fish” (Onchorhynchus mykiss and Tilapia nilotica) in the AUNAP Resolution Number 2287 (Autoridad Nacional de Acuicultura y Pesca AUNAP, 2015). The past indirect driver (Politics interest) also influences the promotion of the introduced fish culture in reservoirs, with under quantified consequences in water pollution.
Driver 9: Hydropower development
Dams for hydropower in the Magdalena basin since 1987 were built downstream of 600 m elevation (Jiménez-Segura et al., 2014). Because of elevation, most of these dams cause major changes in spawning conditions (migrating routes to spawning areas and flood pulse as a cue for spawning for White fish, isolates fish populations, turn over the original fish assemblage structure, and reduce the influence of the floods on lateral floodplain because of regulation of floods (Jiménez-Segura et al., 2014; De Fex-Wolf et al., 2019; Moreno et al., 2021; Valencia-Rodriguez et al., 2022). Reservoirs also retain between 20% and −40% of sediment transported from the Magdalena drainage basin (Vörösmarty et al., 2003; Angarita et al., 2018). Thus, sediment retention by dams changes the geomorphic behavior of rivers. Downstream of the dams, river channels become deeper and some move laterally and change direction (lateral avulsion); mainly due to the sedimentological character of these fine sediment channels. This effect and the floods reduction act synergistically on the hydrological connectivity of the river with its floodplain lakes. This effect has already been observed in the Sogamoso River floodplain; the main river channel has shifted to the north because accelerated lateral avulsion and the floods have been regulated by hydropower. Nowadays, its floodplain lake (El Llanito lake), the former most biodiverse lake in terms of fish fauna (Granado-Lorencio et al., 2012), is being gradually isolated from the Sogamoso River (personal observation based in historic LANDSAT images) with unpredictable consequences on its biological productivity. By 2023, the Hidroituango reservoir will retain sediment from the Cauca River (Figure 1). The Cauca River floodplains and the Momposina floodplain wetlands downstream will experience major physical and biological changes (Angarita et al., 2018). This scenario may be a new knockout for floodplain sustainability and artisanal fisheries downstream of this dam if proper management strategies are not implemented to avoid this expected and negative change.
Recommendations for fish and fisheries conservation in the Magdalena River
If we could recommend what landscape or ecological characteristic must be preserved over all of those we have described in order to preserve fish and artisanal fisheries, certainly we will be in a trouble. In nature, biota has synchronized their dynamics with environmental characteristics and some of them are definitive cues to activate the vital process for reproduction and embryo survival. In the Magdalena River basin, there is a “Chrono-sequence” between changes in the Sun bright, rains, discharge variability, and floods (Figure 4); the order for when they happen are definitive for aquatic biota dynamics (Figure 8) because they are fundamental for maintaining the flooded area and the connection in the fluvial net throughout the dispersion of nutrients and carbon. Marín-Avendaño (2012) suggests that the high resilience of the fish assemblage in the floodplain lakes may be associated with the high number of functional groups and also mentions that flood pulse and connectivity are two characteristics in the river basin that are fundamental for this resilience.
Researchers in the fisheries science need to improve their language and increase the ways of sharing the knowledge more than that in scientific article in order to influence common people, stakeholders, and political decisions. Fishery landings can be seen as the peak of an iceberg that emerges over the water line (López-Casas et al., 2020). The height of the peak represents the landings magnitude, and the submerged height of that iceberg, the intensity of the interactions between physical, chemical, biological, and human variables (and factors); changes in that submerged iceberg size will affect the height of the iceberg over the water line. Variables and their interactions in that submersed iceberg are most of the times unknown and it is common that management decisions in fisheries do not include the environmental context that provide that production, and most of the regulations have been focused on the fisher (Hernandez-Barrero et al., 2021).
Identifying natural drivers for fisheries sustainability are useful to understand how the different compartments in the riverine system interact and let us to take management decisions on the anthropogenic and indirect drivers in order to improve river restoration and conservation. This manuscript constructed a story using knowledge coming from questions made from different Colombian limnologist and fishery ecologist in the last 70 years and new data in the last decade. We focused on explaining how those natural and anthropological drivers influence the fisheries production in the Magdalena River, a tropical Andean river. However, we need to advance looking for answers to many new questions about the functional role of the fish in the aquatic food web, and the connectivity and hydrological functioning of these floodplains (the “submerged iceberg”) in Andean rivers; the more we know, the more we realize we really do not know anything, and it is necessary to continue exploring these particular systems in the world. We must remain working on: 1) how the floodplain morphology changed during the last decades in terms of connectivity with the main river and its flooded areas? 2) What are the hydrological dynamics of floodplains in terms of recharge water level thresholds in a heterogeneous landscape with three Andean Cordilleras and how these dynamics changed over time? 3) What are the spatial and temporal dynamics of suspended sediment concentration and how does this process impact the carbon flux through the food webs in the floodplain lakes? 4) How the hydropower development of the basin changes the river-floodplain connection? 5) Which response variables (physical and/or biological) are important for future monitoring programs? 6) What will be the future of artisanal fishermen, their traditions, and food support under the observed increasing trends of land cover change in the Magdalena floodplains, environmental scientists. Decision makers face new scientific challenges, including: a) obtain the capacity building to monitor and manage floodplain wetlands, b) recognize their ecosystem services and value, and c) establish the scientific needs toward its sustainable hydrologic and biologic functioning. Certainly, the well-being of the floodplain ecosystems, the seasonal flooding and the fluvial network connectivity are the main tools to preserve and manage the ecosystem services of the Magdalena River and its floodplains lakes (Usma-Oviedo et al., 2013; Autoridad Nacional de Acuicultura y Pesca AUNAP, 2015; Jiménez-Segura et al., 2016a; Reis et al., 2016; Herrera-Pérez et al., 2019; Cacua-Ortiz et al., 2020; Williams-Subiza and Epele, 2021).
Author contributions
LJ-S, AH-S, and JR-Á conceptualized and design the manuscript, wrote the draft and revised it for important intellectual content, approved the final version and were in agree for all the aspects mentioned by reviewers in order to improve the accuracy and integrity of manuscript.
Acknowledgments
We are grateful to all our students for their support and to the institutions that have funded scientific research for the knowledge construction we present. Gratitude to the reviewers for suggesting relevant changes; certainly, they improved the quality of this manuscript. Analyses of satellite-derived data of water discharge and floodplain connectivity were carried out with support from the US National Academy of Sciences under the funded PEER project (grant number 70) “Satellite-based estimations of River discharge in the Magdalena River”: Capacity building to mitigate sources of upstream runoff and associated risks of pollution.”
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors, and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Aalto, R., Maurice-Bourgoin, L., Dunne, T., Montgomery, D. R., Nittrouer, C. A., and Guyot, J. L. (2002). Episodic sediment accumulation on amazonian floodplains influenced by el niño/southern oscillation. Nature 425, 493–497. doi:10.1038/nature02002
Alvarez, A., Kolok, A., Jiménez-Segura, L. F., Granados, C., and Palacio, J. (2012). Mercury concentrations in muscle and liver tissue of fish from marshes along the Magdalena River, Colombia. Bull. Environ. Contam. Toxicol. 89, 836–840. doi:10.1007/s00128-012-0782-9
Angarita, H., Wickel, A. J., Sieber, J., Chavarro, J., Maldonado-Ocampo, J. A., Herrera-R., G. A., et al. (2018). Basin-scale impacts of hydropower development on the Mompós Depression wetlands, Colombia. Hydrol. Earth Syst. Sci. 22, 2839–2865. doi:10.5194/hess-22-2839-2018
Angarita, H., Santos-Fleischmann, A., Rogéliz, C., Campo, F., Narváez-Campo, G., Delgado, J., et al. (2020). “Modificación del hábitat para los peces para la cuenca del río Magdalena, Colombia,” in Peces de la cuenca del río Magdalena, Colombia: diversidad, conservación y uso sostenible. Editors L. Jiménez-Segura, and C. Lasso. Serie Editorial Recursos Hidrobiológicos y Pesqueros Continentales de Colombia. (Instituto de Investigación de Recursos Biológicos Alexander von Humboldt). doi:10.21068/B2020RRHHXIX07
AUNAP-UNIMAGDALENA (2014). “Caracterización de los principales artes de pesca de Colombia y reporte del consolidado del tipo y número de artes, embarcaciones y UEP’s empleadas por los pescadores vinculados a la actividad pesquera,” in Contrato de Prestación de Servicios No. 190, suscrito entre la Autoridad Nacional de Acuicultura y Pesca y la Universidad del Magdalena (Santa Marta y Bogotá, Colombia. Avaliable at: http://sepec.aunap.gov.co/Archivos/Informe%20Caracterizaci%C3%B3n%20de%20artes.pdf (last view October 29, 2018.
Autoridad Nacional de Acuicultura y Pesca AUNAP (2015). Resolución No. 2287 del 29 de diciembre de 2015, por la cual se declaran unas especies de peces como domesticadas para el desarrollo de la Acuicultura y se dictan otras disposiciones. Avaliable at: https://sioc.minagricultura.gov.co/Acuicultura/Normatividad/Resoluci%C3%B3n%202287%20de%202015%20AUNAP.pdf (last view April 18 2022).
Barletta, M., Jaureguizar, A. J., Baigun, C., Fontoura, N. F., Agostinho, A. A., Almeida-Val, V. M. F., et al. (2010). Fish and aquatic habitat conservation in South America: A continental overview with emphasis on neotropical systems. J. Fish. Biol. 76, 2118–2176. doi:10.1111/j.1095-8649.2010.02684.x
Baumgartner, G., Nakatani, K., Gomes, L., Bialetzki, A., and Sanches, P. (2004). Identification of spawning sites and natural nurseries of fishes in the upper Paraná River, Brazil. Environ. Biol. Fishes 71 (2), 115–125. doi:10.1007/s10641-004-0098-z
Beach, T., and Luzzadder-Beach, S. (2012). “Pre-columbian people and the wetlands in central and south America,” in The oxford handbook of wetland archaeology. Editors F. Menotti, and A. O’Sullivan (London. UK: Oxford University Press). doi:10.1093/oxfordhb/9780199573493.013.0007
Bray, W. (2006). “Searching for environmental stress: Climatic and anthropogenic influences on the landscape of Colombia,” in Archaeology in the lowland American tropics: Current analytical methods and applications. Editor P. W. Stahl (Cambridge: Cambridge University Press).
Bulla, C. K., Gomes, L. C., Miranda, L. E., and Agostinho, A. A. (2011). The ichthyofauna of drifting macrophyte mats in the Ivinhema River, upper Paraná River basin, Brazil. Neotrop. Ichthyol. 9 (2), 403–409. doi:10.1590/S1679-62252011005000021
Cacua-Ortiz, S. M., Aguirre, N. J., and Peñuela, G. A. (2020). Methyl paraben and carbamazepine in water and striped catfish (Pseudoplatystoma magdaleniatum) in the Cauca and magdalena rivers. Bull. Environ. Contam. Toxicol. 105 (6), 819–826. doi:10.1007/s00128-020-03028-z
Campbell, J. C. (1974). “Colombian Andes,” in Mesozoic-cenozoic orogenic belts. Editor A. M. Spencer (London, UK: Geological Society London, Special Publication).
Carpenter, S., Bennett, E., and Peterson, G. (2006). Scenarios for ecosystem services: An overview. Ecol. Soc. 11, art29. doi:10.5751/es-01610-110129
Carvajal-Quintero, J. D., Escobar, F., Alvarado, F., Villa-Navarro, F. A., Jaramillo-Villa, Ú., and Maldonado-Ocampo, J. A. (2015). Variation in freshwater fish assemblages along a regional elevation gradient in the northern Andes, Colombia. Ecol. Evol. 5 (13), 2608–2620. doi:10.1002/ece3.1539
Castellanos-Mejía, M., Herrera, J., Noguera-Urbano, E., Parra, E., and Jiménez-Segura, L. (2021). Potential distribution in Colombia of the introduced fish Pangasianodon hypophthalmus (Siluriformes: Pangasiidae) and implications for endangered native fish. Rev. Biol. Trop. 69 (2), 573–587. doi:10.15517/rbt.v69i2.44223
De Fex-Wolf, D., López-Casas, S., and Jiménez-Segura, L. (2019). Hydropower effects on Prochilodus magdalenae (prochilodontidae) reproduction: Evidence from endocrine response. Rev. MVZ Cordoba 24 (2), 7180–7187. doi:10.21897/rmvz.1606
Dunne, T., Mertes, L. A. K., Meade, R. H., Richey, J. E., and Forsberg, B. R. (1998). Exchanges of sediment be- tween the flood plain and channel of the Amazon River in Brazil. Geol. Soc. Am. Bull. 110, 450–466. doi:10.1130/0016-7606(1998)110<0450:EOSBTF>2.3.CO;2
Escobar, R., Restrepo, J. D., Brakenridge, G. R., Kettner, A. J., and Kettner, A. (2016). “Satellite-based estimation of water discharge and runoff in the Magdalena river, northern Andes of Colombia,” in Remote sensing of hydrological extremes. Editor V. Lakshmi (Switzerland: Springer International Publishing). doi:10.1007/978-3-319-43744-6_1
Flecker, A. S. (1996). Ecosystem engineering by a dominant detritivore in a diverse tropical stream. Ecology 77 (6), 1845–1854. doi:10.2307/2265788
Gallego, S., Peñuela, G., and Martínez-López, E. (2021). Enzymatic activity changes in striped catfish Pseudoplatystoma magdaleniatum, induced by exposure to different concentrations of ibuprofen and triclosan. Chemosphere 271, 129399. doi:10.1016/j.chemosphere.2020.129399
Gallo-Sánchez, L. N., Aguirre-Ramírez, N., Palacio-Baena, J., and Ramírez-Restrepo, J. J. (2009). Zooplancton (Rotífera y Microcrustácea) y su relación con los cambios del nivel del agua en la ciénaga de Ayapel, Córdoba, Colombia. Caldasia 31, 327–341. doi:10.15446/caldasia
Galvis, G., and Mojica, J. (2007). The Magdalena River fresh water fishes and fisheries. Aquat. Ecosyst. Health Manage. 10, 127–139. doi:10.1080/14634980701357640
García-Alzate, C., DoNascimiento, C., Villa-Navarro, F. A., García-Melo, J. E., and Herrera, R.-G. (2020). “Diversidad de peces de la cuenca del río Magdalena Colombia,” in Peces de la cuenca del río Magdalena, Colombia: diversidad, conservación y uso sostenible. Editors L. Jiménez-Segura, and C. Lasso. Serie Editorial Recursos Hidrobiológicos y Pesqueros Continentales de Colombia. Instituto de Investigación de Recursos Biológicos Alexander von Humboldt. doi:10.21068/B2020RRHHXIX02
Glantz, M. H. (1997). Currents of change, el niño’s impact on climate and society. Cambridge: Cambridge University Press.
Goodbred, S. L., and Kuehl, S. A. (1998). Floodplain processes in the bengal basin and the storage of ganges- brahmaputra river sediment: An accretion study using 137Cs and 210Pb geochronology. Sediment. Geol. 121, 239–258. doi:10.1016/S0037-0738(98)00082-7
Granado-Lorencio, C., Gulfo, A., Alvarez, F., Jiménez-Segura, L. F., Carvajal-Quintero, J., and Hernández-Serna, A. (2012). Fish assemblages in floodplain lakes in a Neotropical river during the wet season (Magdalena River, Colombia). J. Trop. Ecol. 28 (3), 271–279. S0266467412000181. doi:10.1017/s0266467412000181
Gutiérrez, F., Barreto, C., and Mancilla, B. (2011). “Diagnóstico de la pesquería en la cuenca del río Magdalena-Cauca,” in Catalogo de los recursos pesqueros continentales de Colombia. Serie Editorial Recursos Hidrobiológicos y Pesqueros Continentales de Colombia. Editors C. A. Lasso, F. P. Gutiérrez, M. A. Morales-Betancourt, E. Agudelo, H. Ramírez-Gil, and R. E. Ajiaco-Martínez (Bogotá, Colombia: Instituto de Investigación de Recursos Biológicos Alexander von Humboldt).
Gutiérrez-Moreno, L. C., and De la Parra-Guerra, A. C. (2020). “Contaminación en la cuenca del río Magdalena (Colombia) y su relación con los peces”, in Peces de la cuenca del río Magdalena, Colombia: diversidad, conservación y uso sostenible. Editors L. Jiménez-Segura, and C. Lasso. Serie Editorial Recursos Hidrobiológicos y Pesqueros Continentales de Colombia. (Instituto de Investigación de Recursos Biológicos Alexander von Humboldt). doi:10.21068/B2020RRHHXIX06
Hahn, L. (2012). “The application of radio telemetry to fisheries research in Brazil´s large rivers,” in Telemetry techniques A user guide for fisheries research. Editors S. Noah, N. Adams, J. Beeman, and H. John Eiler (American Fisheries Society). ISBN: 978-1-934874-26-4.
Hernández-Atilano, E., Aguirre, N. J., Palacio, J. A., and Ramírez-Restrepo, J. J. (2008). Variación espacio-temporal de la asociación fitoplanctónica en diferentes momentos del pulso hidrológico en la ciénaga de Ayapel (córdoba), Colombia. Rev. Act. Biol. 30 (88), 67–81. Available at: http://www.scielo.org.co/scielo.php?script=sci_arttext&pid=S0304-35842008000100006&lng=en
Hernández-Barrero, S., Valderrama-Barco, M., Barreto-Reyes, C., and Stotz, W. (2021). Effects of selective fishing on a small-scale multi-species and multi-gear freshwater fishery in the Magdalena River Basin (Colombia). Can. J. Fish. Aquat. Sci. 78 (12), 1841–1854. doi:10.1139/cjfas-2020-0216
Herrera-Pérez, J., Parra, J. L., Restrepo-Santamaría, D., and Jiménez-Segura, L. F. (2019). The influence of abiotic environment and connectivity on the distribution of diversity in an Andean fish fluvial network. Front. Environ. Sci. 7. doi:10.3389/fenvs.2019.00009
HIMAT (1977). HIMAT, bogotá, Colombia. Available from: http://agris.fao.org/agris-search/search.do?recordID=CO20000013309 (last view October 28, 2018.Proyecto colombo-Holandes de la Cuenca magdalena-cauca. Consulting report.
INGETEC S.A Ingenieros Consultores. (1998). Actualización del Estudio de Impacto Ambiental. Manejo del medio biótico. 5 (6), 7–69.
Instituto Nacional de Pesca y Acuicultura INPA (1998). Boletín estadístico pesquero. República de Colombia: Ministerio de Agricultura. Available from: http://sepec.aunap.gov.co/Home/BoletinesEstadisticos. (Accessed April 5, 2022).
Instituto Nacional de Pesca y Acuicultura INPA (1999). Boletín estadístico pesquero. República de Colombia: Ministerio de Agricultura. Available from: http://sepec.aunap.gov.co/Home/BoletinesEstadisticos. (Accessed April 5, 2022).
Instituto Alexander von Humboldt (2019). Respuestas socioecológicas en el marco de la navegabilidad en el río magdalena: Criterios técnico—científicos de definición de áreas priorizadas de alta vulnerabilidad al conflicto socioecológico en el marco del proyecto de recuperación de la navegabilidad. Informe técnico. Available from: http://repository.humboldt.org.co/handle/20.500.11761/35497. (Accessed September 5, 2022).
Irving, E. M. (1975). Structural evolution of the northernmost Andes, Colombia. Washington: U.S. Geol. Surv., Prof. Pap., 846. doi:10.1017/S0016756800050780
IUCN (2022). Barometer of life. Available at: https://www.iucnredlist.org/about/barometer-of-life. (Accessed September 5, 2022).
Jaramillo, U., Pelaez, S., Aponte, C., Flórez-Ayala, C., Avella, C., Manrique, O., et al. (2015). “Hacía un inventario completo de humedales,” in Colombia anfibia. Un país de humedales Vol I. Editors U. Jaramillo, J. Cortes-Duque, and C. Flórez (Bogotá, Colombia: Instituto de investigación de recursos hidrobiológicos Alexander von Humboldt). ISBN 978-958-8889-48-1.
Jaramillo, U., Cortes-Duque, J., and Flórez, C. (2015a). Colombia anfibia. Un país de humedales, Vols. I and Vol. II. Bogotá, Colombia: Instituto de investigación de recursos hidrobiológicos Alexander von Humboldt. Available at: http://www.humboldt.org.co/es/estado-de-los-recursos-naturales/item/802-colombiaanfibiav1 (last view November 28, 2021).
Jaramillo-Villa, U., Maldonado-Ocampo, J. A., and Escobar, F. (2010). Altitudinal variation in fish assemblage diversity in streams of the central Andes of Colombia. J. Fish. Biol. 76 (10), 2401–2417. doi:10.1111/j.1095-8649.2010.02629.x
Jiménez-Segura, L. F. (2007). Ictioplancton y reproducción de los peces en la cuenca media del río Magdalena (sector de Puerto Berrio, Antioquia). Colombia: Unpublished Doctoral Thesis. Universidad de Antioquia.
Jiménez-Segura, L. F., Carvajal-Quintero, J. D., and Aguirre, N. (2010a). Las ciénagas como hábitat para los peces: Estudio de caso en la ciénaga de Ayapel (córdoba), Colombia. Rev. Act. Biol. 32, 53–64. Available at: http://www.scielo.org.co/pdf/acbi/v32n92/v32n92a5.pdf.
Jiménez-Segura, L. F., Palacio, J., and Leite, R. (2010b). River flooding and reproduction of migratory fish species in the Magdalena River Basin, Colombia. Ecol. Freshw. Fish. 19, 178–186. doi:10.1111/j.1600-0633.2009.00402.x
Jiménez-Segura, L. F., Restrepo-Santamaría, D., López-Casas, S., Delgado, J., Valderrama, M., Álvarez, J., et al. (2014). Ictiofauna y desarrollo del sector hidroeléctrico en la cuenca del río Magdalena – Cauca, Colombia. Biota Colomb. 15, 3–25. doi:10.21068/c0001
Jiménez-Segura, L. F., Galvis-Vergara, G., Cala-Cala, P., García-Alzate, C. A., López-Casas, S., Rios-Pulgarin, M. I., et al. (2016). Freshwater fish faunas, habitats and conservation challenges in the Caribbean river basins of north-western South America. J. Fish. Biol. 89, 65–101. doi:10.1111/jfb.13018
Jiménez-Segura, L. F., Ortega, H., Chuctaya, J., Jiménez-Prado, P., Carvajal-Vallejos, F., Rivadeneira, J. F., et al. (2016a). “Estado de conservación y distribución de los peces de agua dulce de los Andes Tropicales,” in Estado de Conservación y Distribución de la Biodiversidad de Agua Dulce en los Andes Tropicales. Editors M. F. Tognelli, C. A. Lasso, C. A. Bota-Sierra, L. F. Jiménez-Segura, and N. A. Cox (Gland, Suiza, Cambridge, UK and Arlington, USA: UICN). doi:10.2305/IUCN.CH.2016.02.es
Jiménez-Segura, L., Herrera-Pérez, J., Valencia-Rodríguez, D., Castaño-Tenorio, I., López-Casas, S., Ríos, M. I., et al. (2020). “Ecología e historias de vida de los peces en la cuenca del río Magdalena, Colombia,” in Peces de la cuenca del río Magdalena, Colombia: diversidad, conservación y uso sostenible. Editors L. Jiménez-Segura, and C. Lasso. Serie Editorial Recursos Hidrobiológicos y Pesqueros Continentales de Colombia. (Instituto de Investigación de Recursos Biológicos Alexander von Humboldt). doi:10.21068/B2020RRHHXIX04
Kettner, A., Restrepo, J. D., and Syvitski, J. P. M. (2010). A spatial simulation experiment to replicate fluvial sediment fluxes within the Magdalena river basin, Colombia. J. Geol. 118, 363–379. doi:10.1086/652659
Lasso, C. A., Escobar, M. D., Herrera, J., Castellanos, M. C., Valencia-Rodríguez, D., Campuzano, J., et al. (2020). “Peces introducidos en el río magdalena y cuencas vecinas, Colombia,” in Peces de la cuenca del río Magdalena, Colombia: Diversidad, conservación y uso sostenible. Serie Editorial Recursos Hidrobiológicos y Pesqueros Continentales de Colombia. Editors L. Jiménez-Segura, and C. A. Lasso (Colombia: Instituto de Investigación de Recursos Biológicos Alexander von Humboldt. Bogotá, D. C.). doi:10.21068/B2020RRHHXIX08
Lasso, C. A., Gutiérrez, F. P., Morales-Betancourt, M. A., Agudelo, E., Ramírez-Gil, H., and Ajiaco-Martinez, R. E. (2011). Catálogo de los recursos pesqueros continentales de Colombia. Bogotá, Colombia: Instituto de Investigación de Recursos Biológicos Alexander von Humboldt.
Latrubesse, E. M. (2015). Large rivers, megafans and other quaternary avulsive fluvial systems: A potential who’s who in the geological record. Earth. Sci. Rev. 146, 1–30. doi:10.1016/j.earscirev.2015.03.004
Latrubesse, E. M., Stevauxs, J. C., and Sinha, R. (2005). Tropical rivers. Geomorphology 70, 187–206. doi:10.1016/j.geomorph.2005.02.005
Latrubesse, E., and Restrepo, J. D. (2014). Sediment yield along the Andes: Continental budget, regional variations, and comparisons with other basins from orogenic mountain belts. Geomorphology 216, 225–233. doi:10.1016/j.geomorph.2014.04.007
López-Casas, S., Jiménez-Segura, L. F., Agostinho, A. A., and Pérez, C. M. (2016). Potamodromous migrations in the Magdalena river basin: Bimodal reproductive patterns in neotropical rivers. J. Fish. Biol. 89, 157–171. doi:10.1111/jfb.12941
López-Casas, S. (2015). Magdalena potadromous migrations: Effects of regulated and natural hydrological regimes. Medellín, Colombia: Unpublished Doctoral thesis. Instituto de Biología, Universidad de Antioquia.
López-Casas, S., Rondón-Martínez, Y. F., Gutiérrez-Cortés, A., Escobar-Cardona, J. L, Muñoz-Duque, S., Valencia-Rodríguez, D., et al. (2020). “Diagnóstico del grado de amenaza y medidas de manejo para los peces del río Magdalena, Colombia,” in Peces de la cuenca del río Magdalena, Colombia: diversidad, conservación y uso sostenible. Editors L. Jiménez-Segura, and C. Lasso. Serie Editorial Recursos Hidrobiológicos y Pesqueros Continentales de Colombia. (Instituto de Investigación de Recursos Biológicos Alexander von Humboldt). doi:10.21068/B2020RRHHXIX10
Lundberg, J. G., Marshall, L. G., Guerrero, J., Horton, B., Malabarbarba, M. C., and Wesselingh, F. (1998). “The stage for neotropical fish diversification: A history of tropical South American rivers,” in Phylogeny and classification of neotropical fishes. Editors L. R. Malabarba, R. E. Reis, R. P. Vari, C. A. S. Lucena, Z. M. S. Lucena, P. U. C. R. S. Museu de Ciências e Tecnologiaet al.
Mancera-Rodríguez, N. J., and Álvarez-León, R. (2006). Estado del conocimiento de las concentraciones de metales pesados en los peces dulceacuícolas de Colombia. Acta Biolo. Colomb. 11, 3–23.
Marín-Avendaño, C. M. (2012). Interacciones tróficas y productividad íctica en el sistema cenagoso de Ayapel, Córdoba Colombia. Doctor thesis. Medellín, Colombia: Faculty of Engineering. University of Antioquia. Available at: https://bibliotecadigital.udea.edu.co/bitstream/10495/7404/1/MarinCarlos_2017_InteraccionesTroficasProductividad.pdf (Accessed September 5, 2022).
Moreno, C., López-Casas, S., Rogeliz-Prada, C., and Jiménez-Segura, L. (2021). Protection of spawning habitat for potamodromous fish, an urgent need for the hydropower planning in the Andes. Ecol. Soc. 19, 1–23. doi:10.5751/es-01826-110229
Nelson, G. C., Bennett, E., Berhe, A. A., Cassman, K., DeFries, R., Dietz, T., et al. (2006). Anthropogenic drivers of ecosystem change: An overview. Ecol. Soc. 11 (2), 29. [online] URL: http://www.ecologyandsociety.org/vol11/iss2/art29/.
Ochoa-Orrego, L., Jiménez-Segura, L. F., and Palacio, J. (2015). Ictioplancton en la ciénaga de ayapel, río san jorge (Colombia): Cambios espacio-tempo. Mus. Cient. Hist. Nat. 19 (1), 103–114. doi:10.17151/bccm.2015.19.1.7
Patino, J. E., and Estupiñan-Suarez, L. M. (2016). Hotspots of wetland area loss in Colombia. Wetlands 36 (5), 935–943. doi:10.1007/s13157-016-0806-z
Pickens, A. H., Hansen, M. C., Hancher, M., Stehman, S. V., Tyukavina, A., Potapov, P., et al. (2020). Mapping and sampling to characterize global inland water dynamics from 1999 to 2018 with full Landsat time-series. Remote Sens. Environ. 243, 111792. doi:10.1016/j.rse.2020.111792
Plazas, C., Falchetti, A. M., Van der Hammen, T., and Botero, P. J. (1988). Cambios ambientales y desarrollo cultural en el Bajo Río San Jorge. Bol. Mus. Oro Banco Rep. 20, 58–59. Available at: https://publicaciones.banrepcultural.org/index.php/bmo/article/view/7167
Potapov, P., Hansen, M. C., Kommareddy, I., Kommareddy, A., Turubanova, S., Pickens, A., et al. (2020). Landsat analysis ready data for global land cover and land cover change mapping. Remote Sens. (Basel). 12, 426. doi:10.3390/rs12030426
Poveda, G. (1997). Atractores extraños (CAOS) en la hidroclimatología de Colombia? Rev. Acad. Colomb. Cienc. 21, 431–444. ISSN 0370-3908
Poveda, G., Jaramillo, A., Gil, M. M., Quiceno, N., and Mantilla, R. (2001). Seasonally in ENSO-related precipitation, river discharges, soil moisture, and vegetation index in Colombia. Water Resour. Res. 37 (8), 2169–2178. doi:10.1029/2000WR900395
Poveda, G., and Mesa, O. J. (1997). Feedbacks between hydrological processes in tropical South America and large-scale ocean- atmospheric phenomena. J. Clim. 10, 2690–2702. doi:10.1175/1520-0442(1997)010<2690:fbhpit>2.0.co;2
Ramírez, A., and Viña, G. (1998). Limnología colombiana: Aportes a su conocimiento y estadísticas de análisis. Bogotá, Colombia: Editorial Panamericana. ISBN 958-902906-X.
Reid, A. J., Carlson, A. K., Creed, I. F., Eliason, E. J., Gell, P. A., Johnson, P. T. J., et al. (2019). Emerging threats and persistent conservation challenges for freshwater biodiversity. Biol. Rev. 94, 849–873. doi:10.1111/brv.12480
Reis, R. E., Albert, J. S., Di Dario, F., Mincarone, M. M., Petry, P., and Rocha, L. A. (2016). Fish biodiversity and conservation in South America. J. Fish. Biol. 89, 12–47. doi:10.1111/jfb.13016
Reis, A. S., Albrecht, M. P., and Bunn, S. E. (2020). Food web pathways for fish communities in small tropical streams. Freshw. Biol. 65, 893–907. doi:10.1111/fwb.13471
Restrepo, J. D., Kjerfve, B., Restrepo, J. C., and Hermelin, M. (2006a). Factors controlling sediment yield in a major South American drainage basin: The Magdalena river, Colombia. J. Hydrol. X. 316, 213–232. doi:10.1016/j.jhydrol.2005.05.002
Restrepo, C. A., Toro, M., and Aguirre, N. (2006b). Aproximación a la dinámica del fósforo y del nitrógeno en la ciénaga de Ayapel. Av. Rec. Hidrául. 13, 7–22. Available at: https://revistas.unal.edu.co/index.php/arh/article/view/9336
Restrepo, J. D., Kettner, A. J., and Brackenridge, R. (2020a). Monitoring water discharge and floodplain connectivity for the northern Andes utilizing satellite data: A tool for river planning and science-based decisión-making. J. Hydrology 586, 125123. doi:10.1016/j.jhydrol.2020.125123
Restrepo, J. D., Cárdenas-Rozo, A., Paniagua-Arroyave, J. F., and Jiménez-Segura, L. (2020b). “Aspectos físicos de la cuenca del río magdalena, Colombia: Geología, hidrología, sedimentos, conectividad, ecosistemas acuáticos e implicaciones para la biota,” in Peces de la cuenca del río Magdalena, Colombia: Diversidad, conservación y uso sostenible. Serie Editorial Recursos Hidrobiológicos y Pesqueros Continentales de Colombia. Editors L. Jiménez-Segura, and C. A. Lasso (Bogotá, D. C., Colombia: Instituto de Investigación de Recursos Biológicos Alexander von Humboldt). doi:10.21068/B2020RRHHXIX01
Restrepo, J. D., and Syvitski, J. P. M. (2006). Assessing the effect of natural controls and land use chane on sediment yield in a major Andean river: The magdalena drainage basin, Colombia. Ambio 35, 44–53. doi:10.1579/0044-7447(2006)35[65:ATEONC]2.0.CO;2
Restrepo, J., Kettner, A., and Syvitski, J. (2015). Recent deforestation causes rapid increase in river sediment load in the Colombian Andes. Anthropocene 10, 13–28. doi:10.1016/j.ancene.2015.09.001
Ríos-Pulgarín, M. I., Jiménez-Segura, L. F., Palacio, J. A., and Ramírez-Restrepo, J. J. (2008). Comunidad de peces en la Ciénaga de Ayapel, río magdalena (córdoba) Colombia: Cambios espacio-temporales en su asociación. Rev. Act. Biol. 30, 29–53. Available at: http://www.scielo.org.co/scielo.php?script=sci_abstract&pid=S0304-35842008000100004
Schiesari, L., Zuanon, J., Azevedo-Ramos, C., Garcia, M., Gordo, M., Messias, M., et al. (2003). Macrophyte rafts as dispersal vectors for fishes and amphibians in the lower solimões river, central Amazon. J. Trop. Ecol. 19, 333–336. doi:10.1017/S0266467403003365
Senado de la República de Colombia (2020). Proyecto de ley número 298 de 2019 por medio del cual se declara al río Grande de la Magdalena como hidrovía y se dictan otras disposiciones. Avaliable at: http://www.secretariasenado.gov.co/legibus/legibus/gacetas/2020/GC_0204_2020.pdf (last view April 18 2022).
Smith, D. G. (1986). Anastomosing river deposits, sedimentation rates and basin subsidence, Magdalena River, northwestern Colombia, South America. Sediment. Geol. 46, 177–196. doi:10.1016/0037-0738(86)90058-8
Strayer, D. L., and Dudgeon, D. (2010). Freshwater biodiversity conservation: Recent progress and future challenges. J. North Am. Benthol. Soc. 29, 344–358. doi:10.1899/08-171.1
Tanner, W. F. (1974). The incomplete flood plain. Geol. 2, 105–106. doi:10.1130/0091-7613(1974)2<105:tifp>2.0.co;2
The Nature Conservancy, Fundación Alma, Fundación Humedales and Autoridad Nacional de Pesca (2016). Estado de las planicies inundables y el recurso pesquero en la macrocuenca Magdalena-Cauca y propuesta para su manejo integrado. (The Nature Conservancy. Bogotá, Colombia. Bogotá, Colombia). Available at: https://www.mundotnc.org/nuestro-trabajo/donde-trabajamos/america/colombia/estado-y-propuesta-para-el-manejo-de-las-planicies-inundables-y-el-recurso-p.xml (last view November 29, 2021).
Tung, N. T., Du, N. N., An, V. V., and Tuan, T. T. (2006). “Monitoring of fish larvae during the annual flood of the Mekong and Bassac rivers, Mekong Delta, Viet Nam,” in MRC conference series (No. 6) (Vietnam: Mekong River Commission, Vientiane, Lao PDR).
Usma-Oviedo, S., Villa-Navarro, F., Lasso, C. A., Castro, F., Zuñiga, P. T., Cipamocha, C. A., et al. (2013). “Peces dulceacuícolas migratorios,” in Guía de las especies migratorias de la biodiversidad en Colombia. Editors I. A. Zapata, and J. S. Usma (WWF-Colombia: Ministerio de Ambiente y Desarrollo Sostenible), Vol. 2. ISBN 978-958-8353-61-6
Valderrama, M., Hernández, S., Pinilla, M., and Barreto, C. (2016). “Estado de las Pesquerías,” in Estado de las planicies inundables y el recurso pesquero en la macrocuenca Magdalena-Cauca y propuesta para su manejo integrado. The nature conservancy, Fundación Alma, Fundación Humedales and Autoridad nacional de Pesca (Bogotá, Colombia: The Nature Conservancy).
Valderrama, M., Escobar, C. J. L., Pardo, B. R., Toro, S. M., Gutiérrez, C. J. C., and López, C. S. (2020). “Servicios ecosistémicos generados por los peces de la cuenca del río Magdalena, Colombia,” in Peces de la cuenca del río Magdalena, Colombia: diversidad, conservación y uso sostenible. Editors L. Jiménez-Segura, and C. Lasso. Serie Editorial Recursos Hidrobiológicos y Pesqueros Continentales de Colombia. (Instituto de Investigación de Recursos Biológicos Alexander von Humboldt). doi:10.21068/B2020RRHHXIX05
Valderrama-Barco, M., Jiménez-Segura, L. F., López-Casas, S., Rivas, T., Rincón, , Nieto-Torres, S., et al. (2011). “Prochilodus magdalenae Steindachner 1879 (Characiformes, Prochilodontidae)” in Catálogo de los recursos pesqueros continentales de Colombia,” in Serie Editorial Recursos Hidrobiológicos y Pesqueros Continentales de Colombia. Editors C. Lasso, E. Agudelo-Córdoba, L. F. Jiménez-Segura, H. Ramírez-Gil, M. Morales- Betancourt, R. E. Ajiaco-Martínezet al. (Bogotá, Colombia: Instituto de Investigación de los Recursos Biológicos Alexander von Humboldt).
Valencia-Rodríguez, D., Herrera-Pérez, J., Restrepo-Santamaria, D., Galeano, A., Winton, R., and Jiménez-Segura, L. (2022). Fish community turnover in a dammed Andean River over time. Neotrop. Ichthyol. 20, 91. doi:10.1590/1982-0224-2021-0091
Vargas-Licona, S. P., and Marrugo-Negrete, J. L. (2019). Mercurio, metilmercurio y otros metales pesados en peces de Colombia: Riesgo por ingesta. Acta Biol. Colomb. 24 (2), 232–242. doi:10.15446/abc.v24n2.74128
Vörösmarty, C., Meybeckc, M., Feketea, B., Sharmad, K., Greena, P., and Syvitsk, J. (2003). Anthropogenic sediment retention: Major global impact from registered river impoundments. Glob. Planet. Change 39, 169–190. doi:10.1016/S0921-8181(03)00023-7
Welcomme, R. L., Winemiller, K. O., and Cowx, I. G. (2006). Fish environmental guilds as a tool for assessment of ecological condition of rivers. River Res. Appl. 22, 377–396. doi:10.1002/rra.914
Williams-Subiza, E. A., and Epele, L. B. (2021). Drivers of biodiversity loss in freshwater environments: A bibliometric analysis of the recent literature. Aquat. Conserv. 31 (9), 2469–2480. doi:10.1002/aqc.3627
Winemiller, K., and Rose, K. (1992). Patterns of life-history diversification in north American fishes: Implications for population regulation. Can. J. Fish. Aquat. Sci. 49, 2196–2218. doi:10.1139/f92-242
Winemiller, K., Welcomme, R., and Petr, T. (2004). “Floodplain river food webs: Generalizations and implications for fisheries management,” in Proceedings of the second international symposium on the management of larger rivers for fisheries, volume I. FAO regional office for Asia and the pacific. Editors R. Welcomme, and T. Petr (Bangkok, Thailand: RAP Publication), 16, 285–309.
Keywords: Andean river, Magdalena, connectivity, floodplains, sediment load, potamodromous, artisanal fisheries, habitat loss
Citation: Jiménez-Segura L, Restrepo-Ángel JD and Hernandez-Serna A (2022) Drivers for the artisanal fisheries production in the Magdalena River. Front. Environ. Sci. 10:866575. doi: 10.3389/fenvs.2022.866575
Received: 31 January 2022; Accepted: 16 August 2022;
Published: 06 October 2022.
Edited by:
Claudio Baigun, Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET), ArgentinaReviewed by:
Raphael Ligeiro, Federal University of Pará, BrazilJorge Salgado Bonnet, University of Nottingham, United Kingdom
Copyright © 2022 Jiménez-Segura, Restrepo-Ángel and Hernandez-Serna. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Luz Jiménez-Segura, luz.jimenez@udea.edu.co