
Redescription of Hargeria rapax (Harger, 1879) and description of H. chetumalensis a new species from the Mexican Caribbean (Crustacea, Peracarida, Tanaidacea, Leptocheliidae) based upon morphological and molecular evidence
- Published
- Accepted
- Received
- Academic Editor
- James Reimer
- Subject Areas
- Biodiversity, Taxonomy, Zoology
- Keywords
- Taxonomy, COI-barcoding, Morphology, Cryptic species, New species
- Copyright
- © 2019 Jarquín-González and Carrera-Parra
- Licence
- This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, reproduction and adaptation in any medium and for any purpose provided that it is properly attributed. For attribution, the original author(s), title, publication source (PeerJ) and either DOI or URL of the article must be cited.
- Cite this article
- 2019. Redescription of Hargeria rapax (Harger, 1879) and description of H. chetumalensis a new species from the Mexican Caribbean (Crustacea, Peracarida, Tanaidacea, Leptocheliidae) based upon morphological and molecular evidence. PeerJ 7:e7472 https://doi.org/10.7717/peerj.7472
Abstract
Until now, Hargeria was considered a monospecific leptocheliid genus, with the species Hargeria rapax considered a taxon with a wide distribution, from the Northwestern Atlantic to the Mexican Caribbean. Herein, after a detailed revision of type and topotype materials and specimens collected from the Mexican Caribbean, a new species H. chetumalensis sp. nov. is described, and the redescription of H. rapax is provided. Also, we found a significant genetic divergence between the two species based on the nucleotide sequences of cytochrome oxidase subunit I, which support the morphological data. The morphological features used to recognize both species are also adequate to link males, females, and juvenile stages, although these species have a high intraspecific polymorphism.
Introduction
In 1879, Oscar Harger described a new species of tanaidacean from Annisquam, Massachusetts, USA, Leptochelia rapax; unfortunately, the description lacks illustrations. The females of this species were distinguished from other leptocheliids by the last segment of the antennule, which is scarcely longer than the preceding one; while the males, by having the elongate and slender antennules and chelipeds (Harger, 1879). Later, Harger (1880) gave a new characterization for this species and provided drawings of the cheliped and full body of the male. Much later, this species was transferred to a new genus, Hargeria, by Lang (1973) because the males have a unique feature within the tanaidaceans, an anal plate, as well as by the presence of two pointed spines on the maxilliped endite in females. Furthermore, Lang (1973) drew some of the female’s appendages; however, no accompanying description was provided.
For over 30 years, Hargeria Lang, 1973 was considered a monospecific genus. Nevertheless, Larsen & Wilson (2002), using phylogenetic methods based on morphological data, found that Hargeria rapax and L. dubia (Krøyer, 1842) formed a sister-group relationship. This relationship between Hargeria and Leptochelia was later corroborated by Bird & Larsen (2009) and also regarded Hargeria as a synonym of Leptochelia. Likewise, Drumm (2010), based on molecular data (COI, H3, and 28S), tested the hypothesis that H. rapax is the sister species of L. dubia and suggested that Hargeria should be considered a junior synonym of Leptochelia since both genera can only be differentiated by presence of the anal plate in males of Hargeria that is absent in Leptochelia.
We point out that the results and conclusions above described were obtained using a very limited number of leptocheliids species because those analyses were done with the main task of elucidating the phylogeny of higher taxa (superfamily Paratanaidoidea by Larsen & Wilson, 2002, and Bird & Larsen, 2009, or Tanaidacea by Drumm, 2010) not only the Leptocheliidae. Recently, Guţu (2016) reestablished Hargeria based on an extensive morphological revision of numerous taxa belonging to Leptocheliidae; he found that Hargeria can be recognized from other leptocheliid genera by having some particular features: both sexes have merus of pereopod 1 with one ventrodistal and one dorsodistal seta; merus of pereopods 2 and 3 with two ventrodistal setae; uropodal endopod five-articulate; females with two flat spines and one distal seta on maxilliped endite; the presence of an anal plate in the male, as well as the presence of a small proximal process, and a large distal process on the fixed finger of the male cheliped.
In the context of geographic distribution, H. rapax is an estuarine species that has been considered with a wide distribution along the Northeastern Atlantic coast from Massachusetts, USA to Chetumal Bay, Mexico (Suárez-Morales et al., 2004; García-Madrigal, Heard & Suárez-Morales, 2005). Herein, we provide a detailed redescription of male and female of H. rapax based on type and topotypical materials. Also, we describe the male and female of a new species of Hargeria based on morphological and molecular evidence. This species represents the first tanaidacean described from the Mexican Caribbean.
Materials and Methods
We studied type and topotypical specimens of H. rapax deposited in the collections of the National Museum of the Natural History, Smithsonian Institution, Washington (USNM); also, specimens identified as H. rapax deposited in the Reference Collection of Benthos (ECOSUR) of El Colegio de la Frontera Sur, Chetumal, Mexico were reviewed.
The tanaids were examined under a stereomicroscope Carl Zeiss SV6. The total length of specimens was measured from the anterior end of the cephalothorax to the posterior margin of the pleotelson. Dissection of appendages was generally performed on the right side of the body; the pieces were mounted in glycerol and sealed with transparent nail varnish. Drawings of taxonomically features were made with a camera lucida at 4–40×. In the description section, we follow Larsen (2003), Bird (2011, 2012), and Guţu (2016) for the general morphology and setal terminology. For morphological analysis of setal and cuticular structures of adult (female and male), juvenile, and manca specimens were processed to be observed by SEM. Specimens were dehydrated in a series of different concentration of hexamethyldisilazane. Once air-dried, they were mounted on aluminum stubs and coated with gold for observation using a JEOL JSM-6010Plus-LA scanning electron microscopy at the Scanning Electron Microscopy Laboratory (LMEB), ECOSUR-Chetumal.
For molecular analysis, we use the whole organism. DNA barcoding was carried out at the Canadian Center for DNA Barcoding (University of Guelph), following the standard protocols of the program “Barcode of the Life.” Cytochrome oxidase subunit I (COI) nucleotide sequences between 579–658 bp were amplified using M13F (5′-TGTAAAACGACGGCCAGT-3′) and M13R (5′-CAGGAAACAGCTATGAC-3′) primers (Messing, 1983). Sequence data, electropherograms, trace files, primer details, photographs, life stages, and collection localities for specimens are available within the project “Tanaidacea from Mexico” at Barcode of Life Data System. Some sequences of H. rapax, L. forresti (Stebbing, 1896), L. longichelipes (Lang, 1973), and Chondrochelia dubia (Krøyer, 1842) (as L. dubia) were obtained from the GenBank database (Table 1).
| Species | Locality | BOLD system # process ID | GenBank accession no. | Reference |
|---|---|---|---|---|
| H. chetumalensis sp. nov. | Guerrero Lagoon, Q. Roo, Mexico |
TANAI104-15 TANAI105-15 TANAI106-15 TANAI107-15 TANAI109-15 |
||
| Raudales, Q. Roo, Mexico |
TANAI071-15 TANAI072-15 TANAI073-15 TANAI074-15 TANAI075-15 TANAI076-15 TANAI077-15 TANAI078-15 TANAI079-15 TANAI110-15 TANAI111-15 TANAI112-15 TANAI113-15 TANAI115-15 TANAI116-15 TANAI117-15 |
Current work | ||
| H. rapax | Dania Beach, FL, USA | HM016214 | Drumm (2010) | |
| Indian River Lagoon, FL, USA | KP254749 | Leray & Knowlton (2015) | ||
| Sheepshead Cove, MD, USA |
KU905783 KU905876 |
R Aguilar et al., unpublished data | ||
| White Marsh Creek, MD, USA | KU906052 | |||
| C. dubia | Fort Lauderdale, FL, USA | HM016215 | Drumm (2010) | |
| L. forresti | Dania Beach, FL, USA | HM016206 | Drumm (2010) | |
| L. longichelipes | Belize | HM016201 | Drumm (2010) |
All sequences were aligned using ClustalW method; the selection of the best model of substitution was determined according with the lowest Bayesian information criterion score. As result, we used Hasegawa–Kishino–Yano, using a discrete Gamma distribution (+G) with five rate categories and by assuming that a certain fraction of sites is evolutionarily invariable (+I) as model to construct a tree using the maximum likelihood analysis. Furthermore, we used the Kimura 2-Parameter model to estimate the average evolutionary divergence over sequence pairs within and between species. All analyses were carried out with Mega 7 (Kumar, Stecher & Tamura, 2015).
The electronic version of this article in portable document format will represent a published work according to the International Commission on Zoological Nomenclature (ICZN), and hence the new names contained in the electronic version are effectively published under that Code from the electronic edition alone. This published work and the nomenclatural acts it contains have been registered in ZooBank, the online registration system for the ICZN. The ZooBank LSIDs (Life Science Identifiers) can be resolved and the associated information viewed through any standard web browser by appending the LSID to the prefix http://zoobank.org/. The LSID for this publication is: urn:lsid:zoobank.org:pub:B55A046E-55AA-491A-ABED-C9BA3D7300DE. The online version of this work is archived and available from the following digital repositories: PeerJ, PubMed Central, and CLOCKSS.
Systematics
Superorder Peracarida Calman, 1904
Order Tanaidacea Dana, 1849
Suborder Tanaidomorpha Sieg, 1980
Family Leptocheliidae Lang, 1973
Subfamily Leptocheliinae Lang, 1973
Genus Hargeria Lang, 1973
Hargeria Lang, 1939: 225.
Type species. Leptochelia rapax Harger, 1879, by original designation (Lang, 1973).
Diagnosis. Amended from Lang (1973). Both sexes. Pereopods 2 and 3 with two dorsodistal setae and one ventrodistal spine on propodus. Pereopod 4 with three distal spines and two setae on carpus. Uropodal endopod with five articles; exopod small, unarticulate. Males. With an anal plate longer than pleotelson. Antennule almost as long as body. Cheliped slender, slightly longer than body; carpus five to eight times as long as broad; fixed finger with one small proximal process and one large distal process on cutting edge. Females. Maxillipedal endite with three flat spines, the subdistal smaller.
Hargeria rapax (Harger, 1879)
Leptochelia rapax: Harger, 1879: 163–164; Harger, 1880: 424–426, figs. 89–90.
Hargeria rapax: Lang, 1973: 202; Sieg, 1983: 422–424; Guţu, 2016: 19, 22, 27, 32, 56–57.
Type material. Syntypes, two males, USNM 32176, Annisquam, Massachusetts, USA, in muddy bottom, four m, coll. Gardiner, August 1879.
Topotypical material. USNM 101781, one ovigerous female, two non-ovigerous females, three males, Pocasset River, Cape Cod, Massachusetts, USA, in seagrass and algae, coll. Burbank, W. D., June 16, 1958. USNM 65779, nine ovigerous females, 26 non-ovigerous females, 11 males, Potomac River, Cobb Island, Maryland, USA, in algae, coll. Cochran, D. M., June 21, 1931.
Diagnosis. Male. Rostrum convex, broad, extending beyond eyes. Anal plate capitate, less than three times as long as broad, do not exceed the length of peduncle and first article of uropodal endopod combined. Cheliped carpus five to seven times as long as broad; fixed finger with pointed proximal process on cutting edge. Pereopod 1 with two distal setae on merus; with six distal setae on carpus. Pereopod 4 with two long distal setae that do not exceed the median length of propodus. Female. Right mandible with incisor smooth and pointed apex. Maxillipedal endite with three flat spines (pointed terminally). Pereopod 1 with one dorsodistal setae and two ventrodistal setae on merus.
Description. Adult male. Topotype USNM 65779, 2.5 mm (Figs. 1B and 3D–3E). Body 5.2 times as long as broad. Cephalothorax oval, longer than broad, slightly longer than the first three pereonites combined. Rostrum convex, broad, exceeding beyond eyes. Pereon about 2.5 times as long as cephalothorax; pereonites 1 and 6 narrower than other pereonites; pereonite 4 broader than other pereonites. Pleon less than 0.2 times the body length. Pleotelson about 0.3 times as long as pleon, with two dorsolateral setae; posterior end with four small dorsodistal setae. Anal plate capitate, less than three times as long as broad, do not exceed the length of peduncle and first article of uropodal endopod combined.
Antennule (Fig. 1C) 0.7 times as long as total body length, first peduncle article longer than cephalothorax, about seven times as long as broad, with three distal setae (one of them small). Second peduncle article about 0.4 times as long as first peduncle article, 5.4 times as long as broad, with two distal setae. Third peduncle article about 0.1 times as long as first peduncle article, 2.5 times as long as broad, with two distal setae. Flagellum 0.6 times as long as first peduncle article; with nine articles; first eight articles longer than broad, with two or three aesthetascs (not illustrated); ninth article minute, with five distal setae.
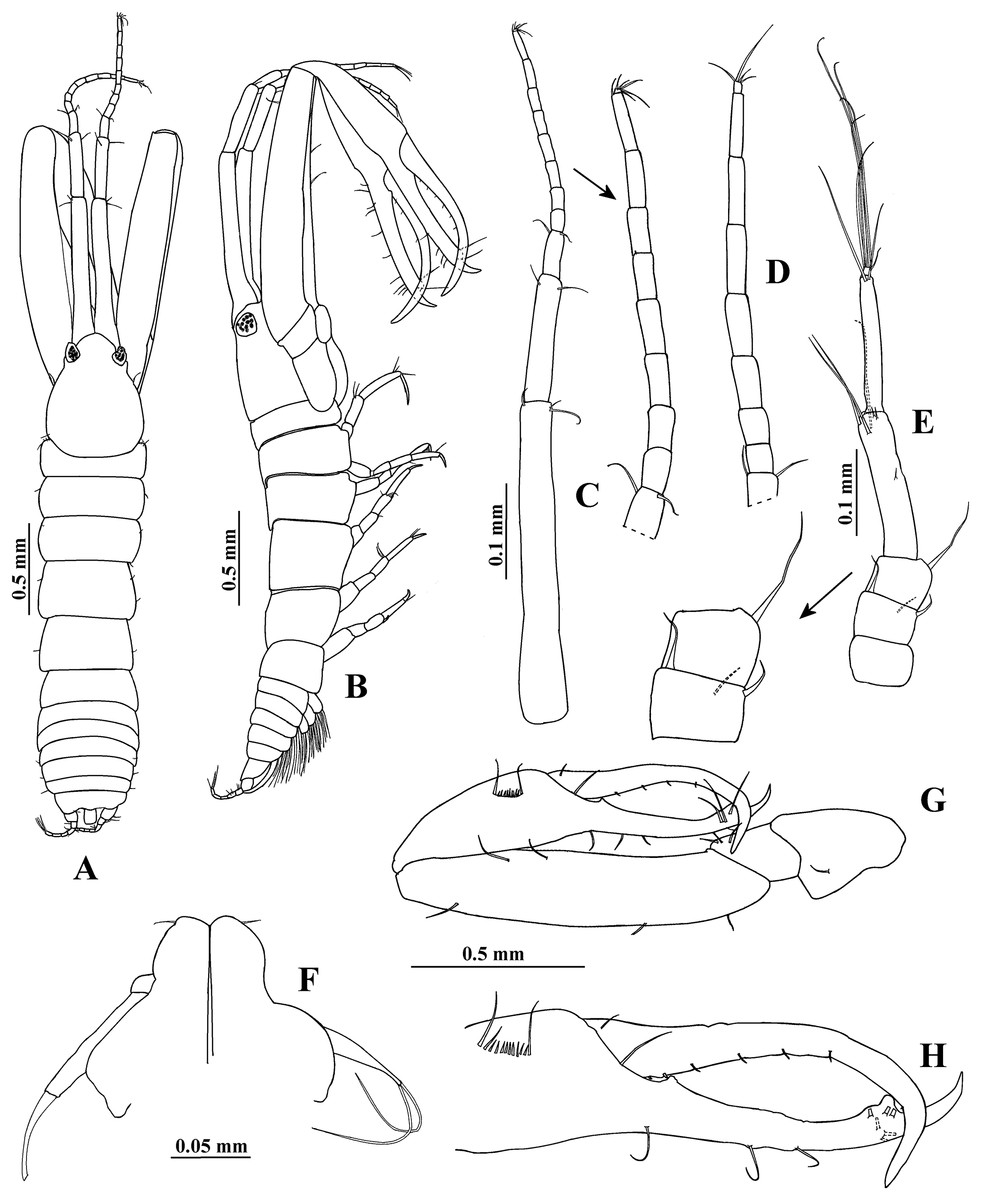
Figure 1: Hargeria rapax, type and topotype specimens.
(A) Habitus. Topotype USNM 65779, adult male, 2.5 mm. (B) Habitus. (C) Antennule. (E) Antenna. (F) Maxilliped and maxillules. (G) Cheliped closed. (H) Detail of chelipedal cutting edge. Topotype USNM 101781, adult male, 2.3 mm. (D) Detail of antennular flagellum; asthetasc were omitted for clarity.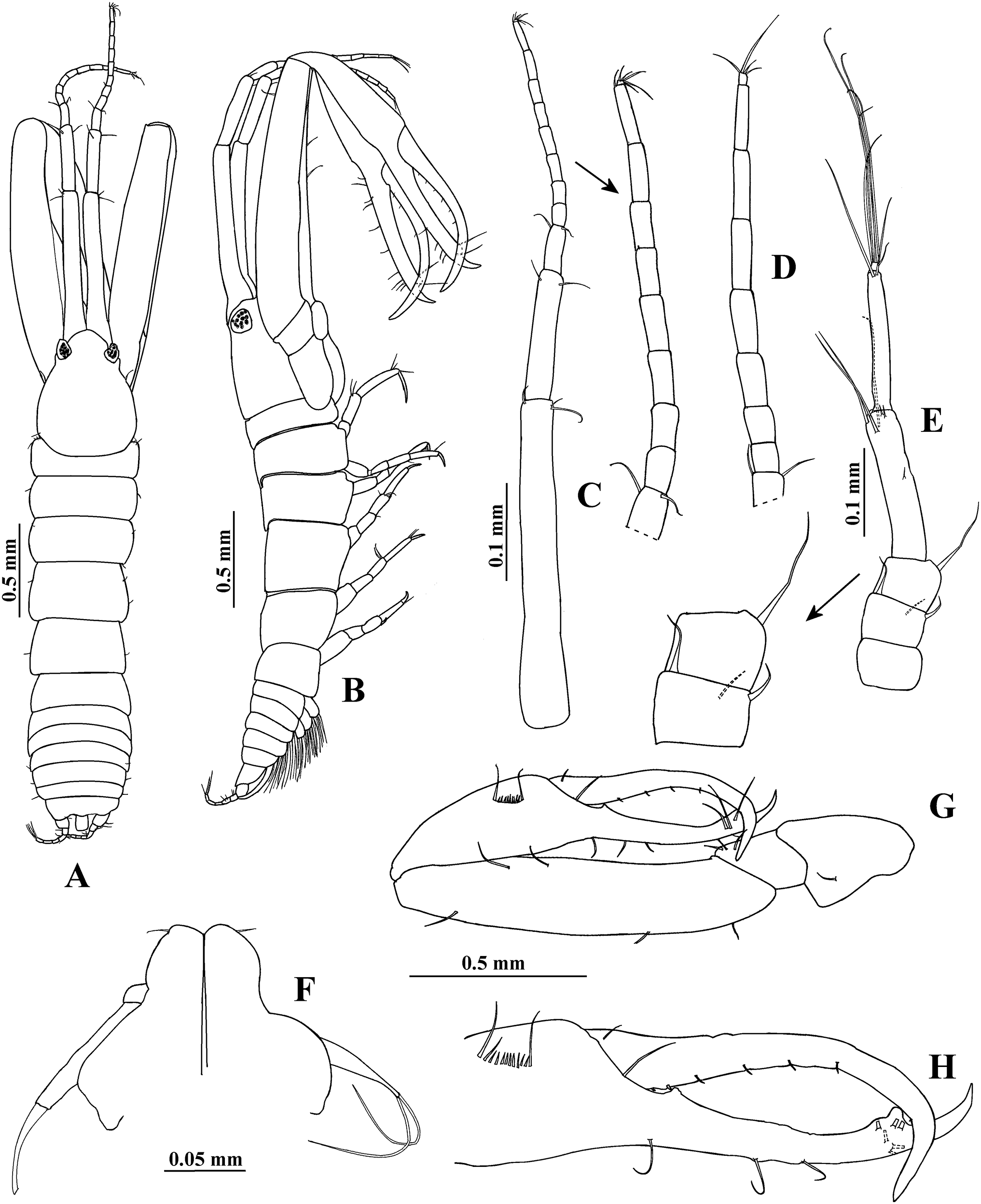{kind=link}
Antenna (Fig. 1E). First article as long as broad, naked. Second article 0.7 times as long as broad, with two distal setiform spines and one simple seta. Third article 0.8 times as long as broad, with one dorsodistal setiform spine. Fourth article 4.7 times as long as broad, with six setae on distal half (three of them small). Fifth article 0.9 times as long as fourth article, about eight times as long as broad, with two distal setae. Sixth article minute, with six distal setae.
Mouthparts reduced. Maxilliped (Fig. 1F) rudimentary, with two proximal setae; maxillule palps with two setae.
Cheliped (Figs. 1G–1H) 1.3 times as long as total body length. Basis 1.5 times as long as broad, with one dorsolateral seta. Merus longer than broad, with three ventrodistal setae. Carpus about five times as long as broad, with two ventrodistal setae and three dorsal setae. Propodus with seta near dactylus articulation; comb-row of 13 setae. Fixed finger with two processes on cutting edge, proximal small, triangular, with pointed apex, distal long, tubular, with blunt apex; with nine setae, six of them situated on ventral side. Dactylus with one proximal seta and seven ventral minute spines.
Pereopod 1 (Fig. 2B) with basis 4.1 times as long as broad, with two dorsoproximal setae, one of them small. Ischium with one seta. Merus 1.2 times as long as carpus, 2.4 times as long as broad, with two distal setae. Carpus twice as long as broad, with six distal setae. Propodus 3.9 times as long as broad, with distal setae (three dorsal and one ventral). Dactylus and unguis combined as long as propodus. Dactylus with one proximal seta.

Figure 2: Hargeria rapax, topotype specimens.
Adult male, 2.3 mm, USNM 101781. (A) Cheliped open. Adult male, 2.5 mm, USNM 65779. (B) Pereopod 1. (C) Pereopod 2. (D) Pereopod 3. (E) Pereopod 4.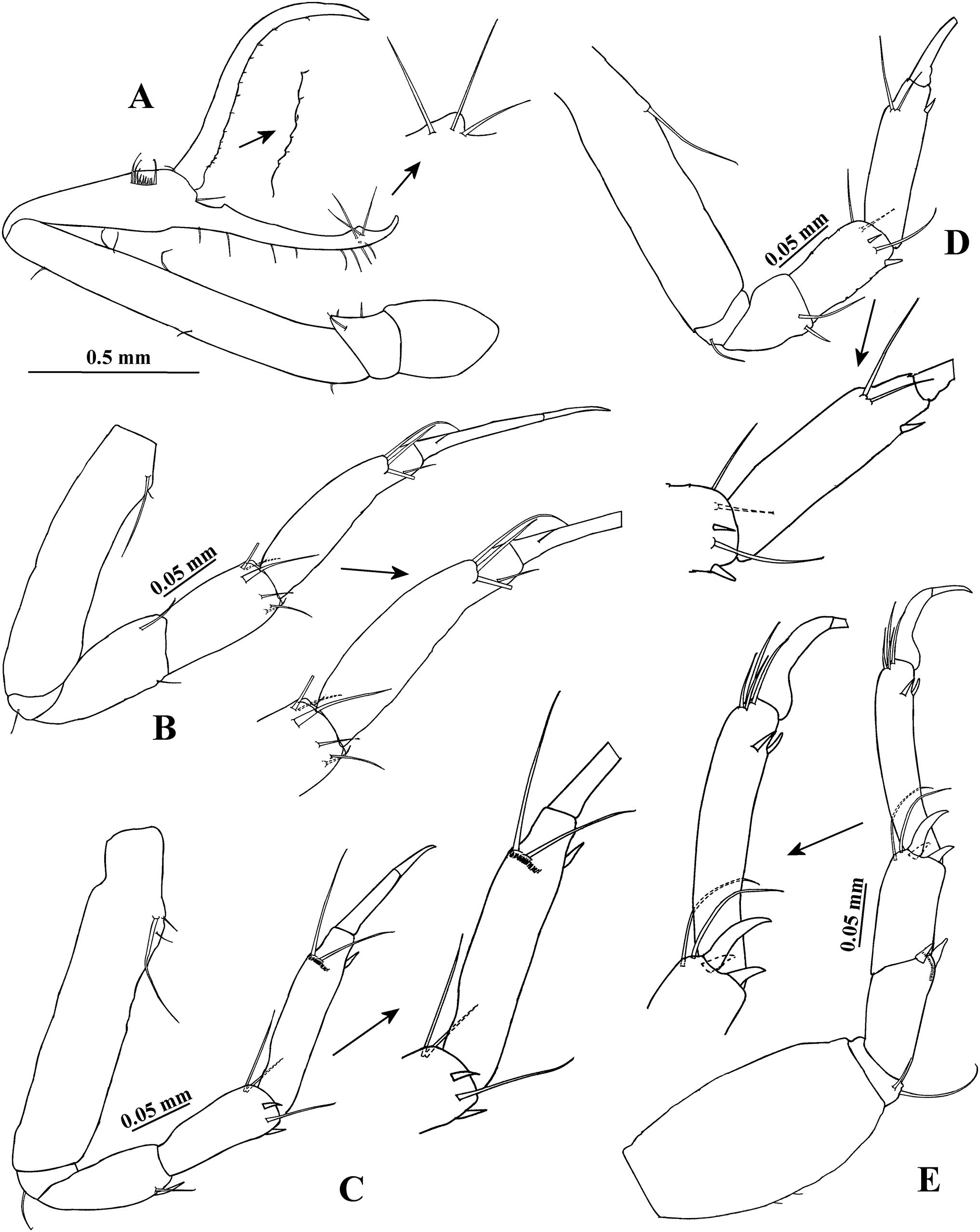{kind=link}
Pereopod 2 (Fig. 2C) with basis 4.9 times as long as broad, with three dorsoproximal setae, two of them small. Ischium with one seta. Merus as long as carpus, 2.4 times as long as broad, with distoventral seta and spine. Carpus 2.4 times as long as broad, with three distal setae and two ventrodistal spines. Propodus 4.6 times as long as broad, with two dorsodistal setae, one ventrodistal spine and scales. Dactylus and unguis combined about 0.6 times as long as propodus, naked.
Pereopod 3 (Fig. 2D) similar to pereopod 2, but basis four times as long as broad. Merus 1.4 times as long as broad. Carpus 1.7 times as long as broad. Propodus 3.2 times as long as broad.
Pereopod 4 (Fig. 2E) with basis 1.8 times as long as broad, with two small ventral setae. Ischium with two setae. Merus as long as carpus, twice as long as broad, with two distal spines and scales. Carpus 2.1 times as long as broad, with three distal spines and two long setae that do not exceed the median length of propodus. Propodus 5.3 times as long as broad, with four dorsodistal setae and two ventrodistal spines. Dactylus and unguis combined about 0.5 times as long as propodus, naked.
Pereopod 5 (Fig. 3A) similar to pereopod 4, but basis 1.6 times as long as broad, with one ventrodistal seta. Propodus with five dorsodistal setae.

Figure 3: Hargeria rapax, topotype specimens.
Adult male, 2.5 mm, USNM 65779. (A) Pereopod 5. (B) Pereopod 6. (C) Pleopod 1. (D) Pleotelson and anal plate. (E) Anal plate. (F) Uropod. Adult male, 2.3 mm, USNM 101781. (G) Uropod.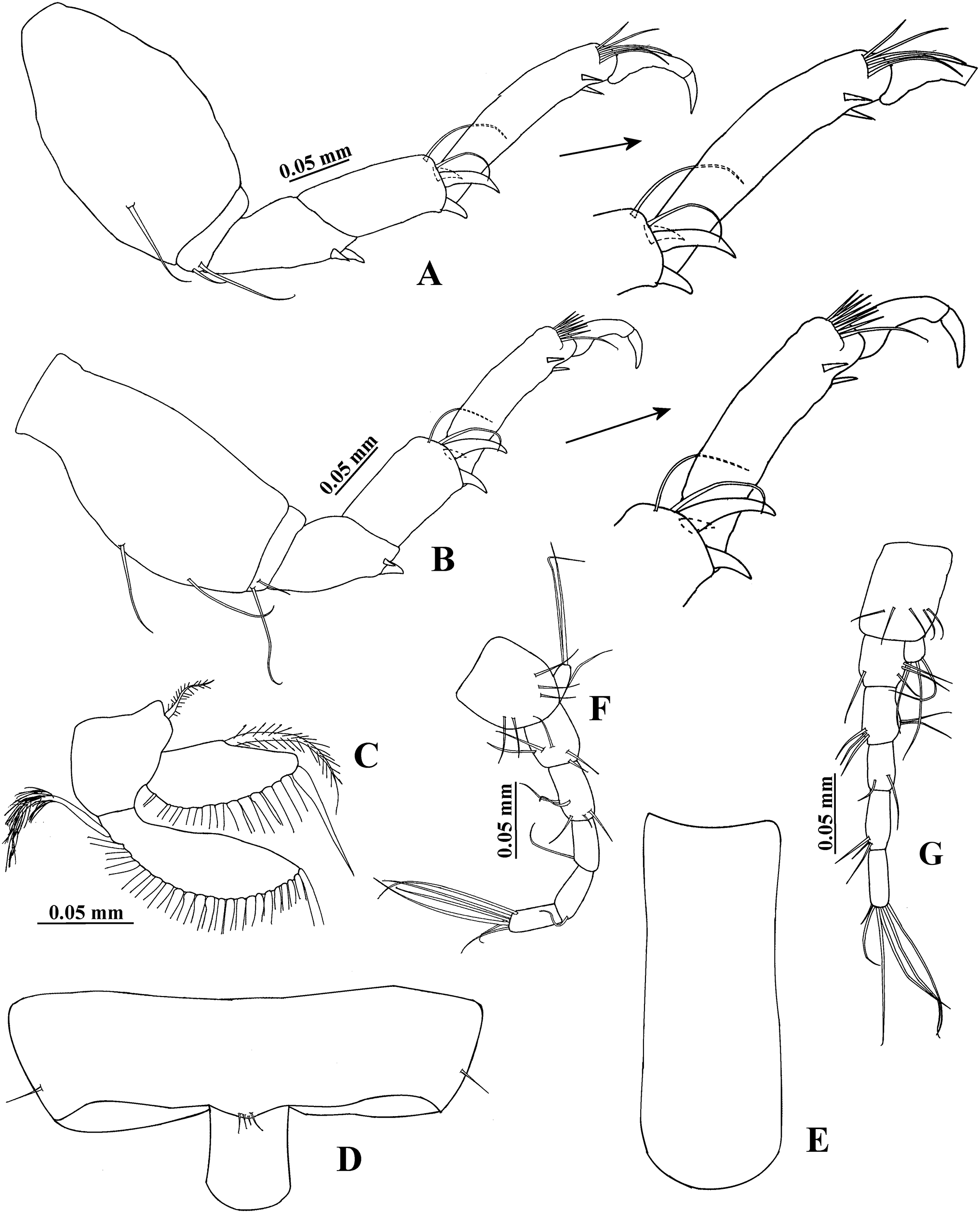{kind=link}
Pereopod 6 (Fig. 3B) similar to pereopods 4 and 5, but basis 1.9 times as long as broad, with two ventral setae. Carpus 1.9 times as long as broad. Propodus 4.3 times as long as broad, with eight dorsodistal setae.
Pleopod 1 (Fig. 3C) with one circumplumose dorsal seta on peduncle. Exopod with one proximoventral circumplumose seta and 21 ventral setae. Endopod with one circumplumose dorsal seta and 13 ventral setae.
Uropod (Fig. 3F) with five dorsal setae on peduncle. Exopod 0.6 times as long as first article of uropodal endopod, with three setae. Endopod first article broader than other articles; first and second article with five distal setae; third article smaller than other articles, with one distal seta; fourth and fifth articles subequal; fifth article with six distal setae.
Non-ovigerous female, Topotype USNM 65779, 3.7 mm (Fig. 4A). Body 6.2 times as long as broad. Cephalothorax oval, 1.3 times as long as broad, 0.7 times as long as first three pereonites combined. Pereon three times as long as cephalotorax; pereonites 2 and 3 subequal in length; pereonite 4 longer than other pereonites. Pleon about 0.2 times as long as body length. Pleotelson (Fig. 6E) 0.3 times as long as pleon, with four dorsolateral setae; posterior end rounded, slightly expanded, with four distal setae and two small plumose sensory setae.
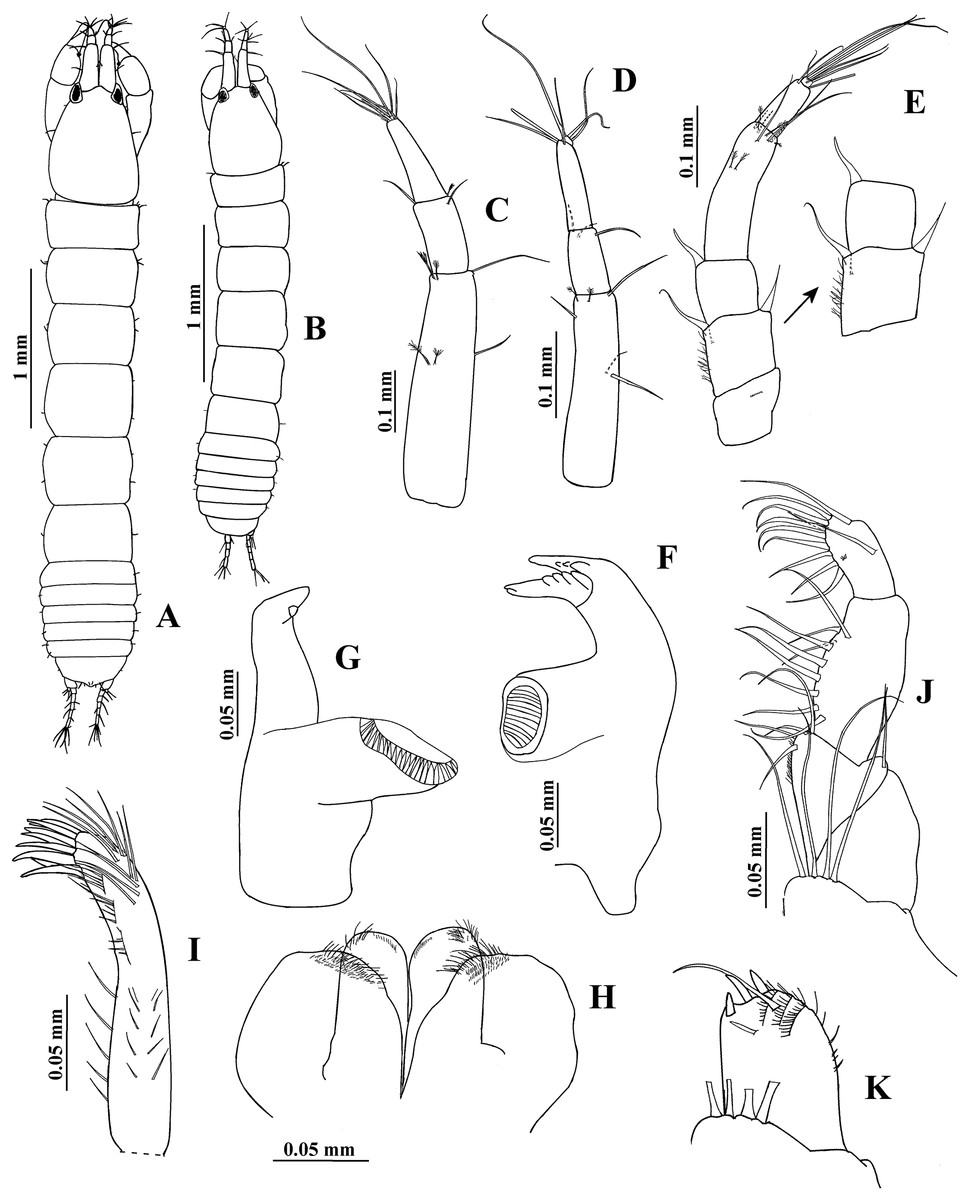
Figure 4: Hargeria rapax, topotype specimens.
Non-ovigerous female, 3.7 mm, USNM 65779. (A) Habitus. (C) Antennule. (E) Antenna. (F) Left mandible. (G) Right mandible. (H) Labium. (I) Maxillule. (J) Maxillipedal palp. (K) Maxillipedal endite. Ovigerous female, 2.9 mm, USNM 101781. (B) Habitus. (D) Antennule.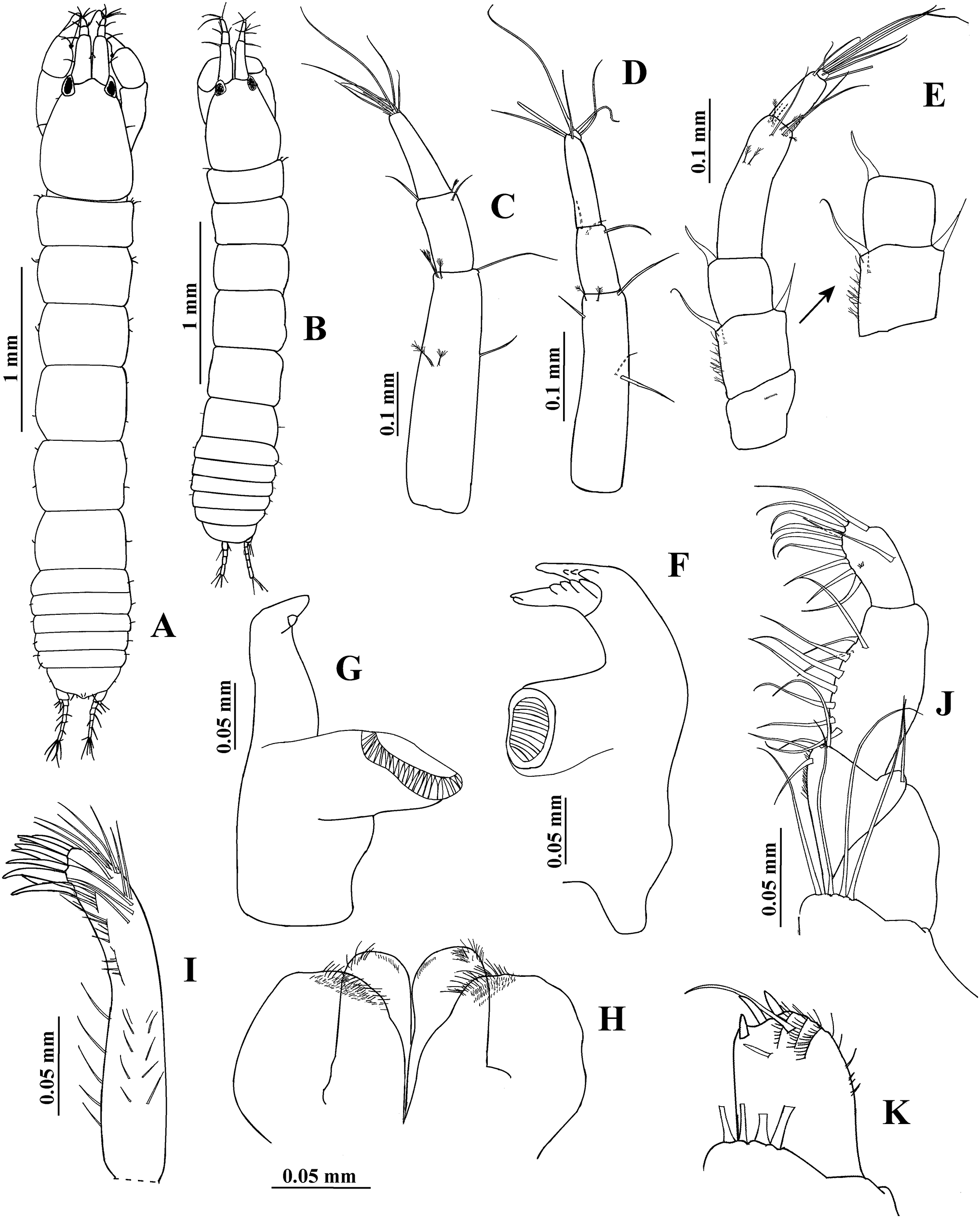{kind=link}
Antennule (Fig. 4C) with four articles. First article 3.7 times as long as broad, with two setae and four plumose sensory setae on distal half. Second article 0.3 times as long as first article, 1.9 times as long as broad, with two distal setae and one plumose sensory seta. Third article 0.4 times as long as first article, three times as long as broad, with two distal setae and one aesthetasc. Fourth article minute, with four distal setae.
Antenna (Fig. 4E). First article 1.2 times as long as broad, with scales. Second article slightly longer than broad, with two distal spines and small distolateral setae. Third article as long as broad, with one distal spine. Fourth article 3.3 times as long as broad, with three setae and plumose sensory setae on distal half. Fifth article 0.4 times as long as fourth article, 2.2 times as long as broad, with two distal setae. Sixth article minute, with five distal setae.
Mouthparts. Left mandible (Fig. 4F) with incisor shorter than lacinia mobilis, with three small processes; lacinia mobilis with four small processes and bifurcate apex; molar cylindrical, with triturative ridges. Right mandible (Fig. 4G) with incisor smooth and pointed apex; molar thick, with triturative ridges. Labium (Fig. 4H) distally setose, with outer lobe shorter but broader than inner lobe. Maxillule (Fig. 4I) setose, with nine spines. Maxilliped (Fig. 4J) with four long setae on basis; first palp article 1.2 times as long as broad, naked; second palp article 1.5 times as long as broad, with one dorsodistal seta, four ventrodistal seta and small lateral setae; third palp article 1.9 times as long as broad, with nine ventral setae; fourth palp article 2.1 times as long as broad, with 10 ventral setae, one dorsolateral seta and scales; endite (Fig. 4K) with three flat spines (pointed terminally), the subdistal smaller, one dorsodistal seta, coupling hook, lateral setae and scales.
Cheliped (Figs. 5A and 5B) 0.2 times as long as total body length. Basis 1.4 times as long as broad, with one dorsal seta. Merus longer than broad, with three ventral setae. Carpus 1.8 times as long as broad, with three ventrodistal setae and three dorsal setae. Propodus with seta near dactylus articulation; comb-row with five setae. Fixed finger with cutting edge raised, with eight setae on distal half. Dactylus with proximodorsal seta and eight lamellae on ventral edge.
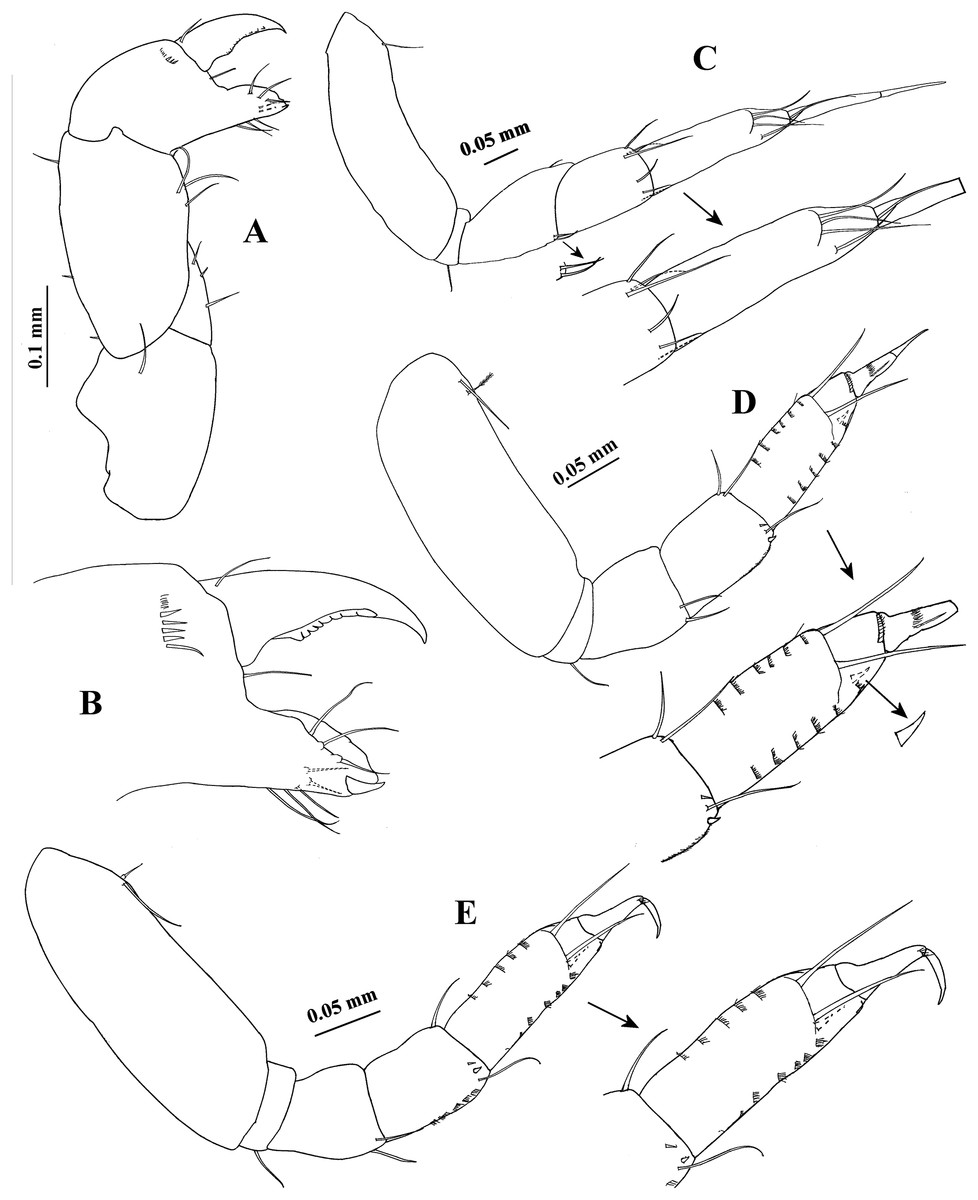
Figure 5: Hargeria rapax, topotype specimen USNM 65779.
Non-ovigerous female, 3.7 mm. (A) Cheliped. (B) Detail of cutting edge of cheliped. (C) Pereopod 1. (D) Pereopod 2. (E) Pereopod 3.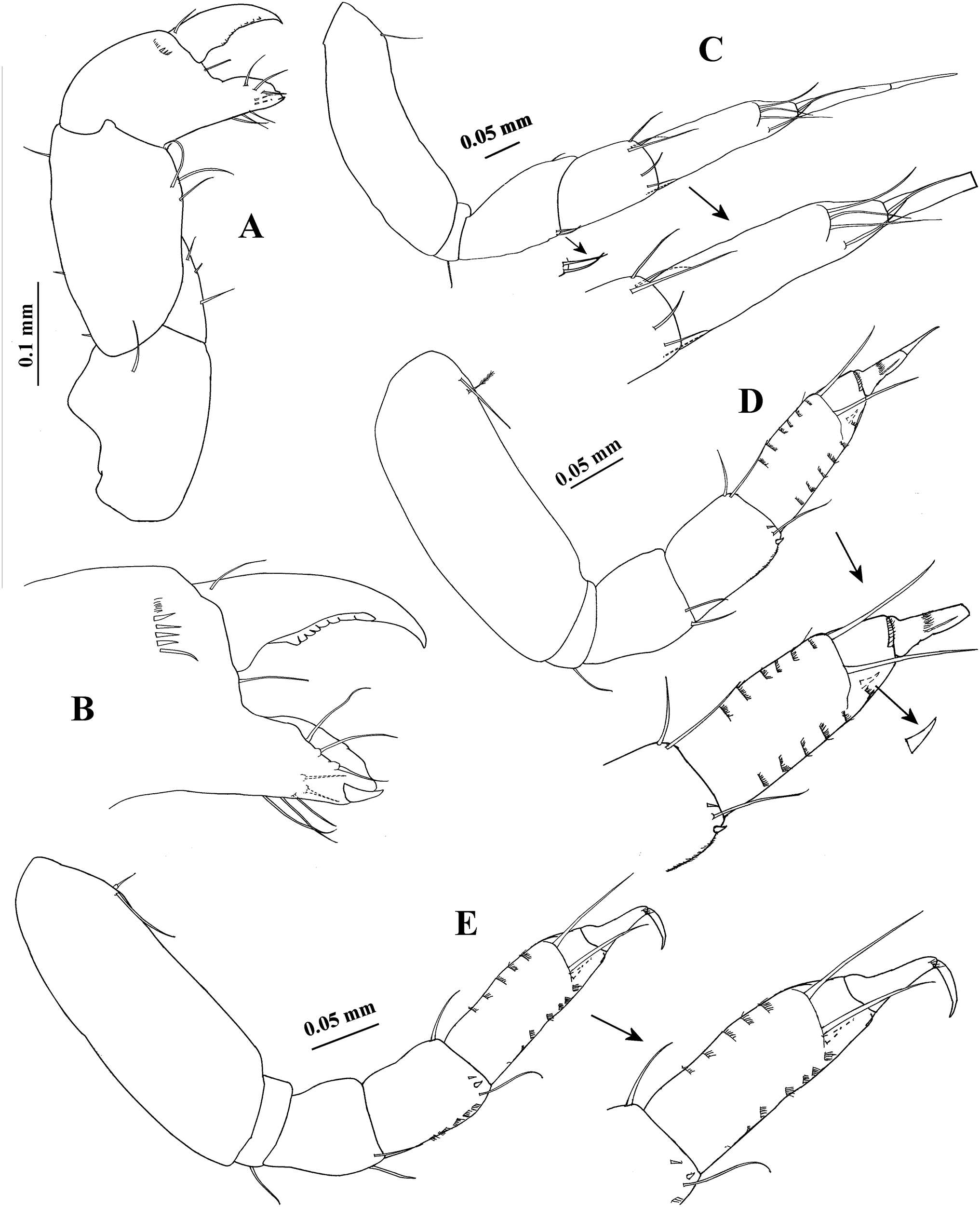{kind=link}
Pereopod 1 (Fig. 5C) with basis 3.6 times as long as broad, with one proximal seta. Ischium with one seta. Merus 1.2 times as long as carpus, 1.6 times as long as broad, with small setae (one dorsal, two ventral). Carpus 1.5 times as long as broad, with six distal setae. Propodus 3.2 times as long as broad, with distal setae (three dorsodistal and one ventral). Dactylus and unguis combined as long as propodus; dactylus with one proximal seta.
Pereopod 2 (Fig. 5D) with basis 2.5 times as long as broad, with one proximal seta and one plumose sensory seta. Ischium with one seta. Merus as long as carpus, 1.3 times as long as broad, with two distoventral setae. Carpus 1.5 times as long as broad, with three setae, two ventrodistal minute spines and scales. Propodus 2.5 times as long as broad, with two dorsodistal setae, small distoventral spine and scales. Dactylus and unguis combined 0.6 times as long as propodus; dactylus with subdistal seta and scales.
Pereopod 3 (Fig. 5E) similar to pereopod 2, but basis 2.7 times as long as broad. Propodus 2.4 times as long as broad.
Pereopod 4 (Fig. 6A) with basis 1.8 times as long as broad, naked. Ischium with two setae. Merus 1.7 times as long as carpus, with two ventrodistal spines. Carpus 1.2 times as long as broad, with two distal setae, three spines and scales. Propodus 3.2 times as long as broad, with three dorsodistal setae, one pinnate seta, two ventrodistal serrate spines and scales. Dactylus and unguis combined 0.7 times as long as propodus; dactylus and unguis joint with scales.

Figure 6: Hargeria rapax, topotype specimens USNM 65779.
Non-ovigerous female, 3.7 mm. (A) Pereopod 4. (B) Pereopod 5. (C) Pereopod 6. (D) Pleopod 1. (E) Pleotelson. (F) Uropod. Ovigerous female, 2.9 mm. (G) Uropod.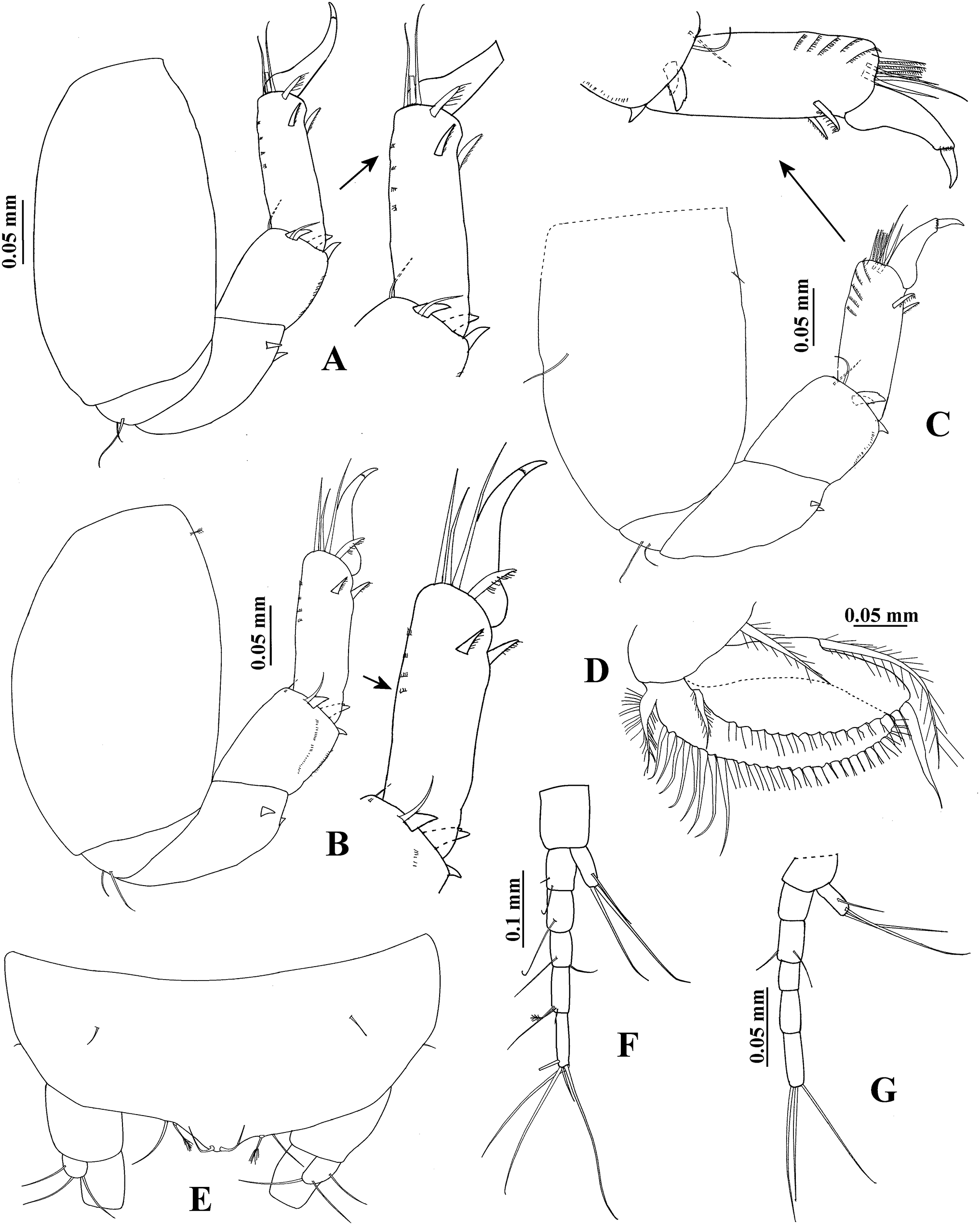{kind=link}
Pereopod 5 (Fig. 6B) similar to pereopod 4.
Pereopod 6 (Fig. 6C) similar to pereopods 4 and 5, but basis with small dorsal seta and one ventroproximal seta. Merus twice times as long as broad. Propodus 2.7 times as long as broad, with three dorsodistal simple setae, four pinnate setae, and two ventrodistal serrate spines.
Pleopod 1 (Fig. 6D) with one circumplumose dorsal seta on peduncle. Exopod with one proximoventral circumplumose seta and 28 ventral setae. Endopod with one dorsal circumplumose seta, one ventroproximal circumplumose seta, and 15 ventral setae.
Uropod (Fig. 6F) with naked peduncle. Exopod uniarticulate, as long as first article of endopod, with three setae. Endopod with five articles, first two articles with subequal length, first article with two setae, second article with one seta; third article shorter than other articles, with two setae; fourth article longer than other articles, with two setae and one plumose sensory seta; fifth article more slender than other articles, with five setae.
Variability. Males. Syntype USNM 32176, 2.6 mm. Body (Fig. 1A) 4.5 times as long as broad. Topotype USNM 101781, 2.3 mm. Antennule (Fig. 1D) with eight flagellar articles; cheliped (Fig. 2A) with carpus about seven times as long as broad, dactylus with 14 min spines, fixed finger with 11 setae, eight of them situated on ventral side; first article of uropodal endopod with four setae (Fig. 3G). Ovigerous female. Topotype USNM 101781, 2.9 mm (Fig. 4B). Antennule (Fig. 4D), first article with four setae and two plumose sensory setae on distal half; third article 0.5 times as long as first article; maxilliped with three long setae on basis, with six ventral setae on third palp article, with eight ventral setae on fourth article of palp; pereopod 1 with four distal setae on carpus; second article of uropodal endopod with two setae (Fig. 6G).
Distribution. Northwestern Atlantic, from Massachusetts to Florida.
Habitat. Shallow-water, in muddy bottom and sandy bottom with seagrass and algae.
Remarks. Historically, H. rapax has been considered a species with a wide distribution, from the Northwestern Atlantic to the Mexican Caribbean. However, the morphological and molecular analysis of specimens from Mexican Caribbean, have shown that they differ from H. rapax (see below remarks section of H. chetumalensis sp. nov.).
Hargeria rapax, as noted above, was regarded as a junior synonym of Leptochelia based on phylogenetic studies where the sister relationships between H. rapax and L. dubia (currently C. dubia) was observed. Recently, Guţu (2016) reestablished it based on a morphological revision of Leptocheliidae. Herein, we agree with this idea by the following reasons; the male of Hargeria is mainly characterized by having an anal plate, which is absent in Leptochelia and Chondrochelia Guţu, 2016. Also, the cheliped fixed finger of Hargeria has a small proximal process and a large distal process on cutting edge, whereas in Leptochelia can be observed only a medial process and in Chondrochelia one large proximal process and one small distal process. Moreover, Hargeria has uropodal exopod uniarticulate, while in Leptochelia it is biarticulate and in Chondrochelia uni- or biarticulate. In Hargeria the length of cheliped carpus is up to 8.0 times as long as broad, whereas in Leptochelia the carpus is more than eight times as long as broad; Hargeria has uropodal endopod five-articulate and the first two articles without long setae, while Leptochelia has uropodal endopod six-articulate and the first two articles with long setae.
Regarding females, the maxillipedal endite of Hargeria has three flat spines (the subdistal smaller); while Leptochelia only has two flat spines and Chondrochelia three to five flat spines. Other differences between these genera can be seen in propodus of Pereopods 2 and 3. For example, although Hargeria and Leptochelia have the same setation on propodus of Pereopod 2 (two dorsodistal setae and one ventrodistal spine), they differ on propodus of Pereopod 3, Hargeria has two dorsodistal setae and one ventrodistal spine, whereas Leptochelia has one dorsodistal seta and one ventrodistal spine. A similar condition occurs when compare Hargeria with Chondrochelia; both genera have two dorsodistal setae and one ventrodistal spine on propodus of Pereopod 3, but they differ because Hargeria has two dorsodistal setae and one ventrodistal spine on propodus of Pereopod 2, while Chondrochelia has three dorsodistal setae and one ventrodistal spine. Furthermore, these morphological differences are supported by the results of the molecular analysis (see below).
Hargeria chetumalensis sp. nov.
urn:lsid:zoobank.org:act:A8D8DCA7-E71C-4124-83B7-F5E66CDA31A1
Hargeria rapax: Carrera-Parra, González & Salazar-Vallejo, 1997: 61–64; Suárez-Morales et al., 2004: 47–53, figs. 21–24; García-Madrigal, Heard & Suárez-Morales, 2005: 1165. (non Harger, 1879).
Type material. Holotype adult male, ECOSUR 0209, 2.9 mm, Raudales, 18°42′25.2″N 88°15′18″W, Quintana Roo, Mexico, in Bathophora sp. on rock, 0.5 m, May 28, 2015. Paratypes: 13 ovigerous females ECOSUR 0210, 44 non-ovigerous females ECOSUR 0211, 17 adult males ECOSUR 0212, same data as holotype.
Additional material. ECOSUR-C1079, five ovigerous females, 10 non-ovigerous female, three males, Calderitas, 18°29′13.2″N 87°45′25.2″W, Chetumal Bay, Quintana Roo, Mexico, in Bathophora sp. on rocks, June 28, 2016. ECOSUR-C1080, 20 ovigerous females, 32 non-ovigerous females, nine males, mouth of Guerrero Lagoon, 18°42′30.4″N 88°09′05.2″W, Quintana Roo, Mexico, in Bathophora sp. on rocks, April 23, 2016.
Molecular material. ECOSUR TANAI104-15, TANAI105-15, TANAI106-15, TANAI107-15, TANAI109-15 (one manca, two juvenile females, two non-ovigerous females): Guerrero Lagoon, 18°41′19.3″N 88°15′50.4″W, Quintana Roo, Mexico, in Bathophora sp. on rocks, 0.5 m, May 28, 2015. ECOSUR TANAI071-15, TANAI072-15, TANAI073-15, TANAI074-15, TANAI075-15, TANAI076-15, TANAI077-15, TANAI078-15, TANAI079-15, TANAI110-15, TANAI111-15, TANAI112-15, TANAI113-15, TANAI115-15, TANAI116-15, TANAI117-15 (two mancae, four juvenile females, four non-ovigerous females, two ovigerous females, two juvenile males, two adult males): Raudales, 18°42′25.2″N 88°15′18″W, Quintana Roo, Mexico, in Bathophora sp. on rocks, 0.5 m, May 28, 2015.
Diagnosis. Male. Rostrum pointed, narrow, not extending beyond eyes. Anal plate tongue-shaped, more than times as long as broad, extending beyond length of peduncle and first article of uropodal endopod combined. Cheliped with carpus about eight times as long as broad; fixed finger with blunt proximal process on cutting edge. Pereopod 1 with one small ventrodistal serrate seta on merus; with three distal serrate setae on carpus. Pereopod 4 with two long distal serrate setae extending beyond the median length of propodus. Female. Right mandible with incisor crenulate and slightly bifurcate apex. Maxillipedal endite with three flat spines (two of them rounded terminally, the smaller pointed). Pereopod 1 with one small ventrodistal serrate seta on merus.
Etymology. The specific epithet is derived from the locality name of the Chetumal Bay System, where the species was found.
Description. Adult male. Holotype ECOSUR0209, 2.9 mm (Figs. 7A–7B and 7G). Body 4.2 times as long as broad. Cephalothorax oval, 1.3 times as long as broad, 0.8 times as long as first three pereonites combined. Rostrum pointed, narrow, not extending beyond eyes. Pereon about 2.8 times as long as cephalothorax; pereonite 1 shorter than other pereonites; pereonites 2, 3, and 6 with subequal length, pereonites 4 and 5 with subequal length. Pleon about 0.2 times as long as total body length. Pleotelson about 0.2 time as long as pleon, with eight dorsolateral setae; posterior end with four small dorsodistal setae. Anal plate (Fig. 9C) tongue-shaped, more than three times as long as broad, extending beyond length of peduncle and first article of uropodal endopod combined.
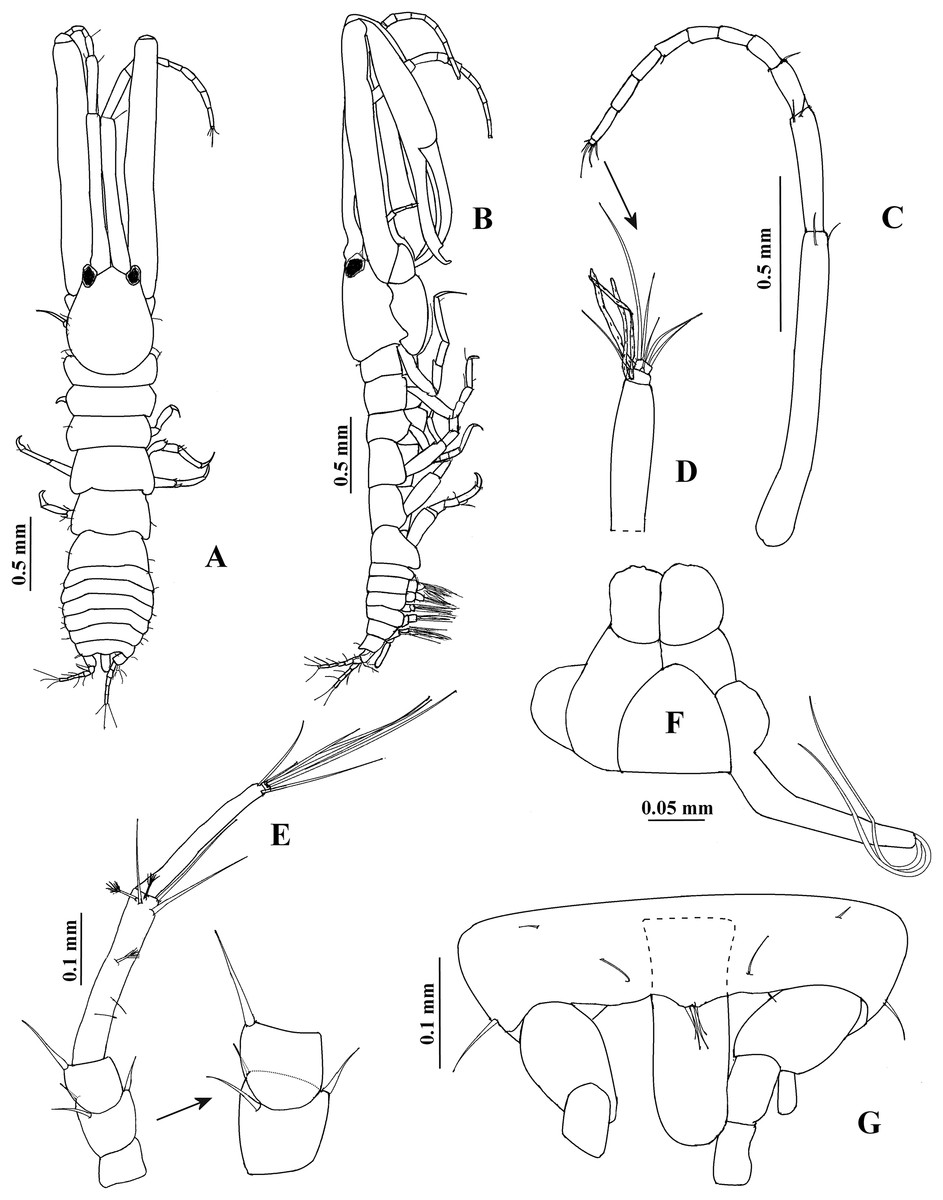
Figure 7: Hargeria chetumalensis sp. nov. Holotype ECOSUR0209, adult male, 2.9 mm.
(A) Habitus, dorsal view. (B) Habitus, lateral view. (C) Antennule, (D) Distal article, antennule. (E) Antenna. (F) Maxilliped and maxillules. (G) Pleotelson with anal plate.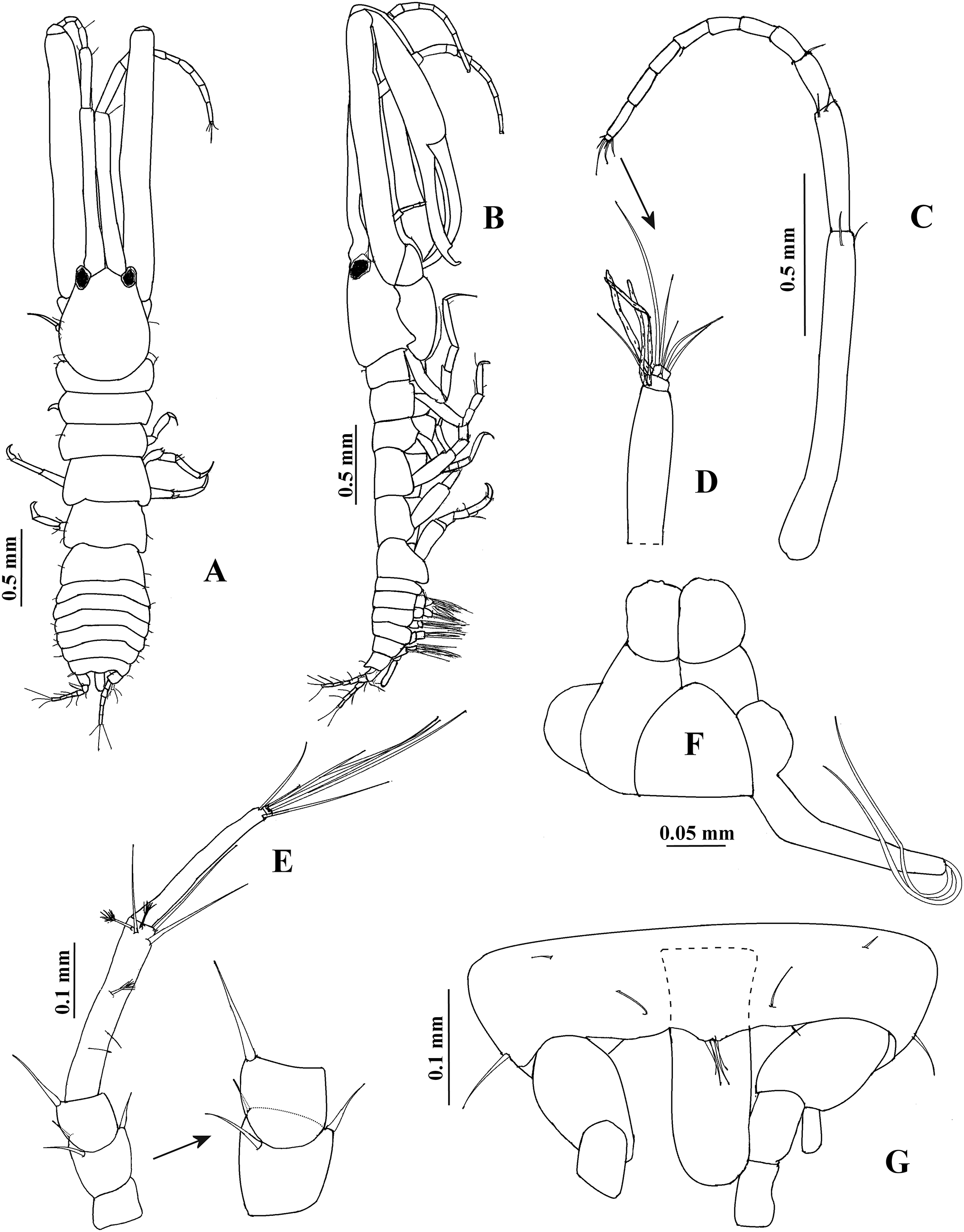{kind=link}
Antennule (Figs. 7C and 7D) 0.9 times as long as total body length; first peduncle article 1.5 times as long as cephalothorax, more than eight times as long as broad, with two distal setae; second peduncle article 0.4 times as long as first peduncle article, 5.4 times as long as broad, with two distal setae; third peduncle article 0.2 times as long as first peduncle article, 2.8 times as long as broad, with two distal setae. Flagellum 0.8 times as long as first article, with nine articles; first seven articles longer than broad, each with 1–2 aesthetascs; last two articles smallest; last article with three distal setae.
Antenna (Fig. 7E). First article about 0.8 times as long as broad, naked. Second article as long as broad, with two distal setiform spines and one simple seta. Third article 0.9 times as long as broad, with one dorsodistal setiform spine. Fourth article 6.6 times as long as broad, with two small subproximal setae, three simple setae and three plumose sensory setae on distal half. Fifth article 0.9 times as long as fourth article, 8.5 times as long as broad, with two distal setae. Sixth article minute, with five distal setae.
Mouthparts reduced. Maxilliped rudimentary (Fig. 7F), maxillule palp with two setae.
Cheliped (Figs. 8A–8C) 1.6 times as long as total body length. Basis 2.1 times as long as broad, naked. Merus longer than broad, with two ventrodistal setae. Carpus eight times as long as broad, with two subdistal ventral setae and three dorsal setae. Propodus with seta near dactylus articulation, comb-row with 24 setae (not illustrated). Fixed finger with two processes on cutting edge; proximal small, triangular, striate, with blunt apex; distal long, tubular, striate, with blunt apex; distally crenulate, with ten setae, six of them situated on ventral side. Dactylus with ten ventral minute spines.

Figure 8: Hargeria chetumalensis sp. nov. Holotype ECOSUR0209, adult male, 2.9 mm.
(A) Cheliped. (B) Proximal chelipedal process. (C) Distal chelipedal process. (D) Pereopod 1. (E) Pereopod 2. (F) Pereopod 3. (G) Pereopod 4.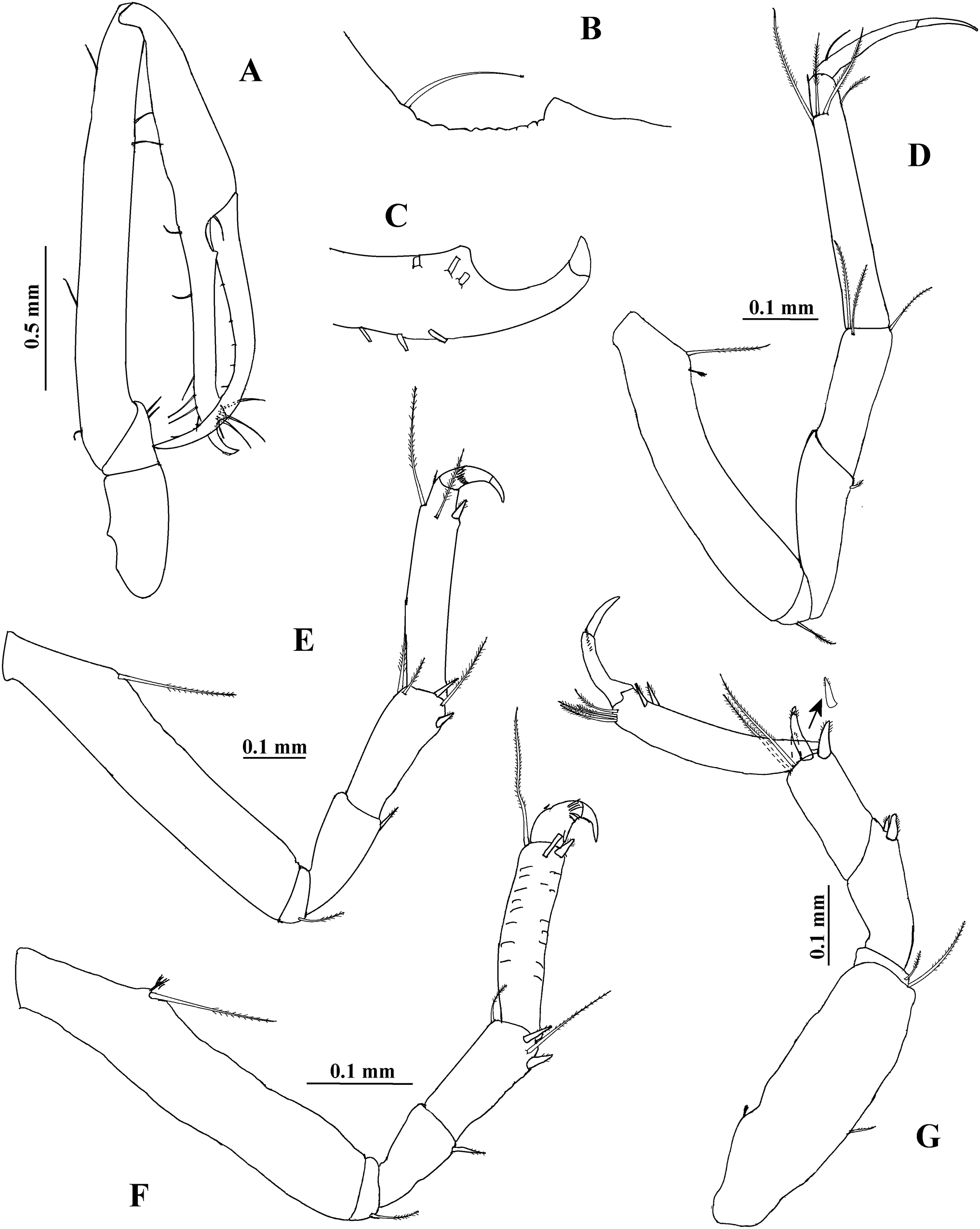{kind=link}
Pereopod 1 (Fig. 8D) with basis 5.1 times as long as broad, with one proximal serrate seta and one small plumose sensory seta. Ischium with one serrate seta. Merus 1.2 times as long as carpus, 3.1 times as long as broad, with one small ventrodistal serrate seta. Carpus 3.2 times as long as broad, with three serrate setae. Propodus 5.6 times as long as broad, with distal serrate setae (three dorsal and one ventral). Dactylus and unguis combined 0.8 times as long as propodus; dactylus with one dorsoproximal seta.
Pereopod 2 (Fig. 8E) with basis 5.5 times as long as broad, with one dorsoproximal serrate seta. Ischium with one serrate seta. Merus 0.9 times as long as carpus, twice as long as broad, with one ventrodistal serrate spine. Carpus 2.5 times as long as broad, with three distal serrate setae and two ventrodistal serrate spines. Propodus 4.9 times as long as broad, with two dorsodistal serrate setae and one ventrodistal serrate spine. Dactylus and unguis combined 0.3 times as long as propodus; dactylus with subdistal scales.
Pereopod 3 (Fig. 8F) similar to pereopod 2, but carpus 2.1 times as long as broad and with two distal serrate setae.
Pereopod 4 (Fig. 8G) with basis 3.2 times as long as broad, with one proximodorsal small plumose sensory seta and one mesioventral serrate seta. Ischium with two unequal serrate setae. Merus 1.2 times as long as carpus, 2.6 times as long as broad, with two distal serrate spines. Carpus 2.4 times as long as broad, with three spines and two long distal serrate setae extending beyond median length of propodus. Propodus 5.5 times as long as broad, with four dorsodistal serrate setae and two ventrodistal serrate spines. Dactylus and unguis combined about 0.5 times as long as propodus; dactylus and unguis joint with scales.
Pereopod 5 (Fig. 9A) similar to pereopod 4, but basis 2.6 times as long as broad, with one ventrolateral serrate seta and three plumose sensory setae on proximal half. Serrate setae on carpus not extending beyond median length of propodus. Propodus with three dorsodistal serrate setae.
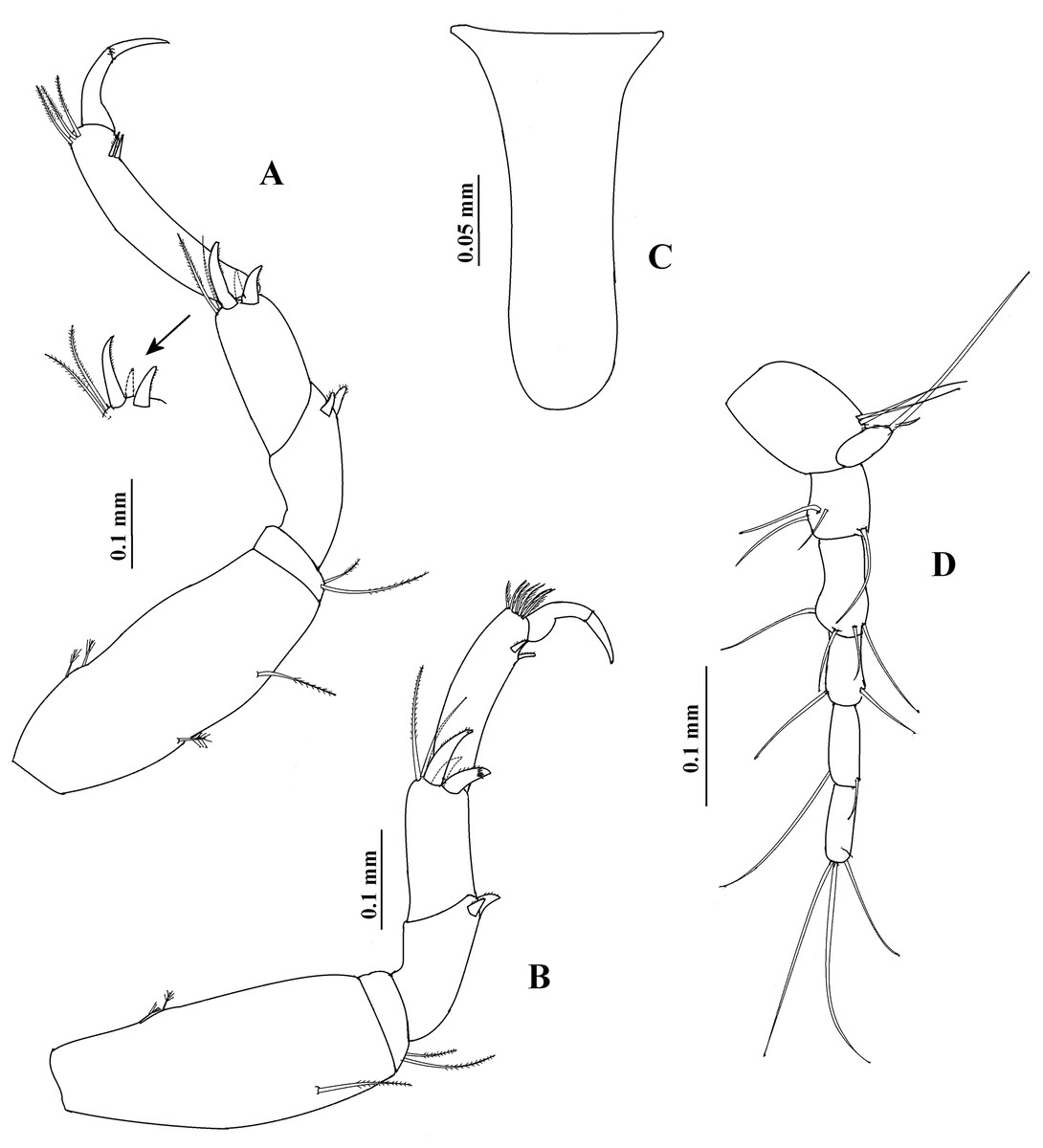
Figure 9: Hargeria chetumalensis sp. nov. Holotype ECOSUR0209, adult male, 2.9 mm.
(A) Pereopod 5. (B) Pereopod 6. (C) Anal plate. (D) Uropod.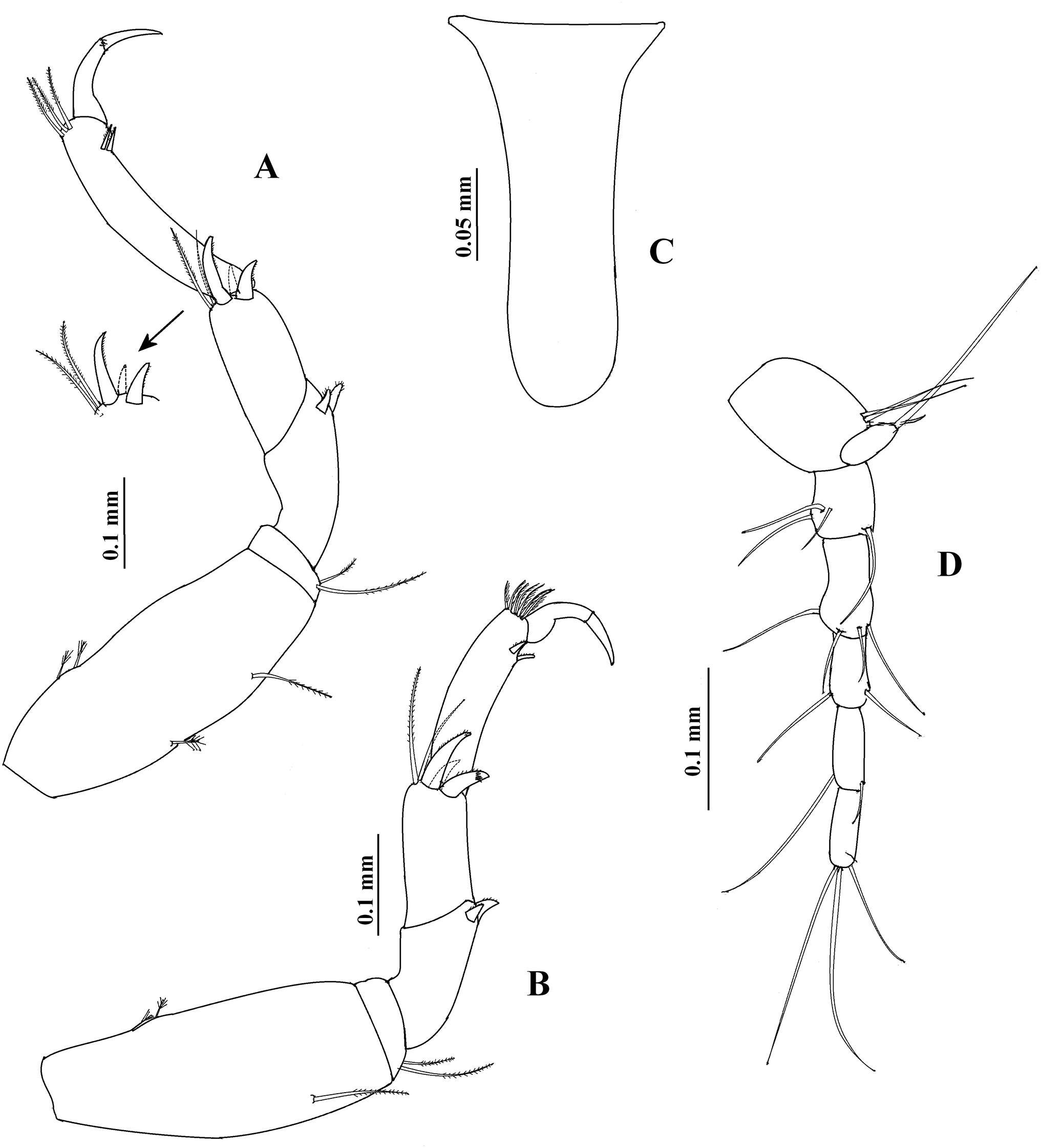{kind=link}
Pereopod 6 (Fig. 9B) similar to pereopods 4 and 5, but basis 2.3 as long as broad. Merus as long as broad. Propodus 3.9 times as long as broad, with seven short dorsodistal serrate setae.
Uropod (Fig. 9D) with three dorsal setae on peduncle. Exopod uniarticulate, 0.8 times as long as first article of uropodal endopod, with three distal setae (two missing). Endopod with five articles; first, fourth, and fifth article of similar length; second article longer than other articles; with four distal setae; third article shorter than other articles; last article with three distal setae.
Non-ovigerous female. Paratype ECOSUR0210, 3.1 mm (Fig. 10A). Body 4.9 times as long as broad. Cephalothorax oval, 1.2 times as long as broad, as long as first three pereonites combined. Pereon about twice as long as cephalothorax; pereonites 4 and 6 subequal in length; pereonite 5 shorter than other pereonites. Pleon less than 0.2 times the total body length. Pleotelson about 0.3 times as long as pleon; posterior end with eight setae, four of them small (Fig. 12B).
Figure 10: Hargeria chetumalensis sp. nov. Paratype ECOSUR0210, non-ovigerous female, 3.1 mm.
(A) Habitus. (B) Antennule. (C) Antenna. (D) Left mandible. (E) Right mandible. (F) Maxillule. (G) Maxilliped. (H) Maxillipedal endite. (I) Cheliped. (J) Details of propodius and dactylus.{kind=link}
Antennule (Fig. 10B) with four articles. First article 2.7 times as long as broad, with three setae and three plumose sensory setae on distal half. Second article 0.4 times as long as first article, 1.9 times as long as broad, with two distal setae. Third article 0.4 times as long as first article, 2.5 as long as broad. Fourth article minute, with three distal setae and one aesthetasc.
Antenna (Fig. 10C). First article as long as broad, naked. Second article 1.4 times as long as broad, with two distal spines and small distolateral seta. Third article 1.2 times as long as broad, with one distal spine. Fourth article 4.7 times as long as broad, with three simple setae and one plumose sensory seta. Fifth article 0.5 times as long as fourth article, 4.3 times as long as broad, with two distal setae and one plumose sensory seta. Sixth article minute, with six distal setae.
Mouthparts. Left mandible, ventral view, (Fig. 10D) with incisor shorter than lacinia mobilis, with four small processes; lacinia mobilis with four small processes and bifurcate apex; molar cylindrical, thick, with triturative ridges. Right mandible (Fig. 10E) with incisor crenulate, with slightly bifurcate apex; molar with triturative ridges. Labium (not illustrated) distally setose, with scales. Maxillule (Fig. 10F) setose, with nine spines. Right maxilliped (Fig. 10G) with five serrate setae on basis; first palp article 1.6 times as long as broad, naked; second palp article twice as long as broad, with one dorsodistal serrate seta, four ventrodistal setae and small ventrolateral setae; third palp article 3.2 times as long as broad, with eight ventrolateral serrate setae; fourth palp article 2.4 times as long as broad, with nine ventrolateral serrate setae and scales; endite (Fig. 10H) with three flat spines (two of them rounded terminally, the subdistal smaller, pointed), one dorsodistal seta, coupling hooks and scales.
Cheliped (Figs. 10I–10J) 0.3 times as long as total body length. Basis 1.3 times as long as broad, with one dorsol seta. Merus longer than broad, with three ventral setae. Carpus twice as long as broad, with three ventrodistal setae and two dorsal setae. Propodus with seta near dactylus articulation, comb-row with four setae (not illustrated). Fixed finger with three weak processes on cutting edge; proximal small; with eight setae, five of them situated on ventral side. Dactylus with proximodorsal seta and three lamellae on ventral edge.
Pereopod 1 (Fig. 11A) with basis 4.3 times as long as broad, with one proximal serrate seta. Ischium with one serrate seta. Merus 1.3 times as long as carpus, 2.2 times as long as broad, with one small ventrodistal serrate seta. Carpus 2.4 times as long as broad, with six distal serrate setae. Propodus 3.8 times as long as broad, with distal serrate setae (three dorsal and one ventral). Dactylus and unguis combined as long as propodus; dactylus with one proximal serrate seta.
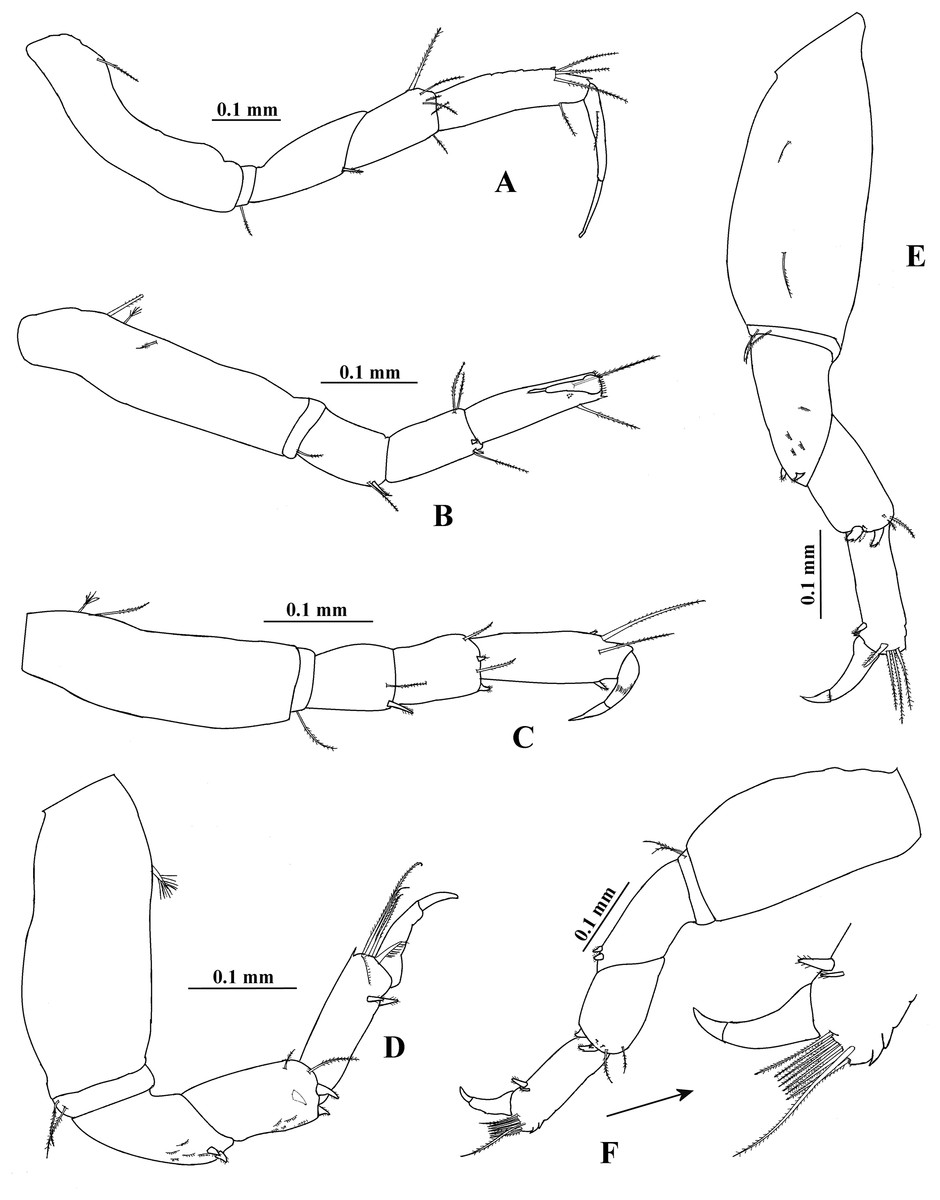
Figure 11: Hargeria chetumalensis sp. nov. Paratype ECOSUR0210, non-ovigerous female, 3.1 mm.
(A) Pereopod 1. (B) Pereopod 2. (C) Pereopod 3. (D) Pereopod 4. (E) Pereopod 5. (F) Pereopod 6.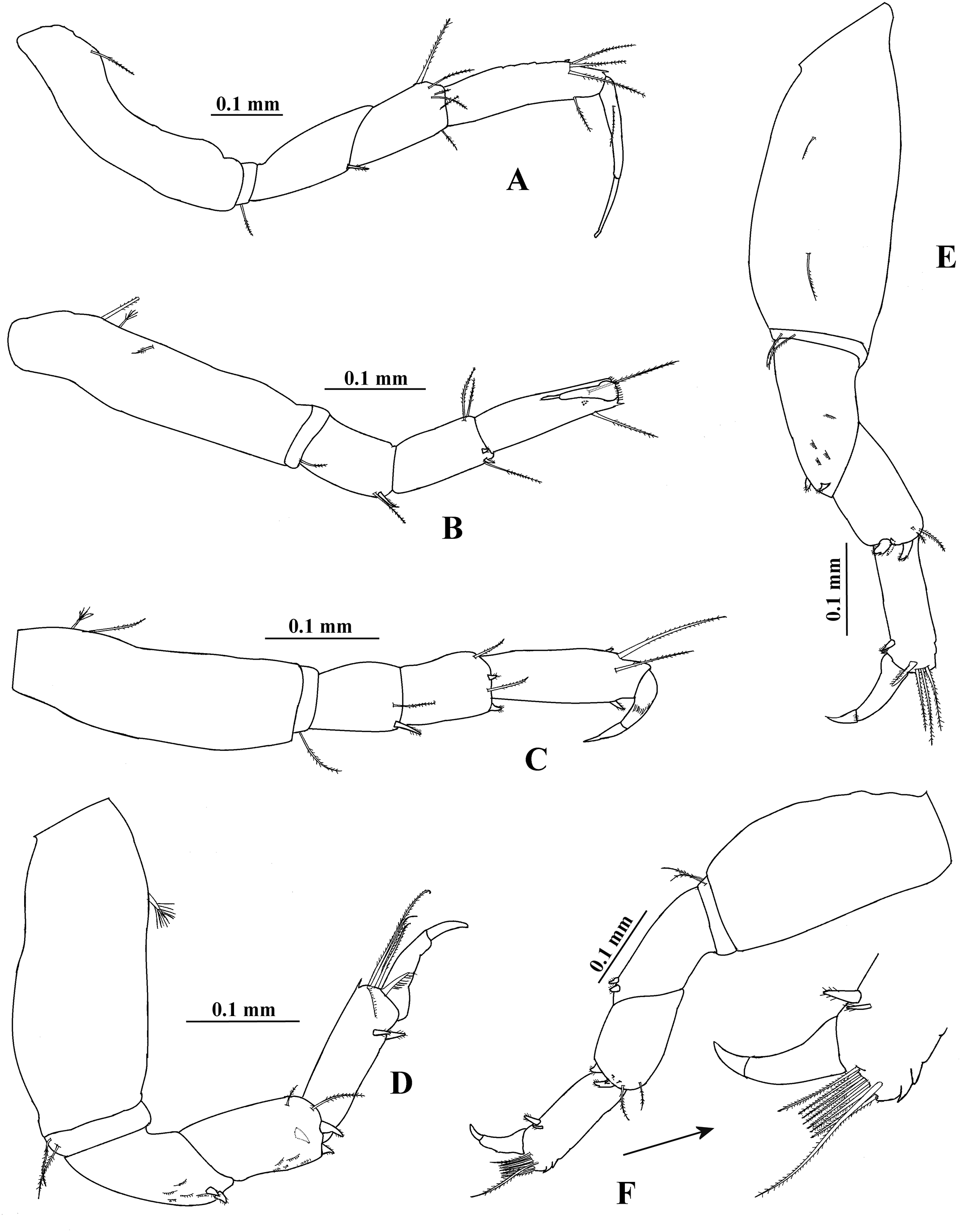{kind=link}
Pereopod 2 (Fig. 11B) with basis four times as long as broad, with one proximal serrate seta and two dorsodistal plumose sensory setae. Ischium with one serrate setae. Merus 0.9 times as long as carpus, 1.7 times as long as broad, with one ventrodistal serrate seta and one slender serrate spine. Carpus twice as long as broad, with three distal setae and two ventrodistal minute spines. Propodus 3.3 times as long as broad, with two dorsodistal serrate setae and small distoventral spine. Dactylus and unguis combined about 0.5 times as long as propodus.
Pereopod 3 (Fig. 11C) similar to pereopod 2, but basis three times as long as broad, with one dorsoproximal plumose sensory seta. Merus 1.2 times as long as broad. Carpus 1.4 times as long as broad.
Pereopod 4 (Fig. 11D) with basis 2.4 times as long as broad, with one dorsal plumose sensory seta. Ischium with two serrate setae. Merus 1.2 times as long as carpus, 2.3 times as long as broad, with two ventrodistal serrate spines and scales. Carpus twice as long as broad, with two distal serrate setae, three serrate spines and scales. Propodus 3.3 times as long as broad, with three dorsodistal serrate setae, one pinnate seta, two ventrodistal serrate spines and scales. Dactylus and unguis combined 0.7 times as long as propodus.
Pereopod 5 (Fig. 11E) similar to pereopod 4, but basis with two dorsal serrate setae. Merus 1.9 times as long as broad.
Pereopod 6 (Fig. 11F) similar to pereopods 4 and 5, but basis apparently naked. Carpus 1.7 times as long as broad. Propodus 2.8 times as long as broad, with eight dorsodistal serrate setae (one longer) and two ventrodistal serrate spines.
Pleopod 1 (Fig. 12A) with one serrate seta on peduncle. Exopod with one proximoventral circumplumose seta and 22 ventral setae. Endopod with one dorsal circumplumose outer seta, one short proximoventral circumplumose inner seta, and 13 ventral setae.
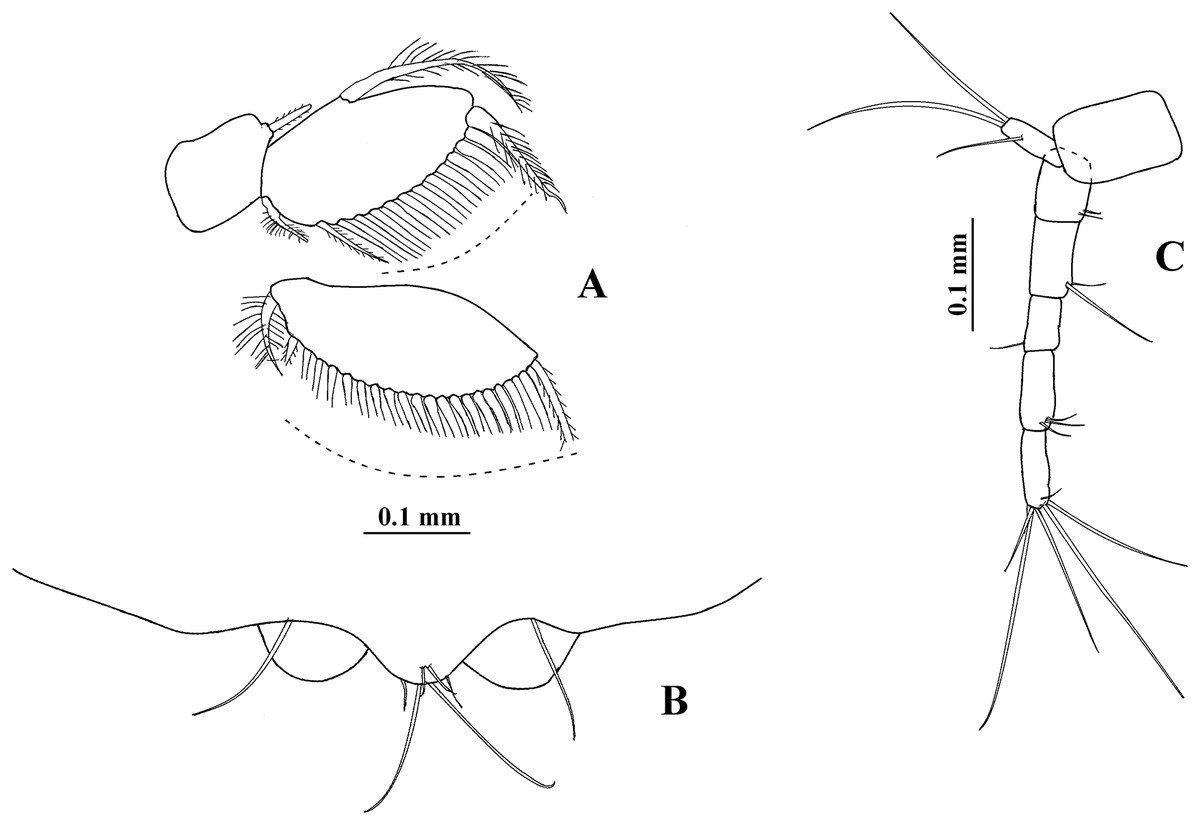
Figure 12: Hargeria chetumalensis sp. nov. Paratype ECOSUR0210, non-ovigerous female, 3.1 mm.
(A) Pleopod. (B) Posterior end of pleotelson. (C) Uropod.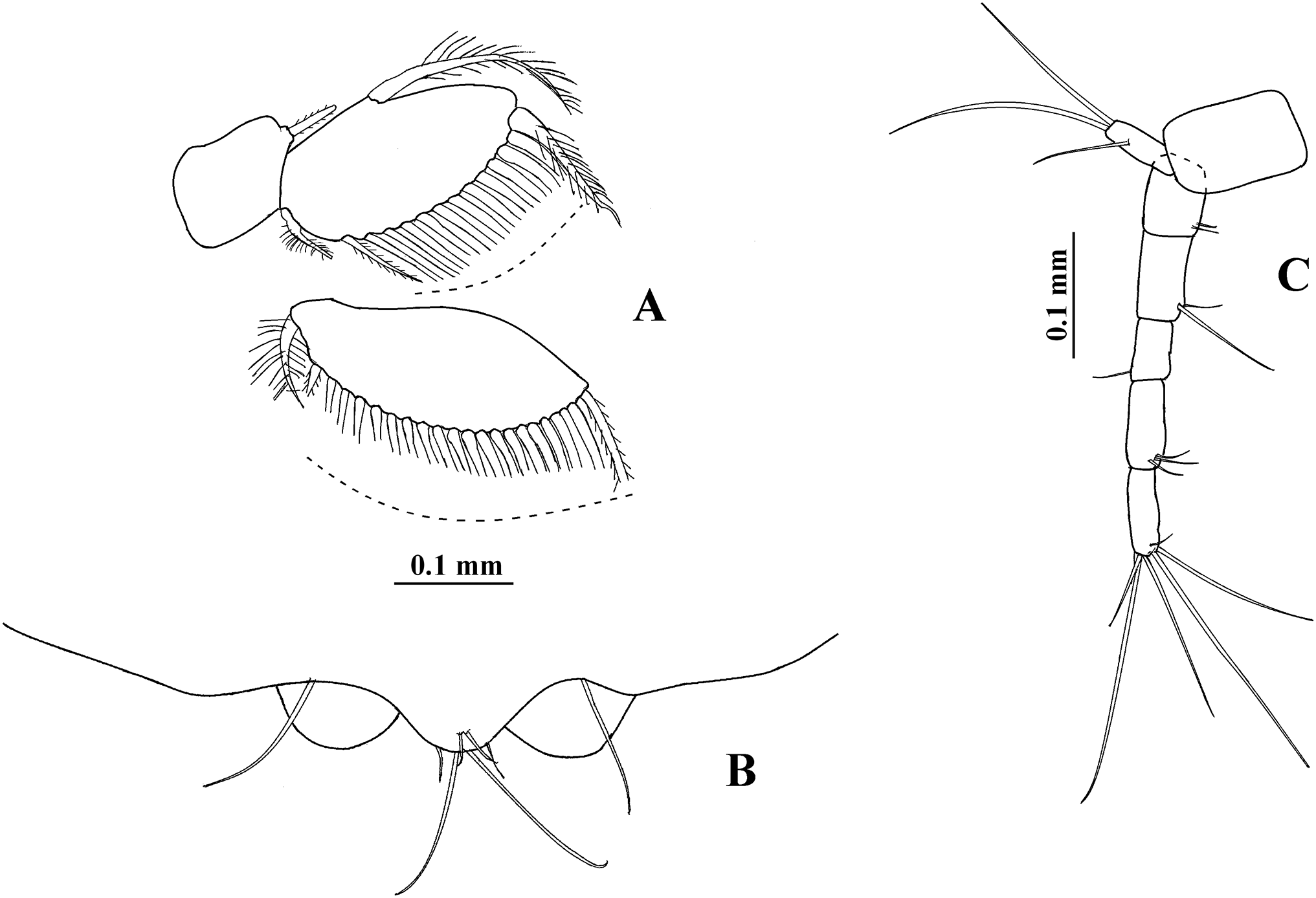{kind=link}
Uropod (Fig. 12C) with naked peduncle. Exopod uniarticulate, 0.9 as long as first endopodal article, with three setae. Endopod with five articles; first article shorter than second article, with two distal setae; third article shorter than other articles, with one distal seta; fourth and fifth articles of subequal length, with three and six distal setae, respectively.
Type locality. Raudales, Guerrero Lagoon, Quintana Roo, Mexico.
Distribution. Chetumal Bay and Guerrero Lagoon Bay System, Quintana Roo, Mexico.
Habitat. Shallow-water in rocks with sediment and alga Batophora sp.
Remarks. Comparing males of H. chetumalensis sp. nov. with H. rapax, interspecific differences can be seen in the rostrum, chelipeds, antennules, antennae, mandibles, pereopods (1, 2, 4, 6) and the anal plate. For example, males of H. chetumalensis have a pointed, narrow rostrum that does not extend beyond the eyes; in contrast, H. rapax has a convex and broad rostrum that does extend beyond eyes. H. chetumalensis has the first peduncle article of the antennule more than eight times as long as broad, while in H. rapax it is up to seven times as long as broad; H. chetumalensis has the carpus of cheliped about eight times as long as broad, whereas in H. rapax it is five to seven times as long as broad. Also, in H. chetumalensis the two long distal setae on merus of pereopod 4 exceed the median length of propodus, while in H. rapax such setae do not. Furthermore, H. chetumalensis has a tongue-shaped anal plate, which is more than three times as long as broad, whereas H. rapax has a capitate anal plate that is less than three times as long as broad (other differences see Table 2).
| H. rapax | H. chetumalensis sp. nov. | |
|---|---|---|
| Total length (mm) | 2.3–2.6 | 1.9–2.9 |
| Rostrum length vs eyes | Exceeding | Not exceeding |
| Fourth article, antenna | Up to 4.7 times as long as broad | Up to 6.6 times as long as broad |
| Chelipedal carpus | Five to seven times as long as broad | About eight times as long as broad |
| Proximal process, fixed finger of cheliped | Pointed | Rounded |
| No. distal setae on merus, pereopod 1 | Two simple | One serrate |
| No. distal setae on carpus, pereopod 1 | Six simple | Three serrate |
| Propodus, pereopod 1 | Up to 3.9 times as long as broad | Up to 5.6 times as long as broad |
| No. setae on merus, pereopods 2 and 3 | One simple | 0 |
| No. spines on merus, pereopods 2 and 3 | One simple | One serrate |
| No. setae on propodus, pereopod 6 | Eight simple | Seven serrate |
| Shape of anal plate | Capitate | Tongue-shaped |
Regarding females, H. chetumalensis is stouter, the body is 4.9 times as long as broad, unlike H. rapax where the body is 6.2 times as long as broad; H. chetumalensis has the first three pereonites of similar length, whereas in H. rapax only the second and third pereonite are similar. Also, H. chetumalensis has three lamellae on the cutting edge of the cheliped dactylus, while H. rapax has eight. Furthermore, H. chetumalensis has the third maxillipedal article 3.2 times as long as broad, whereas in H. rapax it is 2.1 times as long as broad (other differences see Table 3).
| H. rapax | H. chetumalensis sp. nov. | |
|---|---|---|
| Total length (mm) | 2.9–3.7 | 3.1 |
| First antennular article | Up to 3.7 times as long as broad | Up to 2.7 times as long as broad |
| Fourth antennal article | Up to 3.3 times as long as broad | Up to 4.7 times as long as broad |
| Fifth antennal article | Up to 2.2 times as long as broad | Up to 4.3 times as long as broad |
| No. small processes on incisor, left mandible | 3 | 4 |
| No. inner lamellae on chelipedal dactylus | 8 | 3 |
| No. distal setae on merus, pereopod 1 | Two to three simple | One serrate |
| No. setae on merus, peropod 2 and 3 | Two simple | One serrate |
| No. spines on merus, peropods 2 and 3 | 0 | One serrate |
Molecular analyses
The morphological differences found between H. rapax and H. chetumalensis, and also between Hargeria, Chondrochelia, and Leptochelia genera are supported by the molecular analyses based on the COI (Fig. 15). Our results show a genetic divergence of 14.7% between H. rapax and H. chetumalensis, which according to Costa et al. (2007) and Montiel-Martínez et al. (2008), exceeds the lowest value (4.9%) of interspecific divergence found in crustaceans. Also, the intraspecific divergence among the populations of H. rapax along the coast of the USA was 1.2% (n = 5), while in Mexico (for H. chetumalensis) it was 0.2% (n = 21). It is probable that the larger genetic divergence between the populations of Florida and Maryland is due to greater geographic distance between them than those found in the Chetumal Bay and Guerrero Lagoon System.
On the other hand, with regards of genetic divergence between Hargeria species and species of the other genera analyzed, both species of Hargeria have a genetic divergence higher to 40% concerning C. dubia and 35–38% with Leptochelia species. These values are higher than those considered for intrageneric divergence on Crustacea (19–32%) (Costa et al., 2007; Matzen Da Silva et al., 2011). Our molecular and morphological data support Guţu’s proposal to recognize Hargeria as a valid genus.
Morphological analyses of setal and cuticular structures
The SEM analysis showed that the cuticle of all structures, as well as setae of pereopods and mouthparts are more ornate than they can appear using the optical microscope, where they may appear to be simple; however, they comprise scales, pores, and small setae, giving the “serrated” appearance on such features. For example, Fig. 13 shows the details of pores, scales, and serrated setae on the propodus of pereopods 1, 2, and 6 (male, 2.4 mm); the processes and cutting edge of the cheliped fixed finger (male, 1.9 mm); as well as the pleotelson and anal plate (male, three mm) of H. chetumalensis. With regards to female and manca, Fig. 14 shows the seta, spiniform setae and scales of antennules and antennae (manca, 0.9 mm, Fig. 14A; non-ovigerous female, 2.7 mm, Fig. 14D); pereopods (manca, Figs. 14B and 14C), and mouthparts (non-ovigerous female, Figs. 14E–14G). It is important to note that these features are present in all life stages, from manca to adult.
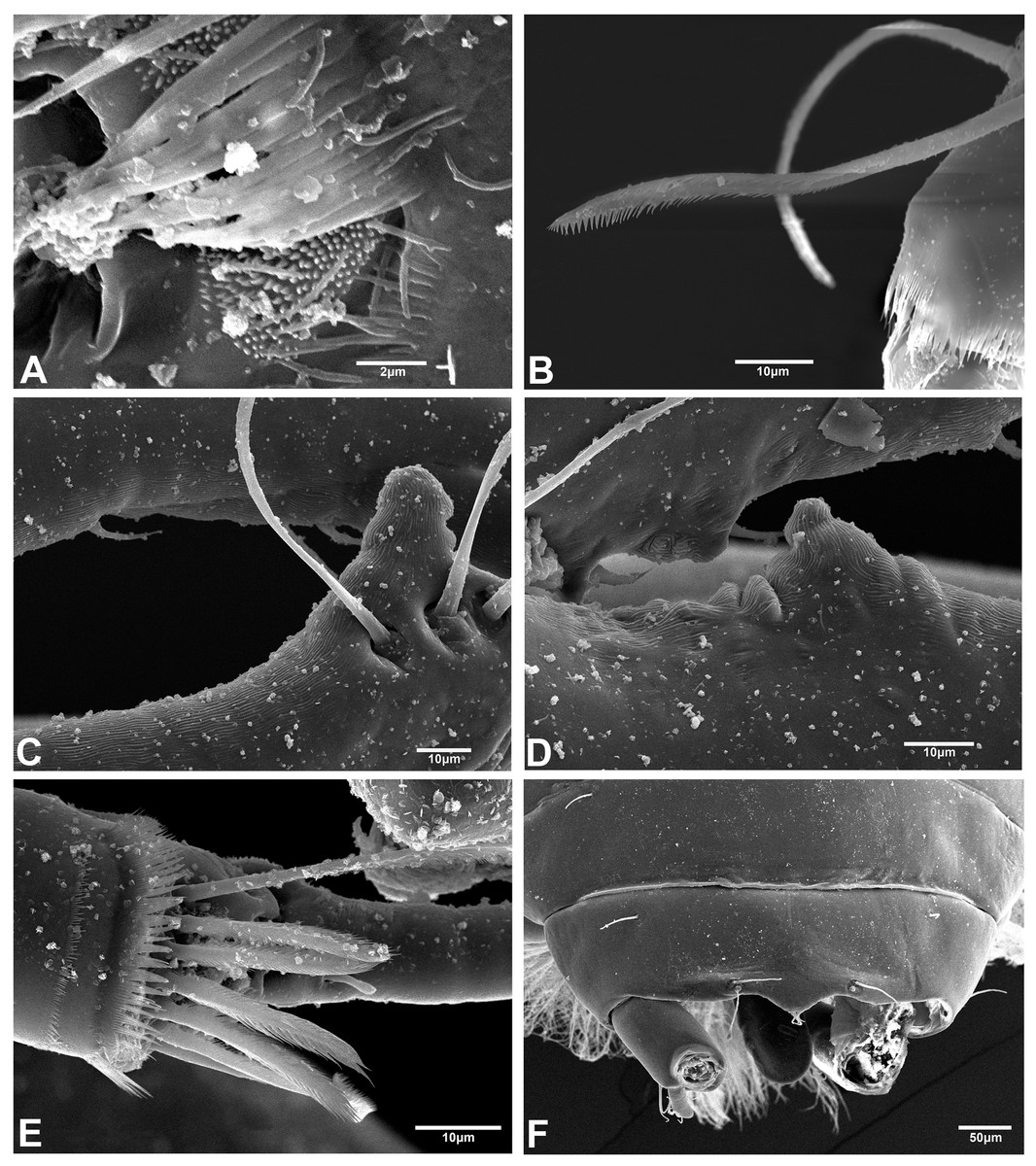
Figure 13: Hargeria chetumalensis sp. nov. males from Raudales, Quintana, Roo. SEM images.
Males, 2.4 mm, ECOSUR. (A) Pores and scales on propodus, pereopod 1. (B) Serrate setae with terminal pore, pereopod 2. Male, 1.9 mm, ECOSUR. (C) Distal chelipedal process. (D) Proximal chelipedal processes. (E) Serrate setae on propodus, pereopod 6. Male, three mm. ECOSUR. (F) Pleotelson, uropods broken.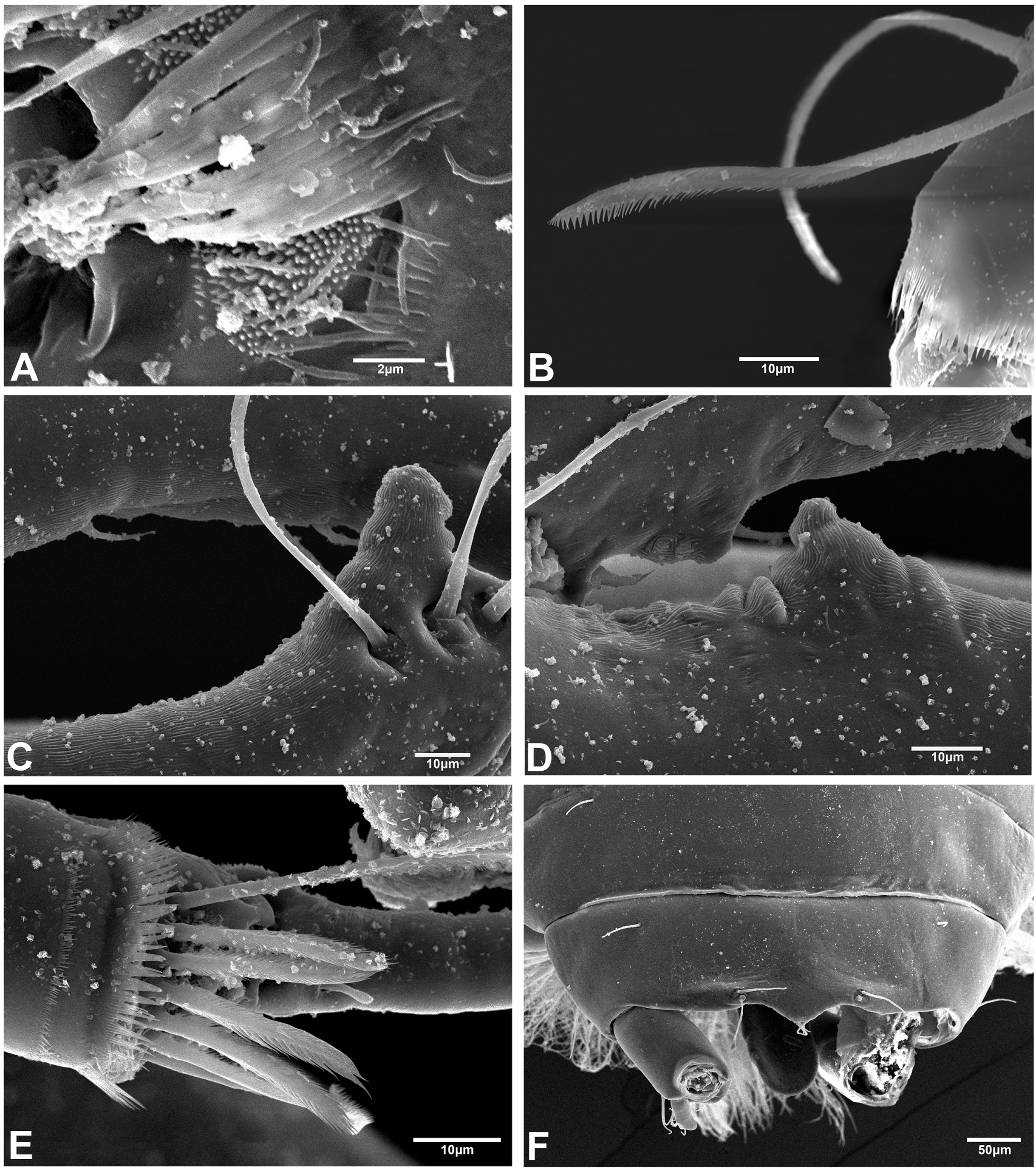{kind=link}

Figure 14: Hargeria chetumalensis sp. nov. manca and female from Raudales, Quintana, Roo. SEM images.
Manca, 0.9 mm, ECOSUR. (A) Antennules and antennae, lateral view. (B) Serrate setae, scales and pores on propodus, pereopod 2. (C) Serrate and plumose sensory setae, spinulate scales and pores on propodus, pereopod 5. Non-ovigerous female, 2.7 mm, ECOSUR. (D) Serrate spiniform seta on second antennal article. (E) Scales and serrate setae on labium. (F) Serrate setae and plumose sensory setae in maxillule. (G) Scales and serrate setae on maxillipedal fourth article.{kind=link}
It is the first time that this type of ornamentation of the cuticle and setae is reported for species belonging to the Leptocheliidae. Therefore, we consider that a more detailed examination of these structures will allow us a better characterization of all structures. With this kind of information, we will have the robustness to clarify the morphological differences of the supposed species complexes; furthermore, we will obtain more information for phylogenetic studies.
Discussion
The use of the integrative taxonomy in tanaids, according to Larsen (2001), Bamber (2010), Drumm (2010), and Larsen & Froufe (2010, 2013), is decisive in demonstrating the differences among cryptic species; i.e., discriminating new species or reinstating those species considered as junior synonyms. Furthermore, according to Błażewicz-Paszkowycz et al. (2014) the use of molecular technics is advantageous to link the different stages of life in tanaidaceans. When we analyzed the genetic sequences of H. chetumalensis sp. nov. and H. rapax, the data allowed us to confirm that the morphological features used for the recognition of both species were adequate to link males, females, and juvenile stages, although these species have a strong polymorphism (Fig. 15).
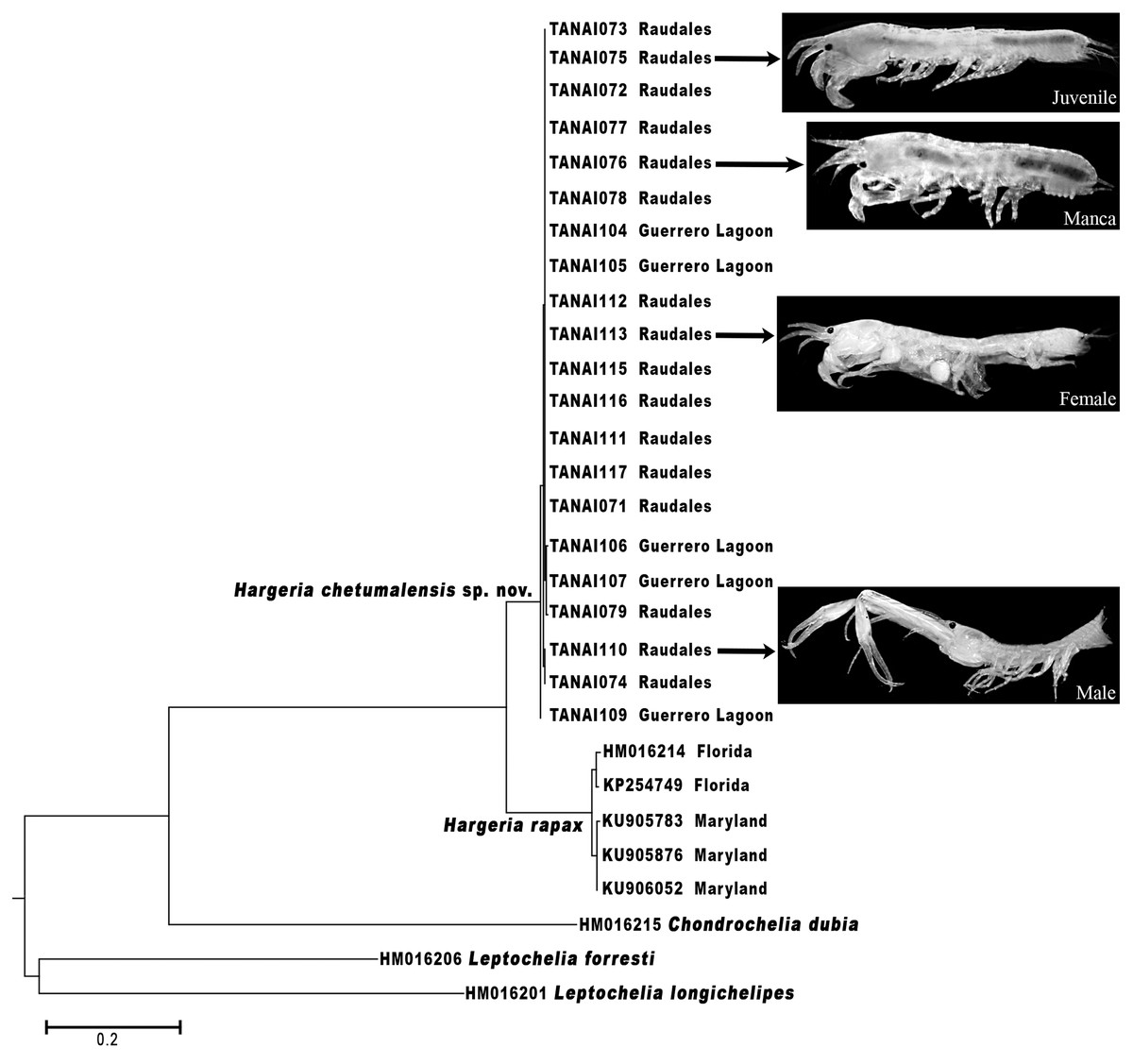
Figure 15: Maximum likelihood tree of COI sequences.
Using Hasegawa–Kishino–Yano with a discrete Gamma distribution with five rate categories and by assuming that a certain fraction of sites is evolutionarily invariable (HKY+G+I).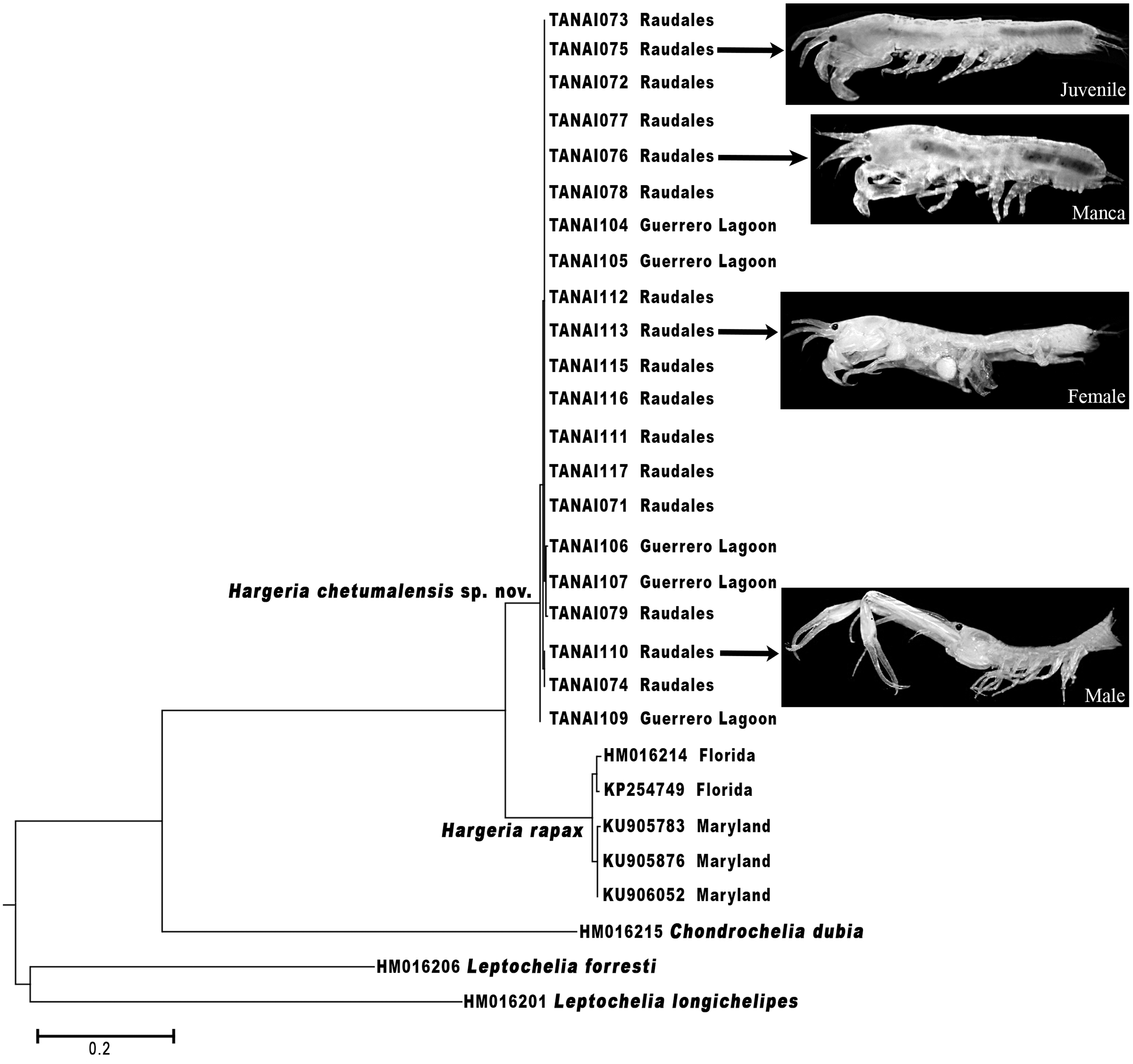{kind=link}
Besides, the use of the morphological and molecular information is advantageous to determine, with some reliability, the geographic range of the species. On the evidence provided in this study, H. rapax is restricted to the east coast of the USA, while H. chetumalensis is only known in the eastern part of the Chetumal Bay and Guerrero Lagoon, Mexico.
As we noted above, H. rapax constitute a cryptic species along the Northwestern American Atlantic. Due to the ecological conditions (e.g., ph, oxygen, and salinity concentrations, current velocity) along the extensive coastline varies considerably, it would be unlikely that an estuarine species could have a wide distribution. A detailed revision of the other specimens reported in this region as H. rapax need to be revaluated to corroborate this idea; however, it is beyond the scope of this study.
Conclusions
The taxonomy of tanaidaceans represents a great challenge, since their small size (one to three mm on average), high intraspecific polymorphisms, and the lack of specialized regional literature have increased the degree of complexity for the recognition of genera and species. In recent years, the use of morphology, genetic, and morphometric data have allowed to detect and solve some taxonomic problems related to cryptic species of leptocheliids. Herein, based on morphological and molecular data, we give new evidence to support the recognition of Hargeria as a valid genus, different from Chondrochelia and Leptochelia. Besides, we redescribed H. rapax based upon type and topotype materials; also, we described the second species belonging to Hargeria, which is the first tanaidacean described from the Mexican Caribbean. We consider that the information provided in this work will help as the baseline to clarify the taxonomic status of the specimens that have been reported under the name of H. rapax within the wide geographical distribution that was attributed to the species.