
Multilocus molecular systematics of the circumtropical reef-fish genus Abudefduf (Pomacentridae): history, geography and ecology of speciation
- Published
- Accepted
- Received
- Academic Editor
- Tomas Hrbek
- Subject Areas
- Biodiversity, Biogeography, Zoology
- Keywords
- Abudefdufinae, Sergeant-majors, Isthmus of Panama, Vicariance, Feeding mode, Phylogenetics
- Copyright
- © 2018 Campbell et al.
- Licence
- This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, reproduction and adaptation in any medium and for any purpose provided that it is properly attributed. For attribution, the original author(s), title, publication source (PeerJ) and either DOI or URL of the article must be cited.
- Cite this article
- 2018. Multilocus molecular systematics of the circumtropical reef-fish genus Abudefduf (Pomacentridae): history, geography and ecology of speciation. PeerJ 6:e5357 https://doi.org/10.7717/peerj.5357
Abstract
We investigated a pantropical sub-family and genus of damselfishes, the sergeant-majors (Pomacentridae: Abudefdufinae: Abudefduf), to identify the tempo and mechanisms of speciation in the lineage. We examined sequence capture data from 500 loci and 20 species, with multiple individuals sampled from across the geographic ranges of widespread species. Utilizing a maximum likelihood framework, as well as a time-calibrated Bayesian phylogeny, the following key questions are addressed: What is the historical tempo of speciation? What are the relative contributions of vicariant, peripatric and parapatric speciation to sergeant-major diversity? How is speciation related to major variation in trophic ecology? The approximately 20 species of sergeant-majors fall into three main lineages. The ancestral condition appears to be benthivory, which is predominant in two lineages comprising six species. The remaining species of sergeant-majors, of which there are at least 15, fall within a clade composed entirely of planktivores. This clade is sister to a benthivore clade that included one species, Abudefduf notatus, in transition to planktivory. Most speciation of sergeant-majors, which appeared ∼24 million years ago, occurred in the last 10 million years. Present distributional patterns indicate vicariant speciation precipitated by the closure of land barriers between both sides of the Atlantic and the Pacific, and the emergence of land between the Indian and Pacific Oceans. Within this backdrop, frequent oscillations in sea level over the last 10 million years also appear to have generated conditions suitable for both peripatric and vicariant speciation, and most speciation within the genus appears linked to these changes in sea level. Diversification within the genus has been concentrated in planktivorous seargeant-majors rather than benthivores. The root cause is unclear, but does not appear to be related to differences in dispersal potential, which is greater in the planktivorous species, due to the ability of their post-larval juveniles to raft with floating debris. This elevated speciation rate in planktivores and their propensity to form local endemics may reflect relaxation of selective pressures (e.g., on crypticity) that limit speciation in benthivorous sergeant-majors. Finally, our data allow us to clarify relationships of geminate sergeant-major species, indicating that there are subdivisions within the Atlantic for both benthivore and planktivore geminate pairs that may have misled previous studies.
Introduction
Sergeant-majors (Pomacentridae: Abudefdufinae: Abudefduf) are a pantropically distributed genus and subfamily of damselfishes that represent typical members of the fish faunas of coral and rocky reefs in all tropical regions (Cooper, Smith & Westneat, 2009). Although the genus has been subject to much recent phylogenetic study, those analyses used few genetic loci and did not include all known species within the genus. As a consequence, uncertainty persists regarding interspecific relationships and how many species of Abudefduf exist (Bertrand, Borsa & Chen, 2017; Cooper, Smith & Westneat, 2009; Cooper et al., 2014; Wibowo, Toda & Motomura, 2017).
An early and long-standing hypothesis was that the genus contained two clades with worldwide distributions divided along ecological lines, the Abudefduf saxatilis clade and the Abudefduf sordidus clade (Hensley, 1978). The A. saxatilis clade of Hensley (1978) comprised planktivorous species while the A. sordidus clade was made up of benthivorous species. Recent genetic studies indicate that there are in fact three broadly pantropical clades within Abudefduf, (1) an A. saxatilis clade, (2) an A. sordidus clade, and, (3) and another clade of benthivores, the Abudefduf taurus clade (Aguilar-Medrano & Barber, 2016; Frédérich et al., 2013). Divisions within the genus along ecological lines revealed in those genetic analyses are broadly in line with Hensley (1978). However, two clades with different feeding ecologies are most closely related to each other, the A. saxatilis and A. sordidus clades. Members of the A. saxatilis clade have smaller body sizes, live in (often large) aggregations and feed above the bottom in the water column on zooplankton (Aguilar-Medrano & Barber, 2016; Emery, 1973; Randall, 1967) (See also Range Maps 1–3). The pantropical A. saxatilis clade, which is the most species-rich clade, is most closely related to the Indo-Pacific A. sordidus clade (Range Map 4). Members of the A. sordidus and Atlantic/East Pacific A. taurus (Range Map 5) clades have thicker, deeper bodies and are benthivores that consume large amounts of benthic algae (Aguilar-Medrano & Barber, 2016; Emery, 1973; Randall, 1967). Members of the A. sordidus clade apparently also consume slightly more animal material that those in the A. taurus clade (Aguilar-Medrano & Barber, 2016).
Within both the A. saxatilis clade and the A. taurus clade, which have alternative ecologies, there are species pairs that were derived from the rise of the Isthmus of Panama. Species originating as a result of vicariance by the closure of the Isthmus of Panama were termed geminates by Jordan (1908). Although some recent studies suggest that the Isthmus formed more than 10 million years ago (MYA), the current general consensus is that the final closure occurred approximately 3 MYA (Cooper, Smith & Westneat, 2009 O’Dea et al., 2016). Geminate species often are morphologically similar and are studied across many taxa to understand molecular evolution (Marko, 2005). Consequently, correctly identifying geminate relationships has important consequences for broader evolutionary research. The two hypothesized geminate species pairs of these fish are Abudefduf concolor (East Pacific—EP) and A. taurus (Atlantic—A) (Lessios et al., 1995) in the A. taurus clade, and Abudefduf troschelii (EP) and the trans-Atlantic (TA) A. saxatilis in the A. saxatilis clade (Bermingham, McCafferty & Martin, 1997), although more recent work indicates A. troschelii is most closely related to East Atlantic (EA) species Abudefduf hoefleri (Frédérich et al., 2013), a relationship similar to that observed in Scarus (Labridae) (Choat et al., 2012).
Despite ongoing and contemporary study of the evolution of sergeant-majors, a fully representative, time-calibrated phylogenetic hypothesis that includes all described species has not yet been produced for the genus (Allen, 1991; Frédérich et al., 2013; Quenouille, Bermingham & Planes, 2004; Randall & Earle, 1999). Here, we generate the most comprehensive phylogenetic treatment to date of Abudefduf. In this study we explored sequence variation across approximately 500 conserved loci while targeting multiple individuals from each of the 19 currently recognized species. In addition, we generally sampled individuals from sites widely scattered across the geographic range of many species. From our sequencing of hundreds of loci we generated a data matrix that produces high-resolution molecular phylogenies not only through providing numerous independent samples of genetic variation from fish genomes, but also by representing many species across Abudefduf with multiple individuals. Such data produce robust molecular phylogenies, both with and without time-calibration, of Abudefduf upon which to test hypotheses about geographical, historical and ecological variation in diversification among lineages. Our results also clarify issues of geminate-species relationships and highlight the need to expand examination for cryptic diversity in the genus with further molecular study.
Methods
Sample collection
We obtained samples from the Smithsonian Tropical Research Institute (STRI) cryocollections at Naos Laboratories, Panama City, Panama and the Natural History Museum, Washington D.C., USA. Tissues from multiple sampling locations across a wide range of described species were targeted, with successfully sequenced tissues described in Table S1. Species involved in possible trans-isthmian geminate species pairs or clades were sampled more heavily (A. troschelii, A. saxatilis, A. hoefleri, A. taurus, and A. concolor) across the widest geographic range for which samples were available. Tissues from the putative geminate species-pair Chromis atrilobata and Chromis multilineata (Bermingham, McCafferty & Martin, 1997) were obtained from STRI cryocollections for further evaluation of the geminate-species concept. We also included Abudefduf luridus, which was recently reclassified as a member of the East Atlantic genus Similiparma (Cooper et al., 2014), to provide representation of an additional divergent pomacentrid lineage. Tissue samples were extracted with Qiagen DNEasy extraction kits (https://www.qiagen.com, Hilden, Germany), electrophoresed for an estimation of quality, and quantified by fluorometric quantitation with a Qubit (Thermo Fisher Scientific Inc., https://www.thermofisher.com, Waltham, MA, USA).
Sequencing
We followed the basic procedures outlined for the 500 ultraconserved element (UCE) acanthopterygian probe set by Faircloth et al. (2013). DNA was sheared on a Covaris S2 (Covaris Ltd., http://www.covaris.com, Woburn, MA, USA) to obtain average fragment sizes between 500 to 600 base pairs (bp). We filtered archived samples for the highest quality DNA; however, because many species did not have alternative tissue samples and were of low fragment size, fragmentation times were adjusted depending on initial sample quality. Illumina DNA sequencing libraries were prepared from fragmented DNA using KAPA library preparation kits (KAPA Biosystems Inc., http://www.kapabiosystems.com, Wilmington, MA, USA). Sequence capture was performed by target enrichment (Blumenstiel et al., 2010), incorporating custom adapter blockers with capture probes for 500 loci from actinopterygian fishes (MYbaits_Actinopts-UCE-0.5Kv1, MYcroarray Inc., http://www.mycroarray.com). Pre-enrichment PCR length was 12 cycles with post-enrichment PCR of 15 cycles. Eight sample libraries were sequenced on an Illumina MiSeq with paired-end (PE) 300 bp sequencing.
Generation of alignments from raw sequence data
Demultiplexed reads were cleaned using Trimmomatic 0.32 (Bolger, Lohse & Usadel, 2014) driven by the illumiprocessor wrapper script (Faircloth, 2013). A subset samples of cleaned reads were assembled for various kmers with Velvet 1.2.10 (Zerbino & Birney, 2008) to establish a range of suitable kmers for assembly. Due to the heterogeneity of the input DNA and enrichment success, we wrote a custom script to drive VelvetOptimiser 2.2.5 for kmers across overlapping sets of kmers between 95 and 185 (https://github.com/MacCampbell/scripts/driveVelvetOptimiser.pl). A final range of kmers (within the span of 95–185) based on the best assemblies indicated by VelvetOptimiser was then applied to each sample and a single optimized assembly retained for further analyses. From here, scripts from the phyluce package were employed (Faircloth et al., 2012; Faircloth, 2016). We filtered samples for overall enrichment and assembly success and aimed to retain at least two samples from each taxon. A description of the number of assembled contigs and number of UCEs detected are presented in Table S1. We performed sequence alignment with MAFFT 7.130b (Katoh et al., 2002). Four different alignments were generated. First, we included as many samples that enriched and assembled well for Abudefduf species and provided near relatives as outgroups from Similiparma luridus, C. atrilobata and C. multilineata. This “phylogenetic placement” alignment was used to identify distinct lineages, which then informed additional analyses. A second alignment, an “A. saxatilis alignment” focusing on A. saxatilis and A. hoefleri, was made to increase the number of UCE loci available for analysis to clarify relationships within and between these two species because they were unclear from the phylogenetic placement alignment. A third alignment, the “A. taurus alignment,” was generated focusing on the A. taurus and A. concolor species to increase geographic sampling without reducing the overall number of loci in the phylogenetic placement alignment. The goal of this alignment was to evaluate phylogeographic divisions within both A. taurus and A. concolor. A fourth alignment, the “time-calibrated alignment,” used a single individual from each unique lineage of Abudefduf identified, a single C. atrilobata, C. multilineata, and S. luridus, and sequences from Faircloth et al. (2013) that were informative for fossil calibration. These additional pomacentrid lineages were chosen in part by overall completeness in terms of number of loci.
Phylogenetic estimation
For analysis of the phylogenetic placement alignment, a maximum likelihood analysis was conducted in RAxML 8.2.6 (Stamatakis, 2014). We partitioned the analysis with PartitionFinder 2.0 (Lanfear et al., 2012; Lanfear et al., 2014) by specifying the General Time Reversible (GTR) model with gamma-distributed rate variation (Γ) to be evaluated across each UCE locus with the “hcluster” search method. The partition supported by a Bayesian Informative Criterion (BIC) selection method was then specified in RAxML with the GTR + Γ model of sequence evolution and 1,000 rapid bootstrap replicates. Analyses of the A. saxatilis alignment and A. taurus alignment were conducted with an identical approach to the phylogenetic placement analysis.
A time-calibrated phylogenetic tree was generated in a Bayesian framework by importing nexus alignments into BEAUTi 2 and using BEAST 2 to run the BEAUTi output (Bouckaert et al., 2014). Due to uncertainty in the number of independent lineages of A. vaigiensis, all samples from this taxon were retained for this analysis. A single partition with a GTR + Γ model of evolution and exponential relaxed clock with a Yule prior were specified. Fossil constraints are described in Table S2. Sufficient effective sample size (ESS > 200) was reached by combining 11 chains of 100 million generations with 10% burnin (990,110,000 states).
Testing of diversification rates
Differences in diversification rates (average number of species produced MY−1) within the three main Abudefduf lineages were evaluated by reducing the time-calibrated phylogeny from BEAST 2 to independent lineages of Abudefduf. Outgroup species along with repeat sampling of Abudefduf were removed for this analysis. The tree was used as input into BAMMTools (Rabosky et al., 2014) and differences in speciation/extinction rate were tested (modeltype = speciationextinction), where an expected number of shifts was set to one. A run length of 1 million generations with a 10% burnin was specified. After the run, ESSs were verified to all be sufficient (>1,000).
Results
Sampling, sequence alignment characteristics, partitioning
We obtained sequence data from all 19 species of Abudefduf that had been described when the study was initiated. Sample species, collection locality and number of successfully enriched UCE loci are described in Table S1. The phylogenetic placement alignment contains 361 UCE loci assembled in 49/59 of samples for a total alignment length of 241,893 bases, with 54,757 distinct alignment patterns and 17.93% missing data including gaps. Partitioning by PartitionFinder indicates 12 partitions (Data S1). The A. saxatilis alignment has 12 samples representing A. saxatilis from Sao Tome (n = 2), A. saxatilis from the West Atlantic (n = 5) and A. hoefleri samples from Cape Verde and Senegal (n = 3). Two A. troschelii samples for rooting are also present. Requiring a UCE locus to be present in all samples resulted in 94 UCE loci objectively partitioned into two subsets by PartitionFinder (Data S1). A total of 59,853 sites, 1,269 distinct alignment patters and 8.63% gaps or missing data characterize the A. saxatilis alignment. Focusing on A. taurus and A. concolor utilizing samples that did not enrich well and were previously excluded from the phylogenetic placement analysis results in a geographic sampling from the Galapagos Islands (n = 3), Panama (n = 2) and Costa Rica (n = 1) while including a wide sampling of A. taurus previously included in the phylogenetic placement alignment (Panama (n = 2), Venezuela (n = 2), Cape Verde (n = 2), Sao Tome (n = 2)). With the addition of the two Abudefduf declivifrons samples, 16 samples are present in the A. taurus alignment. Permitting one missing sequence per locus allows 118 UCE loci for analysis. The total number of sites in this alignment is 78,074 with 3,701 distinct alignment patterns and 11.01% gaps or missing data. The time-calibrated alignment contained 35 taxa and 22 Abudefduf samples and it is characterized by 85,838 sites, 28,506 distinct alignment patterns and 22.41% missing data.
Molecular phylogenetics of Abudefduf
Both the phylogenetic placement analysis and Bayesian timetree support the existence of three major clades of Abudefduf (Figs. 1 and 2, tree files provided in Data S1). An alternative depiction of Fig. 1 in which specific sample identifiers are appended to labels in the phylogenetic tree is presented as Fig. S1. Clade A / “taurus clade” contains A. declivifrons of the Eastern Pacific and the putative geminate species pair A. taurus and A. concolor. The monophyly of the Clade A and placement of species is well supported with bootstrap support (BS) = 100% and posterior probability (PP) = 1.00, except for A. concolor, with monophyly supported by BS=90% (PP = 1.00). Clade A is entirely composed of benthivorous species. A second well-supported clade (B / “sordidus clade”) contains three Indo-Pacific (IP) species—A. sordidus, Abudefduf septemfasciatus, and A. notatus. Support for the monophyly and placement of lineages within clade B are high (BS = 100%, PP = 1.00). Clade B contains two benthivores, and one species in transition to planktivory (A. notatus). The majority of Abudefduf species belong to the pantropical Clade C (“saxitilis clade”). Monophyly of this clade is well-supported (BS = 100%, PP = 1.00). Numerous nodes receive moderate to low support in the ML analysis throughout Clade C and a single node in the Bayesian timetree is supported by a posterior probability of <1.00 within Clade C. All Clade C species are planktivores.
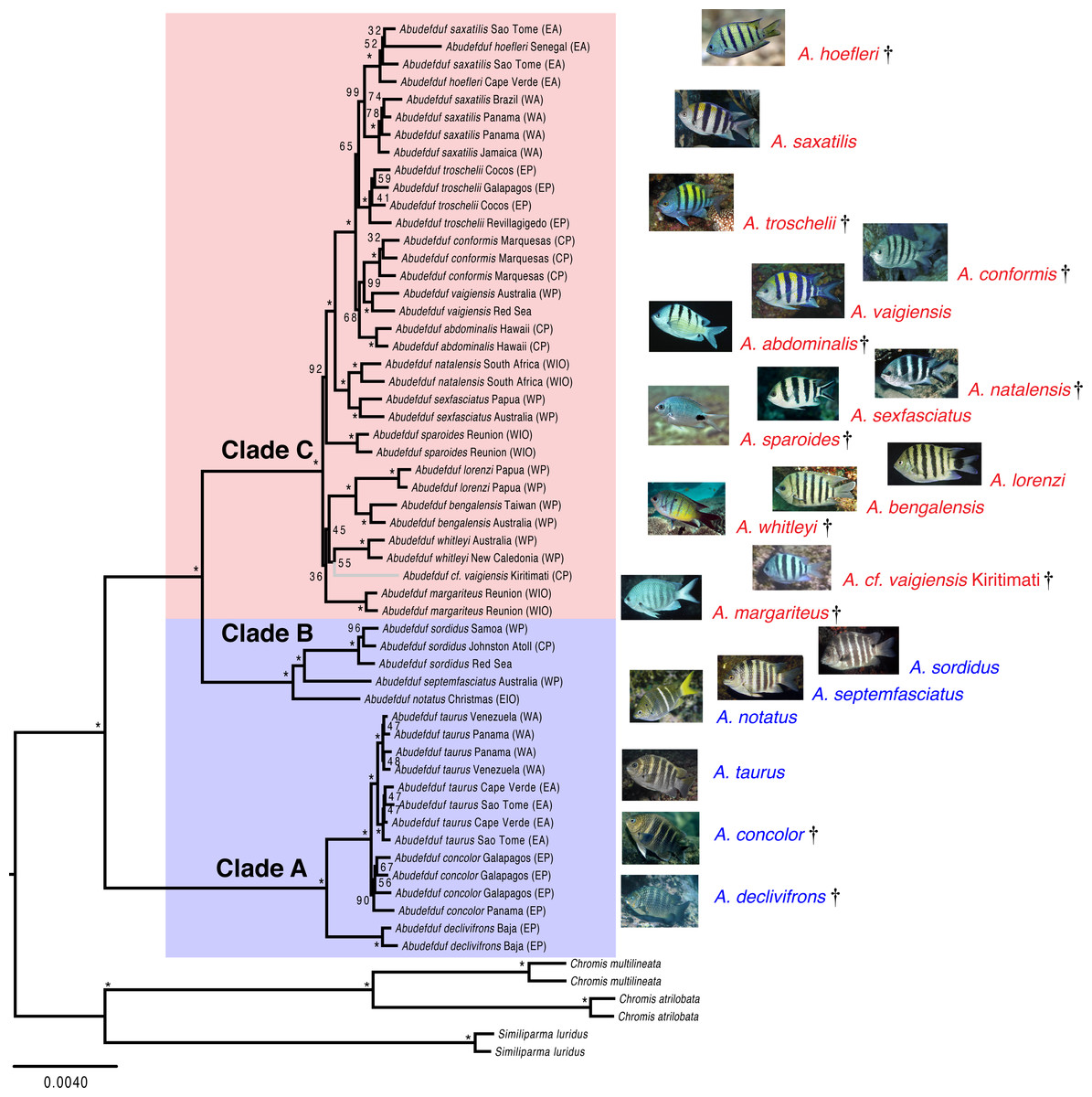
Figure 1: Maximum likelihood phylogenetic tree of Abudefduf.
A maximum likelihood phylogenetic tree of Abudefduf generated from partitioned analysis of 361 ultraconserved element (UCE) loci. An optimal partitioning strategy was implemented (Lanfear et al., 2014; Lanfear et al., 2012). Each partition was modeled under the General Time Reversible (GTR) model of sequence evolution with gamma-distributed rate variation (Γ). Bootstrap support values are indicated with an asterisk (*) if equal to 100. The three lineages of Abudefduf are indicated (Clade A, B and C). Planktivorous lineages are highlighted and named in red with benthivorous lineages highlighted and named in blue. The tree is rooted by pomacentrid outgroups of the genera Chromis and Similiparma. The branch leading to an undescribed species, A. cf. vaigiensis, is colored gray. Appended to each leaf in the tree is the approximate geographic location of the sequenced individual using these abbreviations: EA, East Atlantic; WA, West Atlantic; EP, East Pacific; CP, Central Pacific; WP, West Pacific; EIO, East Indian Ocean; and WIO, West Indian Ocean. Individual identifiers are appended to sample names in Fig. S1. Photo credits: A. hoefleri S. Floeter, A. saxatilis DRR, A. troschelii GRA, A. conformis J. Randall, A. vaigiensis GRA, A. abdominalis GRA, A. sparoides GRA, A. sexfasciatus J. Greenfield (CC BY), A. natalensis J. Randall, A. whitleyi GRA, A. bengalensis G. Edgar (CC BY), A. lorenzi GRA, A. margariteus GRA, A. cf. vaigiensis J. Earle, A. notatus GRA, A. septemfasciatus GRA, A. sordidus GRA, A. taurus DRR, A. concolor GRA, A. declivifrons GRA.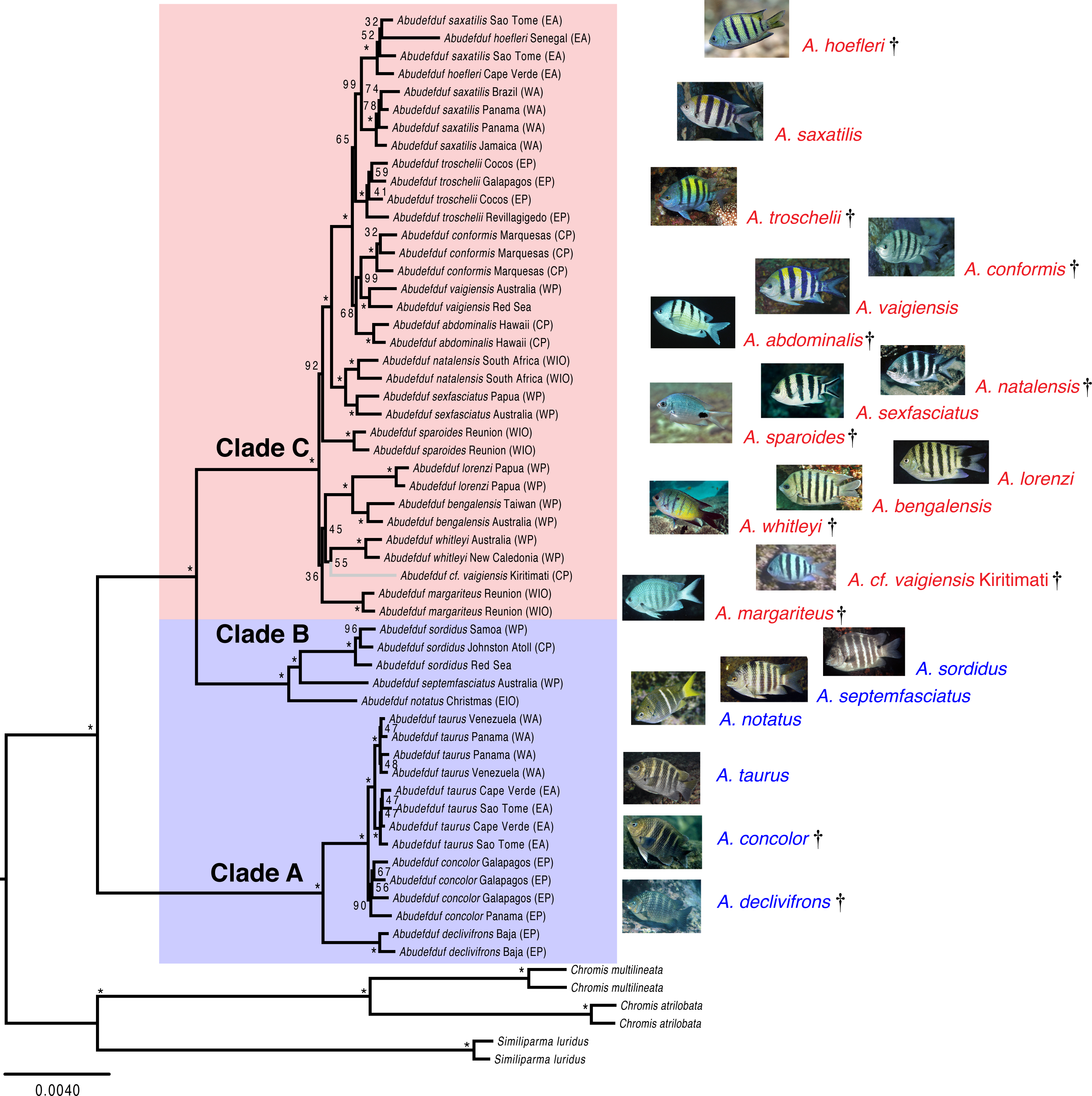{kind=link}
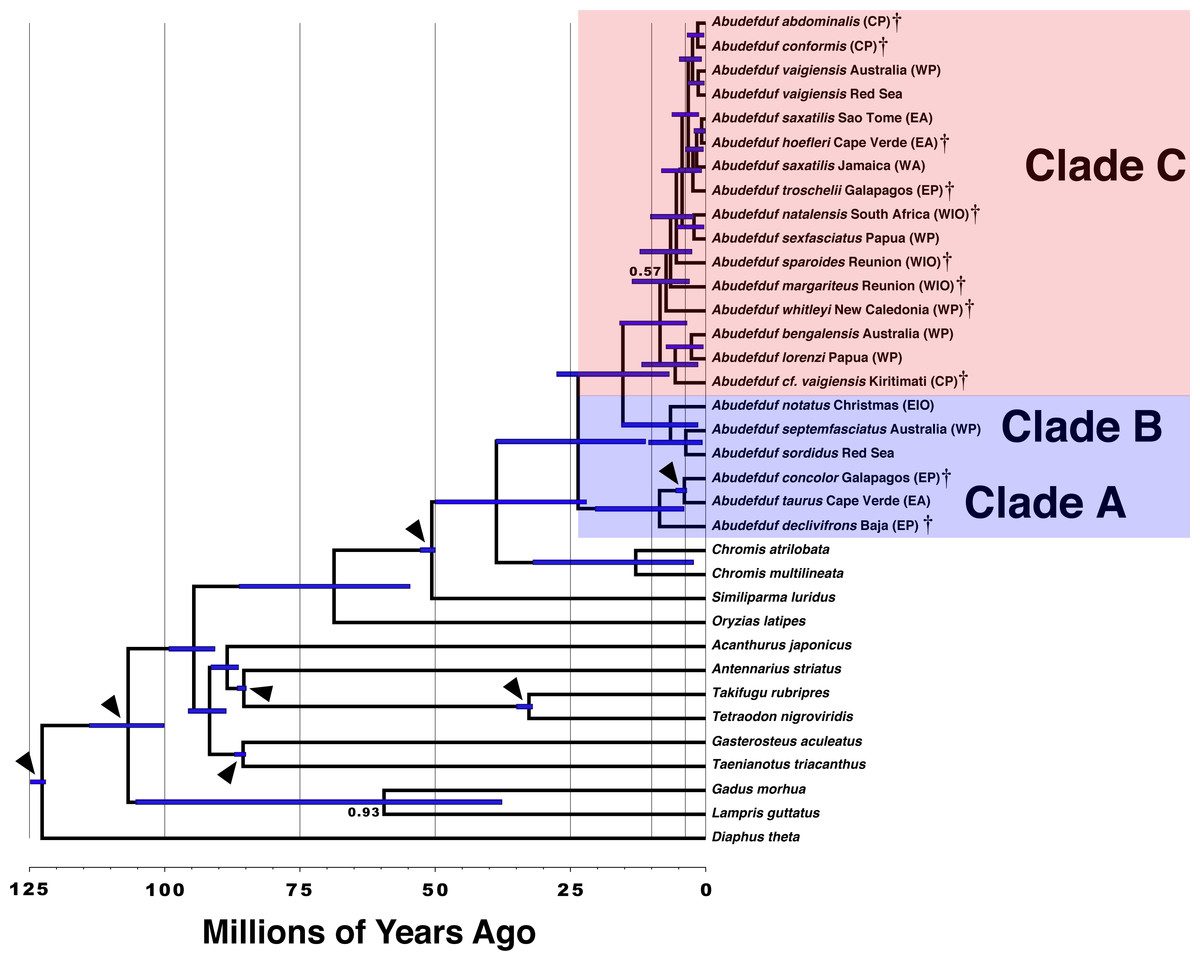
Figure 2: Time-calibrated phylogenetic tree of Abudefduf.
A time-calibrated phylogenetic tree of Abudefduf generated from 137 ultraconserved element (UCE) loci modeled under a single partition with the General Time Reversible (GTR) model with gamma-distributed rate variation (Γ). Fossil-calibrated nodes are indicated by black triangles and are described in Table S2. Posterior support at nodes is 1.00 unless otherwise indicated. Blue bars indicate 95% highest posterior density. The major trophic guilds are indicated by shading of red for planktivores and blue for benthivores. Regional endemics are indicated by a dagger (†). Invidual samples included in this analysis are indicated in Table S1. Vertical lines indicate 25 million year time divisions with 10 million years and 3 million years also indicated. Appended to each leaf in the tree is the approximate geographic location of the sequenced individual using these abbreviations: EA, East Atlantic; WA, West Atlantic; EP, East Pacific; CP, Central Pacific; WP, West Pacific; EIO, East Indian Ocean, and WIO, West Indian Ocean.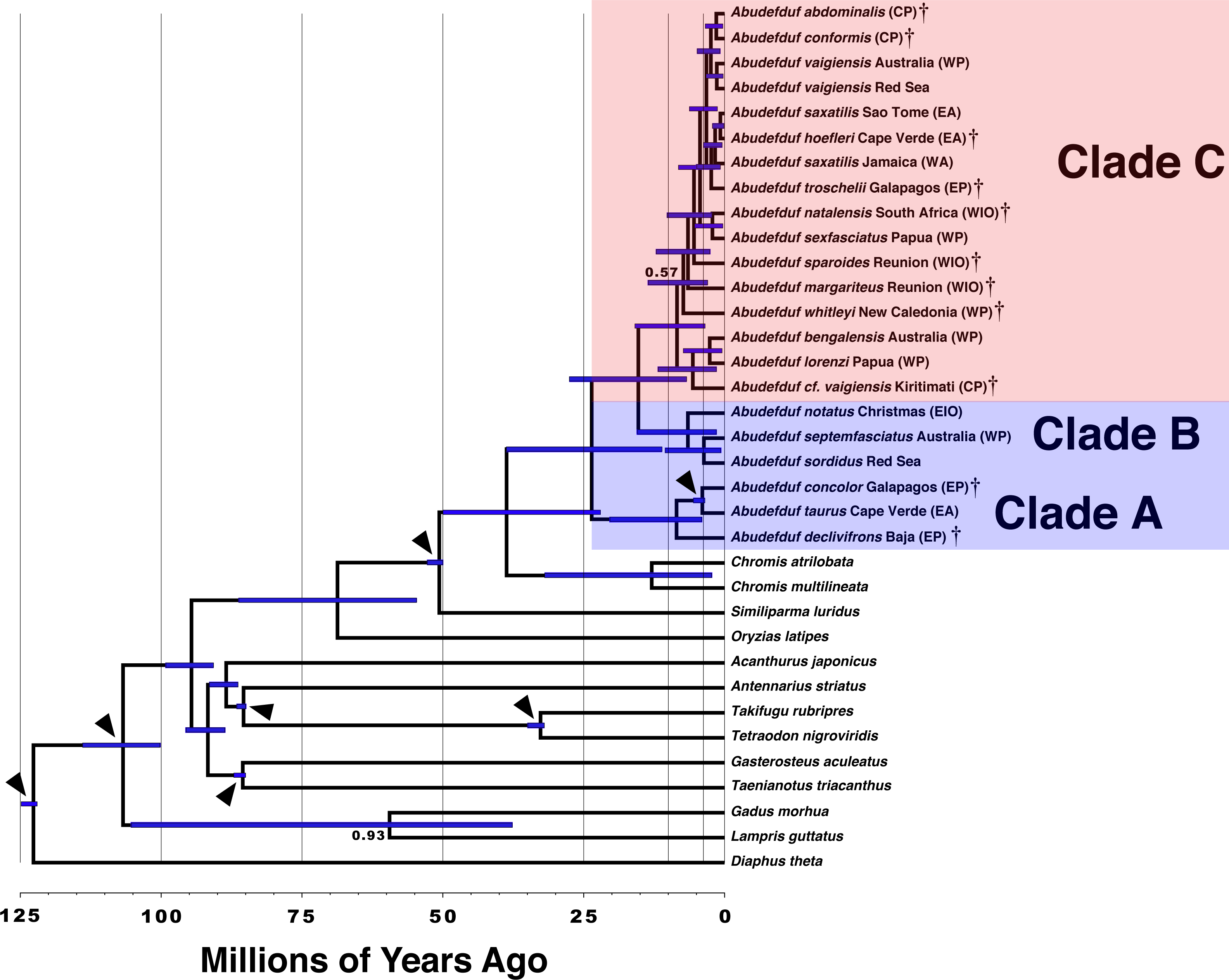{kind=link}
The phylogenetic results indicate the existence of an undescribed species of Abudefduf within Clade C: a sample labelled Abudefduf vaigiensis (STRI-x-6065) from Christmas Island (Kiritimati) in the central Pacific (Figs. 1 and 2). A. cf. vaigiensis Kiritimati is most closely related to non-A. vaigiensis samples, and distantly so to A. vaigiensis. The other samples of A. vaigiensis (Red Sea and Australia) are most closely related to each other, although sampled from a great distance apart (Figs. 1 and 2; Range Map 1). Within the phylogenetic placement analysis, the relationships among A. cf. vaigiensis, Abudefduf margariteus, Abudefduf whitleyi, and (Abudefduf bengalensis + Abudefduf lorenzi) are not clearly resolved, including low support for a sister relationship between A. whitleyi and A. cf. vaigiensis (BS = 55%). A. bengalensis and A. lorenzi are the closest relatives of A. cf. vaigiensis Kiritimati in the Bayesian analysis with high (PP = 1.00) support. Low support for early-branching nodes in Clade C (Fig. 1) prevents an unambiguous placement of A. cf. vaigiensis that is clearly supported by both the maximum likelihood and Bayesian analyses; however, there is support from both analysis frameworks for a close relationship between A. cf. vaigiensis and A. bengalensis + A. lorenzi.
Divergence time estimation
The median estimate of time to most recent common ancestor (TMRCA) of all Abudefduf species is 24 MYA (95% Highest Posterior Density (HPD), 11–39 MYA) (Fig. 2 & Fig. S2). The median TMRCA of Clade B and C is 15 MYA (95% HPD 6.7–28 MYA). Median TMRCAs for each of the three main clades are Clade A 8.5 MYA (95% HPD 4.1–20 MYA), Clade B 6.5 MYA (95% HPD 1.5–15 MYA), and Clade C 8.4 MYA (95% HPD 6.7–28 MYA). TMRCA of the geminate species-pair of A. taurus and A. concolor is 4.0 MYA (95% HPD 3.5–5.5 MYA). The unconstrained TMRCA of the geminate species clade of A. troschelii + (A. saxatilis, A. hoefleri) is estimated with a median of 2.4 MYA (95% HPD 0.75–4.9 MYA) and the TMRCA of East Atlantic and West Atlantic lineages of A. saxatilis + A. hoefleri and A. saxatilis respectively is 1. 7 MYA (95% HPD 0.46–3.8 MYA). The timetree indicates that the putative species A. cf. vaigiensis Kiritimati has been independent for approximately 5.6 million years (95% HPD 1.5–12 MYA) while the TMRCA of the Chromis geminate species pair, C. atrilobata and C. multilineata, is 13 MYA (95% HPD 2.3-32 MYA).
Diversification rates
From the 9,001 posterior samples generated by a total run length of 1,000,000 steps, the ESS of the number of shifts is 1,449.15 and the ESS of log(Likelihood) is 3,426.83. The posterior distribution of shifts in the rate of diversification indicated that there are likely no shifts. Posterior probabilities of rate shifts are zero-rate shifts, 0.90, one-rate shift 0.08 and two-rate shifts 0.01. These results indicate uniform diversification rates within the three lineages of Abudefduf under the framework of BAMMTools. However, the lack of significant differences between rates in those three lineages likely reflects low power of the test due to small sample sizes (cf. Agapow & Purvis, 2002; Kubo & Iwasa, 1995). Despite the results of the test in BAMMTOOLS, there are obvious differences in that rate, which, based on the TMRCAs shown above, is 0.12 species MY−1 in Clade A (the oldest, benthivore clade), 0.19 species MY−1 in Clade B (the younger benthivore clade), and 1.13 species MY−1 in Clade C (the equally young planktivore clade). There are no obvious differences in the tempo of diversification between planktivores and benthivores, as most speciation in both ecotypes occurred during the last 10 million years (Fig. 2).
Atlantic-East Pacific geminate lineages
The phylogenetic analyses indicate that A. concolor and A. taurus are a geminate-species pair (Figs. 1 & 2, Fig. S3, tree files provided in Data S1). As a calibration point in the timetree with a mean divergence time of 3.5 MYA, the posterior estimate of divergence time between the two species was slightly older (4.0 MYA). The widespread sampling of A. taurus, however, did reveal that there is clear geographic structuring across the Atlantic of this benthivore (Fig. 1, Fig. S3). A. taurus segregates into East Atlantic (EA: Sao Tome/Cape Verde) and West Atlantic (WA: Panama/Venezuela) clades (BS = 100%). The A. taurus alignment of 118 UCE loci (as opposed to 361 UCE loci in the phylogenetic placement alignment) supports the same division into EA and WA clades within A. taurus (BS = 100%) but does not indicate any geographic subdivisions within A. concolor in the East Pacific (EP; Fig. S3). Therefore, Clade A geminates are A. concolor and the ancestor of the EA and WA populations of A. taurus, which separated long after the closure of the Isthmus of Panama.
Our phylogenetic analyses also demonstrate that the relationships among A. troschelii, A. saxatilis, and A. hoefleri are less straightforward than previously thought. Those analyses indicate that A. saxatilis is paraphyletic and that A. hoefleri is most closely related to EA A. saxatilis (Figs. 1 and 2). Increased sampling of individuals and loci with the A. saxatilis alignment (Fig. 3, tree file provided in Data S1) produces strong support for the paraphyly of A. saxatilis with a split between WA A. saxatilis and EA A. saxatilis + A. hoefleri (BS = 100%), while A. hoefleri monophyly is only weakly supported (BS = 20%). A. saxatilis (including A. hoefleri), is most closely related to A. troschelii, and forms separate EA (Sao Tome, Cape Verde, Senegal) and WA clades (Panama, Jamaica, Brazil) (Fig. 3). Thus EA (Sao Tome) A. saxatilis is most closely related to EA A. hoefleri (Cape Verde and Senegal). Within the West Atlantic, there appears to be no geographic structuring of A. saxatilis between the Caribbean and Brazil (cf. Piñeros & Gutiérrez-Rodríguez, 2017). Divergence time estimates for the TMRCA of the geminate-clade of (A. saxatilis WA (A. saxatilis EA + A. hoefleri)) + A. troschelii are not constrained by any calibrations and are compatible with the closure of the Isthmus of Panama within the last 5 million years (2.4 MYA, 95% HPD 0.75–4.9 MYA). A late closure date for the Isthmus of Panama is widely supported, around 3 MYA (O’Dea et al., 2016). Our data indicate a transatlantic division of the common ancestor of A. saxatilis and A. hoefleri approximately 1.7 MYA (95% HPD 0.46–3.8 MYA) into WA and EA lineages. Subsequently, within the East Atlantic lineage 0.76 MYA (95% HPD 0.12-2.2 MYA) A. hoefleri split from EA A. saxatilis.
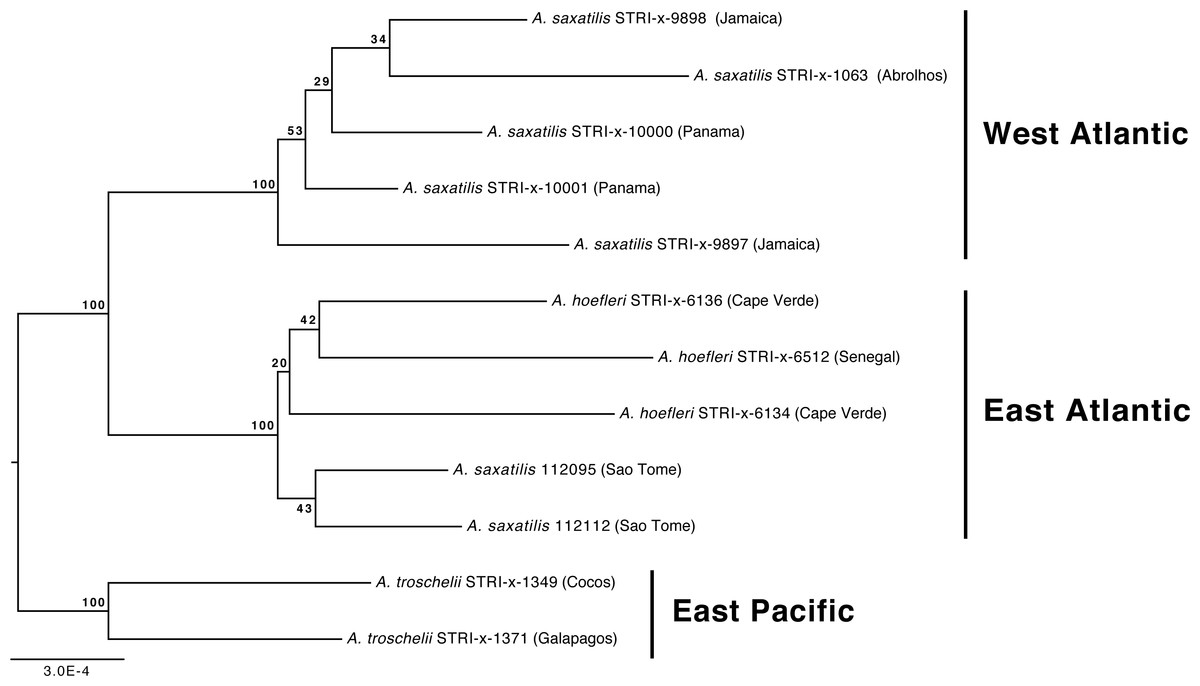
Figure 3: Phylogenetic relationships of Abudefduf saxatilis and A. hoefleri samples sequenced in this study rooted by A. troschelii.
A maximum likelihood tree was generated by optimal partitioning (Lanfear et al., 2014; Lanfear et al., 2012) with each partition modeled with the General Time Reversible (GTR) model of sequence evolution and gamma-distributed rate variation (Γ). Values at nodes are bootstrap support values. Collection sites of samples are indicated after names and the general geographic area of collection indicated.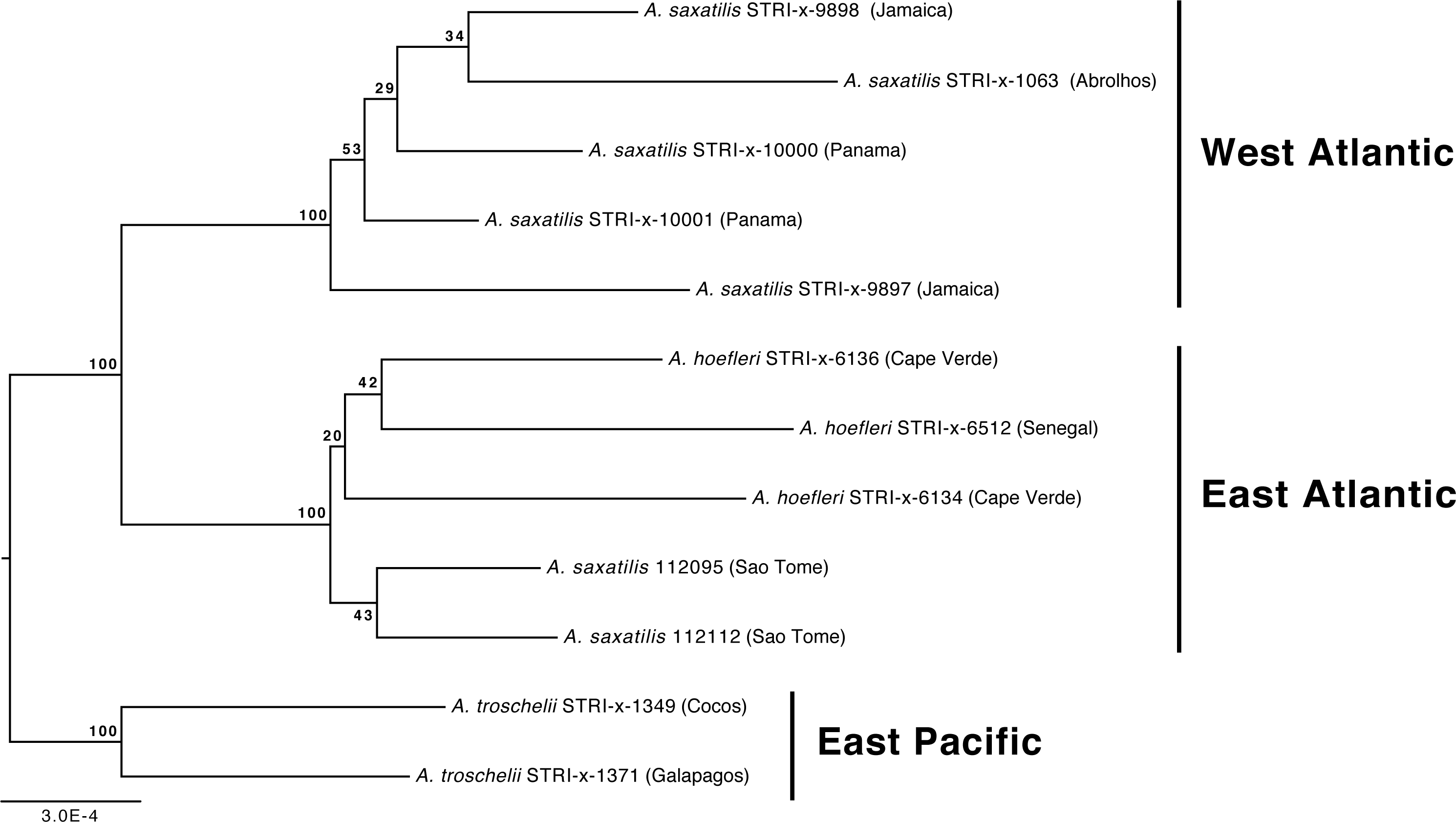{kind=link}
Discussion
General structure of major Abudefduf clades
Our data set is the most comprehensive phylogenetic treatment to date of sergeant-majors, in terms of species examined, geographic range and number of loci. We targeted 500 UCEs for sequencing from 19 described sergeant-major species, and, after limiting missing data, were able to use 361 loci for phylogenetic inference. In many cases, we sampled individuals from within the same nominal species from multiple geographic locations. Among other things, this resulted in the identification of an undescribed species (A. cf. vaigiensis Kiritimati). The increased taxonomic breath, coupled with the large number of loci, increases the accuracy of our phylogenetic analysis and provides a robust framework for exploring diversification within the group.
Broadly, our maximum likelihood and time-calibrated trees corroborate previous work on this group by Frédérich et al. (2013), who examined the Pomacentridae as a whole, and used in part previously published DNA sequence data. We identified three major clades (A, B, and C) (Figs. 1 and 2), relationships among them agree in both studies with the overall arrangement of major clades being (A(B,C)) and the divergence times of major clades are comparable to those of Frédérich et al. (2013) as well. However, relationships we found within Clade C, the most species-rich lineage, differ from some of those of Frédérich et al. (2013). These differences are related to the additional species we examined, the geographic range of our sampling and, perhaps, due to an increased number of variable sites. Greater representation of species within Abudefduf Clade C with substantially more loci in our study (up to 361 here versus four for Frédérich et al., 2013) not only changed relationships among some clade members, but, more importantly, allowed examination of the tempo of diversification within each of the three major lineages of the genus.
Paraphyly in Abudefduf vaigiensis
Recently, genetic variation of mitochondrial and two nuclear loci was examined across much of the geographic range of A. sexfasciatus and A. vaigiensis by Bertrand, Borsa & Chen (2017). That study identified four highly distinct A. vaigiensis lineages, which did not form a monophyletic group relative to other closely related species those authors examined. Bertrand, Borsa & Chen (2017) attributed this pattern to widespread, albeit rare, hybridization among species and the presence of a number of cryptic species within A. vaigiensis. Although we sampled far fewer individuals from fewer locations, there are strong indications that A. vaigiensis is indeed paraphyletic and is composed of several distinct lineages.
Our dataset resolves an issue about one of the four A. vaigiensis clades examined by Bertrand, Borsa & Chen (2017), the identity of an “A. vaigiensis” specimen from Kiritimati (STRI-X-6064; cytochrome b sequence AY208557). The Bertrand, Borsa & Chen (2017) study indicated that this individual fell on a distinct lineage sister to A. bengalensis, but with weak support (BS <50%). We assembled a partial mitochondrial genome from our Kiritimati specimen of A. cf. vaigiensis (STRI-X-6065) and this individual possessed an identical mtDNA haplotype across 1141 base pairs of cytochrome b to the STRI-X-6064 sequence in the NCBI nucleotide collection database. Our phylogeny clearly indicates this individual is only distantly related to the lineage of A. vaigiensis for which we have samples (see below). Nonetheless, a photo of A. cf. vaigiensis taken at Kiritimati (see Fig. 1) shows that the color pattern of five black bars on a pale body and pale fins is more similar to that of A. vaigiensis than that of A. bengalensis, or other potential near relatives (Fig. 1). This discrepancy between color pattern and phylogenetic placement may reflect similar patterns having evolved independently in this lineage or the “vaigiensis”-like pattern being ancestral. The color pattern of A. cf. vaigiensis also is similar to that of A. conformis (Fig. 1), which is endemic to the isolated Marquesas Islands (Range Map 1), and not likely to also occur in Kiritimati, ∼2,200 km away. For these reasons, and the fact that A. vaigiensis has been recorded in the central Pacific (Range Map 1), DRR labeled the specimens which he collected at Kiritimati in 1996 (STRI-X-6064 and STRI-X-6065) as A. vaigiensis. The mitochondrial phylogeny of Bertrand, Borsa & Chen (2017) and phylogenies in this paper from nuclear gene sequence data both indicate that A. cf. vaigiensis Kiritimati is a distinct species.
Unfortunately, all the remainder of our wide geographic sampling of A. vaigiensis occurred in the area occupied only by A. vaigiensis lineage “A” of Bertrand, Borsa & Chen (2017). Fragmentary cytochrome b data from STRI-X-1443 and STRI-X-1497 are greater than 99% similar to the A. vaigiensis reference mitochondrial genome AP006016 and A. vaigiensis A sequences from Bertrand, Borsa & Chen (2017). Thus, based on the partial mitochondrial genome data from our assemblies, we lack samples corresponding to A. vaigiensis lineages B and C of Bertrand, Borsa & Chen (2017). Recently, A. caudobimaculatus, (Okada & Ikeda, 1939), was resurrected from synonymy with A. vaigiensis (Allen, 1991) by Wibowo, Toda & Motomura (2017), on the basis of morphological variation, including relatively minor differences in its color pattern. While no genetic data were examined in that study, the geographic distribution of A. caudobimaculatus corresponds most closely to that of A. vaigiensis- lineage B of Bertrand, Borsa & Chen (2017). Bertrand, Borsa & Chen (2017: Fig. 2), also provide some support for the notion that their lineages A and C represent two other cryptic species within the A. vaigiensis clade. Overall, the emerging data indicate that A. vaigiensis actually comprises three species (not including the unrelated species from Kiritimati) with largely allopatric distributions. If substantiated, the division of A. vaigiensis into additional species would increase the number of localized endemics in planktivore Clade C.
Relationships of, and paraphyly in A. sexfasciatus
Our inclusion of the local-endemics, A. natalensis and A. conformis, which were not represented in Bertrand, Borsa & Chen (2017), alters the placement of A. sexfasciatus to be most closely related to A. natalensis with high support, which is consistent with Hensley & Randall’s (1983) discussion of morphological similarities between them. Our study also indicates that A. vaigiensis “A” of Bertrand, Borsa & Chen (2017) is most closely related to A. conformis. Through wide sampling of A. sexfasciatus, Bertrand, Borsa & Chen (2017) identified shallow subdivisions between the Indian Ocean, Coral Triangle and Western Pacific, as well as the genetic distinctiveness of more peripheral populations. These geographic divisions provide further evidence for the propensity of planktivores to form genetically isolated populations that ultimately may lead to the formation of new species. Our sampling of A. sexfasciatus was limited to two adjacent sites within the Coral Triangle of Bertrand, Borsa & Chen (2017) and is too limited to provide any further insight into this question.
Atlantic-East Pacific geminate lineages
Our examination of the benthivorous Clade A indicates that A. taurus is divided into WA and EA populations that represent divergent allopatric populations or (perhaps) cryptic species. No phylogeographic subdivisions were evident in the EP A. concolor, where distances isolating sampled populations are much smaller than separating EA and WA (see Range Map 5 and Fig. S3). The A. concolor and A. taurus geminate pair arose when the closure of the Isthmus of Panama divided their common ancestor. Subsequently the common ancestor of A. taurus lineages divided into EA and WA A. taurus populations (or species).
The geminate species relationship involving A. saxatilis (EA and WA), A. hoefleri (EA), and A. troschelii (EP) are clarified here. Rather than A. troschellii and A. hoefleri being sisters (see Frédérich et al., 2013), our analyses indicate that the sister of A. troschelii is the ancestor of an Atlantic “species” that now comprises A. saxatilis and A. hoefleri, and in which the EA A. saxatilis is more closely related to the EA A. hoefleri than to WA A. saxatilis. The phylogeographic pattern reported by Frédérich et al. (2013) resulted from a lack of separate EA and WA A. saxatilis representatives in their dataset, and the use of fewer loci (largely GenBank derived super-matrix). Support values for monophyly of the A. troschelii + (A. hoefleri + A. saxatilis) geminate clade are not consistently strong in our study (BS = 65%, PP = 1.00), which perhaps reflects underlying challenges to identifying these three species as a clade with a molecular phylogenetic framework. Monophyly of A. hoefleri was only supported, but weakly so, by using many UCEs (59,853 characters in alignment), while A. saxatilis is paraphyletic in both our phylogenetic placement and A. saxatilis alignment analyses. Analysis of genetic variation across the WA range of A. saxatilis indicated a lack of genetic structuring in either mtDNA or microsatellites within the Caribbean, and a break between the Caribbean and Brazil populations of A. saxatilis in microsatellite data, but not mtDNA data (Piñeros & Gutiérrez-Rodríguez, 2017). Our A. saxatilis alignment-phylogenetic-analysis (Fig. 3) indicates strong support for WA monophyly, and a lack of phylogenetic structure within that area. Our estimate of the TMRCA of the A. troschelii + (A. saxatilis, A. hoefleri) geminate clade is very recent, 2.4 MYA (95% HPD 0.75 –4.9 MYA). Given the capability for long distance gene flow via rafting in A. saxatilis (Luiz et al., 2012; Piñeros & Gutiérrez-Rodríguez, 2017), a population genetics analysis that includes both sides of the Atlantic and the mid-Atlantic islands and that examines these three species as a geminate clade would be useful for demonstrating the extent of gene flow and isolation among them.
Lifestyle transitions and diversification rates
Except for A. notatus, all species in both Clades A (taurus) and B (sordidus) are benthic-feeding herbivores that are characterized by large body size, chunky bodies and blunt fins (Aguilar-Medrano & Barber, 2016). They also have uniformly dark brownish bodies and unpaired fins with indistinct pale vertical bars on the body (Fig. 1). Clade C comprises planktivorous fishes that feed in the water column near the surface and that have more slender bodies and longer, more pointed fins than the benthivores (Aguilar-Medrano & Barber, 2016). Species in Clade C have more conspicuous and variable color patterns that often are essentially the reverse of the benthivore pattern, with pale, silvery to silvery-yellow bodies and strong dark vertical bars (Fig. 1). Other planktivores have pale bodies with indistinct dark bars, while some lack dark bars and have large blotches of dark pigment, and others have black stripes along the upper and lower edges of the caudal fin. A. notatus, a member of Clade B, is intermediate in its trophic ecology between benthivores and planktivores (Masuda & Allen, 1993) and, like planktivores of Clade C, occurs in (sometimes large) aggregations that feed on plankton in midwater (see: https://www.peerintoyourworld.com/species/pomacentridae/abudefduf-notatus-yellowtail-sergeant/, accessed January 12, 2018). This species, which evidently is in transition from benthivory to planktivory, the second such a transition within Abudefduf, also has a color pattern intermediate between that of benthivores and planktivores (see Fig. 1).
Statistical testing with BAMMTools to determine whether diversification rates among the three Abudefduf lineages were different was inconclusive. The relatively small number of species in our dataset results contributes to low power which is typical of tests of this type (Agapow & Purvis, 2002; Kubo & Iwasa, 1995). The use of BAMMTools to accurately detect rate variation also has been questioned (Moore et al., 2016; Rabosky, Mitchell & Chang, 2017). In general, being able to assert that a key innovation is causing increased diversification is problematic (De Queiroz, 2002) and variation in the numbers of extant species in different clades of the same age can, for example, be produced by random speciation and extinction events (Gould et al., 1977; Raup et al., 1973; Slowinski, 1990).
Nevertheless, Clade C contains more than twice as many planktivores as there are benthivores in Clades A and B combined (>17 versus six), and 4-8 times the number of benthivores in either of those two clades. Furthermore, although Clade A is distinctly older than clade C (Fig. 2), the diversification rate is notably higher in Clade C than either Clades A and B: 0.12 species MY−1 in Clade A, and 0.19 species MY−1 in Clade B (0.13 if A. notatus is excluded), versus 1.13 species MY−1 in Clade C. There is an important caveat to this conclusion, that our geographically limited sampling regime may have underestimated the number of species in Clade B. For widespread species, widespread sampling is important, as indicated by our capture of only one of the three major lineages known to exist in A. vaigiensis (plus the undescribed species from Kiritimati). One of those three lineages has recently been shown to be morphologically distinct (Wibowo, Toda & Motomura, 2017) and needs to be genetically assessed. Evidence that A. bengalensis is composed of at least two species was published during review of this manuscript, supporting enhanced diversity of planktivores, as well as emphasizing the importance of geographically dispersed sampling (Wibowo et al., 2018). In our study, we relied on prior knowledge of the existence of various named and morphologically distinct species, leading to much more geographically dispersed sampling of Clade C. In contrast, our sampling of one of the benthivorous clades, which lack obvious morphological differences, was geographically much more restricted. Clade A was sampled effectively across its range. However, that is not the case for Clade B, in which we used only samples from a single location for both A. septemfasciatus and A. notatus, which have large Indo-Central Pacific and Indo-West Pacific ranges, respectively. Thus, we certainly missed any differentiation that might exist across the ranges of these two species. However, our sampling of populations of A. sordidus at two central Pacific sites 3,300 km apart, and the Red Sea, 15,000 km from either of those on the opposite side of that species’ range, produced levels of divergence similar to those within many others of the species we sampled (Fig. 1, Fig. S4). One of those sites (Johnston Island) is a location where a local-endemic planktivore (A. abdominalis) lives (Randall, 2007). To help overcome these limitations in our sampling of A. sordidus and A. septemfasciatus we examined existing mitochondrial data on those species in GenBank merged with mitochondrial sequence data generated as a by-product of sequence capture with samples in this study. That dataset is much more extensive in terms of its geographic coverage for both species. Analysis of mitochondrial data from A. sordidus and A. septemfasciatus (Appendix S1) also indicates that each species represents a single widespread, Indo-central Pacific species rather an aggregate of multiple allopatric species that include local endemics. Similarly, divergence of A. taurus across the Atlantic is relatively small, and less than that within many other species. Moreover, unlike the situation among members of Clade C, (minor) morphological differences between local endemics has been described in only one case among Clades A and B species (A. declivifrons vs A. concolor: Lessios et al., 1995).
With these caveats in mind, the higher number of species within Clade C remains striking and suggests an increase in speciation rate associated with a transition to planktivory. Herbivorous fishes were instrumental in the creation of modern coral reefs, and have occupied coral reefs for at least the last 50 million years (Bellwood, 1996; Bellwood, 2003). High availability of planktonic food resulting from increased coastal upwelling is a more recent phenomenon, starting in the Late Miocene (∼10 MYA) and has been linked to other species radiations (Jacobs, Haney & Louie, 2004). Transitions from benthivory to planktivory are widespread across reef fishes (Floeter et al., 2018; Tavera, Acero & Wainwright, 2018), and have occurred rapidly in various damselfishes (Cooper & Westneat, 2009). Our results are consistent with the general trend across perciform fishes, in which herbivorous lineages show lower levels of diversification relative to the more recently derived and species-rich invertebrate feeders (see also Clements et al., 2004; Gomon & Paxton, 1985).
We propose the hypothesis that, in Abudefduf, transitions to planktivory and increased diversification among planktivores may be due to a combination of (i) trophic niches for benthivores generally being pre-occupied by other damselfishes that successfully defend a predictable resource against heterospecifics, and (ii) zooplankton being a high-quality, easily digestible food that is less predictably available and, hence, economically less controllable by species already resident on reefs when the transition to planktivory began in Abudefduf. Speciation among benthivorous Abudefduf may have been limited by competition from the stegastinine damselfishes, which are common on reefs worldwide and which have many species that defend benthic algal resources. The TMRCA of Stegastinae predates the origin of Abudefdufinae by approximately 20 million years (Frédérich et al., 2013). Consequently, prior occupancy of benthivorous niches by stegastinine fishes may have reduced the ability of Abudefduf species to diversify into that ecological role (Lobato et al., 2014). Observations by DRR on both sides of the Isthmus of Panama show that A. taurus and A. concolor live in intertidal and upper subtidal areas, above the zones occupied by dense, multispecies assemblages of benthic-feeding stegastinine damselfishes (species of Microspathodon and Stegastes) (see also information on depth ranges those two Abudefduf species in their IUCN Red List reports: http://www.iucnredlist.org). A. sordidus and A. septemfasciatus in the Indo-Pacific also have similarly narrow, very shallow depth-ranges (Allen, 1991). Benthivorous Abudefduf species may have not only few available niches to diversify into, but also sufficient gene flow to counteract any neutral divergence may occur even across large spatial scales in these species.
The tempo of diversification
Although Clade C is the most species-rich, all three clades of Abudefduf increased in speciation rate during the last 10 MY (Fig. 2). This increase may be linked to sea level changes. Since the Late Miocene (i.e., ∼10 MYA) sea level has trended downwards and become more variable than during the previous history of Abudeduf, when it was more stable and higher than currently exists (Miller, 2009). The start of diversification of Clade C coincides with the onset of this environmental change. An association between increased speciation and sea-level changes has previously been noted in other reef fishes (e.g., Ludt & Rocha, 2015; McCafferty et al., 2002; McMillan & Palumbi, 1995). The creation of numerous islands/reefs to which dispersal and then speciation occurred (peripatric speciation), isolation across the Sahul and Sunda shelves as they were exposed (allopatric speciation) or isolation due to reduction and fragmentation of coastal habitat and populations during low sea level stands (allopatric speciation) may be the underlying mechanisms of this diversification. Thus, the diversification of the plantivores in Clade C may be driven both by the switching to a different food source, and the sea level oscillations that separated populations.
Local endemism
Both planktivorous and benthivorous Abudefduf are pantropical and have broadly overlapping distributions; however, there is a discrepancy in the level of local endemism between these two trophic guilds. Nine of 16 named (and one unnamed) planktivorous species are local endemics, which occur in the Central Pacific (A. abdominalis, A. conformis, A. cf. vaigiensis; Range Maps 1 & 3), East Pacific (A. troschelii, Range Map 1), East Atlantic (A. hoefleri, Range Map 1), Southwest Pacific (A. whitleyi, Range Map 2), and Southwestern Indian Ocean (SWIO; A. natalensis, A. sparoides, and A. margariteus; Range Maps 2 & 3). In contrast, only two of six named species in either benthivorous clade of Abudefduf clearly are local endemics: A declivifrons and A. concolor, which occupy partly overlapping sections of the EP (Range Map 5). The relatively few species in Abudefduf, particularly in each of the benthivorous clades, makes it difficult to test statistically for differences in levels of regional endemism between trophic guilds and clades. It does appear that the relative proportion of local-endemic benthivores (0.33) is less than that of local-endemic planktivores (0.56); however, a X2 test of these proportions is not significant. If these differences are real, what biological properties could lead to the abundance of local-endemic planktivores?
Dispersal ability can be a key factor affecting genetic differentiation between populations of some marine species (Bohonak, 1999; Slatkin, 1987). Here this raises the question—are increased speciation and the creation of local endemics related to a lower dispersal ability in planktivores (compared to benthivores)? The great majority of coral reef fishes have a pelagic larval stage, the duration of which varies between species (Sale, 1980), and has led to the expectation that range size would be positively related to the duration of the pelagic larval duration (PLD). However, a general relationship to that effect has not been found among tropical reef fishes (Lester & Ruttenberg, 2005). Further, species of Abudefduf examined to date have comparatively short PLDs, which should generally limit dispersal potential, and there is little evidence of a consistent difference in PLDs between planktivorous and benthivorous species (Luiz et al., 2012). Range-size in tropical reef fishes evidently is affected by a suite of characters other than PLD, including the ability of post-larval stages to raft on flotsam, as well as adult-biology characteristics that help establishment following dispersal (Jokiel, 1990; Lester et al., 2007; Luiz et al., 2013; Luiz et al., 2012). Post-larval juveniles and even adults of planktivorous, but not benthivorous, Abudefduf frequently associate with flotsam (Hunter & Mitchell, 1967; Kimura et al., 1998; Nakata, Takeuchi & Hirano, 1988; Luiz et al., 2013; Luiz et al., 2012).
Overlaps in the geographic ranges of sister taxa of planktivorous Abudefduf species that include both widespread species and local endemics, e.g. between A. saxatilis and A. hoefleri in the East Atlantic (Figs. 1 & 2 , and Range Map 1) and between A. natalensis, A. sparoides and A. sexfasciatus in the Southwest Indian Ocean (Figs. 1 & 2, and Range Maps 2 & 3) clearly demonstrate how widespread planktivores have dispersal powers that enable them to repeatedly colonize sites where they previously evolved into local endemics. Differences in dispersal characteristics of planktivorous and benthivorous Abudefduf species indicate that planktivores have greater dispersal capabilities, and hence should have larger ranges, greater connectivity and fewer local endemics. This is the reverse of what is observed.
If not dispersal ability, then what may explain the difference in endemism levels between planktivores and benthivores? The general appearance of benthivores and planktivores is strikingly different (Fig. 1) and may provide insight into this question. Benthivorous Abudefduf are very similar in morphology and color, remarkably so in some cases (Lessios et al., 1995), suggesting restrictions on the variation that can be exhibited in such species. Cryptic coloration may be sufficiently important for shallow-living benthivores to constrain color variation among them. Planktivorous Abudefduf, in contrast, are less cryptically colored than the benthivores and also show much more interspecific variation in color patterns, including among local endemics (Fig. 1). This variation may be the key factor indicating how differences in diversity between benthivores and planktivores evolved. Small isolated populations often develop odd color forms (e.g., Feitoza et al., 2003), and fixation of color variation may occur very early in the speciation process, promoting the rapid development of allopatric color variants (e.g., Gaither et al., 2014). One well-studied example highlights the importance of color variation in speciation. The Caribbean hamlets (Serranidae: Hypoplectrus) contain many species that vary only in color, and often share the same reef (Fischer, 1980; Holt et al., 2008; Mccartney et al., 2003; Puebla et al., 2007; Puebla, Bermingham & McMillan, 2014; Whiteman, Côté & Reynolds, 2007). Color is of paramount importance for hamlets as the vast majority of matings are between individuals of the same color (Barreto & McCartney, 2008; Fischer, 1980; Puebla et al., 2007), and both natural and sexual selection can act to generate assortative mating that will in turn lead to reproductive isolation and speciation even with gene flow (Puebla, Bermingham & Guichard, 2012). Presence of color-assortative mating in Chaetodontidae (McMillan, Weigt & Palumbi, 1999), Serranidae (see references above) and Cirrhitidae (Whitney, Bowen & Karl, 2018) indicates that the process is generally distributed among reef fishes. Hence, the idea that within planktivorous Abudefduf, an ability to vary color may have promoted assortative color-based mating that facilitate speciation and the creation of regional endemics is not unreasonable.
Conclusions
Two of the three major clades of Abudefduf (Clades A and B) are primarily benthivores, apparently have few regional endemics, and many of their few species are widely distributed. In contrast, Clade C comprises planktivores and includes both wide-ranging species and a higher proportion of regional endemics. Past instability in sea level over the last 10 MYA appears to be linked to increased recent speciation in all three clades.
Paradoxically, there are ∼three times as many species of planktivores in a single clade as benthivores in two well-separated clades. Although neither differences in diversification rates nor the relative abundance of local endemics were statistically different between clades or trophic guilds of Abudefduf, this likely reflects small sample sizes. This pattern exists even though dispersal by planktivores through flotsam-rafting (Luiz et al., 2012; Luiz et al., 2013) may lead to higher dispersal rates likely to promote gene flow and reduce isolation. Benthivorous Abudefduf species, in contrast, are not known to participate in flotsam dispersal. Despite the wide, Indo-central Pacific ranges of some benthivores, i.e., A. sordidus and A. septemfasciatus, they do not appear to have produced local endemics, particularly local endemics with distinctive color patterns. Thus a large question remains open in Abudefduf: why do planktivores, with greater dispersal powers, have a much stronger tendency than benthivores to produce morphologically distinct local endemics? In another genus of damselfishes it has been proposed that biological factors such as ecological pressures and sexual selection can generate new species even when dispersal barriers are absent (Leray et al., 2010). We suggest that an ability of planktivores to vary in morphology and color facilitates assortative mating that leads to speciation, while natural selection for crypticity constrains coloration of benthivores, and restricts speciation through assortative mating.
Future research directions
To test our hypothesis that planktivorous Abudefduf have a greater capacity to diversify and form local endemics than do benthivorous congeners, a multilocus analysis based on broad geographic sampling is needed for all Clade B species throughout their large Indo-central Pacific ranges, particularly at sites occupied by local-endemic planktivores. That will demonstrate the extent to which benthivores represent broadly distributed species versus collections of allopatric cryptic species. Further sampling of broadly distributed planktivores in the Indo-central Pacific (A. sexfasciatus and A. vaigiensis) and parts of the Atlantic (A. saxatilis) also are necessary to clarify the extent to which they constitute a collection of local endemics. In addition, detailed information on the trophic and community ecology of all benthivores is needed to understand how they fit into speciose assemblages of benthivorous damselfishes, many of which aggressively control benthic resources and arose long before Abudefduf evolved.
Supplemental Information
Sample name, location, number of assembled contigs and UCEs with indication if used in Bayesian time tree analysis
For each sample included in analyses the species name and unique identifier are given. We also indicate the collection information associated with each sample and how many contigs and UCEs were assembled, and if the sample was included in the Bayesian simultaneous divergence time estimation and phylogenetic inference analysis.
Calibration points for divergence time estimation
Characteristics and sources for calibrations used for divergence time estimation.
Maximum likelihood phylogeny of Abudefduf with individual identifiers
A maximum likelihood phylogenetic of Abudefduf generated from partitioned analysis of 361 ultraconserved element (UCE) loci. An optimal partitioning strategy was implemented (Lanfear et al., 2014; Lanfear et al., 2012) . Each partition was modeled under the General Time Reversible (GTR) model of sequence evolution with Gamma distributed rate variation (G). Bootstrap support values are indicated with an asterisk (*) if equal to 100. The three lineages of Abudefduf are indicated (Clade A, B and C). Planktivorous lineages are highlighted and named in red with benthivorous lineages highlighted and named in blue. The tree is rooted by pomacentrid outgroups of the genera Chromis and Similiparma. Appended to each leaf is the identifier of the particular sample examined. Photo credits: A. hoefleri S. Floeter, A. saxatilis DRR, A. troschelii GRA, A. conformis J. Randall, A. vaigiensis GRA, A. abdominalis GRA, A. sparoides GRA, A. sexfasciatus J. Greenfield (CC BY), A. natalensis J. Randall, A. whitleyi GRA, A. bengalensis G. Edgar (CC BY), A. lorenzi GRA, A. margariteus GRA, A. cf. vaigiensis J. Earle, A. notatus GRA, A. septemfasciatus GRA, A. sordidus GRA, A. taurus DRR, A. concolor GRA, A. declivifrons GRA.
Time-calibrated phylogenetic tree of Abudefduf with age of nodes labeled
Time-calibrated phylogenetic tree of Abudefduf generated from 137 ultraconserved element (UCE) loci modeled under a single partition with the General Time Reversible (GTR) model with Gamma distributed rate variation (Γ). Fossil calibration points are described in Table S2. Nodes are labeled with posterior medians of divergence time estimates with blue bars indicating 95% highest posterior density. The major trophic guilds are indicated by shading of red for planktivores and blue for benthivores. Regional endemics are indicated by a dagger (†). Individual samples included in this analysis are indicated in Table S1.
Maximum likelihood phylogenetic tree of Abudefduf taurus and A. concolor samples examined for this study
A maximum likelihood phylogenetic tree of Abudefduf taurus and A. concolor samples examined for this study. The tree was generated from 118 ultraconserved element (UCE) loci modeled under three partitions as defined by PartitionFinder (see Methods). Each partition was modeled with the General Time Reversible (GTR) model with Gamma distributed rate variation (Γ) and bootstrapping was conducted to assess confidence, with bootstrap values shown at nodes. Individual tissue identifiers and the general geographic collection area are appended to taxon names. For A. taurus, the clear geographic division between the East and West Atlantic is indicated. The other two species examined are both from the East Pacific which is also indicated on the figure.
Clade B sampling locations compared to the ranges of Abudefduf endemics
Comparison of sampling locations of the Abudefduf sordidus Clade, “Clade B” to the distribution of regional endemic Abudefduf species. Clade B sampling points are plotted color-coded to species as indicated in the key, while endemic distribution points are plotted from data points as described in Range Map Methods.docx.