. Introduction
The Paraguayan ka’a he’ ẽ, sweet herb, Stevia rebaudiana Bertoni (Asteraceae) has been used as a sweetener since ancient times by the indigenous people of Amambay, which is the place where the seedlings of that plant were taken from to start large-scale cultivation in various areas of the eastern region (Levy, 1984; Soejarto et al., 1983). Nowadays, the Ministry of Agriculture and Livestock considers the cultivation of S. rebaudiana as an important means of diversification of agricultural production (Casaccia & Álvarez, 2006).
Products based on S. rebaudiana have a number of properties that make them more attractive than sugar and the artificial sweeteners available on the market; hence, their demand exceeds the agricultural production (González-Moralejo, 2011). Therefore, in the same way as the demand increases, the production and optimization of the plant must increase as well (Brandle et al., 1998).
In addition, stevia is a valuable medicinal plant with wide application in the pharmaceutical and food industry. The food industry is increasingly interested in replacing artificial sweeteners with other types of natural sugars in order to offer the consumer a wider range of options and meet the needs of a segment of the population that does not want or cannot eat sucrose. Stevia leaves have been used as a low calorie sweetener for centuries and are now consumed worldwide. The sweetness of this plant is due to the accumulation of bioactive compounds, especially steviol diterpenoid glycosides, which constitute up to 30% of the dry weight of the leaf (Geuns, 2003). Steviosides are 300 times sweeter than regular table sugar, with the added benefits of zero calories, zero carbohydrates, no spikes in blood sugar, the non-ferment ability thermal stability at 100 °C, and a long service life (Singh & Rao, 2005).
One of the methods of asexual propagation is through in vitro propagation, which has been successful due to its effectiveness and produces homogeneous material that preserves the characteristics of the mother plant (Vilchez & Albany, 2014). Additionally, through this technique, the behaviour of varieties exposed to different stressors controlled conditions can be evaluated, and it is thus possible to determine which ones are more suitable for different field conditions.
Crop yield depends on irrigation and other agronomic practices (Aladakatti et al., 2012; Fronza & Folegatti, 2003; Jahan et al., 2020; Abd El-Samad & Taha, 2022). According to the agroecological requirements, the crop must be established in soils with an adequate level of humidity guaranteed by the installation of irrigation systems, which constitute 34% of the total production cost (Casaccia & Álvarez, 2006; Niño et al., 2013).
This study will determine the response of KH/IAN VC-142 (Eirete variety) of S. rebaudiana in hydric and saline stress conditions and evaluate biochemical and growth parameters during the process. Since the reduction of the use of water in agricultural systems constitutes a fundamental priority given its strong repercussion on the total amount of water used in agriculture and the high cost of irrigation systems, the identification of the mechanisms involved in these responses would allow the implementation of new strategies for the management and improvement of crops in conditions of hydric and saline stress (Rodriguez-Péres, 2006; Medrano et al., 2007).
. Materials and methods
Vitroplants of KH/IAN VC – 142 (Eirete variety; honey in Guaraní) of S. rebaudiana from the Biotechnology Laboratory of the Multidisciplinary Center for Technological Research (CEMIT-UNA) were used in the study. Plants were grown in Murashige and Skoog (MS) (Murashige & Skoog, 1962) semi-solid culture medium conditions of 25 °C, 3000 Luxes, and 16 h photoperiod (without the addition of growth regulators).
Nodal segments of S. rebaudiana were transferred to the MS culture medium supplemented with 0 (control), 25, 50, 75, 100, or 150 mM NaCl to determine the concentration at which the growth of the individuals was reduced by 50% (RD 50) (100 nodal segments per treatment). Another part of the segments was supplemented with 0, 50, 100, 150, or 200 mM NaCl to determine the concentration at which death occurred in 50% (LD 50) of the individuals 25 days after treatment (100 nodal segments per treatment). The design was completely randomized, each treatment consisted of 5 repetitions, and each repetition consisted of 20 nodal segments, thus totalling 100 segments/vitroplants per treatment.
The evaluation of the survival of the vitroplants and the measurement of the height, fresh and dry weight of the aerial parts, length of the roots, and fresh and dry weight of the roots in each plant were carried out on individuals treated with the concentration corresponding to 50 mM NaCl. To obtain the dry weight, the samples were deposited in an oven at 60 °C for 24 h.
The selected biochemical parameters were determined in individuals treated with the concentration corresponding to 0 (the control) or 50 mM NaCl. The proline concentration was analysed using the Bates method (Bates, 1973). Lyophilized plant samples (50 mg) taken from the different treatments were mashed with 5 mL of sulphosalicylic acid in an aqueous solution (3% w/v). The samples were centrifuged at 1,200 rpm for 10 min at 5 °C. Then, 2 mL of the supernatant were placed in an assay tube; the control was prepared by replacing of the supernatant by 3% sulphosalicylic acid. 2 mL of glacial acetic acid and 2 mL of acid ninhydrin were added to each tube, the mixture was homogenized by a vortex, and the assay tubes were put into double boil for 1 h. The reaction was ended by cooling in ice. When the samples achieved room temperature, 1 mL of the mixture was extracted with 1 mL of toluene and placed in the vortex mixer for 20 s. Then, the supernatant absorbance was read at 520 nm with a spectrophotometer.
The chlorophyll concentration was determined using the Lichtenthaler equations (Lichtenthaler, 1987). According to this method, 0.25 g of fresh tissue was extracted using 5 mL of 80% acetone, and the extract was centrifuged at 11,000 rpm for 10 min. The absorbance of the obtained extracts was measured at wavelengths of 646 and 663 nm to estimate chlorophyll a (Chla) and chlorophyll b (Chlb) concentrations using a spectrophotometer.
The total soluble sugar concentration was determined using the method proposed by Yemm and Willis (1954). Fresh samples of stevia (0 and 50 of mM NaCl) were weighed (0.2 g) and homogenized using 70% ethanol. Then, they were filtered and pigments were removed with the use of benzene. An aliquot of 0.2 mL of the leaf extract was added to 1.0 mL of 0.2% anthrone to react in a water bath for 10 min at 100 °C. After heating, the test tube was cooled in an ice bath and then the absorbance was read at 620 nm.
All data were analyzed with BioStat®; 2009 and Past®; software for Windows and comparisons of means were performed by the Tukey test (p < 0.05); the significance level was determined by Student’s test (p < 0.05), Mann–Whitney U test (p < 0.05), and Monte Carlo Permutation (p < 0.05) test.
. Results and discussion
Both drought and saline stresses affect various physiological and metabolic processes in plants depending on the development stage, species, time, and degree of exposure to conditions of water deficit or excessive salinity (Jones et al., 1989).
Evaluation of survival of salt-stressed S. rebaudiana in vitro plants
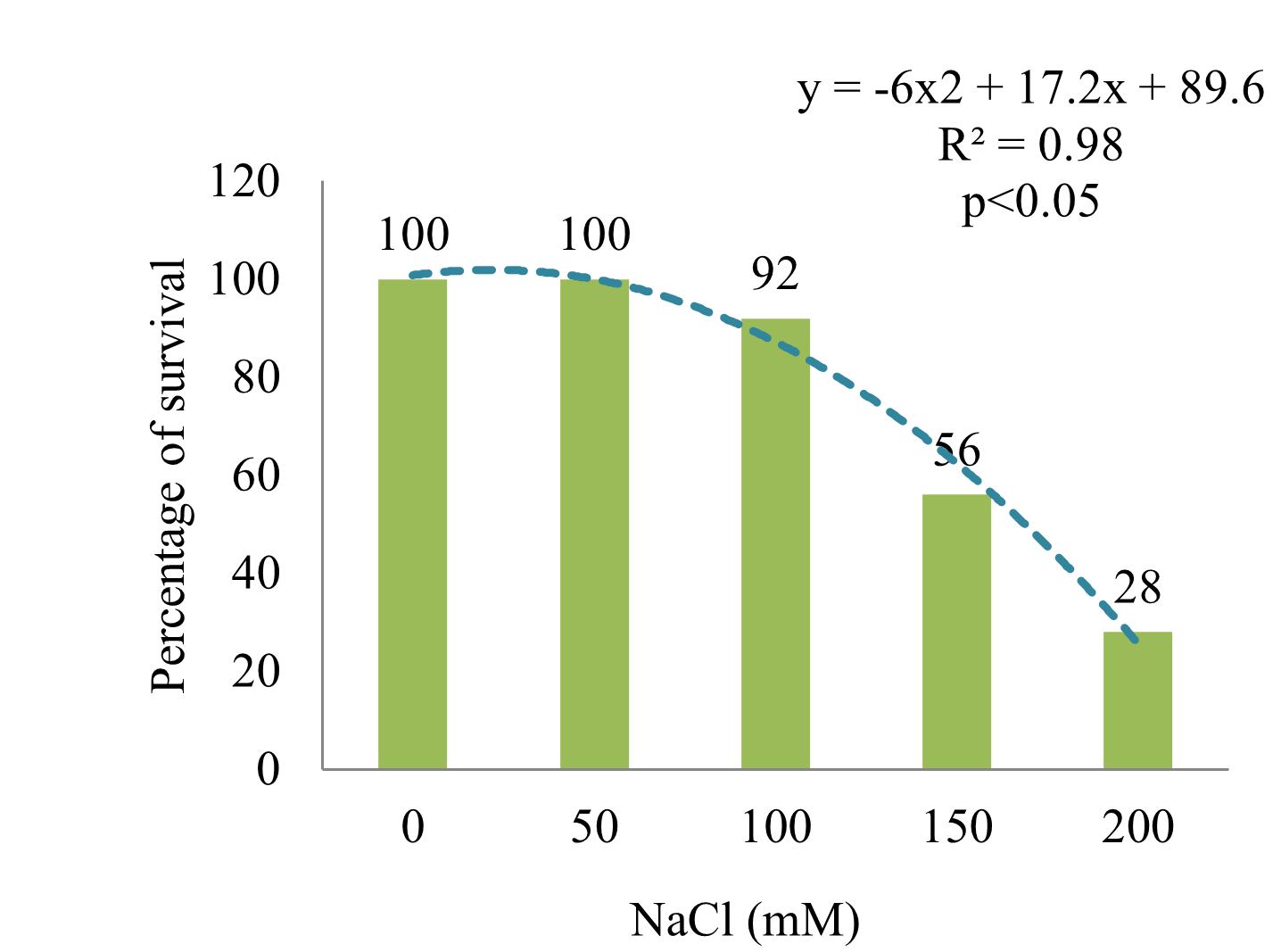
In the Eirete variety of S. rebaudiana, one of the parameters affected was the survival of the plant exposed to high concentrations of NaCl. The percentages of survival after the salt treatment (Figure 1) follow a second order polynomial trend line with a value of R2 = 0.98 and p < 0.05, indicating that plant survival decreases as the concentration of NaCl in the culture medium increases. According to the equation of the curve, the dose that caused the death of 50% (LD 50) of the individuals hada value close to 150 mM NaCl. Plants treated with 200 mM NaCl were not taken into consideration due to their low survival rates.
Evaluation of the height of the aerial parts of salt-stressed S. rebaudiana in vitro plants
The NaCl simulated stress affected the growth of S. rebaudiana (Figure 2). It was found that the mean height of the plants decreased in the presence of the higher NaCl concentrations, except for the plants treated with 25 mM NaCl, which were, on average, higher than the control plants (0 mM NaCl). Despite this tendency, the Mann–Whitney U test (p < 0.05) indicated that there were no statistically significant differences between them. The increase in the average height of the plants treated with 25 mM NaCl compared to the control plants could be due to the variability of the species or to the fact that NaCl at low concentrations can act as a nutrient for plants. An analogous phenomenon occurred in a similar test with sesame, where an increase in the size of the plants treated with 100 mM NaCl was observed in comparison to the control (Koca et al., 2007).
Figure 2
Effect of NaCl exposure on the height of the above-ground parts of S. rebaudiana. Bars represent the mean (±SD) height of the plants after 25 days of treatment with 0, 25, 50, 75, or 100 mM NaCl. Bars represented by different letters indicate significant differences, ANOVA (p < 0.01), Tukey post-hoc test (p < 0.05).
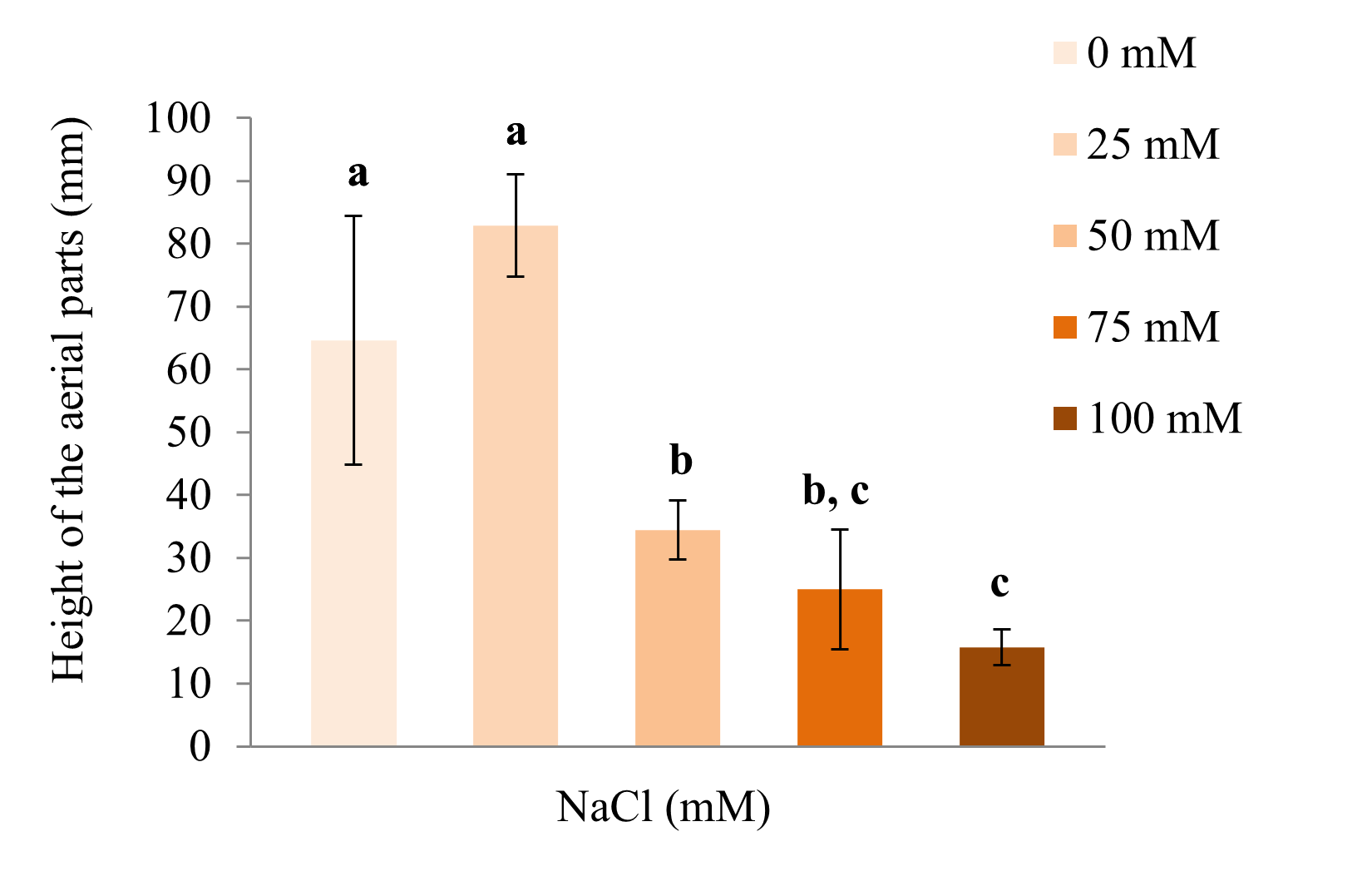
The average height of the plants treated with 50 mM NaCl showed a significant decrease compared to the plants treated with 0 or 25 mM NaCl. The concentration of NaCl that reduced the growth of the shoots of the plants by 50% (RD 50) was close to this concentration.
Plants treated with 75 mM NaCl showed a significant decrease in height compared to the treatment with 0 and 25 mM NaCl; however, it was not highly different from plants treated with 50 mM NaCl. In the treatment with 100 mM NaCl, a significant reduction in the height of the plants was observed with respect to the treatments with 0, 25, or 50 mM NaCl. However, no significant differences were observed in comparison with plants treated with 75 mM NaCl.
When the NaCl concentrations exceeded 25 mM, the growth was inhibited as a mechanism of plant adaptation to adverse conditions. This agrees with various studies, which reported greater growth reduction in higher drought or saline stress conditions. A lower elongation of the stems of S. rebaudiana was also observed as water availability decreased (Niño et al., 2013).
Fresh and dry weight of the aerial parts of salt-stressed S. rebaudiana
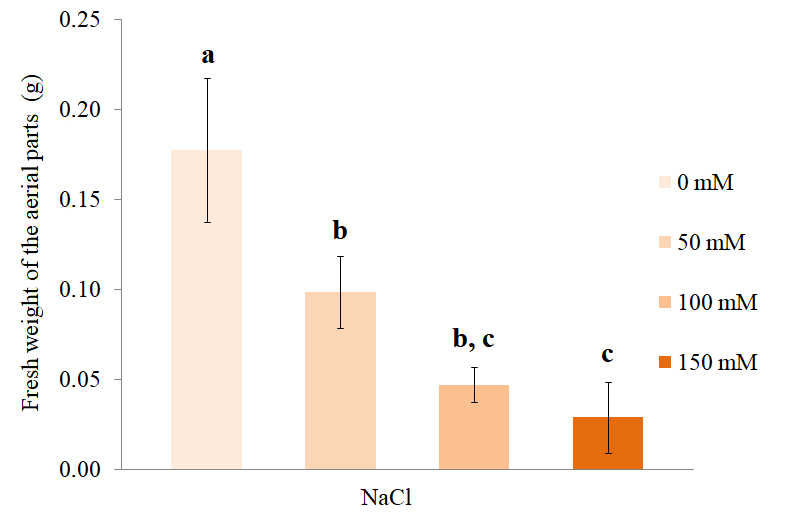
The fresh weight of the aerial parts of S. rebaudiana decreased gradually as the concentration of NaCl in the culture medium increased (Figure 3). Plants treated with 50 mM NaCl showed a significant reduction in the fresh weight of the above-ground parts compared to the control individuals. Plants treated with 150 mM NaCl showed a significant reduction in the fresh weight compared to those treated with 0 and 50 mM NaCl, but not compared to those treated with 100 mM NaCl (Tukey p < 0.05).
Figure 3
Fresh weight of the aerial parts at different concentrations of NaCl. The bars represent the mean (±SD) of the fresh weight of the aerial parts of S. rebaudiana after 25 days of growth in MS culture medium supplemented with 0, 50, 100, or 150 mM NaCl. The bars marked with different letters show significant differences, ANOVA (p < 0.01), post-hoc Tukey test (p < 0.05).
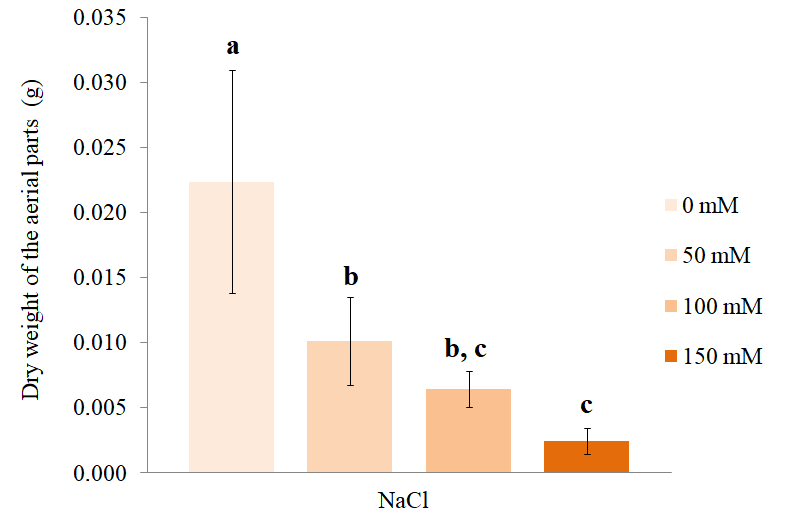
As in the case of the fresh weight, the dry weight of the aerial parts also decreased with an increase in the NaCl concentrations in the culture medium (Figure 4). The S. rebaudiana vitroplants treated with 50 mM NaCl showed a significant reduction in the dry weight of the aerial parts compared to the control individuals. The reduction of the dry weight of the aerial parts of the vitroplants treated with 100 mM NaCl was statistically significant with respect to the control individuals, but not with respect to those treated with 50 mM NaCl. Plants exposed to 150 mM NaCl showed almost 90% reduction in the dry weight of the shoots compared to the control, but this decrease was not significant compared to plants treated with 100 mM NaCl (Tukey p < 0.05).
Figure 4
Dry weight of the aerial parts at different concentrations of NaCl. The bars represent the mean (±SD) dry weight of the aerial parts of S. rebaudiana plants after 25 days of growth in MS culture medium supplemented with 0, 50, 100, or 150 mM NaCl. The bars marked with different letters show significant differences, ANOVA (p < 0.01), post-hoc Tukey test (p < 0.05).
The decrease in the fresh and dry weight of the aerial parts is a reflection of the inhibition of plant growth. This is consistent with various studies demonstrating that the total fresh and dry weight of plants is significantly reduced as the level of stress to which they are subjected increases, either due to the reduction of photosynthesis and carbon fixation or as a mechanism of adaptation (Demiral, 2005; García et al., 2010; González et al., 2005; Niño et al., 2013). Studies in Phaseolus vulgaris L. confirm that upon addition of 40 mM NaCl, a significant reduction in the biomass of shoots can be observed (García et al., 2010). In turn, a study conducted by Shahverdi et al. (2019) has shown a NaCl threshold and EC50 (the NaCl level that causes a 50% decrease in growth) for S. rebaudiana of 77 and 157 mM, respectively.
Length of the roots of salt-stressed S. rebaudiana
According to the Mann–Whitney U test and the Monte Carlo Permutation test, S. rebaudiana exposed to 0 or 50 mM NaCl (Figure 5) did not exhibit significant differences in the length of their roots. This agrees with a study carried out on Perilla frutescens, where it was observed that, between 0 and 50 mM NaCl, there were no significant differences in the root length (Zhang et al., 2012). Moreover, the lengths of the roots showed a great dispersion between individuals that received the same treatment.
Figure 5
Length of roots of S. rebaudiana at different concentrations of NaCl. Each point indicates the total root length (mm) of S. rebaudiana subjected to 0 or 50 mM NaCl, representing the dispersion of the data in each treatment.
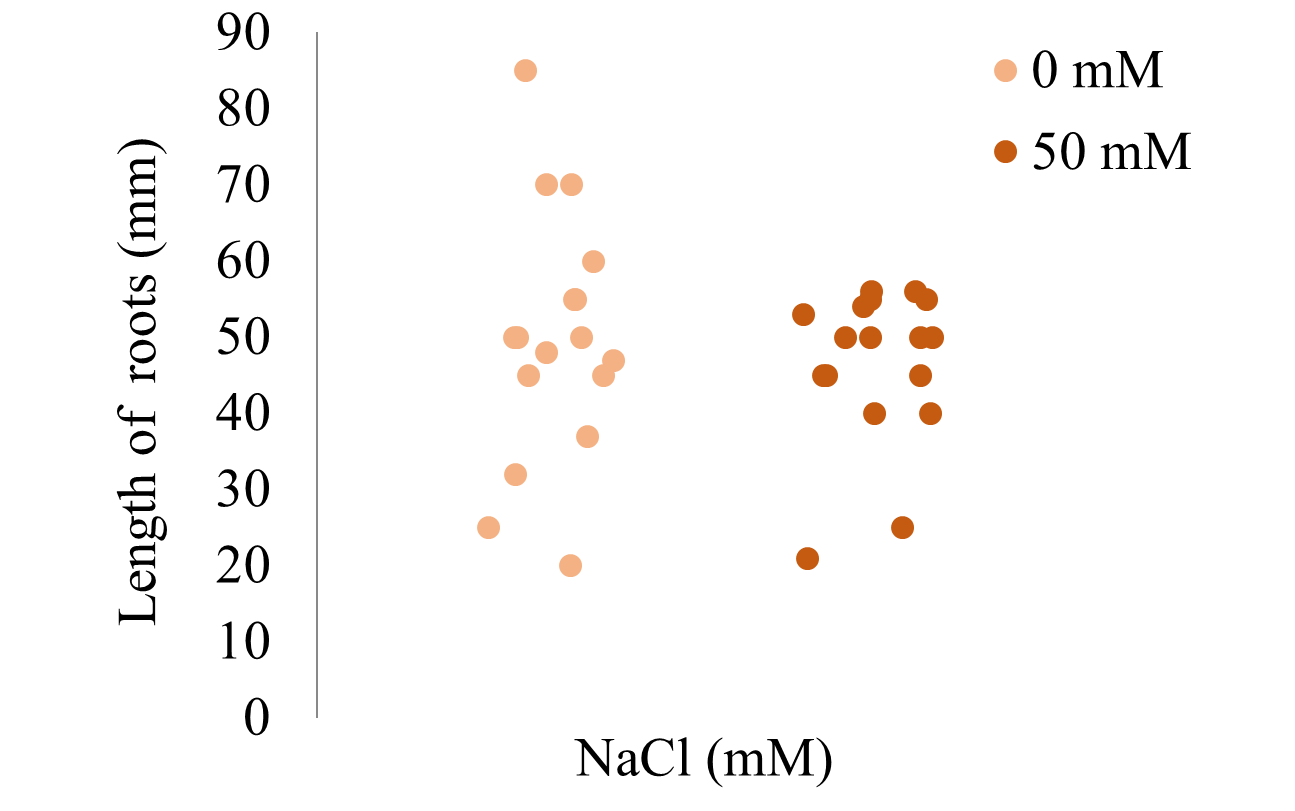
At 50 mM NaCl, the plants of S. rebaudiana developed the same length of roots as the control (Figure 5). However, simultaneously the growth inhibition of the aerial parts was observed (Figure 2), indicating an increase in the root/shoot ratio. This increase may be due to the effect of abscisic acid on root growth in adverse conditions in order to tolerate it (Taíz & Zeiger, 2006). The increase in the root/shoot ratio matches with studies carried out on corn and cotton, where a significant decrease in the growth of the aerial parts was observed at 75 mM NaCl, but root growth was not affected (Mühling & Läuchli, 2002).
Plants treated with 100 or 150 mM NaCl did not develop roots. This inhibition of root development in the plants could be due to the fact that drought stress induces root growth up to specific water potentials. However, once this threshold is surpassed, root development is inhibited. In studies with drought-tolerant species, such as sesame, a marked reduction in root length was observed at concentrations above 100 mM NaCl (Koca et al., 2007).
Fresh and weight of the roots of salt-stressed S. rebaudiana
According to the Mann–Whitney U test, the S. rebaudiana plants treated with 50 mM NaCl showed a significant reduction in the fresh weight of their roots compared to the control individuals (Figure 6). This result agrees with a study on Perilla frutescens, where the roots became thinner with the increase in NaCl concentrations, which was reflected in the decrease in their fresh weight (Zhang et al., 2012).
Figure 6
Fresh weight of S. rebaudiana roots at different concentrations of NaCl. The points indicate the fresh weight of the roots of S. rebaudiana after 25 days of growth in MS culture medium supplemented with 0 and 50 mM NaCl. They also represent the dispersion of the data between individuals that received the same treatment, Mann–Whitney U test (p < 0.05), Monte Carlo Permutation (p < 0.05) test.
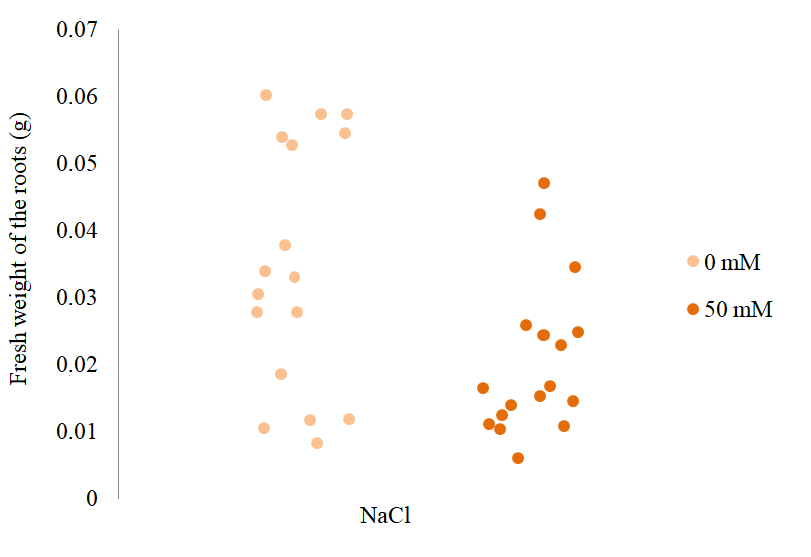
Although no significant differences were observed between the length of the roots of plants treated with 0 and 50 mM NaCl, a significant decrease was found in the fresh weight of the roots. It indicated a clear relationship between the reduction in the fresh weight of the roots and the increase in stress to which the plants were subjected. Unlike the fresh weight of the roots of S. rebaudiana, the statistical tests performed on the values of the dry weight of the roots treated with 0 or 50 mM NaCl (Figure 7) indicated no significant differences between them.
Figure 7
Effect on the dry weight of S. rebaudiana roots at different concentrations of NaCl. The points indicate the dry weight of the roots (g) per S. rebaudiana plant after 25 days of growth in MS culture medium supplemented with 0 and 50 mM NaCl. They also represent the dispersion of the data between individuals corresponding to the same treatment.
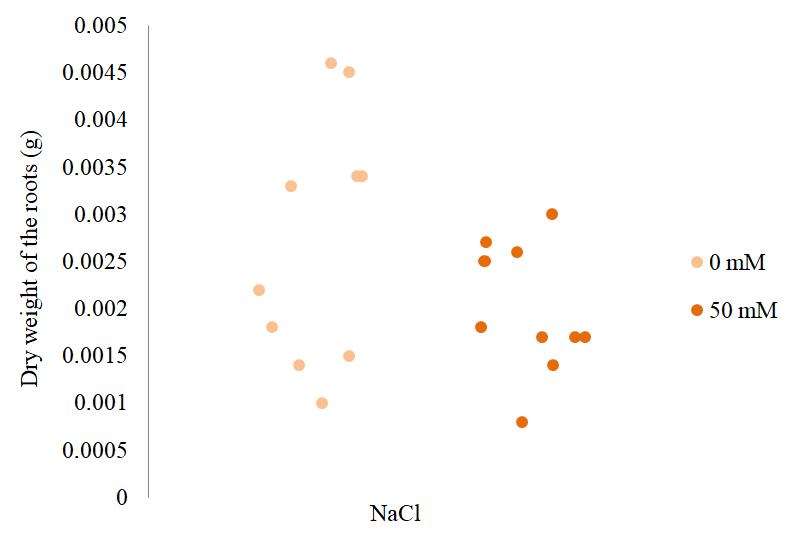
The difference observed between the statistical significance of the parameters in the fresh and dry weight of the roots could be due to the state of hydration of the roots (Valladares, 2004). It was considered that, in this test, the roots of S. rebaudiana treated with 0 mM NaCl had higher water content than those treated with 50 mM NaCl.
Chlorophyll concentration in salt-stressed S. rebaudiana
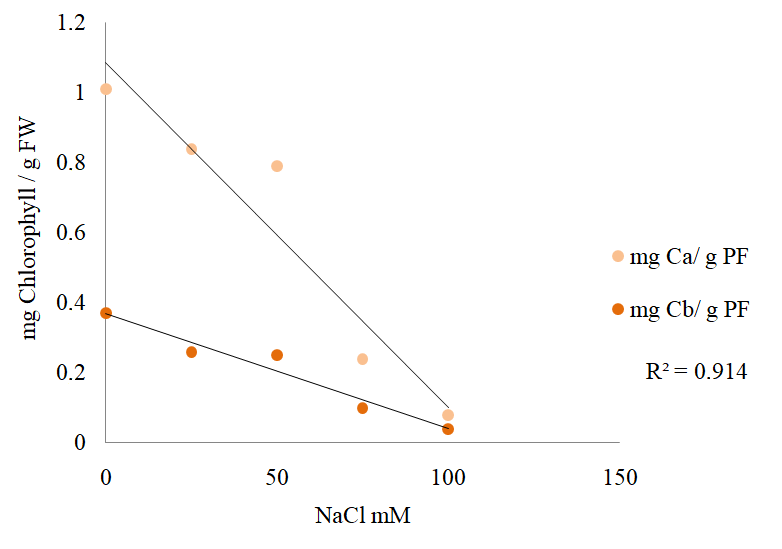
Multivariate linear regression performed on the concentrations of chlorophyll a (Chla) and chlorophyll b (Chlb) in the leaves of S. rebaudiana treated with 0, 50, 100, or 150 mM NaCl indicates a linear reduction of these values as the NaCl concentration in the culture medium increased, with R2 = 0.9149 (Figure 8).
Figure 8
Chlorophyll a and chlorophyll b concentrations in the leaves of S. rebaudiana after 25 days of growth in MS culture medium supplemented with 0, 50, 100, or 150 mM NaCl.
Despite the variation observed, the Student’s t-test performed on the content of Chl a and Chl bin the plants treated with 0 and 50 mM NaCl indicated that there was no significant variation in the content of chlorophyll between both groups (Figure 9). In a study conducted by Shahverdi et al. (2019), at the low concentration of NaCl (30 mM), the level of Chla and carotene id sin S. rebaudiana was not reduced, where as the level of all photosynthetic pigments significantly decreased under the influence of 60 mM NaCl.
Figure 9
Chlorophyll a and chlorophyll b concentrations (±SD) in the leaves of S. rebaudiana after 25 days of growth in MS culture medium supplemented with 0 and 50 mM NaCl.
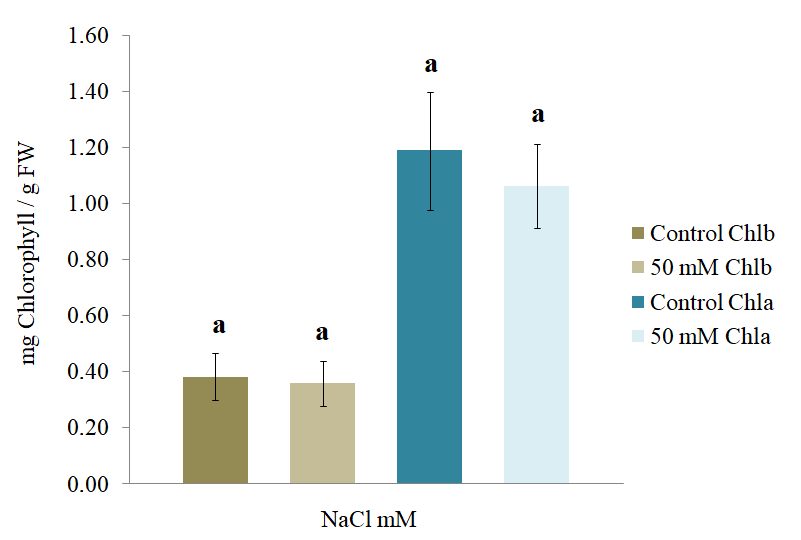
Since the reduction in the chlorophyll content in plants is mainly related to drought stress and this is considered a consequence of oxidative damage, it could be stated that the oxidative damage to the plant sat 50 mM NaCl was not significant (Alves da Costa et al., 2005; García et al., 2010), probably due to the efficiency of the enzymatic antioxidant system, whose purpose is the protection of the plant against oxidative stress (Alves da Costa et al., 2005).
Free proline accumulation in salt-stressed S. rebaudiana
Proline accumulation in stressed plants is one of the most important indicators of stress resistance (Sarmast et al., 2015). The statistical analysis performed on proline concentrations in the leaves of S. rebaudiana treated with 0 or 50 mM NaCl (Figure 10) indicates significant differences between both treatments, being higher in plants treated with 50 mM NaCl.
Figure 10
Proline concentration in the leaves of S. rebaudiana after 25 days of growth in MS culture medium supplemented with 0 or 50 mM NaCl. The whiskers represent the highest and lowest values of the mean proline concentration and the boxes represent the standard error, Student’s t test (p < 0.05).
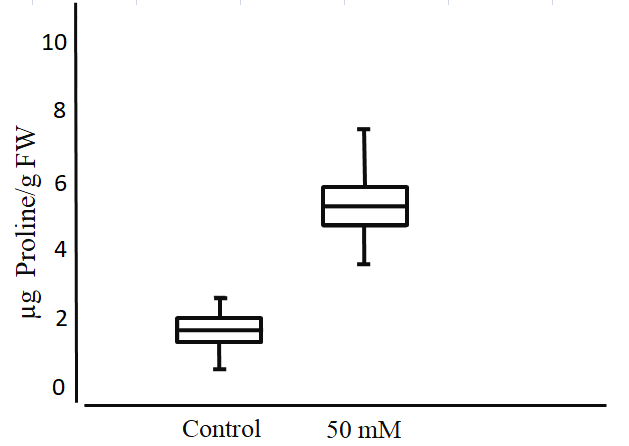
This amino acid is considered one of the main solutes increasing during stress (Díaz et al., 1999). The development of tolerance to drought and saline stress is related to the increase in the production of metabolites such as proline. For this reason, efforts are focused on obtaining transgenic plants capable of optimizing proline synthesis or inhibiting its degradation (Morales, 2006). Studies in transgenic tobacco (Nicotiana tabacum) affirm that the increase in the concentration of proline produced by plants subjected to drought or saline stress gives the plants the ability to grow even at 200 mM NaCl (Hong et al., 2000). In another study on the salt-stress response of S. rebaudiana, it was found that the proline level in salt-exposed plants (60–120 mM NaCl) was 17–42-fold higher than that in the control (Zeng et al., 2013).
Accumulation of soluble sugars in salt-stressed S. rebaudiana
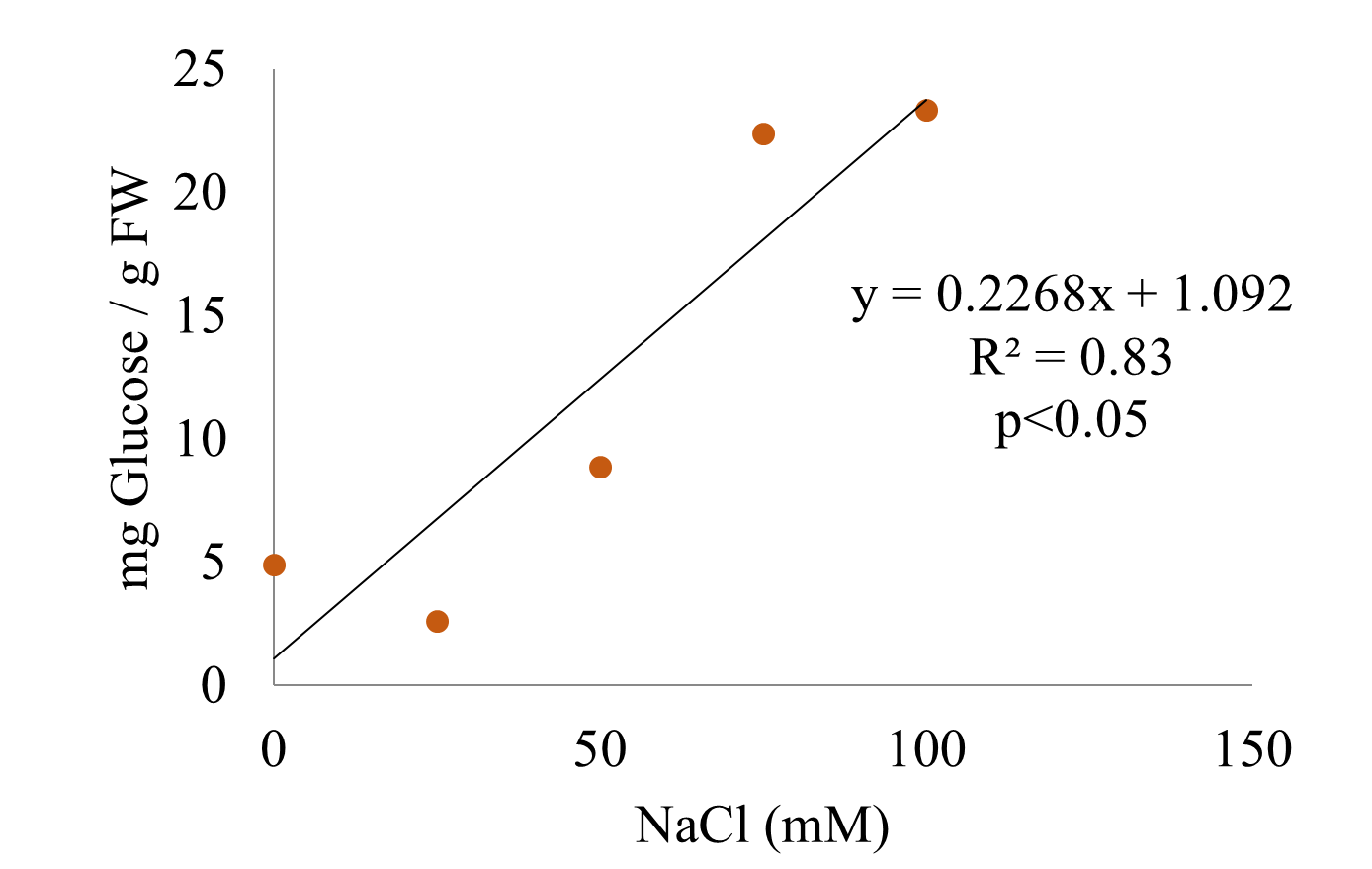
Soluble sugars, represented in this work by the glucose concentration, showed a tendency towards a linear increase with the increasing salt stress in the plants (Figure 11).
Figure 11
Glucose concentration in the leaves of S. rebaudiana after 25 days of growth in MS culture medium supplemented with 0, 25, 50, 75, or 100 mM NaCl.
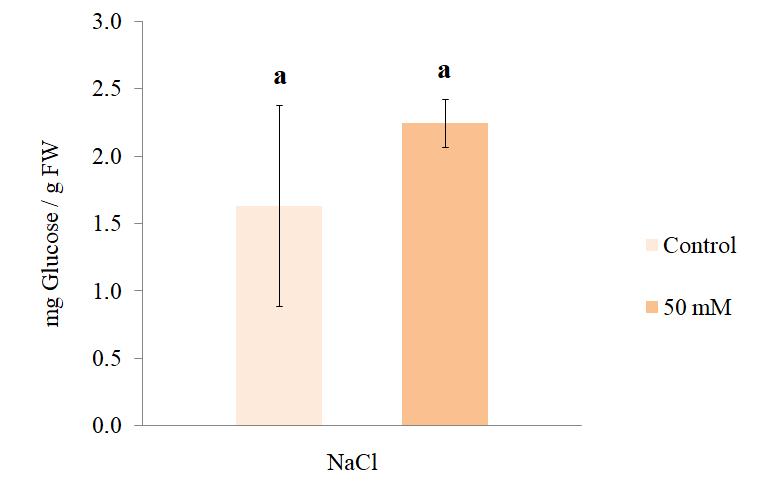
Regarding the concentration of soluble sugars in the plants treated with 0 or 50 mM NaCl (Figure 12), it was found that the means were not statistically different (1 tailed Student’s t p < 0.05). However, the total sugar content in this species exposed to 30–150 mM NaCl was first increased (at 30 or 60 mM) and then reduced (at 150 mM NaCl) (Shahverdi et al., 2019). In these experiments, the percentage of steviosides and rebaudioside. A also increased at 30 or 60 mM NaCl.
Figure 12
Soluble sugar content (±SD) in the leaves of S. rebaudiana plants after 25 days of growth in MS culture medium supplemented with 0 or 50 mM NaCl, Student’s t test at p < 0.05.
This test coincides with studies on Perilla frutescens that indicate that the content of soluble sugars in this species does not exhibit significant variations at concentrations lower than 150 mM NaCl (Zhang et al., 2012). Probably, it is necessary to surpass the NaCl concentration threshold to achieve a significant increase in soluble sugars. In recent studies, the deleterious effects of salinity on stevia have been demonstrated. In these studies, it was found that stevia plants were affected by different levels of salinity and this plant is relatively sensitive to salt stress (Forouzi et al., 2020). Despite this, some researchers consider that stevia is a relatively tolerant plant to salinity (Cantabella et al., 2017).
. Conclusions
Based on the obtained results, it can be concluded that the development of vitroplants of S. rebaudiana Bertoni var. Eirete in these experimental conditions is negatively affected as the concentration of NaCl increases. As indicated by the parameters analyzed in this study, at the concentration corresponding to 50 mM NaCl, stevia uses such strategies as a reduction of the growth of the aerial parts, an increase in the root/shoot ratio, and an increase in the free proline accumulation to cope with salt stress. It canals o be concluded that certain biochemical and growth responses that could contribute to the tolerance of the plant to adverse conditions, such as the increase in the sugar content and the dry weight of the roots, do not occur in the conditions tested, which may explain the relatively high sensitivity of this species to salinity. Further studies could be focused on improving the responses of S. rebaudiana to water deficiency and saline stress by activating tolerance mechanisms at the biochemical level, such as increasing the concentration of soluble sugars, or at the structural level, inducing the development of roots in order to avoid the reduction of crop yield.