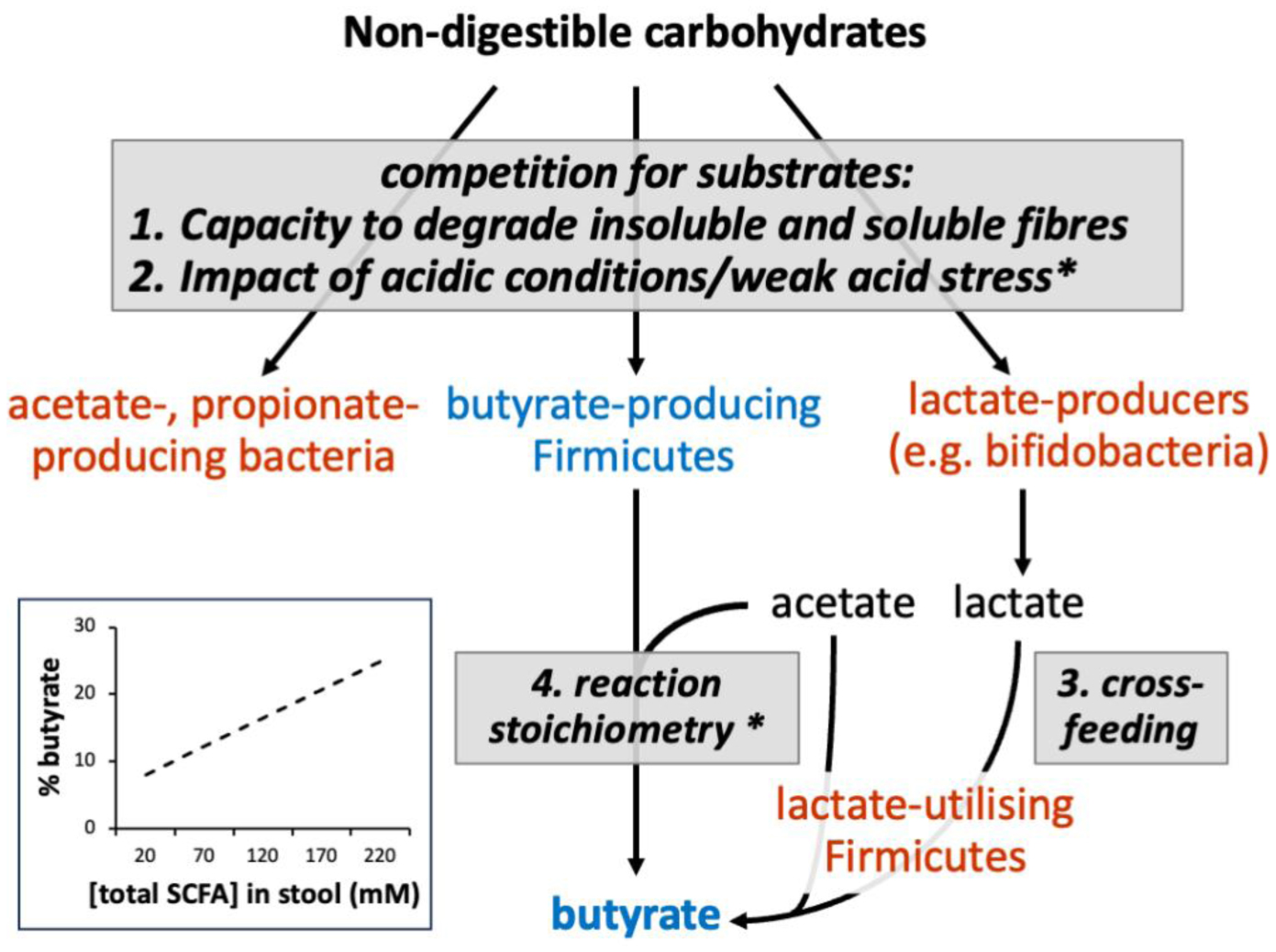
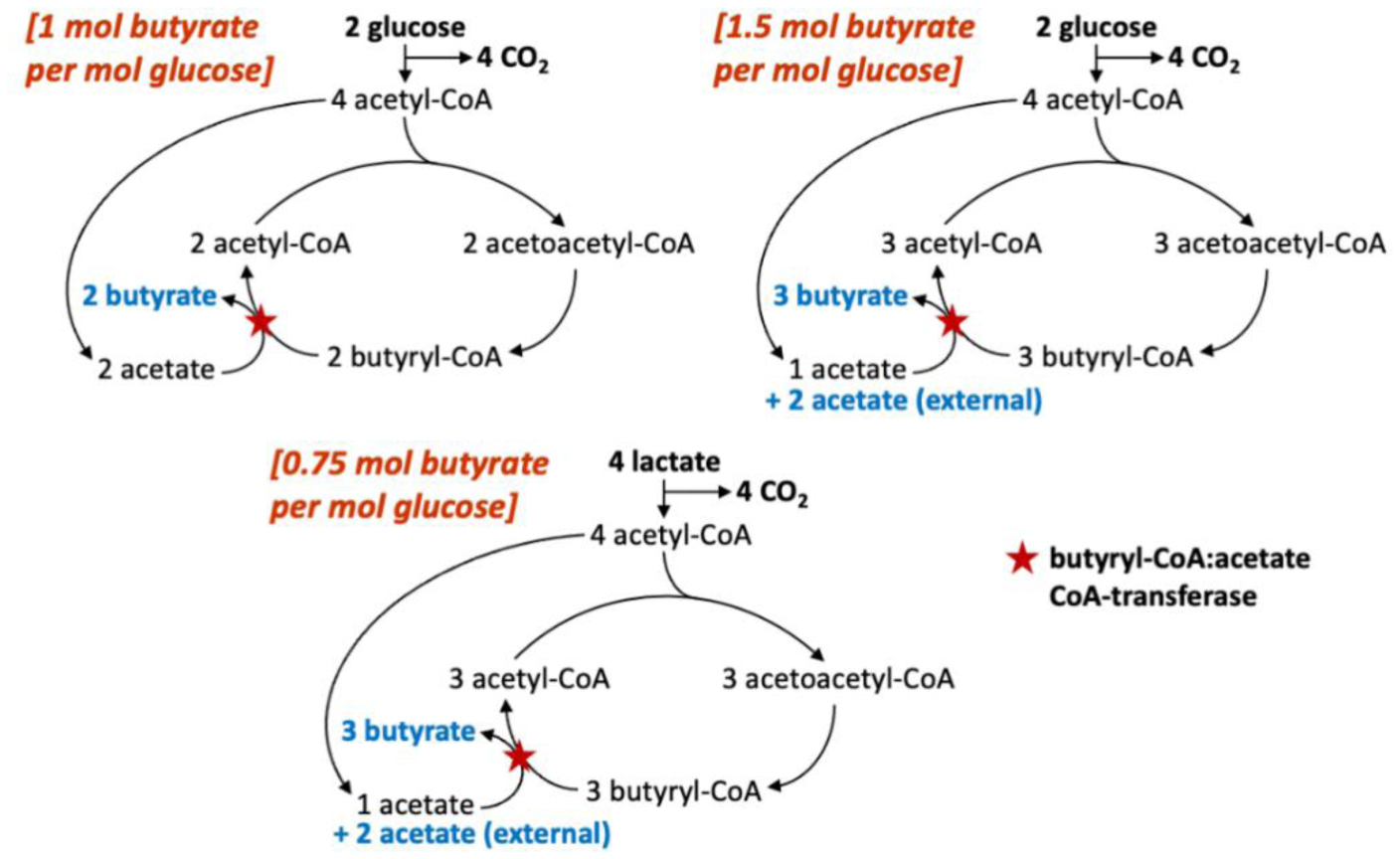
The microbial community of the human large intestine mainly ferments dietary fiber to short chain fatty acids (SCFAs), which are efficiently absorbed by the host. The three major SCFAs (acetate, propionate, and butyrate) have different fates within the body and different effects on health. A recent analysis of 10 human volunteer studies established that the proportions of these SCFA in fecal samples significantly shifted towards butyrate as the overall concentration of SCFA increased. Butyrate plays a key role in gut health and is preferentially utilized as an energy source by the colonic epithelium. Here we discuss possible mechanisms that underlie this ‘butyrate shift’; these include the selection for butyrate-producing bacteria within the microbiota by certain types of fiber, and the possibility of additional butyrate formation from lactate and acetate by metabolite cross-feeding. However, a crucial factor appears to be the pH in the proximal colon, which decreases as the SCFA concentrations increase. A mildly acidic pH has been shown to have an important impact on microbial competition and on the stoichiometry of butyrate production. Understanding these complex interactions has been greatly aided by the refinement of theoretical models of the colonic microbiota that assume a small number (10) of microbial functional groups (MFGs).
Citation: Harry J. Flint, Petra Louis, Sylvia H. Duncan. Why does increased microbial fermentation in the human colon shift toward butyrate?[J]. AIMS Microbiology, 2024, 10(2): 311-319. doi: 10.3934/microbiol.2024016
The microbial community of the human large intestine mainly ferments dietary fiber to short chain fatty acids (SCFAs), which are efficiently absorbed by the host. The three major SCFAs (acetate, propionate, and butyrate) have different fates within the body and different effects on health. A recent analysis of 10 human volunteer studies established that the proportions of these SCFA in fecal samples significantly shifted towards butyrate as the overall concentration of SCFA increased. Butyrate plays a key role in gut health and is preferentially utilized as an energy source by the colonic epithelium. Here we discuss possible mechanisms that underlie this ‘butyrate shift’; these include the selection for butyrate-producing bacteria within the microbiota by certain types of fiber, and the possibility of additional butyrate formation from lactate and acetate by metabolite cross-feeding. However, a crucial factor appears to be the pH in the proximal colon, which decreases as the SCFA concentrations increase. A mildly acidic pH has been shown to have an important impact on microbial competition and on the stoichiometry of butyrate production. Understanding these complex interactions has been greatly aided by the refinement of theoretical models of the colonic microbiota that assume a small number (10) of microbial functional groups (MFGs).

| [1] | Liu P, Wang Y, Yang G, et al. (2020) The role of short chain fatty acids in intestinal barrier function, inflammation, oxidative stress, and colonic carcinogenesis. Pharm Res 165: 1054200. https://doi.org/10.1016/j.phrs.2021.105420 |
| [2] |
LaBouyer M, Holtrop G, Horgan G, et al. (2022) Higher total faecal short-chain fatty acid concentrations correlate with increasing proportions of butyrate and decreasing proportions of branched-chain fatty acids across multiple human studies. Gut Microbiome 3: e2. https://doi.org/10.1017/gmb.2022.1 
|
| [3] |
Hamer HM, Jonkers D, Venema K, et al. (2008) Research article: the role of butyrate on colonic function. Aliment Pharm Therap 27: 104-119. https://doi.org/10.1111/j.1365-2036.2007.03562.x
|
| [4] | Macfarlane GT, Gibson GR, Cummings JH (1992) Comparison of fermentation reactions in different regions of the human colon. J Appl Bacteriol 72: 57-64. https://doi.org/10.1111/j.1365-2672.1992.tb04882.x |
| [5] |
Bown RL, Gibson JA, Sladen PJ, et al. (1974) Effects of lactulose and other laxatives on ileal and colonic pH as measured by radiotelemetry device. Gut 15: 999-1004. https://doi.org/10.1136/gut.15.12.999
|
| [6] |
Yamamura R, Inoue KM, Nishino K, et al. (2023) Intestinal and fecal pH in human health. Front Microbiomes 2: 1192316. https://doi.org/10.3389/frmbi.2023.1192316
|
| [7] |
Louis P, Flint HJ (2017) Formation of propionate and butyrate by the human colonic microbiota. Environ Microbiol 19: 29-41. https://doi.org/10.1111/1462-2920.13589
|
| [8] |
Flint HJ (2020) How gut microorganisms make use of available carbohydrates. Why Gut Microbes Matter.Springer Fascinating Life Sciences: 81-96. https://doi.org/10.1007/978-3-030-43246-1_7
|
| [9] |
Fu XD, Lin ZM, Zhu CL, et al. (2019) Nondigestible carbohydrates, butyrate and butyrate-producing bacteria. Crit Rev Food Sci Nutr 59: S130-S152. https://doi.org/10.1080/10408398.2018.1542587
|
| [10] |
Salonen A, Lahti L, Salojärvi J, et al. (2014) Impact of diet and individual variation on intestinal microbiota composition and fermentation products in obese men. ISME J 8: 2218-2230. https://doi.org/10.1038/ismej.2014.63
|
| [11] |
Duncan SH, Russell WR, Quartieri A, et al. (2016) Wheat bran promotes enrichment within the human colonic microbiota of butyrate-producing bacteria that release ferulic acid. Environ Microbiol 18: 2214-2225. https://doi.org/10.1111/1462-2920.13158
|
| [12] |
Duncan SH, Belenguer A, Holtrop G, et al. (2007) Reduced dietary intake of carbohydrate by obese subjects results in decreased concentrations of butyrate and butyrate-producing bacteria in feces. Appl Environ Microbiol 73: 1073-1078. https://doi.org/10.1128/AEM.02340-06
|
| [13] |
Kleessen B, Schwarz S, Boehm A, et al. (2007) Jerusalem artichoke and chicory inulin in bakery products affect faecal microbiota in healthy volunteers. Br J Nutr 98: 540-549. https://doi.org/10.1017/S0007114507730751
|
| [14] |
Ramaneda J, Stallard-Olivera E, Hoffert M, et al. (2023) Building a genome-based understanding of bacterial pH preferences. Sci Adv 9: eadf8998. https://doi.org/10.1126/sciadv.adf8998
|
| [15] |
Duncan SH, Louis P, Thomson JM, et al. (2009) The role of pH in determining the species composition of the human colonic microbiota. Environ Microbiol 11: 2112-2122. https://doi.org/10.1111/j.1462-2920.2009.01931.x
|
| [16] | Schwartz J, Schumaker K, Bremeyer S, et al. (2022) Bacterial battle against acidity. FEMS Microbiol Revs : 1-27. https://doi.org/10.1093/femsre/fuac037 |
| [17] |
Trcek J, Mira NP, Jarboe LR (2015) Adaptation and tolerance of bacteria against acetic acid. Appl Microbiol Biotechnol 99: 6215-6229. https://doi.org/10.1007/s00253-015-6762-3
|
| [18] |
Walker AW, Duncan SH, McWilliam Leitch E, et al. (2005) pH and peptide supply can radically alter bacterial populations and short chain fatty acid ratios within human colonic microbial communities. Appl Environ Microbiol 71: 3692-3700. https://doi.org/10.1128/AEM.71.7.3692-3700.2005
|
| [19] |
Chung WCF, Walker AW, Louis P, et al. (2016) Modulation of the human gut microbiota by dietary fibres occurs at the species level. BMC Biol 14: 3. https://doi.org/10.1186/s12915-015-0224-3
|
| [20] |
Wang SP, Rubio LA, Duncan SH, et al. (2020) Pivotal roles for pH, lactate, and lactate-utilizing bacteria in the stability of a human colonic microbial ecosystem. mSystems 5: e00645-20. https://doi.org/10.1128/mSystems.00645-20
|
| [21] |
Duncan SH, Louis P, Flint HJ (2004) Lactate-utilising bacteria from human feces that produce butyrate as a major fermentation product. Appl Environ Microbiol 70: 5810-5817. https://doi.org/10.1128/AEM.70.10.5810-5817.2004
|
| [22] |
Louis P, Duncan SH, Sheridan PO, et al. (2022) Microbial lactate utilisation and the stability of the gut microbiome. Gut Microbiome 3: e3. https://doi.org/10.1017/gmb.2022.3
|
| [23] |
Belenguer A, Duncan SH, Calder G, et al. (2006) Two routes of metabolic cross-feeding between bifidobacteria and butyrate-producing anaerobes from the human gut. Appl Environ Microbiol 72: 3593-3599. https://doi.org/10.1128/AEM.72.5.3593-3599.2006
|
| [24] |
Duncan SH, Zuur G, Lobley GE, et al. (2004) Contribution of acetate to butyrate formation by human faecal bacteria. Br J Nutr 91: 915-923. https://doi.org/10.1079/BJN20041150
|
| [25] |
Kettle H, Louis P, Holtrop G, et al. (2015) Modelling the emergent dynamics and major metabolites of the human colonic microbiota. Environ Microbiol 17: 1615-1630. https://doi.org/10.1111/1462-2920.12599
|
| [26] |
Reichardt N, Vollmer M, Holtrop G, et al. (2018) Specific substrate-driven changes in human faecal microbiota compostion contrasts with functional redundancy in short chain fatty acid production. ISME J 12: 610-622. https://doi.org/10.1038/ismej.2017.196
|
| [27] |
Campbell A, Gdanetz K, Schmidt AW, et al. (2023) H2 generated by fermentation in the human gut microbiome influences metabolism and competitive fitness of gut butyrate producers. Microbiome 11: 133. https://doi.org/10.1186/s40168-023-01565-3
|
| [28] | Abell GCJ, Conlon MA, McOrist AL (2009) Methanogenic archaea in adult human faecal samples are inversely related to butyrate concentrations. Microb Ecol Health Dis 18: 154-160. https://doi.org/10.1080/08910600601048969 |
| [29] |
Soto-Martin EC, Warnke I, Farquharson FM, et al. (2020) Vitamin biosynthesis by human gut butyrate-producing bacteria and cross-feeding in synthetic microbial communities. mBio 11: e00886-20. https://doi.org/10.1128/mBio.00886-20
|
| [30] |
Khan MT, Duncan SH, Stams AJM, et al. (2012) The gut anaerobe Faecalibacterium prausnitzii uses an extracellular electron shuttle to grow at oxic-anoxic interphases. ISME J 6: 1578-1585. https://doi.org/10.1038/ismej.2012.5
|
| [31] |
Colgan SP, Wang RX, Hall CHT, et al. (2023) Revisiting the ‘starved gut’ hypothesis in inflammatory bowel disease. Immunometabolism 5: e0016. https://doi.org/10.1097/IN9.0000000000000016
|
| [32] |
Kettle H, Louis P, Flint HJ (2022) Process-based modelling of microbial community dynamics in the human colon. J Royal Soc Interface 19: 20220489. https://doi.org/10.1098/rsif.2022.0489
|
| [33] |
Gawenis LR, Bradford EM, Prasad V, et al. (2007) Colonic anion secretory defects and metabolic acidosis in mice lacking NBC1 Na+/HCO3− cotransporter. J Biol Chem 283: 9042-52. https://doi.org/10.1074/jbc.M607041200
|
| [34] |
Roediger WEW (1982) Utilization of nutrients by isolated epithelial cells of the rat colon. Gastroenterology 83: 424-429. https://doi.org/10.1016/S0016-5085(82)80339-9
|
| [35] |
Louis P, Hold GL, Flint HJ (2014) The gut microbiota, bacterial metabolites and colorectal cancer. Nat Rev Microbiol 12: 661-672. https://doi.org/10.1038/nrmicro3344
|
| [36] |
Roediger WEW (1980) The colonic epithelium in ulcerative colitis: an energy deficiency disease?. Lancet 116: 712-715. https://doi.org/10.1016/S0140-6736(80)91934-0
|
| [37] |
Manichanh C, Borruel N, Cassillas F, et al. (2012) The gut microbiota in IBD. Nat Rev Gastroenterol Hepatol 9: 599-608. https://doi.org/10.1038/nrgastro.2012.152
|
| [38] |
Hove H, Nordgaard-Andersen I, Mortensen PB (1994) Fecal DL-lactate concentration in 100 gastrointestinal patients. Scand J Gastroenterol 29: 255-259. https://doi.org/10.3109/00365529409090473
|
| [39] |
Dear KLE, Elia M, Hunter JO (2005) Do interventions which reduce colonic fermentation improve symptoms of irritable bowel syndrome?. Dig Dis Sci 50: 758-766. https://doi.org/10.1007/s10620-005-2570-4
|
Figures(2)

Harry J. Flint, Petra Louis, Sylvia H. Duncan. Why does increased microbial fermentation in the human colon shift toward butyrate?[J]. AIMS Microbiology, 2024, 10(2): 311-319. doi: 10.3934/microbiol.2024016







 DownLoad:
DownLoad: