
Spatio-Temporal Dynamics of crAssphage and Bacterial Communities in an Algerian Watershed Impacted by Fecal Pollution

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
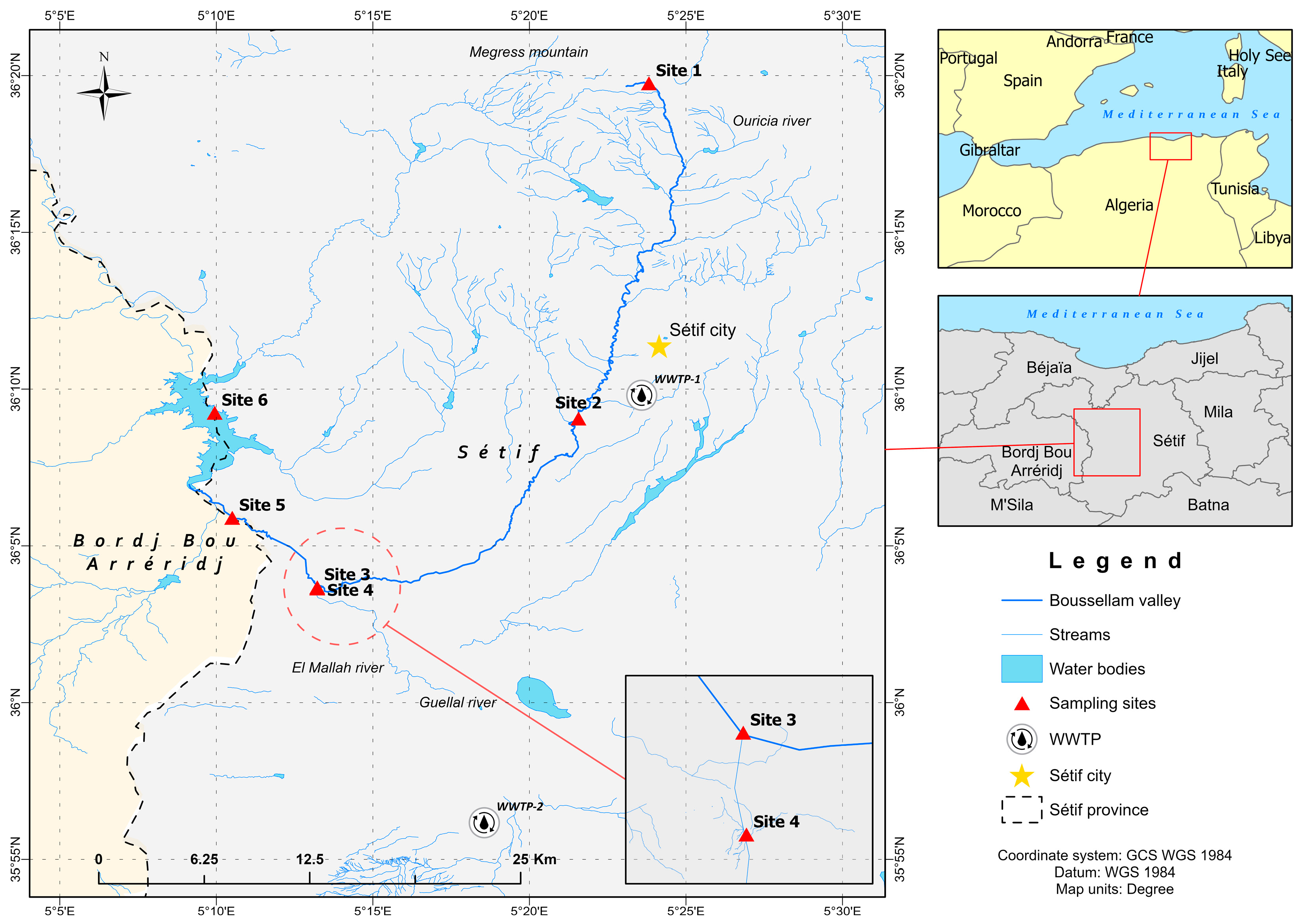
2.1. Study Area and Sample Collection
2.2. Chemical Characterization of the Water
2.3. Enumeration of Fecal Microbial Indicators Using Culture Methods
2.4. DNA Extraction
2.5. Quantification of crAssphage via qPCR
2.6. Bacterial Community Characterization
2.7. Statistical Analyses
3. Results and Discussion
3.1. Physicochemical Parameters
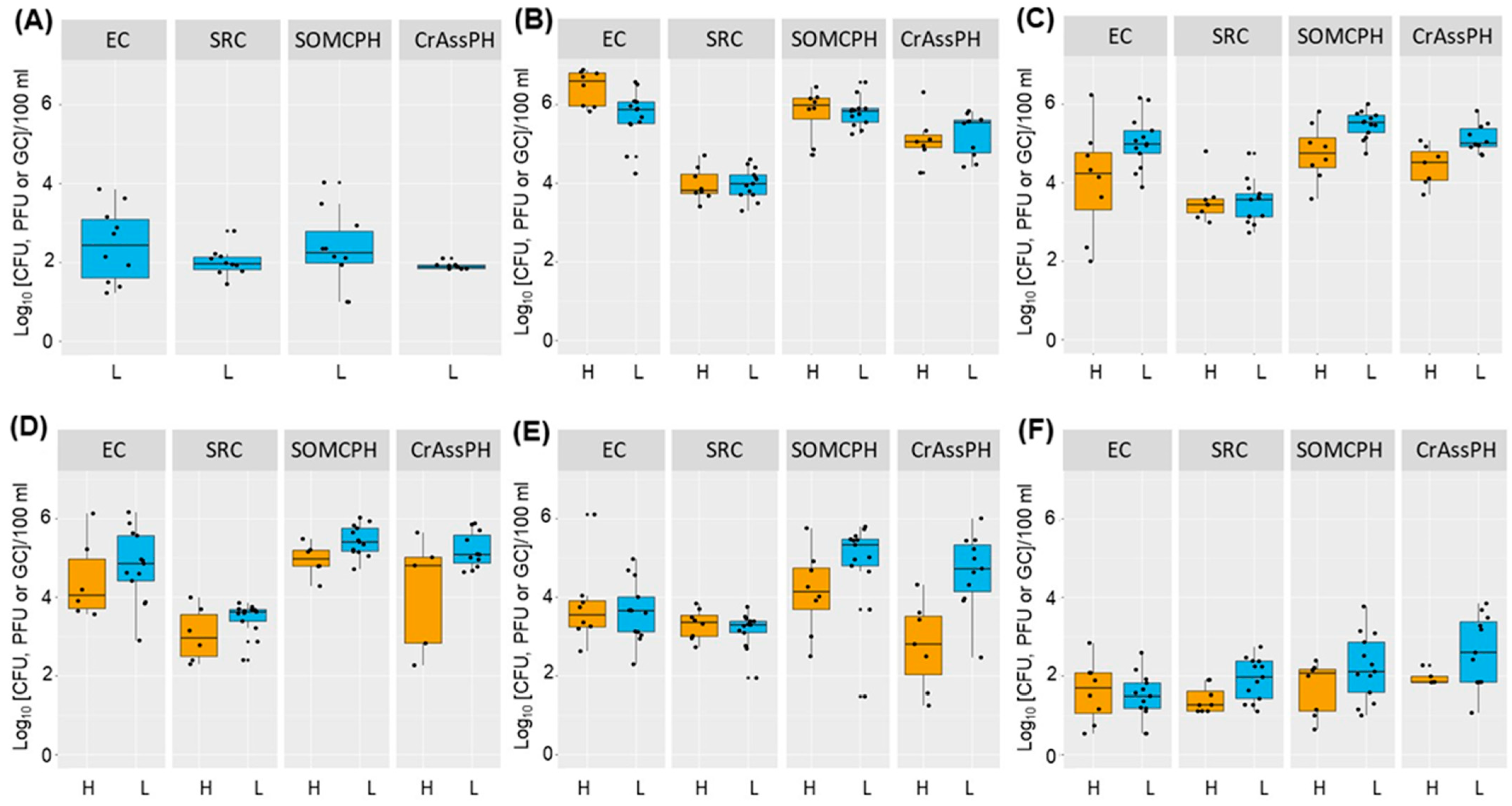
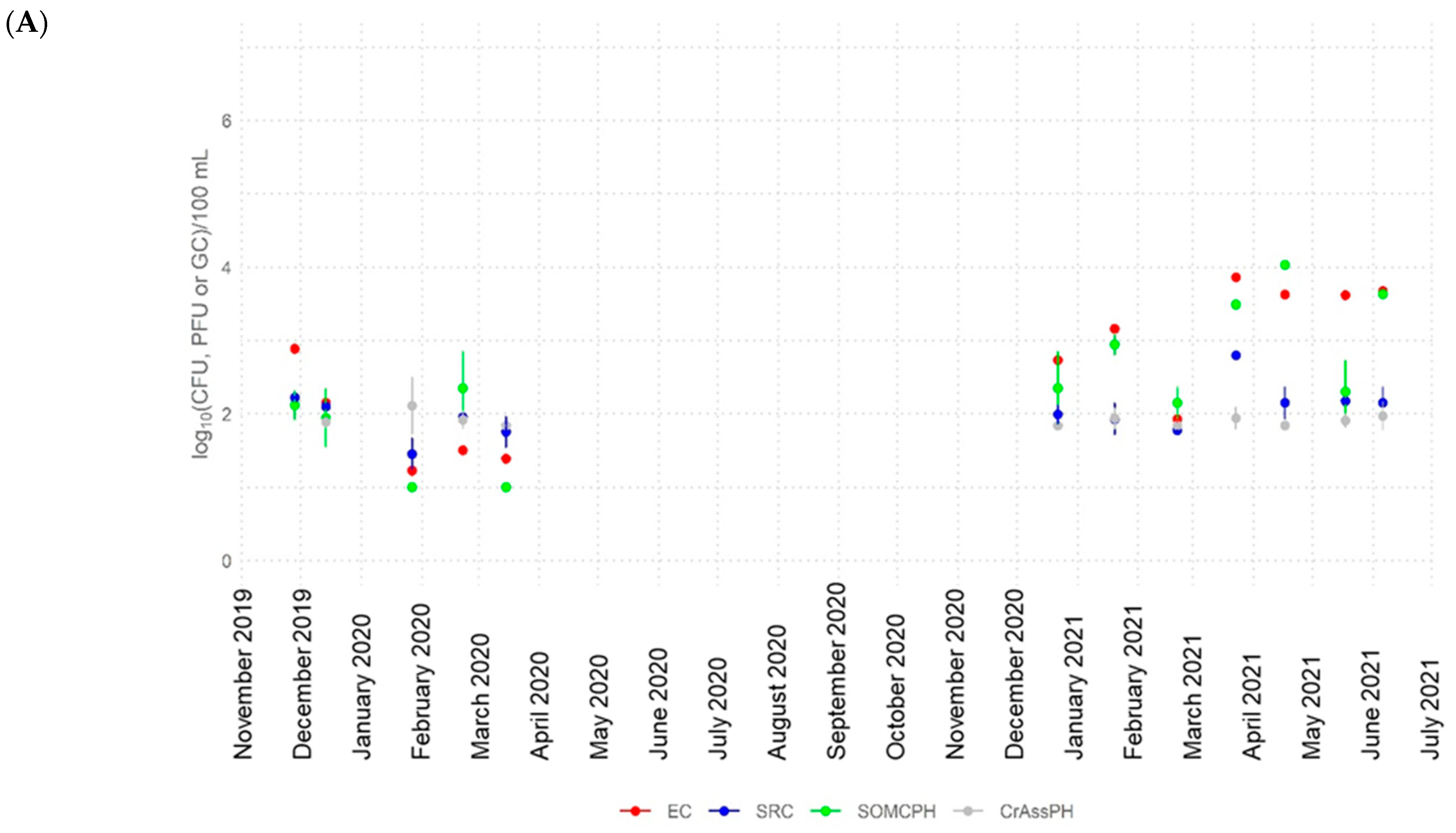
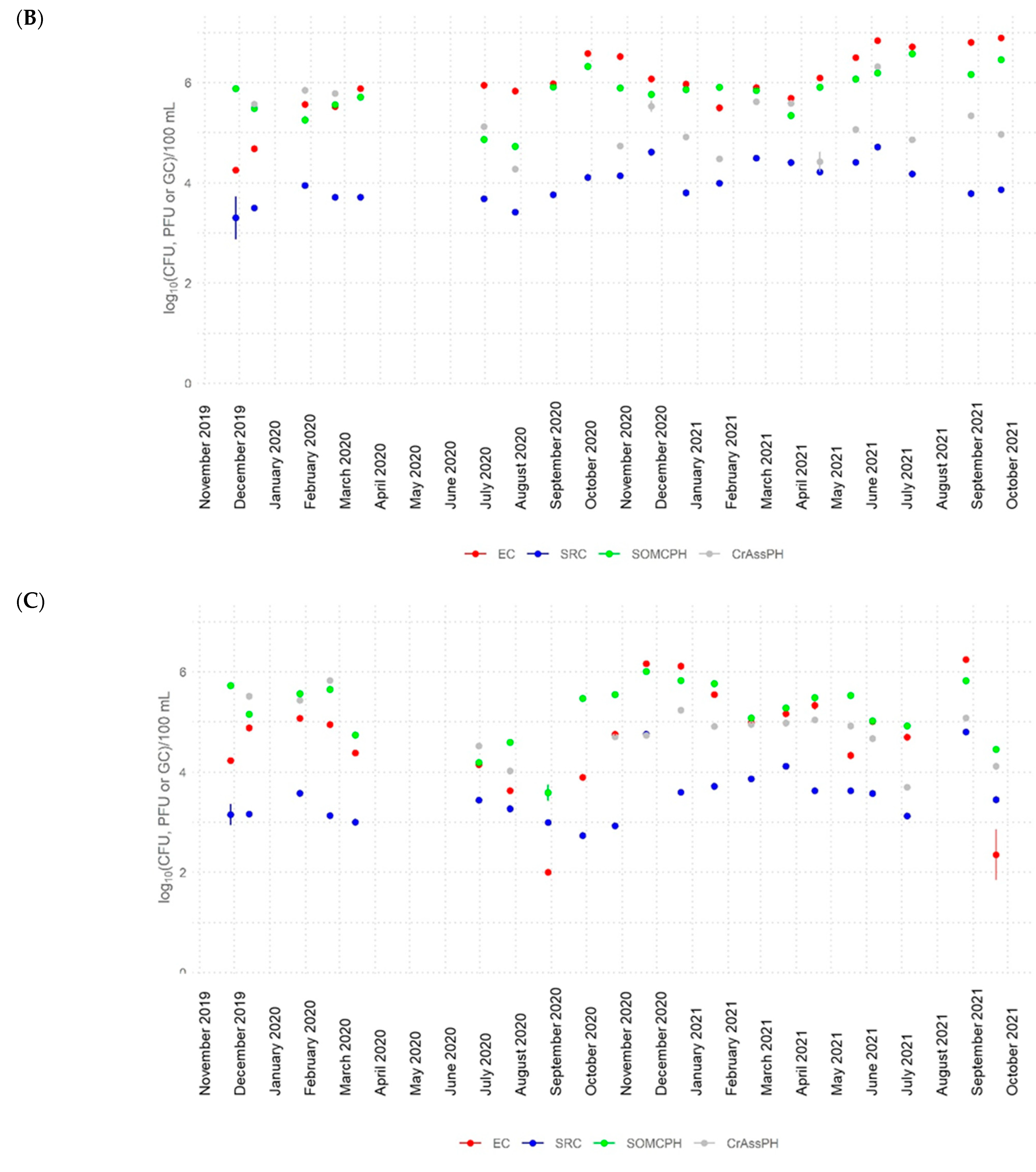
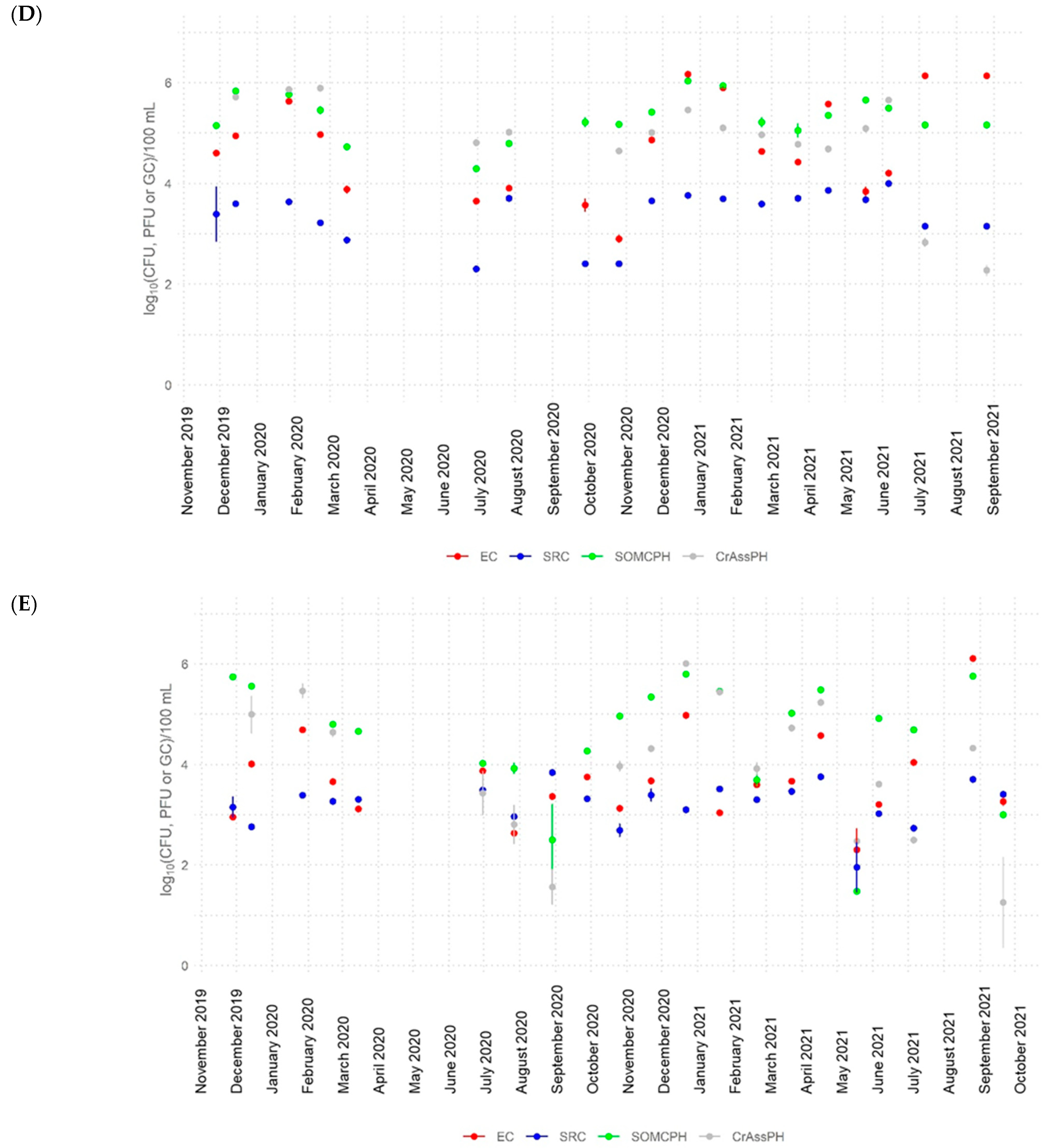
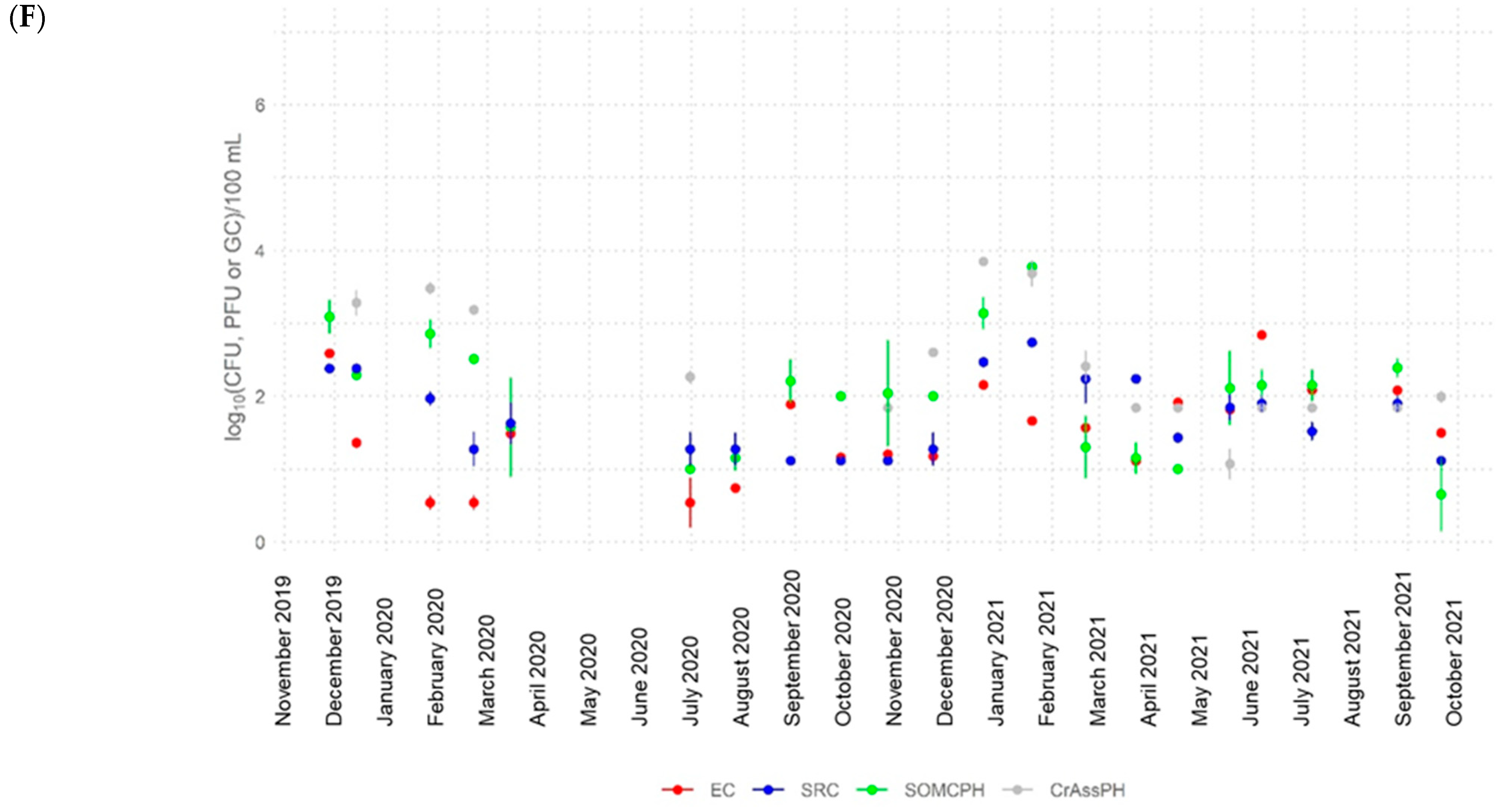
3.2. Dynamics of Fecal Indicators along the River
3.3. Correlation between the Fecal Indicators and CrAssPH
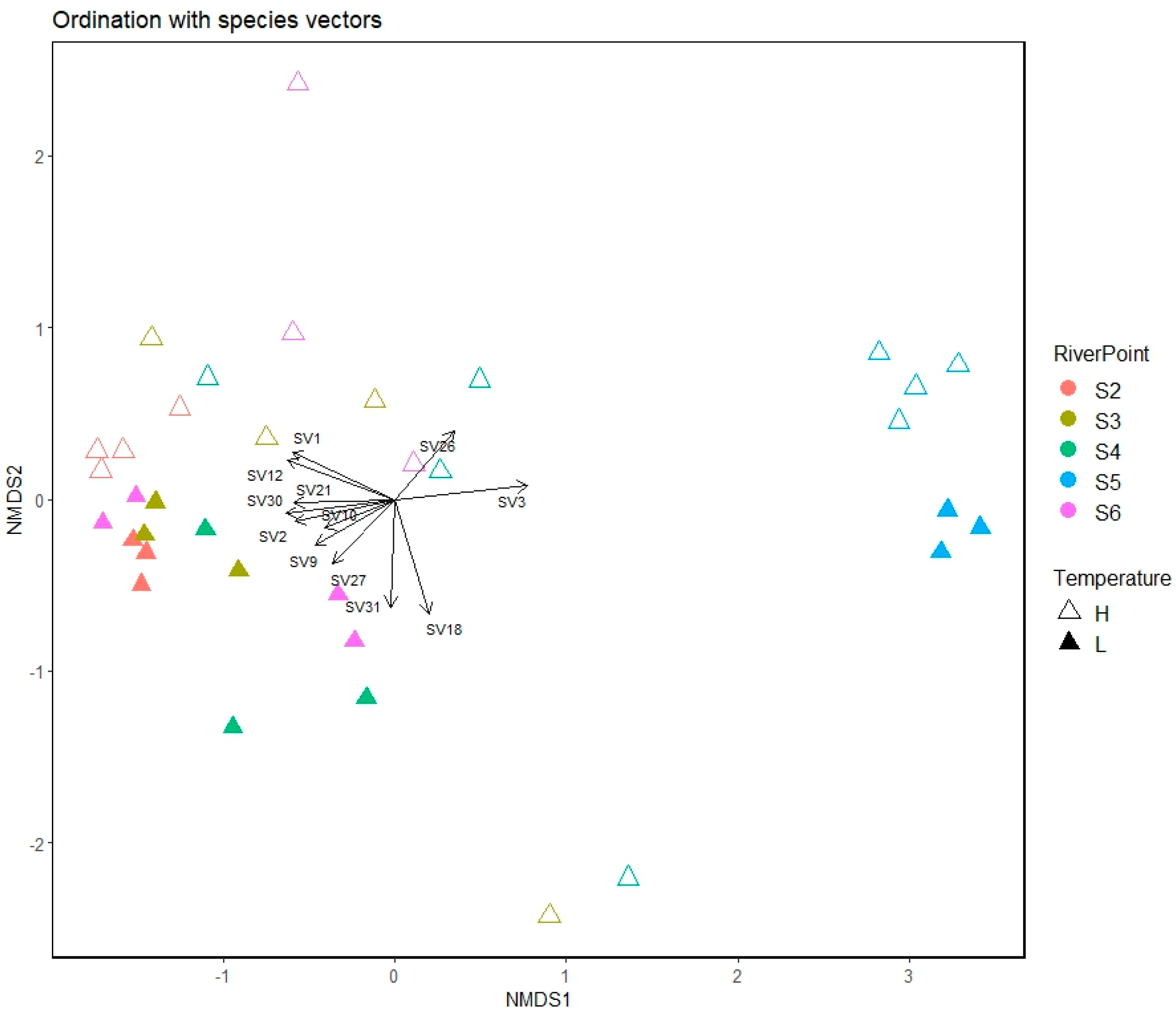
3.4. Dynamics of Bacterial Communities along the River
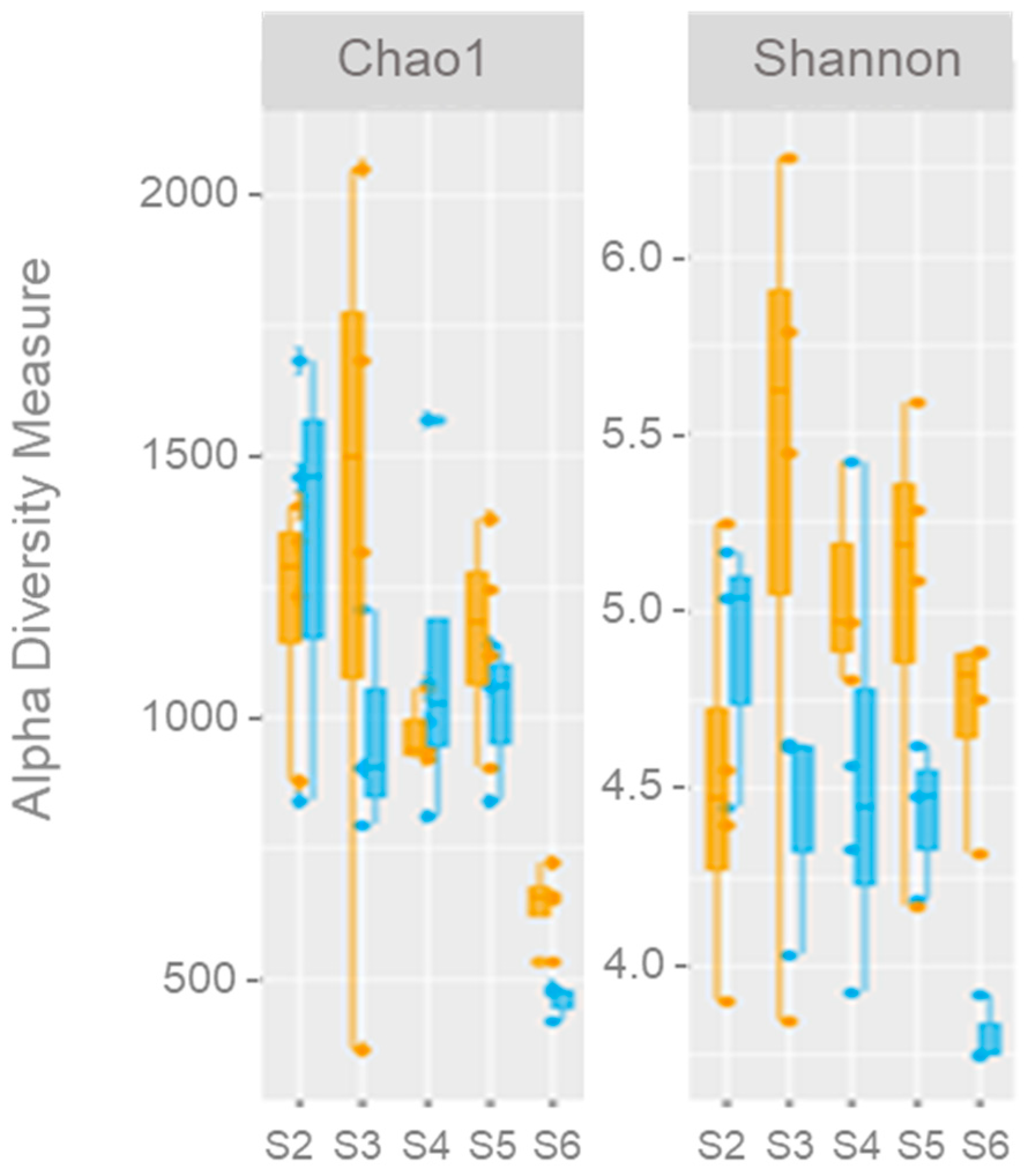
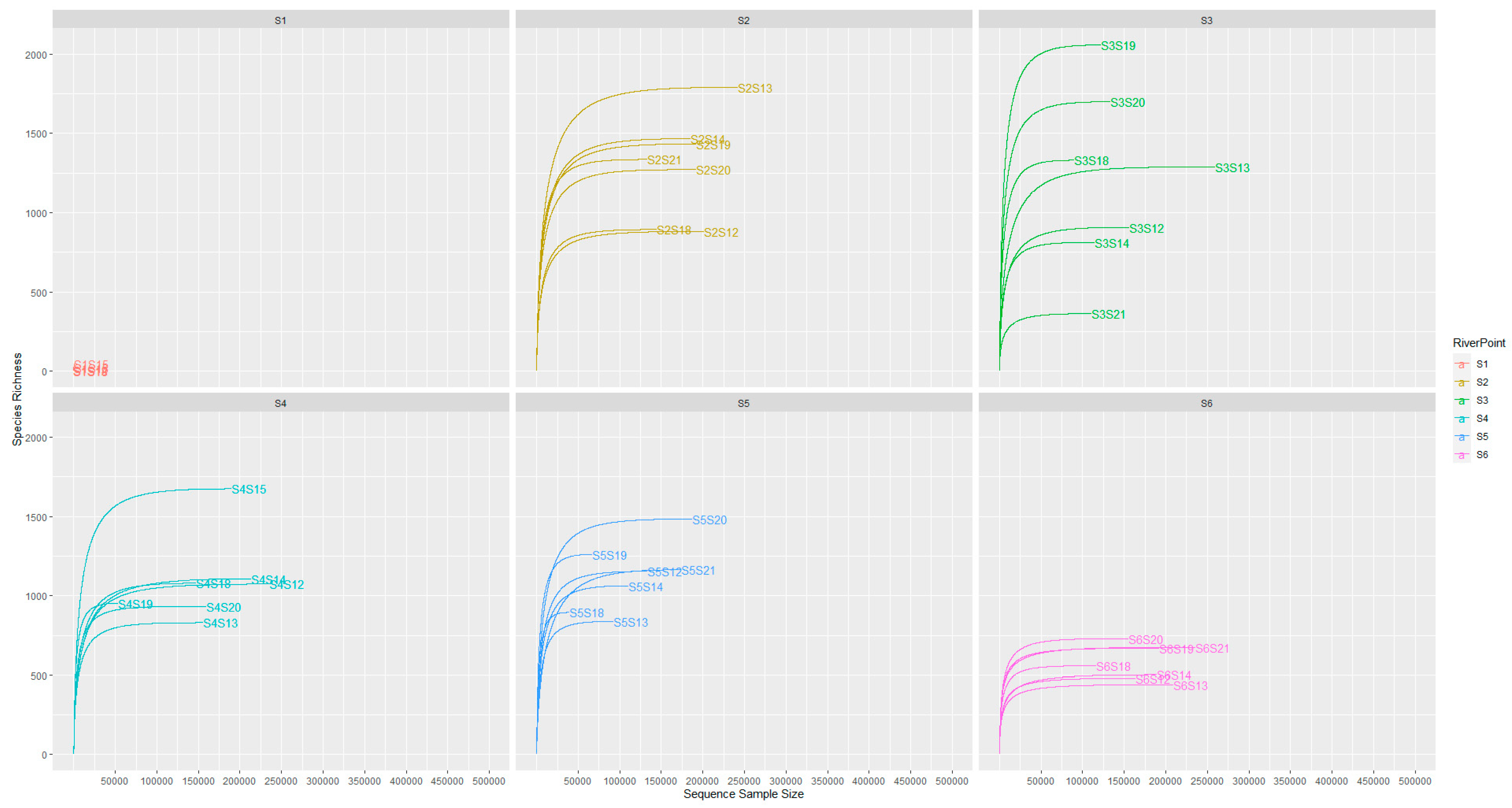
3.4.1. General Microbial Diversity
3.4.2. Bacterial Diversity
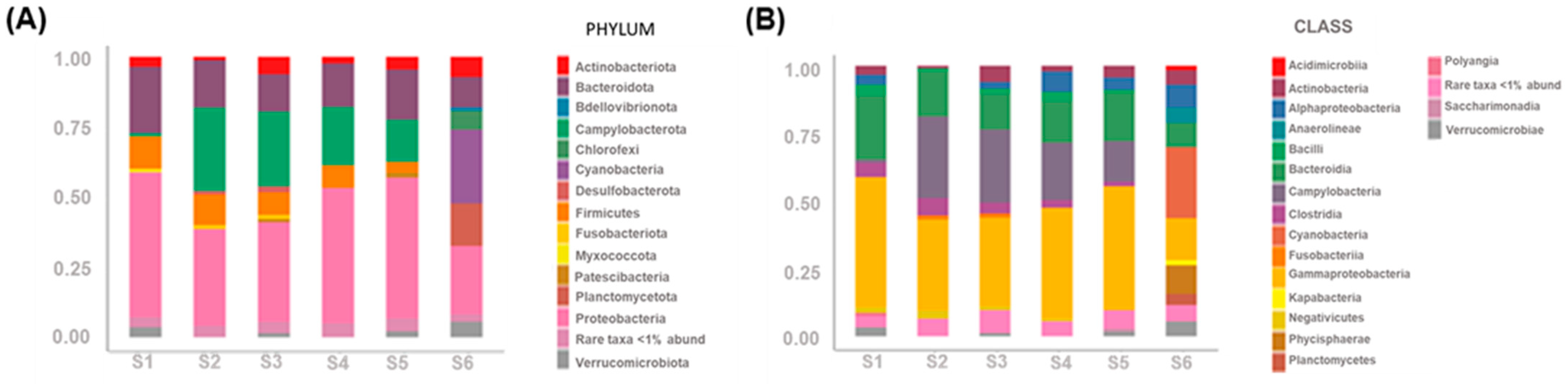
3.4.3. Bacterial Communities’ Taxonomy
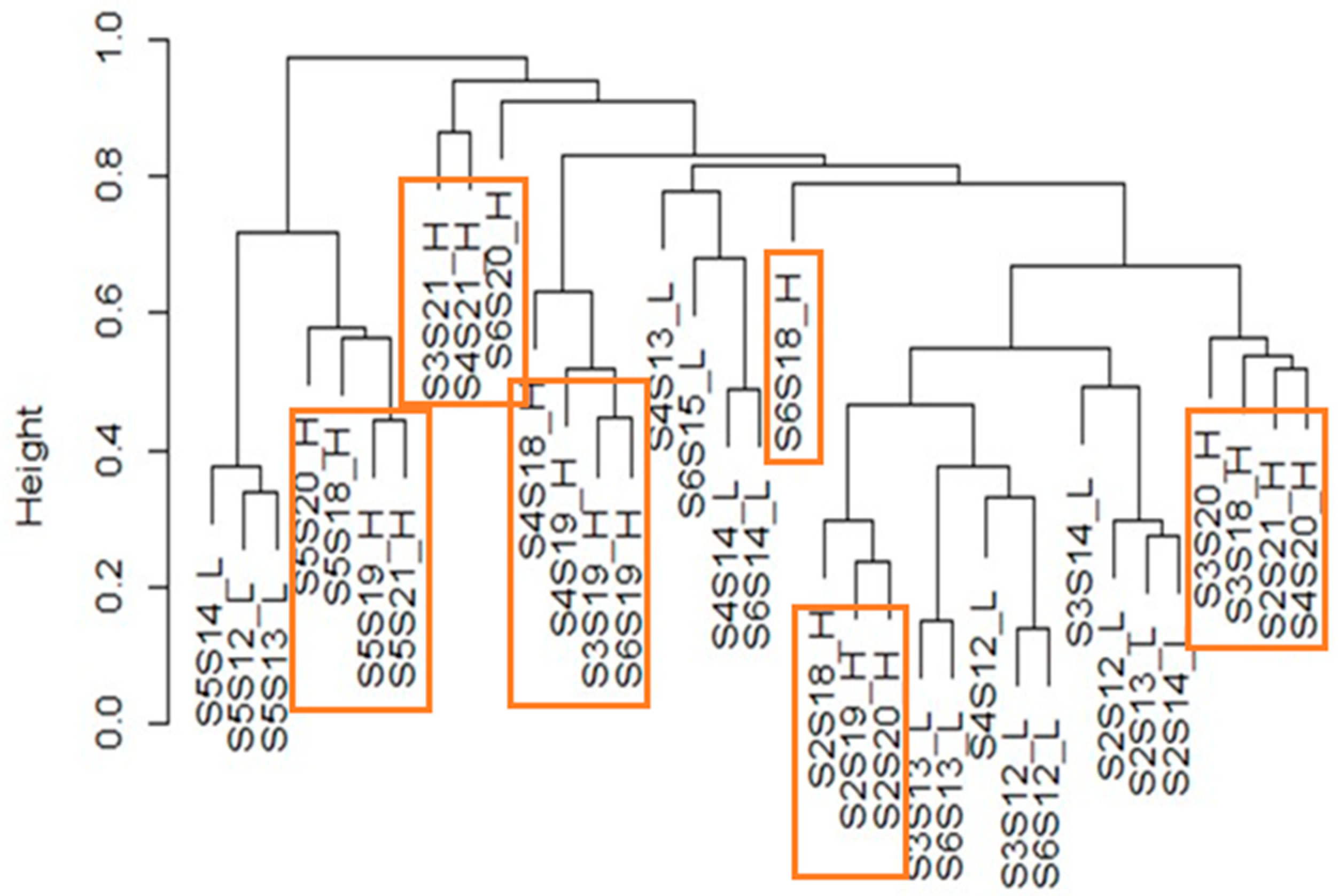
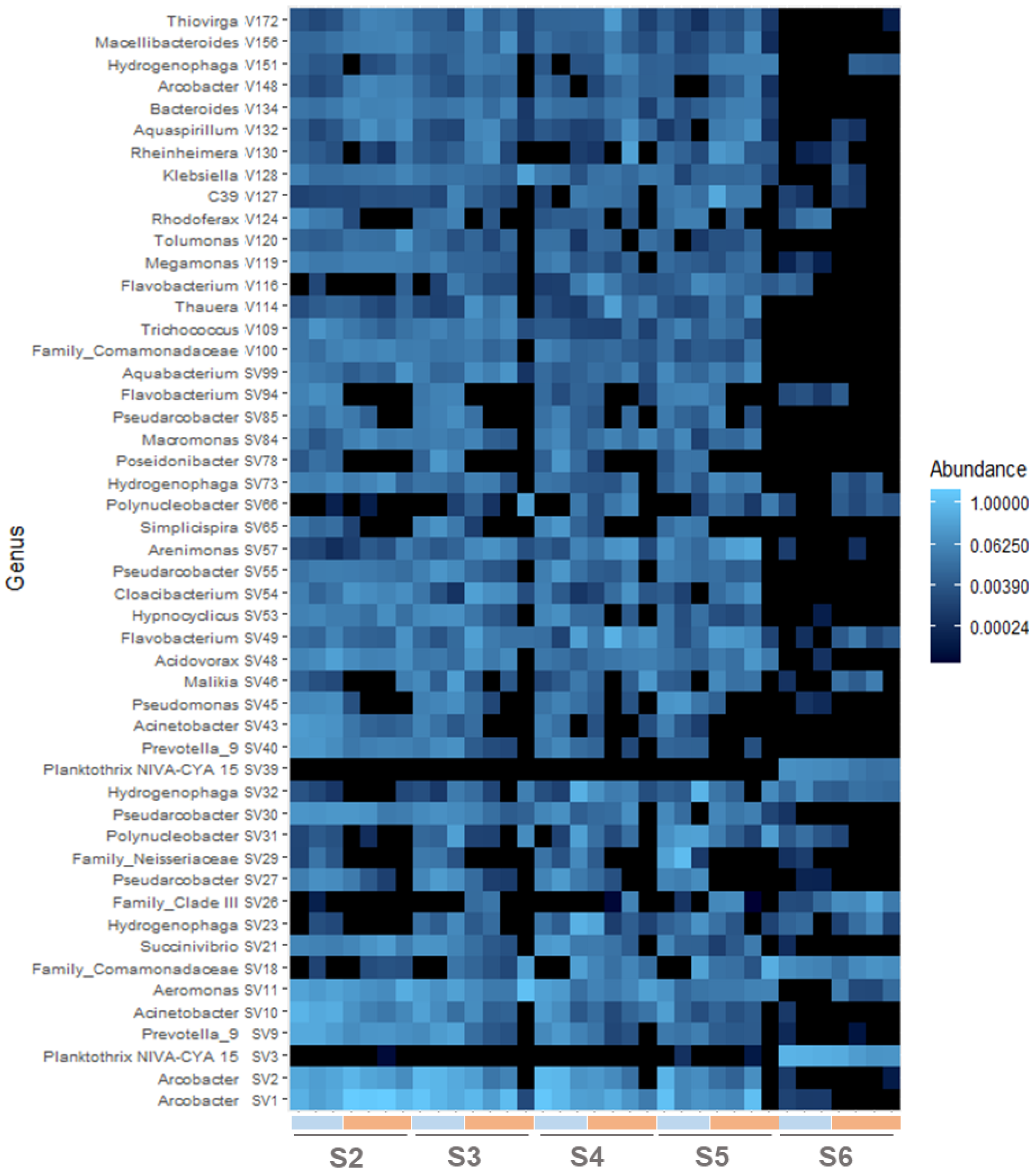
3.4.4. Tracking the ASVs along the Water Course
4. Conclusions
- This study presented data on the prevalence and abundance of crAssphage in environmental waters in Algeria for the first time.
- CrAssphage was frequently detected in river water at all sites (>5 log10 GC/100 mL at the most polluted site) and in the water reservoir at similar abundance values to other fecal indicators, showing strong correlations with somatic coliphages in the river (rho up to 0.88), with the signal fluctuating to lower levels along the river. These findings support the application of crAssphage as a human fecal pollution indicator for aged pollution in Algeria because of its high concentrations and persistence in the environment.
- The high-temperature season produced a higher decrease in the numbers of the fecal indicators (>4 log10 for E. coli and somatic coliphages and >3 log10 for crAssphage). During this season, the bacterial communities in the river also varied, becoming more diverse.
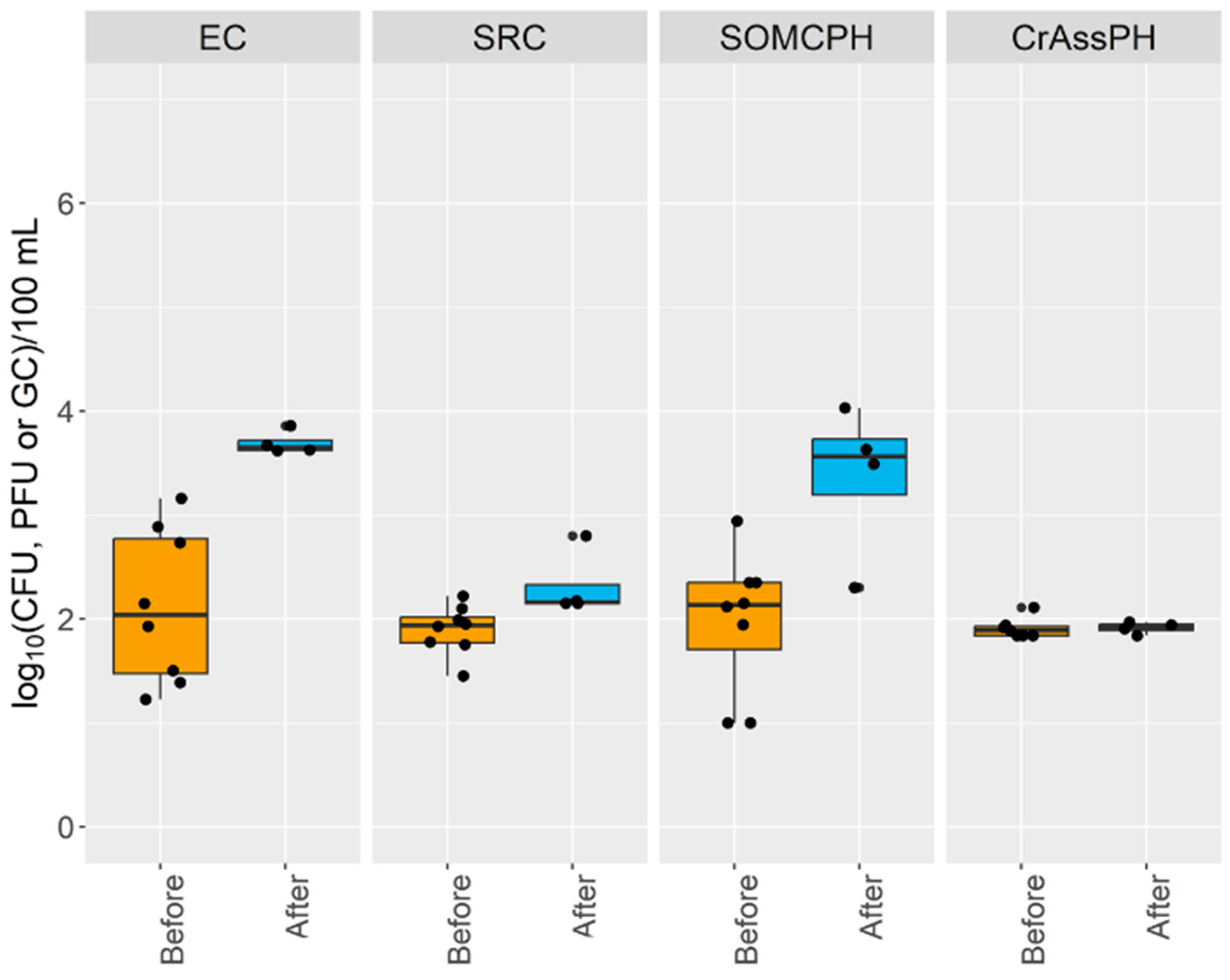
- The self-depuration capacity of the river reduced the impact of municipal wastewater from a microbiological point of view along its water course, as evidenced by the decrease in the number of fecal indicators, as well as crAssphage levels, to almost undetectable levels in the water reservoir. This self-purification was more pronounced during the high-temperature season, except for E. coli at site 5, where possible regrowth or contamination with animal fecal matter was detected.
- The high bacterial diversity values during the high-temperature period suggest a capacity for adaptation and resilience among microbial populations to disturbances, which should be considered when addressing anthropogenic impacts within the context of global warming.
- In summary, this study provides valuable insights into the complex interplay between anthropogenic activities, wastewater discharge, and microbial pollution in a highly urban-impacted watershed in Algeria, where limited information is available. The irrigation of crops with the river water, especially in the near highly populated areas, as a common practice in the region, poses a risk for the transmission of fecal-oral pathogens to the population. The continued monitoring and implementation of appropriate management measures are essential to safeguard water quality and public health in these vulnerable environments.
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sampling Campaigns | Sampling Dates | Season | Rain | Rain B1 | T Min | T Max | Humidity | Humidity B1 |
|---|---|---|---|---|---|---|---|---|
| 1 | 28 November 2019 | A | 0 | 0 | 6 | 15 | 81 | 74 |
| 2 | 14 December 2019 | W | 0 | 0 | 6.1 | 15 | 75 | 73 |
| 3 | 27 January 2020 | W | 2 | 0.3 | 1 | 10.2 | 73 | 82 |
| 4 | 22 February 2020 | W | 0 | 0 | −0.4 | 16 | 39 | 29 |
| 5 | 15 March 2020 | SP | 0 | 0 | 5.3 | 19 | 67 | 67 |
| 6 | 30 June 2020 | S | 0 | 0 | 19.7 | 36.2 | 19 | 22 |
| 7 | 27 July 2020 | S | 0 | 0 | 24 | 38.2 | 24 | 19 |
| 8 | 29 August 2020 | S | 0 | 0 | 22.4 | 33.2 | 21 | 30 |
| 9 | 28 September 2020 | A | 0 | 0 | 5.9 | 22 | 33 | 35 |
| 10 | 26 October 2020 | A | 0 | 0 | 5.3 | 19 | 52 | 49 |
| 11 | 22 November 2020 | A | 3 | 9.9 | 5 | 9 | 91 | 85 |
| 12 | 22 December 2020 | W | 5.3 | 15.6 | 2 | 12 | 86 | 88 |
| 13 | 20 January 2021 | W | 0 | 0 | −1.4 | 12 | 60 | 43 |
| 14 | 21 February 2021 | W | 0 | 0 | 4.1 | 17.8 | 38 | 34 |
| 15 | 23 March 2021 | SP | 5.1 | 4.1 | 1 | 8.5 | 85 | 78 |
| 16 | 17 April 2021 | SP | 3.1 | 1 | 4.6 | 10 | 81 | 70 |
| 17 | 18 May 2021 | SP | 0 | 0 | 13.9 | 28.3 | 36 | 43 |
| 18 | 06 June 2021 | S | 1 | 0 | 15 | 23 | 46 | 61 |
| 19 | 06 July 2021 | S | 0 | 0 | 23.5 | 40 | 15 | 17 |
| 20 | 26 August 2021 | S | 3.1 | 7.1 | 17 | 31 | 42 | 48 |
| 21 | 21 September 2021 | A | 1 | 28.1 | 15 | 25 | 59 | 69 |
| pH | C (µS/CM) | T (NTU) | PO43− (mg/mL) | NH4+ (mg/mL) | NO2− (mg/mL) | OM (mg/L) | |
|---|---|---|---|---|---|---|---|
| Site 1 | 8.1 ± 0.3 | 953 ± 119 | 8.3 ± 10.5 | 1.6 ± 1.6 | 2.3 ± 3.6 | 0.2 ± 0.1 | 12.2 ± 6.1 |
| Site 2 | 7.7 ± 0.3 | 1398 ± 224 | 38.0 ± 16.5 | 4.1 ± 2.3 | 6.9 ± 3.7 | 0.3 ± 0.2 | 19.7 ± 10.3 |
| Site 3 | 7.9 ± 0.3 | 2275 ± 769 | 37.2 ± 9.5 | 4.3 ± 1.8 | 6.7 ± 2.5 | 1.1 ± 1.3 | 26.0 ± 21.1 |
| Site 4 | 8.1 ± 0.5 | 3284 ± 1208 | 31.9 ± 20.7 | 5.3 ± 2.1 | 5.3 ± 2.1 | 1.1 ± 1.8 | 23.3 ± 19.4 |
| Site 5 | 7.5 ± 0.6 | 3165 ± 1691 | 41.0 ± 46.8 | 2.0 ± 1.4 | 4.1 ± 3.4 | 0.6 ± 1.4 | 16.0 ± 10.7 |
| Site 6 | 7.5 ± 0.6 | 1621 ± 73 | 32.2 ± 13.1 | 1.2 ± 1.2 | 2.4 ± 1.0 | 0.6 ± 0.5 | 21.2 ± 16.9 |
| Sample | N | Input | Filtered | denoisedF | denoisedR | Merged | Nonchimeric |
|---|---|---|---|---|---|---|---|
| Site 1 | 2 | 4178 | 3247 | 1938 | 1932 | 1402 | 1402 |
| Site 2 | 7 | 1,567,837 | 1,431,866 | 1,400,922 | 1,404,568 | 1,328,441 | 1,300,807 |
| Site 3 | 7 | 1,225,785 | 1,117,851 | 1,089,082 | 1,091,091 | 1,024,133 | 1,003,202 |
| Site 4 | 7 | 1,403,653 | 1,273,239 | 1,245,568 | 1,249,113 | 1,184,875 | 1,161,627 |
| Site 5 | 7 | 1,056,645 | 954,990 | 922,552 | 924,741 | 862,151 | 848,908 |
| Site 6 | 7 | 1,539,280 | 1,381,830 | 1,366,507 | 1,367,413 | 1,316,790 | 1,293,487 |
| Negative DNA ext #1 | 1 | 695 | 48 | 16 | 8 | 6 | 6 |
| Negative DNA ext #2 | 1 | 824 | 28 | 5 | 1 | 0 | 0 |
| Negative sequencing | 1 | 427 | 24 | 7 | 7 | 7 | 7 |
| Positive control | 1 | 114,168 | 103,850 | 103,432 | 103,651 | 101,735 | 98,655 |
| SV | Phylum | Class | Order | Family | Genus |
|---|---|---|---|---|---|
| 1 | Campylobacterota | Campylobacteria | Campylobacterales | Arcobacteraceae | Arcobacter |
| 2 | Campylobacterota | Campylobacteria | Campylobacterales | Arcobacteraceae | Arcobacter |
| 3 | Cyanobacteria | Cyanobacteriia | Cyanobacteriales | Phormidiaceae | Planktothrix NIVA-CYA 15 |
| 9 | Bacteroidota | Bacteroidia | Bacteroidales | Prevotellaceae | Prevotella |
| 10 | Proteobacteria | Gammaproteobacteria | Pseudomonadales | Moraxellaceae | Acinetobacter |
| 11 | Proteobacteria | Gammaproteobacteria | Enterobacterales | Aeromonadaceae | Aeromonas |
| 12 | Proteobacteria | Gammaproteobacteria | Enterobacterales | Enterobacteriaceae | Escherichia-Shigella |
| 18 | Proteobacteria | Gammaproteobacteria | Burkholderiales | Comamonadaceae | Family_ Comamonadaceae |
| 21 | Proteobacteria | Gammaproteobacteria | Enterobacterales | Succinivibrionaceae | Succinivibrio |
| 26 | Proteobacteria | Alphaproteobacteria | SAR11 clade | Clade III | Family_Clade III |
| 27 | Campylobacterota | Campylobacteria | Campylobacterales | Arcobacteraceae | Pseudarcobacter |
| 30 | Campylobacterota | Campylobacteria | Campylobacterales | Arcobacteraceae | Pseudarcobacter |
| 31 | Proteobacteria | Gammaproteobacteria | Burkholderiales | Burkholderiaceae | Polynucleobacter |
| Sample | Conditions | S2 | S3 | S4 | S5 | Unknown |
|---|---|---|---|---|---|---|
| S6S12 | L | 0.00 ± 0.00 | 0.32 ± 0.32 | 0.76 ± 0.29 | 1.71 ± 0.59 | 97.21 ± 0.46 |
| S6S14 | L | 0.15 ± 0.10 | 0.20 ± 0.12 | 1.03 ± 0.21 | 2.14 ± 0.42 | 96.50 ± 0.35 |
| S6S13 | L | 0.07 ± 0.07 | 0.24 ± 0.17 | 0.27 ± 0.12 | 1.35 ± 0.42 | 98.00 ± 0.50 |
| S6S18 | L | 0.18 ± 0.15 | 1.15 ± 0.32 | 0.67 ± 0.32 | 1.89 ± 0.41 | 96.11 ± 0.40 |
| S6S19 | H | 0.21 ± 0.05 | 0.65 ± 0.31 | 1.55 ± 0.25 | 5.67 ± 0.54 | 91.92 ± 0.54 |
| S6S20 | H | 0.04 ± 0.16 | 0.6 ± 0.32 | 1.25 ± 0.34 | 2.9 ± 0.66 | 95.22 ± 0.62 |
| S6S21 | H | 0.01 ± 0.03 | 0.44 ± 0.14 | 0.49 ± 0.22 | 2.62 ± 0.25 | 96.44 ± 0.41 |






References
- Zittis, G.; Hadjinicolaou, P.; Klangidou, M.; Proestos, Y.; Lelieveld, J. A Multi-Model, Multi-Scenario, and Multi-Domain Analysis of Regional Climate Projections for the Mediterranean. Reg. Environ. Chang. 2019, 19, 2621–2635. [Google Scholar] [CrossRef]
- IPCC. Climate Change 2022: Impacts, Adaptation and Vulnerability; Pörtner, H.-O.D.C., Roberts, M., Tignor, E.S., Poloczanska, K., Mintenbeck, A., Alegría, M., Craig, S., Langsdorf, S., Löschke, V., Möller, A., et al., Eds.; Cambridge University Press: Cambridge, UK, 2022; ISBN 9789291691623. [Google Scholar]
- Mohammed, T.; Al-Amin, A.Q. Climate Change and Water Resources in Algeria: Vulnerability, Impact and Adaptation Strategy. Econ. Environ. Stud. 2018, 18, 411–429. [Google Scholar] [CrossRef]
- Khalid, S.; Shahid, M.; Natasha; Bibi, I.; Sarwar, T.; Shah, A.H.; Niazi, N.K. A Review of Environmental Contamination and Health Risk Assessment of Wastewater Use for Crop Irrigation with a Focus on Low and High-Income Countries. Int. J. Environ. Res. Public Health 2018, 15, 895. [Google Scholar] [CrossRef] [PubMed]
- Edberg, S.C.; Rice, E.W.; Karlin, R.J.; Allen, M.J. Escherichia Coli: The Best Biological Drinking Water Indicator for Public Health Protection. J. Appl. Microbiol. 2000, 88, 106S–116S. [Google Scholar] [CrossRef]
- Saingam, P.; Li, B.; Yan, T. Fecal Indicator Bacteria, Direct Pathogen Detection, and Microbial Community Analysis Provide Different Microbiological Water Quality Assessment of a Tropical Urban Marine Estuary. Water Res. 2020, 185, 116280. [Google Scholar] [CrossRef] [PubMed]
- NHMRC. Australian Drinking Water Guidelines Paper 6: National Water Quality Management Strategy, Version 3.4 Updated October 2017; NHMRC: Canberra, Australia, 2011; ISBN 1864965118. [Google Scholar]
- European Commission. Directive of The European Parliament and of The Council on the Quality of Water Intended for Human. J. Chem. Inf. Model. 2018, 53, 1689–1699. [Google Scholar]
- Health Canada. Guidelines for Canadian Drinking Water Quality. Guideline Technical Document. Enteric Protozoa: Giardia and Cryptosporidium; Health Canada: Vancouver, BC, Canada, 2019; ISBN 6139572991. [Google Scholar]
- Jofre, J.; Lucena, F.; Blanch, A.R.; Muniesa, M. Coliphages as Model Organisms in the Characterization and Management of Water Resources. Water 2016, 8, 199. [Google Scholar] [CrossRef]
- Jebri, S.; Muniesa, M.; Jofre, J. Part Two. Indicators and Microbial Source Tracking Markers General and Indicators of Fecal Pollution. In Global Water Pathogen Project; Rose, J.B., Jiménez-Cisneros, B., Eds.; UNESCO: Paris, France, 2017; Volume 7, pp. 1–43. [Google Scholar]
- Harwood, V.J.; Staley, C.; Badgley, B.D.; Borges, K.; Korajkic, A. Microbial Source Tracking Markers for Detection of Fecal Contamination in Environmental Waters: Relationships between Pathogens and Human Health Outcomes. FEMS Microbiol. Rev. 2014, 38, 1–40. [Google Scholar] [CrossRef] [PubMed]
- Paruch, L.; Paruch, A.M. An Overview of Microbial Source Tracking Using Host-Specific Genetic Markers to Identify Origins of Fecal Contamination in Different Water Environments. Water 2022, 14, 1809. [Google Scholar] [CrossRef]
- Mauffret, A.; Caprais, M.P.; Gourmelon, M. Relevance of Bacteroidales and F-Specific RNA Bacteriophages for Efficient Fecal Contamination Tracking at the Level of a Catchment in France. Appl. Environ. Microbiol. 2012, 78, 5143–5152. [Google Scholar] [CrossRef] [PubMed]
- Hsu, F.C.; Shieh, Y.S.C.; Van Duin, J.; Beekwilder, M.J.; Sobsey, M.D. Genotyping Male-Specific RNA Coliphages by Hybridization with Oligonucleotide Probes. Appl. Environ. Microbiol. 1995, 61, 3960–3966. [Google Scholar] [CrossRef] [PubMed]
- Schaper, M.; Durán, A.E.; Jofre, J. Comparative Resistance of Phage Isolates of Four Genotypes of F-Specific RNA Bacteriophages to Various Inactivation Processes. Appl. Environ. Microbiol. 2002, 68, 3702–3707. [Google Scholar] [CrossRef] [PubMed]
- Mieszkin, S.; Caprais, M.P.; Le Mennec, C.; Le Goff, M.; Edge, T.A.; Gourmelon, M. Identification of the Origin of Faecal Contamination in Estuarine Oysters Using Bacteroidales and F-Specific RNA Bacteriophage Markers. J. Appl. Microbiol. 2013, 115, 897–907. [Google Scholar] [CrossRef] [PubMed]
- Yahya, M.; Blanch, A.R.; Meijer, W.G.; Antoniou, K.; Hmaied, F.; Ballesté, E. Comparison of the Performance of Different Microbial Source Tracking Markers among European and North African Regions. J. Environ. Qual. 2017, 46, 760–766. [Google Scholar] [CrossRef] [PubMed]
- Dutilh, B.E.; Cassman, N.; McNair, K.; Sanchez, S.E.; Silva, G.G.Z.; Boling, L.; Barr, J.J.; Speth, D.R.; Seguritan, V.; Aziz, R.K.; et al. A Highly Abundant Bacteriophage Discovered in the Unknown Sequences of Human Faecal Metagenomes. Nat. Commun. 2014, 5, 4498. [Google Scholar] [CrossRef]
- Edwards, R.A.; Vega, A.A.; Norman, H.M.; Ohaeri, M.; Levi, K.; Dinsdale, E.A.; Cinek, O.; Aziz, R.K.; McNair, K.; Barr, J.J.; et al. Global Phylogeography and Ancient Evolution of the Widespread Human Gut Virus CrAssphage. Nat. Microbiol. 2019, 4, 1727–1736. [Google Scholar] [CrossRef]
- Shkoporov, A.N.; Khokhlova, E.V.; Fitzgerald, C.B.; Stockdale, S.R.; Draper, L.A.; Ross, R.P.; Hill, C. ΦCrAss001 Represents the Most Abundant Bacteriophage Family in the Human Gut and Infects Bacteroides Intestinalis. Nat. Commun. 2018, 9, 4781. [Google Scholar] [CrossRef] [PubMed]
- Ramos-Barbero, M.D.; Gómez-Gómez, C.; Sala-Comorera, L.; Rodríguez-Rubio, L.; Morales-Cortes, S.; Mendoza-Barberá, E.; Vique, G.; Toribio-Avedillo, D.; Blanch, A.R.; Ballesté, E.; et al. Characterization of CrAss-like Phage Isolates Highlights Crassvirales Genetic Heterogeneity and Worldwide Distribution. Nat. Commun. 2023, 14, 4295. [Google Scholar] [CrossRef] [PubMed]
- García-Aljaro, C.; Martín-Díaz, J.; Viñas-Balada, E.; Calero-Cáceres, W.; Lucena, F.; Blanch, A.R. Mobilisation of Microbial Indicators, Microbial Source Tracking Markers and Pathogens after Rainfall Events. Water Res. 2017, 112, 248–253. [Google Scholar] [CrossRef]
- Stachler, E.; Kelty, C.; Sivaganesan, M.; Li, X.; Bibby, K.; Shanks, O.C. Quantitative CrAssphage PCR Assays for Human Fecal Pollution Measurement. Environ. Sci. Technol. 2017, 51, 9146–9154. [Google Scholar] [CrossRef]
- Ballesté, E.; Pascual-Benito, M.; Martín-Díaz, J.; Blanch, A.R.; Lucena, F.; Muniesa, M.; Jofre, J.; García-Aljaro, C. Dynamics of CrAssphage as a Human Source Tracking Marker in Potentially Faecally Polluted Environments. Water Res. 2019, 155, 233–244. [Google Scholar] [CrossRef] [PubMed]
- Crank, K.; Li, X.; North, D.; Ferraro, G.B.; Iaconelli, M.; Mancini, P.; La Rosa, G.; Bibby, K. CrAssphage Abundance and Correlation with Molecular Viral Markers in Italian Wastewater. Water Res. 2020, 184, 116161. [Google Scholar] [CrossRef] [PubMed]
- Malla, B.; Ghaju Shrestha, R.; Tandukar, S.; Sherchand, J.B.; Haramoto, E. Performance Evaluation of Human-Specific Viral Markers and Application of Pepper Mild Mottle Virus and CrAssphage to Environmental Water Samples as Fecal Pollution Markers in the Kathmandu Valley, Nepal. Food Environ. Virol. 2019, 11, 274–287. [Google Scholar] [CrossRef]
- Kongprajug, A.; Mongkolsuk, S.; Sirikanchana, K. CrAssphage as a Potential Human Sewage Marker for Microbial Source Tracking in Southeast Asia. Environ. Sci. Technol. Lett. 2019, 6, 113859. [Google Scholar] [CrossRef]
- Ahmed, W.; Lobos, A.; Senkbeil, J.; Peraud, J.; Gallard, J.; Harwood, V.J. Evaluation of the Novel CrAssphage Marker for Sewage Pollution Tracking in Storm Drain Outfalls in Tampa, Florida. Water Res. 2018, 131, 142–150. [Google Scholar] [CrossRef]
- Stachler, E.; Akyon, B.; De Carvalho, N.A.; Ference, C.; Bibby, K. Correlation of CrAssphage QPCR Markers with Culturable and Molecular Indicators of Human Fecal Pollution in an Impacted Urban Watershed. Environ. Sci. Technol. 2018, 52, 7505–7512. [Google Scholar] [CrossRef] [PubMed]
- Jennings, W.C.; Gálvez-Arango, E.; Prieto, A.L.; Boehm, A.B. CrAssphage for Fecal Source Tracking in Chile: Covariation with Norovirus, HF183, and Bacterial Indicators. Water Res. X 2020, 9, 100071. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, W.; Zhang, Q.; Lobos, A.; Senkbeil, J.; Sadowsky, M.J.; Harwood, V.J.; Saeidi, N.; Marinoni, O.; Ishii, S. Precipitation Influences Pathogenic Bacteria and Antibiotic Resistance Gene Abundance in Storm Drain Outfalls in Coastal Sub-Tropical Waters. Environ. Int. 2018, 116, 308–318. [Google Scholar] [CrossRef] [PubMed]
- Destoumieux-Garzón, D.; Mavingui, P.; Boetsch, G.; Boissier, J.; Darriet, F.; Duboz, P.; Fritsch, C.; Giraudoux, P.; Roux, F.L.; Morand, S.; et al. The One Health Concept: 10 Years Old and a Long Road Ahead. Front. Vet. Sci. 2018, 5, 14. [Google Scholar] [CrossRef] [PubMed]
- Webster, J.P.; Gower, C.M.; Knowles, S.C.L.; Molyneux, D.H.; Fenton, A. One Health—An Ecological and Evolutionary Framework for Tackling Neglected Zoonotic Diseases. Evol. Appl. 2016, 9, 313–333. [Google Scholar] [CrossRef]
- Pascual-Benito, M.; Ballesté, E.; Monleón-Getino, T.; Urmeneta, J.; Blanch, A.R.; García-Aljaro, C.; Lucena, F. Impact of Treated Sewage Effluent on the Bacterial Community Composition in an Intermittent Mediterranean Stream. Environ. Pollut. 2020, 266, 115254. [Google Scholar] [CrossRef] [PubMed]
- Azaroual, S.E.; Kasmi, Y.; Aasfar, A.; El Arroussi, H.; Zeroual, Y.; El Kadiri, Y.; Zrhidri, A.; Elfahime, E.; Sefiani, A.; Meftah Kadmiri, I. Investigation of Bacterial Diversity Using 16S RRNA Sequencing and Prediction of Its Functionalities in Moroccan Phosphate Mine Ecosystem. Sci. Rep. 2022, 12, 3741. [Google Scholar] [CrossRef] [PubMed]
- ISO 8467:1993; Water Quality—Determination of Permanganate Index. ISO: Geneve, Switzerland, 1993.
- ISO 6878:2004; Water Quality—Determination of Phosphorus—Ammonium Molybdate Spectrometric Method. International Organization for Standardization: Geneva, Switzerland, 2004.
- ISO 5171/1; Water Quality—Determination of Ammonium—Part 1: Manual Spectrometric of Ammonium Method. International Organization for Standardization: Geneva, Switzerland, 1984.
- ISO 10406-1; Water Quality—Detection and Enumeration of Escherichia coli and Coliform Bacteria—Part 1: Membrane Filtration Method. International Organization for Standardization: Geneva, Switzerland, 2000.
- Ruiz-Hernando, M.; Martín-Díaz, J.; Labanda, J.; Mata-Alvarez, J.; Llorens, J.; Lucena, F.; Astals, S. Effect of Ultrasound, Low-Temperature Thermal and Alkali Pre-Treatments on Waste Activated Sludge Rheology, Hygienization and Methane Potential. Water Res. 2014, 61, 119–129. [Google Scholar] [CrossRef] [PubMed]
- ISO 10705-2; ISO Water Quality—Detection and Enumeration of Bacteriophages—Part 2: Enumeration of Somatic Coliphages. International Organization for Standardization: Geneva, Switzerland, 2000.
- Gourmelon, M.; Caprais, M.P.; Ségura, R.; Le Mennec, C.; Lozach, S.; Piriou, J.Y.; Rincé, A. Evaluation of Two Library-Independent Microbial Source Tracking Methods to Identify Sources of Fecal Contamination in French Estuaries. Appl. Environ. Microbiol. 2007, 73, 4857–4866. [Google Scholar] [CrossRef] [PubMed]
- Parada, A.E.; Needham, D.M.; Fuhrman, J.A. Every Base Matters: Assessing Small Subunit rRNA Primers for Marine Microbiomes with Mock Communities, Time Series and Global Field Samples. Environ. Microbiol. 2016, 18, 1403–1414. [Google Scholar] [CrossRef] [PubMed]
- Apprill, A.; Mcnally, S.; Parsons, R.; Weber, L. Minor Revision to V4 Region SSU RRNA 806R Gene Primer Greatly Increases Detection of SAR11 Bacterioplankton. Aquat. Microb. Ecol. 2015, 75, 129–137. [Google Scholar] [CrossRef]
- Dębska, K.; Rutkowska, B.; Szulc, W.; Gozdowski, D. Changes in Selected Water Quality Parameters in the Utrata River as a Function of Catchment Area Land Use. Water 2021, 13, 2989. [Google Scholar] [CrossRef]
- Kongprajug, A.; Denpetkul, T.; Chyerochana, N.; Mongkolsuk, S.; Sirikanchana, K. Human Fecal Pollution Monitoring and Microbial Risk Assessment for Water Reuse Potential in a Coastal Industrial–Residential Mixed-Use Watershed. Front. Microbiol. 2021, 12, 647602. [Google Scholar] [CrossRef] [PubMed]
- Ishii, S.; Sadowsky, M.J. Escherichia Coli in the Environment: Implications for Water Quality and Human Health. Microbes Environ. 2008, 23, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, K.H.; Smith, S.; Roundtree, A.; Feistel, D.J.; Kirby, A.E.; Levy, K.; Mattioli, M.C. Fecal Indicators and Antibiotic Resistance Genes Exhibit Diurnal Trends in the Chattahoochee River: Implications for Water Quality Monitoring. Front. Microbiol. 2022, 13, 1029176. [Google Scholar] [CrossRef] [PubMed]
- Pascual-Benito, M.; Nadal-Sala, D.; Tobella, M.; Ballesté, E.; García-Aljaro, C.; Sabaté, S.; Sabater, F.; Martí, E.; Gracia, C.A.; Blanch, A.R.; et al. Modelling the Seasonal Impacts of a Wastewater Treatment Plant on Water Quality in a Mediterranean Stream Using Microbial Indicators. J. Environ. Manag. 2020, 261, 110220. [Google Scholar] [CrossRef] [PubMed]
- Omar, K.B.; Barnard, T.G. The Occurrence of Pathogenic Escherichia Coli in South African Wastewater Treatment Plants as Detected by Multiplex PCR. Water SA 2010, 36, 172–176. [Google Scholar] [CrossRef]
- Akiba, M.; Senba, H.; Otagiri, H.; Prabhasankar, V.P.; Taniyasu, S.; Yamashita, N.; Lee, K.; Yamamoto, T.; Tsutsui, T.; Ian Joshua, D.; et al. Impact of Wastewater from Different Sources on the Prevalence of Antimicrobial-Resistant Escherichia Coli in Sewage Treatment Plants in South India. Ecotoxicol. Environ. Saf. 2015, 115, 203–208. [Google Scholar] [CrossRef] [PubMed]
- WHO. Indicators of Microbial Water Quality. In Water Quality: Guidelines, Standards and Health: Assessment of Risk and Risk Management for Water-Related Infectious Diseases; Chapter13; Lorna, F., Jamie, B., World Health Organization, Eds.; IWA Publishing: London, UK, 2001; pp. 1–431. [Google Scholar]
- Rose, J.B.; Farrah, S.R.; Harwood, V.J.; Levine, A.; Lukasik, J.; McLaughlin, M.R.; Chivukula, V.; Gennaccaro, A.; Scott, T.M. Reduction of Pathogens, Indicator Bacteria, and Alternative Indicators by Wastewater Treatment and Reclamation Processes. Water Supply 2004, 3, 247–252. [Google Scholar] [CrossRef]
- Hamaidi, M.S.; Hamaidi-chergui, F.; Errahmani, M.B. Efficiency of Indicator Bacteria Removal in a Wastewater Treatment Plant (Algiers, Algeria). In Proceedings of the 2nd International Conference—Water Resources and Wetlands, Tulcea, Romania, 11–13 September 2014; pp. 503–509. [Google Scholar]
- Grabow, W.; Holtzhausen, C.; Villiers, J. De Research on Bacteriophages as Indicators of Water Quality; Water Research Commission: Pretoria, South Africa, 1993; p. 148. [Google Scholar]
- Yahya, M.; Hmaied, F.; Jebri, S.; Jofre, J.; Hamdi, M. Bacteriophages as Indicators of Human and Animal Faecal Contamination in Raw and Treated Wastewaters from Tunisia. J. Appl. Microbiol. 2015, 118, 1217–1225. [Google Scholar] [CrossRef] [PubMed]
- Blanch, A.R.; Belanche-Muñoz, L.; Bonjoch, X.; Ebdon, J.; Gantzer, C.; Lucena, F.; Ottoson, J.; Kourtis, C.; Iversen, A.; Kühn, I.; et al. Integrated Analysis of Established and Novel Microbial and Chemical Methods for Microbial Source Tracking. Appl. Environ. Microbiol. 2006, 72, 5915–5926. [Google Scholar] [CrossRef] [PubMed]
- Farkas, K.; Adriaenssens, E.M.; Walker, D.I.; McDonald, J.E.; Malham, S.K.; Jones, D.L. Critical Evaluation of CrAssphage as a Molecular Marker for Human-Derived Wastewater Contamination in the Aquatic Environment. Food Environ. Virol. 2019, 11, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Sabar, M.A.; Honda, R.; Haramoto, E. CrAssphage as an Indicator of Human-Fecal Contamination in Water Environment and Virus Reduction in Wastewater Treatment. Water Res. 2022, 221, 118827. [Google Scholar] [CrossRef] [PubMed]
- Martín-Díaz, J.; García-Aljaro, C.; Pascual-Benito, M.; Galofré, B.; Blanch, A.R.; Lucena, F. Microcosms for Evaluating Microbial Indicator Persistence and Mobilization in Fluvial Sediments during Rainfall Events. Water Res. 2017, 123, 623–631. [Google Scholar] [CrossRef] [PubMed]
- U.S. Environmental Protection Agency Office of Water. EPA Method 1611: Enterococci in Water by TaqMan® Quantitative Polymerase Chain Reaction (QPCR) Assay; EPA-821-R-12-008; U.S. Environmental Protection Agency Office of Water: Washington, DC, USA, 2012. [Google Scholar]
- U.S. Environmental Protection Agency Office of Water. EPA Office of Water Method 1697: Characterization of Human Fecal Pollution in Water by HF183/BacR287 TaqMan® Quantitative Polymerase Chain Reaction (QPCR) Assay I; U.S. Environmental Protection Agency Office of Water: Washington, DC, USA, 2019. [Google Scholar]
- Ishii, S.; Ksoll, W.B.; Hicks, R.E.; Sadowsky, M.J. Presence and Growth of Naturalized Escherichia Coli in Temperate Soils from Lake Superior Watersheds. Appl. Environ. Microbiol. 2006, 72, 612–621. [Google Scholar] [CrossRef] [PubMed]
- Bumadian, M.; Almansury, H.H.; Bozakouk, I.H.; Lawgali, Y.F.; Bleiblo, F.A. Detection and Enumeration of Coliform Bacteria in Drinking Water at Hospital of Benghazi/Libya. J. Exp. Biol. Agric. Sci. 2013, 1, 436–440. [Google Scholar]
- Ishii, S.; Hansen, D.L.; Hicks, R.E.; Sadowsky, M.J. Beach Sand and Sediments Are Temporal Sinks and Sources of Escherichia coli in Lake Superior. Environ. Sci. Technol. 2007, 41, 2203–2209. [Google Scholar] [CrossRef] [PubMed]
- Jofre, J. Is the Replication of Somatic Coliphages in Water Environments Significant? J. Appl. Microbiol. 2009, 106, 1059–1069. [Google Scholar] [CrossRef] [PubMed]
- Tandukar, S.; Sherchand, J.; Bhandari, D.; Sherchan, S.; Malla, B.; Ghaju Shrestha, R.; Haramoto, E. Presence of Human Enteric Viruses, Protozoa, and Indicators of Pathogens in the Bagmati River, Nepal. Pathogens 2018, 7, 38. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Greaves, J.; Arp, L.; Stone, D.; Bibby, K. Comparative Fate of CrAssphage with Culturable and Molecular Fecal Pollution Indicators during Activated Sludge Wastewater Treatment. Environ. Int. 2020, 136, 105452. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, W.; Gyawali, P.; Feng, S.; McLellan, S.L. Host Specificity and Sensitivity of Established and Novel. Appl. Environ. Microbiol. 2019, 85, e00641-19. [Google Scholar] [CrossRef] [PubMed]
- Stachler, E.; Crank, K.; Bibby, K. Co-Occurrence of CrAssphage with Antibiotic Resistance Genes in an Impacted Urban Watershed. Environ. Sci. Technol. Lett. 2019, 6, 216–221. [Google Scholar] [CrossRef]
- Ebdon, J.; Muniesa, M.; Taylor, H. The Application of a Recently Isolated Strain of Bacteroides (GB-124) to Identify Human Sources of Faecal Pollution in a Temperate River Catchment. Water Res. 2007, 41, 3683–3690. [Google Scholar] [CrossRef] [PubMed]
- Farkas, K.; Cooper, D.M.; McDonald, J.E.; Malham, S.K.; de Rougemont, A.; Jones, D.L. Seasonal and Spatial Dynamics of Enteric Viruses in Wastewater and in Riverine and Estuarine Receiving Waters. Sci. Total Environ. 2018, 634, 1174–1183. [Google Scholar] [CrossRef] [PubMed]
- Prevost, B.; Lucas, F.S.; Goncalves, A.; Richard, F.; Moulin, L.; Wurtzer, S. Large Scale Survey of Enteric Viruses in River and Waste Water Underlines the Health Status of the Local Population. Environ. Int. 2015, 79, 42–50. [Google Scholar] [CrossRef] [PubMed]
- Rusiñol, M.; Fernandez-Cassi, X.; Timoneda, N.; Carratalà, A.; Abril, J.F.; Silvera, C.; Figueras, M.J.; Gelati, E.; Rodó, X.; Kay, D.; et al. Evidence of Viral Dissemination and Seasonality in a Mediterranean River Catchment: Implications for Water Pollution Management. J. Environ. Manag. 2015, 159, 58–67. [Google Scholar] [CrossRef] [PubMed]
- Sassi, H.P.; Tuttle, K.D.; Betancourt, W.Q.; Kitajima, M.; Gerba, C.P. Persistence of Viruses by QPCR Downstream of Three Effluent-Dominated Rivers in the Western United States. Food Environ. Virol. 2018, 10, 297–304. [Google Scholar] [CrossRef] [PubMed]
- Sibanda, T.; Okoh, A.I. Assessment of the Incidence of Enteric Adenovirus Species and Serotypes in Surface Waters in the Eastern Cape Province of South Africa: Tyume River as a Case Study. Sci. World J. 2012, 2012, 949216. [Google Scholar] [CrossRef] [PubMed]
- Venegas, C.; Diez, H.; Blanch, A.R.; Jofre, J.; Campos, C. Microbial Source Markers Assessment in the Bogotá River Basin (Colombia). J. Water Health 2015, 13, 801–810. [Google Scholar] [CrossRef] [PubMed]
- Wangkahad, B.; Mongkolsuk, S.; Sirikanchana, K. Integrated Multivariate Analysis with Nondetects for the Development of Human Sewage Source-Tracking Tools Using Bacteriophages of Enterococcus Faecalis. Environ. Sci. Technol. 2017, 51, 2235–2245. [Google Scholar] [CrossRef] [PubMed]
- Besemer, K.; Peter, H.; Logue, J.B.; Langenheder, S.; Lindström, E.S.; Tranvik, L.J.; Battin, T.J. Unraveling Assembly of Stream Biofilm Communities. ISME J. 2012, 6, 1459–1468. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Gao, Y.; Huang, X.; Ni, P.; Wu, Y.; Deng, Y.; Zhan, A. Adaptive Shifts of Bacterioplankton Communities in Response to Nitrogen Enrichment in a Highly Polluted River. Environ. Pollut. 2019, 245, 290–299. [Google Scholar] [CrossRef] [PubMed]
- Oren, A.; Garrity, G.M. Valid Publication of the Names of Forty-Two Phyla of Prokaryotes. Int. J. Syst. Evol. Microbiol. 2021, 71, 005056. [Google Scholar] [CrossRef] [PubMed]
- Newton, R.J.; Jones, S.E.; Eiler, A.; McMahon, K.D.; Bertilsson, S. A Guide to the Natural History of Freshwater Lake Bacteria. Microbiol. Mol. Biol. Rev. 2011, 75, 14–49. [Google Scholar] [CrossRef]
- Mou, X.; Lu, X.; Jacob, J.; Sun, S.; Heath, R. Metagenomic Identification of Bacterioplankton Taxa and Pathways Involved in Microcystin Degradation in Lake Erie. PLoS ONE 2013, 8, e61890. [Google Scholar] [CrossRef]
- Salcher, M.M.; Pernthaler, J.; Posch, T. Seasonal Bloom Dynamics and Ecophysiology of the Freshwater Sister Clade of SAR11 Bacteria ‘That Rule the Waves’ (LD12). ISME J. 2011, 5, 1242–1252. [Google Scholar] [CrossRef] [PubMed]
- Staff, T.P.O. Correction: Control of Temperature on Microbial Community Structure in Hot Springs of the Tibetan Plateau. PLoS ONE 2014, 9, e99751. [Google Scholar] [CrossRef]
- Weltzer, M.L.; Miller, S.R. Ecological Divergence of a Novel Group of Chloroflexus Strains along a Geothermal Gradient. Appl. Environ. Microbiol. 2013, 79, 1353–1358. [Google Scholar] [CrossRef] [PubMed]
- Ward, D.M.; Castenholz, R.W.; Miller, S.R. Ecology of Cyanobacteria II; Whitton, B.A., Ed.; Springer: Dordrecht, The Netherlands, 2012; Volume 9789400738, ISBN 978-94-007-3854-6. [Google Scholar]
- Milan, M.; Carraro, L.; Fariselli, P.; Martino, M.E.; Cavalieri, D.; Vitali, F.; Boffo, L.; Patarnello, T.; Bargelloni, L.; Cardazzo, B. Microbiota and Environmental Stress: How Pollution Affects Microbial Communities in Manila Clams. Aquat. Toxicol. 2018, 194, 195–207. [Google Scholar] [CrossRef] [PubMed]
- Pascual-Benito, M.; Emiliano, P.; Casas-Mangas, R.; Dacal-Rodríguez, C.; Gracenea, M.; Araujo, R.; Valero, F.; García-Aljaro, C.; Lucena, F. Assessment of Dead-End Ultrafiltration for the Detection and Quantification of Microbial Indicators and Pathogens in the Drinking Water Treatment Processes. Int. J. Hyg. Environ. Health 2020, 230, 113628. [Google Scholar] [CrossRef] [PubMed]







| EC | SSRC | SOMCPH | CrAssPH | EC | SSRC | SOMCPH | CrAssPH | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| EC | * | 0.82 | 0.81 | 0.15 | EC | * | 0.31 | 0.5 | 0.52 | ||
| Site 1 | SSRC | 0.00 | * | 0.50 | 0.13 | Site 4 | SSRC | 0.17 | * | 0.09 | 0.16 |
| SOMCPH | 0.00 | 0.10 | * | 0.09 | SOMCPH | 0.02 | 0.71 | * | 0.88 | ||
| CrAssPH | 0.65 | 0.79 | 0.71 | * | CrAssPH | 0.03 | 0.53 | 0.00 | * | ||
| EC | * | 0.50 | 0.73 | −0.16 | EC | * | 0.36 | 0.32 | −0.31 | ||
| Site 2 | SSRC | 0.00 | * | 0.39 | 0.25 | Site 5 | SSRC | 0.11 | * | 0.54 | 0.43 |
| SOMCPH | 0.00 | 0.08 | * | −0.18 | SOMCPH | 0.15 | 0.01 | * | 0.50 | ||
| CrAssPH | 0.55 | 0.34 | 0.50 | * | CrAssPH | 0.25 | 0.09 | 0.05 | * | ||
| EC | * | 0.72 | 0.73 | 0.46 | EC | * | 0.34 | 0.46 | 0.06 | ||
| Site 3 | SSRC | 0.00 | * | 0.43 | 0.27 | Site 6 | SSRC | 0.15 | * | 0.48 | 0.33 |
| SOMCPH | 0.00 | 0.05 | * | 0.61 | SOMCPH | 0.04 | 0.04 | * | 0.70 | ||
| CrAssPH | 0.06 | 0.29 | 0.01 | * | CrAssPH | 0.82 | 0.22 | 0.002 | * |
| Sampling Site | n | Total ASVs | Core ASVs | Chao1 | Shannon |
|---|---|---|---|---|---|
| 1 | 2 | 62 | NA | NA | NA |
| 2 | 7 | 3836 | 300 | 1262 ± 308 | 4.7 ± 0.5 |
| 3 | 7 | 4636 | 34 | 1189 ± 565 | 4.9 ± 0.9 |
| 4 | 7 | 4479 | 44 | 1049 ± 245 | 4.8 ± 0.6 |
| 5 | 7 | 4835 | 27 | 1098 ± 187 | 4.8 ± 0.5 |
| 6 | 7 | 1757 | 91 | 563 ± 114 | 4.3 ± 0.5 |
| Sample | Conditions | S2 | S4 | Unknown |
|---|---|---|---|---|
| S3S12 | L | 66.23 ± 2.4 | 5.36 ± 2.3 | 8.41 ± 0.3 |
| S3S14 | L | 32.44 ± 2.4 | 56.67 ± 2.3 | 10.89 ± 0.4 |
| S3S13 | L | 67.96 ± 2.0 | 18.91 ± 2.0 | 13.13 ± 0.3 |
| S3S18 | L | 35.85 ± 0.9 | 37.95 ± 1.0 | 26.2 ± 0.5 |
| S3S19 | H | 14.7 ± 1.2 | 11.86 ± 1.6 | 73.44 ± 1.9 |
| S3S20 | H | 37.5 ± 0.9 | 8.69 ± 0.6 | 53.81 ± 0.5 |
| S3S21 | H | 0.75 ± 0.3 | 19.03 ± 0.8 | 80.22 ± 0.8 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boulainine, D.; Benhamrouche, A.; Ballesté, E.; Mezaache-Aichour, S.; García-Aljaro, C. Spatio-Temporal Dynamics of crAssphage and Bacterial Communities in an Algerian Watershed Impacted by Fecal Pollution. Water 2024, 16, 1123. https://doi.org/10.3390/w16081123
Boulainine D, Benhamrouche A, Ballesté E, Mezaache-Aichour S, García-Aljaro C. Spatio-Temporal Dynamics of crAssphage and Bacterial Communities in an Algerian Watershed Impacted by Fecal Pollution. Water. 2024; 16(8):1123. https://doi.org/10.3390/w16081123
Chicago/Turabian StyleBoulainine, Dalal, Aziz Benhamrouche, Elisenda Ballesté, Samia Mezaache-Aichour, and Cristina García-Aljaro. 2024. "Spatio-Temporal Dynamics of crAssphage and Bacterial Communities in an Algerian Watershed Impacted by Fecal Pollution" Water 16, no. 8: 1123. https://doi.org/10.3390/w16081123