
Temporal Environmental Status of a Shallow Lake Using Alpha and Beta Diversity on Phytoplankton Communities

1
Laboratorio de Limnología, Departamento de Acuicultura y Recursos Agroalimentarios, Universidad de Los Lagos, Av. Fuchslocher 1305, Osorno 5290000, Chile
2
Department of Ecology and Ecosystem Modelling, University of Potsdam, Am Neuen Palais 10, 14469 Potsdam, Germany
*
Author to whom correspondence should be addressed.
Water 2024, 16(2), 274; https://doi.org/10.3390/w16020274
Submission received: 29 November 2023
/
Revised: 21 December 2023
/
Accepted: 25 December 2023
/
Published: 12 January 2024
(This article belongs to the Special Issue Ecohydrological Processes, Environmental Effects, and Integrated Regulation of Wetland Ecosystems, Volume II)
Abstract
:Phytoplankton communities are excellent indicators of the environmental status of shallow lakes since changes in diversity alert us to potential deterioration. To assess the environmental condition of a shallow lake with strong anthropogenic pressure from forest monocultures, alpha and beta diversities were determined. Three monitoring stations were established in the shallow Kusrüpuyewe Lake, southern Chile, which were monitored during the 2021 austral summer and winter. Alpha diversity was high indicating a stable environmental condition. In total, 103 species in summer, 96 species in winter and a high species diversity (Shannon index: H′ > 5 bits/ind) were recorded. Beta diversity indicated that this ecosystem is seasonally heterogeneous, is potentially productive and has a high species turnover. Based on the large species inventory, we conclude that this shallow lake is stable and diverse. Although it is threatened by the presence of forest monocultures, it maintains a stable environmental condition, mainly due to the ability of the phytoplankton present to self-regulate through the species turnover. This study provides the relevant background information to establish conservation measures around the shallow lake, an ecosystem that provides various ecosystem services for the local indigenous communities.
1. Introduction
A phytoplankton community constitutes a polyphyletic and highly diverse group of organisms [1] that contribute to the structure and functioning of aquatic ecosystems as one of the essential components at the base of the trophic chain [2]. They have the ability to respond quickly to changes in nutrient concentration and other environmental factors, such as alkalinity and temperature, forcing taxonomic changes of phytoplankton communities along the trophic gradient. Therefore, they are regarded as excellent indicators of the trophic state and ecological functioning [3,4], providing unique information on the environmental condition [5].
It is important to select the right tools to assess the environmental and ecological status in aquatic ecosystems, given the existence of various measures, which may present different degrees of sensitivity to environmental changes. Among metrics, diversity is a community attribute that reflects different aspects of a community, which are determined by different ecological processes [6]. The maintenance of biodiversity is essential to ensure the sustainability of natural resources. High levels of biodiversity allow for the functioning of ecosystems, a high resilience to external disturbances, both natural and anthropogenic, and an optimal adaptation in environmental change scenarios [7].
Alpha and beta diversity are ecological estimators that can be obtained from the density and richness of species [4,8,9], and the environmental state of the ecosystem can be evaluated with these tools [10]. While alpha diversity indicates the heterogeneity or stability of the ecosystem, beta diversity indicates productivity and variation in species richness using the presence/absence data [9,10,11]. Baselga et al. [12,13,14] proposed including density and richness in the calculation of beta diversity, using the Sørensen dissimilarity index to determine the balance of the ecosystem and the Simpson dissimilarity index to determine the percentage of species turnover. Environmental and climatic conditions drive changes in species turnover. Therefore, an ecosystem exposed to greater anthropogenic impacts would probably induce the homogenization of phytoplankton communities [9].
To understand the stability and properties of an ecosystem, it is necessary to determine the temporal variability of alpha and beta diversities [15,16,17,18], which is very useful to establish effective conservation strategies. These environmental indicators have been applied to phytoplankton communities mainly in shallow coastal lagoons, coastal wetlands and lakes with alluvial plains [19,20,21]; however, they have not been applied in shallow lakes with a strong anthropogenic pressure. At a global level, shallow lakes are highly threatened by anthropogenic disturbances that increase the drift of sediments [22], which are deposited in these shallow water bodies, in some cases clogging them, and have also been affected by the reduction in precipitation associated with global climate change [23].
Shallow lakes are ecosystems well-known for their diverse flora and fauna, which provide important ecosystem services and are therefore of high conservation interest [24,25]. These valuable ecosystems often exhibit large fluctuations in their hydrological regimes and other abiotic characteristics [26], providing microhabitats of ecological interest, and have an important role in the nitrogen and phosphorus cycles [25,27,28]. These ecosystems are currently highly threatened by global warming and anthropogenic pressures [25,29,30,31], especially due to land use change, which affects the sustainability of these relevant natural water reservoirs [10,16,20].
Considering the above, this study attempts to evaluate the environmental status of the Kusrüpuyewe Lake by determining the alpha and beta diversities of the phytoplankton communities and to formulate conservation strategies. We hypothesize that (1) the alpha and beta diversity of phytoplankton species present in the shallow lake indicate stable environmental health during summer and winter; (2) there will be a high turnover of phytoplankton species between seasons, which would indicate that the shallow lake is stable.
2. Materials and Methods
2.1. Study Area
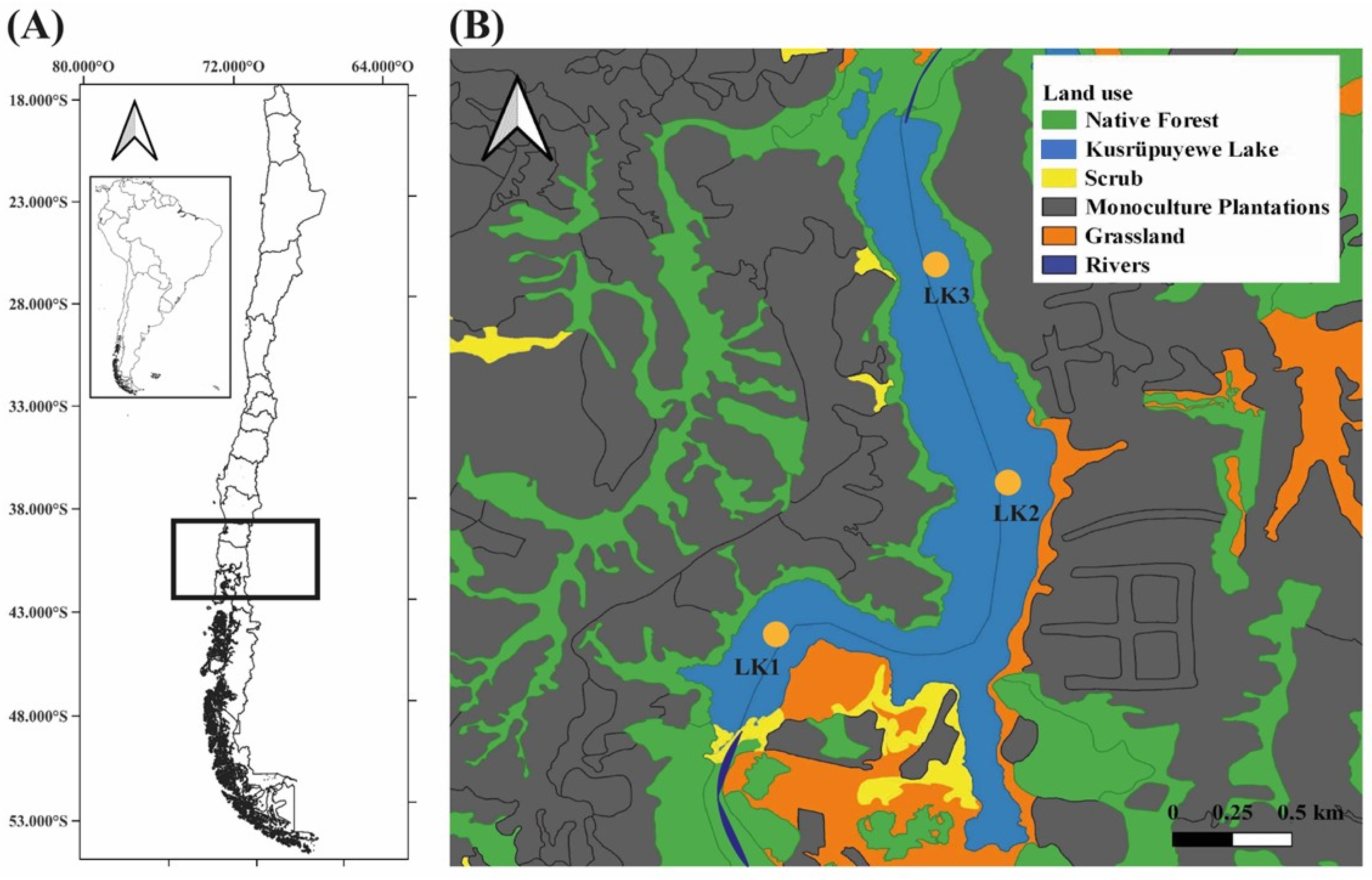
Kusrüpuyewe shallow lake (40°20′ S; 73°26′ W) is located in southern Chile (Figure 1A), in the coastal mountain range at approx. 50 m above sea level. Maximum depth is 9.5 m. It has a surface area of 145.62 ha. The volume is lowest in summer and highest in winter. Its main tributary is the Pucopio River (Figure 1B). It belongs to the sub-basin of the Pucopio River and is inserted within the native evergreen forest, dominated by Temu-Pitra. The ecological status of the area is threatened by the presence of exotic forest monocultures (e.g., pine and eucalyptus) surrounding this shallow lake and small-scale land use for agriculture and livestock farming (Figure 1B). This shallow lake provides habitat and shelter for mammals and waterfowl [22]. The area has a temperate-rainy climate, with a precipitation of 2500 mm year−1. The prevailing winds from the north and northwest bring abundant precipitation in winter, about 75% accumulates between April and November. Indigenous communities live around this shallow lake, which benefit from the various ecosystem services such as provision of water for irrigation, recreation, fishing and ceremonial events [22].
2.2. Sampling, Phytoplankton Analysis and Physicochemical Parameters
Three monitoring stations, about 1 km apart from each other, were established in the shallow lake during the summer and winter 2021, aiming for a broad spatial coverage of the entire lake: LK1 = located near the mouth of the Pucopio River, with the presence of native forest; LK2 = center of the lake without direct influence of the river, less presence of native forest and abundant presence of monocultures on the shores of the lake; LK3 = close to the river outlet, scarce presence of native forest and a strong influence of monocultures on the shores of the lake (Figure 1B).
Six water samples were obtained from the top (0–1 m) and bottom zone (one meter above the bottom) of each station (Figure 1B, Table 1) with a 3 L Van Dorn hydrographic bottle, for a total of 12 samples to ensure representative sample [32,33]. Moreover, 1 L Sub-samples were fixed with non-acid Lugol. The quantification and identification of the phytoplankton species was carried out using the Utermöhl method [34], using a 25 mL sedimentation chamber and with direct observation in a LEITZ Diavert inverted microscope. Density was expressed in cells L−1. Taxonomic identification was carried out to the genus/species level using the specialized literature [35,36]. To characterize the environmental condition at each sampling station, a vertical profile using a Garmin GPSMAP echo sounder and EXO-1 autonomous probe was recorded. We measured depth, temperature, pH (accuracy ±0.2 pH units for entire temp range), DO (mg/L: 0 to 20 mg/L: ±0.1 mg/L or 1% of reading and% sat: 0 to 200%: ±1% of reading or 1% saturation) and conductivity (0 to 100: ±0.5% of reading or 0.001 µS/cm). Chlorophyll “a” (three replicates) was determined following Nusch’s methods using hot ethanol extraction [37], and total suspended solids were determined by gravimetric methods [38].
2.3. Alpha and Beta Diversities Metrics
To determine spatial and temporal environmental health in Kusrüpuyewe Lake, alpha and beta diversities were used in this study. Alpha diversity was obtained using the Chao-1 species richness estimators and Shannon index; we consider a Shannon index of H′ > 2.6 bits/ind high for phytoplankton assemblages [39]. Alpha diversity indicated spatial and temporal ecosystem stability [40]. The beta diversity proposed by Baselga [13] was calculated from the density matrix of phytoplankton species, obtaining the Sørensen dissimilarity index (βSØR) to estimate ecological stability and the Simpson dissimilarity index (βSIM) to calculate the percentage of species turnover within Kusrüpuyewe Lake spatially among sampling sites and temporally between summer and winter. With this, we can quantify the degree of similarity among sites and seasons. In addition, Pielou’s evenness, to assess diversity/density equality, and Hill’s effective number of species indicate that the number of species having the same abundance were determined. All these analyses were carried out with the free PAST software v4.03.
2.4. Statistical Analyses
To estimate whether the sampling effort was appropriate to analyze species richness, the rarefaction curves were calculated for each monitoring station. To quantify differences between summer and winter phytoplankton abundance, the Bray–Curtis dissimilarity index of the biological matrix was calculated and used for the analyses of similarity (ANOSIM; p = 0.05) and a similarity percentage (SIMPER) to estimate phytoplankton turnover. We can understand turnover as the replacement of species along a spatial or temporal gradient [41]. In lakes, turnover in phytoplankton communities is related to the annual seasonal succession, e.g., the community in summer can be very different from that in winter. Species can appear, disappear or be replaced by other species [42].
To evaluate if physicochemical parameters influenced the patterns of alpha and beta diversity and density, a BIO-ENV was conducted. All analyses were performed with PRIMER v6.
3. Results
3.1. Physicochemical Parameters in Kusrüpuyewe Lake
The maximum depth recorded for each sampling station was in LK1 = 3 m, LK2 = 4 m and LK3 = 5 m at the austral summer, and during winter water depth increased (LK1 = 5 m, LK2 = 7 m and LK3 = 6 m). As shown in Table 1 and Figure S1, a temperature variation in the water column during summer and winter 2021 can be observed. Likewise, Table 1 indicates the average and variation ranges recorded for each sampling station in both climatic seasons. During summer, dissolved oxygen in the shallow lake was recorded as fluctuating between 64.6 and 103.3% saturation, and concentration fluctuated between 7.8 and 9.1 mg L−1. pH variation was 5.9–7.2. Temperature ranged from 18.6 to 23.5 °C. Conductivity ranged from 11.5 to 51.6 µS cm−1. Chlorophyll “a” variation was 1.4–4.9 µg L−1, and total suspended solids ranged from 1.6 to 5.8 mg L−1. On the other hand, in winter dissolved oxygen was recorded to fluctuate between 61.2 and 101.1% saturation with a concentration of 7.0–11.5 mg L−1. pH fluctuated between 6.0 and 7.9. Temperature ranged from 8.0 to 10.9 °C. Conductivity ranged from 10.1 to 37.6 µS cm−1. Chlorophyll “a” variation was 0.3–3.2 µg L−1, and the total suspended solids ranged from 1.3 to 3.9 mg/L−1.
3.2. Spatial Environmental Stability and Phytoplankton Assemblage in Kusrüpuyewe Lake
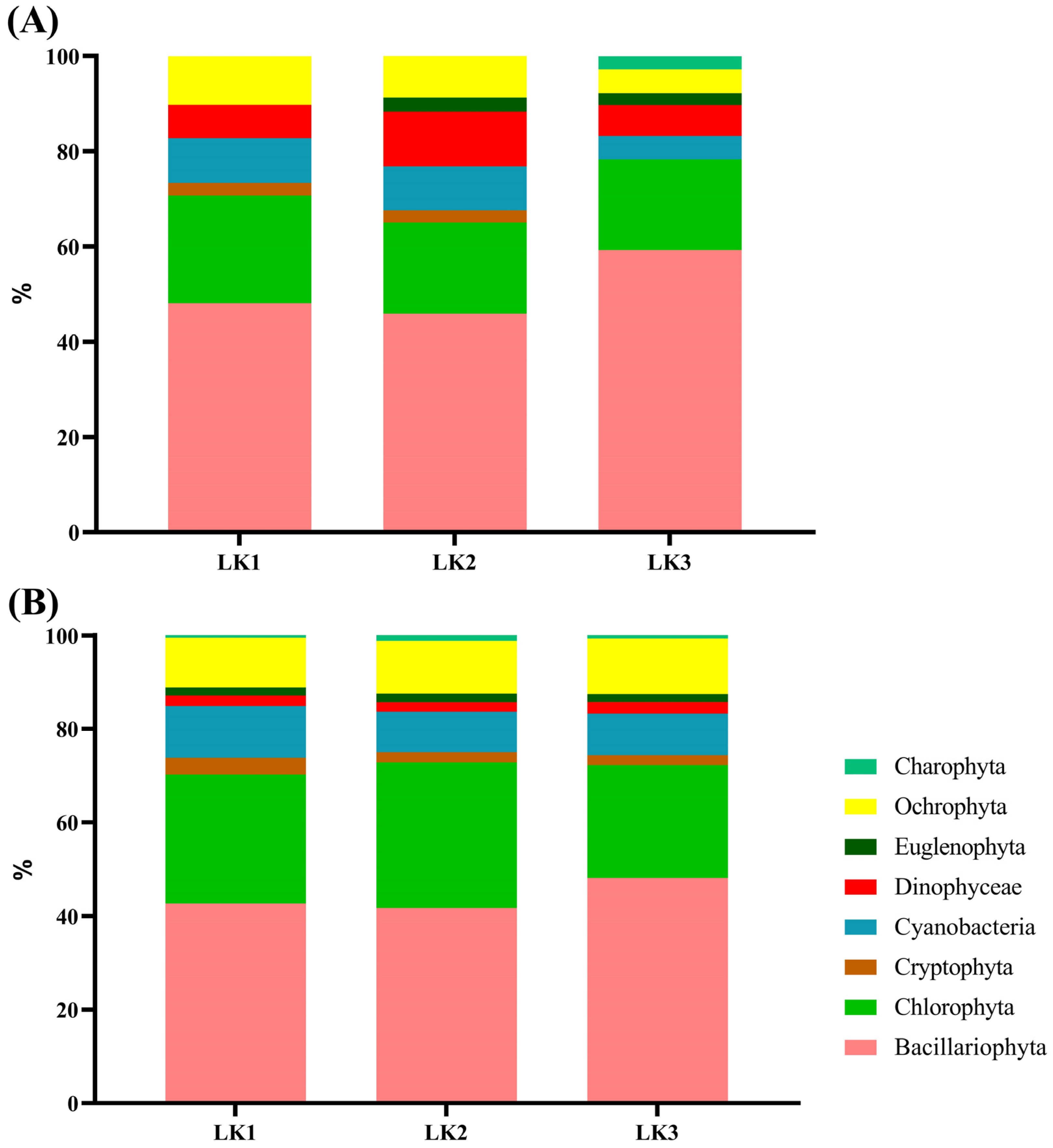
Of the 103 species recorded during summer, 50 belonged to the Phylum Bacillariophyta, 27 to Chlorophyta, 8 to Cyanobacteria, 7 to Ochrophyta, 4 to Charophyta, 2 to Cryptophyta, 2 to Euglenophyta and 3 to the Class Dinophyceae (Table 2 and Figure 2 and Figure 3). Alpha diversity obtained from the Chao-1 richness index was higher at station LK3 bottom (67 species). Shannon index was high (H′ > 4.5) at all stations, with the highest recorded at the station LK3 bottom (H′ = 6.0). Pielou’s evenness was J′ > 0.9 at all stations sampled. The effective number of Hill species (N1) varied between 22 and 66 species (Table 3).
During winter, 96 species were recorded; 36 were Bacillariophyta, 24 Chlorophyta, 16 Cyanobacteria, 9 Ochrophyta, 5 Dinophyceae, 3 Euglenophyta, 2 Charophyta and 1 Cryptophyta (Table 2 and Figure 3 and Figure 4). The alpha diversity was higher in the LK3 top (Chao-1 = 54 species), and the Shannon index was high at all the stations (H′ > 5). Pielou’s evenness was J′ > 0.9 at all stations. The effective number of Hill species (N1) varied between 36 and 46 (Table 3).
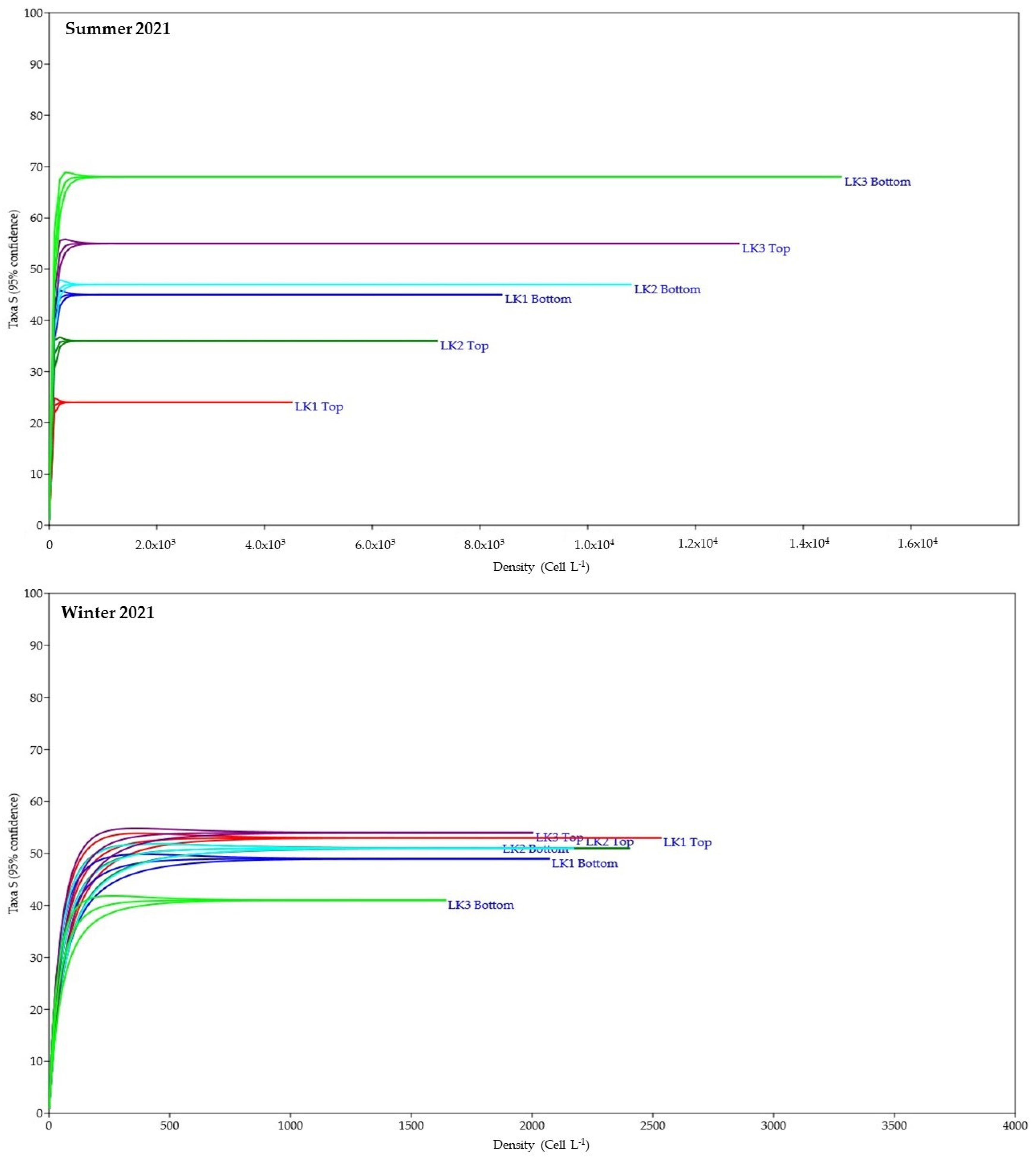
The major density was registered during the summer in LK3 bottom (584,520 cells L−1), and the winter major density was in LK1 top (101,800 cells L−1). The rarefaction curve indicated that the sampling effort was appropriate for both seasons and all stations, reaching the saturation at 24 species and >400 cells L−1 in the summer and 42 species and >500 cells L−1 in winter (Figure 5).
3.3. Spatial Environmental Heterogeneity and Species Turnover in Kusrüpuyewe Lake
We found slight differences in the beta diversity between the stations. In summer, the beta diversity, determined by the Sørensen dissimilarity index, was higher at station LK3, with an ecological stability of βSØR = 67.2%, while the Simpson dissimilarity index was higher in the LK1 station with a turnover percentage of βSIM = 87.5% (Table 3). Beta diversity in winter indicated that ecological stability (βSØR = 71.3%) and species turnover (βSIM = 70.7%) were higher at station LK3 (Table 3).
3.4. Temporal Variability of the Phytoplankton Assemblage
A total of 144 species of phytoplankton were recorded in the two seasons; taxa from the Phylum Bacillariophyta were the predominant group (63 species). Alpha diversity varied only slightly between seasons: species richness was 103 phytoplankton in summer and 96 in winter, and the Shannon index was H′ = 5.8 in summer and 5.7 in winter (Table 1). Furthermore, the Sørensen dissimilarity index indicated that the ecological stability of the Kusrüpuyewe Lake was similar in summer (βSØR = 62.9%) and winter (βSØR = 60.6%); the Simpson dissimilarity index indicated that species turnover was greater in summer (βSIM = 74.4%) than in winter (βSIM = 67.6%) (Table 3). The ANOSIM analysis showed significant differences in phytoplankton composition between seasons (p = 0.02). According to the SIMPER analysis, 40 species recorded in winter were not observed during the summer, while 49 species recorded in the summer were not observed during the winter, indicating a substantial turnover of species. We found no correlation between physicochemical parameters with abundance, alpha diversity and beta diversity (Table 2).
4. Discussion
4.1. Environmental Conditions and Heterogeneity
Kusrüpuyewe Lake is considered a wetland due to its low depth [43]. Maximum depth recorded in this study was four meters in summer and six meters in winter. Alpha and beta diversity indicated that Kusrüpuyewe shallow lake is stable and heterogeneous in both seasons (summer and winter). The high richness (144 species), diversity (H′ > 4) and evenness (J′ > 0.9) on phytoplankton species recorded indicate that we are in the presence of a pristine ecosystem. Pristine ecosystems are highly diverse and heterogeneous aquatic ecosystems [44], unlike homogeneous ecosystems with low diversity that present a poor environmental condition mainly due to anthropogenic factors such as modification of the hydrological regime, disconnection with contributing river, chemical contamination, fragmentation of the landscape and land use change [44,45]. Although less frequent, natural disturbances also affect the environmental quality of aquatic ecosystems, decimating biodiversity and deteriorating their chemical quality [7,46]. In India, shallow lakes with strong anthropogenic pressure recorded only 47 phytoplankton species [47], far below the 144 species recorded in our study.
Species turnover was high, especially in station LK1 (βSIM = 87.5%), located at the mouth of the main contributing, oligotrophic river. The connection to a major contributing source supports a stable environmental condition of the lake [22]. Studies have shown that contributing rivers positively influence the heterogeneity of an ecosystem, since they constantly export species and nutrients, increasing richness and favoring species turnover of the receiving water body [10,21]. However, during the winter, phytoplankton species turnover decreases, even though the contribution of the river is higher (rainfall regime); the slight decrease in turnover during the winter can be addressed to the phytoplankton ecology [44]. The high number of species is advantageous for the ecosystem since it increases the resilience and resistance of ecosystems to different disturbances [48,49]. This information is very useful for identifying areas with high conservation potential [50].
4.2. Temporal Variability and Species Turnover of Phytoplankton Communities
A temporal study of the alpha diversity in this shallow lake indicated high richness, diversity and evenness of phytoplankton communities (Chao-1 = 103 species; H′ = 5.8 and J = 0.9 respectively, Table 2), especially in summer when the light condition was optimal [51]. Studies involving temporal scales provide a better assessment of the heterogeneity and environmental health of the species diversity [52]. In both seasons, the dominant group was diatoms; the constant dominance of a taxonomic group is due to the constant contribution of a river, favoring the development of the species of that group [21]. In general, diatoms are the most diverse group with the greatest contribution in freshwater ecosystems. They are considered the most important group in phytoplankton communities, with a high plasticity giving them the ability to acclimatize to environmental fluctuations [53,54]. However, in ecosystems with a strong anthropic pressure, diatoms constituted the second dominant group while Cyanobacteria rise as the dominant group [47]. The fact that we recorded diatoms as the dominant group indicates the stable environmental status of this shallow lake. In ecosystems with deteriorated health, high diversities and a richness in Cyanobacteria and Euglenophyta are recorded. There is a direct relationship between the increase in density and the richness of cyanobacteria with land use changes [55]. In our study, we recorded a low richness and density of Cyanobacteria, attributable to the physicochemical and hydrological characteristics of Kusrüpuyewe Lake as it is an ecosystem that does not present a marked thermal stratification (Figure S1). Recent reports indicate that this ecosystem is mesotrophic in phosphorus and oligotrophic in nitrogen [56].
The minor dissimilarity in beta diversity observed in the monitoring stations during winter indicated differences in the phytoplankton assemblage between seasons (Table 3). These results are in line with a study carried out in Brazilian reservoirs with distinct dry and wet seasons [57] which is mainly due to changes in the ecosystem’s hydrological regimes [4,58]. Hydrological changes are relevant in the phytoplankton structuring in shallow ecosystems [21], since hydrological pulses cause important variations in the water level, influencing the dynamics of the system [59]. Rainfall increases between April and July (austral winter), increasing flows in the Pucopio River sub-basin, deepening this shallow lake [60]. As flow rates increase, erosion and, hence, turbidity increase, decreasing transparency and, thus, photosynthetic activity [61,62].
The dissimilarity of species in the Kusrüpuyewe Lake did not exceed βSØR 72% during the summer season, while the French lakes had a βSØR of 88.8% and Swedish lakes reached βSØR 88% [10,63]. It is worth mentioning that a strong dissimilarity in the phytoplankton communities is caused by the high turnover of species, since βSIM values greater than 80% are considered a stable ecosystem [10]. Temporal fluctuations with a high species turnover play an important role in the stability of the community structure and, therefore, in the ecosystem [21]. The interactions and temporal changes within the phytoplankton communities (density, richness and lack of a dominant species) make the aquatic ecosystem more constant over time; when richness does not present variation over time, it is related to low environmental variability (homogeneous environment) [54,59]. These results coincide with studies carried out by Cardoso [64] in shallow lakes of the Pantanal River basin, where the species richness was high in summer, with a slight decrease in winter due to the water increase.
In addition, the highest species turnover recorded was located in areas with native forest; at stations close to forest monocultures, the species turnover was βSIM < 70% (Figure 1B). It has been shown that sites with a forest presence play an important role in maintaining a high-water quality in shallow lakes, in contrast to agricultural grasslands or others [65]. Landscape fragmentation and land use changes are directly related to changes in phytoplankton communities, as well as in the turnover percentage [10,59,66]. The presence of unique species of phytoplankton is associated with the presence of forest monocultures or agricultural use [20]. It has been proven that changes in land use cause a decrease in beta diversity, negatively affecting diversity patterns [20,67,68]. In Kusrüpuyewe Lake, the phytoplankton communities are not strongly affected by forest monocultures. Pielou’s evenness and the effective number of species (N1) indicate that most of the species have similar densities; there is no dominance of a single species (Table 2 and Table 3). In ecosystems where the anthropic pressure from monocultures is strongest, the presence of dominant species is recorded, diversity decreases and species turnover is less than 45% [66]. However, the negative effects of the land use change in our ecosystems are not evident, as demonstrated by the results of the alpha and beta diversities. Peng et al. [55] indicates that monoculture plantations also have the capacity to absorb excess nutrients. The main effect on the Kusrüpuyewe Lake is the contribution of sediment to the water column [7,56].
Community functioning is more stable when there are heterogeneous conditions, i.e., rainfall regimes, hydrological changes, the presence of macrophytes [69]. SIMPER analysis, using Bray–Curtis dissimilarity, showed the species that were replaced during the study seasons (Table 2), demonstrating that Kusrüpuyewe Lake is temporarily stable, since a community is regulated through the turnover or replacement of species, which implies colonization and extinction of species, keeping the ecosystem stable or in balance [70].
4.3. Application of Alpha and Beta Diversities in Shallow Ecosystems
We recommend the use of alpha and beta diversities in environmental health studies to assess the ecological potential of the ecosystem, as we did with Kusrüpuyewe Lake. The application of both diversities provided valuable information on the heterogeneity and stability of the shallow lake. The use of different coefficients of dissimilarity has been very useful to evaluate different patterns of change in the community structure of phytoplankton [20], since these coefficients complement each other. While beta diversity provides information on the spatial and temporal variation of species composition throughout the ecosystem, alpha diversity provides information on a part of the ecosystem, that is, information is obtained from the richness or Shannon index from the monitored stations [41]. Both diversities measures, applied independently, do not provide sufficient information to make conservation decisions. Just as beta diversity cannot be based only on presence/absence (richness), it is important to incorporate the density values of each species recorded [12,13,14]. The application of beta diversity in freshwater ecosystems related to phytoplankton is still scarce; although they have been developed, they are not yet sufficient to understand the processes in time and space [10,41,71,72]. High beta diversity can be considered as an ecological insurance, allowing the maintenance of alpha diversity even under strong environmental fluctuations, thus maintaining the functioning of the ecosystem [10].
Shallow lakes, such as the Kusrüpuyewe Lake, are excellent models for ecological research and allow for the evaluation of the effects of climate change on biological communities through the determination of community structure and ecosystem stability [59]. Therefore, a solid knowledge on the near-pristine status is necessary. The application of alpha diversity indicators and beta diversity indicators was very useful because these ecosystems have discrete boundaries, are small (facilitating sampling) and are representative, which is demonstrated by applying the rarefaction curve (Figure 5). In diversity studies, it is important to determine that the sampling effort was adequate, avoiding an underestimation of the population; with an adequate sampling effort, all members of the population would be considered [73,74,75]. In addition, they can be used as sentinel ecosystems or early warning systems for the long-term effects of climate change, warning about a potential deterioration of the ecosystem [6,73].
5. Conclusions
Alpha diversity and beta diversity, as ecological indicators, helped us to understand the degree of heterogeneity in the Kusrüpuyewe shallow lake. Phytoplankton communities can reflect the trophic conditions of water bodies, and this shallow lake appears to be in a stable environmental condition. It is important to maintain these conditions; therefore, it is necessary and imperative to establish conservation policies in this ecosystem, by protecting the catchment and by maintaining or restoring the land cover with native forest. Native forests have a higher capacity to absorb pollutants and prevent them from entering than exotic monocultures. High forest diversity has been shown to increase the buffering capacity in aquatic ecosystems by reducing the effects of environmental impacts such as extreme weather or the deposition of various pollutants. The application of beta diversity temporally in this ecosystem indicated that the turnover was not as evident as expected. However, at present, forest monocultures have not been harvested, and it would be interesting to carry out a temporary post-harvest assessment of the effects on the alpha and beta diversities of the phytoplankton communities in this ecosystem.
Supplementary Materials
The following supporting information can be downloaded at https://www.mdpi.com/article/10.3390/w16020274/s1: Figure S1. Temperature profile in each station monitoring during summer and winter. Letters correspond to the following: (a) LK1 summer; (b) LK2 summer; (c) LK3 summer; (d) LK1 winter; (e) LK2 winter; (f) LK3 winter.
Author Contributions
Conceptualization, methodology, investigation, formal analysis, writing—original draft preparation, writing—review and editing, C.R.-H. and N.F. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the collaboration agreement Ulagos/CONADI, grant number “DU1815”. The APC was funded by Vicerrectoria de Investigación y Postgrado of the Universidad de Los Lagos.
Data Availability Statement
Data are contained within the article and Supplementary Materials.
Acknowledgments
We thank Víctor Cuchipe, a member of the Rayen Ko-Lafquen indigenous community and Limnology Laboratory members for supporting us in the field campaigns; Lafayette Eaton and Guntram Weithoff for improving the English and their valuable comments; Vicerrectoria de Investigación y Postgrado of the Universidad de Los Lagos and National Corporation for Indigenous Development (CONADI for initials in Spanish) for funding this study.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Morales, M.; Aflalo, C.; Bernard, O. Microalgal lipids: A review of lipids potential and quantification for 95 phytoplankton species. Biomass Bioenergy 2021, 150, 106108. [Google Scholar] [CrossRef]
- Özkan, K.; Jeppesen, E.; Søndergaard, M.; Lauridsen, T.L.; Liboriussen, L.; Svenning, J.C. Contrasting roles of water chemistry, lake morphology, land-use, climate and spatial processes in driving phytoplankton richness in the Danish landscape. Hydrobiologia 2013, 710, 173–187. [Google Scholar] [CrossRef]
- Phillips, G.; Lyche-Solheim, A.; Skjelbred, B.; Mischke, U.; Drakare, S.; Free, G.; Järvinen, M.; de Hoyos, C.; Morabito, G.; Poikane, S.; et al. A phytoplankton trophic index to assess the status of lakes for the Water Framework Directive. Hydrobiologia 2013, 704, 75–95. [Google Scholar] [CrossRef]
- Rodrigues, L.C.; Simões, N.R.; Bovo-Scomparin, V.M.; Jati, S.; Santana, N.F.; Roberto, M.C.; Train, S. Phytoplankton alpha diversity as an indicator of environmental changes in a neotropical floodplain. Ecol. Indic. 2015, 48, 334–341. [Google Scholar] [CrossRef]
- Yang, B.; Jiang, Y.J.; He, W.; Liu, W.X.; Kong, X.Z.; Jørgensen, S.E.; Xu, F.-L. The tempo-spatial variations of phytoplankton diversities and their correlation with trophic state levels in a large eutrophic Chinese lake. Ecol. Indic. 2016, 66, 153–162. [Google Scholar] [CrossRef]
- Zhang, M.; Yu, Y.; Yang, Z.; Kong, F. Deterministic diversity changes in freshwater phytoplankton in the Yunnan-Guizhou Plateau lakes in China. Ecol. Indic. 2016, 63, 273–281. [Google Scholar] [CrossRef]
- Fuentes, N.; Arriagada, A. Long-term responses of macroinvertebrate assemblages to the 2011 eruption of the Puyehue-Cordón Caulle volcanic complex, Chile. Sci. Total Environ. 2022, 807, 150978. [Google Scholar] [CrossRef]
- Carvalho, L.; Poikane, S.; Lyche, S.A.; Phillips, G.; Borics, G.; Catalan, J.; De Hoyos, C.; Drakare, S.; Dudley, B.J.; Järvinen, M.; et al. Strength and uncertainty of phytoplankton metrics for assessing eutrophication impacts in lakes. Hydrobiologia 2013, 704, 127–140. [Google Scholar] [CrossRef]
- Whittaker, R.H. Evolution and Measurement of Species Diversity. Taxon 1972, 21, 213–251. [Google Scholar] [CrossRef]
- Maloufi, S.; Catherine, A.; Mouillot, D.; Louvard, C.; Couté, A.; Bernard, C.; Troussellier, M. Environmental heterogeneity among lakes promotes hyper β-diversity across phytoplankton communities. Freshw. Biol. 2016, 61, 633–645. [Google Scholar] [CrossRef]
- Siqueiros-Beltrones, D.A. Una paradoja sobre uniformidad vs. Orden y estabilidad en la medida de la diversidad de especies según la teoría de la información. Ludus Vitalis 2005, 13, 83–92. [Google Scholar]
- Baselga, A. Partitioning the turnover and nestedness components of beta diversity. Glob. Ecol. Biogeogr. 2010, 19, 134–143. [Google Scholar] [CrossRef]
- Baselga, A. Separating the two components of abundance-based dissimilarity: Balanced changes in abundance vs. abundance gradients. Methods Ecol. Evol. 2013, 4, 552–557. [Google Scholar] [CrossRef]
- Baselga, A.; Gómez-Rodríguez, C. Diversidad alfa, beta y gamma: ¿cómo medimos diferencias entre comunidades biológicas? Nov. Acta Científica Compostel. 2019, 26, 39–45. [Google Scholar]
- Alves-De-Souza, C.; Benevides, T.S.; Santos, J.B.O.; Von Dassow, P.; Guillou, L.; Menezes, M. Does environmental heterogeneity explain temporal β diversity of small eukaryotic phytoplankton? Example from a tropical eutrophic coastal lagoon. J. Plankton Res. 2017, 39, 698–714. [Google Scholar] [CrossRef]
- Zorzal-Almeida, S.; Bini, L.M.; Bicudo, D.C. Beta diversity of diatoms is driven by environmental heterogeneity, spatial extent and productivity. Hydrobiologia 2017, 800, 7–16. [Google Scholar] [CrossRef]
- Loreau, M.; Naeem, S.; Inchausti, P.; Bengtsson, J.; Grime, J.P.; Hector, A.; Hooper, D.U.; Huston, M.A.; Raffaelli, D.; Schmid, B.; et al. Biodiversity and Ecosystem Functioning: Current Knowledge and Future Challenges. Sci. Compass 2001, 294, 804–808. [Google Scholar] [CrossRef]
- Gonzalez, A.; Loreau, M. The causes and consequences of compensatory dynamics in ecological communities. Annu. Rev. Ecol. Evol. Syst. 2009, 40, 393–414. [Google Scholar] [CrossRef]
- Ríos-Henríquez, C.; Fuentes, N.; Tobar, C.N.; Rau, J.R.; Cruces, F. Planktonic diatom assemblage seasonal diversity used to assess environmental health in a coastal wetland of southern Chile. Gayana Bot. 2020, 77, 139–151. [Google Scholar] [CrossRef]
- Tavares-Costa, A.P.; Oliveira-Crossetti, L.; Hartz, S.M.; Becker, F.G.; Hepp, L.U.; Bohnenberger, J.E.; Lima, M.S.; Guimarães, T.; Schneck, F. Land cover is the main correlate of phytoplankton beta diversity in subtropical coastal shallow lakes. Aquat. Ecol. 2020, 54, 1015–1028. [Google Scholar] [CrossRef]
- Frau, D.; Pineda, A.; Mayora, G.; Devercelli, M. Phytoplankton taxonomic and functional diversity in two shallow alluvial lakes with contrasting river connectivity. Aquat. Sci. 2022, 84, 26. [Google Scholar] [CrossRef]
- Fuentes, N.; Arriagada, A. New records of the endangered southern river otter Lontra provocax, with notes on its diet, in threatened wetlands of southern Chile. Oryx 2023, 57, 76–79. [Google Scholar] [CrossRef]
- Santibáñez, F. El cambio climático y los recursos hídricos de Chile. In Agricultura Chilena, Reflexiones y Desafíos Al 2030, 1st ed.; Apey, A., Barrera, D., Rivas, T., Eds.; Oficina de Estudios y Políticas Agrarias (ODEPA): Santiago, Chile, 2016; pp. 147–177. [Google Scholar]
- Lemmens, P.; Mergeay, J.; de Bie, T.; Van Wichelen, J.; de Meester, L.; Declerck, S.A.J. How to Maximally Support Local and Regional Biodiversity in Applied Conservation? Insights from Pond Management. PLoS ONE 2013, 8, e72538. [Google Scholar] [CrossRef] [PubMed]
- Celewicz-Goldyn, S.; Kuczynska-Kippen, N. Ecological value of macrophyte cover in creating habitat for microalgae (diatoms) and zooplankton (rotifers and crustaceans) in small field and forest water bodies. PLoS ONE 2017, 12, e0177317. [Google Scholar] [CrossRef] [PubMed]
- Kollár, J.; Fránková, M.; Hašler, P.; Letáková, M.; Poulíčková, A. Epiphytic diatoms in lotic and lentic waters-Diversity and representation of species complexes. Fottea 2015, 15, 259–271. [Google Scholar] [CrossRef]
- Downing, J.A. Emerging global role of small lakes and ponds: Little things mean a lot. Limnetica 2010, 29, 9–24. [Google Scholar] [CrossRef]
- Céréghino, R.; Boix, D.; Cauchie, H.M.; Martens, K.; Oertli, B. The ecological role of ponds in a changing world. Hydrobiologia 2014, 723, 1–6. [Google Scholar] [CrossRef]
- Biggs, J.; Williams, P.; Whitfield, M.; Nicolet, P.; Weatherby, A. 15 Years of pond assessment in Britain: Results and lessons learned from the work of Pond Conservation. Aquat. Conserv. Mar. Freshw. Ecosyst. 2005, 15, 693–714. [Google Scholar] [CrossRef]
- Bella, V.D.; Mancini, L. Freshwater diatom and macroinvertebrate diversity of coastal permanent ponds along a gradient of human impact in a Mediterranean eco-region. Hydrobiologia 2009, 634, 25–41. [Google Scholar] [CrossRef]
- Pereira, H.M.; Navarro, L.M.; Martins, I.S. Global biodiversity change: The Bad, the good, and the unknown. Annu. Rev. Environ. Resour. 2012, 37, 25–50. [Google Scholar] [CrossRef]
- Gotelli, N.J.; Colwell, R.K. Quantifying biodiversity: Procedures and pitfalls in the measurement and comparison of species richness. Ecol. Lett. 2001, 4, 379–391. [Google Scholar] [CrossRef]
- Cermeño, P.; Teixeira, I.G.; Branco, M.; Figueiras, F.G.; Marañón, E. Sampling the limits of species richness in marine phytoplankton communities. J. Plankton Res. 2014, 36, 1135–1139. [Google Scholar] [CrossRef]
- Utermöhl, H. Zur vervollkommnung der quantitativen phytoplankton methodic. Int. Ver. Theor. Angew. Limnol. 1958, 9, 1–38. [Google Scholar]
- Parra, O.O.; González, M.A.; Dellarossa, V.; Rivera, P.; Orellana, M. Manual Taxonómico del Fitoplancton de Agua Continentales Con Especial Referencia al Fitoplancton en Chile; Editorial Universidad de Concepción: Concepción, Chile, 1982; pp. 1–20. [Google Scholar]
- Bahls, L.; Boynton, B.; Johnston, B. Atlas of diatoms (Bacillariophyta) from diverse habitats in remote regions of western Canada. PhytoKeys 2018, 186, 1–186. [Google Scholar] [CrossRef] [PubMed]
- Nusch, E. Comparison of different methods for chlorophyll and phaeopigment determination. Arch. Hydrobiol. 1980, 14, 14–36. [Google Scholar]
- APHA. Standard Methods for the Examination of Water and Wastewater, 21st ed.; American Public Health Association; American Water Works Association; Water Environment Federation: Washington, DC, USA, 2005; p. 541. [Google Scholar]
- Siqueiros-Beltrones, D.A.; Argumedo-Hernández, U.; Hernández-Almeida, O.U. High species diversity (H) of benthic diatoms in a coastal lagoon located within a natural protected area. Hidrobiologica 2017, 27, 293–300. [Google Scholar] [CrossRef]
- Brown, J.H.; Ernest, S.K.M.; Parody, J.M.; Haskell, J.P. Regulation of diversity: Maintenance of species richness in changing environments. Oecologia 2001, 126, 321–332. [Google Scholar] [CrossRef]
- Legendre, P. Interpreting the replacement and richness difference components of beta diversity. Glob. Ecol. Biogeogr. 2014, 23, 1324–1334. [Google Scholar] [CrossRef]
- Zhang, M.; Shi, X.; Chen, F.; Yang, Z.; Yu, Y. The underlying causes and effects of phytoplankton seasonal turnover on resource use efficiency in freshwater lakes. Ecol. Evol. 2021, 11, 8897–8909. [Google Scholar] [CrossRef]
- Secretaría de la Convención de Ramsar. Designación de Sitios Ramsar: Marco Estratégico y Lineamientos Para el Desarrollo Futuro de la Lista de Humedales de Importancia Internacional, 4th ed.; Manuales Ramsar Para el Uso Racional de Los Humedales: Ramsar Gland, Switzerland, 2010; Volume 17, p. 130. [Google Scholar]
- Rahel, F.J. Homogenization of freshwater faunas. Annu. Rev. Ecol. Syst. 2002, 33, 291–315. [Google Scholar] [CrossRef]
- Lougheed, V.L.; Mcintosh, M.D.; Parker, C.A.; Stevenson, R.J. Wetland degradation leads to homogenization of the biota at local and landscape scales. Freshw. Biol. 2008, 53, 2402–2413. [Google Scholar] [CrossRef]
- Fuentes, N.; Gómez, L.; Venegas, H.; Rau, J.R. Total devastation of river macroinvertebrates following a volcanic eruption in southern Chile. Ecosphere 2020, 11, e03105. [Google Scholar] [CrossRef]
- Jindal, R.; Thakur, R.K.; Singh, U.B.; Ahluwalia, A.S. Phytoplankton dynamics and species diversity in a shallow eutrophic, natural mid-altitude lake in Himachal Pradesh (India): Role of physicochemical factors. Chem. Ecol. 2014, 30, 328–338. [Google Scholar] [CrossRef]
- Downing, A.L.; Leibold, M.A. Species richness facilitates ecosystem resilience in aquatic food webs. Freshw. Biol. 2010, 55, 2123–2137. [Google Scholar] [CrossRef]
- Mori, A.S.; Isbell, F.; Seidl, R. β-Diversity, Community Assembly, and Ecosystem Functioning. Trends Ecol. Evol. 2018, 33, 549–564. [Google Scholar] [CrossRef]
- Andermann, T.; Antonelli, A.; Barrett, R.L.; Silvestro, D. Estimating Alpha, Beta, and Gamma Diversity through Deep Learning. Front. Plant Sci. 2022, 13, 839407. [Google Scholar] [CrossRef]
- Roelke, D.L.; Spatharis, S. Phytoplankton Succession in Recurrently Fluctuating Environments. PLoS ONE 2015, 10, e0121392. [Google Scholar] [CrossRef]
- Gering, J.C.; Crist, T.O.; Veech, J.A. Additive partitioning of species diversity across multiple spatial scales: Implications for regional conservation of biodiversity. Conserv. Biol. 2003, 17, 488–499. [Google Scholar] [CrossRef]
- Bussard, A.; Corre, E.; Hubas, C.; Duvernois-Berthet, E.; Le Corguillé, G.; Jourdren, L.; Coulpier, F.; Claquin, P.; Lopez, P.J. Physiological adjustments and transcriptome reprogramming are involved in the acclimation to salinity gradients in diatoms. Environ. Microbiol. 2017, 19, 909–925. [Google Scholar] [CrossRef]
- Stefanidou, N.; Katsiapi, M.; Tsianis, D.; Demertzioglou, M.; Michaloudi, E.; Moustaka-Gouni, M. Patterns in Alpha and Beta Phytoplankton Diversity along a Conductivity Gradient in Coastal Mediterranean Lagoons. Diversity 2020, 12, 38. [Google Scholar] [CrossRef]
- Peng, X.; Zhang, L.; Li, Y.; Lin, Q.; Chao He, C.; Huang, S.; Li, H.; Zhang, X.; Liu, B.; Ge, F.; et al. The changing characteristics of phytoplankton community and biomass in subtropical shallow lakes: Coupling effects of land use patterns and lake morphology. Water Res. 2021, 200, 117235. [Google Scholar] [CrossRef]
- Fuentes, N.; Arriagada, A.; Araos, F.; Ríos-Henríquez, C.; Riquelme, W.; Iwama, A.; Cárdenas, L.; Soto, R.; Escobar, A.; Henríquez, G. Caracterización Ambiental y Social de Laguna Trinidad y Estero Pucopio, Comunas de San Pablo y San Juan de la Costa, Provincia de Osorno, 1st ed.; Universidad de Los Lagos y Corporación Nacional de Desarrollo Indígena: Osorno, Chile, 2021; p. 128. [Google Scholar]
- Santana, L.M.; Weithoff, G.; Ferragut, C. Seasonal and spatial functional shifts in phytoplankton communities of five tropical reservoirs. Aquat. Ecol. 2017, 51, 531–543. [Google Scholar] [CrossRef]
- Simões, N.R.; Lansac-Tôha, F.A.; Velho, L.F.M.; Bonecker, C.C. Intra and inter-annual structure of zooplankton communities in floodplain lakes: A long-term ecological research study. Rev. Biol. Trop. 2012, 60, 1819–1836. [Google Scholar] [CrossRef]
- Pineda, A.; Peláez, Ó.; Dias, J.D.; Segovia, B.T.; Bonecker, C.C.; Velho, L.F.M.; Rodrigues, L.C. The El Niño Southern Oscillation (ENSO) is the main source of variation for the gamma diversity of plankton communities in subtropical shallow lakes. Aquat. Sci. 2019, 81, 49. [Google Scholar] [CrossRef]
- Nabout, J.C.; Nogueira, I.S.; Oliveira, L.G. Phytoplankton community of floodplain lakes of the Araguaia River, Brazil, in the rainy and dry seasons. J. Plankton Res. 2006, 28, 181–193. [Google Scholar] [CrossRef]
- Rasconi, S.; Gall, A.; Winter, K.; Kainz, M.J. Increasing water temperature triggers dominance of small freshwater plankton. PLoS ONE 2015, 10, e0140449. [Google Scholar] [CrossRef]
- Train, S.; Rodrigues, L.C. Temporal fluctuations of the phytoplankton community of the Baia River, in the upper Parana River floodplain, Mato Grosso do Sul, Brazil. Hydrobiologia 1997, 361, 125–134. [Google Scholar] [CrossRef]
- Angeler, D.G. Revealing a conservation challenge through partitioned long-term beta diversity: Increasing turnover and decreasing nestedness of boreal Lake Metacommunities. Divers. Distrib. 2013, 19, 772–781. [Google Scholar] [CrossRef]
- Cardoso, S.J.; Roland, F.; Loverde-Oliveira, S.M.; Huszar, V.L. Phytoplankton abundance, biomass and diversity within and between Pantanal wetland habitats. Limnologica 2012, 42, 235–241. [Google Scholar] [CrossRef]
- Leon-Múñoz, J.; Echeverría, C.; Marcé, R.; Riss, W.; Sherman, B.; Iriarte, J.L. The combined impact of land use change and aquaculture on sediment and water quality in oligotrophic Lake Rupanco (North Patagonia, Chile, 40.8 S). J. Environ. Manag. 2013, 128, 283–291. [Google Scholar] [CrossRef]
- Lu, W.; Zhang, S.; Zhou, Z.; Wang, Y.; Wang, S. Effects of Land Use and Physicochemical Factors on Phytoplankton Community Structure: The Case of Two Fluvial Lakes in the Lower Reach of the Yangtze River, China. Diversity 2023, 15, 180. [Google Scholar] [CrossRef]
- Siqueira, T.; Lacerda, C.G.L.T.; Saito, V.S. How Does Landscape Modification Induce Biological Homogenization in Tropical Stream Metacommunities? Biotropica 2015, 47, 509–516. [Google Scholar] [CrossRef]
- Winegardner, A.K.; Legendre, P.; Beisner, B.E.; Gregory-Eaves, I. Diatom diversity patterns over the past c. 150 years across the conterminous United States of America: Identifying mechanisms behind beta diversity. Glob. Ecol. Biogeogr. 2017, 26, 1303–1315. [Google Scholar] [CrossRef]
- De Boeck, H.J.; Bloor, J.M.G.; Kreyling, J.; Ransijn, J.C.G.; Nijs, I.; Jentsch, A.; Zeiter, M. Patterns and drivers of biodiversity–stability relationships under climate extremes. J. Ecol. 2018, 106, 890–902. [Google Scholar] [CrossRef]
- Gotelli, N.J.; Shimadzu, H.; Dornelas, M.; McGill, B.; Moyes, F.; Magurran, A.E. Community-level regulation of temporal trends in biodiversity. Sci. Adv. 2017, 3, e1700315. [Google Scholar] [CrossRef]
- Stomp, M.; Huisman, J.; Mittelbach, G.G.; Litchman, E.; Klausmeier, C.A. Large-scale biodiversity patterns in freshwater phytoplankton. Ecology 2011, 92, 2096–2107. [Google Scholar] [CrossRef]
- Stein, A.; Gerstner, K.; Kreft, H. Environmental heterogeneity as a universal driver of species richness across taxa, biomes and spatial scales. Ecol. Lett. 2014, 17, 866–880. [Google Scholar] [CrossRef]
- De Meester, L.; Declerck, S.; Stoks, R.; Louette, G.; Van De Meutter, F.; De Bie, T.; Michels, E.; Brendonck, L. Ponds and pools as model systems in conservation biology, ecology and evolutionary biology. Aquat. Conserv. Mar. Freshw. Ecosyst. 2005, 15, 715–725. [Google Scholar] [CrossRef]
- Hortal, J.; Nabout, J.C.; Calatayud, J.; Carneiro, F.M.; Padial, A.; Santos, A.M.C.; Siqueira, T.; Bokma, F.; Bini, L.M.; Ventura, M. Perspectives on the use of lakes and ponds as model systems for macroecological research. J. Limnol. 2014, 73, 46–60. [Google Scholar] [CrossRef]
- Almeida-Gomes, M.; Valente-Neto, F.; Pacheco, E.O.; Ganci, C.C.; Leibold, M.A.; Melo, A.S.; Provete, D.B. How Does the Landscape Affect Metacommunity Structure? A Quantitative Review for Lentic Environments. Curr. Landsc. Ecol. Rep. 2020, 5, 68–75. [Google Scholar] [CrossRef]
Figure 1.
Map of the location of the Kusrüpuyewe Lake water network and land use coverage. (A) Map of South America and Chile, location of the study area; (B) location of sampling points and land use.
Figure 1.
Map of the location of the Kusrüpuyewe Lake water network and land use coverage. (A) Map of South America and Chile, location of the study area; (B) location of sampling points and land use.

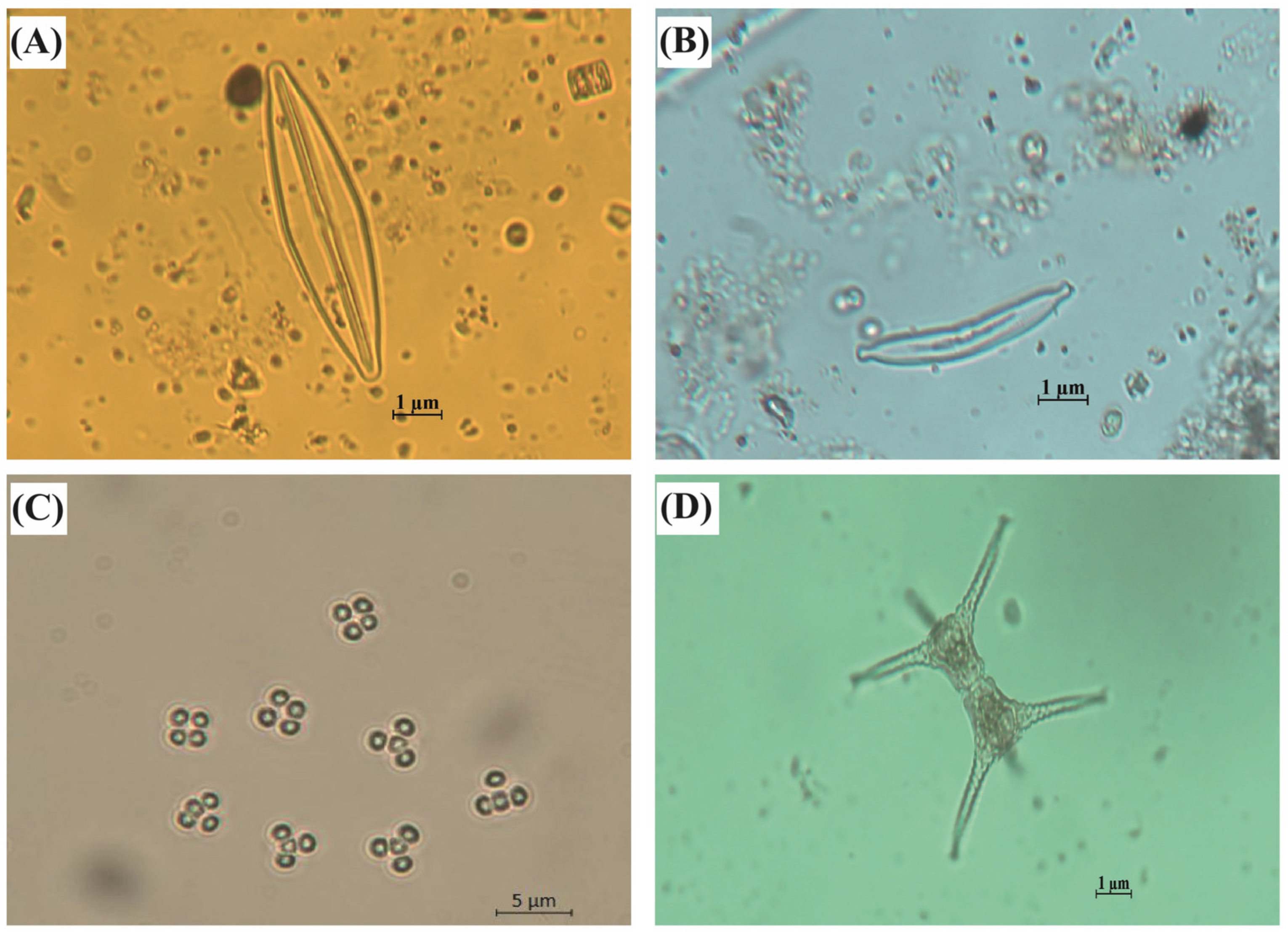
Figure 2.
Photographs of principal phytoplankton representatives during the austral summer 2021 season in Kusrüpuyewe Lake. (A) Amphipleura lindheimeri; (B) Hannaea arcus; (C) Tetraspora lacustris; (D) Staurastrum paradoxum.
Figure 2.
Photographs of principal phytoplankton representatives during the austral summer 2021 season in Kusrüpuyewe Lake. (A) Amphipleura lindheimeri; (B) Hannaea arcus; (C) Tetraspora lacustris; (D) Staurastrum paradoxum.

Figure 3.
Percentages of phytoplankton present in the Kusrüpuyewe Lake: (A) corresponds to the austral summer 2021 season; (B) the austral winter 2021 season.
Figure 3.
Percentages of phytoplankton present in the Kusrüpuyewe Lake: (A) corresponds to the austral summer 2021 season; (B) the austral winter 2021 season.

Figure 4.
Photographs of principal phytoplankton representatives during the austral winter 2021 season in Kusrüpuyewe Lake. (A) Mallomonas alpina; (B) Dolichospemum sp; (C) Neidium bisulcatum; (D) Pleurosgima sp.
Figure 4.
Photographs of principal phytoplankton representatives during the austral winter 2021 season in Kusrüpuyewe Lake. (A) Mallomonas alpina; (B) Dolichospemum sp; (C) Neidium bisulcatum; (D) Pleurosgima sp.

Figure 5.
Rarefaction curve to estimate the sampling effort at each monitoring station in summer and winter 2021 in Kusrüpuyewe Lake.
Figure 5.
Rarefaction curve to estimate the sampling effort at each monitoring station in summer and winter 2021 in Kusrüpuyewe Lake.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Results of physicochemical parameters: dissolved oxygen (DO percentage% and concentration mg L−1); pH; temperature (T °C), conductivity (Con µS cm−1), chlorophyll “a” (Chl a µg L−1) and total suspended solid (TSS mg L−1) at monitoring stations in Kusrüpuyewe Lake during summer and winter.
Table 1.
Results of physicochemical parameters: dissolved oxygen (DO percentage% and concentration mg L−1); pH; temperature (T °C), conductivity (Con µS cm−1), chlorophyll “a” (Chl a µg L−1) and total suspended solid (TSS mg L−1) at monitoring stations in Kusrüpuyewe Lake during summer and winter.
| Depth | DO% | DO mg L−1 | pH | T °C | Con µS cm −1 | Chl a µg L−1 | TSS mg L−1 | ||
|---|---|---|---|---|---|---|---|---|---|
| Summer | |||||||||
| LK1 | Top | 0–1 m | 100.6 (98.2–102) | 8.6 (8.4–8.7) | 6.9 (6.5–7.2) | 23.3 (23.1–23.4) | 46.5 (46.4–46.5) | 3.9 (2.6–4.9) | 2.1 (1.6–2.7) |
| Bottom | 2–3 m | 98.1 (91.4–100.8) | 8.5 (7.8–9.1) | 7.1 (7.0–7.2) | 22.6 (22.1–23.1) | 45.9 (45.5–46.6) | 3.0 (2.7–3.4) | 2.0 (1.6–2.8) | |
| LK2 | Top | 0–1 m | 97.9 (97.8–98.1) | 8.3 (8.3–8.4) | 6.7 (6.5–7.2) | 23.4 (23.3–23.5) | 43.5 (11.7–51.6) | 1.9 (1.4–2.3) | 2.7 (2.6–3.1) |
| Bottom | 3–4 m | 90.2 (83.4–95.8) | 7.8 (7.2–8.2) | 7.2 (7.1–7.2) | 22.7 (22.4–22.9) | 46.1 (45.9–46.3) | 2.7 (2.4–3.1) | 4.4 (3.3–5.8) | |
| LK3 | Top | 0–1 m | 94.8 (84.4–103.2) | 8.0 (7.2–8.7) | 6.8 (6.3–7.1) | 21.7 (21.5–21.9) | 33.7 (11.5–46.6) | 3.3 (2.3–4.1) | 3.9 (3.7–4.4) |
| Bottom | 4–5 m | 83.5 (76.2–89.9) | 7.9 (5.9–8.7) | 6.9 (6.9–7.1) | 18.7 (18.6–19.1) | 44.4 (43.6–46.1) | 2.5 (1.5–3.4) | 3.9 (3.5–4.2) | |
| Winter | |||||||||
| LK1 | Top | 0–1 m | 85.4 (80.1–92.9) | 8.9 (7.4–10.4) | 6.9 (6.1–7.1) | 10.6 (10.5–10.7) | 20.4 (10.1–25.4) | 0.8 (0.3–1.2) | 2.1 (1.9–2.5) |
| Bottom | 4–5 m | 65.5 (62.0–72.8) | 7.5 (7.1–8.2) | 6.9 (6.8–7.1) | 9.5 (9.3–9.9) | 24.7 (24.6–25.1) | 0.7 (0.5–0.9) | 1.6 (1.5–1.6) | |
| LK2 | Top | 0–1 m | 90.3 (80.9–101.1) | 10.0 (8.9–11.3) | 7.8 (7.5–7.8) | 10.7 (10.6–10.7) | 21.7 (10.1–27.1) | 2.8 (2.3–3.2) | 2.7 (2.6–2.8) |
| Bottom | 6–7 m | 68.4 (66.9–69.8) | 8.1 (7.9–8.2) | 7.9 (7.8–7.9) | 8.1 (8.1–8.2) | 22.6 (22.6–22.7) | 0.9 (0.8–1.1) | 1.5 (1.3–1.8) | |
| LK3 | Top | 0–1 m | 89.2 (80.1–99.3) | 9.9 (9.1–11.5) | 6.5 (6.2–7.1) | 10.2 (10–10.3) | 24.2 (19.1–37.6) | 2.3 (1.7–3.2) | 2.9 (2.5–3.9) |
| Bottom | 5–6 m | 70.4 (69.2–71.7) | 8.1 (8.1–8.2) | 6.9 (6.9–7.1) | 9.0 (8.9–9.7) | 23.4 (23.1–25.4) | 1.4 (1.1–1.8) | 3.2 (3–3.4) | |
Table 2.
Density of phytoplankton species (×103 cells L−1) in Kusrüpuyewe Lake, results of analysis of similitude (ANOSIM), BIO-ENV and similitude percentage analyses (SIMPER) between summer and winter. Letters correspond to the following: (a) phytoplankton species present in summer only; (b) phytoplankton species present in winter only; (c) phytoplankton species present in both seasons.
Table 2.
Density of phytoplankton species (×103 cells L−1) in Kusrüpuyewe Lake, results of analysis of similitude (ANOSIM), BIO-ENV and similitude percentage analyses (SIMPER) between summer and winter. Letters correspond to the following: (a) phytoplankton species present in summer only; (b) phytoplankton species present in winter only; (c) phytoplankton species present in both seasons.
| Summer | Winter | SIMPER | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Phylum | Class | Species | LK1 | LK2 | LK3 | LK1 | LK2 | LK3 | |||||||
| Top | Bottom | Top | Bottom | Top | Bottom | Top | Bottom | Top | Bottom | Top | Bottom | ||||
| Bacillariophyta | Bacillariophyceae | Achnanthes affinis | 6.1 | 6.6 | 5.8 | 9.5 | 9.8 | 8.6 | a | ||||||
| Achnanthes exigua | 6.6 | 7.4 | 7.9 | 9.8 | 8.7 | 10.6 | 2.6 | 3.3 | 1.1 | 2.7 | c | ||||
| Achnanthes hauckiana | 8.8 | 9.3 | 8.7 | a | |||||||||||
| Achnanthes lanceolata | 2.8 | 2.8 | 0.8 | 2.0 | b | ||||||||||
| Amphora veneta | 1.3 | 0.5 | b | ||||||||||||
| Amphipleura lindheimeri | 5.4 | 6.8 | a | ||||||||||||
| Asterionella formosa | 21.6 | 12.8 | 11.4 | 9.6 | 10.2 | 8.8 | 2.4 | 1.8 | 3.1 | 3.2 | 2.8 | c | |||
| Cocconeis placentula | 10.3 | 3.5 | 3.2 | c | |||||||||||
| Cymbella lanceolata | 6.6 | 7.5 | a | ||||||||||||
| Cymbella minuta | 8.0 | 1.9 | 2.0 | 2.2 | 2.8 | 2.4 | c | ||||||||
| Cymbella naviculiforme | 8.2 | 9.1 | 8.0 | a | |||||||||||
| Diatoma tenue | 7.1 | 0.6 | 0.6 | c | |||||||||||
| Epithemia adnata | 8.0 | a | |||||||||||||
| Eunotia bidens | 7.9 | a | |||||||||||||
| Eunotia major | 4.8 | 6.4 | 7.5 | 9.1 | 11.6 | 8.1 | a | ||||||||
| Eunotia juettrerae | 0.8 | 1.4 | b | ||||||||||||
| Fragilaria construens | 4.4 | 2.4 | 2.6 | 1.1 | b | ||||||||||
| Frustulia rhomboides | 7.1 | 6.7 | 1.1 | 1.8 | c | ||||||||||
| Gomphonema affini | 6.2 | a | |||||||||||||
| Gomphonema parvulum | 8.0 | a | |||||||||||||
| Gomphonema subclavatum | 6.3 | 1.5 | 2.0 | 2.2 | 1.2 | 1.3 | c | ||||||||
| Gomphonema tenellum | 4.9 | 8.8 | 9.3 | a | |||||||||||
| Gomphonema angusticephalum | 1.2 | b | |||||||||||||
| Gomphonema herculeanum | 0.7 | a | |||||||||||||
| Gomphonema micropus | 1.5 | 1.3 | b | ||||||||||||
| Gyrosigma terryanum | 6.4 | a | |||||||||||||
| Hannaea arcus | 7.9 | 7.4 | 8.2 | 9.7 | 9.0 | 9.4 | a | ||||||||
| Hantzschia virgata | 7.6 | 5.9 | 8.7 | 8.1 | 1.1 | c | |||||||||
| Meridion circulare | 1.6 | 1.4 | 2.1 | 2.0 | 1.8 | 2.5 | b | ||||||||
| Navicula cryptocephala | 7.1 | 8.1 | 9.4 | 8.1 | a | ||||||||||
| Navicula dicephala | 8.7 | 9.8 | 7.9 | a | |||||||||||
| Navicula directa | 8.5 | 10.8 | 5.9 | 0.8 | c | ||||||||||
| Navicula gotlandica | 9.1 | 8.9 | 8.8 | 2.2 | 1.6 | 1.2 | c | ||||||||
| Navicula lateropunctata | 8.1 | 8.6 | 9.9 | a | |||||||||||
| Navicula pseudoseinhardti | 8.8 | 5.0 | 5.6 | a | |||||||||||
| Navicula ryncocephala | 9.4 | 13.4 | 8.0 | a | |||||||||||
| Navicula viridula var. rotellata | 8.2 | 8.1 | 8.8 | a | |||||||||||
| Neidium bisulcatum | 3.4 | b | |||||||||||||
| Nitzschia apiculata | 7.2 | 9.2 | 8.8 | 2.6 | c | ||||||||||
| Nitzschia ignorata | 7.8 | 9.0 | 8.8 | 8.9 | a | ||||||||||
| Nitzschia rumpens | 5.6 | 8.6 | 1.1 | c | |||||||||||
| Nitzschia parva | 1.3 | b | |||||||||||||
| Nitzschia robusta | 1.0 | 1.6 | b | ||||||||||||
| Opephora martyi | 2.2 | 1.6 | 2.5 | b | |||||||||||
| Pinnularia intermedia | 7.9 | a | |||||||||||||
| Pinnularia major | 11.4 | 8.2 | 8.6 | 8.8 | 9.5 | 7.5 | 0.6 | 1.6 | 0.8 | c | |||||
| Pinnularia minor | 9.9 | 7.5 | 0.6 | 1.2 | 0.8 | 2.5 | 1.4 | 2.3 | c | ||||||
| Pleurosgima sp. | 0.6 | 1.4 | b | ||||||||||||
| Rhoicosphenia curvata | 8.7 | 2.5 | 3.1 | 2.7 | 3.0 | 4.0 | c | ||||||||
| Synedra acicularis | 7.8 | a | |||||||||||||
| Synedra acus | 8.2 | 3.9 | 4.4 | 3.5 | 2.2 | c | |||||||||
| Synedra rumpens | 8.2 | 9.9 | 8.7 | 10.2 | a | ||||||||||
| Tabellaria fenestrata | 8.0 | 2.0 | 2.6 | 3.0 | 4.6 | 4.4 | 5.0 | c | |||||||
| Tabellaria floculosa | 9.2 | 1.5 | 2.4 | 4.4 | 4.8 | 2.2 | c | ||||||||
| Coscinodiscophyceae | Actinocyclus curvatulus | 11.4 | 9.8 | a | |||||||||||
| Actinocyclus subtilis | 9.9 | 8.5 | a | ||||||||||||
| Aulacoseira granulata | 8.7 | 8.9 | 1.8 | 2.0 | 1.2 | 1.7 | 1.5 | 1.6 | c | ||||||
| Melosira hustedttii | 7.5 | 8.6 | 9.9 | 8.7 | 1.4 | 2.4 | c | ||||||||
| Melosira italica | 9.3 | 10.6 | 9.9 | 3.3 | 2.8 | c | |||||||||
| Melosira sol | 6.8 | 7.7 | 7.9 | 10.6 | 9.6 | 10.3 | 0.4 | 0.8 | c | ||||||
| Melosira varians | 8.2 | 8.3 | 10.9 | 8.5 | 2.2 | 2.5 | 1.8 | 2.6 | c | ||||||
| Rhizosolenia longiseta | 9.6 | 8.7 | 7.6 | 9.6 | 11.4 | 9.5 | a | ||||||||
| Mediophyceae | Cyclotella meneghiniana | 5.8 | 6.3 | 5.5 | 8.8 | 8.3 | 8.5 | a | |||||||
| Chlorophyta | Chlorophyceae | Chlamydocapsa bacillus | 6.0 | 1.4 | 3.4 | 1.4 | 1.8 | 1.2 | c | ||||||
| Chlamydocapsa planctonica | 1.6 | 0.8 | b | ||||||||||||
| Chlamydomonas dinobryonii | 7.8 | 2.7 | 2.1 | c | |||||||||||
| Chlamydomonas reinhardtii | 8.8 | 9.2 | a | ||||||||||||
| Coelastrum cambricum | 0.8 | 1.3 | b | ||||||||||||
| Coelastrum crenatum var. cubicum | 1.2 | 1.6 | b | ||||||||||||
| Cylindrocapsa geminella | 1.4 | b | |||||||||||||
| Eudorina elegans | 5.9 | 6.2 | 7.6 | 2.8 | 0.9 | 2.5 | 0.8 | 1.6 | 0.5 | c | |||||
| Gonium sociale | 8.1 | 8.7 | 7.9 | 2.2 | 1.3 | 0.8 | c | ||||||||
| Hemaetococcus pluvialis | 0.7 | 0.8 | 1.1 | b | |||||||||||
| Microspora tumidula | 8.5 | 7.9 | 9.1 | 9.7 | 8.9 | a | |||||||||
| Monoraphidium irregulare | 4.2 | 2.6 | 3.0 | 1.8 | b | ||||||||||
| Monoraphidium saxatile | 6.1 | 1.7 | 2.6 | 1.8 | 1.4 | c | |||||||||
| Pseudosphaerocystis lacustris | 8.4 | a | |||||||||||||
| Radiofilum conjunctivum | 6.7 | 5.9 | 8.2 | 8.7 | a | ||||||||||
| Scenedesmus armatus | 6.8 | 7.5 | a | ||||||||||||
| Scenedesmus brevispina | 7.7 | a | |||||||||||||
| Scenedesmus denticulatus | 7.4 | 0.6 | 0.7 | 0.6 | c | ||||||||||
| Scenedesmus quadriculata var. quadrispina | 7.9 | 5.7 | 7.3 | a | |||||||||||
| Sphaerocystis schroeteri | 10.2 | 2.8 | 0.8 | 3.5 | 1.1 | 0.8 | 0.4 | c | |||||||
| Stauridium tetras | 6.8 | 7.7 | 7.3 | 1.8 | 1.3 | 1.8 | 1.1 | 1.2 | c | ||||||
| Tetraspora lacustris | 7.5 | 7.5 | a | ||||||||||||
| Tetraedron sp. | 0.8 | b | |||||||||||||
| Volvox globator | 8.0 | 8.3 | 0.8 | 1.3 | 0.8 | 0.4 | 1.0 | 0.6 | c | ||||||
| Rhaphidiella fascicularis | 6.9 | a | |||||||||||||
| Trebouxiophyceae | Willea irregularis | 5.9 | a | ||||||||||||
| Botryococcus braunii | 2.4 | 1.7 | 1.2 | b | |||||||||||
| Chlorella fusca | 8.5 | 8.1 | 12.7 | 10.2 | 3.8 | 2.1 | 4.0 | 3.1 | 3.7 | 2.0 | c | ||||
| Chlorella saccharophila | 8.6 | a | |||||||||||||
| Chlorella vugaris | 8.2 | 8.1 | 8.1 | 9.4 | 14.2 | 10.6 | 5.7 | 2.4 | 6.7 | 1.8 | 3.2 | 2.5 | c | ||
| Nephrocytium limneticum | 5.8 | 5.8 | a | ||||||||||||
| Oocystis elliptica | 8.9 | 8.7 | 8.1 | 8.6 | a | ||||||||||
| Oocystis lacustris | 8.0 | 1.2 | 1.9 | 1.5 | 1.2 | 0.8 | 1.6 | c | |||||||
| Oocystis solitaria | 8.6 | a | |||||||||||||
| Ulvophyceae | Ulothrix tenuissima | 0.6 | b | ||||||||||||
| Ulothrix variabilis | 7.9 | 1.0 | c | ||||||||||||
| Ulothrix zonata | 1.5 | 1.2 | b | ||||||||||||
| Cryptophyta | Cryptophyceae | Cryptomonas curvata | 6.2 | 7.6 | 10.2 | a | |||||||||
| Cryptomonas ovata | 8.1 | 3.5 | 3.1 | 2.7 | 1.4 | 1.4 | 1.7 | c | |||||||
| Cyanobacteria | Cyanophyceae | Anabaena constricta | 6.6 | 6.9 | 8.0 | 7.4 | a | ||||||||
| Anabaena solitaria | 5.9 | 6.3 | 6.9 | 5.6 | 0.6 | c | |||||||||
| Aphanocapsa parietina | 7.4 | 8.1 | 1.5 | 1.9 | 0.8 | 0.6 | c | ||||||||
| Aphanothece microscopica | 6.7 | 8.9 | 15.6 | 13.0 | 2.6 | 2.0 | 1.4 | 1.6 | 0.9 | 1.6 | c | ||||
| Chroococcus minutus | 10.6 | 12.2 | 1.3 | 1.8 | 1.4 | c | |||||||||
| Dolicospermum sp. | 2.3 | 0.5 | 0.9 | b | |||||||||||
| Gomphosphaeria aponina | 0.9 | b | |||||||||||||
| Limnothrix redekei | 2.1 | 1.9 | 1.8 | 1.2 | 1.3 | b | |||||||||
| Lyngbya martensiana | 1.1 | b | |||||||||||||
| Merismopedia punctata | 0.5 | b | |||||||||||||
| Microcystis botrys | 0.8 | 0.8 | b | ||||||||||||
| Microcystis flosaquae | 0.4 | 0.4 | 1.4 | b | |||||||||||
| Nodularia spumigena | 5.6 | 4.8 | 5.5 | 6.4 | 0.9 | 0.7 | 0.8 | 1.4 | c | ||||||
| Nostoc kihlman | 5.2 | a | |||||||||||||
| Oscillatoria lacustris | 0.9 | b | |||||||||||||
| Raphidiopsis curvata | 0.4 | b | |||||||||||||
| Rivularia sp. | 0.8 | 0.9 | 1.8 | b | |||||||||||
| Spirulina platensis | 7.1 | 1.1 | 1.3 | c | |||||||||||
| Miozoa | Dinophyceae | Ceratium furcoide | 7.9 | 8.4 | 12.9 | 13.9 | 18.1 | 19.2 | 0.7 | c | |||||
| Ceratium tetraceros | 1.1 | 0.6 | 0.6 | 0.7 | 0.7 | b | |||||||||
| Gymnodinium fuscum | 5.4 | 15.4 | 16.6 | 11.8 | 19.5 | 14.8 | 1.1 | 0.6 | 0.3 | 0.7 | 0.6 | 0.8 | c | ||
| Peridium cingulum | 15.5 | 12.1 | 0.4 | 0.9 | c | ||||||||||
| Protoperidinium | 0.8 | 1.0 | b | ||||||||||||
| Euglenophyta | Euglenophyceae | Euglena gracilis | 5.3 | 7.8 | 10.2 | 8.4 | 0.6 | 0.8 | 0.9 | 0.4 | 0.7 | 0.5 | c | ||
| Phacus sp. | 1.3 | 0.4 | b | ||||||||||||
| Trachelomonas volvocina | 8.3 | 8.9 | 0.8 | 0.9 | 0.8 | 0.9 | c | ||||||||
| Ochrophyta | Chrysophyceae | Dinobryon divergens | 9.0 | 9.2 | 8.5 | 9.0 | 12.1 | 9.2 | 4.6 | 3.6 | 3.9 | 3.4 | 2.0 | 3.2 | c |
| Ochromonas elegans | 5.6 | 1.7 | 1.3 | 2.2 | 0.9 | c | |||||||||
| Mallomonas alpina | 2.0 | b | |||||||||||||
| Mallomonas longiseta | 4.4 | 8.8 | 8.3 | 1.7 | 1.2 | c | |||||||||
| Mallomonas areolata | 10.6 | 7.2 | 10.5 | 1.3 | 3.8 | 1.8 | 0.8 | c | |||||||
| Mallomonas caudata | 5.4 | 10.1 | 9.7 | 1.2 | 1.6 | 2.6 | 2.9 | c | |||||||
| Mallomonas elongiseta | 8.4 | 10.3 | 7.9 | 1.3 | 1.0 | 1.6 | 1.3 | 1.6 | 1.3 | c | |||||
| Synura uvella | 0.6 | b | |||||||||||||
| Chlorodendrophyceae | Tetraselmis limnetis | 0.4 | 0.4 | 0.5 | 0.8 | b | |||||||||
| Eustigmatophyceae | Goniochloris fallax | 1.4 | b | ||||||||||||
| Xantophyceae | Ophiocytium capitatum | 8.1 | a | ||||||||||||
| Charophyta | Zygnematophyceae | Closterium parvulum | 8.7 | a | |||||||||||
| Staurastrum orbiculare | 7.9 | 0.9 | 1.3 | 1.0 | c | ||||||||||
| Staurastrum paradoxum | 7.9 | a | |||||||||||||
| Staurastrum rotula | 0.8 | b | |||||||||||||
| Klebsormidiophyceae | Elakatothrix gelatinosa | 6.3 | a | ||||||||||||
| BIO-ENV | p = 0.08 | p = 0.51 | |||||||||||||
| ANOSIM between season | p = 0.02 | ||||||||||||||
Table 3.
Results of alpha diversity using the Shannon index (H′), richness index (Chao-1), Pielou’s evenness (J′) and Hill’s effective numbers (N1). Beta diversity with the Sorensen dissimilarity index (βSØR) and Simpson dissimilarity index (βSIM) at monitoring stations in Kusrüpuyewe Lake during summer and winter.
Table 3.
Results of alpha diversity using the Shannon index (H′), richness index (Chao-1), Pielou’s evenness (J′) and Hill’s effective numbers (N1). Beta diversity with the Sorensen dissimilarity index (βSØR) and Simpson dissimilarity index (βSIM) at monitoring stations in Kusrüpuyewe Lake during summer and winter.
| H′ | Chao-1 | J′ | N1 | βSØR (%) | βSIM (%) | ||
|---|---|---|---|---|---|---|---|
| Summer | |||||||
| LK1 | Top | 4.5 | 24 | 0.9 | 22 | 60.9 | 87.5 |
| Bottom | 5.5 | 45 | 0.9 | 44 | |||
| LK2 | Top | 5.1 | 36 | 0.9 | 35 | 53.7 | 61.1 |
| Bottom | 5.5 | 46 | 0.9 | 45 | |||
| LT3 | Top | 5.7 | 55 | 0.9 | 53 | 67.2 | 74.6 |
| Bottom | 6.1 | 67 | 0.9 | 66 | |||
| Winter | |||||||
| LK1 | Top | 5.5 | 53 | 0.9 | 45 | 62.7 | 65.3 |
| Bottom | 5.4 | 49 | 0.9 | 42 | |||
| LK2 | Top | 5.4 | 51 | 0.9 | 42 | 55.0 | 66.7 |
| Bottom | 5.4 | 51 | 0.9 | 42 | |||
| LK3 | Top | 5.5 | 54 | 0.9 | 46 | 71.3 | 70.7 |
| Bottom | 5.2 | 41 | 0.9 | 36 | |||
| Between Seasons | |||||||
| Summer | 5.8 | 103 | 0.9 | 98 | 62.9 | 74.4 | |
| Winter | 5.7 | 96 | 0.9 | 87 | 60.6 | 67.6 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Ríos-Henríquez, C.; Fuentes, N. Temporal Environmental Status of a Shallow Lake Using Alpha and Beta Diversity on Phytoplankton Communities. Water 2024, 16, 274. https://doi.org/10.3390/w16020274
AMA Style
Ríos-Henríquez C, Fuentes N. Temporal Environmental Status of a Shallow Lake Using Alpha and Beta Diversity on Phytoplankton Communities. Water. 2024; 16(2):274. https://doi.org/10.3390/w16020274
Chicago/Turabian StyleRíos-Henríquez, Catalina, and Norka Fuentes. 2024. "Temporal Environmental Status of a Shallow Lake Using Alpha and Beta Diversity on Phytoplankton Communities" Water 16, no. 2: 274. https://doi.org/10.3390/w16020274
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.