
Response of Phytoplankton Communities to Variation in Salinity in a Small Mediterranean Coastal Lagoon: Future Management and Foreseen Climate Change Consequences
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
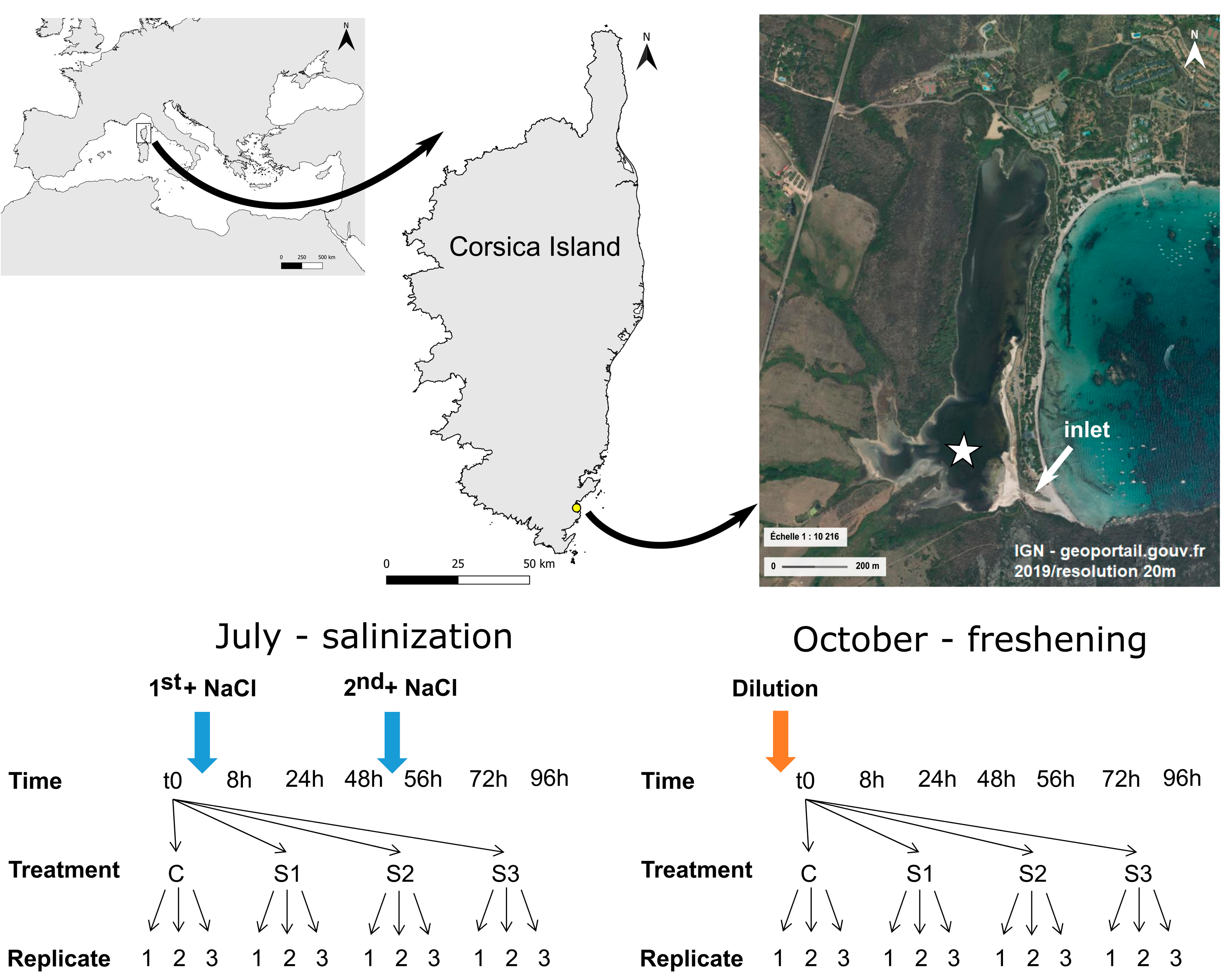
2.1. Study Site
2.2. Microcosm Experiments
2.2.1. Microcosm Setup for Salinization Experiment
2.2.2. Microcosm Setup for Freshening Experiment
2.3. Phytoplankton Community
2.4. Statistical Analysis
3. Results
3.1. Physico-Chemical Parameters
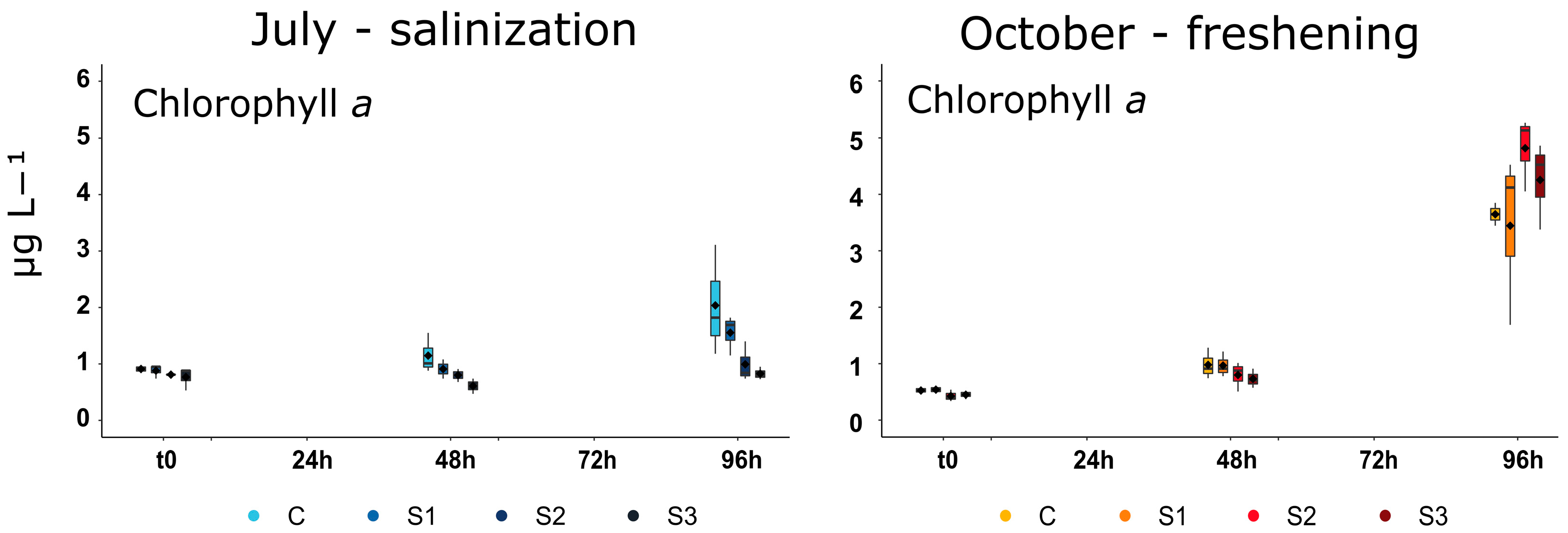
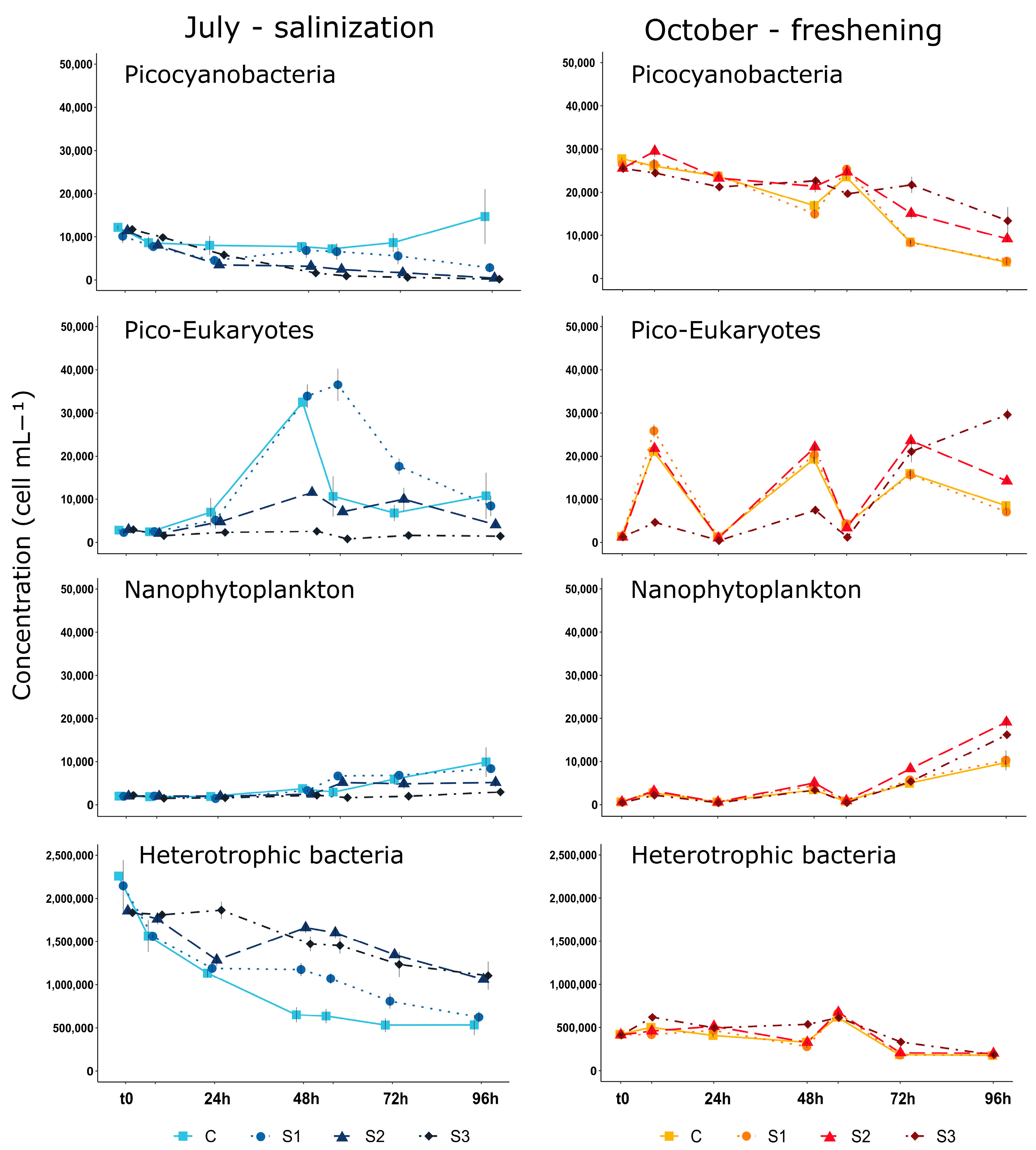
3.2. Phytoplankton Biomass and Small-Sized Phytoplankton Structure
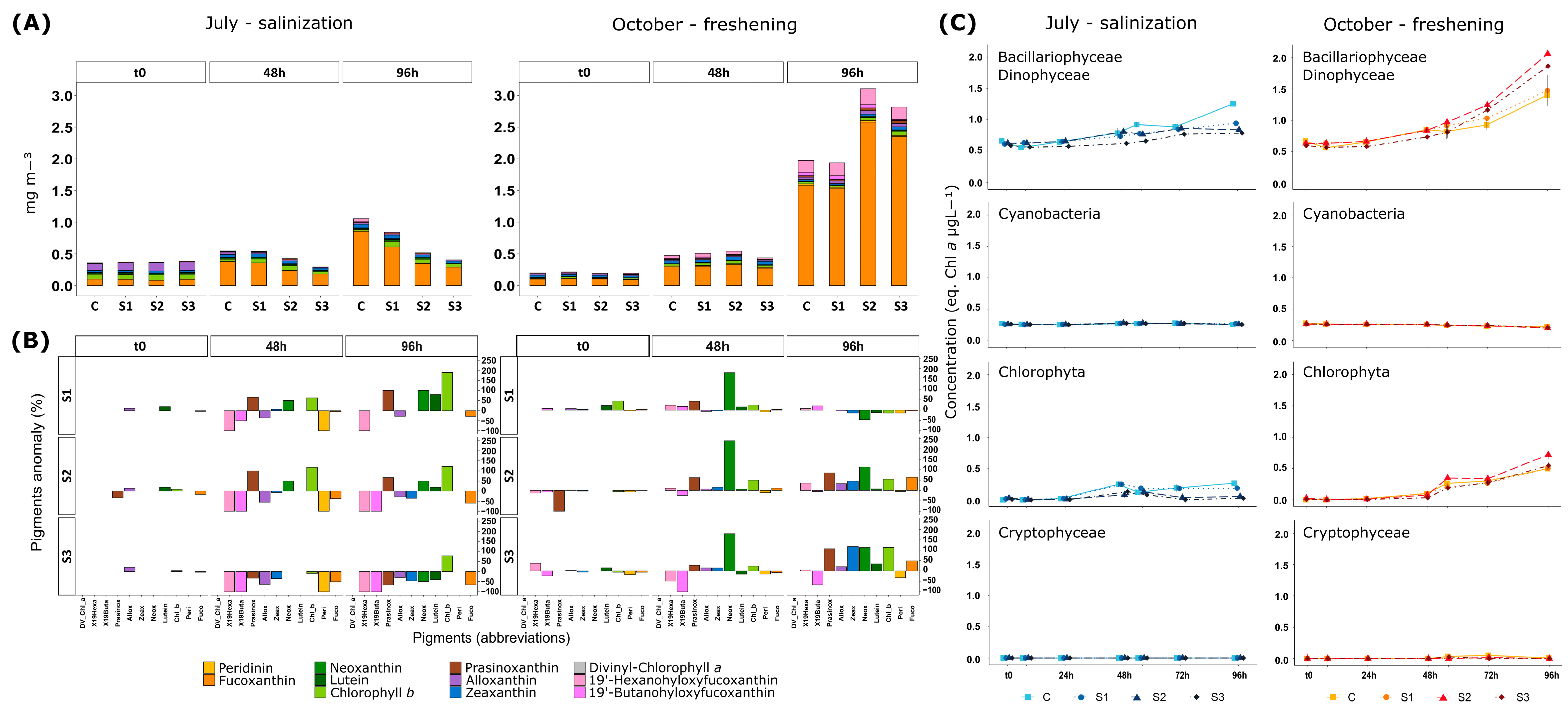
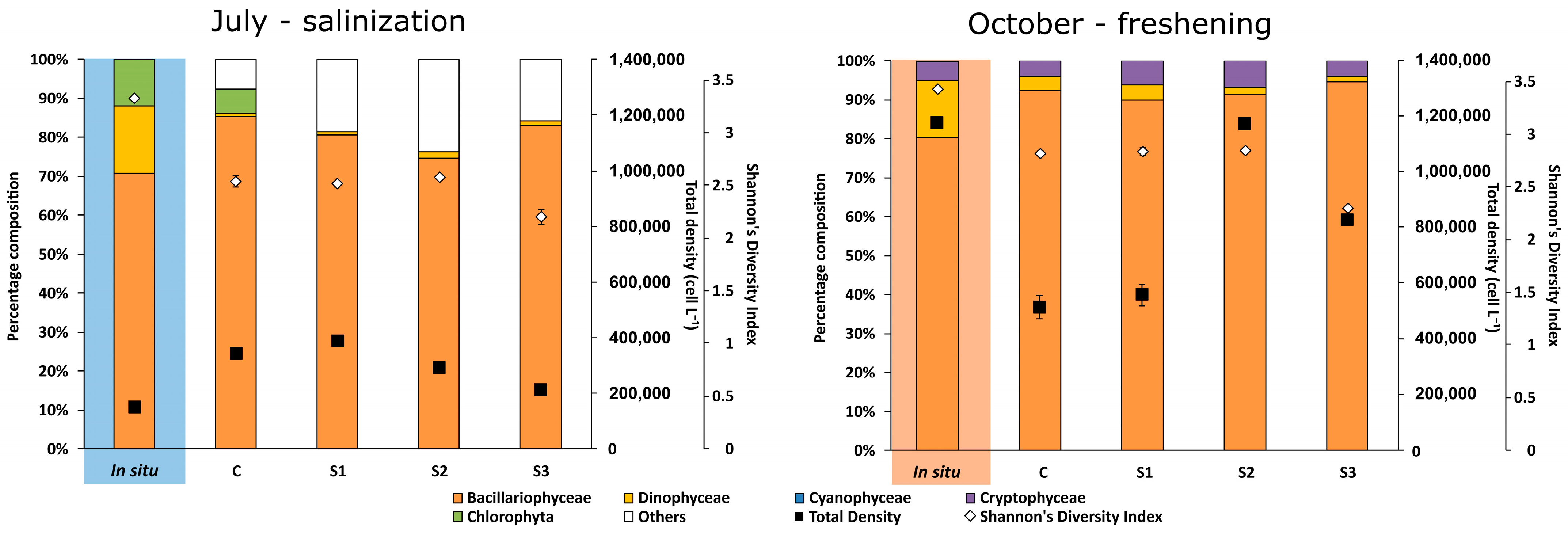
3.3. Phytoplankton Communities’ Composition by Pigment Analyses, Fluorometry, and Microscopy
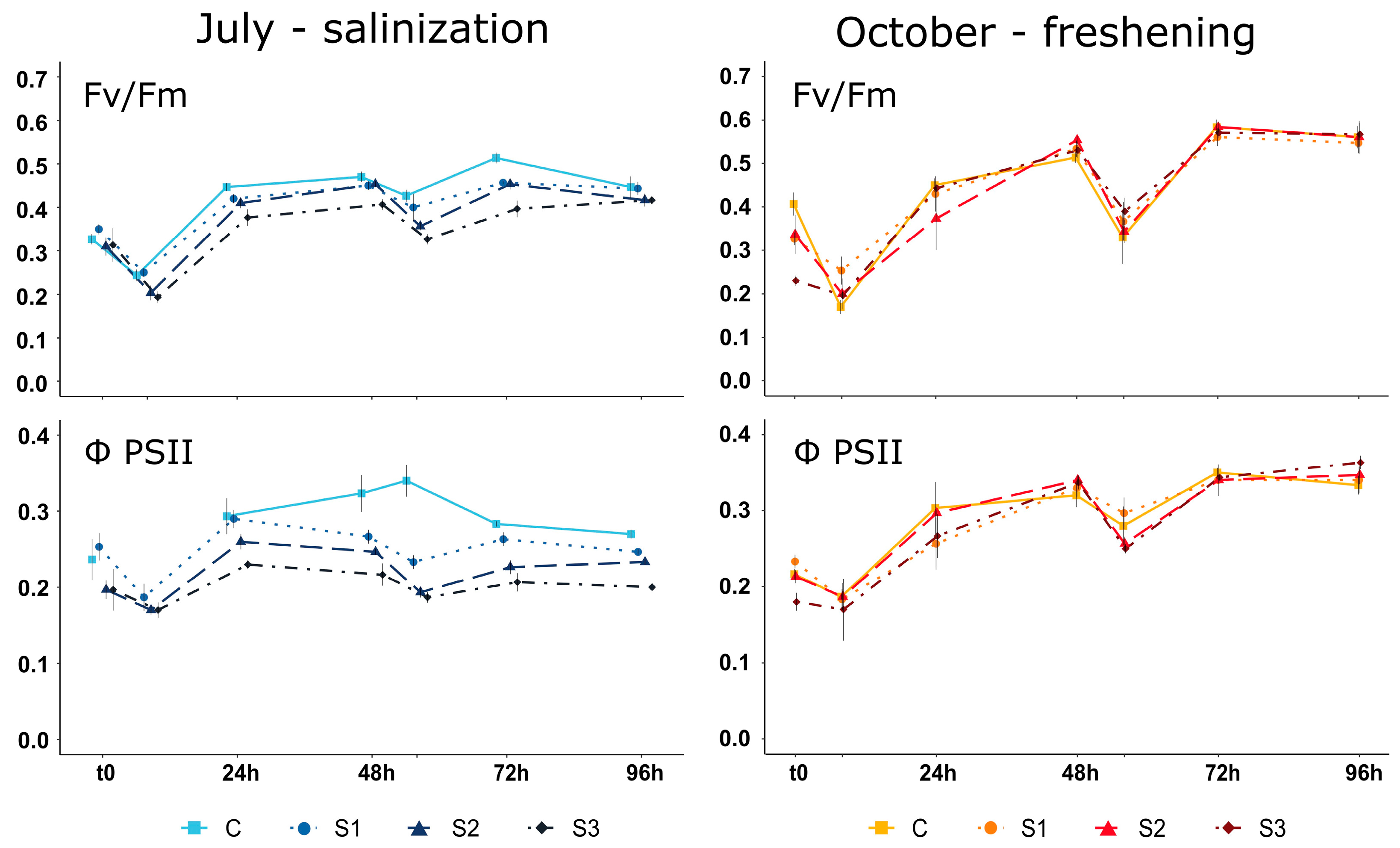
3.4. Phytoplankton Communities’ Metabolism and Status
4. Discussion
4.1. Global Effects of Short-Term Salinity Changes
4.2. Perspectives and Implications on Future Management of Small Mediterranean Lagoons
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Aurelle, D.; Thomas, S.; Albert, C.; Bally, M.; Bondeau, A.; Boudouresque, C.-F.; Cahill, A.E.; Carlotti, F.; Chenuil, A.; Cramer, W.; et al. Biodiversity, Climate Change, and Adaptation in the Mediterranean. Ecosphere 2022, 13, e3915. [Google Scholar] [CrossRef]
- MedECC. Climate and Environmental Change in the Mediterranean Basin—Current Situation and Risks for the Future. First Mediterranean Assessment Report; Zenodo: Marseille, France, 2020. [Google Scholar] [CrossRef]
- Cos, J.; Doblas-Reyes, F.; Jury, M.; Marcos, R.; Bretonnière, P.-A.; Samsó, M. The Mediterranean Climate Change Hotspot in the CMIP5 and CMIP6 Projections. Earth Syst. Dyn. 2022, 13, 321–340. [Google Scholar] [CrossRef]
- MWO. Mediterranean Wetlands Outlook 2: Solutions for Sustainable Mediterranean Wetlands; Mediterranean Wetlands Observatory: Arles, France, 2018. [Google Scholar]
- Chacón Abarca, S.; Chávez, V.; Silva, R.; Martínez, M.L.; Anfuso, G. Understanding the Dynamics of a Coastal Lagoon: Drivers, Exchanges, State of the Environment, Consequences and Responses. Geosciences 2021, 11, 301. [Google Scholar] [CrossRef]
- Loureiro, S.; Newton, A.; Icely, J. Boundary Conditions for the European Water Framework Directive in the Ria Formosa Lagoon, Portugal (Physico-Chemical and Phytoplankton Quality Elements). Estuar. Coast. Shelf Sci. 2006, 67, 382–398. [Google Scholar] [CrossRef]
- Cataudella, S.; Crosetti, D.; Ciccotti, E.; Massa, F. Sustainable Management in Mediterranean Coastal Lagoons: Interactions among Capture Fisheries, Aquaculture and the Environment. GFCM Stud. Rev. 2015, 95, 7–49. [Google Scholar]
- Renzi, M.; Provenza, F.; Pignattelli, S.; Cilenti, L.; Specchiulli, A.; Pepi, M. Mediterranean Coastal Lagoons: The Importance of Monitoring in Sediments the Biochemical Composition of Organic Matter. Int. J. Environ. Res. Public Health 2019, 16, 3466. [Google Scholar] [CrossRef]
- Ligorini, V.; Crayol, E.; Huneau, F.; Garel, E.; Malet, N.; Garrido, M.; Simon, L.; Cecchi, P.; Pasqualini, V. Small Mediterranean Coastal Lagoons under Threat: Hydro-Ecological Disturbances and Local Anthropogenic Pressures (Size Matters). Estuaries Coasts 2023. [Google Scholar] [CrossRef] [PubMed]
- European Union (EU). Directive 2000/60/EC of the European Parliament and of the Council of 23 October 2000 Establishing a Framework for Community Action in the Field of Water Policy. Official Journal L 327; European Union (EU): Maastricht, The Netherlands, 2000; Volume 327. [Google Scholar]
- SDAGE. Projet de Schéma Directeur d’Amenagement et Gestion Des Eaux 2022–2027—Bassin de Corse—3 Decémbre 2021; Comité de Bassin de Corse: Ajaccio, France, 2021; Available online: https://www.corse.eaufrance.fr/ (accessed on 17 March 2022).
- ISPRA. GdL “Reti di Monitoraggio e Reporting Direttiva 2000/60/CE”: Progettazione di Reti e Programmi Di Monitoraggio Delle Acque ai Sensi del D.Lgs. 152/2006 e Relativi Decreti Attuativi—ISPRA—Manuali e Linee Guida 116/2014; ISPRA: Roma, Italy, 2014. [Google Scholar]
- Doughty, C.L.; Cavanaugh, K.C.; Ambrose, R.F.; Stein, E.D. Evaluating Regional Resiliency of Coastal Wetlands to Sea Level Rise through Hypsometry-Based Modeling. Glob. Chang. Biol. 2019, 25, 78–92. [Google Scholar] [CrossRef] [PubMed]
- Ferrarin, C.; Bajo, M.; Bellafiore, D.; Cucco, A.; De Pascalis, F.; Ghezzo, M.; Umgiesser, G. Toward Homogenization of Mediterranean Lagoons and Their Loss of Hydrodiversity. Geophys. Res. Lett. 2014, 41, 5935–5941. [Google Scholar] [CrossRef]
- Robert, R.C.; Mazzilli, S. Defining the Coast and Sentinel Ecosystems for Coastal Observations of Global Change. Hydrobioligia 2007, 577, 55–70. [Google Scholar] [CrossRef]
- Cloern, J.E. The Relative Importance of Light and Nutrient Limitation of Phytoplankton Growth: A Simple Index of Coastal Ecosystem Sensitivity to Nutrient Enrichment. Aquat. Ecol. 1999, 33, 3–16. [Google Scholar] [CrossRef]
- Armi, Z.; Trabelsi, E.; Turki, S.; Béjaoui, B.; Maïz, N.B. Seasonal Phytoplankton Responses to Environmental Factors in a Shallow Mediterranean Lagoon. J. Mar. Sci. Technol. 2010, 15, 417–426. [Google Scholar] [CrossRef]
- Goberville, E.; Beaugrand, G.; Sautour, B.; Tréguer, P.; Somlit, T. Climate-Driven Changes in Coastal Marine Systems of Western Europe. Mar. Ecol. Prog. Ser. 2010, 408, 129–147. [Google Scholar] [CrossRef]
- Birk, S.; Bonne, W.; Borja, A.; Brucet, S.; Courrat, A.; Poikane, S.; Solimini, A.; van de Bund, W.; Zampoukas, N.; Hering, D. Three Hundred Ways to Assess Europe’s Surface Waters: An Almost Complete Overview of Biological Methods to Implement the Water Framework Directive. Ecol. Indic. 2012, 18, 31–41. [Google Scholar] [CrossRef]
- Dube, A.; Jayaraman, G.; Rani, R. Modelling the Effects of Variable Salinity on the Temporal Distribution of Plankton in Shallow Coastal Lagoons. J. Hydro-Environ. Res. 2010, 4, 199–209. [Google Scholar] [CrossRef]
- Pulina, S.; Padedda, B.M.; Satta, C.T.; Sechi, N.; Lugliè, A. Long-Term Phytoplankton Dynamics in a Mediterranean Eutrophic Lagoon (Cabras Lagoon, Italy). Plant Biosyst.—Int. J. Deal. All Asp. Plant Biol. 2012, 146, 259–272. [Google Scholar] [CrossRef]
- Stefanidou, N.; Genitsaris, S.; Lopez-Bautista, J.; Sommer, U.; Moustaka-Gouni, M. Effects of Heat Shock and Salinity Changes on Coastal Mediterranean Phytoplankton in a Mesocosm Experiment. Mar. Biol. 2018, 165, 154. [Google Scholar] [CrossRef]
- Greenwald, G.M.; Hurlbert, S.H. Microcosm Analysis of Salinity Effects on Coastal Lagoon Plankton Assemblages. Hydrobiologia 1993, 267, 307–335. [Google Scholar] [CrossRef]
- Pilkaitytë, R.; Schoor, A.; Schubert, H. Response of Phytoplankton Communities to Salinity Changes—A Mesocosm Approach. Hydrobiologia 2004, 513, 27–38. [Google Scholar] [CrossRef]
- Pulina, S.; Suikkanen, S.; Padedda, B.M.; Brutemark, A.; Grubisic, L.M.; Satta, C.T.; Caddeo, T.; Farina, P.; Lugliè, A. Responses of a Mediterranean Coastal Lagoon Plankton Community to Experimental Warming. Mar. Biol. 2020, 167, 22. [Google Scholar] [CrossRef]
- Barnes, B.D.; Wurtsbaugh, W.A. The Effects of Salinity on Plankton and Benthic Communities in the Great Salt Lake, Utah, USA: A Microcosm Experiment. Can. J. Fish. Aquat. Sci. 2015, 72, 807–817. [Google Scholar] [CrossRef]
- Schallenberg, M.; Larned, S.T.; Hayward, S.; Arbuckle, C. Contrasting Effects of Managed Opening Regimes on Water Quality in Two Intermittently Closed and Open Coastal Lakes. Estuar. Coast. Shelf Sci. 2010, 86, 587–597. [Google Scholar] [CrossRef]
- D’ors, A.; Bartolomé, M.C.; Sánchez-Fortún, S. Repercussions of Salinity Changes and Osmotic Stress in Marine Phytoplankton Species. Estuar. Coast. Shelf Sci. 2016, 175, 169–175. [Google Scholar] [CrossRef]
- Flöder, S.; Jaschinski, S.; Wells, G.; Burns, C.W. Dominance and Compensatory Growth in Phytoplankton Communities under Salinity Stress. J. Exp. Mar. Biol. Ecol. 2010, 395, 223–231. [Google Scholar] [CrossRef]
- Hernando, M.; Schloss, I.R.; Malanga, G.; Almandoz, G.O.; Ferreyra, G.A.; Aguiar, M.B.; Puntarulo, S. Effects of Salinity Changes on Coastal Antarctic Phytoplankton Physiology and Assemblage Composition. J. Exp. Mar. Biol. Ecol. 2015, 466, 110–119. [Google Scholar] [CrossRef]
- Nche-Fambo, F.A.; Scharler, U.M.; Tirok, K. Resilience of Estuarine Phytoplankton and Their Temporal Variability along Salinity Gradients during Drought and Hypersalinity. Estuar. Coast. Shelf Sci. 2015, 158, 40–52. [Google Scholar] [CrossRef]
- Barroso, H.d.S.; Tavares, T.C.L.; Soares, M.d.O.; Garcia, T.M.; Rozendo, B.; Vieira, A.S.C.; Viana, P.B.; Pontes, T.M.; Ferreira, T.J.T.; Pereira Filho, J.; et al. Intra-Annual Variability of Phytoplankton Biomass and Nutrients in a Tropical Estuary during a Severe Drought. Estuar. Coast. Shelf Sci. 2018, 213, 283–293. [Google Scholar] [CrossRef]
- Hernando, M.; Varela, D.E.; Malanga, G.; Almandoz, G.O.; Schloss, I.R. Effects of Climate-Induced Changes in Temperature and Salinity on Phytoplankton Physiology and Stress Responses in Coastal Antarctica. J. Exp. Mar. Biol. Ecol. 2020, 530–531, 151400. [Google Scholar] [CrossRef]
- Pecqueur, D.; Vidussi, F.; Fouilland, E.; Le Floc’h, E.; Mas, S.; Roques, C.; Salles, C.; Tournoud, M.-G.; Mostajir, B. Dynamics of Microbial Planktonic Food Web Components during a River Flash Flood in a Mediterranean Coastal Lagoon. Hydrobiologia 2011, 673, 13–27. [Google Scholar] [CrossRef]
- Chapman, P.M. Management of Coastal Lagoons under Climate Change. Estuar. Coast. Shelf Sci. 2012, 110, 32–35. [Google Scholar] [CrossRef]
- Pérez-Ruzafa, A.; Pérez-Ruzafa, I.M.; Newton, A.; Marcos, C. Coastal Lagoons: Environmental Variability, Ecosystem Complexity, and Goods and Services Uniformity. In Coasts and Estuaries; Elsevier: Amsterdam, The Netherlands, 2019; pp. 253–276. ISBN 978-0-12-814003-1. [Google Scholar]
- Taylor, N.G.; Grillas, P.; Al Hreisha, H.; Balkız, Ö.; Borie, M.; Boutron, O.; Catita, A.; Champagnon, J.; Cherif, S.; Çiçek, K.; et al. The Future for Mediterranean Wetlands: 50 Key Issues and 50 Important Conservation Research Questions. Reg. Environ. Chang. 2021, 21, 33. [Google Scholar] [CrossRef]
- Pergent-Martini, C.; Fernandez, C.; Agostini, S.; Pergent, G. Les Étangs de Corse, Bibliographie—Synthèse 1997. Contrat Equipe Ecosystèmes Littoraux. Univ. Corse/Off. de l’Environnement de la Corse Ifremer 1997, 269. [Google Scholar]
- WMO-No. 1203 © World Meteorological Organization; Chairperson, Publications Board World Meteorological Organization (WMO): Geneva, Switzerland, 2017; ISBN 978-92-63-11203-3. [Google Scholar]
- Lafabrie, C.; Garrido, M.; Leboulanger, C.; Cecchi, P.; Grégori, G.; Pasqualini, V.; Pringault, O. Impact of Contaminated-Sediment Resuspension on Phytoplankton in the Biguglia Lagoon (Corsica, Mediterranean Sea). Estuar. Coast. Shelf Sci. 2013, 130, 70–80. [Google Scholar] [CrossRef]
- de la Broise, D.; Palenik, B. Immersed in Situ Microcosms: A Tool for the Assessment of Pollution Impact on Phytoplankton. J. Exp. Mar. Biol. Ecol. 2007, 341, 274–281. [Google Scholar] [CrossRef]
- Holmes, R.M.; Aminot, A.; Kérouel, R.; Hooker, B.A.; Peterson, B.J. A Simple and Precise Method for Measuring Ammonium in Marine and Freshwater Ecosystems. Can. J. Fish. Aquat. Sci. 1999, 56, 1801–1808. [Google Scholar] [CrossRef]
- Aminot, A.; Kérouel, R. Dosage Automatique des Nutriments Dans les Eaux Marines: Méthodes en Flux Continu; Méthodes d’analyse En Milieu Marin; IFREMER: Brest, France, 2007; 188p. [Google Scholar]
- Leruste, A.; Garrido, M.; Malet, N.; Bec, B.; De Wit, R.; Cecchi, P.; Pasqualini, V. Impact of Nutrient Availability on the Trophic Strategies of the Planktonic Protist Communities in a Disturbed Mediterranean Coastal Lagoon. Hydrobiologia 2021, 848, 1101–1119. [Google Scholar] [CrossRef]
- Collos, Y.; Vaquer, A.; Bibent, B.; Souchu, P.; Slawyk, G.; Garcia, N. Response of Coastal Phytoplankton to Ammonium and Nitrate Pulses: Seasonal Variations of Nitrogen Uptake and Regeneration. Aquat. Ecol. 2003, 37, 227–236. [Google Scholar] [CrossRef]
- Cecchi, P.; Garrido, M.; Collos, Y.; Pasqualini, V. Water Flux Management and Phytoplankton Communities in a Mediterranean Coastal Lagoon. Part II: Mixotrophy of Dinoflagellates as an Adaptive Strategy? Mar. Pollut. Bull. 2016, 108, 120–133. [Google Scholar] [CrossRef]
- Neveux, J.; Lantoine, F. Spectrofluorometric Assay of Chlorophylls and Phaeopigments Using the Least Squares Approximation Technique. Deep Sea Res. Part I Oceanogr. Res. Pap. 1993, 40, 1747–1765. [Google Scholar] [CrossRef]
- Marie, D.; Rigaut-Jalabert, F.; Vaulot, D. An Improved Protocol for Flow Cytometry Analysis of Phytoplankton Cultures and Natural Samples. Cytometry 2014, 85, 962–968. [Google Scholar] [CrossRef]
- Fouilland, E.; Trottet, A.; Alves-de-Souza, C.; Bonnet, D.; Bouvier, T.; Bouvy, M.; Boyer, S.; Guillou, L.; Hatey, E.; Jing, H.; et al. Significant Change in Marine Plankton Structure and Carbon Production after the Addition of River Water in a Mesocosm Experiment. Microb. Ecol. 2017, 74, 289–301. [Google Scholar] [CrossRef]
- Ras, J.; Claustre, H.; Uitz, J. Spatial Variability of Phytoplankton Pigment Distributions in the Subtropical South Pacific Ocean: Comparison between in Situ and Predicted Data. Biogeosciences 2008, 5, 353–369. [Google Scholar] [CrossRef]
- Garrido, M.; Cecchi, P.; Malet, N.; Bec, B.; Torre, F.; Pasqualini, V. Evaluation of FluoroProbe® Performance for the Phytoplankton-Based Assessment of the Ecological Status of Mediterranean Coastal Lagoons. Environ. Monit. Assess. 2019, 91, 204. [Google Scholar] [CrossRef]
- Utermöhl, H. Zur Vervollkommnung Der Quantitativen Phytoplankton-Methodik. Int. Ver. Theor. Angew. Limnol. 1958, 9, 1–38. [Google Scholar] [CrossRef]
- NF EN 15204; Norme Guide Pour le Dénombrement du Phytoplancton Par Microscopie Inversée; Méthode Utermöhl. AFNOR: Paris, France, 2006.
- Lund, J.W.G.; Kipling, C.; Le Cren, E.D. The Inverted Microscope Method of Estimating Algal Numbers and the Statistical Basis of Estimations by Counting. Hydrobiologia 1958, 11, 143–170. [Google Scholar] [CrossRef]
- Uehlinger, V. Étude Statistique Des Méthodes de Dénombrement Planctonique. Archives Des Sciences, Soc. de Phys. et d’Hist. Nat. de Genève. Int. Rev. Der Gesamten Hydrobiol. Und Hydrogr. 1964, 17, 121–233. [Google Scholar] [CrossRef]
- Chrétiennot-Dinet, M.J. Atlas du Phytoplancton Marin, Volume 3: Chlorarachniophycées, Chlorophycées, Chrysophycées, Cryptophycées, Euglénophycées, Eustigmatophycées, Prasinophycées, Prymnésiophycées, Rhodophycées, Tribophycées; Ed. CNRS: Paris, France, 1990; 260p. [Google Scholar]
- Throndsen, J. The Planktonic Marine Flagellates. In Identifying Marine Phytoplankton; Thomas, C.R., Ed.; Academic Press: San Diego, CA, USA, 1997. [Google Scholar]
- Horner, R.A. A Taxonomic Guide to Some Common Marine Phytoplancton; Biopres Ltd.: Bristol, UK, 2002; 195p. [Google Scholar]
- Avancini, M.; Cicero, A.M.; Innamorati, M.; Magaletti, E.; Sertorio Zunini, T. Guida al Riconoscimento Del Plancton Nei Mari Italiani, Volume I—Fitoplancton; Ministero Dell’Ambiente e Della Tutela Del Territorio e Del Mare—ICRAM: Roma, Italy, 2006; 503p. [Google Scholar]
- Hoppenrath, M.; Elbrachter, M.; Drebes, G. Marine Phytoplankton: Selected Microphytoplankton Species from the North Sea around Helgoland and Sylt; E. Schweizerbart’sche Verlagsbuchhandlung (Nagele U. Obermiller): Stuttgart, Germany, 2009. [Google Scholar]
- Steidinger, K.A.; Meave del Castillo, M.E. Guide to the Identification of Harmful Microalgae in the Gulf of Mexico, Volume I: Taxonomy; Florida Fish and Wildlife Research Institute: St. Petersburg, FL, USA, 2018; 384p, Available online: http://myfwc.com/research/redtide/research/scientific-products/ (accessed on 12 July 2021).
- Claustre, H. The Trophic Status of Various Oceanic Provinces as Revealed by Phytoplankton Pigment Signatures. Limnol. Oceanogr. 1994, 39, 1206–1210. [Google Scholar] [CrossRef]
- Vidussi, F.; Claustre, H.; Manca, B.B.; Luchetta, A.; Marty, J.-C. Phytoplankton Pigment Distribution in Relation to Upper Thermocline Circulation in the Eastern Mediterranean Sea during Winter. J. Geophys. Res. Ocean. 2001, 106, 19939–19956. [Google Scholar] [CrossRef]
- Leruste, A.; Malet, N.; Munaron, D.; Derolez, V.; Hatey, E.; Collos, Y.; De Wit, R.; Bec, B. First Steps of Ecological Restoration in Mediterranean Lagoons: Shifts in Phytoplankton Communities. Estuar. Coast. Shelf Sci. 2016, 180, 190–203. [Google Scholar] [CrossRef]
- Garrido, M.; Cecchi, P.; Vaquer, A.; Pasqualini, V. Effects of Sample Conservation on Assessments of the Photosynthetic Efficiency of Phytoplankton Using PAM Fluorometry. Deep Sea Res. Part I Oceanogr. Res. Pap. 2013, 71, 38–48. [Google Scholar] [CrossRef]
- Genty, B.; Briantais, J.-M.; Baker, N.R. The Relationship between the Quantum Yield of Photosynthetic Electron Transport and Quenching of Chlorophyll Fluorescence. Biochim. Biophys. Acta BBA—Gen. Subj. 1989, 990, 87–92. [Google Scholar] [CrossRef]
- Maxwell, K.; Johnson, G.N. Chlorophyll Fluorescence—A Practical Guide. J. Exp. Bot. 2000, 51, 659–668. [Google Scholar] [CrossRef] [PubMed]
- RStudio Team. RStudio: Integrated Development for R; RStudio Inc.: Boston, MA, USA, 2016; Available online: https://www.rstudio.com/ (accessed on 5 August 2022).
- Underwood, A.J. Experiments in Ecology: Their Logical Design and Interpretation Using Analysis of Variance; Cambridge University Press: Cambridge, UK, 1996; ISBN 9780521553292/9780521556965/9780511806407. [Google Scholar]
- Lamont, T.; Tutt, G.C.O.; Barlow, R.G. Phytoplankton Biomass and Photophysiology at the Sub-Antarctic Prince Edward Islands Ecosystem in the Southern Ocean. J. Mar. Syst. 2022, 226, 103669. [Google Scholar] [CrossRef]
- Delaune, K.D.; Nesich, D.; Goos, J.M.; Relyea, R.A. Impacts of Salinization on Aquatic Communities: Abrupt vs. Gradual Exposures. Environ. Pollut. 2021, 285, 117636. [Google Scholar] [CrossRef]
- Redden, A.M.; Rukminasari, N. Effects of Increases in Salinity on Phytoplankton in the Broadwater of the Myall Lakes, NSW, Australia. Hydrobiologia 2008, 608, 87–97. [Google Scholar] [CrossRef]
- Ersoy, Z.; Abril, M.; Cañedo-Argüelles, M.; Espinosa, C.; Vendrell-Puigmitja, L.; Proia, L. Experimental Assessment of Salinization Effects on Freshwater Zooplankton Communities and Their Trophic Interactions under Eutrophic Conditions. Environ. Pollut. 2022, 313, 120127. [Google Scholar] [CrossRef]
- Gregor, J.; Maršálek, B. Freshwater Phytoplankton Quantification by Chlorophyll a: A Comparative Study of in Vitro, in Vivo and in Situ Methods. Water Res. 2004, 38, 517–522. [Google Scholar] [CrossRef]
- Catherine, A.; Escoffier, N.; Belhocine, A.; Nasri, A.B.; Hamlaoui, S.; Yéprémian, C.; Bernard, C.; Troussellier, M. On the Use of the FluoroProbe®, a Phytoplankton Quantification Method Based on Fluorescence Excitation Spectra for Large-Scale Surveys of Lakes and Reservoirs. Water Res. 2012, 46, 1771–1784. [Google Scholar] [CrossRef]
- Garrido, M.; Cecchi, P.; Collos, Y.; Agostini, S.; Pasqualini, V. Water Flux Management and Phytoplankton Communities in a Mediterranean Coastal Lagoon. Part I: How to Promote Dinoflagellate Dominance? Mar. Pollut. Bull. 2016, 104, 139–152. [Google Scholar] [CrossRef]
- Dhib, A.; Denis, M.; Ziadi, B.; Barani, A.; Turki, S.; Aleya, L. Assessing Ultraphytoplankton and Heterotrophic Prokaryote Composition by Flow Cytometry in a Mediterranean Lagoon. Environ. Sci. Pollut. Res. 2017, 24, 13710–13721. [Google Scholar] [CrossRef]
- Xia, X.; Guo, W.; Tan, S.; Liu, H. Synechococcus Assemblages across the Salinity Gradient in a Salt Wedge Estuary. Front. Microbiol. 2017, 8, 1254. [Google Scholar] [CrossRef]
- Rajaneesh, K.M.; Mitbavkar, S. Factors Controlling the Temporal and Spatial Variations in Synechococcus Abundance in a Monsoonal Estuary. Mar. Environ. Res. 2013, 92, 133–143. [Google Scholar] [CrossRef]
- Xiao, W.; Liu, X.; Irwin, A.J.; Laws, E.A.; Wang, L.; Chen, B.; Zeng, Y.; Huang, B. Warming and Eutrophication Combine to Restructure Diatoms and Dinoflagellates. Water Res. 2018, 128, 206–216. [Google Scholar] [CrossRef] [PubMed]
- Caroppo, C. Ecology and Biodiversity of Picoplanktonic Cyanobacteria in Coastal and Brackish Environments. Biodivers. Conserv. 2015, 24, 949–971. [Google Scholar] [CrossRef]
- Le Rouzic, B. Changes in Photosynthetic Yield (Fv/Fm) Responses of Salt-Marsh Microalgal Communities along an Osmotic Gradient (Mont-Saint-Michel Bay, France). Estuar. Coast. Shelf Sci. 2012, 115, 326–333. [Google Scholar] [CrossRef]
- Nikitashina, V.; Stettin, D.; Pohnert, G. Metabolic Adaptation of Diatoms to Hypersalinity. Phytochemistry 2022, 201, 113267. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Callier, M.D.; Blancheton, J.-P.; Galès, A.; Nahon, S.; Triplet, S.; Geoffroy, T.; Menniti, C.; Fouilland, E.; Roque d’orbcastel, E. Bioremediation of Fishpond Effluent and Production of Microalgae for an Oyster Farm in an Innovative Recirculating Integrated Multi-Trophic Aquaculture System. Aquaculture 2019, 504, 314–325. [Google Scholar] [CrossRef]
- Bonilla, S.; Conde, D.; Aubriot, L.; Pérez, M.d.C. Influence of Hydrology on Phytoplankton Species Composition and Life Strategies in a Subtropical Coastal Lagoon Periodically Connected with the Atlantic Ocean. Estuaries 2005, 28, 884–895. [Google Scholar] [CrossRef]
- Dhib, A.; Frossard, V.; Turki, S.; Aleya, L. Dynamics of Harmful Dinoflagellates Driven by Temperature and Salinity in a Northeastern Mediterranean Lagoon. Environ. Monit. Assess. 2013, 185, 3369–3382. [Google Scholar] [CrossRef]
- Bravo, I.; Fraga, S.; Isabel Figueroa, R.; Pazos, Y.; Massanet, A.; Ramilo, I. Bloom Dynamics and Life Cycle Strategies of Two Toxic Dinoflagellates in a Coastal Upwelling System (NW Iberian Peninsula). Deep Sea Res. Part II Top. Stud. Oceanogr. 2010, 57, 222–234. [Google Scholar] [CrossRef]
- Yang, C.; Li, Y.; Zhou, Y.; Zheng, W.; Tian, Y.; Zheng, T. Bacterial Community Dynamics during a Bloom Caused by Akashiwo Sanguinea in the Xiamen Sea Area, China. Harmful Algae 2012, 20, 132–141. [Google Scholar] [CrossRef]
- Islabão, C.A.; Odebrecht, C. Influence of Salinity on the Growth of Akashiwo sanguinea and Prorocentrum micans (Dinophyta) under Acclimated Conditions and Abrupt Changes. Mar. Biol. Res. 2015, 11, 965–973. [Google Scholar] [CrossRef]
- Clavero, E.; Hernández-Mariné, M.; Grimalt, J.O.; Garcia-Pichel, F. Salinity Tolerance of Diatoms from Thalassic Hypersaline Environments. J. Phycol. 2000, 36, 1021–1034. [Google Scholar] [CrossRef]
- Varona-Cordero, F.; Gutiérrez-Mendieta, F.J.; Meave del Castillo, M.E. Phytoplankton Assemblages in Two Compartmentalized Coastal Tropical Lagoons (Carretas-Pereyra and Chantuto-Panzacola, Mexico). J. Plankton Res. 2010, 32, 1283–1299. [Google Scholar] [CrossRef]
- Astorg, L.; Gagnon, J.; Lazar, C.S.; Derry, A.M. Effects of Freshwater Salinization on a Salt-naïve Planktonic Eukaryote Community. Limnol. Oceanogr. Lett. 2023, 8, 38–47. [Google Scholar] [CrossRef]
- Ganguly, D.; Robin, R.S.; Vardhan, K.V.; Muduli, P.R.; Abhilash, K.R.; Patra, S.; Subramanian, B.R. Variable Response of Two Tropical Phytoplankton Species at Different Salinity and Nutrient Condition. J. Exp. Mar. Biol. Ecol. 2013, 440, 244–249. [Google Scholar] [CrossRef]
- Pereira Coutinho, M.T.; Brito, A.C.; Pereira, P.; Gonçalves, A.S.; Moita, M.T. A Phytoplankton Tool for Water Quality Assessment in Semi-Enclosed Coastal Lagoons: Open vs Closed Regimes. Estuar. Coast. Shelf Sci. 2012, 110, 134–146. [Google Scholar] [CrossRef]
- Collos, Y.; Bec, B.; Jauzein, C.; Abadie, E.; Laugier, T.; Lautier, J.; Pastoureaud, A.; Souchu, P.; Vaquer, A. Oligotrophication and Emergence of Picocyanobacteria and a Toxic Dinoflagellate in Thau Lagoon, Southern France. J. Sea Res. 2009, 61, 68–75. [Google Scholar] [CrossRef]
- Trombetta, T.; Vidussi, F.; Mas, S.; Parin, D.; Simier, M.; Mostajir, B. Water Temperature Drives Phytoplankton Blooms in Coastal Waters. PLoS ONE 2019, 14, e0214933. [Google Scholar] [CrossRef]
- Fischer, A.D.; Hayashi, K.; McGaraghan, A.; Kudela, R.M. Return of the “Age of Dinoflagellates” in Monterey Bay: Drivers of Dinoflagellate Dominance Examined Using Automated Imaging Flow Cytometry and Long-Term Time Series Analysis. Limnol. Oceanogr. 2020, 65, 2125–2141. [Google Scholar] [CrossRef]
- Carrada, G.C.; Casotti, R.; Modigh, M.; Saggiomo, V. Presence of Gymnodinium catenatum (Dinophyceae) in a Coastal Mediterranean Lagoon. J. Plankton Res. 1991, 13, 229–238. [Google Scholar] [CrossRef]
- Fraga, S.; Bravo, I.; Delgado, M.; Franco, J.M.; Zapata, M. Gyrodinium impudicum Sp. Nov. (Dinophyceae), a Non Toxic, Chain-Forming, Red Tide Dinoflagellate. Phycologia 1995, 34, 514–521. [Google Scholar] [CrossRef]
- Skarlato, S.; Filatova, N.; Knyazev, N.; Berdieva, M.; Telesh, I. Salinity Stress Response of the Invasive Dinoflagellate Prorocentrum minimum. Estuar. Coast. Shelf Sci. 2018, 211, 199–207. [Google Scholar] [CrossRef]
- Ligorini, V.; Malet, N.; Garrido, M.; Four, B.; Etourneau, S.; Leoncini, A.S.; Dufresne, C.; Cecchi, P.; Pasqualini, V. Long-Term Ecological Trajectories of a Disturbed Mediterranean Coastal Lagoon (Biguglia Lagoon): Ecosystem-Based Approach and Considering Its Resilience for Conservation? Front. Mar. Sci. 2022, 9, 937795. [Google Scholar] [CrossRef]
- Coelho, S.; Pérez-Ruzafa, A.; Gamito, S. Phytoplankton Community Dynamics in an Intermittently Open Hypereutrophic Coastal Lagoon in Southern Portugal. Estuar. Coast. Shelf Sci. 2015, 167, 102–112. [Google Scholar] [CrossRef]
- Camacho, A.; Peinado, R.; Santamans, A.C.; Picazo, A. Functional Ecological Patterns and the Effect of Anthropogenic Disturbances on a Recently Restored Mediterranean Coastal Lagoon. Needs for a Sustainable Restoration. Estuar. Coast. Shelf Sci. 2012, 114, 105–117. [Google Scholar] [CrossRef]
- Jones, H.F.E.; Özkundakci, D.; McBride, C.G.; Pilditch, C.A.; Allan, M.G.; Hamilton, D.P. Modelling Interactive Effects of Multiple Disturbances on a Coastal Lake Ecosystem: Implications for Management. J. Environ. Manag. 2018, 207, 444–455. [Google Scholar] [CrossRef]
- Fichez, R.; Archundia, D.; Grenz, C.; Douillet, P.; Gutiérrez Mendieta, F.; Origel Moreno, M.; Denis, L.; Contreras Ruiz Esparza, A.; Zavala-Hidalgo, J. Global Climate Change and Local Watershed Management as Potential Drivers of Salinity Variation in a Tropical Coastal Lagoon (Laguna de Terminos, Mexico). Aquat. Sci. 2017, 79, 219–230. [Google Scholar] [CrossRef]
- Biggs, R.; Schlüter, M.; Schoon, M.L. (Eds.) Principles for Building Resilience: Sustaining Ecosystem Services in Social-Ecological Systems; Cambridge University Press: Cambridge, UK, 2015; ISBN 978-1-107-08265-6. [Google Scholar]
- García-Ayllón, S. Integrated Management in Coastal Lagoons of Highly Complexity Environments: Resilience Comparative Analysis for Three Case-Studies. Ocean Coast. Manag. 2017, 143, 16–25. [Google Scholar] [CrossRef]
- De Wit, R.; Leruste, A.; Le Fur, I.; Sy, M.M.; Bec, B.; Ouisse, V.; Derolez, V.; Rey-Valette, H. A Multidisciplinary Approach for Restoration Ecology of Shallow Coastal Lagoons, a Case Study in South France. Front. Ecol. Evol. 2020, 8, 108. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| July-Salinization | October-Freshening | |||||||
|---|---|---|---|---|---|---|---|---|
| Treatment | Initial Salinity In Situ | First Salinization (% of Starting Salinity) | Salinity after First Salinization | Second Salinization (% of Starting Salinity) | Salinity after Second Salinization (Final Salinity) | Initial Salinity In Situ | Freshening (% of Starting Salinity) | Salinity after Freshening (Final Salinity) |
| C | 43 | +0% | 43 | +0% | 43 | 40 | −0% | 40 |
| S1 | +12% | 48 | +23% | 53 | −17% | 33 | ||
| S2 | +23% | 53 | +46% | 63 | −33% | 26 | ||
| S3 | +35% | 58 | +69% | 73 | −50% | 20 | ||
| Pigment | Taxonomic Group | Reference |
|---|---|---|
| Alloxanthin | Cryptophyta | 1, 2, 3 |
| Chlorophyll b | Chlorophyta and green flagellates | 1, 2, 3 |
| Divinyl Chlorophyll a | Prochlorophyta | 2 |
| Fucoxanthin | Bacillariophyceae | 1, 2, 3 |
| Lutein | Chlorophyta and Prasinophyta | 3 |
| Neoxanthin | Chlorophyta and Prasinophyta | 3 |
| Peridinin | Dinophyceae | 1, 2, 3 |
| Prasinoxanthin | Prasinophyta | 3 |
| Zeaxanthin | Cyanobacteria | 1, 2, 3 |
| 19′-Butanoyloxyfucoxanthin | Chrysophyta | 1 |
| 19′-Hexanoyloxyfucoxanthin | Prymnesiophyta | 1 |
| July 2021-Salinization (Cellular Abundance-Cell L−1 ± SD) | October 2021-Freshening (Cellular Abundance-Cell L−1 ± SD) | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Taxonomic Unit | In Situ | C | S1 | S2 | S3 | Taxonomic Unit | In Situ | C | S1 | S2 | S3 | ||||||||
| Bacillariophyceae | Bacillariophyceae | ||||||||||||||||||
| Navicula spp. | 30,828 | 158,689 | ±35,288 | 163,999 | ±26,625 | 115,443 | ±22,675 | 105,124 | ±22,128 | Licmophora spp. | 233,609 | 5078 | ±3044 | 1327 | ±2299 | 20,352 | ±1533 | 7669 | ±2703 |
| Fragilariophycideae | 23,549 | 8259 | ±1842 | 5899 | ±1022 | 5870 | ±1752 | 2832 | ±811 | Nitzchia spp. | 196,444 | 105,505 | ±12,743 | 120,698 | ±20,844 | 192,020 | ±19,925 | 129,193 | ±41,391 |
| Surirella sp. | 12,845 | 0 | ±0 | 0 | ±0 | 0 | ±0 | 0 | ±0 | Navicula spp. | 159,279 | 173,955 | ±67,309 | 135,033 | ±62,317 | 185,826 | ±23,143 | 163,409 | ±27,092 |
| Cocconeis sp. | 11,132 | 9144 | ±2554 | 14,158 | ±9365 | 9494 | ±1957 | 10,088 | ±1405 | Fragilariophycideae | 132,733 | 8502 | ±3054 | 6371 | ±1839 | 8849 | ±1533 | 0 | ±0 |
| Ardissonia sp. | 9420 | 0 | ±0 | 0 | ±0 | 0 | ±0 | 0 | ±0 | Small centric | 69,021 | 48,206 | ±10,089 | 61,234 | ±9866 | 143,351 | ±14,047 | 34,216 | ±14,195 |
| Nitzschia sp. | 5994 | 61,057 | ±16,813 | 63,712 | ±19,467 | 42,618 | ±14,367 | 40,882 | ±8692 | Petroneis sp. | 47,784 | 0 | ±0 | 3451 | ±2871 | 0 | ±0 | 0 | ±0 |
| Petroneis sp. | 3854 | 295 | ±511 | 0 | ±0 | 0 | ±0 | 0 | ±0 | Surirella sp. | 37,165 | 0 | ±0 | 354 | ±613 | 0 | ±0 | 0 | ±0 |
| Licmophora sp. | 3425 | 2655 | ±885 | 1770 | ±1770 | 0 | ±0 | 0 | ±0 | Chaetoceros sp. | 18,583 | 77,203 | ±22,813 | 101,585 | ±26,362 | 161,049 | ±12,545 | 87,309 | ±28,224 |
| Stauroneis sp. | 3425 | 0 | ±0 | 0 | ±0 | 0 | ±0 | 0 | ±0 | Cocconeis sp. | 18,583 | 3853 | ±1263 | 3274 | ±668 | 5309 | ±4598 | 0 | ±0 |
| Cylindrotheca closterium | 2141 | 15,633 | ±2703 | 5899 | ±2703 | 4059 | ±6436 | 0 | ±0 | Gyrosigma/Pleurosigma sp. | 13,273 | 2008 | ±1991 | 442 | ±766 | 0 | ±0 | 0 | ±0 |
| Diploneis sp. | 1713 | 0 | ±0 | 0 | ±0 | 0 | ±0 | 0 | ±0 | Diploneis sp. | 7964 | 340 | ±589 | 0 | ±0 | 0 | ±0 | 0 | ±0 |
| Entomoneis sp. | 0 | 31,856 | ±20,809 | 47,194 | ±34,571 | 38,765 | ±8623 | 15,397 | ±10,737 | Amphora sp. | 5309 | 0 | ±0 | 0 | ±0 | 0 | ±0 | 0 | ±0 |
| Amphora sp. | 0 | 5014 | ±2845 | 5309 | ±1770 | 2256 | ±915 | 2478 | ±1105 | Entomoneis sp. | 2655 | 26,315 | ±7830 | 60,084 | ±11,201 | 324,752 | ±19,925 | 355,723 | ±54,863 |
| Chaetoceros sp. | 0 | 2065 | ±3576 | 6489 | ±2044 | 239 | ±414 | 0 | ±0 | Cerataulina sp. | 0 | 2464 | ±2464 | 1770 | ±1533 | 17,698 | ±12,261 | 3540 | ±1770 |
| Grammatophora sp. | 0 | 18,760 | ±9047 | 5752 | ±9962 | 7964 | ±13,794 | 0 | ±0 | ||||||||||
| Dinophyceae | Dinophyceae | ||||||||||||||||||
| Dinophyceae und. | 23,978 | 2360 | ±1022 | 2950 | ±2703 | 4887 | ±4559 | 2124 | ±1062 | Dinophyceae und. | 76,985 | 0 | ±0 | 0 | ±0 | 0 | ±0 | 0 | ±0 |
| Mesoporos sp. | 1713 | 0 | ±0 | 0 | ±0 | 0 | ±0 | 0 | ±0 | Alexandrium sp. | 79,640 | 12,402 | ±807 | 13,539 | ±8281 | 17,698 | ±5526 | 2360 | ±1022 |
| Prorocentrum spp. | 856 | 0 | ±0 | 0 | ±0 | 0 | ±0 | 0 | ±0 | Prorocentrum micans | 13,273 | 6643 | ±1655 | 1681 | ±853 | 3540 | ±1533 | 0 | ±0 |
| Dinophysis sp. | 0 | 0 | ±0 | 0 | ±0 | 239 | ±414 | 0 | ±0 | Akashiwo sanguinea | 0 | 442 | ±766 | 4513 | ±3326 | 2655 | ±2655 | 9439 | ±1022 |
| Chlorophyta | 0 | 0 | ±0 | 0 | ±0 | 0 | ±0 | 0 | ±0 | ||||||||||
| Pyramimonas sp. | 18,411 | 21,827 | ±9210 | 0 | ±0 | 0 | ±0 | 0 | ±0 | ||||||||||
| Cryptophyceae | 58,402 | 20,625 | ±1061 | 34,864 | ±7439 | 79,640 | ±5309 | 33,036 | ±5109 | ||||||||||
| Others | Others | ||||||||||||||||||
| Small flagellates und. | 0 | 20,647 | ±2554 | 29,496 | ±5407 | 26,235 | ±6537 | 8672 | ±2146 | Dictyochophyceae | |||||||||
| Small coccoid und. | 0 | 0 | ±0 | 42,474 | ±3540 | 40,671 | ±7797 | 23,715 | ±8648 | Dictyocha sp. | 2655 | 0 | ±0 | 0 | ±0 | 0 | ±0 | 0 | ±0 |
| Raphidophyceae | 0 | 5899 | ±2703 | 0 | ±0 | 0 | ±0 | 0 | ±0 | ||||||||||
| und.1 | 0 | 0 | ±0 | 0 | ±0 | 2476 | ±1529 | 1239 | ±2146 | ||||||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ligorini, V.; Garrido, M.; Malet, N.; Simon, L.; Alonso, L.; Bastien, R.; Aiello, A.; Cecchi, P.; Pasqualini, V. Response of Phytoplankton Communities to Variation in Salinity in a Small Mediterranean Coastal Lagoon: Future Management and Foreseen Climate Change Consequences. Water 2023, 15, 3214. https://doi.org/10.3390/w15183214
Ligorini V, Garrido M, Malet N, Simon L, Alonso L, Bastien R, Aiello A, Cecchi P, Pasqualini V. Response of Phytoplankton Communities to Variation in Salinity in a Small Mediterranean Coastal Lagoon: Future Management and Foreseen Climate Change Consequences. Water. 2023; 15(18):3214. https://doi.org/10.3390/w15183214
Chicago/Turabian StyleLigorini, Viviana, Marie Garrido, Nathalie Malet, Louise Simon, Loriane Alonso, Romain Bastien, Antoine Aiello, Philippe Cecchi, and Vanina Pasqualini. 2023. "Response of Phytoplankton Communities to Variation in Salinity in a Small Mediterranean Coastal Lagoon: Future Management and Foreseen Climate Change Consequences" Water 15, no. 18: 3214. https://doi.org/10.3390/w15183214