
Zooplankton in the Technogenic Aquatic Ecosystem of the North Crimean Canal, the Longest in Europe

A.O. Kovalevsky Institute of Biology of the Southern Seas of RAS, 2 Nakhimov Ave., 299011 Sevastopol, Russia
*
Author to whom correspondence should be addressed.
Water 2023, 15(13), 2327; https://doi.org/10.3390/w15132327
Submission received: 1 June 2023
/
Revised: 16 June 2023
/
Accepted: 21 June 2023
/
Published: 22 June 2023
(This article belongs to the Special Issue Biology and Ecology of Zooplankton: Latest Advances and Prospects)
Abstract
:The construction and use of navigation and irrigation canals are among the common human alterations in landscapes and ecosystems. The North Crimean Canal (NCC) is the longest in Europe; its main branch is 403 km, and the total of all its branches exceeds 10,000 km. It is a main driver of the significant environmental changes in an all-arid part of Crimea. No zooplankton studies in the canal have been carried out up to this time at all. In 2022, zooplankton was studied in different sites from May to October. Total zooplankton abundance fluctuated between 170 and 19,560 ind. m−3, and wet biomass between 0.75 and 1057 mg m−3. In total, 11 Rotifera species, 11 Cladocera species, and 15 Copepoda species, including 2 Harpacticoida, 5 Calanoida, and 8 Cyclopoida, were found in plankton. The most common Rotifera species were Brachionus calyciflorus, Euchlanis dilatate, and Keratella cochlearis. Among Cladocera, Chydorus sphaericus, Bosmina longirostris, and Coronatella rectangular have the highest frequency of occurrence. In Copepoda, these were Acanthocyclops vernalis, Eurytemora affinis, and Nitocra typica. Among the 37 species found in NCC, 10 were not previously noted in the water bodies of Crimea. Among rotifers, they were Ploesoma hudsoni, Polyarthra dolichoptera, Pompholyx complanata, Synchaeta grandis, and Synchaeta stylata. Bosminopsis zernowi, Rhynchotalona rostrata, and Scapholeberis mucronata were not noted before among Cladocera. Eurytemora lacustris and Cyclops smirnovi were first found in Crimea among Copepoda. New technogenic aquatic ecosystems such as NCC are the “gateway” for the invasion of alien species into the new regions.
1. Introduction
The first technogenic and natural-technogenic water bodies on the planet probably began to appear no later than about 8000–9000 years ago, as irrigation systems. According to archaeological evidence, the irrigation canals in the areas with a lack of rainfall to support crops were about 6000 BCE in Mesopotamia [1,2,3]. The Indus Valley Civilization constructed irrigation canals and water-storage systems as artificial reservoirs at Girnar during 3000 BCE [4,5]. The Egyptian pharaoh Amenemhet IV (about 1800 BCE) built a canal to the natural lake in the Faiyum Oasis (now Lake Qarun) from the Nile to store water for use during dry seasons [6,7]. Currently, this canal works and expands [8]. In the Andes Mountain valley (Peru), remains of irrigation canals were found with radiocarbon-dated from the fourth millennium BCE, and some traces under one canal may be dating from the fifth millennium BCE [9]. Since that time, the demand for irrigated land has increased rapidly, as well as the total length of irrigation canals [10]. Currently, worldwide irrigation water withdrawals were estimated at 2881 km3 year−1 [11].
Navigable canals (artificial waterways) connect rivers, lakes, and seas, and are designed, as a rule, for all kinds of water transport from small boats to huge bulk carriers. Early canals were connected with natural rivers, and one of the first such canals was the Grand Canal of China, which was constructed starting in the fifth century BCE during the Sui Dynasty [3,12]. For thousands of years, humans constructed more and more artificial waterways, and currently, there are more than 63,000 km of navigation canals worldwide [13].
Currently, the construction and use of navigation and irrigation canals are among the common human alterations in landscapes and ecosystems [14,15,16]. To date, a huge amount of evidence has accumulated that, solving social and economic problems, the construction of any canals changes the structure of landscape connectivity and has multiple, mostly negative, impacts on the environment [13,17,18,19]. Every built irrigation canal can lead to changes in hydrology, pollution, soil degradation, biological and ecological alterations, human health, and social–economic values [20,21,22]. All effects are more pronounced in arid and semi-arid areas with irrigation canal systems [21,23,24]. In these cases, the environmental impact can derive due to the water supply as well as the operation of irrigation practices influencing the environment in a variety of ways [13,17,19]. Irrigation generates a large volume of drainage water, usually discharging from agricultural fields into some depressions creating artificial lakes, and also leading to significant landscape alterations [8]. Nutrient discharge is a major issue associated with irrigated agricultural lands around the world, including with the Salton Sea [25,26]. The Salton Sea (USA) is the largest such natural-technogenic lake (area 930–1000 km2) in the world [27]. In Egypt, the Canal Bahr Yousef was constructed in early Pharaonic times to transport fresh water from the Nile to the desert areas [28]. Its branches provide water for agricultural use in the Fayoum oasis. Drainage waters from irrigated lands are discharged, gradually forming several artificial lakes [8]. These man-made lakes significantly change the system of landscape connections in total but are valuable for biodiversity conservation creating new habitats for many species with a high increase in local biodiversity, especially of birds [8].
Every canal system is recognized worldwide as a corridor for invasive species distribution [29,30,31,32,33]. As an example, 443 multicellular alien species were introduced into the Mediterranean Sea through the Suez Canal in total [34]. Constructed canals, and shipping through them, contributed to the introduction of most nonindigenous species in the lake systems such as the Laurentian Great Lakes and the Caspian Sea [35]. By increasing the likelihood of non-native species entering the region, channels also gain a foothold in the region by creating new habitats as an additional opportunity. Therefore, in arid areas, due to the appearance of new types of habitats (wetlands), canal construction can increase species richness—not only aquatic but also birds and mammals [8,36].
Irrigated agricultural lands provide about 40% of the total food produced worldwide [11,37], and therefore, the growth of irrigated lands and canals providing water to them will continue in arid and semi-arid zones, as well as their impact on the environment. Aquatic ecosystems of irrigation canal systems are complicated playing an important role in landscapes and support of total local biodiversity [32,38,39]. They also provide a lot of social and economic services not only for agriculture development [13,40]. All these must be taken into account when developing strategies and plans for a multipurpose irrigation canal system use. Integrated management of the canals involves multiple goals across many agencies and stakeholders, often with different or conflicting objectives [13]. Development of such strategies and plans must be based on adequate scientific background which also contains knowledge of the peculiarities of unique technogenic ecosystems of the canals. Unfortunately, at present, there is a shortage of such knowledge that does not allow making generalizations about these ecosystems, their structure, functioning, and dynamics. There are only scattered ecological and biological data on a small number of irrigation channels [35,39,41,42,43]. Zooplankton plays a key role in different aquatic ecosystems, but few articles were devoted to its study in canals [44,45,46,47,48,49,50]. Therefore, for a better understanding of the characteristics of technogenic ecosystems of irrigation canals, it is necessary to accumulate and comprehend new data for various specific water bodies.
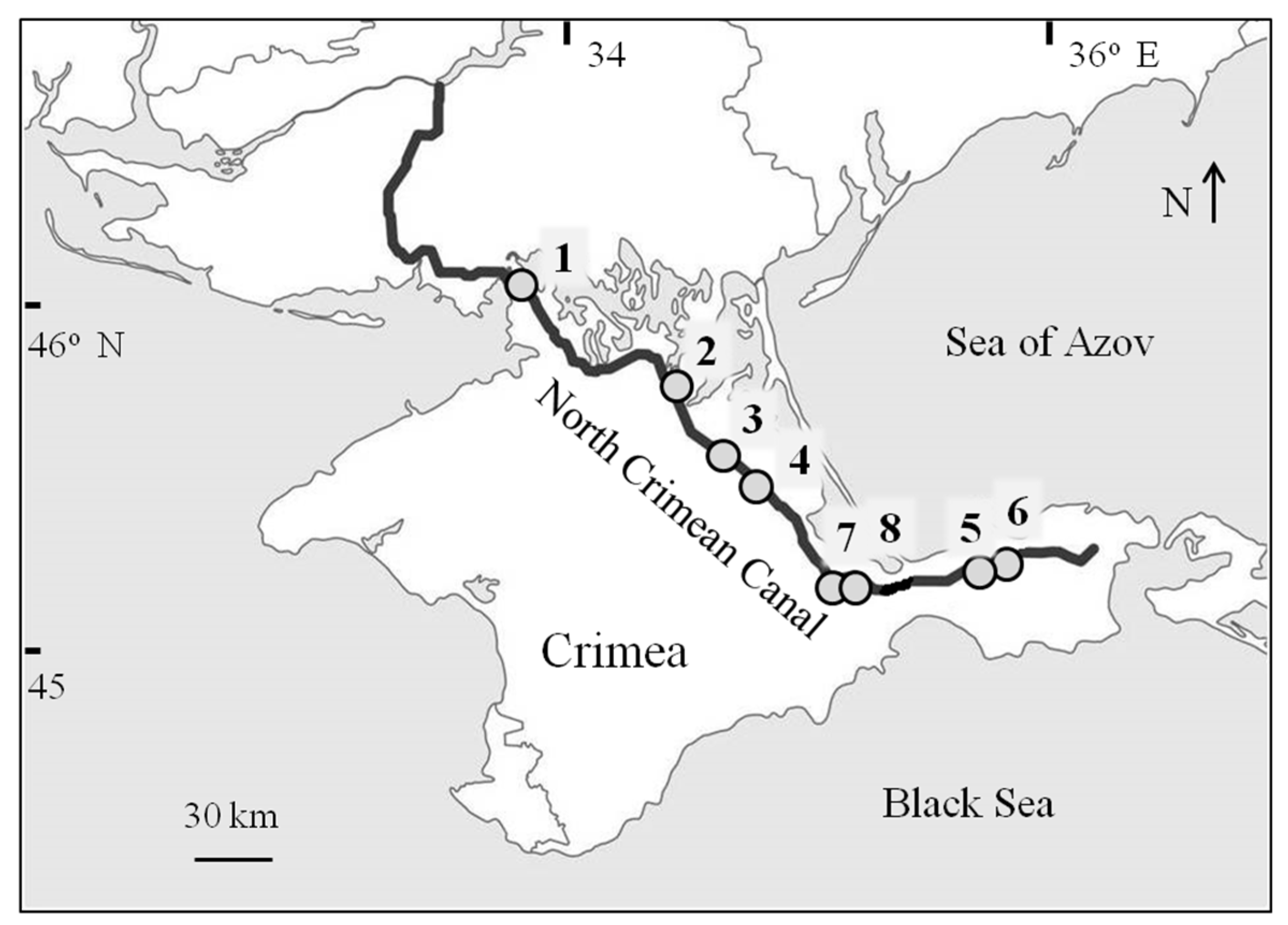
The North Crimean Canal (NCC) is the longest in Europe (Figure 1); the length of its main branch is 403 km, the total of all its branches exceeds 10,000 km, and its construction (1963–1976) has become a main driver of significant environmental changes in an all arid parts of Crimea [51,52,53,54,55].
Despite the huge size of the NCC and its significant impact on the environment, no systematic studies of the canal ecosystem have been carried out up to this time, although separate scattered studies have been done. Studies of zoobenthos [56,57] and fish [36] were occasionally carried out. There are no known studies of zooplankton from this canal system. The conducted studies showed a rather high species richness of fish in NCC 19 [36] and mollusks [54], among which most species are new to Crimea. The NCC construction led to the desalination of most hypersaline lakes and the Sivash Bay, and as a result, to the formation of giant beds of the common reed Phragmites australis (Cav.) Trin. ex Steud. [52,54], which made it possible for the muskrat Ondatra zibethicus (Linnaeus, 1766) and the common raccoon dog Nyctereutes procyonoides (Gray, 1834) to independently settle through NCC in Crimea, and moreover, spread widely in it [58].
For political reasons, in April 2014, the flow of water into the canal was blocked [52], and at the end of February 2022, it was renewed. During the period when the Dnieper water did not enter the canal for a considerable length, the canal dried up (Figure 2), in some sections of the canal, the water remained but its level decreased significantly (Figure 2).
In March 2022, the canal was filled with water (Figure 3); the study of zooplankton was started and continued until October of this year.
The objectives of the article are: 1. to give an overview of the first data on zooplankton of NCC, analyzing them; 2. to test the hypotheses that (a) the structure and seasonal dynamics of zooplankton are similar to those noted in the Dnieper reservoirs, and (b) in the canal, there are zooplankton species, which are new for Crimea.
2. Materials and Methods
2.1. Site Description
The North Crimean Canal (Figure 1, Figure 2 and Figure 3) was built in 1961–1971 to solve the problem of an acute shortage of fresh water in the Crimea (the largest peninsula in the Black Sea, area of 27,000 km2) and the development of irrigated agriculture [52,59]. The main water intake of NCC is located at the Kakhovka Reservoir on the Dnieper River, where water flows by gravity through the main lock for a distance of 208 km. Water runs for 402.6 km in the main canal branch in a generally southeasterly direction. At this distance, there are several water pumping stations, which in total raise water by more than 80 m. Between these stations, water flows by gravity. The width of the canal at its beginning is 150 m; the depth is 7 m. The average annual flow is 380 m3 s−1. The maximum technologically possible flow is up to 500 m3 s−1 (30% of the Dnieper’s flow in its lower reaches, equal to 1670 m3 s−1). The construction of the first stage of the NCC began in 1961; water reached the town of Krasnoperekopsk in 1963, and in 1971, it came to the Kerch Peninsula. Independent main canals of irrigation systems branch off from the main channel of NCC, and 7 reservoirs were also built in Crimea. Before 2014, Crimea satisfied about 85% of its water needs precisely at the expense of NCC, which is the longest irrigation system in Europe. The length of NCC from Novaya Kakhovka to Kerch is 405 km, and the total length of the main canal and all its branches exceeds 10,000 km. In 1986, the area of land irrigated with water from NCC was 3000 km2 and the area of land provided by water was 6000 km2. In the canal—almost all the time—there were high concentrations of phytoplankton (Figure 4), with episodic outbreaks of cyanobacterial blooms.
The NCC activity resulted in the formation and widening of beds of Ph. Australis, which provided a new attraction for Crimea bird species [52,53]. In the canal, good conditions for macrophytes formed, and they began to develop intensively, reaching a length of more than 4–5 m and creating dense thickets (Figure 4). Among the most widespread and abundant aquatic plants, three or four species of Potamogeton, as well as Myriophyllum, Elodea, and others were noted. Part of the plants break off and drifts in the flow of water downstream. This poses a threat to pumping stations, so there are devices in front of them that catch drifting plants and throw them ashore, several tons per day in summer (Figure 5). Various animals are removed together with the plants, including the swimming with them of, for example, mollusks Anodonta, Dreissena, Lymnaea, Unio (Figure 5).
2.2. Sampling and Processing
The collection of zooplankton in the main branch of the canal was carried out at several places from March to October. The distribution of sampling points and dates of sampling are presented (Figure 1 and Table 1). Every time near shores, the water samples of 100–150 L were taken from upper 0.5–0.7 m by filtering through a Judy plankton net (55-µm diameter mesh size), with two duplications. Zooplankton samples were fixed with 4% formalin. In parallel, measurements of salinity, temperature, pH, and Eh (redox potential) were carried out by Kellong WZ212 and an electronic thermometer PHH-830, respectively. Using a Bogorov chamber, samples were analyzed under a stereomicroscope Olympus SZ61TR (Olympus, Tokyo, Japan). Identification of species was carried out under a light microscope Olympus BX53F using different keys [60,61,62,63,64,65]. The size of the animals was determined using an eyepiece micrometer, and their mass was calculated using the corresponding formulas “wet mass – size” [66]. Biomass was calculated as the sum of individuals, taking into account their individual mass.
2.3. Data Processing
Data processing was conducted by standard statistical methods [67]. The standard deviation (SD), coefficients of variability (CV), correlation (R), and determination (R2), as well as the parameters of regression equations, were calculated in MS Excel. The confidence level of correlation coefficients (p) was evaluated by comparing them with critical values [68].
There is a strong relation between the found species richness and the number of analyzed samples [69,70,71]. Power or logarithmic functions are most used to approximate such a relationship between sampling effort (the number of samples) and the observed species richness. To generate a variety of ordering among available samples for calculation of “the number of species found − sample analyzed” curves, random permutations of data were done online (http://www.ebcalculator.co.uk/statistics/rpermute3.htm (accessed on 20 May 2023) Average values of expected species richness were calculated from all made permutations.
3. Results
In total, 11 Rotifera species, 11 Cladocera species, and 15 Copepoda species, including 2 Harpacticoida, 5 Calanoida, and 8 Cyclopoida, were found in plankton (Table 2).
Ostracoda and Gammaridae (juveniles) were found once in March, and veligers of Dreissena (Mollusca) were found once in April. In one sample, there were a maximum of 4 species of Rotifera, 6 Cladocera, and 10 Copepoda; the maximum total species richness in 1 sample was 19 in April, and then it decreased, changing from 5 to 10 species per sample (Table 2). The maximum number of Rotifera species was recorded in March and June; Cladocera and Copepoda in April (Table 2). The most common Rotifera species were Brachionus calyciflorus, Euchlanis dilatate, and Keratella cochlearis (all 30% of total abundance). Among Cladocera, Chydorus sphaericus (70%), Bosmina longirostris (60%), and Coronatella rectangular (40%) have the highest frequency of occurrence. In Copepoda, these were Acanthocyclops vernalis (80%), Eurytemora affinis (50%), and Nitocra typica (50%).
The number of Rotifera species did not correlate with that of Cladocera and Copepoda. There was a significant positive relationship between the number of species in the sample of Cladocera and Copepoda (R = 0.688, p = 0.01):
where X is the number of Cladocera species, and Y is the number of Copepoda species.
Y = 0.95 + 0.45X,
The total abundance of zooplankton varied from 170 (May) to 91,470 ind. m−3 (April) (Table 3).
In mid-March, the total abundance of zooplankton was low with the dominance of Rotifera (79% of the total abundance). From mid-March to the end of April, there was a rapid increase in the total abundance of zooplankton, mainly due to Cladocera (55–80% of the total abundance). Abundance increased in May and was relatively high until October. In total, Rotifera dominated in 2 samples (March and June), Cladocera in 4 samples (April, May, and June), and Copepoda in 4 samples (May, June, August, and October) (Table 4). The largest abundance among Rotifera was noted in Brachionus angularis (1840 ind. m−3), among Cladocera in Chydorus sphaericus (72,000 ind. m−3), and Copepoda in Eurytemora lacustris (1920 ind. m−3) and Eucyclops serrulatus (2240 ind. m−3).
4. Discussion
It is obvious that the found species does not include all available species in the canal. The question arises: to what degree, then, was the species’ richness explored in the canal? Based upon data from this study, the species accumulation curves for Rotifera, Cladocera, and Copepoda were calculated for real ordering of sample taking. For Rotifera, such an equation is (R = 0.988, p = 0.001):
for Cladocera (R = 0.855, p = 0.005):
for Copepoda (R = 0.862, p = 0.005):
where NR, NCl, and NCo are the number of species for Rotifera, Cladocera, and Copepoda, respectively, and K is the number of analyzed samples.
NR = 4.21 K0.484,
NCl = 2.08 K0.858,
NCo = 2.14 K0.861,
These functions correspond to 1 of the many possible orderings of 10 samples. The expected species richness for 300 samples, averaging different orderings of the samples, were: for Rotifera—54 species (SD = 28.5), Cladocera—72 species (SD = 95.8), and Copepoda—85 species (SD = 83.6). Based on the hypothesis that the structure of zooplankton in the canal is similar to that in the reservoirs on the Dnieper River as in other such cases [72], we compared these values and the list of species (Table 2) with what is known about the reservoirs. To do a comparison, we used available zooplankton data from the reservoirs of the Dnieper [73,74,75,76,77,78,79,80,81] and small rivers flowing into it [82,83,84]. The comparison showed that the total species richness of Rotifera, Cladocera, and Copepoda in plankton is very close to the calculated expected values in the canal. For example, as a result of processing 162 samples of zooplankton from the Kakhovka Reservoir, 57 species of Rotifera, 47 species of Cladocera, and 43 species of Copepoda were identified [74]. The expected species richness was calculated by all our equations with a different ordering. The averaged expected species richness for 162 samples were: Rotifera—49 species (SD = 25.5), Cladocera—62 species (SD = 66.8), and Copepoda—64 species (SD = 63.4). No significant differences exist with observed values [74]. All species found in NCC, except Cyclops smirnovi, were previously found in the Dnieper reservoirs and/or in the rivers flowing into the Dnieper. As shown in the works cited above, there is very high spatiotemporal variability in the species structure of zooplankton in the Dnieper reservoirs. The same can be noted for the NCC, but there is still not enough data to discuss this.
The list of species found in the canal (Table 2) was compared with those previously noted in Crimea [85,86,87,88]. Among the 37 species found in NCC, 10 were not previously noted in the water bodies of Crimea. Among rotifers, these include 5 species of Ploesoma hudsoni, Polyarthra dolichoptera, Pompholyx complanata, Synchaeta grandis, and Synchaeta stylata. However, it should be noted that after the work [85], serious studies of this group in the inland waters of the peninsula were not carried out. Bosminopsis zernowi, Rhynchotalona rostrata, and Scapholeberis mucronata were not noted before among Cladocera. A good revision of Bosminopsis deitersi species group was recently made, and it is shown that a widely distributed cryptic species in the Old World is Bosminopsis zernowi Linko, 1901 [89]. At the same time, only one old occurrence of the species in Russia and Europe as a whole was indicated [89,90]. This does not correspond to the fact that earlier B. zernowi was found in the rivers of the upper reaches of the Dnieper [82], the Dnieper reservoirs (Yakovenko, own unpublished data), and now, the species has been found in the NCC. It can be fairly and confidently asserted that the species came to the peninsula because of the canal.
In the present study, Eurytemora lacustris and Cyclops smirnovi were first found in Crimea. E. lacustris showed the highest abundance among the copepods in NCC. Earlier, cladocerans Daphnia cucullate, Leptodora kindtii, copepods Mesocyclops leuckarti, Heterocope appendiculata and E. velox most likely entered Crimea through the canal and became widespread [85,86,87,88]. The estuarine calanoid copepod E. velox also invaded the man-made inland water bodies in Britain [91]. Cyclops smirnovi is a rare valid species [92], which until then was found as an abundant species only in a high mountain lake in Altai (Western Siberia) in the first half of the 20th century [61]. Most likely, it was brought to the new region by migratory birds; earlier, such a path was noted for several Asian cyclopoid copepods, which recently settled in Crimea [70].
It should be noted that, in Crimea, all introduced Asian species of cyclopoids were found in new or catastrophically changed water bodies [70]. In Lugansk (Donbas region) it was noted exactly the same [93]. Several authors, after analyzing numerous data, concluded that new technogenic aquatic ecosystems or significantly altered natural ones are the “gateway” for the invasion of alien species into the new regions [70,91,94,95,96]. NCC is an example of this. A deeper study of this landscape function is necessary for the organization of an integrated multi-purpose sustainable management of NCC.
The NCC has a significant impact on the entire natural environment of Crimea [36,52,53], and its socioeconomic significance is not limited to the value of irrigated agriculture. The canal is a popular place for fishing and recreation of residents, the huge biomass of harvested macrophytes (Figure 5) can be used profitably for the production of fertilizers and feed for farm animals. It is possible to rationally combine all its functions and services only with the transition to integrated sustainable management. This requires, among other things, a comprehensive study of the canal ecosystem, zooplankton being its important component. Only the first step has been taken in this direction.
5. Conclusions
In 2022, zooplankton was present in the canal during all observation periods. Its species composition, abundance and biomass varied widely. Total zooplankton abundance fluctuated between 170 and 19,560 ind. m−3, and wet biomass between 0.75 and 1057 mg m−3. In total, 11 Rotifera species, 11 Cladocera species, and 15 Copepoda species, including 2 Harpacticoida, 5 Calanoida, and 8 Cyclopoida, were found in plankton. Among the 37 species found in NCC, 10 were not previously noted in the water bodies of the Crimean Peninsula.
Author Contributions
Conceptualization, N.S. and E.A.; methodology, N.S.; field studies, N.S., V.Y., I.M. and E.A.; sample processing, V.Y. and E.A.; formal analysis, N.S.; writing—original draft preparation, N.S.; writing—review and editing, N.S., V.Y., I.M. and E.A. All authors have read and agreed to the published version of the manuscript.
Funding
The study was conducted in the framework of the state assignments of A.O. Kovalevsky Institute of Biology of the Southern Seas of RAS (No. 121041500203-3 and 121031500515-8).
Data Availability Statement
All data used in this study are available upon request from the corresponding author.
Acknowledgments
The authors thank Bindy Datson (Australia) for her help to improve English.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Lawton, H.W.; Wilke, P.J. Ancient Agricultural Systems in Dry Regions of the Old World. In Agriculture in Semi-Arid Environments; Hall, A.E., Cannell, G.H., Lawton, H.W., Eds.; Ecological Studies; Springer Science & Business Media: Berlin/Heidelberg, Germany, 1979; Volume 34, pp. 1–44. [Google Scholar]
- Flannery, K.V. Origins and ecological effects of early domestication in Iran and the Near East. In The Domestication and Exploitation of Plants and Animals; Ucko, P.J., Dimbleby, G.W., Eds.; Transaction Publishers: New Brunswick, NJ, USA, 1969; pp. 73–100. [Google Scholar]
- Huang, W.; Xi, M.; Lu, S.; Taghizadeh-Hesary, F. Rise and fall of the grand canal in the ancient Kaifeng city of China: Role of the grand canal and water supply in urban and regional development. Water 2021, 13, 1932. [Google Scholar] [CrossRef]
- Rodda, J.C.; Ubertini, L. The Basis of Civilization—Water Science? International Association of Hydrological Sciences: Oxfordshire, UK, 2004; p. 342. [Google Scholar]
- Thomas, A.R.; Fulkerson, G.M. City and Country: The Historical Evolution of Urban-Rural Systems; Rowman & Littlefield: Lanham, MD, USA, 2021; p. 137. [Google Scholar]
- Baioumy, H.M.; Kayanne, H.; Tada, R. Reconstruction of lake-level and climate changes in Lake Qarun, Egypt, during the last 7000 years. J. Great Lakes Res. 2010, 36, 318–327. [Google Scholar] [CrossRef]
- Shadrin, N.V.; El-Shabrawy, G.M.; Anufriieva, E.V.; Goher, M.E.; Ragab, E. Long-term changes of physicochemical parameters and benthos in Lake Qarun (Egypt): Can we make a correct forecast of ecosystem future? Knowl. Manag. Aquat. Ecosyst. 2016, 417, 18. [Google Scholar] [CrossRef] [Green Version]
- Anufriieva, E.V.; Goher, M.E.; Hussian, A.M.; El-Sayed, S.M.; Hegab, M.H.; Tahoun, U.M.; Shadrin, N.V. Ecosystems of artificial saline lakes. A case of Lake Magic in Wadi El-Rayan depression (Egypt). Knowl. Manag. Aquat. Ecosyst. 2020, 421, 31. [Google Scholar] [CrossRef]
- Dillehay, T.D.; Eling, H.H., Jr.; Rossen, J. Preceramic irrigation canals in the Peruvian Andes. Proc. Natl. Acad. Sci. USA 2005, 102, 17241–17244. [Google Scholar] [CrossRef] [Green Version]
- De Wrachien, D.; Schultz, B.; Goli, M.B. Impacts of population growth and climate change on food production and irrigation and drainage needs: A world-wide view. Irrig. Drain. 2021, 70, 981–995. [Google Scholar] [CrossRef]
- Droppers, B.; Supit, I.; Van Vliet, M.T.; Ludwig, F. Worldwide water constraints on attainable irrigated production for major crops. Environ. Res. Lett. 2021, 16, 055016. [Google Scholar] [CrossRef]
- Harrington, L. The Grand Canal of China; Bailey Brothers and Swinfen Ltd.: Folkestone, UK, 1974; p. 110. [Google Scholar]
- Bergman, J.N.; Beaudoin, C.; Mistry, I.; Turcotte, A.; Vis, C.; Minelga, V.; Neigel, K.; Lin, H.Y.; Bennett, J.R.; Young, N.; et al. Historical, contemporary, and future perspectives on a coupled social–ecological system in a changing world: Canada’s historic Rideau Canal. Environ. Rev. 2022, 30, 72–87. [Google Scholar] [CrossRef]
- Hupp, C.R.; Pierce, A.R.; Noe, G.B. Floodplain geomorphic processes and environmental impacts of human alteration along coastal plain rivers, USA. Wetlands 2009, 29, 413–429. [Google Scholar] [CrossRef]
- Chen, J.; Zeng, X.; Deng, Y. Environmental pollution and shipping feasibility of the Nicaragua Canal. Mar. Pollut. Bull. 2016, 113, 87–93. [Google Scholar] [CrossRef]
- Paul, S.S.; Hasselquist, E.M.; Jarefjäll, A.; Agren, A.M. Virtual landscape-scale restoration of altered channels helps us understand the extent of impacts to guide future ecosystem management. Ambio 2023, 52, 182–194. [Google Scholar] [CrossRef] [PubMed]
- Hatfield, J.L. Environmental impact of water use in agriculture. Agron. J. 2015, 107, 1554–1556. [Google Scholar] [CrossRef] [Green Version]
- Zhuang, W. Eco-environmental impact of inter-basin water transfer projects: A review. Environ. Sci. Pollut. Res. 2016, 23, 12867–12879. [Google Scholar] [CrossRef]
- Li, Q.; Li, X.; Ran, Y.; Feng, M.; Nian, Y.; Tan, M.; Chen, X. Investigate the relationships between the Aral Sea shrinkage and the expansion of cropland and reservoir in its drainage basins between 2000 and 2020. Int. J. Digit. Earth 2021, 14, 661–677. [Google Scholar] [CrossRef]
- Causapé, J. Agro-environmental evaluation of irrigation land. II. Pollution induced by Bardenas Irrigation District (Spain). Agric. Water Manag. 2009, 96, 188–194. [Google Scholar]
- Fernández-Cirelli, A.; Arumí, J.L.; Rivera, D.; Boochs, P.W. Environmental effects of irrigation in arid and semi-arid regions. Review. Chil. J. Agric. Res. 2009, 69 (Suppl. S1), 27–40. [Google Scholar] [CrossRef] [Green Version]
- Li, D.; Zhang, J.; Wang, G.; Wang, X.; Wu, J. Impact of changes in water management on hydrology and environment: A case study in North China. J. Hydro-Environ. Res. 2020, 28, 75–84. [Google Scholar] [CrossRef]
- López-Pomares, A.; López-Iborra, G.M.; Martín-Cantarino, C. Irrigation canals in a semi-arid agricultural landscape surrounded by wetlands: Their role as a habitat for birds during the breeding season. J. Arid Environ. 2015, 118, 28–36. [Google Scholar] [CrossRef]
- Thorslund, J.; Bierkens, M.F.; Oude Essink, G.H.; Sutanudjaja, E.H.; van Vliet, M.T. Common irrigation drivers of freshwater salinisation in river basins worldwide. Nat. Commun. 2021, 12, 4232. [Google Scholar] [CrossRef]
- Tompson, A.F. Born from a flood: The Salton Sea and its story of survival. J. Environ. Sci. 2016, 27, 89–97. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; An, C.; Tan, Q.; Tian, X.; Li, G.; Zhou, Y. Spatiotemporal analysis of land use pattern and stream water quality in southern Alberta, Canada. J. Contam. Hydrol. 2021, 242, 103852. [Google Scholar] [CrossRef]
- Schroeder, R.A.; Orem, W.H.; Kharaka, Y.K. Chemical evolution of the Salton Sea, California: Nutrient and selenium dynamics. Hydrobiologia 2002, 473, 23–45. [Google Scholar] [CrossRef]
- Hassan, F.A. Holocene lakes and prehistoric settlements of the western Fayum, Egypt. J. Archaeol. Sci. 1986, 13, 483–501. [Google Scholar] [CrossRef]
- Gollasch, S.; Galil, B.S.; Cohen, A.N. Bridging Divides: Maritime Canals as Invasion Corridors; Springer: Dordrecht, The Netherlands, 2006; 316p. [Google Scholar]
- Kim, J.; Mandrak, N.E. Assessing the potential movement of invasive fishes through 1143 the Welland Canal. J. Great Lakes Res. 2016, 42, 1102–1108. [Google Scholar] [CrossRef] [Green Version]
- Martel, A.L.; Madill, J.B. Twenty-six years (1990–2015) of monitoring annual 1209 recruitment of the invasive zebra mussel (Dreissena polymorpha) in the Rideau River, a small 1210 river system in Eastern Ontario, Canada. Can. J. Zool. 2018, 96, 1071–1079. [Google Scholar] [CrossRef] [Green Version]
- Lin, H.Y.; Cooke, S.J.; Wolter, C.; Young, N.; Bennett, J.R. On the conservation value of historic canals for aquatic ecosystems. Biol. Conserv. 2020, 251, 108764. [Google Scholar] [CrossRef]
- Asth, M.D.; Rodrigues, R.G.; Zenni, R.D. Canals as invasion pathways in tropical dry forest and the need for monitoring and management. J. Appl. Ecol. 2021, 58, 2004–2014. [Google Scholar] [CrossRef]
- Galil, B.; Marchini, A.; Occhipinti-Ambrogi, A.; Ojaveer, H. The enlargement of the Suez Canal—Erythraean introductions and management challenges. Manag. Biol. Invasions 2017, 8, 141–152. [Google Scholar] [CrossRef] [Green Version]
- Grigorovich, I.A.; MacIsaac, H.J.; Shadrin, N.V.; Mills, E.L. Patterns and mechanisms of aquatic invertebrate introductions in the Ponto-Caspian region. Can. J. Fish. Aquat. Sci. 2002, 59, 1189–1208. [Google Scholar] [CrossRef]
- Kostyushin, V.A.; Bagrikova, N.A.; Kostin, S.Y.; Karpenko, S.A.; Maslov, I.I.; Tovpinets, N.N.; Demchenko, V.A.; Mityai, I.S.; Antonovsky, A.G.; Zagorodnyaya, Y.A.; et al. Irrigated Agriculture and Problems of Biodiversity Conservation Dzhankoy Region of the Autonomous Republic of Crimea; Institute of Zoology I. I. Shmalgauzen NAS of Ukraine: Kiev, Ukraine, 2005; 121p. (In Russian) [Google Scholar]
- Hossain, A.; Krupnik, T.J.; Timsina, J.; Mahboob, M.G.; Chaki, A.K.; Farooq, M.; Bhatt, R.; Fahad, S.; Hasanuzzaman, M. Agricultural land degradation: Processes and problems undermining future food security. In Environment, Climate, Plant and Vegetation Growth; Fahad, S., Hasanuzzaman, M., Alam, M., Ullah, H., Saeed, M., Khan, I.A., Adnan, M., Eds.; Springer International Publishing: Cham, Switzerland, 2020; pp. 17–61. [Google Scholar]
- Aspe, C.; Gilles, A.; Jacqué, M. Irrigation canals as tools for climate change adaptation and fish biodiversity management in Southern France. Reg. Environ. Change 2016, 16, 1975–1984. [Google Scholar] [CrossRef] [Green Version]
- Hill, M.J.; Chadd, R.P.; Morris, N.; Swaine, J.D.; Wood, P.J. Aquatic macroinvertebrate biodiversity associated with artificial agricultural drainage ditches. Hydrobiologia 2016, 776, 249–260. [Google Scholar] [CrossRef] [Green Version]
- Fleming, W.M.; Rivera, J.A.; Miller, A.; Piccarello, M. Ecosystem services of traditional irrigation systems in northern New Mexico, USA. Int. J. Biodivers. Sci. Ecosyst. Serv. Manag. 2014, 10, 343–350. [Google Scholar] [CrossRef] [Green Version]
- Dorotovičová, C. Man-made canals as a hotspot of aquatic macrophyte biodiversity in Slovakia. Limnologica 2013, 43, 277–287. [Google Scholar] [CrossRef]
- Ohira, M.; Fukuda, S. Flow regime shapes seasonal patterns of fish species richness and abundance in main and branch channels of a rice irrigation system. Paddy Water Environ. 2018, 16, 783–793. [Google Scholar] [CrossRef]
- Richart, A.I.; Jiménez, M.N.; Ondoño, E.F.; Navarro, F.B. Plant diversity in traditional irrigation channels and its relations with soil properties. Ecosistemas 2019, 28, 81–91. [Google Scholar] [CrossRef] [Green Version]
- Shivashankar, P.; Venkataramana, G.V. Zooplankton diversity of Chikkadevarayana canal in relation to physico-chemical characteristics. J. Environ. Biol. 2013, 34, 819. [Google Scholar]
- Ismail, A.H.; Zaidin, S.A. A comparative study of zooplankton diversity and abundance from three different types of water body. In Proceedings of the 2nd International Conference on Agriculture, Environment and Biological Sciences (ICAEBS’15), Bali, Indonesia, 16–17 August 2015. [Google Scholar]
- Khalifa, N.; Abd El-Hady, H.H.; Zaher, S.S. The influence of Cyanophyceae bloom on zooplankton populations in El-Bahr El-Pharaony Canal, Egypt. Ecol. Environ. Cons. 2016, 22, 1629–1639. [Google Scholar]
- Khalifa, N.; Bendary, R.E. Composition and biodiversity of zooplankton and macrobenthic populations in El-Rayah El-Menoufy, Egypt. Int. J. Appl. Environ. Sci. 2016, 11, 683–700. [Google Scholar]
- Ashour, M.; Abo-Taleb, H.; Abou-Mahmoud, M.; El-Feky, M.M. Effect of the integration between plankton natural productivity and environmental assessment of irrigation water, El-Mahmoudia Canal, on aquaculture potential of Oreochromis niloticus. Turk. J. Fish. Aquat. Sci. 2018, 18, 1163–1175. [Google Scholar] [CrossRef]
- Buhungu, S.; Donou, M.; Ntakimazi, G.; Bonou, C.A.; Montchowui, E. Identification of characteristic zooplankton species in the Kinyankonge River basin, Burundi. Int. J. Aquat. Biol. 2019, 7, 71–84. [Google Scholar]
- Fishar, R.M.; Mahmoud, N.H.; El-Feqy, F.A.; Gaber, K.M.G. Community Composition of Zooplankton in El-Rayah El-Behery, Egypt. Egypt. J. Aquat. Biol. Fish. 2019, 23, 135–150. [Google Scholar] [CrossRef] [Green Version]
- Zhovtonog, O.; Hoffmann, M.; Polishchuk, V.; Dubel, A. New planning technique to master the future of water on local and regional level in Ukraine. J. Water Clim. Chang. 2011, 2, 189–200. [Google Scholar] [CrossRef]
- Shadrin, N.V.; Anufriieva, E.V.; Kipriyanova, L.M.; Kolesnikova, E.A.; Latushkin, A.A.; Romanov, R.E.; Sergeeva, N.G. The political decision caused the drastic ecosystem shift of the Sivash Bay (the Sea of Azov). Quat. Int. 2018, 475, 4–10. [Google Scholar] [CrossRef]
- Shadrin, N.V.; Simonov, V.G.; Anufriieva, E.V.; Popovichev, V.N.; Sirotina, N.O. Anthropogenic Transformation of Kyzyl-Yar Lake in Crimea: Multiyear Research Findings. Arid Ecosyst. 2018, 8, 299–306. [Google Scholar] [CrossRef]
- Mirzoeva, N.; Shadrin, N.; Arkhipova, S.; Miroshnichenko, O.; Kravchenko, N.; Anufriieva, E. Does Salinity Affect the Distribution of the Artificial Radionuclides 90Sr and 137Cs in Water of the Saline Lakes? A Case of the Crimean Peninsula. Water 2020, 12, 349. [Google Scholar] [CrossRef] [Green Version]
- Anufriieva, E.; Kolesnikova, E.; Revkova, T.; Latushkin, A.; Shadrin, N. Human-Induced Sharp Salinity Changes in the World’s Largest Hypersaline Lagoon Bay Sivash (Crimea) and Their Effects on the Ecosystem. Water 2022, 14, 403. [Google Scholar] [CrossRef]
- Stadnichenko, A.P. Review of the fauna of freshwater molluscs of the Crimea. Vestn. Zool. 1979, 1, 14. (In Russian) [Google Scholar]
- Kharchenko, T.G. Macrobenthos communities of the North Crimean Canal. Hydrobiol. J. 1983, 19, 37–44. (In Russian) [Google Scholar]
- Grinchenko, A.B.; Dulitsky, A.I. Distribution and ecology of the muskrat in the northern Crimea. Vestn. Zool. 1984, 3, 69–71. (In Russian) [Google Scholar]
- Zotiev, A.N.; Mokhnoshchekov, I.G. The Canal of Abundance: North-Crimean Canal; Krymizdat: Simferopol, Crimea, 1964; 40p. (In Russian) [Google Scholar]
- Rylov, V.M. Fauna of the USSR. Crustaceans. Cyclopoida of Fresh Waters; Izdatel’stvo Akademii Nauk SSSR: Moscow, Russia, 1948; Volume 3, Issue 3; 321p. (In Russian) [Google Scholar]
- Manuilova, E.F. Cladocera of the USSR. Guide-Books on the Fauna of the USSR; Nauka: Leningrad, Russia, 1964; Volume 88, 379p. (In Russian) [Google Scholar]
- Kutikova, L.A. Rotifers of the Fauna of the USSR; Nauka: Leningrad, Russia, 1970; 744p. (In Russian) [Google Scholar]
- Korovchinsky, N.M.; Kotov, A.A.; Boykova, O.S.; Smirnov, N.N. Cladocerans (Crustacea: Cladocera) of Northern Eurasia; Association of Scientific Publications KMK: Moscow, Russia, 2021; Volume 1, 481p. (In Russian) [Google Scholar]
- Korovchinsky, N.M.; Kotov, A.A.; Sinev, A.Y.; Neretina, A.N.; Gharibyan, P.G. Cladocerans (Crustacea: Cladocera) of Northern Eurasia; Association of Scientific Publications KMK: Moscow, Russia, 2021; Volume 2, 544p. (In Russian) [Google Scholar]
- Alekseev, V.; Tsalolikhin, S. Key to Zooplankton and Zoobenthos in Fresh Waters of European Russia. Volume 1. Zooplankton; Association of Scientific Publications KMK: Moscow, Russia, 2010; 495p. (In Russian) [Google Scholar]
- Alimov, A.F.; Bogatov, V.V.; Golubkov, S.M. Production Hydrobiology; Nauka: Saint Petersburg, Russia, 2013; 343p. (In Russian) [Google Scholar]
- Sokal, R.R.; Rohlf, F.J. Biometry: The Principles and Practice in Biological Research; Freeman: New York, NY, USA, 1995; 880p. [Google Scholar]
- Müller, P.H.; Neuman, P.; Storm, R. Tafeln der Mathematischen Statistik; VEB Fachbuchverlag: Leipzig, Germany, 2007; 275p. [Google Scholar]
- Sitran, R.; Bergamasco, A.; Decembrini, F.; Guglielmo, L. Temporal succession of tintinnids in the northern Ionian Sea, Central Mediterranean. J. Plankton Res. 2007, 29, 495–508. [Google Scholar] [CrossRef]
- Anufriieva, E.; Hołynska, M.; Shadrin, N. Current invasions of Asian Cyclopid species (Copepoda: Cyclopidae) in Crimea, with taxonomical and zoogeographical remarks on the hypersaline and freshwater fauna. Ann. Zool. 2014, 64, 109–130. [Google Scholar] [CrossRef]
- El-Shabrawy, G.A.; Anufriieva, E.; Shadrin, N. Tintinnina (Ciliophora) and Foraminifera in plankton of hypersaline Lagoon Bardawil (Egypt): Spatial and temporal variability. Turk. J. Zool. 2018, 42, 218–229. [Google Scholar] [CrossRef] [Green Version]
- Zhao, K.; Wang, L.; Riseng, C.; Wehrly, K.; Pan, Y.; Song, K.; Da, L.; Pang, W.; You, Q.; Tian, H.; et al. Factors determining zooplankton assemblage difference among a man-made lake, connecting canals, and the water-origin river. Ecol. Indic. 2018, 84, 488–496. [Google Scholar] [CrossRef]
- Gusynskaya, S.L. Pelagic zooplankton. In Bespozvonochnyye i Ryby Dnepra i Yego Vodokhranilishch; Shcherbak, G.I., Ed.; Naukova Dumka: Kiev, Ukraine, 1989; pp. 21–44. (In Russian) [Google Scholar]
- Dombrovsky, K.O. Zooplankton of the floodwater uppers of the Kakhovka reservoir. Visnyk Zaporiz’koho Derzhavnoho Univ. Biol. Nauk. 1999, 1, 173–178. (In Ukrainian) [Google Scholar]
- Pashkova, O.V. Near-bottom zooplankton of the littoral zone of the Dnieper reservoirs and its seasonal dynamics. Hydrobiol. J. 2005, 41, 42–56. [Google Scholar] [CrossRef]
- Mykolaichuk, T.V. Zooplankton of the Zaporizhzhya Reservoir. Biosyst. Divers. 2006, 2, 23. (In Ukrainian) [Google Scholar]
- Trokhymets, V.M. Littoral zooplankton of the middle part of the Kaniv Reservoir. Rybohospodars’ka Nauka Ukrayiny 2012, 2, 14–23. (In Ukrainian) [Google Scholar]
- Trokhymets, V.M. The current state of the littoral zooplankton of the lower part of the Kyiv Reservoir. Nauk. Zap. Ternopil’s’koho Derzhavnoho Peduniversytetu. Seriya Biol. 2015, 3, 64. (In Ukrainian) [Google Scholar]
- Kruzhilina, S.V.; Kotovs’ka, G.O. Forage base and potential bioproductive possibilities of the Dnieper river reservoirs. Bull. Zaporizhzhia Natl. Univ. Biol. Sci. 2013, 3, 22–31. (In Ukrainian) [Google Scholar]
- Dombrovsky, K.O.; Bychok, S.V. Zooplanktonic community structure of the river area of the Kakhovka reservoir littoral. Bull. Zaporizhzhia Natl. Univ. Biol. Sci. 2016, 1, 127–138. (In Ukrainian) [Google Scholar]
- Yakovenko, V.A.; Zaychenko, E.Y. Study of vertical distribution of zooplankton in Zaporozhskoye Reservoir aiming to calculate bighead carp stocking: Scientific communication on the materials of the reports of All-Russian conference with international participate, devoted to the 110th anniversary of Dr. Viktor Sergeevich Ivlev (1907–1964) and to the 100th anniversary of Dr. Irina Viktorovna Ivleva (1918–1992) “Prospects and directions of aquatic ecology development”(11–15 October, 2017, Sevastopol). Mar. Biol. J. 2018, 3, 57–69. (In Russian) [Google Scholar]
- Kovaleva, O.V. Taxonomic and trophic diversity of zooplankton, water quality of some small rivers of the Dnieper basin. Viesnik Maz. Dziaržaŭnaha Piedahahičnaha Ŭniv. Imia IP Šamiakina 2014, 42, 31–37. (In Russian) [Google Scholar]
- Pashkova, O.V. Development and distribution of zooplankton in anthropogenically modified and natural secondary reservoirs of the upper part of the Kanevsky Reservoir and R. Dnipro. Hydrobiol. J. 2016, 52, 40–55. (In Russian) [Google Scholar] [CrossRef]
- Yakovenko, V.A.; Fedonenko, E.V.; Tushnitskaya, N.I. Assessment of the state of zooplankton and zoobenthos of the Mokraya Sura River. Ribogospodars’ka Nauka Ukraïni 2017, 4, 19–32. (In Russian) [Google Scholar] [CrossRef] [Green Version]
- Ulomsky, S.N. Plankton of inland waters of the Crimea and its biomass. Tr. Karadag. Biol. Stantsii AN USSR 1955, 13, 131–162. (In Russian) [Google Scholar]
- Temirova, S.I. Freshwater ecosystems. In Biological and Landscape Diversity of Crimea: Problems and Prospects; SONAT: Simferopol, Crimea, 1999; pp. 105–107. (In Russian) [Google Scholar]
- Samchyshina, L.V. The state of the fauna of Calanoida (Crustacea, Copepoda) of internal reservoirs of the Crimea. Nauk. Zap. Ternopil’s’koho Derzhavnoho Peduniv. Seriya Biol. Spets. Issue Hidroekol. 2001, 3, 89–90. (In Russian) [Google Scholar]
- Monchenko, V.I. Free-Living Cyclopoid Copepods of the Ponto-Caspian Basin; Naukova Dumka: Kiev, Ukraine, 2003; 350p. (In Russian) [Google Scholar]
- Garibian, P.G.; Karabanov, D.P.; Neretina, A.N.; Taylor, D.J.; Kotov, A.A. Bosminopsis deitersi (Crustacea: Cladocera) as an ancient species group: A revision. PeerJ 2021, 9, e11310. [Google Scholar] [CrossRef] [PubMed]
- Linko, A. Bosminopsis (J. Richard) im europaischen Russland. Zool. Anz. 1901, 24, 345–347. [Google Scholar]
- Duggan, I.C.; Payne, R.J. Revisiting Elton’s copepods: Lake construction has altered the distribution and composition of calanoid copepods in the British Isles. Aquat. Invasions 2017, 12, 159–166. [Google Scholar] [CrossRef]
- Walter, T.C.; Boxshall, G.; World of Copepods Database. Cyclops Smirnovi. World Register of Marine Species. 2023. Available online: https://www.marinespecies.org/aphia.php?p=taxdetails&id=350600 (accessed on 3 May 2023).
- Anufriieva, E.; Shadrin, N. Current invasions of East Asian cyclopoids (Copepoda, Cyclopoida) in Europe: New records from eastern Ukraine. Turk. J. Zool. 2016, 40, 282–285. [Google Scholar] [CrossRef]
- Simons, J.H.; Bakker, C.; Schropp, M.H.; Jans, L.H.; Kok, F.R.; Grift, R.E. Man-made secondary channels along the River Rhine (The Netherlands). results of post-project monitoring. Regul. Rivers Res. Manag. 2001, 17, 473–491. [Google Scholar] [CrossRef]
- Havel, J.E.; Lee, C.E.; Vander Zanden, J.M. Do reservoirs facilitate invasions into landscapes? BioScience 2005, 55, 518–525. [Google Scholar] [CrossRef] [Green Version]
- Anufriieva, E.V.; Shadrin, N.V. Extreme hydrological events destabilize aquatic ecosystems and open doors for alien species. Quat. Int. 2018, 475, 11–15. [Google Scholar] [CrossRef]
Figure 1.
The scheme of the North Crimean Canal and sampling sites (1–8), whose coordinates and other information are given in Table 1.
Figure 1.
The scheme of the North Crimean Canal and sampling sites (1–8), whose coordinates and other information are given in Table 1.

Figure 2.
The North Crimean Canal in October 2014 (a,b)—the sites without water; (c,d)—the sites with the dropped water level.
Figure 2.
The North Crimean Canal in October 2014 (a,b)—the sites without water; (c,d)—the sites with the dropped water level.

Figure 3.
The North Crimean Canal in 2022 (a,b)—the 1st sampling site, 15 March 2022; (c)—the 5th sampling site, 30 May 2022; (d)—the 6th sampling site, 21 June 2022.
Figure 3.
The North Crimean Canal in 2022 (a,b)—the 1st sampling site, 15 March 2022; (c)—the 5th sampling site, 30 May 2022; (d)—the 6th sampling site, 21 June 2022.

Figure 4.
The North Crimean Canal (a)—high concentrations of phytoplankton; (b–d)—thickets of macrophytes.
Figure 4.
The North Crimean Canal (a)—high concentrations of phytoplankton; (b–d)—thickets of macrophytes.

Figure 5.
The North Crimean Canal (a,b)—harvesting macrophytes in front of a pumping station; (c,d)—mollusks in harvesting macrophytes.
Figure 5.
The North Crimean Canal (a,b)—harvesting macrophytes in front of a pumping station; (c,d)—mollusks in harvesting macrophytes.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Coordinates and general characteristics of sampling sites in the North Crimean Canal.
| Station Number | Date | Coordinates | T, °C | S, g L−1 | pH | Eh, mV |
|---|---|---|---|---|---|---|
| 1 | 15 March 2022 | 46°07′12.617″ N, 33°41′25.697″ E | 8.5 | 0.3 | 7.7 | 171 |
| 27 April 2022 | 46°07′12.617″ N, 33°41′25.697″ E | 14.5 | 0.3 | 7.1 | 210 | |
| 2 | 12 August 2022 | 45°57′26.003″ N, 33°49′18.280″ E | 29.6 | 0.4 | 8.6 | - |
| 3 | 6 October 2022 | 45°48′32.020″ N, 33°48′35.522″ E | 19.2 | 0.2 | 6.8 | 235 |
| 4 | 27 April 2022 | 45°42′38.801″ N, 34°26′21.799″ E | 16.9 | 0.3 | 7.0 | 154 |
| 5 | 30 May 2022 | 45°22′58.501″ N, 35°56′46.498″ E | 25.0 | 0.3 | 6.9 | – |
| 23 June 2022 | 45°22′58.501″ N, 35°56′46.498″ E | 20.5 | 0.3 | 7.3 | 138 | |
| 6 | 30 May 2022 | 45°22′58.489″ N, 35°56′46.497″ E | 24.0 | 0.3 | 7.1 | 140 |
| 7 | 21 June 2022 | 45°17′13.399″ N, 34°56′46.497″ E | 29.5 | 0.3 | 7.3 | 145 |
| 8 | 21 June 2022 | 45°17′13.399″ N, 34°56′46.495″ E | 30.0 | 0.3 | 7.7 | 151 |
Note: T—temperature; S—salinity.
Table 2.
Species composition of zooplankton in the North Crimean Canal in 2022.
| Taxon | Frequency of Occurrence, % | The Period of Presence | Abundance, Ind. m−3 | |
|---|---|---|---|---|
| Min | Max | |||
| Rotifera | 80 | March–June | ||
| Brachionus angularis Gosse, 1851 | 20 | June | 60 | 1840 |
| Brachionus calyciflorus Pallas, 1766 | 30 | March, May | 7 | 160 |
| Euchlanis dilatata Ehrenberg, 1832 | 30 | April, June | 10 | 320 |
| Filinia longiseta (Ehrenberg, 1834) | 10 | May | 10 | – |
| Keratella cochlearis (Gosse, 1851) | 30 | April, June | 20 | 240 |
| Keratella quadrata (Müller, 1786) | 10 | April | 240 | – |
| Ploesoma hudsoni (Imhof, 1891) | 10 | June | 40 | – |
| Polyarthra dolichoptera Idelson, 1925 | 20 | April | 160 | 240 |
| Pompholyx complanata Gosse, 1851 | 20 | March, June | 84 | 560 |
| Synchaeta grandis Zacharias, 1893 | 10 | March | 14 | – |
| Synchaeta stylata Wierzejski, 1893 | 20 | March, June | 56 | – |
| Arthropoda | 100 | March–October | ||
| Cladocera | 80 | March–October | ||
| Bosmina longirostris (O.F.Müller, 1776) | 60 | April–June | 10 | 2440 |
| Bosminopsis zernowi Linko, 1901 | 10 | May | 10 | – |
| Ceriodaphnia pulchella Sars, 1862 | 10 | April | 120 | – |
| Chydorus sphaericus (O.F.Müller, 1776) | 70 | March–June | 20 | 72,000 |
| Coronatella rectangula (Sars, 1862) | 40 | April, June | 10 | 160 |
| Daphnia cucullata Sars, 1862 | 10 | April | 240 | – |
| Leptodora kindtii (Focke, 1844) | 10 | June | 10 | – |
| Moina macrocopa (Straus, 1820) | 10 | April | 160 | – |
| Rhynchotalona rostrata (Koch, 1841) | 20 | April, June | 20 | 160 |
| Scapholeberis mucronata (O.F.Müller, 1776) | 30 | April–June | 10 | 240 |
| Simocephalus vetulus (O.F.Müller, 1776) | 10 | April | 680 | – |
| Copepoda | 100 | March–October | Adults only | |
| Calanoida | 60 | March–October | ||
| Eurytemora affinis (Poppe, 1880) | 50 | March, June, October | 20 | 1280 |
| Eurytemora lacustris (Poppe, 1887) | 20 | April | 1760 | 1920 |
| Eurytemora velox (Lilljeborg, 1853) | 10 | August | 72 | – |
| Heterocope appendiculata Sars G.O., 1863 | 10 | April | 320 | – |
| Heterocope caspia Sars G.O., 1897 | 10 | April | 240 | – |
| Cyclopoida | 90 | April–June, October | ||
| Acanthocyclops vernalis (Fischer, 1853) | 80 | April–October | 10 | 840 |
| Cyclops furcifer Claus, 1857 | 10 | April | 60 | – |
| Cyclops strenuus Sars G.O., 1903 | 10 | April | 120 | – |
| Cyclops smirnovi Rylov, 1948 | 10 | April | 60 | – |
| Diacycops bisetosus (Rehberg, 1880) | 10 | April | 120 | – |
| Eucyclops serrulatus (Fischer, 1851) | 30 | April, June | 10 | 2240 |
| Mesocyclops leuckarti (Claus, 1857) | 20 | August, October | 36 | 64 |
| Thermocyclops oithonoides (Sars G.O., 1863) | 10 | April | 240 | – |
| Harpacticoida | 60 | March–August | ||
| Halectinosoma abrau (Krichagin, 1877) | 10 | August | 2488 | – |
| Nitocra typica Boeck, 1865 | 50 | April, May, June | 10 | 160 |
| Nitocra sp. | 10 | March | 14 | – |
| Ostracoda | 10 | March | 7 | – |
| Gammaridae (juvenils) | 10 | March | 42 | – |
| Mollusca | 10 | April | ||
| Dreissena (veligers) | 10 | April | 320 | – |
Table 3.
General characteristics of zooplankton in the North Crimean Canal during 2022.
| Station Number | Date | Rotifera | Cladocera | Copepoda | Total Zooplankton | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Number of Species | N, Ind. m−3 | B, mg m−3 | Number of Species | N, Ind. m−3 | B, mg m−3 | Number of Species | N, Ind. m−3 | B, mg m−3 | Number of Species | N, Ind. m−3 | B, mg m−3 | ||
| 1 | 15 March 2022 | 4 | 161 | 0.20 | 1 | 28 | 0.35 | 1 | 14 | 0.20 | 6 | 203 | 0.75 |
| 27 April 2022 | 3 | 640 | 1.44 | 6 | 10,600 | 184.45 | 6 | 8320 | 356.66 | 15 | 19,560 | 542.55 | |
| 2 | 12 August 2022 | 0 | 0 | 0 | 1 | 36 | 1.93 | 4 | 3066 | 26.78 | 5 | 3102 | 28.71 |
| 3 | 6 October 2022 | 0 | 0 | 0 | 3 | 288 | 4.06 | 3 | 4288 | 56.19 | 6 | 3102 | 60.25 |
| 4 | 27 April 2022 | 3 | 720 | 0.36 | 6 | 72,990 | 686.13 | 10 | 17,760 | 370.42 | 19 | 91,470 | 1056.91 |
| 5 | 30 May 2022 | 1 | 20 | 0.05 | 2 | 220 | 1.39 | 2 | 60 | 1.37 | 5 | 300 | 2.81 |
| 23 June 2022 | 1 | 10 | 0.02 | 4 | 50 | 0.56 | 3 | 110 | 1.27 | 7 | 170 | 1.85 | |
| 6 | 30 May 2022 | 2 | 30 | 0.06 | 4 | 70 | 0.45 | 2 | 80 | 1.54 | 8 | 180 | 2.05 |
| 7 | 21 June 2022 | 4 | 2960 | 2.26 | 4 | 1900 | 8.16 | 2 | 160 | 3.29 | 10 | 5020 | 13.71 |
| 8 | 21 June 2022 | 3 | 120 | 0.14 | 1 | 2440 | 7.61 | 2 | 170 | 2.19 | 6 | 2730 | 9.94 |
Note: N—abundance; B—biomass.
Table 4.
Contribution of Rotifera, Cladocera, and Copepoda to the total abundance of zooplankton.
| Station Number | Date | Rotifera, % | Cladocera, % | Copepoda, % |
|---|---|---|---|---|
| 1 | 15 March 2022 | 79 | 14 | 7 |
| 27 April 2022 | 3 | 54 | 43 | |
| 2 | 12 August 2022 | 0 | 1 | 99 |
| 3 | 6 October 2022 | 0 | 6 | 94 |
| 4 | 27 April 2022 | 1 | 80 | 19 |
| 5 | 30 May 2022 | 7 | 73 | 20 |
| 23 June 2022 | 6 | 29 | 65 | |
| 6 | 30 May 2022 | 17 | 39 | 44 |
| 7 | 21 June 2022 | 59 | 38 | 3 |
| 8 | 21 June 2022 | 4 | 89 | 6 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Shadrin, N.; Yakovenko, V.; Moseychenko, I.; Anufriieva, E. Zooplankton in the Technogenic Aquatic Ecosystem of the North Crimean Canal, the Longest in Europe. Water 2023, 15, 2327. https://doi.org/10.3390/w15132327
AMA Style
Shadrin N, Yakovenko V, Moseychenko I, Anufriieva E. Zooplankton in the Technogenic Aquatic Ecosystem of the North Crimean Canal, the Longest in Europe. Water. 2023; 15(13):2327. https://doi.org/10.3390/w15132327
Chicago/Turabian StyleShadrin, Nickolai, Vladimir Yakovenko, Igor Moseychenko, and Elena Anufriieva. 2023. "Zooplankton in the Technogenic Aquatic Ecosystem of the North Crimean Canal, the Longest in Europe" Water 15, no. 13: 2327. https://doi.org/10.3390/w15132327
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.