
Assessing the Movements, Habitat Use, and Site Fidelity of the Giant Freshwater Whipray (Urogymnus polylepis) with Acoustic Telemetry in the Maeklong River, Thailand
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
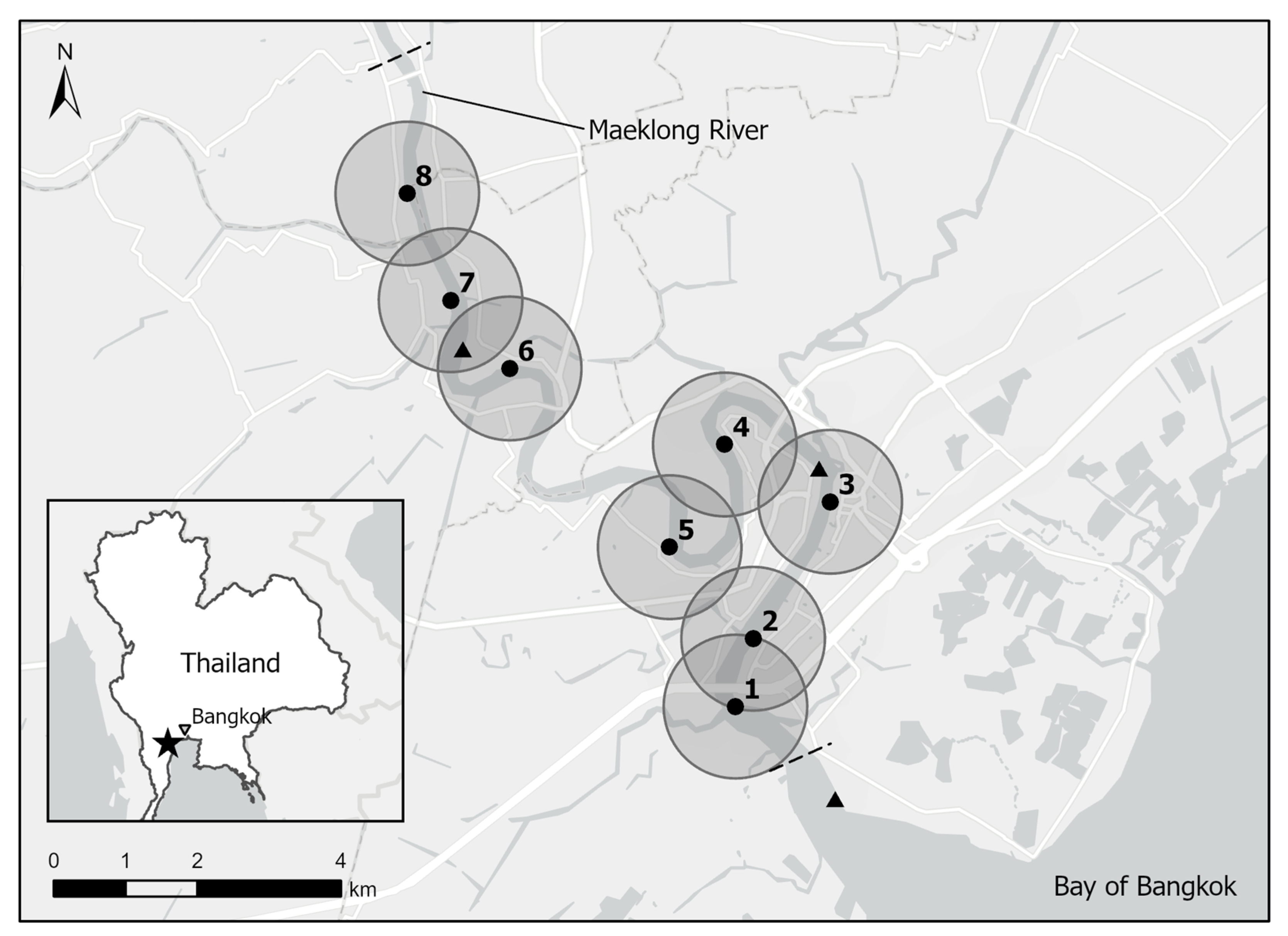
2.1. Study Site and Receiver Deployment
2.2. Animal Capture, Work-Up, and Tagging
2.3. Data Analysis
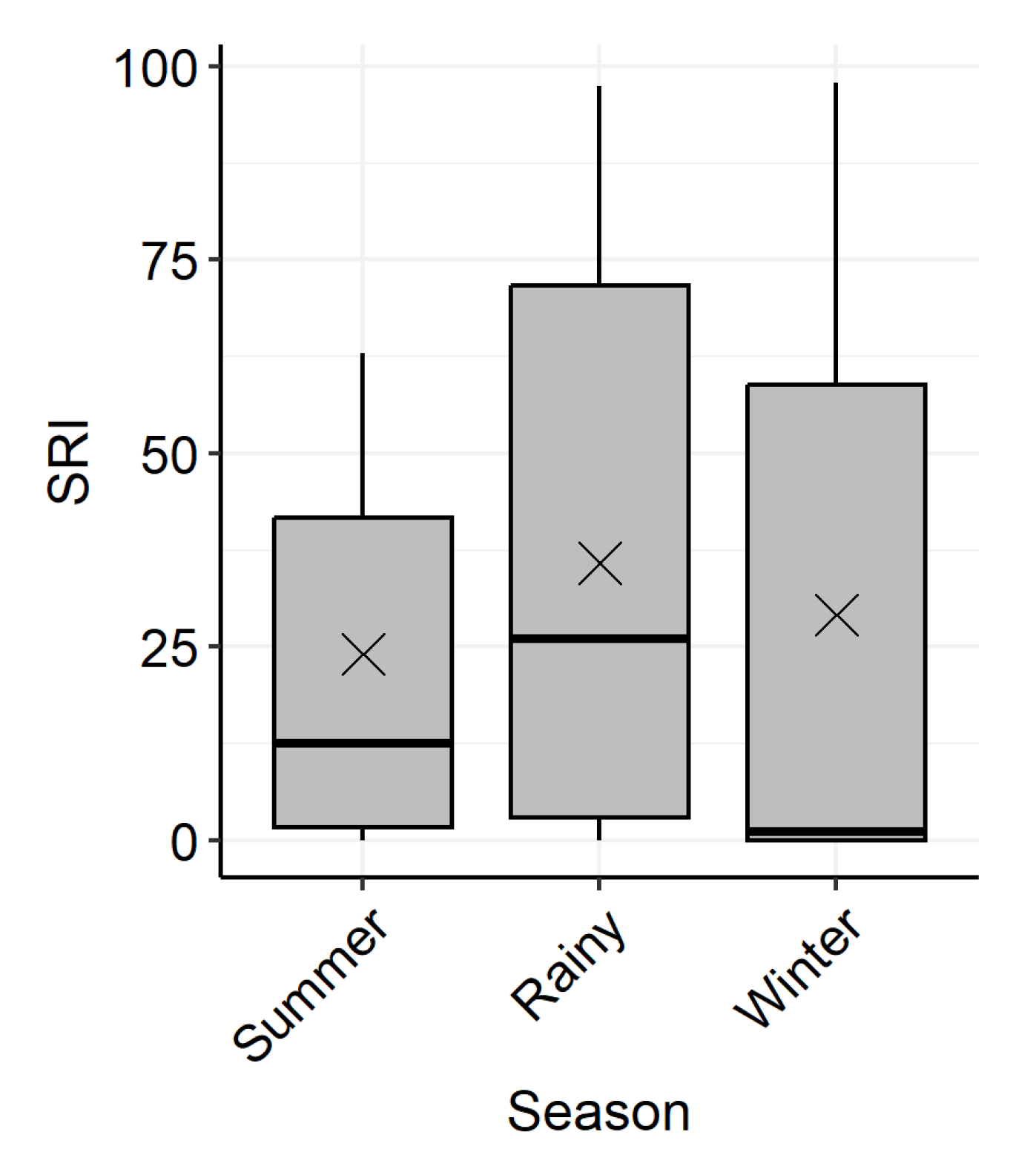
2.3.1. Annual and Seasonal Site Fidelity
2.3.2. Spatial Variation in Site Use
2.3.3. Diel Activity Patterns
2.3.4. Relationship between Temperature and Site Fidelity
3. Results
3.1. Whipray Morphology and Physiology
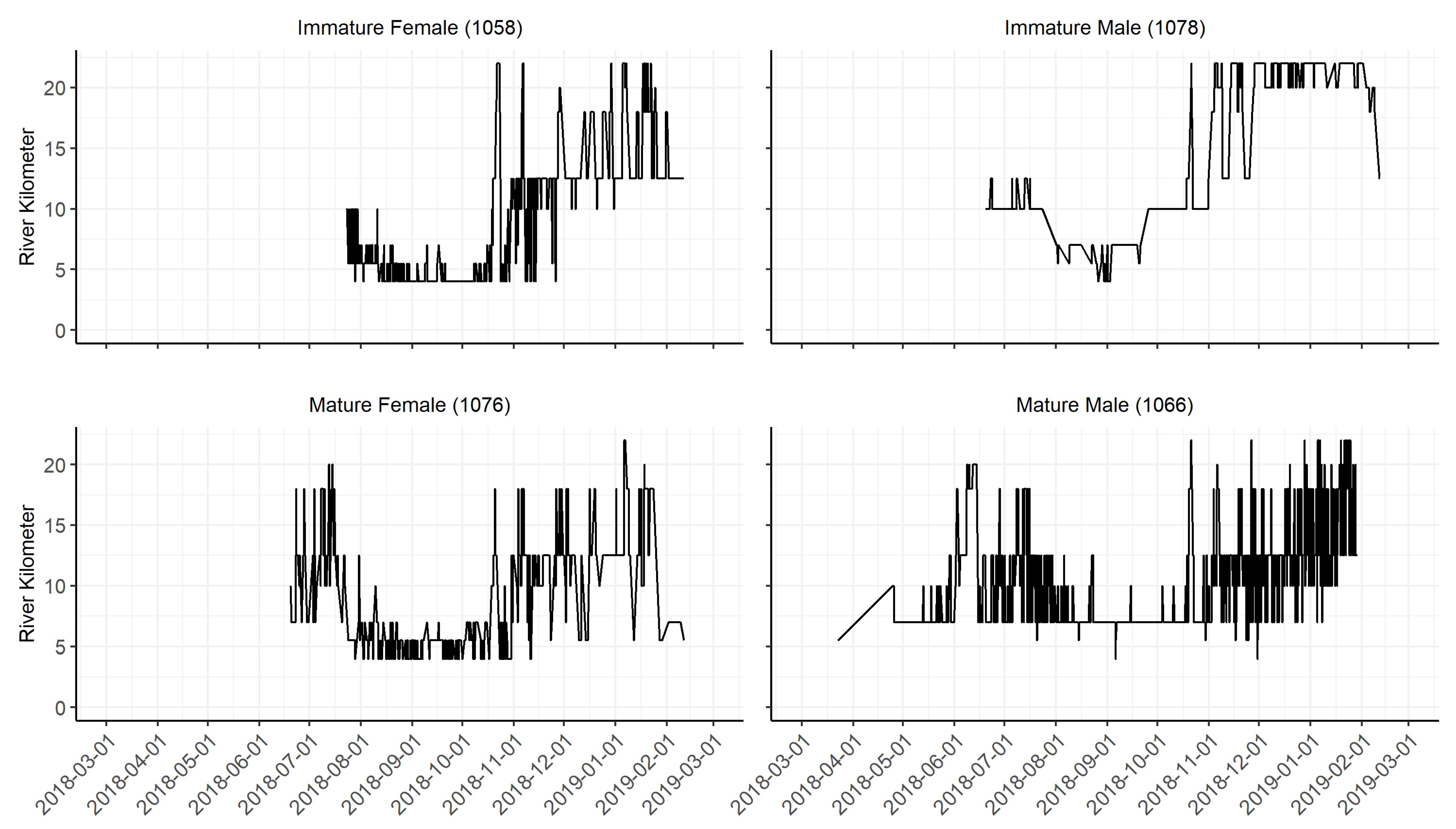
3.2. Spatio-Temporal Patterns of Site Use
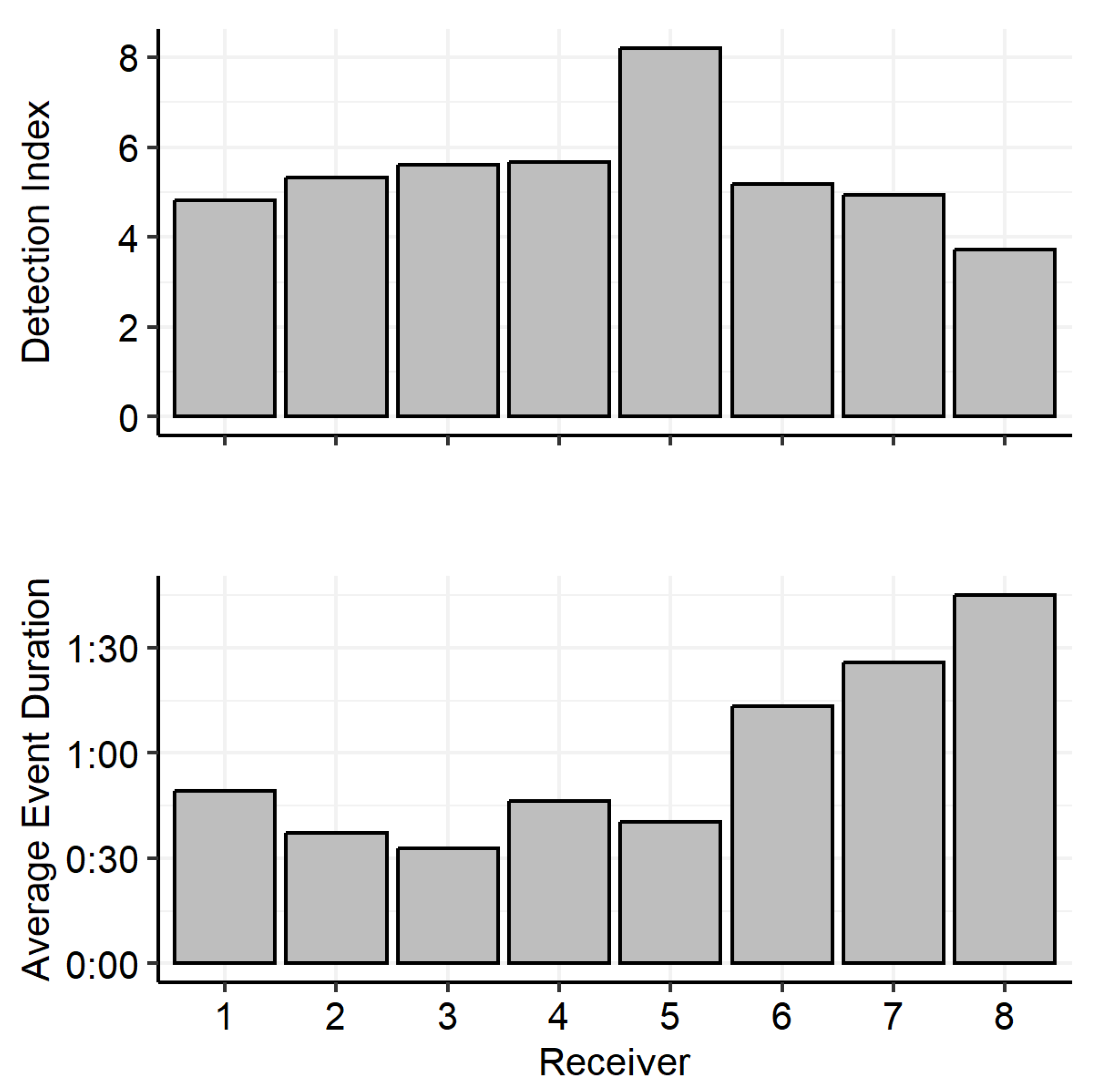
3.3. Diel Activity Patterns
3.4. Environmental Correlates of Site Use
4. Discussion
4.1. Site Fidelity
4.2. Spatio-Temporal Patterns of Site Use
4.3. Life-History Observations
4.4. Environmental Influences on Site Fidelity
4.5. Research Technique
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Campbell, T.; Onboundisane, S.; Kong, H.; Hogan, Z.S. A review of the conservation status and ecology of the giant freshwater whipray (Urogymnus polylepis) across its known distribution. Water, 2023; in press. [Google Scholar]
- Grant, I.; Rigby, C.L.; Bin Ali, A.; Fahmi; Haque, A.B.; Hasan, V.; Sayer, C. Urogymnus polylepis. In The IUCN Red List of Threatened Species, 25 January 2021 ed.; International Union for Conservation of Nature and Natural Resources: Gland, Switzerland, 2021; p. e.T195320A104294071. [Google Scholar]
- Compagno, L.J.V.; Cook, S.F. Urogymnus polylepis (Thailand subpopulation). In The IUCN Red List of Threatened Species, 30 June 2000 ed.; International Union for Conservation of Nature and Natural Resources: Gland, Switzerland, 2000; p. e.T39408A10228071. [Google Scholar]
- Monkolprasit, S.; Roberts, T.R. Himantura chaophraya, a new giant freshwater stingray from Thailand. Jpn. J. Ichthyol. 1990, 37, 203–208. [Google Scholar]
- Millward, A. Record-breaking ray confirmed as world’s largest freshwater fish. In Guinness World Records Limited 2023; Guinness World Records: London, UK, 2022. [Google Scholar]
- Shaw, A.K. Drivers of animal migration and implications in changing environments. Evol. Ecol. 2016, 30, 991–1007. [Google Scholar] [CrossRef]
- Schlaff, A.M.; Heupel, M.R.; Simpfendorfer, C.A. Influence of environmental factors on shark and ray movement, behaviour and habitat use: A review. Rev. Fish Biol. Fish. 2014, 24, 1089–1103. [Google Scholar] [CrossRef]
- Heupel, M.R.; Webber, D.M. Trends in acoustic tracking: Where are the fish going and how will we follow them? In Proceedings of the American Fisheries Society Symposium 76, Auckland, New Zealand, 24–28 February 2008; American Fisheries Society Symposium: Bethesda, MD, USA, 2012; pp. 219–231. [Google Scholar]
- Campbell, H.A.; Hewitt, M.; Watts, M.E.; Peverell, S.; Franklin, C.E. Short-and long-term movement patterns in the freshwater whipray (Himantura dalyensis) determined by the signal processing of passive acoustic telemetry data. Mar. Freshw. Res. 2012, 63, 341–350. [Google Scholar] [CrossRef]
- Ramsden, S.; Cotton, C.F.; Curran, M.C. Using acoustic telemetry to assess patterns in the seasonal residency of the Atlantic stingray Dasyatis Sabina. Environ. Biol. Fishes 2017, 100, 89–98. [Google Scholar] [CrossRef]
- Vaudo, J.J.; Heithaus, M.R. Diel and seasonal variation in the use of a nearshore sandflat by a ray community in a near pristine system. Mar. Freshw. Res. 2012, 63, 1077–1084. [Google Scholar] [CrossRef]
- Davy, L.E.; Simpfendorfer, C.A.; Heupel, M.R. Movement patterns and habitat use of juvenile mangrove whiprays (Himantura granulata). Mar. Freshw. Res. 2015, 66, 481–492. [Google Scholar] [CrossRef]
- Corcoran, M.J.; Wetherbee, B.M.; Shivji, M.S.; Potenski, M.D.; Chapman, D.D.; Harvey, G.M. Supplemental feeding for ecotourism reverses diel activity and alters movement patterns and spatial distribution of the southern stingray, Dasyatis americana. PLoS ONE 2013, 8, e59235. [Google Scholar] [CrossRef] [Green Version]
- Then, A.Y.-H.; Lim, K.C.; Loh, K.-H. Updated distribution of the endangered freshwater stingray Urogymnus polylepis in Malaysia, with notes on biology and genetics. Raffles Bull. Zool. 2022, 70, 534–549. [Google Scholar]
- Sen, S.; Thomas, S.; Joe Kizhakudan, S.; Dash, G.; Pradhan, R.K.; Ghosh, S.; Das, M.; Dash, B.; Zacharia, P.U. New observations of the endangered giant freshwater whipray, Urogymnus polylepis, provide further evidence for its distribution and breeding in the north-east coast of India. J. Fish Biol. 2022, 101, 1611–1616. [Google Scholar] [CrossRef]
- Huveneers, C.; Simpfendorfer, C.A.; Kim, S.; Semmens, J.M.; Hobday, A.J.; Pederson, H.; Stieglitz, T.; Vallee, R.; Webber, D.; Heupel, M.R. The influence of environmental parameters on the performance and detection range of acoustic receivers. Methods Ecol. Evol. 2016, 7, 825–835. [Google Scholar] [CrossRef]
- Loher, T.; Webster, R.A.; Carlile, D. A test of the detection range of acoustic transmitters and receivers deployed in deep waters of Southeast Alaska, USA. Anim. Biotelem. 2017, 5, 27. [Google Scholar] [CrossRef] [Green Version]
- Clements, S.; Jepsen, D.; Karnowski, M.; Schreck, C.B. Optimization of an acoustic telemetry array for detecting transmitter-implanted fish. North Am. J. Fish. Manag. 2005, 25, 429–436. [Google Scholar] [CrossRef]
- Kessel, S.T.; Hussey, N.E. Tonic immobility as an anaesthetic for elasmobranchs during surgical implantation procedures. Can. J. Fish. Aquat. Sci. 2015, 72, 1287–1291. [Google Scholar] [CrossRef]
- Chatchavalvanich, K.; Thongpan, A.; Nakai, M. Structure of the testis and genital duct of freshwater stingray, Himantura signifer (Elasmobranchii: Myliobatiformes: Dasyatidae). Ichthyol. Res. 2005, 52, 123. [Google Scholar] [CrossRef]
- Daochai, C.; Keschumras, N.; Daochai, S.; Chansue, N. Using ultrasonography method for monitoring ovarian status in freshwater stingray (Potamotrygon sp.). In Proceedings of the Biomedical Engineering International Conference, Pattaya, Thailand, 25–27 November 2015; pp. 1–4. [Google Scholar]
- Daochai, C. Using Ultrasonography Method for Monitoring Testis Status in Freshwater Stingray (Potamotrygon motoro). Thai. J. Vet. Med. Suppl. 2016, 46, 330. [Google Scholar]
- Daochai, C.; Chansue, N.; Daochai, S. Application of ultrasound in determination gestation status in captive female freshwater stingray (Potamotrygon sp.). In Proceedings of the Biomedical Engineering International Conference, Singapore, 7–10 December 2016; pp. 1–4. [Google Scholar]
- Freeland, W.J.; Fry, K. Suitability of passive integrated transponder tags for marking live animals for trade. Wildl. Res. 1995, 22, 767–773. [Google Scholar] [CrossRef]
- Wagner, G.N.; Cooke, S.J. Methodological approaches and opinions of researchers involved in the surgical implantation of telemetry transmitters in fish. J. Aquat. Anim. Health 2005, 17, 160–169. [Google Scholar] [CrossRef]
- Bond, M.E.; Babcock, E.A.; Pikitch, E.K.; Abercrombie, D.L.; Lamb, N.F.; Chapman, D.D. Reef sharks exhibit site-fidelity and higher relative abundance in marine reserves on the Mesoamerican Barrier Reef. PLoS ONE 2012, 7, e32983. [Google Scholar] [CrossRef] [Green Version]
- Reubens, J.T.; Pasotti, F.; Degraer, S.; Vincx, M. Residency, site fidelity and habitat use of Atlantic cod (Gadus morhua) at an offshore wind farm using acoustic telemetry. Mar. Environ. Res. 2013, 90, 128–135. [Google Scholar] [CrossRef] [PubMed]
- Vianna, G.M.S.; Meekan, M.G.; Meeuwig, J.J.; Speed, C.W. Environmental influences on patterns of vertical movement and site fidelity of grey reef sharks (Carcharhinus amblyrhynchos) at aggregation sites. PLoS ONE 2013, 8, e60331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pollution Control Department. Water Quality Data. 2018. Available online: https://rwater.mnre.go.th/front/main/WaterQuality (accessed on 2 May 2023).
- Yeiser, B.G.; Heupel, M.R.; Simpfendorfer, C.A. Occurrence, home range and movement patterns of juvenile bull (Carcharhinus leucas) and lemon (Negaprion brevirostris) sharks within a Florida estuary. Mar. Freshw. Res. 2008, 59, 489–501. [Google Scholar] [CrossRef]
- Tilley, A.; López-Angarita, J.; Turner, J.R. Effects of scale and habitat distribution on the movement of the southern stingray Dasyatis americana on a Caribbean atoll. Mar. Ecol. Prog. Ser. 2013, 482, 169–179. [Google Scholar] [CrossRef] [Green Version]
- Vaudo, J.J.; Lowe, C.G. Movement patterns of the round stingray Urobatis halleri (Cooper) near a thermal outfall. J. Fish Biol. 2006, 68, 1756–1766. [Google Scholar] [CrossRef] [Green Version]
- Jirik, K.E.; Lowe, C.G. An elasmobranch maternity ward: Female round stingrays Urobatis halleri use warm, restored estuarine habitat during gestation. J. Fish Biol. 2012, 80, 1227–1245. [Google Scholar] [CrossRef]
- Whitney, N.M.; Pyle, R.L.; Holland, K.N.; Barcz, J.T. Movements, reproductive seasonality, and fisheries interactions in the whitetip reef shark (Triaenodon obesus) from community-contributed photographs. Environ. Biol. Fishes 2012, 93, 121–136. [Google Scholar] [CrossRef]
- Mull, C.G.; Lowe, C.G.; Young, K.A. Seasonal reproduction of female round stingrays (Urobatis halleri): Steroid hormone profiles and assessing reproductive state. Gen. Comp. Endocrinol. 2010, 166, 379–387. [Google Scholar] [CrossRef]
- Shea, O.O.; Van Leeuwen, T.E.; Brien, D.O.; Arrowsmith, L.; McCalman, R.; Griffiths, M.; Exton, D.A. Evidence and description of a nursery habitat for the recently reclassified stingray Styracura schmardae from The Bahamas. Mar. Ecol. Prog. Ser. 2021, 660, 141–151. [Google Scholar]
- Le Port, A.; Lavery, S.; Montgomery, J.C. Conservation of coastal stingrays: Seasonal abundance and population structure of the short-tailed stingray Dasyatis brevicaudata at a Marine Protected Area. ICES J. Mar. Sci. 2012, 69, 1427–1435. [Google Scholar] [CrossRef] [Green Version]
- Nordell, S.E. Observations of the mating behavior and dentition of the round stingray, Urolophus halleri. Environ. Biol. Fishes 1994, 39, 219–229. [Google Scholar] [CrossRef]
- Chaikin, S.; Belmaker, J.; Barash, A. Coastal breeding aggregations of threatened stingrays and guitarfish in the Levant. Aquat. Conserv. Mar. Freshw. Ecosyst. 2020, 30, 1160–1171. [Google Scholar] [CrossRef]
- Pratt Jr, H.L.; Pratt, T.C.; Knotek, R.J.; Carrier, J.C.; Whitney, N.M. Long-term use of a shark breeding ground: Three decades of mating site fidelity in the nurse shark, Ginglymostoma cirratum. PLoS ONE 2022, 17, e0275323. [Google Scholar] [CrossRef] [PubMed]
- Pini-Fitzsimmons, J.; Knott, N.A.; Brown, C. Effects of food provisioning on site use in the short-tail stingray Bathytoshia brevicaudata. Mar. Ecol. Prog. Ser. 2018, 600, 99–110. [Google Scholar] [CrossRef]
- Lyon, B.J.; Dwyer, R.G.; Pillans, R.D.; Campbell, H.A.; Franklin, C.E. Distribution, seasonal movements and habitat utilisation of an endangered shark, Glyphis glyphis, from northern Australia. Mar. Ecol. Prog. Ser. 2017, 573, 203–213. [Google Scholar] [CrossRef]
- Choo-ina, S.; Tharasawatpipata, C.; Kasemsawata, S.; Monprapussornb, S.; Utarasakula, T. Surface Water Quality for Sustainable Water Consumption and Utilization in Samut Songkram Province. In Proceedings of the 2nd Environment Asia International Conference on “Human Vulnerability and Global Environmental Change”, Chonburi, Thailand, 15–17 May 2013. [Google Scholar]
- Koning, A.A.; Perales, K.M.; Fluet-Chouinard, E.; McIntyre, P.B. A network of grassroots reserves protects tropical river fish diversity. Nature 2020, 588, 631–635. [Google Scholar] [CrossRef]
- Mattos, S.M.G.; Broadhurst, M.; Hazin, F.H.V.; Jonnes, D.M. Reproductive biology of the Caribbean sharpnose shark, Rhizoprionodon porosus, from northern Brazil. Mar. Freshw. Res. 2001, 52, 745–752. [Google Scholar] [CrossRef]
- Carlisle, A. Long-term trends in catch composition from elasmobranch derbies in Elkhorn Slough, California. Mar. Fish. Rev. 2007, 69, 25–45. [Google Scholar]
- Thorson, T.B.; Langhammer, J.K.; Oetinger, M.I. Reproduction and development of the South American freshwater stingrays, Potamotrygon circularis and P. motoro. Environ. Biol. Fishes 1983, 9, 3–24. [Google Scholar] [CrossRef]
- Heithaus, M.R. The biology of tiger sharks, Galeocerdo cuvier, in Shark Bay, Western Australia: Sex ratio, size distribution, diet, and seasonal changes in catch rates. Environ. Biol. Fishes 2001, 61, 25–36. [Google Scholar] [CrossRef]
- Cartamil, D.P.; Vaudo, J.J.; Lowe, C.G.; Wetherbee, B.M.; Holland, K.N. Diel movement patterns of the Hawaiian stingray, Dasyatis lata: Implications for ecological interactions between sympatric elasmobranch species. Mar. Biol. 2003, 142, 841–847. [Google Scholar] [CrossRef]
- Collins, A.B.; Heupel, M.R.; Motta, P.J. Residence and movement patterns of cownose rays Rhinoptera bonasus within a south-west Florida estuary. J. Fish Biol. 2007, 71, 1159–1178. [Google Scholar] [CrossRef]
- Brinton, C.P.; Curran, M.C. Tidal and diel movement patterns of the Atlantic stingray (Dasyatis sabina) along a stream-order gradient. Mar. Freshw. Res. 2017, 68, 1716–1725. [Google Scholar] [CrossRef]
- Schieber, N.L.; Collin, S.P.; Hart, N.S. Comparative retinal anatomy in four species of elasmobranch. J. Morphol. 2012, 273, 423–440. [Google Scholar] [CrossRef]
- Morrissey, J.F.; Gruber, S.H. Habitat selection by juvenile lemon sharks, Negaprion brevirostris. Environ. Biol. Fishes 1993, 38, 311–319. [Google Scholar] [CrossRef]
- Wallman, H.L.; Bennett, W.A. Effects of parturition and feeding on thermal preference of Atlantic stingray, Dasyatis sabina (Lesueur). Environ. Biol. Fishes 2006, 75, 259–267. [Google Scholar] [CrossRef]
- Fangue, N.A.; Bennett, W.A. Thermal tolerance responses of laboratory-acclimated and seasonally acclimatized Atlantic stingray, Dasyatis sabina. Copeia 2003, 2003, 315–325. [Google Scholar] [CrossRef]
- Carlisle, A.B.; Starr, R.M. Tidal movements of female leopard sharks (Triakis semifasciata) in Elkhorn Slough, California. Environ. Biol. Fishes 2010, 89, 31–45. [Google Scholar] [CrossRef]
- Grant, M.I.; Kyne, P.M.; Simpfendorfer, C.A.; White, W.T.; Chin, A. Categorising use patterns of non-marine environments by elasmobranchs and a review of their extinction risk. Rev. Fish Biol. Fish. 2019, 29, 689–710. [Google Scholar] [CrossRef]
- Mitamura, H.; Mitsunaga, Y.; Arai, N.; Viputhanumas, T. Movements of immature hatchery-reared Mekong giant catfish Pangasianodon gigas released in the Mekong River, measured using acoustic telemetry. Fish. Sci. 2008, 74, 1034–1039. [Google Scholar] [CrossRef]
- Hahn, L.; Martins, E.G.; Nunes, L.D.; da Câmara, L.F.; Machado, L.S.; Garrone-Neto, D. Biotelemetry reveals migratory behaviour of large catfish in the Xingu River, Eastern Amazon. Sci. Rep. 2019, 9, 8464. [Google Scholar] [CrossRef] [Green Version]
- Naughton, G.P.; Hogan, Z.S.; Campbell, T.; Graf, P.J.; Farwell, C.; Sukumasavin, N. Acoustic Telemetry Monitors Movements of Wild Adult Catfishes in the Mekong River, Thailand and Laos. Water 2021, 13, 641. [Google Scholar] [CrossRef]
- Davie, P.S.; Franklin, C.E.; Grigg, G.C. Blood pressure and heart rate during tonic immobility in the black tipped reef shark, Carcharhinus melanoptera. Fish Physiol. Biochem. 1993, 12, 95–100. [Google Scholar] [CrossRef] [PubMed]
- Brooks, E.J.; Sloman, K.A.; Liss, S.; Hassan-Hassanein, L.; Danylchuk, A.J.; Cooke, S.J.; Mandelman, J.W.; Skomal, G.B.; Sims, D.W.; Suski, C.D. The stress physiology of extended duration tonic immobility in the juvenile lemon shark, Negaprion brevirostris (Poey 1868). J. Exp. Mar. Biol. Ecol. 2011, 409, 351–360. [Google Scholar] [CrossRef]
- Mitamura, H.; Mitsunaga, Y.; Arai, N.; Yamagishi, Y.; Khachaphichat, M.; Viputhanumas, T. Horizontal and vertical movement of Mekong giant catfish Pangasianodon gigas measured using acoustic telemetry in Mae Peum Reservoir, Thailand. Fish. Sci. 2008, 74, 787–795. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Receiver | Lat WGS84 | Long WGS84 | Approximate Distance from River Mouth | Location | Primary Site Use | Deployment Date |
|---|---|---|---|---|---|---|
| 1 | 13.381447 | 99.984996 | 4 | Near estuary | FH | 18 March 18 |
| 2 | 13.389970 | 99.987304 | 5.5 | Mussel Pier | FH | 18 March 18 |
| 3 | 13.407168 | 99.997295 | 7 | Marine Department | PH | 18 March 18 |
| 4 | 13.414427 | 99.983757 | 10 | Baan Tai Had Resort | AQ/RC | 18 March 18 |
| 5 | 13.401533 | 99.976614 | 12.5 | Wat Tai Had | AQ/RC | 18 March 18 |
| 6 | 13.424104 | 99.956114 | 18 | Chaosamran Restaurant | RC/EC | 18 March 18 |
| 7 | 13.432719 | 99.948545 | 20 | Wat Bang Khae Noi | RC | 3 April 18 |
| 8 | 13.446162 | 99.943028 | 22 | Wat Bang Kung | RC | 23 July 18 |
| Tag No. | Maturity | Sex | TL | GL | DW | WT | Tag Date |
|---|---|---|---|---|---|---|---|
| 1058 | Immature | Female | 2.28 | 0.9 | 0.99 | 23 July 2018 | |
| 1059 | Immature | Female | 2.32 | 0.94 | 1.02 | 26 | 22 May 2018 |
| 1063 | Immature | Female | 1.51 | 0.63 | 0.72 | 20 March 2018 | |
| 1069 | Immature | Female | 2.02 | 0.72 | 0.82 | 13 | 21 May 2018 |
| 1071 | Immature | Female | 1.9 | 0.69 | 0.72 | 18 June 2018 | |
| 1073 | Immature | Female | 2.48 | 0.89 | 0.95 | 22 | 23 July 2018 |
| 1057 | Immature | Male | 1.1 | 0.87 | 0.77 | 15 | 5 April 2018 |
| 1075 | Immature | Male | 2.2 | 0.72 | 0.8 | 14.25 | 3 April 2018 |
| 1078 | Immature | Male | 2.24 | 0.87 | 0.92 | 21.5 | 19 June 2018 |
| 1079 | Immature | Male | 2.36 | 0.92 | 0.98 | 28.1 | 19 June 2018 |
| 1061 | Mature | Female | 2.82 | 1.52 | 1.66 | 105 | 23 July 2018 |
| 1062 | Mature | Female | 3.38 | 1.6 | 1.69 | 19 March 2018 | |
| 1067 | Mature | Female | 4.14 | 2 | 2.06 | 21 May 2018 | |
| 1070 | Mature | Female | 3.92 | 1.75 | 2.03 | 19 June 2018 | |
| 1076 | Mature | Female | 3.95 | 1.79 | 1.93 | 19 June 2018 | |
| 1077 | Mature | Female | 2.9 | 1 | 1.1 | 36.7 | 18 June 2018 |
| 1064 | Mature | Male | 2.92 | 1.26 | 1.31 | 62 | 20 March 2018 |
| 1066 | Mature | Male | 2.65 | 1.1 | 1.21 | 44.3 | 24 April 2018 |
| 1068 | Mature | Male | 2.7 | 0.99 | 1.07 | 33 | 4 April 2018 |
| 1074 | Mature | Male | 3.18 | 1.16 | 1.25 | 50 | 23 April 2018 |
| 1080 | Mature | Male | 2.69 | 1.03 | 1.16 | 40 | 20 March 2018 |
| 1081 | Mature | Male | 2.72 | 1.04 | 1.12 | 36 | 4 April 2018 |
| Mat. | Sex | Tag No. | Tag Date | Days Monitored | Days Detected | Total Detections | Presence Events | Presence Duration | RI |
|---|---|---|---|---|---|---|---|---|---|
| I | F | 1058 | 23 July 2018 | 204 | 170 | 5942 | 489 | 335:34:00 | 83 |
| I | F | 1059 | 22 May 2018 | 266 | 122 | 4203 | 279 | 277:05:00 | 46 |
| I | F | 1063 | 20 March 2018 | 329 | 0 | 0 | 0 | 0 | 0 |
| I | F | 1069 | 21 May 2018 | 267 | 50 | 1206 | 107 | 171:04:00 | 19 |
| I | F | 1071 | 18 June 2018 | 239 | 3 | 85 | 6 | 16:50:00 | 1 |
| I | F | 1073 | 23 July 2018 | 204 | 11 | 405 | 32 | 38:07:00 | 5 |
| I | M | 1057 | 5 April 2018 | 313 | 9 | 843 | 16 | 41:04:00 | 3 |
| I | M | 1075 | 3 April 2018 | 315 | 1 | 24 | 2 | 1:12:00 | 0.3 |
| I | M | 1078 | 19 June 2018 | 238 | 147 | 7062 | 328 | 453:53:00 | 62 |
| I | M | 1079 | 19 June 2018 | 238 | 37 | 465 | 61 | 35:10:00 | 16 |
| M | F | 1061 | 23 July 2018 | 204 | 26 | 1765 | 98 | 88:30:00 | 14 |
| M | F | 1062 | 19 March 2018 | 330 | 0 | 0 | 0 | 0 | 0 |
| M | F | 1067 | 21 May 2018 | 267 | 10 | 131 | 20 | 10:39:00 | 4 |
| M | F | 1070 | 19 June 2018 | 238 | 162 | 3983 | 400 | 354:44:00 | 61 |
| M | F | 1076 | 19 June 2018 | 238 | 191 | 3486 | 482 | 250:58:00 | 80 |
| M | F | 1077 | 18 June 2018 | 239 | 81 | 805 | 143 | 54:34:00 | 34 |
| M | M | 1064 | 20 March 2018 | 329 | 2 | 29 | 2 | 2:49:00 | 0.6 |
| M | M | 1066 | 24 April 2018 | 294 | 276 | 9587 | 1084 | 604:33:00 | 94 |
| M | M | 1068 | 4 April 2018 | 314 | 261 | 26,700 | 912 | 1299:29:00 | 83 |
| M | M | 1074 | 23 April 2018 | 295 | 3 | 32 | 7 | 3:14:00 | 1 |
| M | M | 1080 | 20 March 2018 | 329 | 0 | 0 | 0 | 0 | 0 |
| M | M | 1081 | 4 April 2018 | 314 | 21 | 184 | 41 | 15:25:00 | 7 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Haetrakul, T.; Campbell, T.; Daochai, C.; Keschumras, N.; Tantiveerakul, T.; Hogan, Z.; Chansue, N. Assessing the Movements, Habitat Use, and Site Fidelity of the Giant Freshwater Whipray (Urogymnus polylepis) with Acoustic Telemetry in the Maeklong River, Thailand. Water 2023, 15, 2311. https://doi.org/10.3390/w15132311
Haetrakul T, Campbell T, Daochai C, Keschumras N, Tantiveerakul T, Hogan Z, Chansue N. Assessing the Movements, Habitat Use, and Site Fidelity of the Giant Freshwater Whipray (Urogymnus polylepis) with Acoustic Telemetry in the Maeklong River, Thailand. Water. 2023; 15(13):2311. https://doi.org/10.3390/w15132311
Chicago/Turabian StyleHaetrakul, Thanida, Teresa Campbell, Chayanis Daochai, Nitiwadee Keschumras, Tanatporn Tantiveerakul, Zeb Hogan, and Nantarika Chansue. 2023. "Assessing the Movements, Habitat Use, and Site Fidelity of the Giant Freshwater Whipray (Urogymnus polylepis) with Acoustic Telemetry in the Maeklong River, Thailand" Water 15, no. 13: 2311. https://doi.org/10.3390/w15132311