

Retrospective Analysis of the Detection of Pathogens Associated with the Porcine Respiratory Disease Complex in Routine Diagnostic Samples from Austrian Swine Stocks
 , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Design and Sample Collection
2.2. Virological Investigations
2.3. Microbiological Investigations
2.4. Evaluation and Statistical Analyses
3. Results
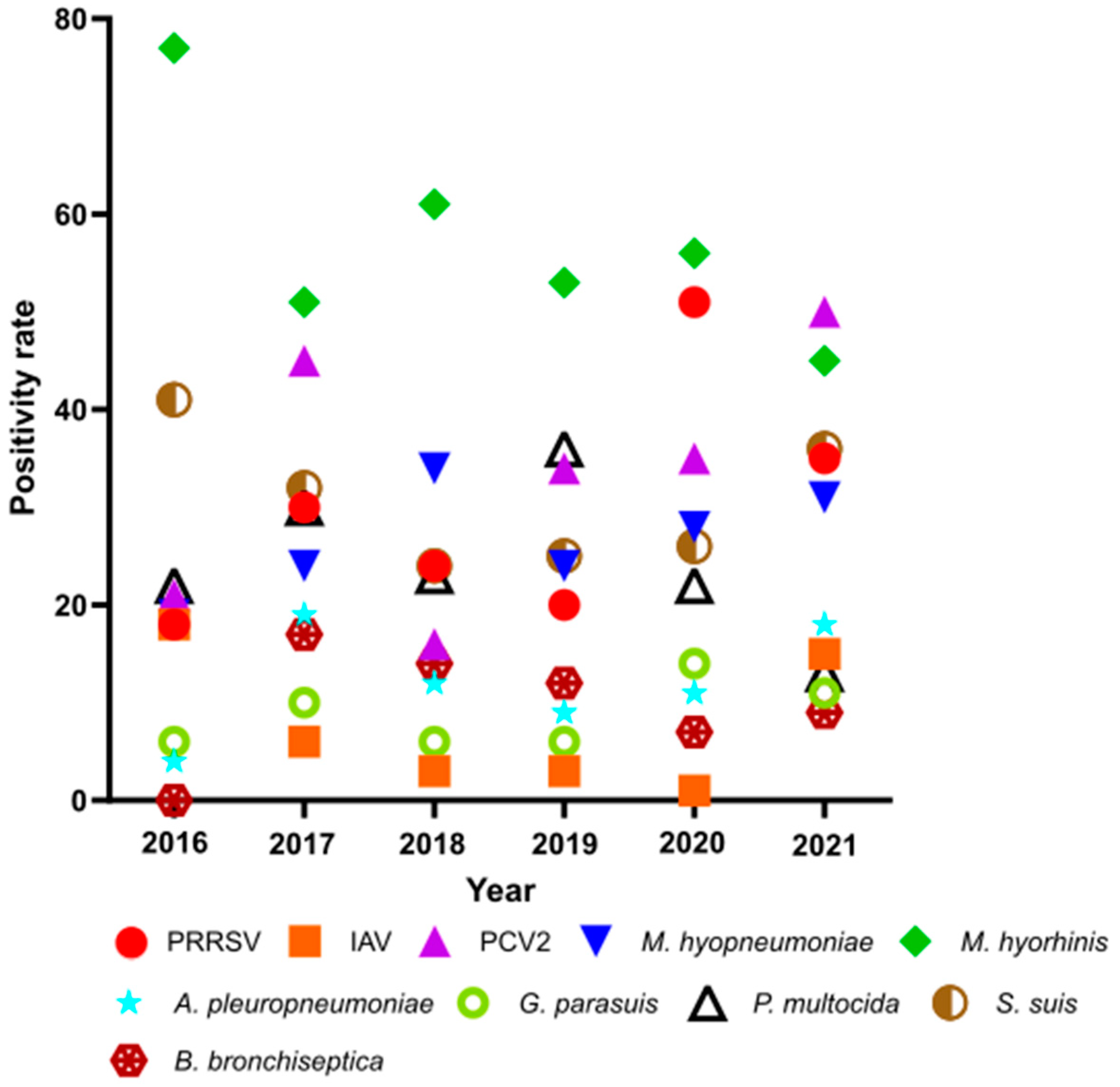
3.1. Positivity Rates
3.2. Positivity Rates of Pathogens in Different Specimens
3.3. Concurrent Infections
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hansen, M.S.; Pors, S.E.; Jensen, H.E.; Bille-Hansen, V.; Bisgaard, M.; Flachs, E.M.; Nielsen, O.L. An Investigation of the Pathology and Pathogens Associated with Porcine Respiratory Disease Complex in Denmark. J. Comp. Pathol. 2010, 143, 120–131. [Google Scholar] [CrossRef]
- Thacker, E.L. Immunology of the Porcine Respiratory Disease Complex. Vet. Clin. N. Am.-Food Anim. Pract. 2001, 17, 551–565. [Google Scholar] [CrossRef]
- Brockmeier, S.L.; Halbur, P.G.; Thacker, E.L. Porcine Respiratory Disease Complex. In Polymicrobial Diseases; Brogden, K.A., Guthmiller, J.M., Eds.; ASM Press: Washington, DC, USA, 2014; pp. 231–258. ISBN 978-1-68367-232-6. [Google Scholar]
- Asai, T.; Okada, M.; Ono, M.; Irisawa, T.; Mori, Y.; Yokomizo, Y.; Sato, S. Increased Levels of Tumor Necrosis Factor and Interleukin 1 in Bronchoalveolar Lavage Fluids from Pigs Infected with Mycoplasma Hyopneumoniae. Vet. Immunol. Immunopathol. 1993, 38, 253–260. [Google Scholar] [CrossRef]
- Maes, D.; Verdonck, M.; Deluyker, H.; de Kruif, A. Enzootic Pneumonia in Pigs. Vet. Q. 1996, 18, 104–109. [Google Scholar] [CrossRef]
- Saade, G.; Deblanc, C.; Bougon, J.; Marois-Créhan, C.; Fablet, C.; Auray, G.; Belloc, C.; Leblanc-Maridor, M.; Gagnon, C.A.; Zhu, J.; et al. Coinfections and Their Molecular Consequences in the Porcine Respiratory Tract. Vet. Res. 2020, 51, 80. [Google Scholar] [CrossRef]
- Bochev, I. Porcine Respiratory Disease Complex (PRDC): A Review. I. Etiology, Epidemiology, Clinical Forms and Pathoanatomical Features. Bulg. J. Vet. Res. 2007, 3, 131–146. [Google Scholar]
- Obradovic, M.R.; Segura, M.; Segalés, J.; Gottschalk, M. Review of the Speculative Role of Co-Infections in Streptococcus Suis-Associated Diseases in Pigs. Vet. Res. 2021, 52, 49. [Google Scholar] [CrossRef]
- Zhu, H.; Chang, X.; Zhou, J.; Wang, D.; Zhou, J.; Fan, B.; Ni, Y.; Yin, J.; Lv, L.; Zhao, Y.; et al. Co-infection Analysis of Bacterial and Viral Respiratory Pathogens from Clinically Healthy Swine in Eastern China. Vet. Med. Sci. 2021, 7, 1815–1819. [Google Scholar] [CrossRef]
- Sassu, E.L.; Bossé, J.T.; Tobias, T.J.; Gottschalk, M.; Langford, P.R.; Henning-Pauka, I. Update on Actinobacillus pleuropneumoniae-knowledge, gaps and challenges. Transbound. Emerg. Dis. 2018, 65, 72–90. [Google Scholar]
- Chiers, K.; Ducatelle, R.; Haesebrouck, F. Actinobacillus Pleuropneumoniae Infections in Closed Swine Herds: Infection Patterns and Serological Profiles. Vet. Microbiol. 2002, 85, 343–352. [Google Scholar]
- Sidibé, M.; Messier, S.; Lariviere, S.; Gottschalk, M.; Mittal, K.R. Detection of Actinobacillus Pleuropneumoniae in the Porcine Upper Respiratory Tract as a Complement to Serological Tests. Can. J. Vet. Res. 1993, 57, 204–208. [Google Scholar]
- Macedo, N.; Rovira, A.; Oliveira, S.; Holtcamp, A.; Torremorell, M. Effect of Enrofloxacin in the Carrier Stage of Haemophilus Parasuis in Naturally Colonized Pigs. Can. J. Vet. Res. 2014, 78, 17–22. [Google Scholar]
- MacInnes, J.I.; Gottschalk, M.; Lone, A.G.; Metcalf, D.S.; Ojha, S.; Rosendal, T.; Watson, S.B.; Friendship, R.M. Prevalence of Actinobacillus Pleuropneumoniae, Actinobacillus Suis, Haemophilus Parasuis, Pasteurella Multocida, and Streptococcus Suis in Representative Ontario Swine Herds. Can. J. Vet. Res. 2008, 72, 242–248. [Google Scholar]
- Eriis, N.E.; Eeenstra, A.A. Mycoplasma Hyorhinis in the Etiology of Serositis among Piglets. Acta Vet. Scand. 1994, 35, 93–98. [Google Scholar] [CrossRef]
- Prickett, J.; Simer, R.; Christopher-Hennings, J.; Yoon, K.-J.; Evans, R.B.; Zimmerman, J.J. Detection of Porcine Reproductive and Respiratory Syndrome Virus Infection in Porcine Oral Fluid Samples: A Longitudinal Study under Experimental Conditions. J. Vet. Diagn. Investig. 2008, 20, 156–163. [Google Scholar] [CrossRef]
- Decorte, I.; Van Campe, W.; Mostin, L.; Cay, A.B.; De Regge, N. Diagnosis of the Lelystad Strain of Porcine Reproductive and Respiratory Syndrome Virus Infection in Individually Housed Pigs: Comparison between Serum and Oral Fluid Samples for Viral Nucleic Acid and Antibody Detection. J. Vet. Diagn. Investig. 2015, 27, 47–54. [Google Scholar] [CrossRef]
- Olsen, C.; Wang, C.; Christopher-Hennings, J.; Doolittle, K.; Harmon, K.M.; Abate, S.; Kittawornrat, A.; Lizano, S.; Main, R.; Nelson, E.A.; et al. Probability of Detecting Porcine Reproductive and Respiratory Syndrome Virus Infection Using Pen-Based Swine Oral Fluid Specimens as a Function of within-Pen Prevalence. J. Vet. Diagn. Investig. 2013, 25, 328–335. [Google Scholar] [CrossRef]
- López, W.A.; Zimmerman, J.J.; Gauger, P.C.; Harmon, K.M.; Bradner, L.; Zhang, M.; Giménez-Lirola, L.; Ramirez, A.; Cano, J.P.; Linhares, D.C.L. Practical Aspects of PRRSV RNA Detection in Processing Fluids Collected in Commercial Swine Farms. Prev. Vet. Med. 2020, 180, 105021. [Google Scholar] [CrossRef]
- Halbur, P.G.; Paul, P.S.; Frey, M.L.; Landgraf, J.; Eernisse, K.; Meng, X.-J.; Andrews, J.J.; Lum, M.A.; Rathje, J.A. Comparison of the Antigen Distribution of Two US Porcine Reproductive and Respiratory Syndrome Virus Isolates with That of the Lelystad Virus. Vet. Pathol. 1996, 33, 159–170. [Google Scholar] [CrossRef]
- Segalés, J.; Sibila, M. Revisiting Porcine Circovirus Disease Diagnostic Criteria in the Current Porcine Circovirus 2 Epidemiological Context. Vet. Sci. 2022, 9, 110. [Google Scholar] [CrossRef]
- Segalés, J. Porcine Circovirus Type 2 (PCV2) Infections: Clinical Signs, Pathology and Laboratory Diagnosis. Virus Res. 2012, 164, 10–19. [Google Scholar] [CrossRef]
- Prickett, J.R.; Johnson, J.; Murtaugh, M.P.; Puvanendiran, S.; Wang, C.; Zimmerman, J.J.; Opriessnig, T. Prolonged Detection of PCV2 and Anti-PCV2 Antibody in Oral Fluids Following Experimental Inoculation: Prolonged Detection of PCV2 Infection by Oral Fluid. Transbound. Emerg. Dis. 2011, 58, 121–127. [Google Scholar] [CrossRef]
- Caprioli, A.; McNeilly, F.; McNair, I.; Lagan-Tregaskis, P.; Ellis, J.; Krakowka, S.; McKillen, J.; Ostanello, F.; Allan, G. PCR Detection of Porcine Circovirus Type 2 (PCV2) DNA in Blood, Tonsillar and Faecal Swabs from Experimentally Infected Pigs. Res. Vet. Sci. 2006, 81, 287–292. [Google Scholar] [CrossRef]
- Hernandez-Garcia, J.; Robben, N.; Magnée, D.; Eley, T.; Dennis, I.; Kayes, S.M.; Thomson, J.R.; Tucker, A.W. The Use of Oral Fluids to Monitor Key Pathogens in Porcine Respiratory Disease Complex. Porc. Health Manag. 2017, 3, 7. [Google Scholar] [CrossRef]
- Pieters, M.; Daniels, J.; Rovira, A. Comparison of Sample Types and Diagnostic Methods for in Vivo Detection of Mycoplasma Hyopneumoniae during Early Stages of Infection. Vet. Microbiol. 2017, 203, 103–109. [Google Scholar] [CrossRef]
- Lin, J.; Chen, S.; Yeh, K.; Weng, C. Mycoplasma Hyorhinis in Taiwan: Diagnosis and Isolation of Swine Pneumonia Pathogen. Vet. Microbiol. 2006, 115, 111–116. [Google Scholar] [CrossRef]
- Makhanon, M.; Tummaruk, P.; Thongkamkoon, P.; Thanawongnuwech, R.; Prapasarakul, N. Comparison of Detection Procedures of Mycoplasma Hyopneumoniae, Mycoplasma Hyosynoviae, and Mycoplasma Hyorhinis in Lungs, Tonsils, and Synovial Fluid of Slaughtered Pigs and Their Distributions in Thailand. Trop. Anim. Health Prod. 2012, 44, 313–318. [Google Scholar] [CrossRef]
- Pohl, S.; Frederiksen, W.; Mannheim’, W. Transfer of Haemophilus Pleuropneumoniae and the Pasteurella Haernolytica-Like Organism Causing Porcine Necrotic Pleuropneumonia to the Genus Actinobacillus (Actinobacillus Pleuropneumoniae Comb. Nov.) on the Basis of Phenotypic and Deoxyribonucleic Acid Relatedness. Int. J. Syst. Microbiol. 1983, 33, 510–514. [Google Scholar]
- Oliveira, S.; Pijoan, C. Haemophilus Parasuis: New Trends on Diagnosis, Epidemiology and Control. Vet. Microbiol. 2004, 99, 1–12. [Google Scholar] [CrossRef]
- Kawashima, K.; Yamada, S.; Kobayashi, H.; Narita, M. Detection of Porcine Reproductive and Respiratory Syndrome Virus and Mycoplasma Hyorhinis Antigens in Pulmonary Lesions of Pigs Suffering from Respiratory Distress. J. Comp. Pathol. 1996, 114, 315–323. [Google Scholar] [CrossRef]
- Luehrs, A.; Siegenthaler, S.; Grützner, N.; Beilage, E.G.; Kuhnert, P.; Nathues, H. Occurrence of Mycoplasma Hyorhinis Infections in Fattening Pigs and Association with Clinical Signs and Pathological Lesions of Enzootic Pneumonia. Vet. Microbiol. 2017, 203, 1–5. [Google Scholar] [CrossRef]
- Lee, J.-A.; Oh, Y.-R.; Hwang, M.-A.; Lee, J.-B.; Park, S.-Y.; Song, C.-S.; Choi, I.-S.; Lee, S.-W. Mycoplasma Hyorhinis Is a Potential Pathogen of Porcine Respiratory Disease Complex That Aggravates Pneumonia Caused by Porcine Reproductive and Respiratory Syndrome Virus. Vet. Immunol. Immunopathol. 2016, 177, 48–51. [Google Scholar] [CrossRef]
- Palzer, A.; Ritzmann, M.; Wolf, G.; Heinritzi, K. Associations between Pathogens in Healthy Pigs and Pigs with Pneumonia. Vet. Rec. 2008, 162, 267–271. [Google Scholar] [CrossRef]
- Vötsch, D.; Willenborg, M.; Weldearegay, Y.B.; Valentin-Weigand, P. Streptococcus Suis—The “Two Faces” of a Pathobiont in the Porcine Respiratory Tract. Front. Microbiol. 2018, 9, 480. [Google Scholar] [CrossRef]
- Zimmerman, J.J.; Karriker, L.A.; Ramirez, A.; Schwartz, K.J.; Stevenson, G.W.; Jianqiang, Z. (Eds.) Diseases of Swine, 11th ed.; Wiley-Blackwell/American Association of Swine Veterinarians: Hoboken, NJ, USA, 2019; p. 397. [Google Scholar]
- Lu, K.; Wang, C.; Shu, J.; Obeng, E.; Wu, Y.; Chen, J.; Shu, J.; He, Y. Co-Infection of Mycoplasma Hyopneumoniae and Other Swine Pathogens. Authorea. 2020. Available online: https://www.authorea.com/users/322723/articles/451679-co-infection-of-mycoplasma-hyopneumoniae-and-other-swine-pathogens?commit=8d40934e7b93a18af20687c0ef77510959ec113a (accessed on 12 January 2023).
- Brockmeier, S.L.; Palmer, M.V.; Bolin, S.R.; Rimler, R.B. Effects of Intranasal Inoculation with Bordetella Bronchiseptica, Porcine Reproductive and Respiratory Syndrome Virus, or a Combination of Both Organisms on Subsequent Infection with Pasteurella Multocida in Pigs. Am. J. Vet. Res. 2001, 62, 521–525. [Google Scholar] [CrossRef]
- Opriessnig, T.; Giménez-Lirola, L.G.; Halbur, P.G. Polymicrobial Respiratory Disease in Pigs. Anim. Health. Res. Rev. 2011, 12, 133–148. [Google Scholar] [CrossRef]
- Fetzer, C.; Pesch, S.; Ohlinger, V. High Risk of False Positive Results in a Widely Used Diagnostic Test for Detection of the Porcine Reproductive and Respiratory Syndrome Virus (PRRSV). Vet. Microbiol. 2006, 115, 21–31. [Google Scholar] [CrossRef]
- Ward, C. Design and Performance Testing of Quantitative Real Time PCR Assays for Influenza A and B Viral Load Measurement. J. Clin. Virol. 2004, 29, 179–188. [Google Scholar] [CrossRef]
- Hoffmann, B. Genomnachweis des Porcinen Circovirus Typ 2 (pCV-2) Mittels Real-Time PCR (FLI Methode). AVID-Methodensammlung. Available online: http://avid.dvg.net/fileadmin/Bilder/PDF_AVID_Alt/website/Methoden_ab_2016/AVID-Methode_VIR02_porcines_Circovirus_2_final.pdf (accessed on 11 February 2023).
- Toussaint, J.F.; Sailleau, C.; Breard, E.; Zientara, S.; De Clercq, K. Bluetongue Virus Detection by Two Real-Time RT-QPCRs Targeting Two Different Genomic Segments. J. Virol. Methods 2007, 140, 115–123. [Google Scholar] [CrossRef]
- Bünger, M.; Brunthaler, R.; Unterweger, C.; Loncaric, I.; Dippel, M.; Ruczizka, U.; Schwarz, L.; Griessler, A.; Voglmayr, T.; Verhovsek, D.; et al. Mycoplasma Hyorhinis as a Possible Cause of Fibrinopurulent Meningitis in Pigs?—A Case Series. Porc. Health Manag. 2020, 6, 38. [Google Scholar] [CrossRef]
- Spergser, J.; Hess, C.; Loncaric, I.; Ramírez, A.S. Matrix-Assisted Laser Desorption Ionization–Time of Flight Mass Spectrometry Is a Superior Diagnostic Tool for the Identification and Differentiation of Mycoplasmas Isolated from Animals. J. Clin. Microbiol. 2019, 57, e00316-19. [Google Scholar] [CrossRef]
- Caron, J.; Ouardani, M.; Dea, S. Diagnosis and Differentiation of Mycoplasma Hyopneumoniae and Mycoplasma Hyorhinis Infections in Pigs by PCR Amplification of the P36 and P46 Genes. J. Clin. Microbiol. 2000, 38, 1390–1396. [Google Scholar] [CrossRef]
- Kurth, K.T.; Hsu, T.; Snook, E.R.; Thacker, E.L.; Thacker, B.J.; Minion, F.C. Use of a Mycoplasma Hyopneumoniae Nested Polymerase Chain Reaction Test to Determine the Optimal Sampling Sites in Swine. J. Vet. Diagn. Investig. 2002, 14, 463–469. [Google Scholar] [CrossRef]
- Cho, W.-S.; Chae, C. PCR Detection of Actinobacillus Pleuropneumoniae apxIV Gene in Formalin-Fixed, Paraffin-Embedded Lung Tissues and Comparison with In Situ Hybridization. Lett. Appl. Microbiol. 2003, 37, 56–60. [Google Scholar] [CrossRef]
- Kang, I.; Kim, D.; Han, K.; Seo, H.W.; Oh, Y.; Park, C.; Lee, J.; Gottschalk, M.; Chae, C. Optimized Protocol for Multiplex Nested Polymerase Chain Reaction to Detect and Differentiate Haemophilus Parasuis, Streptococcus Suis, and Mycoplasma Hyorhinis in Formalin-Fixed, Paraffin-Embedded Tissues from Pigs with Polyserositis. Can. J. Vet. Res. 2012, 76, 195–200. [Google Scholar]
- SPSS Statistics—Überblick. Available online: https://www.ibm.com/de-de/products/spss-statistics (accessed on 12 October 2020).
- Allende, R.; Laegreid, W.W.; Kutish, G.F.; Galeota, J.A.; Wills, R.W.; Osorio, F.A. Porcine Reproductive and Respiratory Syndrome Virus: Description of Persistence in Individual Pigs upon Experimental Infection. J. Virol. 2000, 74, 10834–10837. [Google Scholar] [CrossRef]
- Beyer, J.; Fichtner, D.; Schirrmeir, H.; Polster, U.; Weiland, E.; Wege, H. Porcine Reproductive and Respiratory Syndrome Virus (PRRSV): Kinetics of Infection in Lymphatic Organs and Lung. J. Vet. Med. Ser. B 2000, 47, 9–25. [Google Scholar] [CrossRef]
- Gibert, E.; Martín-Valls, G.; Mateu, E. Comparison of Protocols for the Analysis of Type 1 Porcine Reproductive and Respiratory Syndrome Virus by RT-PCR Using Oral Fluids. J. Virol. Methods 2017, 243, 190–195. [Google Scholar] [CrossRef]
- Savić, B.; Radanović, O.; Jovičić, D.; Nešić, K.; Ivanović, S.; Stevančević, O.; Cvetojević, Đ.; Kasagić, D. Survey of Infectious Agents Associated with Porcine Respiratory Disease Complex (PRDC) in Serbian Swine Herds Using Polymerase Chain Reaction (PCR) Detection. Acta Vet. Beogr. 2015, 65, 79–88. [Google Scholar] [CrossRef]
- Takemae, N.; Tsunekuni, R.; Uchida, Y.; Ito, T.; Saito, T. Experimental Infection of Pigs with H1 and H3 Influenza A Viruses of Swine by Using Intranasal Nebulization. BMC Vet. Res. 2018, 14, 115. [Google Scholar] [CrossRef]
- Sreta, D.; Kedkovid, R.; Tuamsang, S.; Kitikoon, P.; Thanawongnuwech, R. Pathogenesis of Swine Influenza Virus (Thai Isolates) in Weanling Pigs: An Experimental Trial. Virol. J. 2009, 6, 34. [Google Scholar] [CrossRef]
- Valette, M.; Bouscambert-Duchamp, M.; Fanget, R.; Lambert, S.; Lina, B. Comparison of Virocult® Swab, Σ-Swabtm and Σ-Virocult® For Influenza a Viability for Cell Culture and Molecular Detection. Available online: https://www.labshop-online.com/792131256/DWN/LabShop_MWE_Sigma_Liquid_Swabs.pdf (accessed on 11 February 2023).
- Yu, S.; Opriessnig, T.; Kitikoon, P.; Nilubol, D.; Halbur, P.G.; Thacker, E. Porcine Circovirus Type 2 (PCV2) Distribution and Replication in Tissues and Immune Cells in Early Infected Pigs. Vet. Immunol. Immunopathol. 2007, 115, 261–272. [Google Scholar] [CrossRef]
- Opriessnig, T.; Karuppannan, A.K.; Castro, A.M.M.G.; Xiao, C.-T. Porcine Circoviruses: Current Status, Knowledge Gaps and Challenges. Virus Res. 2020, 286, 198044. [Google Scholar] [CrossRef]
- Deutscher, A.T.; Jenkins, C.; Minion, F.C.; Seymour, L.M.; Padula, M.P.; Dixon, N.E.; Walker, M.J.; Djordjevic, S.P. Repeat Regions R1 and R2 in the P97 Paralogue Mhp271 of Mycoplasma Hyopneumoniae Bind Heparin, Fibronectin and Porcine Cilia: Mhp271 Is a Multifunctional Surface Protein. Mol. Microbiol. 2010, 78, 444–458. [Google Scholar] [CrossRef]
- Allan, G.M.; Ellis, J.A. Porcine Circoviruses: A Review. J. Vet. Diagn. Investig. 2000, 12, 3–14. [Google Scholar] [CrossRef]
- Clavijo, M.J.; Hu, D.; Krantz, S.; Cano, J.P.; Pereira Maróstica, T.; Henao-Diaz, A.; Poeta Silva, A.P.S.; Hemker, D.; Tapia, E.; Zimmerman, S.; et al. Mycoplasma Hyopneumoniae Surveillance in Pig Populations: Establishing Sampling Guidelines for Detection in Growing Pigs. J. Clin. Microbiol. 2021, 59, 1128. [Google Scholar] [CrossRef]
- Fablet, C.; Marois, C.; Dorenlor, V.; Eono, F.; Eveno, E.; Jolly, J.P.; Le Devendec, L.; Kobisch, M.; Madec, F.; Rose, N. Bacterial Pathogens Associated with Lung Lesions in Slaughter Pigs from 125 Herds. Res. Vet. Sci. 2012, 93, 627–630. [Google Scholar] [CrossRef]
- Smart, N.L.; Miniats, P. Glasser’s Disease and Prevalence of Subclinical Infection with Haemophilus Parasuis in Swine in Southern Ontario. Can. Vet. J. 1989, 30, 337–340. [Google Scholar]
- Turni, C.; Blackall, P. Comparison of Sampling Sites and Detection Methods for Haemophilus Parasuis. Aust. Vet. J. 2007, 85, 177–184. [Google Scholar] [CrossRef]
- Goodnow, R.A. Biology of Bordetella Bronchiseptica. Microbiol. Rev. 1980, 44, 722–738. [Google Scholar]
- Pallarés, F.J.; Halbur, P.G.; Opriessnig, T.; Sorden, S.D.; Villar, D.; Janke, B.H.; Yaeger, M.J.; Larson, D.J.; Schwartz, K.J.; Yoon, K.J.; et al. Porcine Circovirus Type 2 (PCV-2) Coinfections in US Field Cases of Postweaning Multisystemic Wasting Syndrome (PMWS). J. Vet. Diagn. Investig. 2002, 14, 515–519. [Google Scholar] [CrossRef]
- Fablet, C.; Marois-Créhan, C.; Grasland, B.; Simon, G.; Rose, N. Factors Associated with Herd-Level PRRSV Infection and Age-Time to Seroconversion in Farrow-to-Finish Herds. Vet. Microbiol. 2016, 192, 10–20. [Google Scholar] [CrossRef]
- Choi, Y.K.; Goyal, S.M.; Joo, H.S. Retrospective Analysis of Etiologic Agents Associated with Respiratory Diseases in Pigs. Can. Vet. J. 2003, 44, 3. [Google Scholar]
- Ferreira, M.M.; Mechler-Dreibi, M.L.; Sonalio, K.; Almeida, H.M.d.S.; Ferraz, M.E.S.; Jacintho, A.P.P.; Maes, D.; de Oliveira, L.G. Co-Infections by Mycoplasma Hyopneumoniae, Mycoplasma Hyorhinis and Mycoplasma Flocculare in Macroscopic Lesions of Lung Consolidation of Pigs at Slaughter. Vet. Microbiol. 2021, 258, 109123. [Google Scholar] [CrossRef]


{kind=link}
| Investigations | Lung | Lymph Node | Lung, Tonsil, Lymph Node | BALF | OF | Nasal Swabs | Total |
|---|---|---|---|---|---|---|---|
| Microbiological examination | 590 | 21 | 23 | 634 | |||
| M. hyorhinis cultivation | 142 | 17 | 8 | 167 | |||
| PRRSV-PCR | 278 | 469 | 38 | 136 | 921 | ||
| IAV-PCR | 240 | 51 | 12 | 94 | 82 | 479 | |
| PCV2-PCR | 168 | 329 | 9 | 10 | 2 | 0 | 518 |
| M. hyopneumoniae-PCR | 605 | 10 | 87 | 11 | 713 | ||
| M. hyorhinis-PCR | 139 | 36 | 5 | 180 | |||
| A. pleuropneumoniae-PCR | 193 | 3 | 2 | 198 | |||
| G. parasuis-PCR | 157 | 8 | 165 | ||||
| Samples (Total) | 874 | 329 | 469 | 56 | 142 | 105 | 1975 |
| Virus | Primer/Probe Sequences | PCR Kit | Reference |
|---|---|---|---|
| PRRSV | Primer-F (PRS133): 5′-ATGGCCAGCCAGTCAATC-3′ | One Taq One-Step RT-PCR Kit (NEB) | [40] |
| Primer-R (PRS134): 5′-TCGCCCTAATTGAATAGGTG-3′ | |||
| IAV | FLUAM-1F: 5′-AAGACCAATCCTGTCACCTCTGA-3′ | Luna® Universal Probe One-Step RT-qPCR Kit (NEB) | [41] |
| FLUAM-2F: 5′-CATTGGGATCTTGCACTTGATATT-3′ | |||
| FLUAM-1R: 5′-CAA AGCGTCTACGCTGCAGTCC-3′ | |||
| FLUAM-2R: 5′-AAACCGTATTTAAGGCGACGATAA-3′ | |||
| FLUAM-1P: FAM-5′-TTTGTGTTCACGCTCACCGT-3′-TAMRA | |||
| FLUAM-2P: FAM-5′-TGGATTCTTGATCGTCTTTTCTTCAAATGCA-3′-TAMRA | |||
| PCV2 | qPCV2-F: 5′-GAGTCTGGTGACCGTTGCA-3′ | Luna® Universal Probe One-Step RT-qPCR Kit (NEB) | [42] |
| qPCV2-R: 5′-YCCCGCTCACTTTCAAAAGTTC-3′ | |||
| qPCV2-probe: FAM-5′-CCCTGTAACGTTTGTCAGAAATTTCCGCG-3′-BHQ1 | |||
| ß-Actin | qBeta-Actin-1005-F: 5′-CAGCACAATGAAGATCAAGATCATC-3′ | Luna® Universal Probe One-Step RT-qPCR Kit (NEB) | [43] |
| qBeta-Actin-1135-R: 5′-CGGACTCATCGTACTCCTGCTT-3′ | |||
| qBeta-Actin-1081-probe: HEX-5′-TCGCTGTCCACCTTCCAGCAGATGT-3′-BHQ1 |
| Lung | Lymph Node | Lung, Tonsil, Lymph Node | BALF | OF | Nasal Swabs | Total | |
|---|---|---|---|---|---|---|---|
| PRRSV | 92/278 | 170/469 | 5/38 | 4/136 | 271/921 | ||
| 33.1% | 36.2% | 13.2% | 2.9% | 29.4% | |||
| IAV | 18/240 | 4/51 | 0/12 | 2/94 | 5/82 | 29/479 | |
| 7.5% | 7.8% | 0% | 2.1% | 6.1% | 6.1% | ||
| PCV2 | 79/168 | 111/329 | 7/9 | 0/10 | 0/2 | 197/518 | |
| 47% | 33.7% | 77.8% | 0% | 0% | 38.0% | ||
| M. hyopneumoniae | 185/605 | 0/10 | 3/87 | 0/11 | 188/713 | ||
| 30.6% | 0% | 3.4% | 0% | 26.6% | |||
| M. hyorhinis | 136/257 | 26/46 | 12/13 | 174/316 | |||
| 52.9% | 56.6% | 92.3% | 55.1% | ||||
| A. pleuropneumoniae | 83/679 | 0/21 | 3/23 | 86/723 | |||
| 12.2% | 0% | 13% | 11.9% | ||||
| G. parasuis | 52/641 | 8/23 | 0/23 | 60/ | |||
| 8.1% | 34.8% | 0% | 8.8% | ||||
| P. multocida | 146/590 | 0/21 | 5/23 | 151/634 | |||
| 24.7% | 0% | 21.7% | 23.8% | ||||
| B. bronchiseptica | 62/590 | 3/21 | 1/23 | 66/634 | |||
| 10.5% | 14.3% | 4.3% | 10.4% | ||||
| S. suis | 163/590 | 18/21 | 13/23 | 194/634 | |||
| 27.6% | 85.7% | 56.5% | 30.6% |
| Investigations | Result | Field Samples | Samples from Animals Submitted for Necropsy |
|---|---|---|---|
| Isolation | Negative | 386 | 209 |
| Positive | 9 | 15 | |
| PCR | Negative | 66 | 64 |
| Positive | 27 | 12 |
| Number of Samples Being Positively Tested for Both Pathogens | Number of Samples Being Tested for Both Pathogens | Positivity Rate | p-Value | |
|---|---|---|---|---|
| P. multocida and S. suis | 151 | 634 | 24% | 0.517 |
| PRRSV and PCV2 | 66 | 368 | 18% | 0.001 |
| M. hyorhinis and S. suis | 66 | 193 | 34% | <0.001 |
| PRRSV and P. multocida | 52 | 347 | 15% | <0.001 |
| M. hyopneumoniae and P. multocida | 41 | 330 | 12% | <0.001 |
| PRRSV and M. hyopneumoniae | 41 | 426 | 10% | 0.032 |
| PRRSV and S. suis | 40 | 347 | 12% | 0.388 |
| M. hyopneumoniae and S. suis | 37 | 330 | 11% | 0.046 |
| PRRSV and A. pleuropneumoniae | 31 | 391 | 8% | 0.001 |
| PCV2 and M. hyorhinis | 29 | 125 | 23% | 0.256 |
| PCV2 and S. suis | 29 | 242 | 12% | 0.398 |
| PRRSV and M. hyorhinis | 26 | 191 | 14% | 0.092 |
| M. hyorhinis and P. multocida | 25 | 193 | 13% | 0.378 |
| M. hyorhinis and G. parasuis | 24 | 212 | 11% | 0.509 |
| PCV2 and P. multocida | 24 | 242 | 10% | 0.856 |
| PCV2 and M. hyopneumoniae | 24 | 265 | 9% | 0.397 |
| M. hyopneumoniae and M. hyorhinis | 23 | 218 | 11% | 0.004 |
| PCV2 and A. pleuropneumoniae | 21 | 272 | 8% | 0.002 |
| G. parasuis and S. suis | 18 | 634 | 3% | 0.793 |
| PRRSV and G. parasuis | 15 | 373 | 4% | 0.562 |
| A. pleuropneumoniae and S. suis | 14 | 634 | 2% | 0.017 |
| PRRSV and B. bronchiseptica | 12 | 347 | 3% | 0.667 |
| PCV2 and B. bronchiseptica | 11 | 242 | 5% | 0.412 |
| M. hyopneumoniae and B. bronchiseptica | 11 | 330 | 3% | 0.464 |
| M. hyorhinis and B. bronchiseptica | 11 | 193 | 6% | 0.696 |
| A. pleuropneumoniae and P. multocida | 11 | 634 | 2% | 0.048 |
| B. bronchiseptica and S. suis | 11 | 634 | 2% | 0.009 |
| PCV2 and G. parasuis | 9 | 264 | 3% | 0.144 |
| PRRSV and IAV | 8 | 364 | 2% | 0.727 |
| IAV and S. suis | 8 | 185 | 4% | 0.724 |
| M. hyopneumoniae and G. parasuis | 8 | 358 | 4% | 0.273 |
| G. parasuis and P. multocida | 8 | 634 | 1% | 0.079 |
| IAV and M. hyorhinis | 7 | 109 | 6% | 0.861 |
| IAV and PCV2 | 5 | 149 | 3% | 0.811 |
| IAV and P. multocida | 4 | 185 | 2% | 0.502 |
| P. multocida and B. bronchiseptica | 4 | 634 | 1% | <0.001 |
| IAV and M. hyopneumoniae | 3 | 291 | 1% | 0.681 |
| IAV and G. parasuis | 2 | 194 | 2% | 0.704 |
| IAV and A. pleuropneumoniae | 2 | 213 | 1% | 0.800 |
| M. hyopneumoniae and A. pleuropneumoniae | 2 | 398 | 1% | <0.001 |
| M. hyorhinis and A. pleuropneumoniae | 2 | 209 | 1% | 0.038 |
| A. pleuropneumoniae and B. bronchiseptica | 2 | 634 | 0% | 0.019 |
| G. parasuis and B. bronchiseptica | 2 | 634 | 0% | 0.079 |
| A. pleuropneumoniae and G. parasuis | 1 | 649 | 0% | 0.014 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Renzhammer, R.; Auer, A.; Loncaric, I.; Entenfellner, A.; Dimmel, K.; Walk, K.; Rümenapf, T.; Spergser, J.; Ladinig, A. Retrospective Analysis of the Detection of Pathogens Associated with the Porcine Respiratory Disease Complex in Routine Diagnostic Samples from Austrian Swine Stocks. Vet. Sci. 2023, 10, 601. https://doi.org/10.3390/vetsci10100601
Renzhammer R, Auer A, Loncaric I, Entenfellner A, Dimmel K, Walk K, Rümenapf T, Spergser J, Ladinig A. Retrospective Analysis of the Detection of Pathogens Associated with the Porcine Respiratory Disease Complex in Routine Diagnostic Samples from Austrian Swine Stocks. Veterinary Sciences. 2023; 10(10):601. https://doi.org/10.3390/vetsci10100601
Chicago/Turabian StyleRenzhammer, René, Angelika Auer, Igor Loncaric, Annabell Entenfellner, Katharina Dimmel, Karin Walk, Till Rümenapf, Joachim Spergser, and Andrea Ladinig. 2023. "Retrospective Analysis of the Detection of Pathogens Associated with the Porcine Respiratory Disease Complex in Routine Diagnostic Samples from Austrian Swine Stocks" Veterinary Sciences 10, no. 10: 601. https://doi.org/10.3390/vetsci10100601