
Hepatic Coccidiosis in Wild Rabbits in Greece: Parasite Detection on Liver Imprints and the Associated Biochemical Profile

 , , , , , , and
, , , , , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Blood Samples and Biochemical Analyses
2.3. Liver Imprints
2.4. Statistics
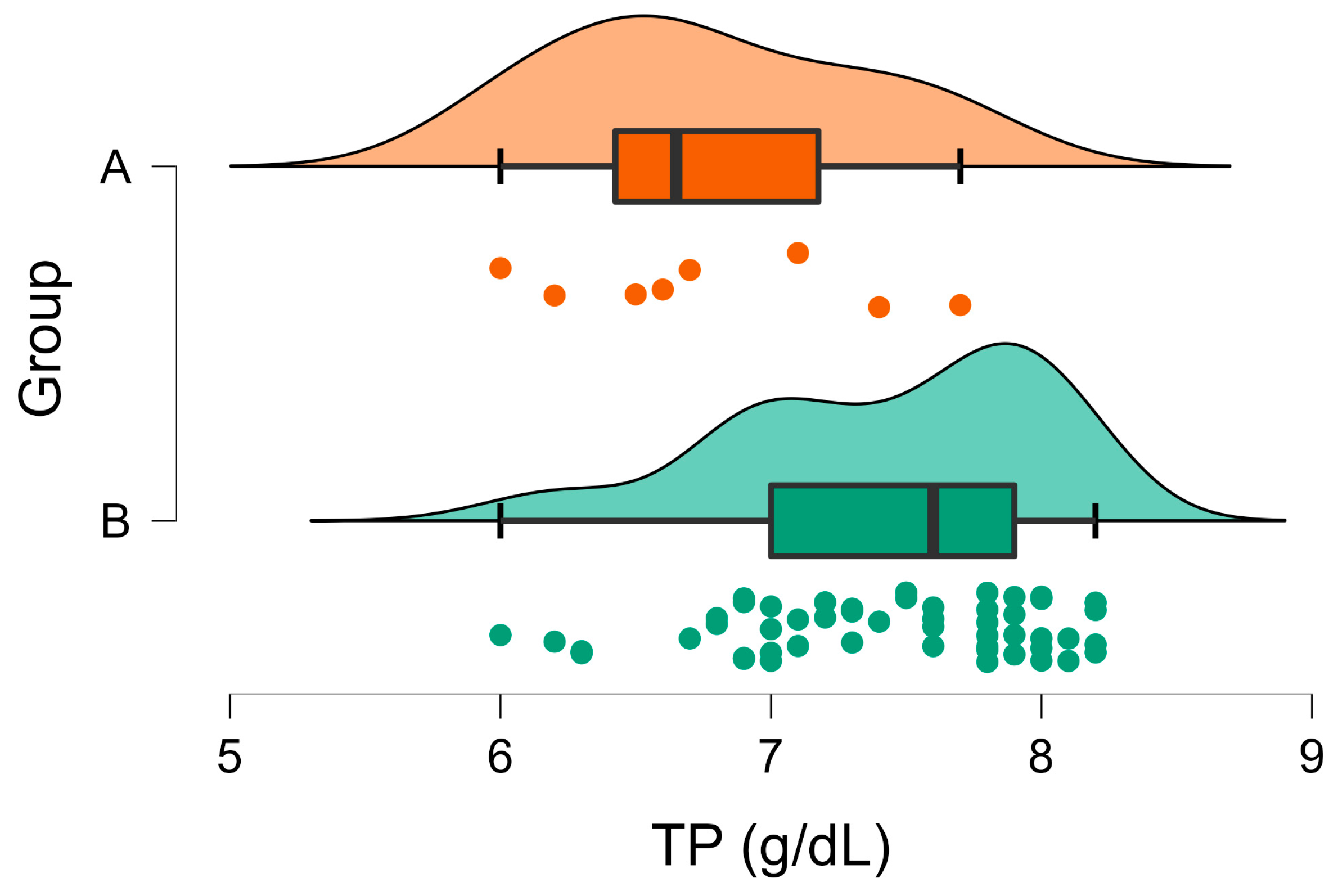
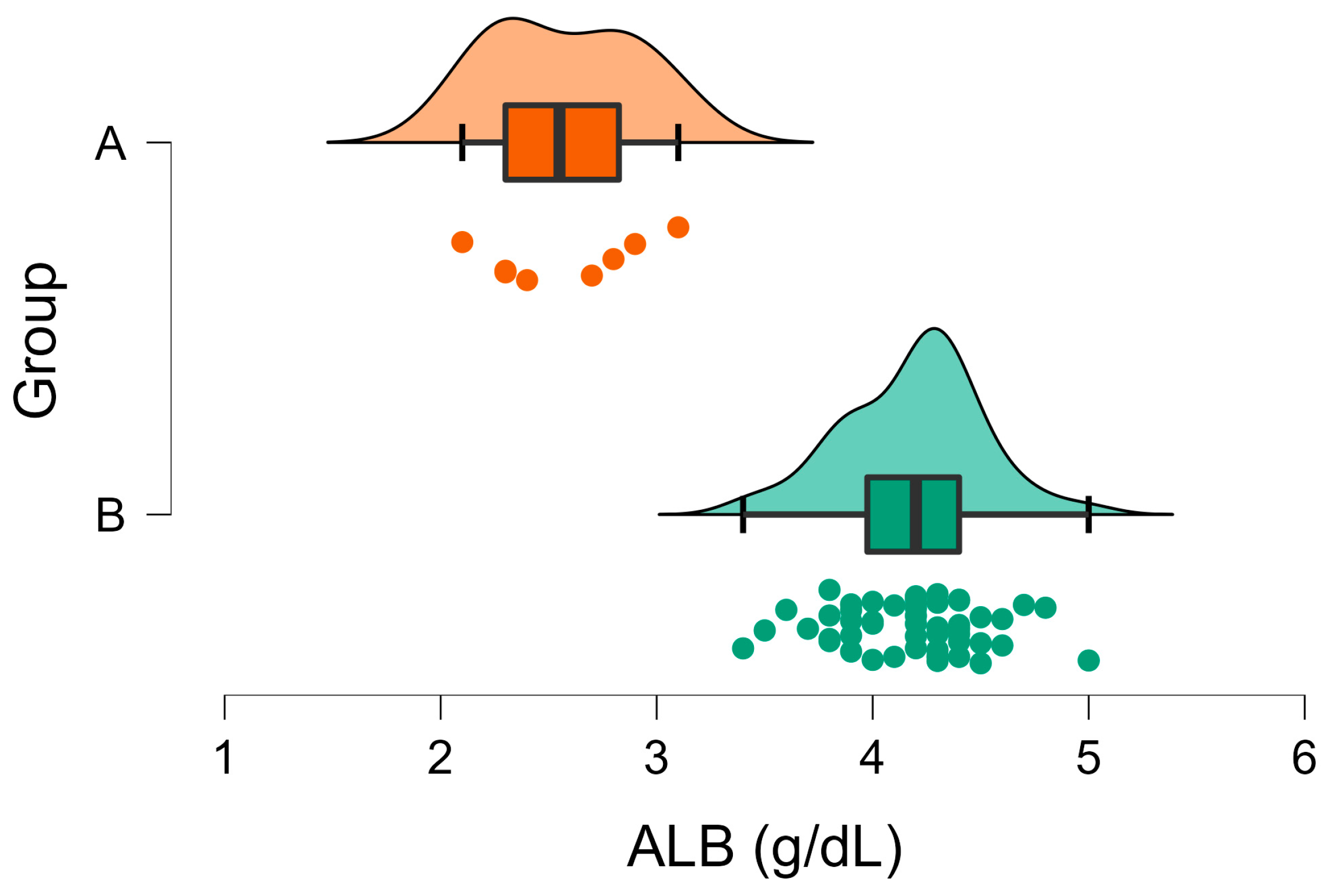
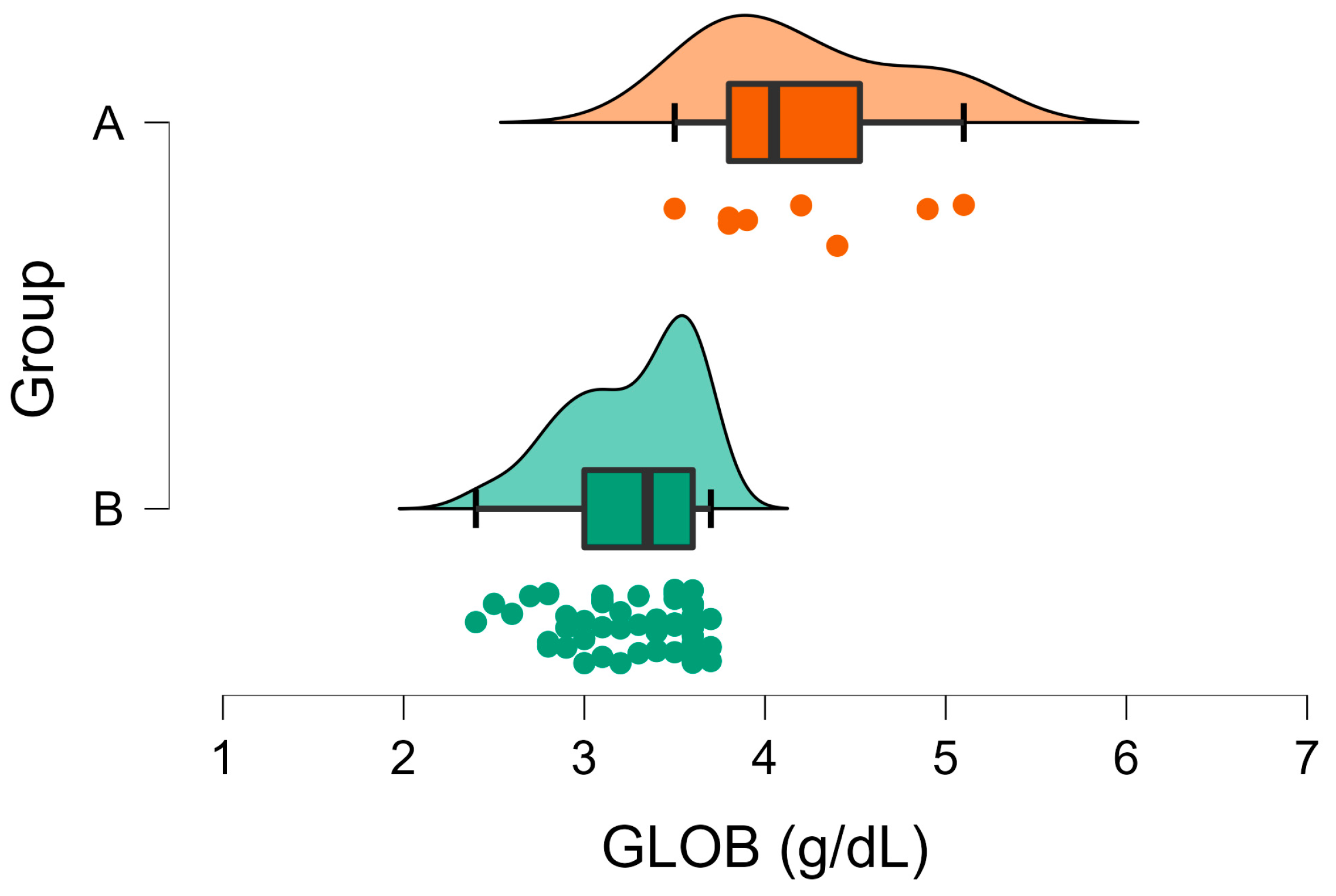
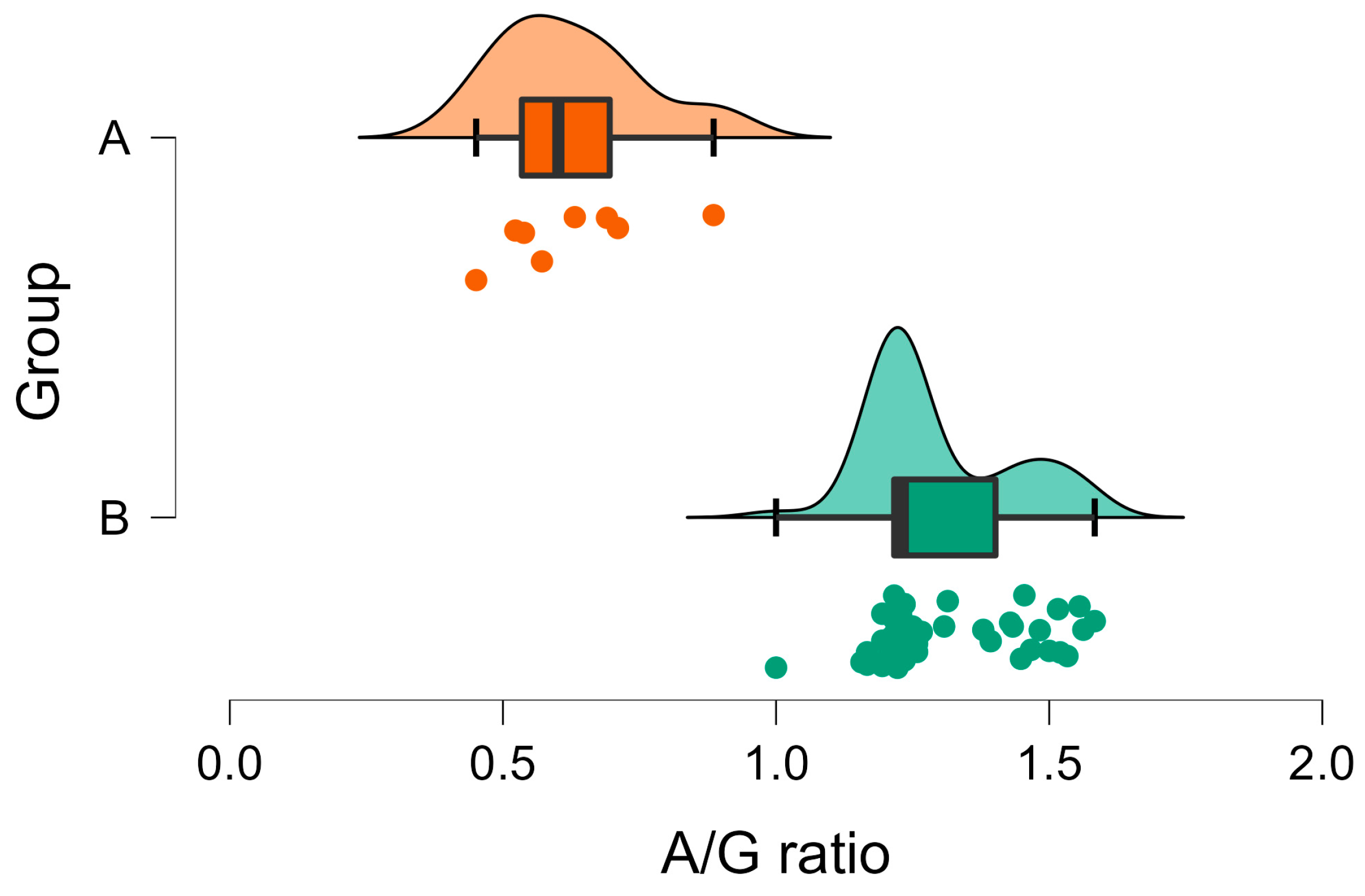
3. Results
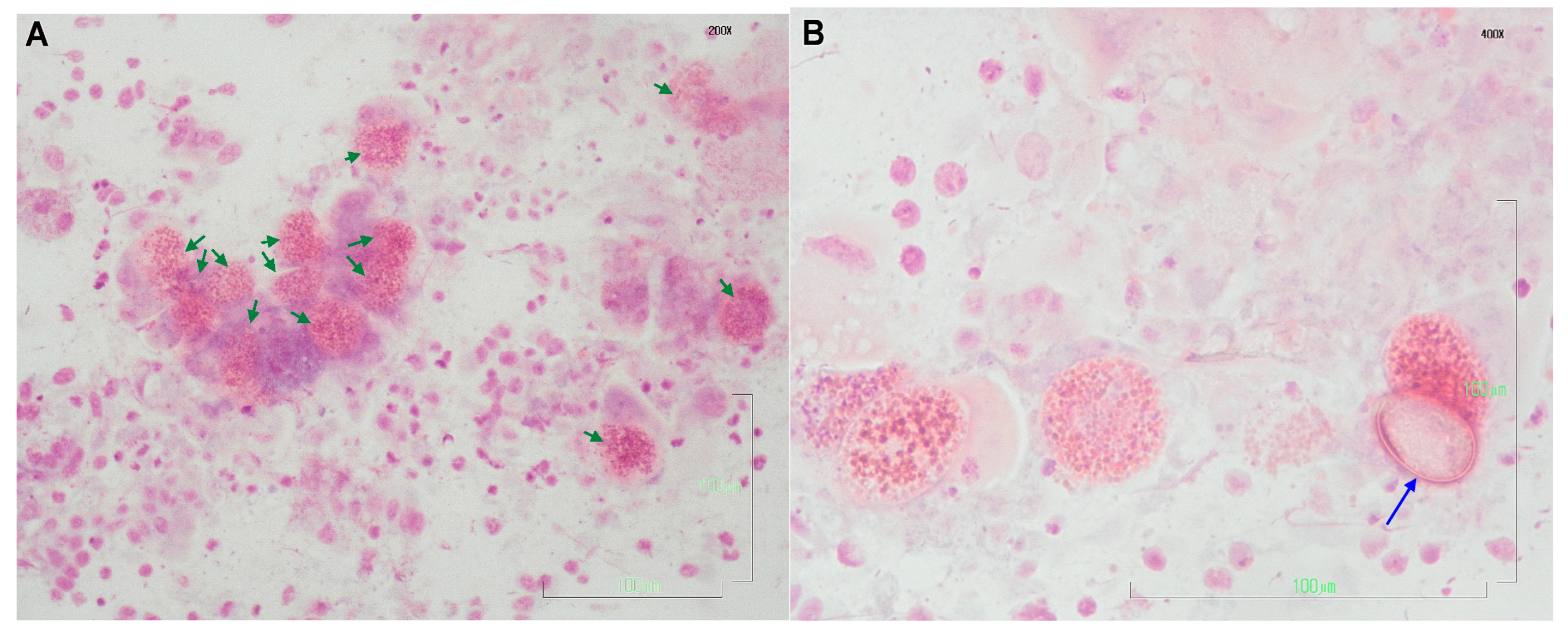
3.1. Liver Imprints
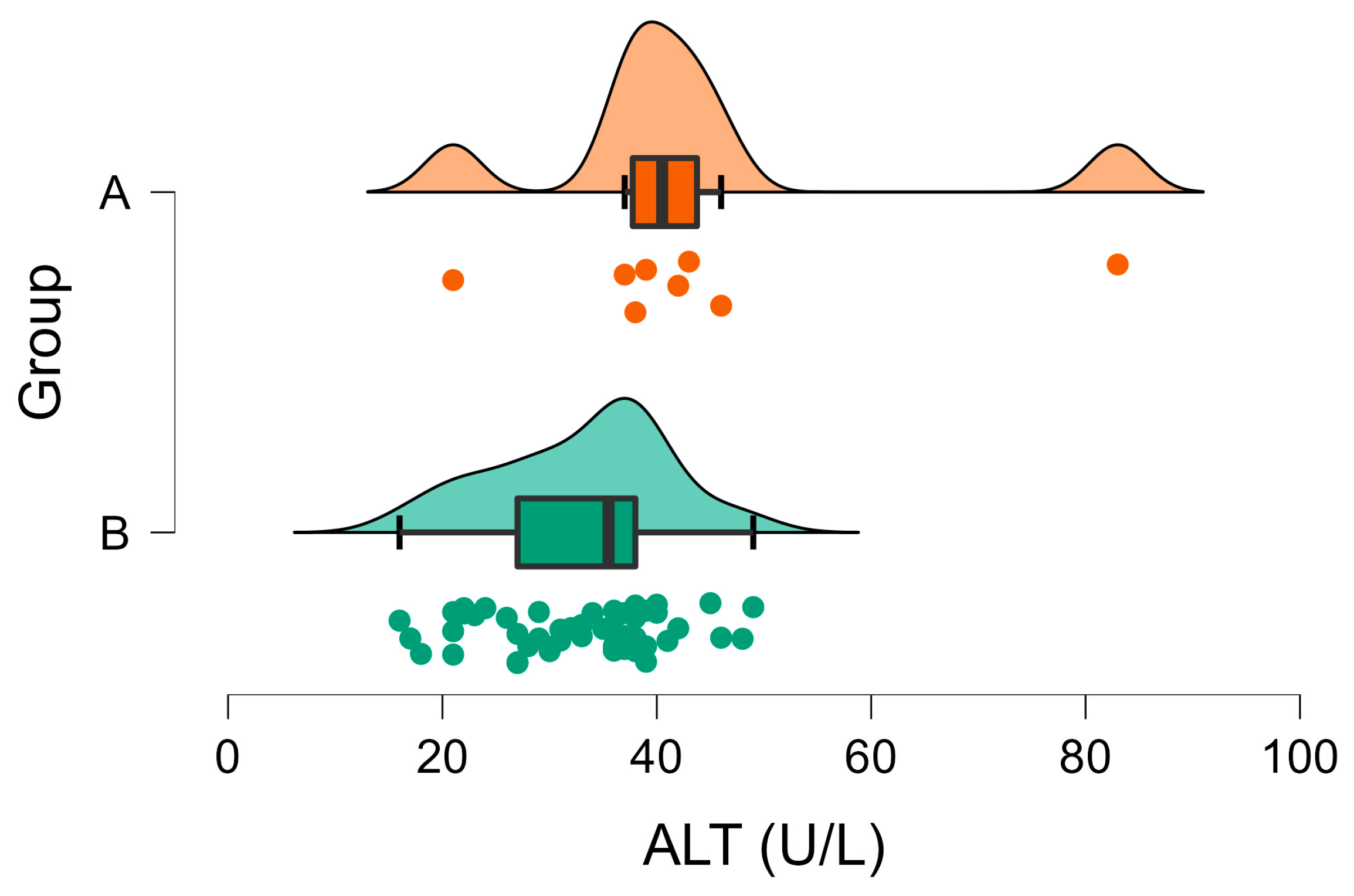
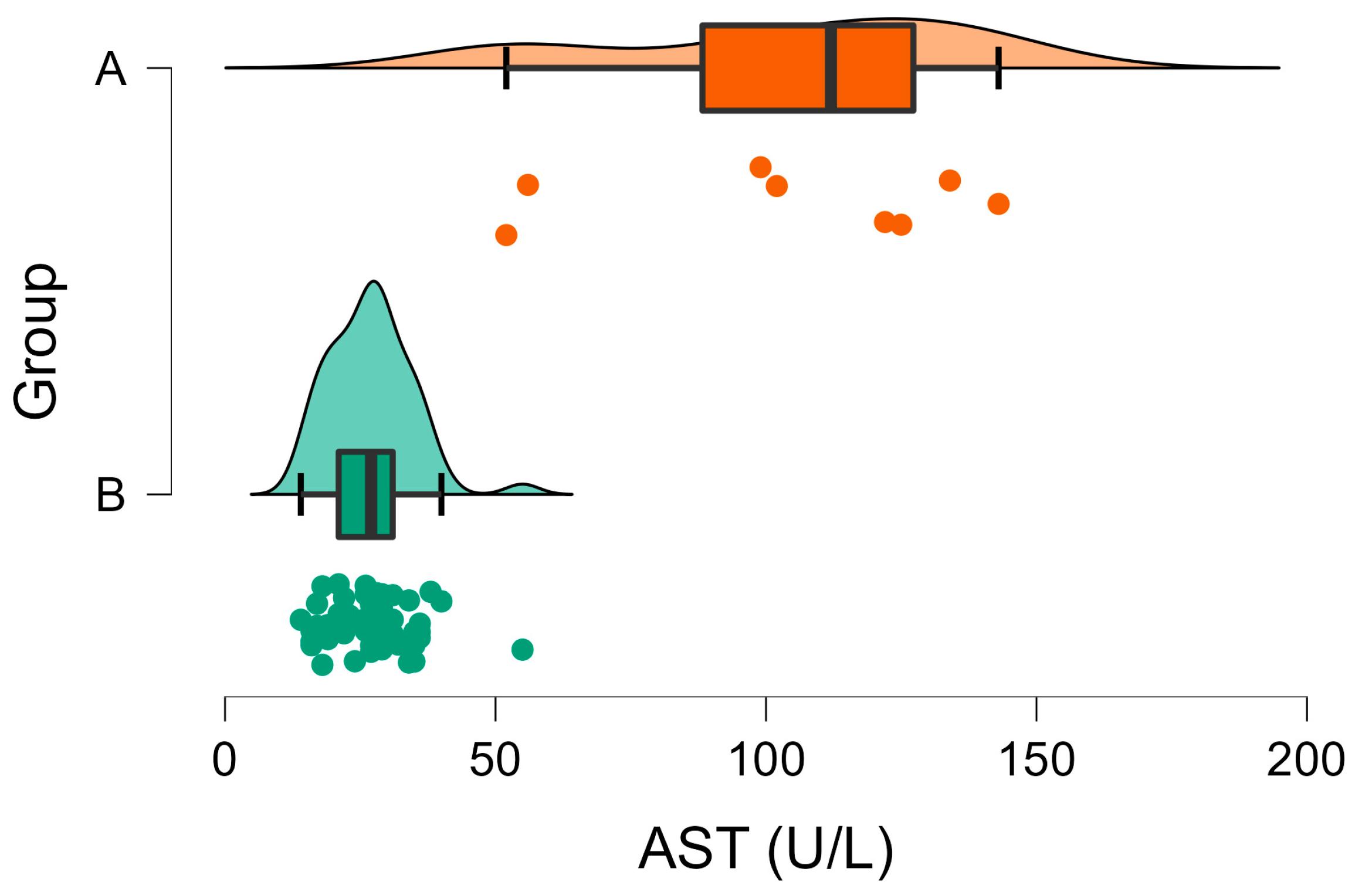
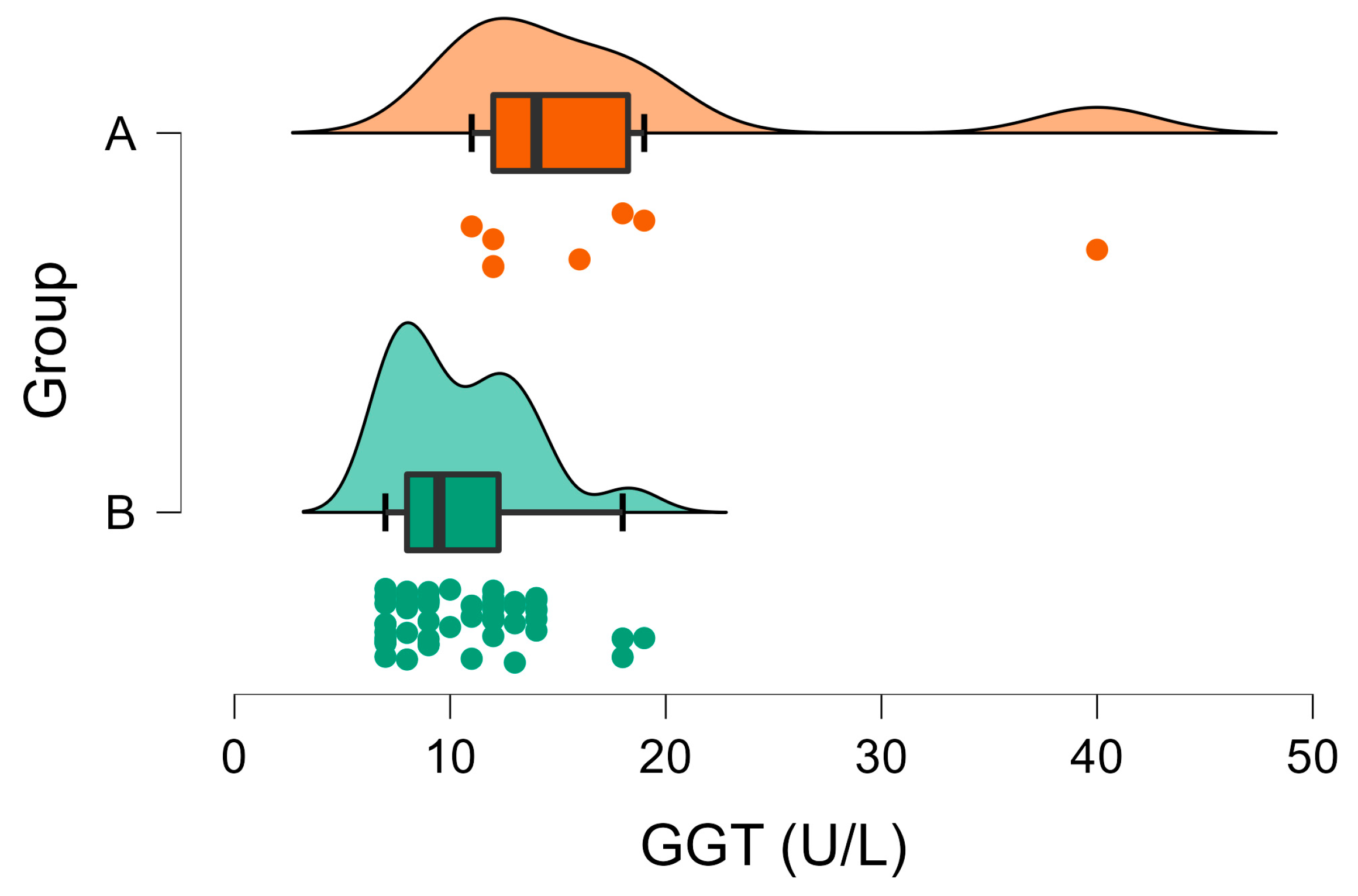
3.2. Biochemical Analyses
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Coudert, P.; Licois, D.; Drouet-Viard, F. Eimeria species and strains of rabbit. In Guidelines on Techniques in Coccidiosis Research: COST 89/820-Biotechnology; Office for Official Publications of the European Communities: Luxembourg, 1995; pp. 52–72. ISBN 978-92-827-4970-8. [Google Scholar]
- Pakandl, M. Coccidia of Rabbit: A Review. Folia Parasit. 2009, 56, 153–166. [Google Scholar] [CrossRef] [PubMed]
- Sivajothi, S.; Reddy, B.S.; Rayulu, V.C. Study on Impression Smears of Hepatic Coccidiosis in Rabbits. J. Parasit. Dis. 2016, 40, 906–909. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jing, J.; Liu, C.; Zhu, S.-X.; Jiang, Y.-M.; Wu, L.-C.; Song, H.-Y.; Shao, Y.-X. Pathological and Ultrastructural Observations and Liver Function Analysis of Eimeria stiedae-Infected Rabbits. Vet. Parasitol. 2016, 223, 165–172. [Google Scholar] [CrossRef] [PubMed]
- Okumu, P.O.; Gathumbi, P.K.; Karanja, D.N.; Mande, J.D.; Wanyoike, M.M.; Gachuiri, C.K.; Kiarie, N.; Mwanza, R.N.; Borter, D.K. Prevalence, Pathology and Risk Factors for Coccidiosis in Domestic Rabbits (Oryctolagus cuniculus ) in Selected Regions in Kenya. Vet. Q. 2014, 34, 205–210. [Google Scholar] [CrossRef]
- Owen, D. Life Cycle of Eimeria stiedae. Nature 1970, 227, 304. [Google Scholar] [CrossRef]
- Horton, R.J. The Route of Migration of Eimeria stiedae (Lindemann, 1865) Sporozoites between the Duodenum and Bile Ducts of the Rabbit. Parasitology 1967, 57, 9–17. [Google Scholar] [CrossRef]
- Çam, Y.; Atasever, A.; Eraslan, G.; Kibar, M.; Atalay, Ö.; Beyaz, L.; İnci, A.; Liman, B.C. Eimeria stiedae: Experimental Infection in Rabbits and the Effect of Treatment with Toltrazuril and Ivermectin. Exp. Parasitol. 2008, 119, 164–172. [Google Scholar] [CrossRef]
- Bochyńska, D.; Lloyd, S.; Restif, O.; Hughes, K. Eimeria stiedae Causes Most of the White-Spotted Liver Lesions in Wild European Rabbits in Cambridgeshire, United Kingdom. J. VET. Diagn. Invest. 2022, 34, 199–205. [Google Scholar] [CrossRef]
- Barriga, O.O.; Arnoni, J.V. Eimeria stiedae: Weight, Oocyst Output, and Hepatic Function of Rabbits with Graded Infections. Exp. Parasitol. 1979, 48, 407–414. [Google Scholar] [CrossRef]
- Yakhchali, M.; Tehrani, A. Eimeriidosis and Pathological Findings in New Zealand White Rabbits. J. Biol. Sci. 2007, 7, 1488–1491. [Google Scholar] [CrossRef] [Green Version]
- Silva, S.M.; Ferreira, C.; Paupério, J.; Silva, R.M.; Alves, P.C.; Lemos, A. Coccidiosis in European Rabbit (Oryctolagus cuniculus Algirus) Populations in the Iberian Peninsula. Acta Parasitol. 2015, 60, 350–355. [Google Scholar] [CrossRef] [PubMed]
- Sorour, S.S.; Abou Asa, S.; Elhawary, N.M.; Ghazy, E.W.; Abd El Latif, A.; El-Abasy, M.A.; Khalifa, H.O. Anticoccidial and Hepatoprotective Effects of Artemisinin Liquid Extract, Cinnamon Essential Oil and Clove Essential Oil against Eimeria stiedae Infection in Rabbits. Trop. Biomed. 2018, 35, 926–943. [Google Scholar] [PubMed]
- Petrova, J.P.; Iliev, P.T.; Ivanov, A.I.; Petrov, V.S.; Kalkanov, I.I.; Kostadinov, N.N.; Ceciliani, F.; Vitiello, T.; Giordano, A.; Georgieva, T.M. Biochemical and Pathomorphological Investigations on Rabbits with Experimentally Induced Hepatic Eimeriosis. Maced. Vet. Rev. 2022, 45, 53–59. [Google Scholar] [CrossRef]
- Aboelhadid, S.M.; El-Ashram, S.; Hassan, K.M.; Arafa, W.M.; Darwish, A.B. Hepato-Protective Effect of Curcumin and Silymarin against Eimeria stiedae in Experimentally Infected Rabbits. Livest. Sci. 2019, 221, 33–38. [Google Scholar] [CrossRef]
- Abd El-Ghany, W.A. Coccidiosis: A Parasitic Disease of Significant Importance in Rabbits. WVJ 2020, 10, 499–507. [Google Scholar] [CrossRef]
- Ball, S.; Pittilo, M.; Snow, K. Observations on Oocyst Development of Eimeria stiedae in Rabbits. Acta Parasitol. 2014, 59, 544–547. [Google Scholar] [CrossRef]
- Wei, W.; Shen, N.; Xiao, J.; Tao, Y.; Luo, Y.; Angel, C.; Gu, X.; Xie, Y.; He, R.; Jing, B.; et al. Expression Analysis and Serodiagnostic Potential of Microneme Proteins 1 and 3 in Eimeria stiedae. Genes 2020, 11, 725. [Google Scholar] [CrossRef]
- Onaga, H.; Saeki, H.; Hoshi, S.; Ueda, S. An Enzyme-Linked Immunosorbent Assay for Serodiagnosis of Coccidiosis in Chickens: Use of a Single Serum Dilution. Avian Dis. 1986, 30, 658–661. [Google Scholar] [CrossRef]
- Hassan, K.M.; Arafa, W.M.; Mousa, W.M.; Shokier, K.A.M.; Shany, S.A.; Aboelhadid, S.M. Molecular Diagnosis of Eimeria stiedae in Hepatic Tissue of Experimentally Infected Rabbits. Exp. Parasitol. 2016, 169, 1–5. [Google Scholar] [CrossRef]
- Yan, W.; Wang, W.; Wang, T.; Suo, X.; Qian, W.; Wang, S.; Fan, D. Simultaneous Identification of Three Highly Pathogenic Eimeria species in Rabbits Using a Multiplex PCR Diagnostic Assay Based on ITS1-5.8S RRNA-ITS2 Fragments. Vet. Parasitol. 2013, 193, 284–288. [Google Scholar] [CrossRef]
- Al-Rukibat, R.K.; Irizarry, A.R.; Lacey, J.K.; Kazacos, K.R.; Storandt, S.T.; DeNicola, D.B. Impression Smear of Liver Tissue from a Rabbit. Vet. Clin. Pathol. 2001, 30, 57–61. [Google Scholar] [CrossRef]
- Razavi, S.M.; Oryan, A.; Rakhshandehroo, E.; Moshiri, A.; Mootabi Alavi, A. Eimeria species in Wild Rabbits (Oryctolagus cuniculus) in Fars Province, Iran. Trop. Biomed. 2010, 27, 470–475. [Google Scholar] [PubMed]
- Mykytowycz, R. Epidemiology of Coccidiosis (Eimeria spp.) in an Experimental Population of the Australian Wild Rabbit, Oryctolagus cuniculus (L.). Parasitology 1962, 52, 375–395. [Google Scholar] [CrossRef]
- Ministerial. Decision HMEE/DFM 71082/539/2019 (GG3137/B/6-8-2019): Hunting Arrangements for the Hunting Season 2019–2020. Available online: https://dasarxeio.com/wp-content/uploads/2019/08/71082_539_2019.pdf (accessed on 10 February 2023).
- Katsoulos, P.D.; Athanasiou, L.V.; Karatzia, M.A.; Giadinis, N.; Karatzias, H.; Boscos, C.; Polizopoulou, Z.S. Comparison of Biuret and Refractometry Methods for the Serum Total Proteins Measurement in Ruminants. Vet. Clin. Pathol. 2017, 46, 620–624. [Google Scholar] [CrossRef] [PubMed]
- Allen, M.; Poggiali, D.; Whitaker, K.; Marshall, T.R.; van Langen, J.; Kievit, R.A. Raincloud Plots: A Multi-Platform Tool for Robust Data Visualization. Wellcome Open Res. 2021, 4, 63. [Google Scholar] [CrossRef]
- Jensen, A.L.; Kjelgaard-Hansen, M. Method Comparison in the Clinical Laboratory. Vet. Clin. Pathol. 2006, 35, 276–286. [Google Scholar] [CrossRef]
- Passing, H.; Bablok, W. A New Biometrical Procedure for Testing the Equality of Measurements from Two Different Analytical Methods. Application of Linear Regression Procedures for Method Comparison Studies in Clinical Chemistry, Part I. J. Clin. Chem. Clin. Biochem. 1983, 21, 709–720. [Google Scholar] [CrossRef] [Green Version]
- Passing, H.; Bablok, W. Comparison of Several Regression Procedures for Method Comparison Studies and Determination of Sample Sizes Application of Linear Regression Procedures for Method Comparison Studies in Clinical Chemistry, Part II. Clin. Chem. Lab. Med. 1984, 22, 431–445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hobbs, R.; Twigg, L. Coccidia (Eimeria spp.) of Wild Rabbits in Southwestern Australia. Aust. Vet. J. 1998, 76, 209–210. [Google Scholar] [CrossRef]
- Grès, V.; Voza, T.; Chabaud, A.; Landau, I. Coccidiosis of the Wild Rabbit (Oryctolagus cuniculus) in France. Parasite 2003, 10, 51–57. [Google Scholar] [CrossRef] [Green Version]
- Mason, S.; Dubey, J.P.; Smith, J.E.; Boag, B. Toxoplasma gondii Coinfection with Diseases and Parasites in Wild Rabbits in Scotland. Parasitology 2015, 142, 1415–1421. [Google Scholar] [CrossRef]
- Jing, F.; Yin, G.; Liu, X.; Suo, X.; Qin, Y. Large-Scale Survey of the Prevalence of Eimeria Infections in Domestic Rabbits in China. Parasitol. Res. 2012, 110, 1495–1500. [Google Scholar] [CrossRef]
- Yin, G.; Goraya, M.U.; Huang, J.; Suo, X.; Huang, Z.; Liu, X. Survey of Coccidial Infection of Rabbits in Sichuan Province, Southwest China. SpringerPlus 2016, 5, 870. [Google Scholar] [CrossRef] [Green Version]
- Katsui, K.; Takami, S.; Ohashi, K.; Otsuka, H.; Uni, S.; Shibahara, T.; Sasai, K.; Matsubayashi, M. Molecular Identification of Eimeria species in Liver and Feces of Naturally Infected Rabbits in Japan. Parasitol. Res. 2022, 121, 2733–2738. [Google Scholar] [CrossRef] [PubMed]
- Balicka-Ramisz, A.; Grupiński, T.; Ramisz, A.; Pilarczyk, B.; Laurans, L. [Prevalence of Trichinella spp. in red foxes and wild boars in the northwestern part of Poland]. Dtsch. Tierarztl. Wochenschr. 2007, 114, 354–357. [Google Scholar]
- Raue, K.; Heuer, L.; Böhm, C.; Wolken, S.; Epe, C.; Strube, C. 10-Year Parasitological Examination Results (2003 to 2012) of Faecal Samples from Horses, Ruminants, Pigs, Dogs, Cats, Rabbits and Hedgehogs. Parasitol. Res. 2017, 116, 3315–3330. [Google Scholar] [CrossRef]
- Hajipour, N.; Zavarshani, M. Ectoparasites and Endoparasites of New Zealand White Rabbits from North West of Iran. Iran. J. Parasitol. 2020, 15, 266–271. [Google Scholar] [CrossRef] [PubMed]
- Hamid, P.H.; Prastowo, S.; Kristianingrum, Y.P. Intestinal and Hepatic Coccidiosis among Rabbits in Yogyakarta, Indonesia. Vet. World 2019, 12, 1256–1260. [Google Scholar] [CrossRef]
- Temim, S.; Ainbaziz, H.; Bachene, A.; Bachene, M.S. Prevalence of Rabbit Coccidia in Medea Province, Algeria. J. World’s Poult. Res. 2019, 9, 123–128. [Google Scholar] [CrossRef]
- Sursal, N.; Gokpinar, S.; Yildiz, K. Prevalence of Intestinal Parasites in Hamsters and Rabbits in Some Pet Shops of Turkey. Turk. J. Parasitol. 2014, 38, 102–105. [Google Scholar] [CrossRef]
- Sioutas, G.; Evangelou, K.; Vlachavas, A.; Papadopoulos, E. Deaths Due to Mixed Infections with Passalurus ambiguus, Eimeria spp. and Cyniclomyces guttulatus in an Industrial Rabbit Farm in Greece. Pathogens 2021, 10, 756. [Google Scholar] [CrossRef]
- Andreopoulou, M.; Chaligiannis, I.; Sotiraki, S.; Daugschies, A.; Bangoura, B. Prevalence and Molecular Detection of Eimeria species in Different Types of Poultry in Greece and Associated Risk Factors. Parasitol. Res. 2022, 121, 2051–2063. [Google Scholar] [CrossRef] [PubMed]
- Lianou, D.T.; Arsenopoulos, K.V.; Michael, C.K.; Papadopoulos, E.; Fthenakis, G.C. Protozoan Parasites in Adult Dairy Small Ruminants and Potential Predictors for Their Presence in Faecal Samples. Microorganisms 2022, 10, 1931. [Google Scholar] [CrossRef] [PubMed]
- Kalfa-Papaioannou, A.M.; Athanassopoulou-Raptopoulou, F. Incidence of Coccidiosis in Horse-Mackerel (Trachurus trachurus, T. mediterraneus, T. picturatus) and Sardines (Clupea pilchardus) from the North Aegean Sea. Zent. Für Veterinärmedizin Reihe B 2010, 31, 530–536. [Google Scholar] [CrossRef]
- Fumeaux, L.; Scarpelli, M.P.; Tettamanti, C.; Palmiere, C. Usefulness of Liver Function Tests in Postmortem Samples. J. Forensic Leg. Med. 2018, 56, 51–54. [Google Scholar] [CrossRef]
- Melillo, A. Rabbit Clinical Pathology. J. Exot. Pet Med. 2007, 16, 135–145. [Google Scholar] [CrossRef]
- Jenkins, J.R. Rabbit Diagnostic Testing. J. Exot. Pet Med. 2008, 17, 4–15. [Google Scholar] [CrossRef]
- Tsai, H.-Y.; Chueh, L.-L.; Lin, C.-N.; Su, B.-L. Clinicopathological Findings and Disease Staging of Feline Infectious Peritonitis: 51 Cases from 2003 to 2009 in Taiwan. J. Feline Med. Surg. 2011, 13, 74–80. [Google Scholar] [CrossRef] [PubMed]
- Cray, C.; Zaias, J.; Altman, N.H. Acute Phase Response in Animals: A Review. Comp. Med. 2009, 59, 517–526. [Google Scholar]
- Gomez-Bautista, M.; Garcia, M.V.; Rojo-Vazquez, F.A. The Levels of Total Protein and Protein Fractions in the Serum of Rabbits Infected with Eimeria stiedai. Ann. Parasitol. Hum. Comp. 1986, 61, 393–400. [Google Scholar] [CrossRef] [Green Version]
- Martinez-Sanchez, L.; Marques-Garcia, F.; Ozarda, Y.; Blanco, A.; Brouwer, N.; Canalias, F.; Cobbaert, C.; Thelen, M.; den Elzen, W. Big Data and Reference Intervals: Rationale, Current Practices, Harmonization and Standardization Prerequisites and Future Perspectives of Indirect Determination of Reference Intervals Using Routine Data. Adv. Lab. Med./Av. Med. Lab. 2021, 2, 9–16. [Google Scholar] [CrossRef]
- Fraser, C.G. Biological Variation: From Principles to Practice; AACC Press: Washington, DC, USA, 2001; ISBN 978-1-890883-49-2. [Google Scholar]
- Meredith, A. Liver Disease in Rabbits. Practice 2013, 35, 291–301. [Google Scholar] [CrossRef]
- Kontsiotis, V.J.; Bakaloudis, D.E.; Xofis, P.; Konstantaras, N.; Petrakis, N.; Tsiompanoudis, A. Modeling the Distribution of Wild Rabbits (Oryctolagus cuniculus) on a Mediterranean Island. Ecol. Res. 2013, 28, 317–325. [Google Scholar] [CrossRef]
- Kontsiotis, V.J.; Bakaloudis, D.E.; Liordos, V. Impact of European Wild Rabbits Foraging in Different Habitat and Vegetation Types in an Insular Environment. Mammalia 2018, 82, 193–196. [Google Scholar] [CrossRef]
- Hadjigeorgiou, I.; Chatzigeorgiou, T.; Kosmas, C.; Kairis, O. Report on Lemnos Wild Rabbit Population; Agricultural University of Athens: Athens, Greece, 2018. [Google Scholar]
- Kontsiotis, V.J.; Bakaloudis, D.E.; Tsiompanoudis, A.C.; Xofis, P. Body Condition Variation of Wild Rabbit Population in the North-East Mediterranean Island of Lemnos—Greece. Folia Zool. 2014, 63, 87–94. [Google Scholar] [CrossRef]
- Vasios, G.K.; Antoniadis, I.; Zevgolis, Y.; Giaginis, C.; Troumbis, A.Y. Turning a Problem into an Opportunity Through Tourism and Marketing: The Case of Wild Rabbits in Lemnos Island, Greece. In Strategic Innovative Marketing and Tourism; Kavoura, A., Kefallonitis, E., Theodoridis, P., Eds.; Springer Proceedings in Business and Economics; Springer International Publishing: Cham, Switzerland, 2020; pp. 677–686. ISBN 978-3-030-36125-9. [Google Scholar]
- Athanasiou, L.V.; Katsogiannou, E.G.; Tsokana, C.N.; Boutsini, S.G.; Bisia, M.G.; Papatsiros, V.G. Wild Rabbit Exposure to Leishmania infantum, Toxoplasma gondii, Anaplasma phagocytophilum and Babesia caballi Evidenced by Serum and Aqueous Humor Antibody Detection. Microorganisms 2021, 9, 2616. [Google Scholar] [CrossRef] [PubMed]
- Tsakmakidis, Ι.; Pavlou, C.; Tamvakis, A.; Papadopoulos, T.; Christodoulou, V.; Angelopoulou, K.; Dovas, C.I.; Antoniou, Μ.; Anastasakis, C.; Diakou, A. Leishmania Infection in Lagomorphs and Minks in Greece. Vet. Parasitol. Reg. Stud. Rep. 2019, 16, 100279. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Biochemical Variable | Group | Mean | SE | p-Value | RI |
|---|---|---|---|---|---|
| ALT (U/L) | A | 32.90 | 1.11 | 0.005 | 31–53 |
| B | 43.63 | 6.22 | |||
| AST (U/L) | A | 26.94 | 1.06 | <001 | 42–98 |
| B | 104.13 | 12.12 | |||
| GGT (U/L) | A | 10.50 | 0.43 | <001 | 0–7 |
| B | 17.50 | 3.39 | |||
| TP (g/dL) | A | 7.44 | 0.08 | 0.003 | 5.5–7.2 |
| B | 6.78 | 0.20 | |||
| ALB (g/dL) | A | 4.19 | 0.04 | <001 | 2.7–4.6 |
| B | 2.58 | 0.12 | |||
| GLOB (g/dL) | A | 3.26 | 0.05 | <001 | 1.5–2.8 |
| B | 4.20 | 0.20 | |||
| A/G ratio | A | 1.30 | 0.02 | <001 | N.A. |
| B | 0.63 | 0.05 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Athanasiou, L.V.; Tsokana, C.N.; Doukas, D.; Kantere, M.C.; Katsoulos, P.D.; Papakonstantinou, G.I.; Katsogiannou, E.G.; Dedousi, A. Hepatic Coccidiosis in Wild Rabbits in Greece: Parasite Detection on Liver Imprints and the Associated Biochemical Profile. Vet. Sci. 2023, 10, 248. https://doi.org/10.3390/vetsci10040248
Athanasiou LV, Tsokana CN, Doukas D, Kantere MC, Katsoulos PD, Papakonstantinou GI, Katsogiannou EG, Dedousi A. Hepatic Coccidiosis in Wild Rabbits in Greece: Parasite Detection on Liver Imprints and the Associated Biochemical Profile. Veterinary Sciences. 2023; 10(4):248. https://doi.org/10.3390/vetsci10040248
Chicago/Turabian StyleAthanasiou, Labrini V., Constantina N. Tsokana, Dimitrios Doukas, Maria C. Kantere, Panagiotis D. Katsoulos, Georgios I. Papakonstantinou, Eleni G. Katsogiannou, and Anna Dedousi. 2023. "Hepatic Coccidiosis in Wild Rabbits in Greece: Parasite Detection on Liver Imprints and the Associated Biochemical Profile" Veterinary Sciences 10, no. 4: 248. https://doi.org/10.3390/vetsci10040248