
Multi-Tissue Transcriptome Study of Innate Immune Gene Expression Profiling Reveals Negative Energy Balance Altered the Defense and Promoted System Inflammation of Dairy Cows

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. RNA Sequencing Data and Data Grouping Information
- RNA Sequencing Data Sources
- Data grouping information
2.2. Reads Mapping and Quantification of Gene Expression
2.3. Innate Immune Gene Sets and Principal Component Analysis
2.4. Gene-Set Enrichment Analysis and Network Architecture
3. Results
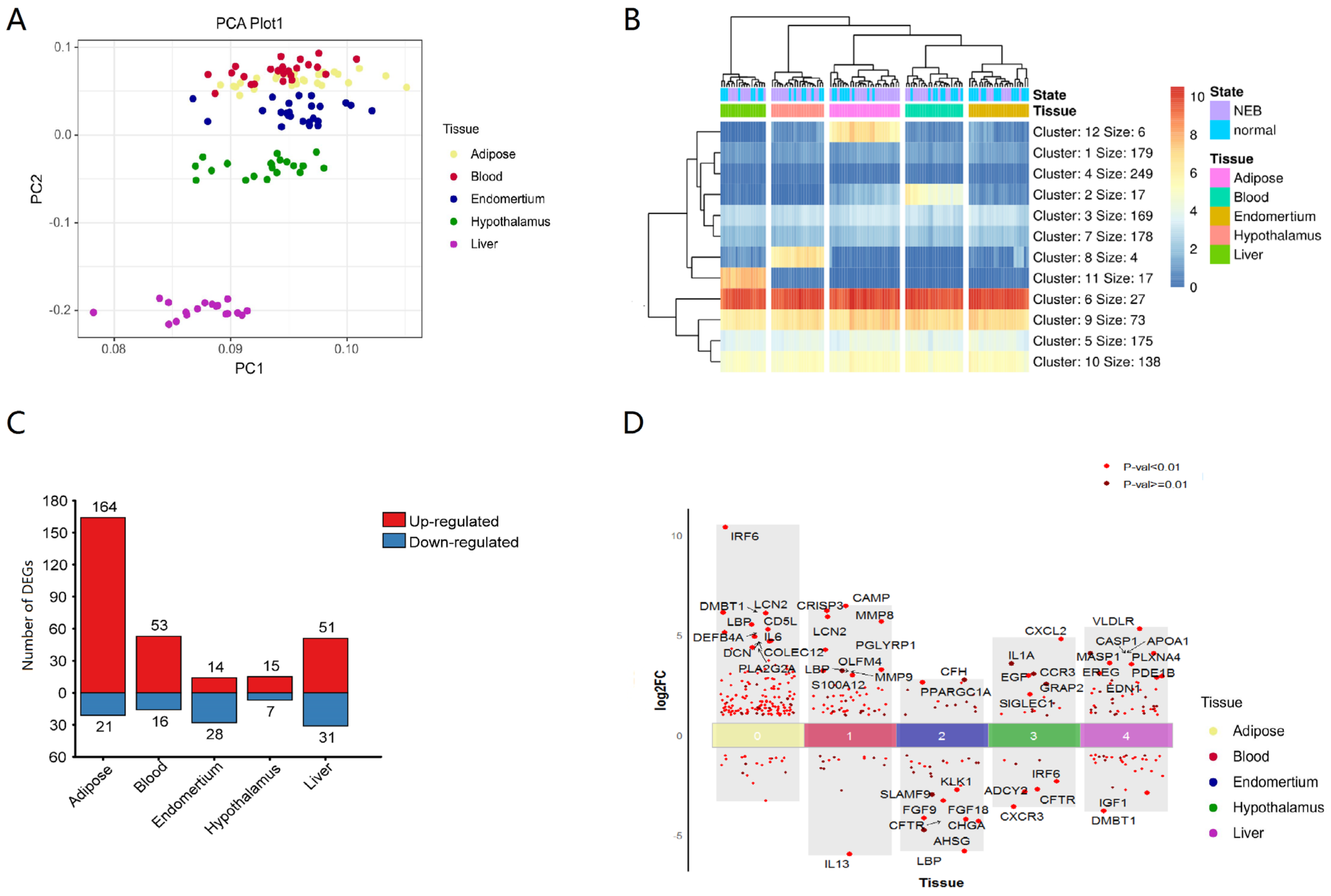
3.1. Expression of Innate Immune Genes in Different Tissues of Dairy Cows
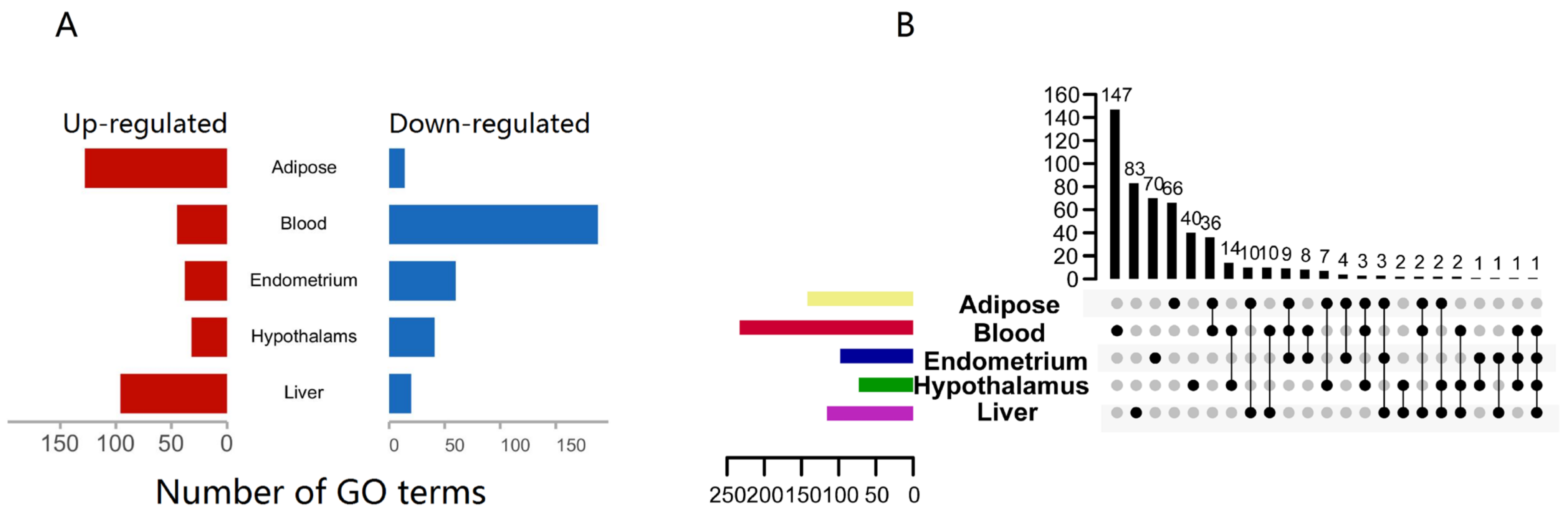
3.2. Functional Analysis Reveals Widespread Alterations of Biological Processes of Innate Immune Gene Post NEB Condition with Different Tissue
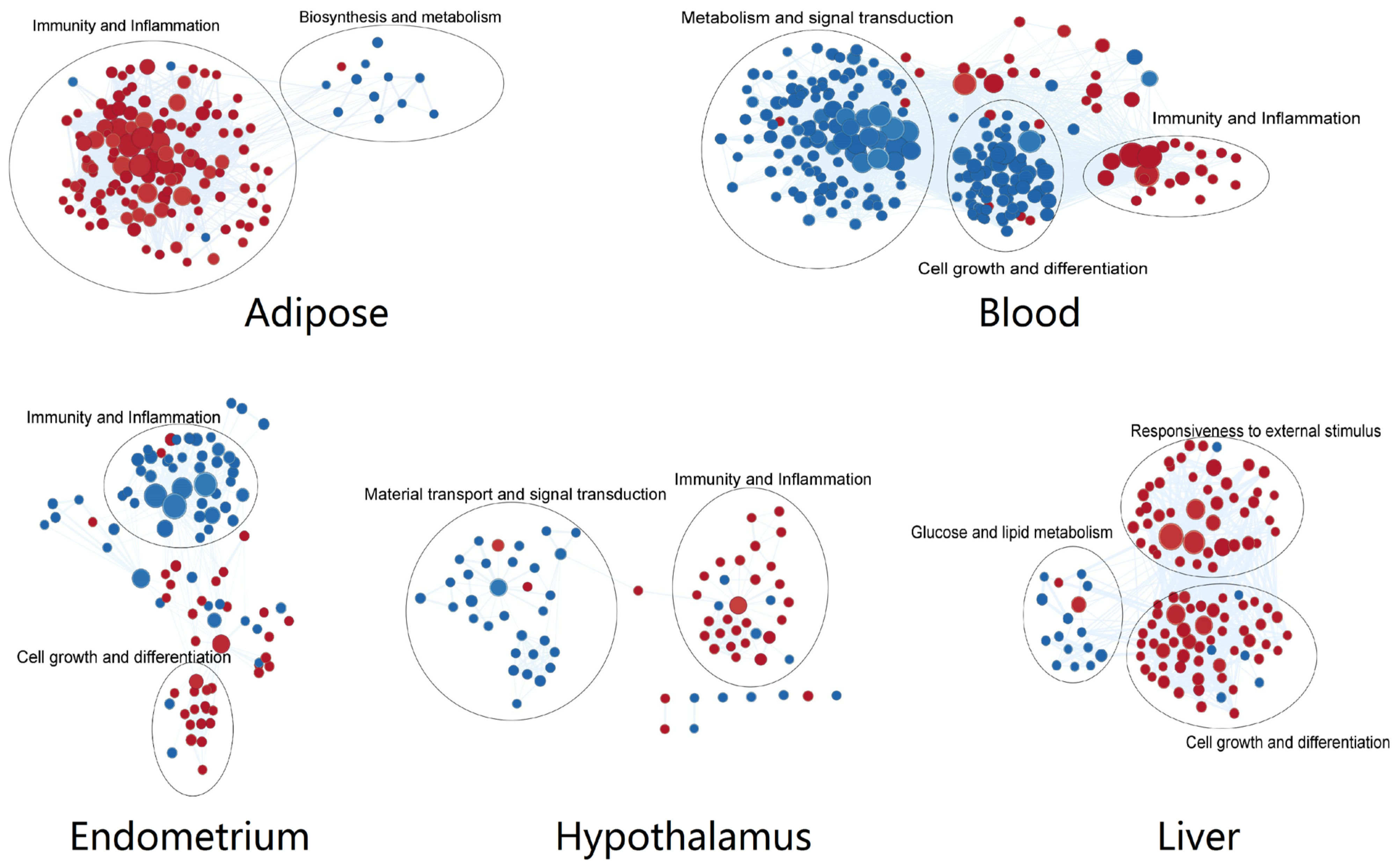
3.3. Tissue-Specific Altered Biological Functions Point to Specificity of Defensive, Metabolic, and Signaling Responses to NEB
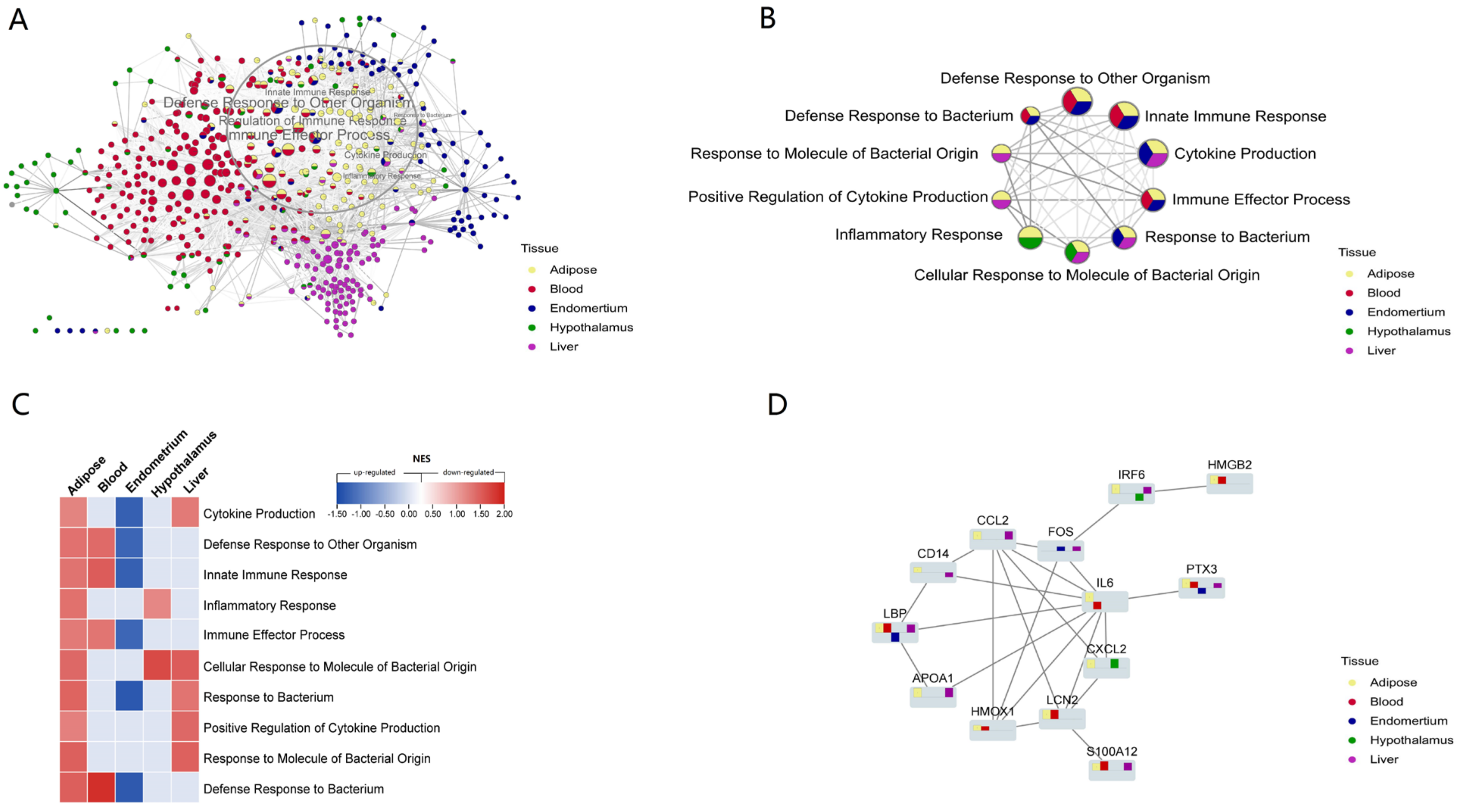
3.4. Network Analyses of Gene Sets from Multi-Tissue Unveils Universal Changes to the Defense Response Caused by NEB
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wankhade, P.R.; Manimaran, A.; Kumaresan, A.; Jeyakumar, S.; Ramesha, K.P.; Sejian, V.; Rajendran, D.; Varghese, M.R. Metabolic and immunological changes in transition dairy cows: A review. Vet. World 2017, 10, 1367–1377. [Google Scholar] [CrossRef] [PubMed]
- Ingvartsen, K.L.; Moyes, K. Nutrition, immune function and health of dairy cattle. Animal 2013, 7 (Suppl. S1), 112–122. [Google Scholar] [CrossRef] [PubMed]
- Fiore, E.; Gianesella, M.; Arfuso, F.; Giudice, E.; Piccione, G.; Lora, M.; Stefani, A.; Morgante, M. Glucose infusion response on some metabolic parameters in dairy cows during transition period. Arch. Anim. Breed. 2014, 57, 3. [Google Scholar] [CrossRef]
- Fiore, E.; Arfuso, F.; Gianesella, M.; Vecchio, D.; Morgante, M.; Mazzotta, E.; Badon, T.; Rossi, P.; Bedin, S.; Piccione, G. Metabolic and hormonal adaptation in Bubalus bubalis around calving and early lactation. PLoS ONE 2018, 13, e0193803. [Google Scholar] [CrossRef] [PubMed]
- Arfuso, F.; Minuti, A.; Liotta, L.; Giannetto, C.; Trevisi, E.; Piccione, G.; Lopreiato, V. Stress and inflammatory response of cows and their calves during peripartum and early neonatal period. Theriogenology 2023, 196, 157–166. [Google Scholar] [CrossRef]
- Gordon, J.L.; Leblanc, S.J.; Duffield, T.F. Ketosis treatment in lactating dairy cattle. Vet. Clin. N. Am. Food Anim. Pract. 2013, 29, 433–445. [Google Scholar] [CrossRef]
- Vanholder, T.; Papen, J.; Bemers, R.; Vertenten, G.; Berge, A.C. Risk factors for subclinical and clinical ketosis and association with production parameters in dairy cows in the Netherlands. J. Dairy Sci. 2015, 98, 880–888. [Google Scholar] [CrossRef]
- Suthar, V.S.; Canelas-Raposo, J.; Deniz, A.; Heuwieser, W. Prevalence of subclinical ketosis and relationships with postpartum diseases in European dairy cows. J. Dairy Sci. 2013, 96, 2925–2938. [Google Scholar] [CrossRef]
- Overton, T.R.; McArt, J.A.A.; Nydam, D.V. A 100-Year Review: Metabolic health indicators and management of dairy cattle. J. Dairy Sci. 2017, 100, 10398–10417. [Google Scholar] [CrossRef]
- Ringseis, R.; Gessner, D.K.; Eder, K. Molecular insights into the mechanisms of liver-associated diseases in early-lactating dairy cows: Hypothetical role of endoplasmic reticulum stress. J. Anim. Physiol. Anim. Nutr. 2015, 99, 626–645. [Google Scholar] [CrossRef]
- Morris, D.G.; Waters, S.M.; McCarthy, S.D.; Patton, J.; Earley, B.; Fitzpatrick, R.; Murphy, J.J.; Diskin, M.G.; Kenny, D.A.; Brass, A.; et al. Pleiotropic effects of negative energy balance in the postpartum dairy cow on splenic gene expression: Repercussions for innate and adaptive immunity. Physiol. Genom. 2009, 39, 28–37. [Google Scholar] [CrossRef] [PubMed]
- LeBlanc, S.J.; Osawa, T.; Dubuc, J. Reproductive tract defense and disease in postpartum dairy cows. Theriogenology 2011, 76, 1610–1618. [Google Scholar] [CrossRef] [PubMed]
- Yasui, T.; McCann, K.; Gilbert, R.O.; Nydam, D.V.; Overton, T.R. Associations of cytological endometritis with energy metabolism and inflammation during the periparturient period and early lactation in dairy cows. J. Dairy Sci. 2014, 97, 2763–2770. [Google Scholar] [CrossRef] [PubMed]
- Kuhla, B. Review: Pro-inflammatory cytokines and hypothalamic inflammation: Implications for insufficient feed intake of transition dairy cows. Animal 2020, 14, s65–s77. [Google Scholar] [CrossRef]
- Stanley, S.; Wynne, K.; McGowan, B.; Bloom, S. Hormonal regulation of food intake. Physiol. Rev. 2005, 85, 1131–1158. [Google Scholar] [CrossRef]
- Ferguson, J.D. Nutrition and reproduction in dairy herds. Vet. Clin. N. Am.-Food A 2005, 21, 325. [Google Scholar] [CrossRef]
- Dervishi, E.; Zhang, G.; Hailemariam, D.; Dunn, S.M.; Ametaj, B.N. Innate immunity and carbohydrate metabolism alterations precede occurrence of subclinical mastitis in transition dairy cows. J. Anim. Sci. Technol. 2015, 57, 46. [Google Scholar] [CrossRef]
- Esposito, G.; Irons, P.C.; Webb, E.C.; Chapwanya, A. Interactions between negative energy balance, metabolic diseases, uterine health and immune response in transition dairy cows. Anim. Reprod. Sci. 2014, 144, 60–71. [Google Scholar] [CrossRef]
- Trevisi, E.; Minuti, A. Assessment of the innate immune response in the periparturient cow. Res. Vet. Sci. 2018, 116, 47–54. [Google Scholar] [CrossRef]
- Salcedo-Tacuma, D.; Parales-Giron, J.; Prom, C.; Chirivi, M.; Laguna, J.; Lock, A.L.; Contreras, G.A. Transcriptomic profiling of adipose tissue inflammation, remodeling, and lipid metabolism in periparturient dairy cows (Bos taurus). BMC Genom. 2020, 21, 824. [Google Scholar] [CrossRef]
- Yan, Z.; Huang, H.; Freebern, E.; Santos, D.J.A.; Dai, D.; Si, J.; Ma, C.; Cao, J.; Guo, G.; Liu, G.E.; et al. Integrating RNA-Seq with GWAS reveals novel insights into the molecular mechanism underpinning ketosis in cattle. BMC Genom. 2020, 21, 489. [Google Scholar] [CrossRef] [PubMed]
- Chankeaw, W.; Lignier, S.; Richard, C.; Ntallaris, T.; Raliou, M.; Guo, Y.; Plassard, D.; Bevilacqua, C.; Sandra, O.; Andersson, G.; et al. Analysis of the transcriptome of bovine endometrial cells isolated by laser micro-dissection (2): Impacts of post-partum negative energy balance on stromal, glandular and luminal epithelial cells. BMC Genom. 2021, 22, 450. [Google Scholar] [CrossRef] [PubMed]
- Matthews, D.; Diskin, M.G.; Kenny, D.A.; Creevey, C.J.; Keogh, K.; Waters, S.M. Effect of short term diet restriction on gene expression in the bovine hypothalamus using next generation RNA sequencing technology. BMC Genom. 2017, 18, 857. [Google Scholar] [CrossRef]
- McCabe, M.; Waters, S.; Morris, D.; Kenny, D.; Lynn, D.; Creevey, C. RNA-seq analysis of differential gene expression in liver from lactating dairy cows divergent in negative energy balance. BMC Genom. 2012, 13, 193. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef]
- Rosen, B.D.; Bickhart, D.M.; Schnabel, R.D.; Koren, S.; Elsik, C.G.; Tseng, E.; Rowan, T.N.; Low, W.Y.; Zimin, A.; Couldrey, C.; et al. De novo assembly of the cattle reference genome with single-molecule sequencing. Gigascience 2020, 9, giaa021. [Google Scholar] [CrossRef]
- Howe, K.L.; Achuthan, P.; Allen, J.; Allen, J.; Alvarez-Jarreta, J.; Amode, M.R.; Armean, I.M.; Azov, A.G.; Bennett, R.; Bhai, J.; et al. Ensembl 2021. Nucleic Acids Res. 2021, 49, D884–D891. [Google Scholar] [CrossRef]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef]
- Pertea, M.; Pertea, G.M.; Antonescu, C.M.; Chang, T.C.; Mendell, J.T.; Salzberg, S.L. StringTie enables improved reconstruction of a transcriptome from RNA-seq reads. Nat. Biotechnol. 2015, 33, 290–295. [Google Scholar] [CrossRef]
- Leek, J.T.; Johnson, W.E.; Parker, H.S.; Jaffe, A.E.; Storey, J.D. The sva package for removing batch effects and other unwanted variation in high-throughput experiments. Bioinformatics 2012, 28, 882–883. [Google Scholar] [CrossRef] [Green Version]
- Lawrence, M.; Huber, W.; Pages, H.; Aboyoun, P.; Carlson, M.; Gentleman, R.; Morgan, M.T.; Carey, V.J. Software for computing and annotating genomic ranges. PLoS Comput. Biol. 2013, 9, e1003118. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Emms, D.M.; Kelly, S. OrthoFinder: Phylogenetic orthology inference for comparative genomics. Genome Biol. 2019, 20, 238. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Miao, B.; Wang, S.; Dong, W.; Xu, H.; Si, C.; Wang, W.; Duan, S.; Lou, J.; Bao, Z.; et al. Hiplot: A comprehensive and easy-to-use web service for boosting publication-ready biomedical data visualization. Brief Bioinform. 2022, 23, bbac261. [Google Scholar] [CrossRef]
- Kolde, R. Pheatmap: Pretty Heatmaps. R Package. Version 1.0.12. 2019. Available online: https://CRAN.R-project.org/package=pheatmap (accessed on 4 January 2019).
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene set enrichment analysis: A knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef] [PubMed]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef] [PubMed]
- Merico, D.; Isserlin, R.; Stueker, O.; Emili, A.; Bader, G.D. Enrichment map: A network-based method for gene-set enrichment visualization and interpretation. PLoS ONE 2010, 5, e13984. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Li, X.; Wang, Q.; Zhang, M.; He, J.; Ming, S.; Wang, Z.; Cao, C.; Zhang, S.; Geng, L.; et al. TLT-1 Promotes Platelet-Monocyte Aggregate Formation to Induce IL-10-Producing B Cells in Tuberculosis. J. Immunol. 2022, 208, 1642–1651. [Google Scholar] [CrossRef]
- Derive, M.; Bouazza, Y.; Sennoun, N.; Marchionni, S.; Quigley, L.; Washington, V.; Massin, F.; Max, J.P.; Ford, J.; Alauzet, C.; et al. Soluble TREM-like transcript-1 regulates leukocyte activation and controls microbial sepsis. J. Immunol. 2012, 188, 5585–5592. [Google Scholar] [CrossRef]
- Harder, J.; Meyer-Hoffert, U.; Teran, L.M.; Schwichtenberg, L.; Bartels, J.; Maune, S.; Schroder, J.M. Mucoid Pseudomonas aeruginosa, TNF-alpha, and IL-1beta, but not IL-6, induce human beta-defensin-2 in respiratory epithelia. Am. J. Respir. Cell Mol. Biol. 2000, 22, 714–721. [Google Scholar] [CrossRef]
- Dahlstrand Rudin, A.; Khamzeh, A.; Venkatakrishnan, V.; Basic, A.; Christenson, K.; Bylund, J. Short chain fatty acids released by Fusobacterium nucleatum are neutrophil chemoattractants acting via free fatty acid receptor 2 (FFAR2). Cell. Microbiol. 2021, 23, e13348. [Google Scholar] [CrossRef] [PubMed]
- Degols, G.; Eldin, P.; Mechti, N. ISG20, an actor of the innate immune response. Biochimie 2007, 89, 831–835. [Google Scholar] [CrossRef] [PubMed]
- Unsworth, D.J. Complement deficiency and disease. J. Clin. Pathol. 2008, 61, 1013–1017. [Google Scholar] [CrossRef] [PubMed]
- Friedman, J.R.; Kaestner, K.H. The Foxa family of transcription factors in development and metabolism. Cell. Mol. Life Sci. 2006, 63, 2317–2328. [Google Scholar] [CrossRef] [PubMed]
- Alphonse, N.; Wanford, J.J.; Voak, A.A.; Gay, J.; Venkhaya, S.; Burroughs, O.; Mathew, S.; Lee, T.; Evans, S.L.; Zhao, W.; et al. A family of conserved bacterial virulence factors dampens interferon responses by blocking calcium signaling. Cell 2022, 185, 2354–2369.e17. [Google Scholar] [CrossRef]
- Nair, J.S.; DaFonseca, C.J.; Tjernberg, A.; Sun, W.; Darnell, J.E., Jr.; Chait, B.T.; Zhang, J.J. Requirement of Ca2+ and CaMKII for Stat1 Ser-727 phosphorylation in response to IFN-gamma. Proc. Natl. Acad. Sci. USA 2002, 99, 5971–5976. [Google Scholar] [CrossRef]
- Mahata, S.K.; Corti, A. Chromogranin A and its fragments in cardiovascular, immunometabolic, and cancer regulation. Ann. N. Y. Acad. Sci. 2019, 1455, 34–58. [Google Scholar] [CrossRef] [PubMed]
- Rotondo, F.; Butz, H.; Syro, L.V.; Yousef, G.M.; Di Ieva, A.; Restrepo, L.M.; Quintanar-Stephano, A.; Berczi, I.; Kovacs, K. Arginine vasopressin (AVP): A review of its historical perspectives, current research and multifunctional role in the hypothalamo-hypophysial system. Pituitary 2016, 19, 345–355. [Google Scholar] [CrossRef] [PubMed]
- Falix, F.A.; Aronson, D.C.; Lamers, W.H.; Gaemers, I.C. Possible roles of DLK1 in the Notch pathway during development and disease. Biochim. Biophys. Acta 2012, 1822, 988–995. [Google Scholar] [CrossRef]
- Dankel, S.N.; Fadnes, D.J.; Stavrum, A.K.; Stansberg, C.; Holdhus, R.; Hoang, T.; Veum, V.L.; Christensen, B.J.; Vage, V.; Sagen, J.V.; et al. Switch from stress response to homeobox transcription factors in adipose tissue after profound fat loss. PLoS ONE 2010, 5, e11033. [Google Scholar] [CrossRef] [Green Version]
- Hjort, L.; Jorgensen, S.W.; Gillberg, L.; Hall, E.; Brons, C.; Frystyk, J.; Vaag, A.A.; Ling, C. 36 h fasting of young men influences adipose tissue DNA methylation of LEP and ADIPOQ in a birth weight-dependent manner. Clin. Epigenetics 2017, 9, 40. [Google Scholar] [CrossRef] [PubMed]
- Ning, M.; Zhao, Y.; Li, Z.; Cao, J. Ketosis Alters Transcriptional Adaptations of Subcutaneous White Adipose Tissue in Holstein Cows during the Transition Period. Animals 2022, 12, 2238. [Google Scholar] [CrossRef]
- Wang, Y.H.; Liu, C.L.; Chiu, W.C.; Twu, Y.C.; Liao, Y.J. HMGCS2 Mediates Ketone Production and Regulates the Proliferation and Metastasis of Hepatocellular Carcinoma. Cancers 2019, 11, 1876. [Google Scholar] [CrossRef] [PubMed]
- Contreras, G.A.; Thelen, K.; Schmidt, S.E.; Strieder-Barboza, C.; Preseault, C.L.; Raphael, W.; Kiupel, M.; Caron, J.; Lock, A.L. Adipose tissue remodeling in late-lactation dairy cows during feed-restriction-induced negative energy balance. J. Dairy Sci. 2016, 99, 10009–10021. [Google Scholar] [CrossRef] [PubMed]
- Egger-Danner, C.; Cole, J.B.; Pryce, J.E.; Gengler, N.; Heringstad, B.; Bradley, A.; Stock, K.F. Invited review: Overview of new traits and phenotyping strategies in dairy cattle with a focus on functional traits. Animal 2015, 9, 191–207. [Google Scholar] [CrossRef]
- Rauw, W.M.; Gomez-Raya, L. Genotype by environment interaction and breeding for robustness in livestock. Front. Genet. 2015, 6, 310. [Google Scholar] [CrossRef] [PubMed]
- Contreras, G.A.; Sordillo, L.M. Lipid mobilization and inflammatory responses during the transition period of dairy cows. Comp. Immunol. Microbiol. Infect. Dis. 2011, 34, 281–289. [Google Scholar] [CrossRef]
- Dervishi, E.; Zhang, G.; Hailemariam, D.; Goldansaz, S.A.; Deng, Q.; Dunn, S.M.; Ametaj, B.N. Alterations in innate immunity reactants and carbohydrate and lipid metabolism precede occurrence of metritis in transition dairy cows. Res. Vet. Sci. 2016, 104, 30–39. [Google Scholar] [CrossRef]
- Zhang, G.; Hailemariam, D.; Dervishi, E.; Goldansaz, S.A.; Deng, Q.; Dunn, S.M.; Ametaj, B.N. Dairy cows affected by ketosis show alterations in innate immunity and lipid and carbohydrate metabolism during the dry off period and postpartum. Res. Vet. Sci. 2016, 107, 246–256. [Google Scholar] [CrossRef]
- McFadden, J.W. Review: Lipid biology in the periparturient dairy cow: Contemporary perspectives. Animal 2020, 14, s165–s175. [Google Scholar] [CrossRef] [Green Version]
- Dadarwal, D.; Palmer, C.; Griebel, P. Mucosal immunity of the postpartum bovine genital tract. Theriogenology 2017, 104, 62–71. [Google Scholar] [CrossRef] [PubMed]
- LeBlanc, S.J. Interactions of metabolism, inflammation, and reproductive tract health in the postpartum period in dairy cattle. Reprod. Domest. Anim. 2012, 47 (Suppl. S5), 18–30. [Google Scholar] [CrossRef] [PubMed]
- Moyes, K.M.; Drackley, J.K.; Morin, D.E.; Loor, J.J. Greater expression of TLR2, TLR4, and IL6 due to negative energy balance is associated with lower expression of HLA-DRA and HLA-A in bovine blood neutrophils after intramammary mastitis challenge with Streptococcus uberis. Funct. Integr. Genom. 2010, 10, 53–61. [Google Scholar] [CrossRef]
- Esposito, G.; Raffrenato, E.; Lukamba, S.D.; Adnane, M.; Irons, P.C.; Cormican, P.; Tasara, T.; Chapwanya, A. Characterization of metabolic and inflammatory profiles of transition dairy cows fed an energy-restricted diet. J. Anim. Sci. 2020, 98, skz391. [Google Scholar] [CrossRef] [PubMed]
- Bannerman, D.D.; Paape, M.J.; Hare, W.R.; Sohn, E.J. Increased levels of LPS-binding protein in bovine blood and milk following bacterial lipopolysaccharide challenge. J. Dairy Sci. 2003, 86, 3128–3137. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.M.; Lecchi, C.; Sauerwein, H.; Mielenz, M.; Haussler, S.; Restelli, L.; Giudice, C.; Ceciliani, F. Expression of alpha1-acid glycoprotein and lipopolysaccharide binding protein in visceral and subcutaneous adipose tissue of dairy cattle. Vet. J. 2015, 203, 223–227. [Google Scholar] [CrossRef]
- Townsend, M.; Singh, B. Pentraxin 3 expression in lungs and neutrophils of calves. Vet. Immunol. Immunopathol. 2021, 236, 110251. [Google Scholar] [CrossRef]
- Imamura, M.; Kawasaki, T.; Savchenko, A.S.; Ohashi, R.; Jiang, S.; Miyamoto, K.; Ito, Y.; Iwanari, H.; Sagara, M.; Tanaka, T.; et al. Lipopolysaccharide induced expression of pentraxin 3 in human neutrophils and monocyte-derived macrophages. Cell. Immunol. 2007, 248, 86–94. [Google Scholar] [CrossRef]
- Camozzi, M.; Rusnati, M.; Bugatti, A.; Bottazzi, B.; Mantovani, A.; Bastone, A.; Inforzato, A.; Vincenti, S.; Bracci, L.; Mastroianni, D.; et al. Identification of an antiangiogenic FGF2-binding site in the N terminus of the soluble pattern recognition receptor PTX3. J. Biol. Chem. 2006, 281, 22605–22613. [Google Scholar] [CrossRef]
- Genís, S.; Arís, A.; Kaur, M.; Cerri, R.L.A. Effect of metritis on endometrium tissue transcriptome during puerperium in Holstein lactating cows. Theriogenology 2018, 122, 116–123. [Google Scholar] [CrossRef] [Green Version]
- Cozzi, V.; Garlanda, C.; Nebuloni, M.; Maina, V.; Martinelli, A.; Calabrese, S.; Cetin, I. PTX3 as a potential endothelial dysfunction biomarker for severity of preeclampsia and IUGR. Placenta 2012, 33, 1039–1044. [Google Scholar] [CrossRef] [PubMed]
- Hatakeyama, T.; Okada, M.; Shimamoto, S.; Kubota, Y.; Kobayashi, R. Identification of intracellular target proteins of the calcium-signaling protein S100A12. Eur. J. Biochem. 2004, 271, 3765–3775. [Google Scholar] [CrossRef]
- Zhong, K.; Zhang, C.-Y.; Zha, G.-M.; Wang, X.-J.; Jiao, X.-Q.; Zhu, H.-S.; Wang, Y.-Y. S100 calcium-binding protein A12 as a diagnostic index for subclinical mastitis in cows. Reprod. Domest. Anim. 2018, 53, 1442–1447. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.-Y.; Yang, F.; Peng, L.-J.; Li, Y.-B.; Wang, A.P. CXCL2, a new critical factor and therapeutic target for cardiovascular diseases. Clin. Exp. Hypertens. 2020, 42, 428–437. [Google Scholar] [CrossRef] [PubMed]
- Rouault, C.; Pellegrinelli, V.; Schilch, R.; Cotillard, A.; Poitou, C.; Tordjman, J.; Sell, H.; Clement, K.; Lacasa, D. Roles of chemokine ligand-2 (CXCL2) and neutrophils in influencing endothelial cell function and inflammation of human adipose tissue. Endocrinology 2013, 154, 1069–1079. [Google Scholar] [CrossRef] [PubMed]
- Burfeind, K.G.; Zhu, X.; Levasseur, P.R.; Michaelis, K.A.; Norgard, M.A.; Marks, D.L. TRIF is a key inflammatory mediator of acute sickness behavior and cancer cachexia. Brain Behav. Immun. 2018, 73, 64–374. [Google Scholar] [CrossRef] [PubMed]
- Mosialou, I.; Shikhel, S.; Luo, N.; Petropoulou, P.I.; Panitsas, K.; Bisikirska, B.; Rothman, N.J.; Tenta, R.; Cariou, B.; Wargny, M.; et al. Lipocalin-2 counteracts metabolic dysregulation in obesity and diabetes. J. Exp. Med. 2020, 217, e20191261. [Google Scholar] [CrossRef] [PubMed]
- Frances, L.; Tavernier, G.; Viguerie, N. Adipose-Derived Lipid-Binding Proteins: The Good, the Bad and the Metabolic Diseases. Int. J. Mol. Sci. 2021, 22, 10460. [Google Scholar] [CrossRef]
- Petropoulou, P.I.; Mosialou, I.; Shikhel, S.; Hao, L.; Panitsas, K.; Bisikirska, B.; Luo, N.; Bahna, F.; Kim, J.; Carberry, P.; et al. Lipocalin-2 is an anorexigenic signal in primates. eLife 2020, 9, e58949. [Google Scholar] [CrossRef]
- Contreras, G.A.; Strieder-Barboza, C.; De Koster, J. Modulating adipose tissue lipolysis and remodeling to improve immune function during the transition period and early lactation of dairy cows. J. Dairy Sci. 2018, 101, 2737–2752. [Google Scholar] [CrossRef] [Green Version]
- Taniguchi, T.; Ogasawara, K.; Takaoka, A.; Tanaka, N. IRF family of transcription factors as regulators of host defense. Annu. Rev. Immunol. 2001, 19, 623–655. [Google Scholar] [CrossRef] [PubMed]
- Kowalec, K.; Wright, G.E.B.; Drögemöller, B.I.; Aminkeng, F.; Bhavsar, A.P.; Kingwell, E.; Yoshida, E.M.; Traboulsee, A.; Marrie, R.A.; Kremenchutzky, M.; et al. Common variation near IRF6 is associated with IFN-β-induced liver injury in multiple sclerosis. Nat. Genet. 2018, 50, 1081–1085. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Chu, H.; Jiang, Q.; Wang, C.; Tian, Y. Irf6 participates in sevoflurane-induced perioperative neurocognitive disorder via modulating M2, but not M1 polarization of microglia. Brain Res. Bull. 2021, 177, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Ying, W.; Huang, Z.; Brehm, T.; Morin, A.; Vella, A.T.; Zhou, B. IRF6 Regulates Alternative Activation by Suppressing PPARgamma in Male Murine Macrophages. Endocrinology 2017, 158, 2837–2847. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trait Material | Data Grouping | Sequencing Platform | Data Sources | Accession Numbers |
|---|---|---|---|---|
| Adipose | Normal (PreP), n = 12 NEB (PP1), n = 12 | Illumina NextSeq 500 (Bos taurus) | David Salcedo-Tacuma, 2020 | GSE159224 |
| Blood | Normal, n = 10 NEB, n = 13 | Illumina HiSeq 2000 (Bos taurus) | Ze Yan, 2020 | PRJNA605719 |
| Endometrium | Normal (MNEB), n = 15 NEB (SNEB), n = 9 | Illumina HiSeq 4000 (Bos taurus) | Wiruntita Chankeaw, 2021 | GSE169638 |
| Hypothalamus | Normal (Restricted), n = 14 NEB (high level), n = 7 | Illumina HiSeq 4000 (Bos taurus) | Daragh Matthews, 2017 | GSE49540 |
| Liver | Normal (MNEB), n = 5 NEB (SNEB), n = 6 | Illumina Genome Analyzer (Bos taurus) | Matthew McCabe, 2012 | GSE37544 |
| GO Term | NES | Tissue | Regulated |
|---|---|---|---|
| Humoral Immune Response Mediated by Circulating Immunoglobulin | 1.8095 | Adipose | Up |
| Membrane Invagination | 1.8004 | Adipose | Up |
| Regulation of Humoral Immune Response | 1.7919 | Adipose | Up |
| Amyloid Beta Clearance | 1.7888 | Adipose | Up |
| Complement Activation | 1.7642 | Adipose | Up |
| Negative Regulation of Blood Vessel Diameter | −1.6923 | Adipose | Down |
| Regulation of Steroid Biosynthetic Process | −1.702 | Adipose | Down |
| Regulation of Steroid Metabolic Process | −1.7112 | Adipose | Down |
| Fatty Acid Metabolic Process | −1.9598 | Adipose | Down |
| Sterol Metabolic Process | −2.1124 | Adipose | Down |
| Myeloid Leukocyte Mediated Immunity | 2.2295 | Blood | Up |
| Exocytosis | 2.1093 | Blood | Up |
| Myeloid Leukocyte Activation | 1.9157 | Blood | Up |
| Defense Response to Gram-Positive Bacterium | 1.8822 | Blood | Up |
| Cell Activation Involved in Immune Response | 1.8773 | Blood | Up |
| B Cell Activation | −1.9504 | Blood | Down |
| Regulation of Fat Cell Differentiation | −2.0154 | Blood | Down |
| Regulation of Immunoglobulin Production | −2.147 | Blood | Down |
| B Cell Activation Involved in Immune Response | −2.2318 | Blood | Down |
| Immunoglobulin Production | −2.2439 | Blood | Down |
| Lymphocyte Chemotaxis | 1.9792 | Endometrium | Up |
| Protein-DNA Complex Subunit Organization | 1.9613 | Endometrium | Up |
| Organ Growth | 1.949 | Endometrium | Up |
| Heart Growth | 1.8857 | Endometrium | Up |
| Positive Regulation of Heart Growth | 1.8108 | Endometrium | Up |
| Cellular Response to Virus | −1.6434 | Endometrium | Down |
| Positive Regulation of Response to Cytokine Stimulus | −1.6448 | Endometrium | Down |
| Sterol Metabolic Process | −1.6475 | Endometrium | Down |
| Regulation of Jun Kinase Activity | −1.6754 | Endometrium | Down |
| Positive Regulation of Jun Kinase Activity | −1.7641 | Endometrium | Down |
| Response to Retinoic Acid | 1.9746 | Hypothalamus | Up |
| Positive Regulation of CD4-Positive Alpha Beta T-Cell Differentiation | 1.8528 | Hypothalamus | Up |
| Regulation of CD4-Positive Alpha Beta T-Cell Differentiation | 1.7604 | Hypothalamus | Up |
| Leukocyte Homeostasis | 1.6864 | Hypothalamus | Up |
| Response to Chemokine | 1.6836 | Hypothalamus | Up |
| Non-canonical Wnt Signaling Pathway | −1.6246 | Hypothalamus | Down |
| Calcium Ion Transmembrane Import into Cytosol | −1.6279 | Hypothalamus | Down |
| Neuron Projection Organization | −1.6319 | Hypothalamus | Down |
| Cellular Response to Ketone | −1.6627 | Hypothalamus | Down |
| Regulation of Calcium Ion Transmembrane Transport | −1.6661 | Hypothalamus | Down |
| Tissue Migration | 1.695 | Liver | Up |
| Negative Regulation of Cell Adhesion | 1.6623 | Liver | Up |
| Negative Regulation of Coagulation | 1.6527 | Liver | Up |
| Odontogenesis | 1.65 | Liver | Up |
| Positive Regulation of Cell Division | 1.6472 | Liver | Up |
| Calcineurin-Mediated Signaling | −1.656 | Liver | Down |
| Positive Regulation of Lipid Kinase Activity | −1.6596 | Liver | Down |
| Glucose Metabolic Process | −1.6671 | Liver | Down |
| Establishment of Cell Polarity | −1.7294 | Liver | Down |
| Positive Regulation of Phospholipid Metabolic Process | −1.7409 | Liver | Down |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dai, L.; Liu, Z.; Guo, L.; Chai, Y.; Yang, Y.; Wang, Y.; Ma, Y.; Shi, C.; Zhang, W. Multi-Tissue Transcriptome Study of Innate Immune Gene Expression Profiling Reveals Negative Energy Balance Altered the Defense and Promoted System Inflammation of Dairy Cows. Vet. Sci. 2023, 10, 107. https://doi.org/10.3390/vetsci10020107
Dai L, Liu Z, Guo L, Chai Y, Yang Y, Wang Y, Ma Y, Shi C, Zhang W. Multi-Tissue Transcriptome Study of Innate Immune Gene Expression Profiling Reveals Negative Energy Balance Altered the Defense and Promoted System Inflammation of Dairy Cows. Veterinary Sciences. 2023; 10(2):107. https://doi.org/10.3390/vetsci10020107
Chicago/Turabian StyleDai, Lingli, Zaixia Liu, Lili Guo, Yuan Chai, Yanda Yang, Yu Wang, Yanfen Ma, Caixia Shi, and Wenguang Zhang. 2023. "Multi-Tissue Transcriptome Study of Innate Immune Gene Expression Profiling Reveals Negative Energy Balance Altered the Defense and Promoted System Inflammation of Dairy Cows" Veterinary Sciences 10, no. 2: 107. https://doi.org/10.3390/vetsci10020107