
Investigation of the Effects of Monosodium Glutamate on the Embryonic Development of the Eye in Chickens

1
Department of Histology and Embryology, Faculty of Medicine, Aksaray University, 68100 Aksaray, Turkey
2
Department of Histology and Embryology, Faculty of Veterinary Medicine, Selcuk University, 42100 Konya, Turkey
*
Author to whom correspondence should be addressed.
Vet. Sci. 2023, 10(2), 99; https://doi.org/10.3390/vetsci10020099
Submission received: 9 December 2022
/
Revised: 19 January 2023
/
Accepted: 28 January 2023
/
Published: 30 January 2023
(This article belongs to the Section Anatomy, Histology and Pathology)
Abstract
:Simple Summary
Monosodium glutamate (MSG, E62, C5H8NO4Na) is the most widely used food additive in the world for enhancing flavour. The number of flavour-enhancers added to foods is very important for the health of consumers. It has been determined by many researchers that MSG has negative effects on various organs of the body. Since chicken embryos develop without the influence of the maternal organism and allow toxicity to be evaluated very quickly and precisely, they have become a preferred experimental model for investigating the embryotoxic and teratogenic effects of chemicals, toxins, drugs, and numerous food additives and flavour-enhancing agents. This study aimed to investigate the effects of in ovo MSG administration at different doses on the embryonic development of the eye using histological and histometric techniques. The results of the present work have shown that in ovo-administered MSG can adversely affect embryonic eye development.
Abstract
MSG is the most ubiquitous food additive in the food industry. The aim of this report was to investigate the effects of in ovo MSG administration on embryonic chicken eye development using histological and histometric methods. A total of 410 fertilized eggs obtained from Babcock Brown laying hens (Gallus gallus domesticus) were used and divided into 5 groups: I (untreated control), II (vehicle control), III (0.12 mg/g egg MSG), IV (0.6 mg/g egg MSG), and V (1.2 mg/g egg MSG), and injections were performed via the egg yolk. At incubation day 15, 18, and 21, 6 embryos from each group were sacrificed by decapitation and pieces of eye tissue were obtained. In all MSG groups, it was determined that both corneal epithelium thickness and total corneal thickness decreased at incubation time points 15, 18, and 21 days compared with the controls (p < 0.05). The total retinal thickness, thickness of the outer nuclear layer (ONL), inner nuclear layer (INL), ganglion cell layer (GL), and nerve fibre layers (NFL), as well as the number of ganglion cells decreased significantly at incubation days 15, 18, and 21 (p < 0.05), and degenerative changes such as vacuolar degeneration and retinal pigment epithelial detachment were also observed. In conclusion, MSG in ovo administration can affect the cornea and distinct layers of retinal cells.
1. Introduction
Monosodium glutamate (MSG) has been used extensively to enhance the flavour of seasonings in the food industry and restaurants. MSG can be found in processed food without an indication on the label, especially to increase the consumption and flavour of processed food such as snack food, chips, sauces, and soups [1,2]. Natural form glutamic acid in food is not toxic, but industrially produced synthetic glutamic acid is a toxin [3]. MSG is available without limits in a wide variety of processed food. It is also added to meals in unlimited quantities in restaurants, hospitals, nursing homes, and cafeterias. Food manufacturers do not list the amount of MSG on their packaging, so there is no way to know how much MSG an adult or child consumes in a day. According to industrial research, 0.6% MSG is optimally added to food [4]. According to the U.S. Environmental Protection Agency, the use of MSG in food should not be allowed for infants and especially children under one year of age [4].
Glutamate is the primary neurotransmitter stimulant for brain networks and stimulates respective receptors, which have important effects in both physiological and pathological processes [5]. Excessive stimulation of glutamate receptors leads to the development of excitotoxicity [6]. Glutamate is metabolized to non-toxic glutamine by glutamine synthetase after being taken into Müller cells by the glutamate transporter, GLAST [7]. Glutamic acid is responsible for synaptic transmission between bipolar, photoreceptor, and ganglion cells in the retina, and its presence in high amounts is responsible for neuronal cell death [8], such as retinal ganglion cells [9].
The toxic effects of MSG on the central nervous system, adipose tissue, hepatic tissue, reproductive organs, and liver and kidney functions were determined by several studies [2,10,11]. These studies revealed that MSG causes cytotoxicity [12,13] and oxidative damage in the liver, kidney, and various tissues [14,15,16,17,18] and increased the risk of certain cancers [16]. MSG may adversely affect immune system organs [13,18,19], the testes [20,21,22], ovarium [23,24,25,26,27], pancreas [28], spleen [29,30,31], liver [15,32,33], and cardiac tissue [2]. It has been reported that the incidence of certain diseases, such as obesity [34,35], diabetes, and Alzheimer’s is increased in people who consume foods with added MSG [36,37].
MSG can cause neurotoxicity, neurodegeneration [38], and neuroendocrine abnormalities [39]. There have been many studies investigating the negative effects of MSG on the nervous system [40,41,42,43,44,45].
Reports have demonstrated that MSG can be toxic to foetal development, children, and adults [4]. Maternal MSG exposure can result in neurotoxic effects and severe intrauterine growth retardation in rats [46]. In addition, MSG has been found to cause apoptosis and necrosis in the hippocampus of prepubertal rats [47]. Narayanan et al. [48] reported that MSG showed neurotoxic effects when administered to newborn animals at high concentrations. MSG has been reported to cause neurological damage by inducing oxidative stress and neurotoxicity, which is more severe in newborns during brain development than in adults [9]. In addition, Gim et al. [49] reported that MSG injection to newborn rats caused histopathological changes in the brain and degeneration of the cerebral cortex. Bölükbaş and Öznurlu [42] reported that in ovo MSG administration caused histopathological changes such as necrosis, neuronophagia, and gliosis in brain tissue.
Studies have shown that exposure to MSG causes significant degenerative effects on the eye and retinal layers [50,51,52,53,54]. Dénes et al. [55] revealed that subcutaneous injection of MSG into rats causes retinal degeneration and pycnosis of retinal cells. Additionally, in further study, it was reported that the density of ganglion cells in the retina was considerably reduced in rat models exposed to MSG, in comparison with control rats [56]. Praputpittaya and Wililak [57] reported that subcutaneous injection of MSG in different doses to newborn rats caused deficits in visual performance. Swelim [58] reported that low concentrations of subcutaneous MSG injections in neonate mice caused retinal damage.
It has been suggested that MSG administration in newborn rats leads to degeneration of retinal ganglion cells, as well as degeneration of the optic nerve [51,56]. It has also been reported to lead to degeneration of neurons in retinal layers, the arcuate nucleus, and various other brain regions, delaying the emergence of certain reflexes during neurobehavioral development and leading to temporary changes in reflex performance and motor coordination [59,60]. El-Sayyad et al. [61] found that MSG given orally to pregnant rats significantly reduced total retinal thickness, outer and inner nuclear layers, and photoreceptor layer thickness. It has also been reported that mother rats have retinal ganglion degeneration, loss of pigment epithelium, and vacuolization in the plexiform inner layer. The chick eye has been a common model to study embryonic development as well as eye diseases. The chicken eye has similar basic components to the human eye including the cornea, ciliary body, iris, lens, sclera, choroid, retina, and optic nerve [62,63]. While numerous studies have reported the neurotoxic effects of MSG on humans, experimental animals, and chicken embryos [38,41,42,45,64,65], there is insufficient information about the effects of MSG on the cornea and retina during the embryonic period. This study aimed to investigate the effects of in ovo MSG administration at different doses on the cornea and retina in chicken embryos using histological and histometric techniques.
2. Materials and Methods
2.1. Experimental Design and Preperation of Test Solution
For the experiments, 410 Babcock Brown fertilized eggs (50–55 g) obtained from Babcock Brown laying hens (Gallus gallus domesticus) were examined. The MSG doses were adjusted for 55 g egg weight. MSG (Sigma-Aldrich Chemical) was diluted with sterilized distilled water. MSG doses of 0.12 mg/g, 0.6 mg/g, and 1.2 mg/g egg were prepared and given in a volume of 100 μL. Prior to MSG injection, the eggs were disinfected for 15 min under steam obtained from mixing 21 g of potassium permanganate with 42 mL of formaldehyde/m3 in a closed cabin. The eggs were divided into 5 groups: Group I (untreated control, 40 eggs), which includes non-treated eggs; Group II (vehicle control, 62 eggs), which injected only distilled water; and three MSG-injected groups as Group III (0.12 mg/g egg MSG, 80 eggs), Group IV (0.6 mg/g egg MSG, 90 eggs), and Group V (1.2 mg/g egg MSG, 138 eggs). Embryonic deaths were considered in numbers of eggs per group. The eggs were injected just before starting the incubation period. In the injection groups, the injection sites on the eggs were wiped with 96% ethanol for further disinfection. All injections were performed in egg yolks at the beginning of the incubation period. By drilling a hole on the side of the egg with a special egg driller, the test solution was injected through a sterile insulin injector (26 Gx1/2”, Beybi) and then the hole was sealed with liquid paraffin. Incubations were carried out in an incubator (Imza Technical Equipment, Konya, Turkey), under optimal conditions (37.5 °C temperature and 65% relative humidity). The eggs were subjected to turning angles of 45 with a turning frequency of 12 times daily.
2.2. Tissue Sampling and Histologic Procedures
At incubation days 15, 18, and 21, 6 embryos from each group were sacrificed by decapitation. The stages of development of the living embryos were determined according to the Hamburger–Hamilton (1951) scale (H–H scale) [66]. Eye tissue samples were collected from embryos and fixed in 10% formalin for 24 h and then subjected to dehydration and paraffin embedding. For routine histological examination, tissue sections were stained with Crossmon’s trichrome [67], Toluidine blue, and Hematoxylin and Eosin (H&E) staining [68]. Histometric measurements were performed in serial sections taken from the eye tissue obtained on incubation days 15, 18, and 21. The thickness of the total cornea and corneal epithelium were measured from three different regions of the cornea. The thickness of total retina and retinal layers were measured from four different retina regions. Additionally, the ganglion cell number was determined by counting the number on a 100 μm line length in three different fields per section. All evaluations were performed by two researchers blinded to the sample identification.
Next, prepared sections were examined with a Leica DM-2500 model light microscope. Digital images of the required areas were captured using an attached DFC-320 model camera and analysed for histometric measurements. All histometric measurements were performed using the Leica IM50 measurement program (Leica, Leica Microsystems GmbH, Wetzlar, Germany) and numerical data of the investigated parameters were obtained.
2.3. Statistical Analysis
The collected data were given in mean ± SD. A p-value of less than 0.05 was considered statistically significant. Total corneal thickness, corneal epithelial thickness, total retinal thickness, retinal layers thickness, and the number of ganglion cells were evaluated with ANOVA and the Tukey test. This was performed by using SPSS 26 (SPSS, IBM Corp. Released 2019, Armonk, NY, USA).
3. Results
3.1. Incubation Day 15
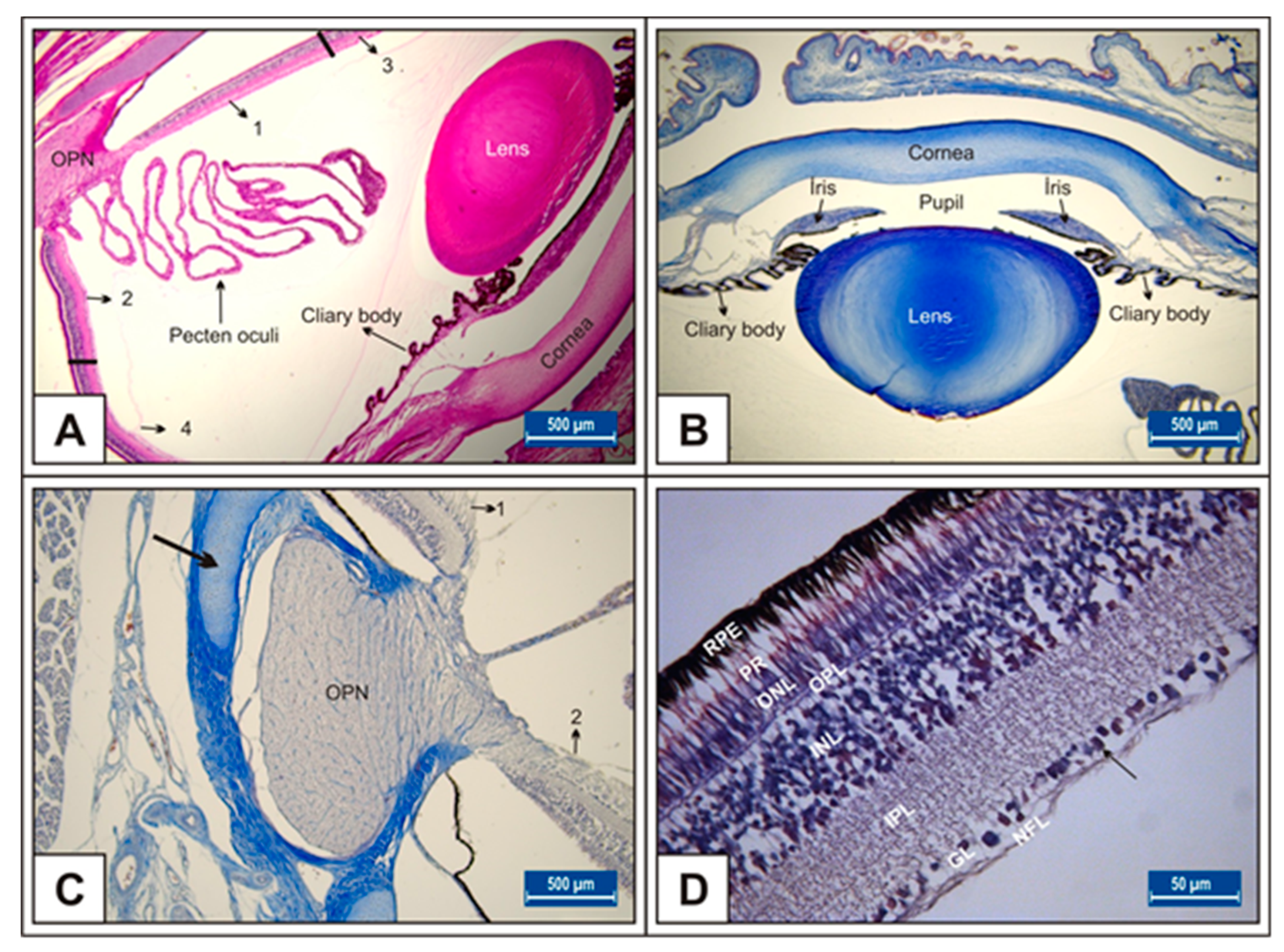
On day 15 of incubation, the transparent structures that refract and focus light (cornea and lens), muscle structures (iris and ciliary body), retina, pecten oculi, and optic nerve have developed (Figure 1A–D).
It was noted that in the MSG-treated groups, corneal epithelium thickness and total corneal thickness decreased depending on the dose in comparison to untreated and vehicle controls (p < 0.05, Figure 2A–E, Table 1).
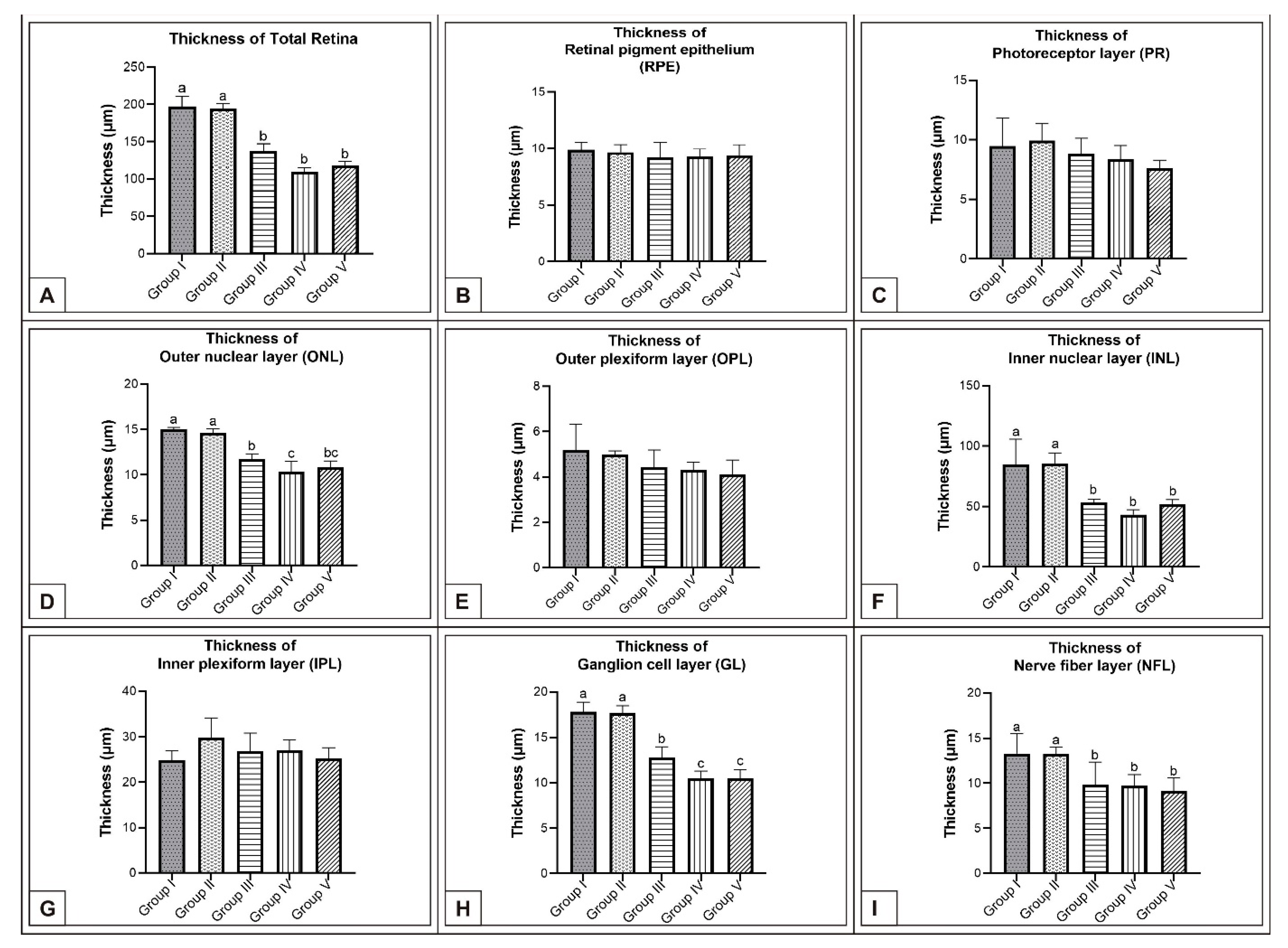
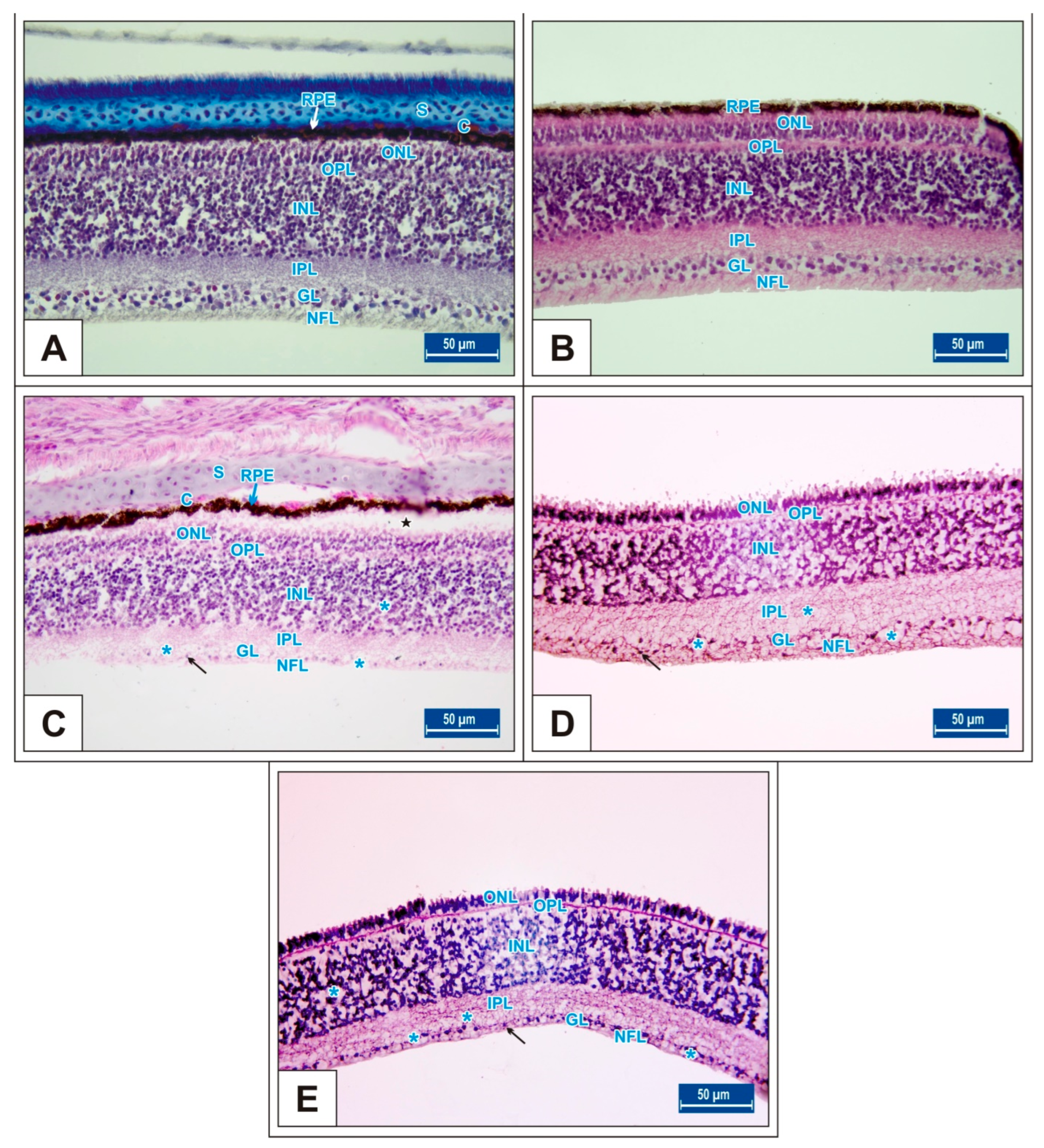
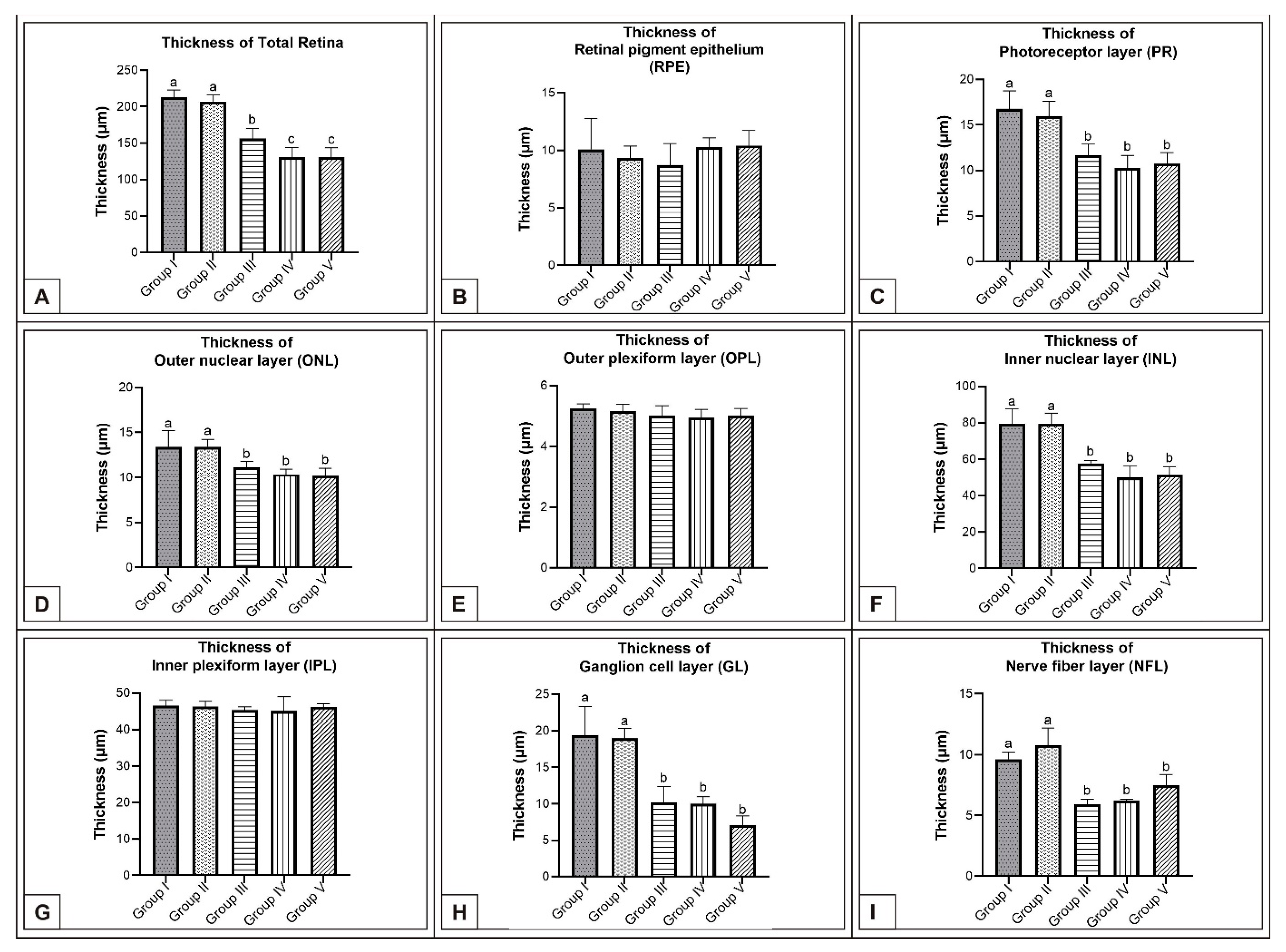
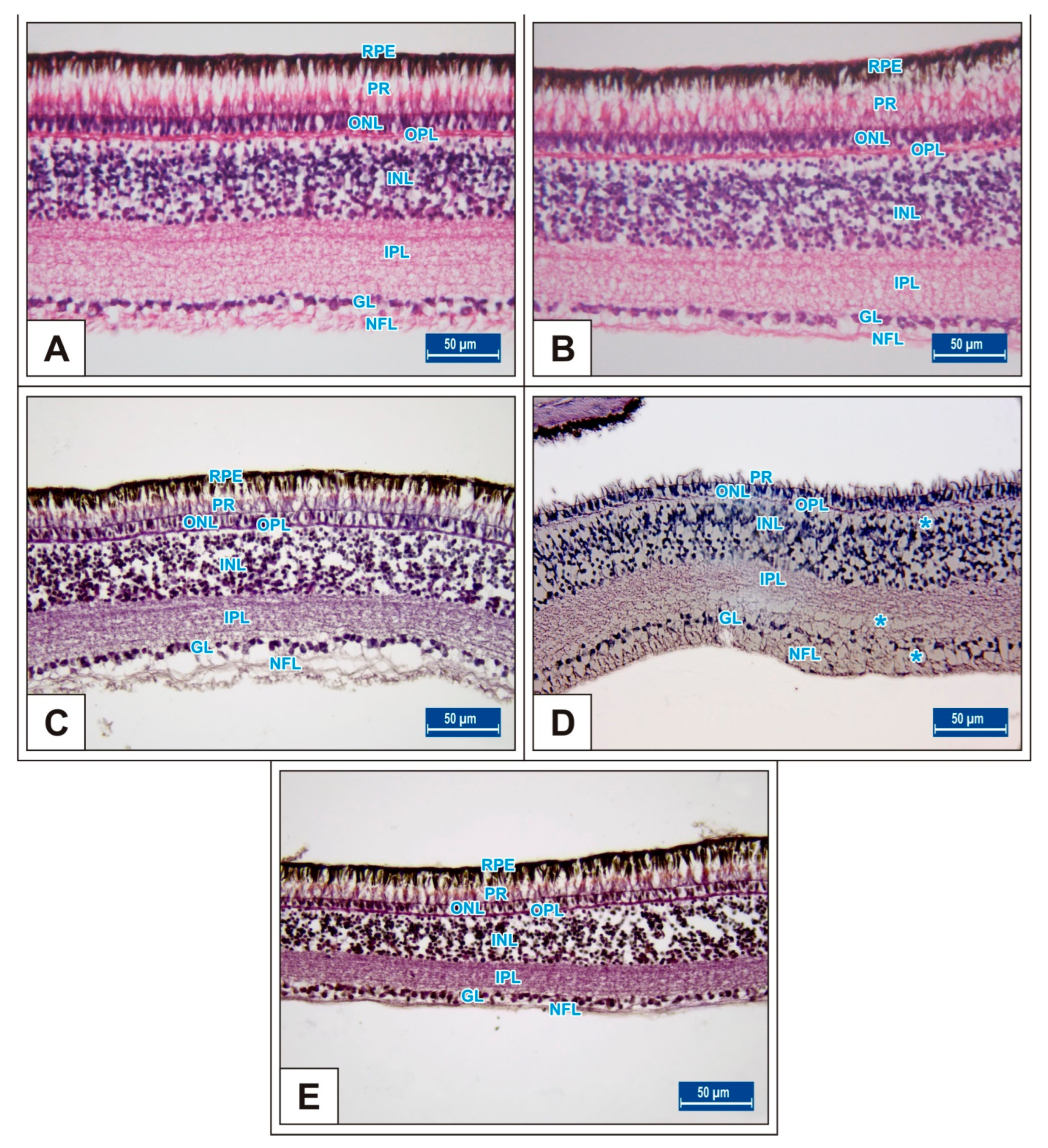
On the 15th day of incubation, it was observed that the retina consisted of 9 layers, respectively, from the outside to the inside: retinal pigment epithelium (RPE), photoreceptor layer (PR), outer limiting membrane, outer nuclear layer (ONL), outer plexiform layer (OPL), inner nuclear layer (INL), inner plexiform layer (IPL), ganglion cell layer (GL), and nerve fibre layer (NFL) (Figure 1D). In the MSG groups, total retinal thickness decreased significantly compared with the controls. Corneal epithelium thickness and total corneal thickness decreased in the MSG groups depending on the dose compared with the controls, and the thickness of the retinal layers, especially the ONL, INL, GL, and NFL layers decreased significantly (p < 0.05, Figure 3A–I). Furthermore, in these groups retinal degeneration such as retinal pigment epithelium detachment (Figure 4C–E) and vacuolization in the IPL, INL, GL, and NFL layers were observed (Figure 4C–E). Moreover, ganglion cell count was significantly reduced in the GL layer and this layer was thinner for all MSG groups (p < 0.05, Figure 4A–D, Table 2).
3.2. Incubation Day 18
On incubation day 18, in MSG-treated groups retinal degenerations such as detachment of the retinal pigment epithelium (Figure 5C,D) and vacuolization in the IPL, INL, GL, and PR layer were observed (Figure 5E). Moreover, ganglion cell numbers were significantly decreased in all MSG groups compared with controls (p < 0.05, Table 2, Figure 5C–E).
In the MSG groups, corneal epithelium and total corneal thickness decreased depending on the dose in comparison with controls (p < 0.05, Figure 2F–J, Table 1). Similarly, the total retinal thickness decreased significantly when compared with controls; in particular, thickness of the PR, ONL, INL, GL, and NFL layers was reduced (p < 0.05, Figure 6A–I).
3.3. Incubation Day 21
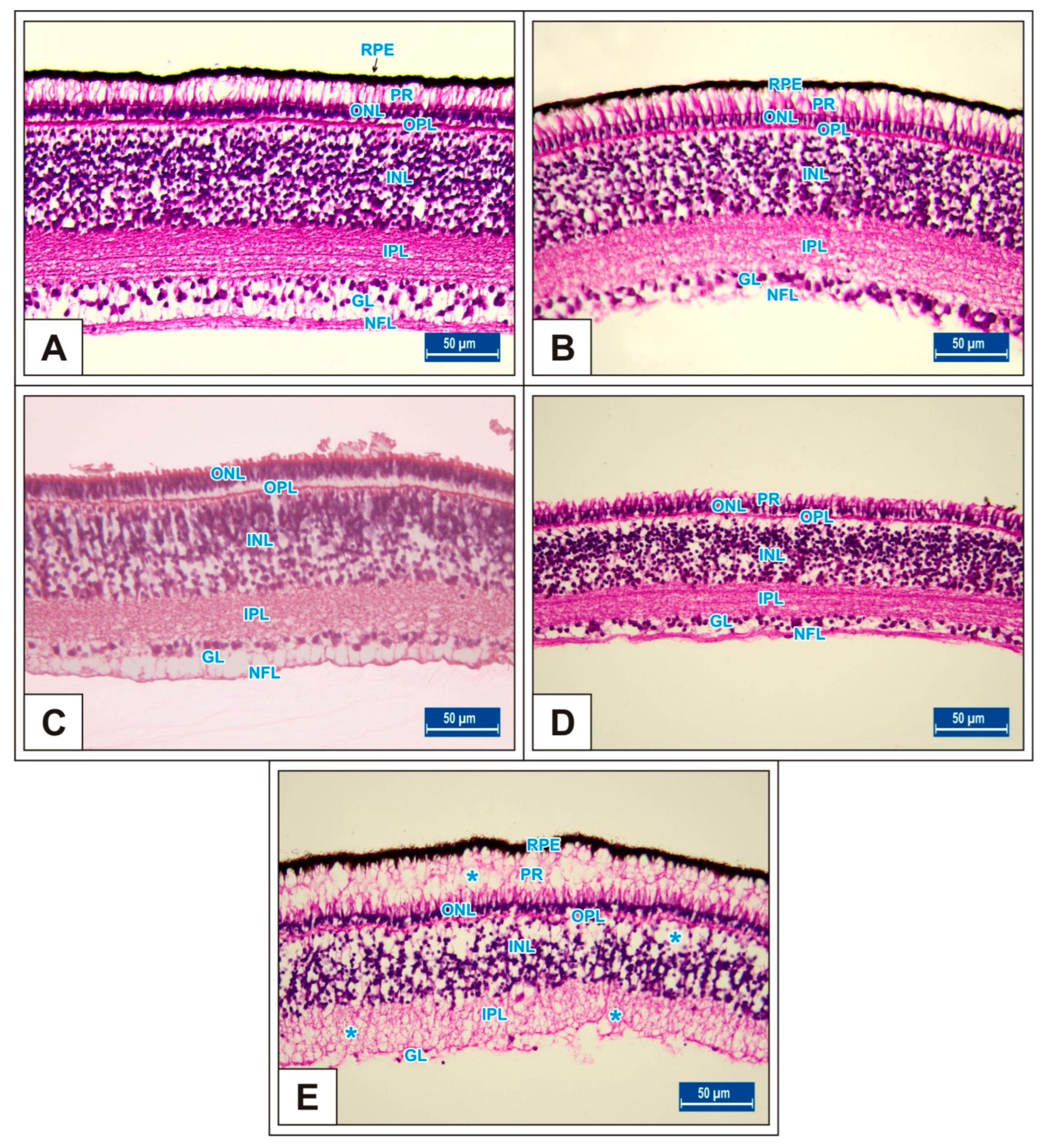
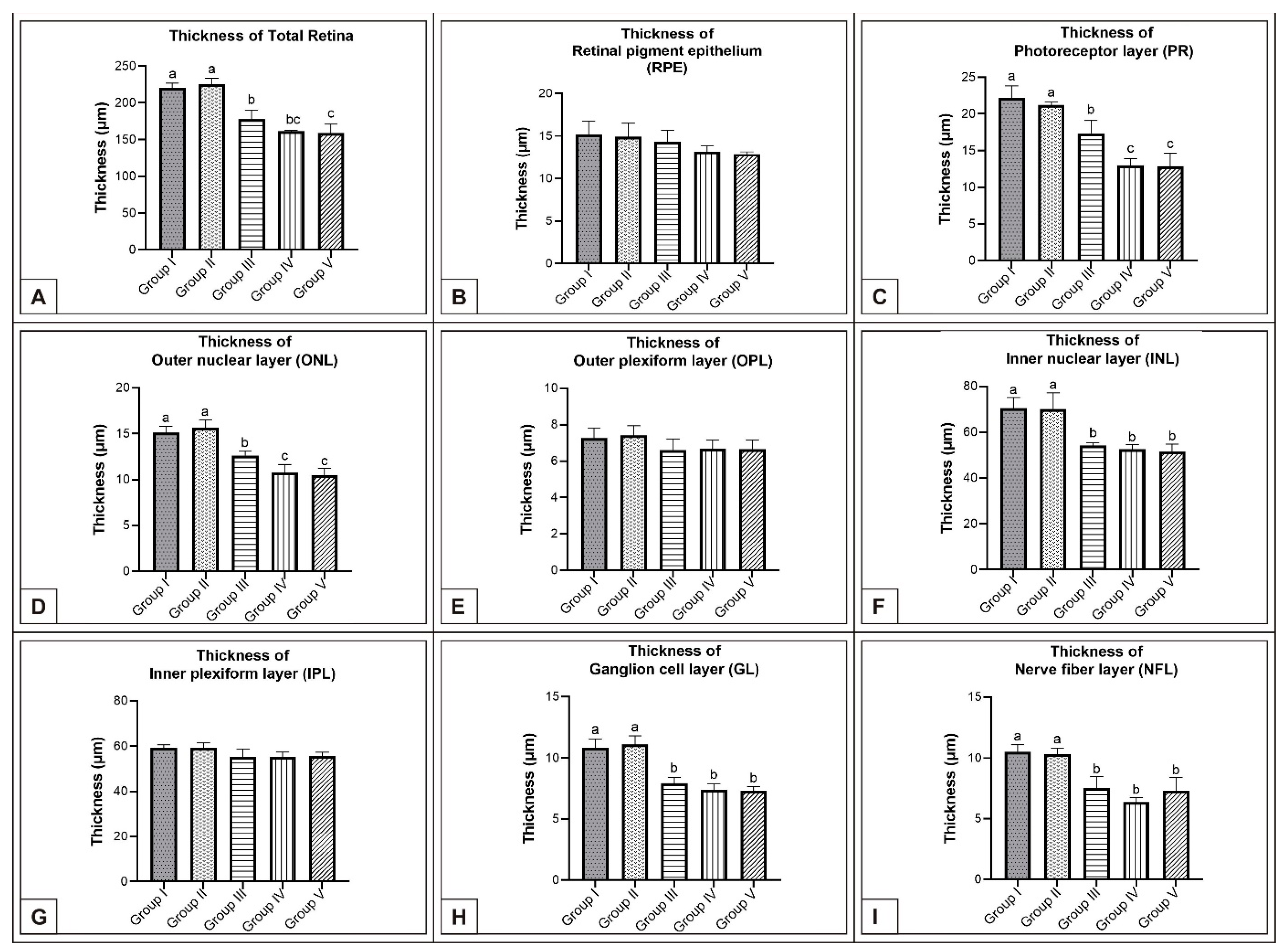
On incubation day 21, it was determined that corneal epithelium and total corneal thickness significantly decreased following MSG treatment in comparison with controls (p < 0.05, Figure 2K–O, Table 1). It was also noted that total RT as well as PR, ONL, INL, GL, and NFL thickness significantly decreased when compared with controls (p < 0.05, Figure 7A–E, Figure 8A–I). These groups were observed to have retinal degenerations such as detachment of the RPE and cellular vacuolization of INL and GL layers (Figure 7D). Additionally, the number of ganglion cells in the GL layer of the MSG groups significantly decreased in comparison with the control groups (p < 0.05, Table 2).
4. Discussion
In this study, we studied the effects of in ovo MSG administered at different doses on the cornea and retina of chicken embryos. While a significant number of studies have reported on the various adverse effects of MSG on humans, experimental animals, and chicken embryos [38,41,42,45,64,65,69,70], no detailed studies have been found in the literature on the effects of MSG on embryonic eye development in chick embryos.
Regulations have been imposed on the use of MSG as a flavour-enhancing food additive. In August 2017, the European Food Safety Authority (EFSA) corrected the acceptable daily intake (ADI) for glutamic acid and salts from 120 mg to 30 mg per kilogram of body weight. While the average daily intake of MSG ranges from 0.3 to 0.5 g per day among European Union member countries, the average daily intake of MSG ranges are higher in Asian countries, ranging from 1.6–2.3 g/day in South Korea, 1.5–3.0 g/day in Taiwan, 1.1–1.6 g/day in Japan, and 4 g/day in China [1,3]. In recent years, large-scale shifts in lifestyle and eating habits across the globe have led to increased consumption of processed food, elevating the risk of exceeding the ADI of MSG and similar chemicals. Many reports have demonstrated the adverse effects of low dose MSG resulting from chronic consumption [1,2,71,72]. MSG doses between 1.25 and 12 gr were used in human experimental studies [73,74], dosages ranging from 0.04 to 100 g in animal studies [2,12,75,76], and dosages between 0.75 and 3 mg/g egg in chicken embryo studies [41,70]. In this study, both human daily MSG intake and previous chicken embryo studies were taken into account in the adjustment of MSG doses (0.12 mg/g, 0.6 mg/g, and 1.2 mg/g eggs).
Plasma glutamate concentration has been reported to increase significantly after MSG supplementation in humans [77]. High oral intake of MSG in rats has been shown to result in oxidative stress in multiple areas of the brain and in the retina [78,79]. This excitotoxicity effect of glutamate was reported to trigger extreme glutamate receptor (GluR) activity [80]. Ganglion cell and retina IPL have been reported to be mostly affected by both in vivo and in vitro MSG administration [55]. Van Rijn et al. [56] showed that the administration of MSG to rats significantly reduced the density of ganglion cells in the retina in comparison with controls. El-Gohari et al. [81] showed that different doses of MSG injection in rats caused retinal degeneration as well as a decrease in retinal thickness and ganglion cell count. Researchers have also reported that MSG causes detachment in the RPE and the irregular presence of the photoreceptor layer in rats. Ohguro et al. [79] reported that retinal layer thickness was thinner and the number of cells in GL, INL, and ONL decreased in rats fed three different doses of MSG. Bellhorn et al. [50] showed that MSG given to newborn rats on days 1 to 10 after birth caused degeneration of the inner retinal layers. In addition, oral administration of MSG has been reported to cause degeneration in rabbit retina [82]. AlThanoon and Abd [83] reported that different doses of MSG injection to pregnant mice caused degeneration and necrosis of the INL and ganglion cells as well as of the optic nerve depending on the dose in mouse embryos on the 14th and 18th days of pregnancy.
Chicken embryos are a common model for investigation of embryotoxicity [84,85], neurotoxicity [42,86,87], eye diseases [88], retinal development [89], and retinal pathologies [90]. Therefore, chicken embryos are good models for investigation of the effects of many food additives, flavour-enhancing additives, and environmental pollutants during embryogenesis because embryonic development is fully described and the individual developmental stages are clearly visible and easily accessible [91,92]. The chicken egg is a closed system that has no interaction with its environment, except for the interchange of gases. Jessl et al. [92] suggested that a single injection of different test solutions into the yolk sac on day one of incubation results in chronic chemical exposure and may be sufficient to influence the developing embryo. In this study, different doses of MSG (0.12 mg/g, 0.6 mg/g, and 1.2 mg/g eggs) were administered to egg yolks as a single injection, thus creating chronic exposure.
Changes in corneal thickness with age were still unclear in humans and animals. While some studies reported no significant change in corneal thickness over time [93,94], other studies showed a decreasing trend of corneal thickness with age [95,96]. Inomata et al. [97] reported that corneal thickness initially increased between 1 and 6 months, reached a maximum at 9 months, and then decreased between 12 and 14 months, while body size (weight) increased with age. AlThanoon and Abd [83] reported that, in the eyes of 18-day-old MSG-treated albino mice foetuses, there was deformation of the lens, thickening of the cornea’s inner lining, necrosis of the cornea’s stroma, and necrosis of the optic nerve. Al-Qudsi and Azzouz [98] showed that electromagnetic mobile radiation caused changes in neural retinal thickness and congenital malformations in chick embryos at 7, 10, and 14 days of incubation. In previous chick embryo retina culture studies, MSG was suggested to contribute to inner retinal layer and ganglion cell damage [99,100]. Kujawa-Hadryś et al. [101] reported that the thickness of the corneal epithelium increased markedly through the end of incubation in chick embryos. Several studies have been conducted examining normal retinal structure in poultry [102,103]. However, there is insufficient information about the effects of MSG on the cornea and retina in chicken embryos.
In this study, it was observed that corneal epithelium and total corneal thickness increased considerably from day 15 to day 21 of incubation. In the MSG groups, both corneal epithelium and total corneal thickness significantly decreased depending on the dose on days 15, 18, and 21 of incubation compared with controls, (p < 0.05, Figure 2, Table 1). In addition, detachment in the corneal epithelium was noted in the MSG groups. It was further noted that in the MSG groups, total retinal thickness decreased significantly on the incubation days examined compared with control groups, and thickness of both the ONL, INL, GL, and NFL layers of the retinal layers on the 15th day, and the PR, ONL, INL, GL, and NFL layers on days 18 and 21, were significantly reduced (p < 0.05, Figure 3A–I, Figure 6A–I and Figure 8A–I). In these groups, degenerative changes such as vacuolar degeneration and retinal pigment epithelial detachment were observed. It was also observed that the number of ganglion cells decreased markedly in all MSG groups in comparison with the control groups on all days tested (p < 0.05, Table 2). These findings were consistent with rat, mouse, and chicken studies [79,81,82,83,97,99,100].
This study has certain limitations that should be noted. First, it is difficult to calculate the rate of MSG uptake from the yolk sac of the developing embryo; however, it can be argued that this rate is proportional to the rate of embryo development. Moreover, the chicken embryo model has the advantage of allowing potentially hazardous chemicals to be investigated directly on the embryo. On the other hand, the detachments seen in the cornea and retina bring to mind artificial separations in the sections. However, the absence of such detachment in the control groups was evaluated as the effect of MSG.
5. Conclusions
It was observed that different doses of in ovo MSG administration caused histometric and histopathological changes in both the cornea and retina. Since the data obtained from studies using chicken embryos can also be adapted to mammals, the findings of this study suggest that animals and humans exposed to MSG during the prenatal period may have increased susceptibility to certain eye diseases during their lifetime. Given these results, our study adds a new perspective to the literature. Nowadays, while exposure to and/or consumption of MSG in various ways is rapidly and unsafely increasing, it may be advisable to minimize or even prohibit eating foods containing MSG, especially during pregnancy.
Author Contributions
F.B.: research procedure design, conceptualization, resources, writing—review, investigation. Y.Ö.: conceptualization, resources, writing—review and editing, statistics. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
For this research, ethical approval was obtained from Selcuk University Veterinary Faculty Experimental Animals Production and Research Center (SÜVDAMEK) and an ethics report for this study was completed (02.06.2022 and 2022/60).
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Nguyen, L.; Salanta, L.-C.; Socaci, S.; Tofana, M.; Fărcaş, A.; Pop, C. A Mini Review About Monosodium Glutamate. Bull. Univ. Agric. Sci. Vet. Med. Cluj-Napoca. Food Sci. Technol. 2020, 77, 2020. [Google Scholar] [CrossRef]
- Zanfirescu, A.; Ungurianu, A.; Tsatsakis, A.M.; Nițulescu, G.M.; Kouretas, D.; Veskoukis, A.; Tsoukalas, D.; Engin, A.B.; Aschner, M.; Margină, D. A Review of the Alleged Health Hazards of Monosodium Glutamate. Compr. Rev. Food Sci. Food Saf. 2019, 18, 1111–1134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beyreuther, K.; Biesalski, H.K.; Fernstrom, J.D.; Grimm, P.; Hammes, W.P.; Heinemann, U.; Kempski, O.; Stehle, P.; Steinhart, H.; Walker, R. Consensus meeting: Monosodium glutamate—An update. Eur. J. Clin. Nutr. 2007, 61, 304–313. [Google Scholar] [CrossRef] [Green Version]
- Chakraborty, S.P. Patho-physiological and toxicological aspects of monosodium glutamate. Toxicol. Mech. Methods 2019, 29, 389–396. [Google Scholar] [CrossRef] [PubMed]
- Sedlak, T.W.; Paul, B.D.; Parker, G.M.; Hester, L.D.; Snowman, A.M.; Taniguchi, Y.; Kamiya, A.; Snyder, S.H.; Sawa, A. The glutathione cycle shapes synaptic glutamate activity. Proc. Natl. Acad. Sci. USA 2019, 116, 2701–2706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, A.; Kalotra, S.; Bajaj, P.; Singh, H.; Kaur, G. Butanol extract of Tinospora cordifolia ameliorates cognitive deficits associated with glutamate-induced excitotoxicity: A mechanistic study using hippocampal neurons. Neuromolecular Med. 2020, 22, 81–99. [Google Scholar] [CrossRef]
- Derouiche, A.; Rauen, T. Coincidence of L-glutamate/L-aspartate transporter (GLAST) and glutamine synthetase (GS) immunoreactions in retinal glia: Evidence for coupling of GLAST and GS in transmitter clearance. J. Neurosci. Res. 1995, 42, 131–143. [Google Scholar] [CrossRef]
- Rauen, T.; Rothstein, J.; Wässle, H. Differential expression of three glutamate transporter subtypes in the rat retina. Cell Tissue Res. 1996, 286, 325–336. [Google Scholar] [CrossRef]
- Gudiño-Cabrera, G.; Ureña-Guerrero, M.E.; Rivera-Cervantes, M.C.; Feria-Velasco, A.I.; Beas-Zárate, C. Excitotoxicity triggered by neonatal monosodium glutamate treatment and blood–brain barrier function. Arch. Med. Res. 2014, 45, 653–659. [Google Scholar] [CrossRef]
- Banerjee, A.; Mukherjee, S.; Maji, B.K. Worldwide flavor enhancer monosodium glutamate combined with high lipid diet provokes metabolic alterations and systemic anomalies: An overview. Toxicol. Rep. 2021, 8, 938–961. [Google Scholar] [CrossRef]
- Nnadozie, J.O.; Chijioke, U.O.; Okafor, O.C.; Olusina, D.B.; Oli, A.N.; Nwonu, P.C.; Mbagwu, H.O.; Chijioke, C.P. Chronic toxicity of low dose monosodium glutamate in albino Wistar rats. BMC Res. Notes 2019, 12, 593. [Google Scholar] [CrossRef] [PubMed]
- Eweka, A.; Igbigbi, P.; Ucheya, R. Histochemical studies of the effects of monosodium glutamate on the liver of adult Wistar rats. Ann. Med. Health Sci. Res. 2011, 1, 21–30. [Google Scholar] [PubMed]
- Pavlovic, V.; Pavlovic, D.; Kocic, G.; Sokolovic, D.; Sarac, M.; Jovic, Z. Ascorbic acid modulates monosodium glutamate induced cytotoxicity in rat thymus. Bratisl. Lek. Listy 2009, 110, 205–209. [Google Scholar] [PubMed]
- Anbarkeh, F.R.; Baradaran, R.; Ghandy, N.; Jalali, M.; Nikravesh, M.R.; Soukhtanloo, M. Effects of monosodium glutamate on apoptosis of germ cells in testicular tissue of adult rat: An experimental study. Int. J. Reprod. Biomed. 2019, 17, 261. [Google Scholar]
- Diab, A.; Hamza, R.Z. Monosodium glutamate induced hepatotoxicity and the possible mitigating effect of vitamin C and propolis. J. Adv. Med. Pharm. Sci. 2016, 7, 1–10. [Google Scholar] [CrossRef]
- Eid, R.A.; Al-Shraim, M.; Zaki, M.S.; Kamar, S.S.; Abdel Latif, N.S.; Negm, S.; Al-Ani, B.; Haidara, M.A. Vitamin E protects against monosodium glutamate-induced acute liver injury and hepatocyte ultrastructural alterations in rats. Ultrastruct. Pathol. 2019, 43, 199–208. [Google Scholar] [CrossRef]
- Farombi, E.; Onyema, O. Monosodium glutamate-induced oxidative damage and genotoxicity in the rat: Modulatory role of vitamin C, vitamin E and quercetin. Hum. Exp. Toxicol. 2006, 25, 251–259. [Google Scholar] [CrossRef]
- Pavlovic, V.; Pavlovic, D.; Kocic, G.; Sokolovic, D.; Jevtovic-Stoimenov, T.; Cekic, S.; Velickovic, D. Effect of monosodium glutamate on oxidative stress and apoptosis in rat thymus. Mol. Cell Biochem. 2007, 303, 161–166. [Google Scholar] [CrossRef]
- Bölükbaş, F.; Öznurlu, Y. The determination of the effect of in ovo administered monosodium glutamate on the embryonic development of thymus and bursa of Fabricius and percentages of alpha-naphthyl acetate esterase positive lymphocyte in chicken. Environ. Sci. Pollut. Res. 2022, 29, 45338–45348. [Google Scholar] [CrossRef]
- Abd-Elkareem, M.; El-Rahman, A.; Mokhless, A.; Khalil, N.S.A.; Amer, A.S. Antioxidant and cytoprotective effects of Nigella sativa L. seeds on the testis of monosodium glutamate challenged rats. Sci. Rep. 2021, 11, 13519. [Google Scholar] [CrossRef]
- Abdul-Hamid, M.; Galaly, S.R.; Ahmed, R.R.; Hamdalla, H.M. Histopathological and biochemical effect of quercetin on monosodium glutamate supplementation-induced testicular toxicity. Beni-Suef Univ. J. Basic Appl. Sci. 2021, 10, 73. [Google Scholar] [CrossRef]
- Hamza, R.Z.; Al-Harbi, M.S. Monosodium glutamate induced testicular toxicity and the possible ameliorative role of vitamin E or selenium in male rats. Toxicol. Rep. 2014, 1, 1037–1045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ali, A.A.; El-Seify, G.H.; El Haroun, H.M.; Soliman, M.A.E.M.M. Effect of monosodium glutamate on the ovaries of adult female albino rats and the possible protective role of green tea. Menoufia Med. J. 2014, 27, 793. [Google Scholar]
- Bojanić, V.; Bojanić, Z.; Najman, S.; Savić, T.; Jakovljević, V.; Najman, S.; Jančić, S. Diltiazem prevention of toxic effects of monosodium glutamate on ovaries in rats. Gen. Physiol. Biophys. 2009, 28, 149–154. [Google Scholar] [PubMed]
- Eweka, A.O.; Eweka, A.; Om’iniabohs, F.A.E. Histological studies of the effects of monosodium glutamate of the fallopian tubes of adult female Wistar rats. N. Am. J. Med. Sci. 2010, 2, 146–149. [Google Scholar] [CrossRef]
- Rohmawati, W.; Istiananingsih, Y.; Nurdiana, N.; Barlianto, W.; Dwijayasa, P.M. Vitamin CE ve Monosodyum Glutamata Bağlı Ovaryan Toksisite. Cukurova Med. J. 2014, 39, 517–524. [Google Scholar] [CrossRef] [Green Version]
- Eweka, A.; Om’Iniabohs, F. Histological studies of the effects of monosodium glutamate on the ovaries of adult wistar rats. Ann. Med. Health Sci. Res. 2011, 1, 37–44. [Google Scholar]
- Ajibade, A.; Fakunle, P.; Adetunji, M. Some effects of monosodium glutamate administration on the histo-architecture of the spleen and pancreas of adult Wistar rats. J. Pharm. Biol. Sci. 2015, 3, 39. [Google Scholar]
- Alsalmi, F.; Hamza, R.; El-Shenawy, N. Effect of Green Tea and Zinc oxide Nanoparticles Complex on Histopathology of Spleen of Male Rats Induced by Monosodium Glutamate. Instant J. Hematol. Oncol. 2019, 1, 4–11. [Google Scholar] [CrossRef]
- Hassan, Z.A.; Arafa, M.H.; Soliman, W.I.; Atteia, H.H.; Al-Saeed, H.F. The effects of monosodium glutamate on thymic and splenic immune functions and role of recovery (biochemical and histological study). J. Cytol. Histol. 2014, 5, 1000283. [Google Scholar] [CrossRef] [Green Version]
- Mohamed, D.S.; Abdelhaliem, N.G.; Zakaria, A.M. Histological and immunohistochemical study of the possible protective effect of ascorbic acid on the toxic effect of monosodium glutamate on the spleen of adult male albino rat. Egypt. J. Histol. 2017, 40, 94–104. [Google Scholar] [CrossRef]
- Al-Ghamdi, F.A. Microscopic study of potential toxic effects of monosodium glutamate on liver of chicken embryos aged 16 days. Egypt. Liver J. 2021, 11, 38. [Google Scholar] [CrossRef]
- Solomon, E.N. Changes in Liver Oxidative Stress Biomarkers, Biochemicals and Histological Assess-ment in Lactating Wistar Rats Following Oral Monosodium Glutamate (MSG) Administration. EC Pharmacol. Toxicol. 2020, 8, 14–25. [Google Scholar]
- Araujo, T.R.; Freitas, I.N.; Vettorazzi, J.F.; Batista, T.M.; Santos-Silva, J.C.; Bonfleur, M.L.; Balbo, S.L.; Boschero, A.C.; Carneiro, E.M.; Ribeiro, R.A. Benefits of L-alanine or L-arginine supplementation against adiposity and glucose intolerance in monosodium glutamate-induced obesity. Eur. J. Nutr. 2017, 56, 2069–2080. [Google Scholar] [CrossRef] [PubMed]
- He, K.; Du, S.; Xun, P.; Sharma, S.; Wang, H.; Zhai, F.; Popkin, B. Consumption of monosodium glutamate in relation to incidence of overweight in Chinese adults: China Health and Nutrition Survey (CHNS). Am. J. Clin. Nutr. 2011, 93, 1328–1336. [Google Scholar] [CrossRef] [Green Version]
- Madhavadas, S.; Kutty, B.M.; Subramanian, S. Amyloid ß Lowering and Cognition Enhancing Effects of Ghrelin Receptor Analog [D-Lys (3)] GHRP-6 in Rat Model of Obesity. NIScPR Publ. 2014, 51, 257–262. [Google Scholar]
- Špolcová, A.; Mikulášková, B.; Holubová, M.; Nagelová, V.; Pirnik, Z.; Zemenová, J.; Haluzík, M.; Železná, B.; Galas, M.-C.; Maletínská, L. Anorexigenic lipopeptides ameliorate central insulin signaling and attenuate tau phosphorylation in hippocampi of mice with monosodium glutamate-induced obesity. J. Alzheimer’s Dis. 2015, 45, 823–835. [Google Scholar] [CrossRef]
- Chaparro-Huerta, V.; Rivera-Cervantes, M.; Torres-Mendoza, B.; Beas-Zarate, C. Neuronal death and tumor necrosis factor-α response to glutamate-induced excitotoxicity in the cerebral cortex of neonatal rats. Neurosci. Lett. 2002, 333, 95–98. [Google Scholar] [CrossRef]
- Moreno, G.; Perelló, M.; Gaillard, R.C.; Spinedi, E. Orexin a stimulates hypothalamic-pituitary-adrenal (HPA) axis function, but not food intake, in the absence of full hypothalamic NPY-ergic activity. Endocrine 2005, 26, 99–106. [Google Scholar] [CrossRef]
- Abass, M.; Abd El-Haleem, M. Evaluation of monosodium glutamate induced neurotoxicity and nephrotoxicity in adult male albino rats. J. Am. Sci. 2011, 7, 264–276. [Google Scholar]
- Al-Qudsi, F.; Al-Jahdali, A. Effect of monosodium glutamate on chick embryo development. J. Am. Sci 2012, 8, 499–509. [Google Scholar]
- Bölükbaş, F.; Öznurlu, Y. Determining the effects of in ovo administration of monosodium glutamate on the embryonic development of brain in chickens. NeuroToxicology 2023, 94, 87–97. [Google Scholar] [CrossRef] [PubMed]
- Dief, A.E.; Kamha, E.S.; Baraka, A.M.; Elshorbagy, A.K. Monosodium glutamate neurotoxicity increases beta amyloid in the rat hippocampus: A potential role for cyclic AMP protein kinase. Neurotoxicology 2014, 42, 76–82. [Google Scholar] [CrossRef] [PubMed]
- Espinar, A.; García-Oliva, A.; Isorna, E.M.; Quesada, A.; Prada, F.A.; Guerrero, J.M. Neuroprotection by melatonin from glutamate-induced excitotoxicity during development of the cerebellum in the chick embryo. J. Pineal Res. 2000, 28, 81–88. [Google Scholar] [CrossRef]
- Hashem, H.E.; Safwat, E.-D.; Algaidi, S. The effect of monosodium glutamate on the cerebellar cortex of male albino rats and the protective role of vitamin C (histological and immunohistochemical study). J. Mol. Histol. 2012, 43, 179–186. [Google Scholar] [CrossRef]
- Miko, A.M.; Shehu, A.M.; Bello, N.; Allyu, I.A.; Tasiu, I.; Abdussalam, A.O.; Isa, A.S. A morphometric study of the teratogenic effect of monosodium glutamate on the developing developing cerebral cortex of Wista Rat(Rattus norvegicus). Niger. J. Sci. Res. 2016, 15, 240–244. [Google Scholar]
- Gurgen, S.G.; Sayın, O.; Çetïn, F.; Sarsmaz, H.Y.; Yazıcı, G.L.N.; Umur, N.; Yucel, A.T. The Effect of Monosodium Glutamate on Neuronal Signaling Molecules in the Hippocampus and the Neuroprotective Effects of Omega-3 Fatty Acids. ACS Chem. Neurosci. 2021, 12, 3028–3037. [Google Scholar] [CrossRef]
- Narayanan, S.N.; Kumar, R.S.; Paval, J.; Nayak, S. Effect of ascorbic acid on the monosodium glutamate-induced neurobehavioral changes in periadolescent rats. Bratisl. Lek Listy 2010, 111, 247–252. [Google Scholar]
- Gim, S.-A.; Park, D.-J.; Kang, J.-B.; Shah, F.-A.; Koh, P.-O. Identification of regulated proteins by resveratrol in glutamate-induced cortical injury of newborn rats. J. Vet. Med. Sci. 2021, 83, 724–733. [Google Scholar] [CrossRef]
- Bellhorn, R.; Lipman, D.; Confino, J.; Burns, M. Effect of monosodium glutamate on retinal vessel development and permeability in rats. Investig. Ophthalmol. Vis. Sci. 1981, 21, 237–247. [Google Scholar]
- Cohen, A.I. An electron microscopic study of the modification by monosodium glutamate of the retinas of normal and “rodless” mice. Am. J. Anat. 1967, 120, 319–355. [Google Scholar] [CrossRef]
- Moustafa, K.A.; Okasha, E.F. The possible protective effect of curcumin on monosodium glutamate-induced retinal changes in adult male albino rats. Egypt. J. Histol. 2016, 39, 87–95. [Google Scholar] [CrossRef]
- Szabadfi, K.; Atlasz, T.; Horváth, G.; Kiss, P.; Hamza, L.; Farkas, J.; Tamás, A.; Lubics, A.; Gábriel, R.; Reglődi, D. Early postnatal enriched environment decreases retinal degeneration induced by monosodium glutamate treatment in rats. Brain Res. 2009, 1259, 107–112. [Google Scholar] [CrossRef] [PubMed]
- Zaghlool, S.S.; Hanaf, L.K.; Afifi, N.M.; Ibrahim, E.R. Histological and immunohistochemical study on the protective effect of Ginkgo biloba extract against glutamate-induced neurotoxicity in male albino rat retinal cells. Egypt. J. Histol. 2012, 35, 176–188. [Google Scholar] [CrossRef]
- Dénes, V.; Lakk, M.; Czotter, N.; Gábriel, R. A precise temporal dissection of monosodium glutamate-induced apoptotic events in newborn rat retina in vivo. Neurochem. Res. 2011, 36, 1464–1474. [Google Scholar] [CrossRef]
- Van Rijn, C.; Marani, E.; Rietveld, W. The neurotoxic effect of monosodium glutamate (MSG) on the retinal ganglion cells of the albino rat. Histol. Histopathol. 1986, 1, 3. [Google Scholar]
- Praputpittaya, C.; Wililak, A. Visual Performance in Monosodium Glutamate-Treated Rats. Nutr. Neurosci. 2003, 6, 301–307. [Google Scholar] [CrossRef] [PubMed]
- Swelim, H.H. Monosodium glutamate (MSG) induced retinopath y in adult and neonate CD-1 mice. Egypt J. Med. Lab Sci. 2004, 13, 45–71. [Google Scholar]
- Horvath, G.; Reglodi, D.; Vadasz, G.; Farkas, J.; Kiss, P. Exposure to enriched environment decreases neurobehavioral deficits induced by neonatal glutamate toxicity. Int. J. Mol. Sci. 2013, 14, 19054–19066. [Google Scholar] [CrossRef] [Green Version]
- Kiss, P.; Hauser, D.; Tamas, A.; Lubics, A.; Racz, B.; Horvath, Z.; Farkas, J.; Zimmermann, F.; Stepien, A.; Lengvari, I. Changes in open-field activity and novelty-seeking behavior in periadolescent rats neonatally treated with monosodium glutamate. Neurotox. Res. 2007, 12, 85–93. [Google Scholar] [CrossRef]
- El-Sayyad, H.; Abou-El-Naga, A.; Khalifa, S.; El-Shahari, E.; Jala, H. Abnormal Retinal Structure and Function of Mother Wistar Rats Supplemented Aspartame, Glutamate and Galactose. J. Drug Metab. Toxicol. 2016, 7, 2. [Google Scholar] [CrossRef] [Green Version]
- Alshareef, M.; Alrafiah, A.; Abed, S.; Basingab, F.; Alrofaidi, A. Effect of e-cigarette flavoring agents on the neural retina of chick embryo: Histological and gene expression study. Folia Histochem. Et Cytobiol. 2021, 59, 245–258. [Google Scholar] [CrossRef] [PubMed]
- Wisely, C.E.; Sayed, J.A.; Tamez, H.; Zelinka, C.; Abdel-Rahman, M.H.; Fischer, A.J.; Cebulla, C.M. The chick eye in vision research: An excellent model for the study of ocular disease. Prog. Retin. Eye Res. 2017, 61, 72–97. [Google Scholar] [CrossRef] [PubMed]
- Airaodion, A.I.; Ogbuagu, E.O.; Osemwowa, E.U.; Ogbuagu, U.; Esonu, C.E. Toxicological Effect of Monosodium Glutamate in Seasonings on Human Health. Glob. J. Nutri. Food Sci. 2019, 1, 522. [Google Scholar] [CrossRef]
- Gao, J.; Wu, J.; Zhao, X.; Zhang, W.; Zhang, Y.; Zhang, Z. Transplacental neurotoxic effects of monosodium glutamate on structures and functions of specific brain areas of filial mice. Sheng Li Xue Bao Acta Physiol. Sin. 1994, 46, 44–51. [Google Scholar]
- Hamburger, V.; Hamilton, H.L. A series of normal stages in the development of the chick embryo. J. Morphol. 1951, 88, 49–92. [Google Scholar] [CrossRef]
- Crossmon, G. A modification of Mallory’s connective tissue stain with a discussion of the principles involved. Anat. Rec. 1937, 69, 33–38. [Google Scholar] [CrossRef]
- Culling, C.F.A.; Allison, R.T.; Barr, W.T. Cellular Pathology Technique, 4th ed.; Culling, C.F.A., Allison, R.T., Barr, W.T., Eds.; Butterworth-Heinemann: Woburn, MA, USA, 1985; pp. 164–180, 214–255, 377–407. [Google Scholar]
- Geha, R.S.; Beiser, A.; Ren, C.; Patterson, R.; Greenberger, P.A.; Grammer, L.C.; Ditto, A.M.; Harris, K.E.; Shaughnessy, M.A.; Yarnold, P.R.; et al. Review of alleged reaction to monosodium glutamate and outcome of a multicenter double-blind placebo-controlled study. J. Nut.r 2000, 130, 1058s–1062s. [Google Scholar] [CrossRef] [Green Version]
- Roongruangchai, J.; Viravud, Y.; Plakornkul, V.; Sripaoraya, K.; Boonmark, W.; Roongruangchai, K. The teratogenic effects of monosodium glutamate (MSG) on the development of chick embryos. Siriraj Med. J. 2018, 70, 514–522. [Google Scholar]
- Hajihasani, M.M.; Soheili, V.; Zirak, M.R.; Sahebkar, A.; Shakeri, A. Natural products as safeguards against monosodium glutamate-induced toxicity. Iran. J. Basic Med. Sci. 2020, 23, 416. [Google Scholar] [CrossRef]
- Kazmi, Z.; Fatima, I.; Perveen, S.; Malik, S.S. Monosodium glutamate: Review on clinical reports. Int. J. Food Prop. 2017, 20, 1807–1815. [Google Scholar] [CrossRef] [Green Version]
- Baad-Hansen, L.; Cairns, B.; Ernberg, M.; Svensson, P. Effect of systemic monosodium glutamate (MSG) on headache and pericranial muscle sensitivity. Cephalalgia Int. J. Headache 2010, 30, 68–76. [Google Scholar] [CrossRef] [PubMed]
- Obayashi, Y.; Nagamura, Y. Does monosodium glutamate really cause headache? A systematic review of human studies. J. Headache Pain. 2016, 17, 54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abu Elnaga, N.A.; Sarhan, M.; Mansour, H. Teratogenicity of Monosodium Glutamate on The Pregnant Rats and Their Fetuses. Egypt. J. Hosp. Med. 2019, 74, 1737–1747. [Google Scholar] [CrossRef]
- Umukoro, S.; Oluwole, G.O.; Olamijowon, H.E.; Omogbiya, A.I.; Eduviere, A.T. Effect of monosodium glutamate on behavioral phenotypes, biomarkers of oxidative stress in brain tissues and liver enzymes in mice. World J. Neurosci. 2015, 5, 339. [Google Scholar] [CrossRef] [Green Version]
- Boutry, C.; Matsumoto, H.; Airinei, G.; Benamouzig, R.; Tomé, D.; Blachier, F.; Bos, C. Monosodium glutamate raises antral distension and plasma amino acid after a standard meal in humans. Am. J. Physiol.-Gastrointest. Liver Physiol. 2011, 300, G137–G145. [Google Scholar] [CrossRef]
- Adebayo, O. Lipid peroxidation and antioxidant status of the cerebrum, cerebellum and brain stem following dietary monosodium glutamate administration in mice. Asian J. Clin. Nutr. 2011, 3, 71–77. [Google Scholar] [CrossRef]
- Ohguro, H.; Katsushima, H.; Maruyama, I.; Maeda, T.; Yanagihashi, S.; Metoki, T.; Nakazawa, M. A high dietary intake of sodium glutamate as flavoring (ajinomoto) causes gross changes in retinal morphology and function. Exp. Eye Res. 2002, 75, 307–315. [Google Scholar] [CrossRef]
- Gu, L.; Liang, X.; Wang, L.; Yan, Y.; Ni, Z.; Dai, H.; Gao, J.; Mou, S.; Wang, Q.; Chen, X. Functional metabotropic glutamate receptors 1 and 5 are expressed in murine podocytes. Kidney Int. 2012, 81, 458–468. [Google Scholar] [CrossRef] [Green Version]
- El-Gohari, K.M.; Bahei-Eldin, I.A.; Habib, E.M.; Saad, S.A.; Rady, H.Y.; Said, A.M. Neuroprotection of the rat’s retinal ganglion cells against glutamate-induced toxicity. J. Egypt. Ophthalmol. Soc. 2016, 109, 135. [Google Scholar]
- Ali, H.S.; El-Gohary, A.A.; Metwally, F.G.; Sabra, N.M.; El Sayed, A.A. Mono sodium glutamate induced damage in rabbit retina: Electroretinographic and histologic studies. Glob. J Pharm. 2012, 6, 148–159. [Google Scholar]
- AlThanoon, S.A.; Abd, A.A. Histopathological Changes Induced by Monosodium Glutamate and Sodium Nitrite in the Development of Eye in Albino Mice Mus musculus. J. Educ. Sci. 2021, 30, 3. [Google Scholar] [CrossRef]
- Öznurlu, Y.; Özaydın, T.; Sur, E.; Özparlak, H. The effects of in ovo administered bisphenol A on tibial growth plate histology in chicken. Birth Defects Res. 2021; in press. [Google Scholar] [CrossRef]
- Bölükbaş, F.; Öznurlu, Y. Yumurtaya verilen monosodyum glutamat’ın tavuk embriyolarında medulla spinalisin servikal bölgesinin embriyonik gelişimi üzerindeki etkilerinin belirlenmesi. J. Adv. VetBio Sci. Tech. 2021, 6, 298–311. [Google Scholar] [CrossRef]
- Atallah, M.N.; Badawy, G.M.; El–Garawani, I.M.; Abdallah, F.S.; El–Borm, H.T. Neurotoxic effect of nalufin on the histology, ultrastructure, cell cycle and apoptosis of the developing chick embryo and its amelioration by selenium. Food Chem. Toxicol. 2021, 158, 112693. [Google Scholar] [CrossRef]
- Kmecick, M.; Vieira da Costa, M.C.; Oliveira Ribeiro, C.A.; Ortolani-Machado, C.F. Morphological evidence of neurotoxic effects in chicken embryos after exposure to perfluorooctanoic acid (PFOA) and inorganic cadmium. Toxicology 2019, 427, 152286. [Google Scholar] [CrossRef] [PubMed]
- Hocking, P.M.; Guggenheim, J.A. The chick as an animal model of eye disease. Drug Discov. Today Dis. Model. 2013, 10, e225–e230. [Google Scholar] [CrossRef]
- Vergara, M.N.; Canto-Soler, M.V. Rediscovering the chick embryo as a model to study retinal development. Neural. Dev. 2012, 7, 22. [Google Scholar] [CrossRef] [Green Version]
- Trejo-Reveles, V.; McTeir, L.; Summers, K.; Rainger, J. An analysis of anterior segment development in the chicken eye. Mech. Dev. 2018, 150, 42–49. [Google Scholar] [CrossRef]
- Berg, C.; Halldin, K.; Brunström, B. Effects of bisphenol A and tetrabromobisphenol A on sex organ development in quail and chicken embryos. Environ. Toxicol. Chem. Int. J. 2001, 20, 2836–2840. [Google Scholar] [CrossRef]
- Jessl, L.; Lenz, R.; Massing, F.G.; Scheider, J.; Oehlmann, J. Effects of estrogens and antiestrogens on gonadal sex differentiation and embryonic development in the domestic fowl (Gallus gallus domesticus). PeerJ 2018, 6, e5094. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, S.B.; Ang, H.-P.; Poh, R.; Chaurasia, S.S.; Peh, G.; Liu, J.; Tan, D.T.; Vithana, E.N.; Mehta, J.S. Mice with a targeted disruption of Slc4a11 model the progressive corneal changes of congenital hereditary endothelial dystrophy. Investig. Ophthalmol. Vis. Sci. 2013, 54, 6179–6189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, Y.; Huang, G.; Huang, W.; He, M. Distribution of central and peripheral corneal thickness in Chinese children and adults: The Guangzhou twin eye study. Cornea 2008, 27, 776–781. [Google Scholar] [CrossRef] [PubMed]
- Nemesure, B.; Wu, S.-Y.; Hennis, A.; Leske, M.C.; Group, B.E.S. Corneal thickness and intraocular pressure in the Barbados eye studies. Arch. Ophthalmol. 2003, 121, 240–244. [Google Scholar] [CrossRef]
- Ueno, Y.; Hiraoka, T.; Miyazaki, M.; Ito, M.; Oshika, T. Corneal thickness profile and posterior corneal astigmatism in normal corneas. Ophthalmology 2015, 122, 1072–1078. [Google Scholar] [CrossRef]
- Inomata, T.; Mashaghi, A.; Hong, J.; Nakao, T.; Dana, R. Scaling and maintenance of corneal thickness during aging. PloS ONE 2017, 12, e0185694. [Google Scholar] [CrossRef] [Green Version]
- Al-Qudsi, F.; Azzouz, S. Effect of electromagnetic mobile radiation on chick embryo development. Life Sci. J. 2012, 9, 983–991. [Google Scholar]
- Blanks, J.C.; Reif-Lehrer, L.; Casper, D. Effects of monosodium glutamate on the isolated retina of the chick embryo as a function of age: A morphological study. Exp. Eye Res. 1981, 32, 105–124. [Google Scholar] [CrossRef]
- Reif-Lehrer, L.; Bergenthal, J.; Hanninen, L. Effects of monosodium glutamate on chick embryo retina in culture. Investig. Ophthalmol. Vis. Sci. 1975, 14, 114–124. [Google Scholar]
- Kujawa-Hadryś, M.; Tosik, D.; Bartel, H. Changes in thickness of each layer of developing chicken cornea after administration of caffeine. Folia Histochem. Cytobiol. 2010, 48, 273–277. [Google Scholar] [CrossRef] [Green Version]
- Moayed, A.A.; Hariri, S.; Song, E.S.; Choh, V.; Bizheva, K. In vivo volumetric imaging of chicken retina with ultrahigh-resolution spectral domain optical coherence tomography. Biomed. Opt. Express 2011, 2, 1268–1274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zareen, N.; Khan, M.; Minhas, L. Histological stages of retinal morphogenesis in chicken–a descriptive laboratory research. Ital. J. Zool. 2011, 78, 45–52. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Eye histological section in Group I on 15th day of incubation (A); 18th day of incubation (B); 21st day of incubation (C,D); retinal layers (D); OPN: optic nerve, thick arrow: sclera with visible hyaline cartilage; arrow: Ganglion cells. Toluidine blue (B,C) and H&E (A,D) stains. Bar: 500 μm (A–C). Bar: 50 μm (D).
Figure 1.
Eye histological section in Group I on 15th day of incubation (A); 18th day of incubation (B); 21st day of incubation (C,D); retinal layers (D); OPN: optic nerve, thick arrow: sclera with visible hyaline cartilage; arrow: Ganglion cells. Toluidine blue (B,C) and H&E (A,D) stains. Bar: 500 μm (A–C). Bar: 50 μm (D).

Figure 2.
Comparison of chicken corneal layer thicknesses among Group I, II, III, IV, and V on the 15th (A–E), 18th (F–J), and 21st (K–O) days of incubation. Groups I (A,F,K), II (B,G,L), III (C,H,M), IV (D,I,N), and V (E,J,O) on days 15, 18, and 21 of incubation, respectively. Arrowhead: endothelium; E→: epithelium, S: stroma; stars (*): detachment of the epithelium layers. Crossmon’s triple staining. Bar: 100 μm.
Figure 2.
Comparison of chicken corneal layer thicknesses among Group I, II, III, IV, and V on the 15th (A–E), 18th (F–J), and 21st (K–O) days of incubation. Groups I (A,F,K), II (B,G,L), III (C,H,M), IV (D,I,N), and V (E,J,O) on days 15, 18, and 21 of incubation, respectively. Arrowhead: endothelium; E→: epithelium, S: stroma; stars (*): detachment of the epithelium layers. Crossmon’s triple staining. Bar: 100 μm.

Figure 3.
Comparison of chicken retina layer thicknesses (μm) among Group I, Group II, Group III, Group IV, and Group V on the 15th day of incubation in chicken embryos. Retinal thickness (A); thickness of RPE (B); thickness of PR (C); thickness of ONL (D); thickness of OPL (E); thickness of INL (F); thickness of IPL (G); thickness of GL (H); thickness of NFL (I). (a–c) Various superscript letters on the columns indicate a statistical difference (mean ± SD, p < 0.05).
Figure 3.
Comparison of chicken retina layer thicknesses (μm) among Group I, Group II, Group III, Group IV, and Group V on the 15th day of incubation in chicken embryos. Retinal thickness (A); thickness of RPE (B); thickness of PR (C); thickness of ONL (D); thickness of OPL (E); thickness of INL (F); thickness of IPL (G); thickness of GL (H); thickness of NFL (I). (a–c) Various superscript letters on the columns indicate a statistical difference (mean ± SD, p < 0.05).

Figure 4.
Sections from the retina of the Groups I (A), II (B), III (C), IV (D), and V (E) at day 15 of incubation. Blue stars (*): vacuole formation in the IPL, INL, GL, and NFL layers; black stars (*): retinal pigment epithelium detachment; arrow: ganglion cells; S: scleral cartilage; C: choroid; retinal pigment epithelium (RPE); nerve fibre layer (NFL); ganglion cell layer (GL); inner plexiform layer (IPL); inner nuclear layer (INL); outer plexiform layer (OPL); outer nuclear layer (ONL). H&E staining.
Figure 4.
Sections from the retina of the Groups I (A), II (B), III (C), IV (D), and V (E) at day 15 of incubation. Blue stars (*): vacuole formation in the IPL, INL, GL, and NFL layers; black stars (*): retinal pigment epithelium detachment; arrow: ganglion cells; S: scleral cartilage; C: choroid; retinal pigment epithelium (RPE); nerve fibre layer (NFL); ganglion cell layer (GL); inner plexiform layer (IPL); inner nuclear layer (INL); outer plexiform layer (OPL); outer nuclear layer (ONL). H&E staining.

Figure 5.
Sections from the retina of Groups I (A), II (B), III (C), IV (D), and V (E) at incubation day 18. Blue stars (*): vacuole formation in the PR, INL, IPL, and GL layer. Retinal pigment epithelium (RPE); photoreceptor layer (PR); nerve fibre layer (NFL); ganglion cell layer (GL); inner plexiform layer (IPL); inner nuclear layer (INL); outer plexiform layer (OPL); outer nuclear layer (ONL). H&E staining.
Figure 5.
Sections from the retina of Groups I (A), II (B), III (C), IV (D), and V (E) at incubation day 18. Blue stars (*): vacuole formation in the PR, INL, IPL, and GL layer. Retinal pigment epithelium (RPE); photoreceptor layer (PR); nerve fibre layer (NFL); ganglion cell layer (GL); inner plexiform layer (IPL); inner nuclear layer (INL); outer plexiform layer (OPL); outer nuclear layer (ONL). H&E staining.

Figure 6.
Comparison of retinal layer thicknesses (μm) among Groups I, II, III, IV, and V on incubation day 18 in chicken embryos. Retinal thickness (A); thickness of RPE (B); thickness of PR (C); thickness of ONL (D); thickness of OPL (E); thickness of INL (F); thickness of IPL (G); thickness of GL (H); and thickness of NFL (I). (a–c) Various superscript letters on the columns indicate a statistical difference (mean ± SD, p < 0.05).
Figure 6.
Comparison of retinal layer thicknesses (μm) among Groups I, II, III, IV, and V on incubation day 18 in chicken embryos. Retinal thickness (A); thickness of RPE (B); thickness of PR (C); thickness of ONL (D); thickness of OPL (E); thickness of INL (F); thickness of IPL (G); thickness of GL (H); and thickness of NFL (I). (a–c) Various superscript letters on the columns indicate a statistical difference (mean ± SD, p < 0.05).

Figure 7.
Sections from the retina of Groups I (A), II (B), III (C), IV (D), and V (E) at incubation day 21. Blue stars (*): vacuole formation in the INL, IPL, and NFL layers. Retinal pigment epithelium (RPE); photoreceptor layer (PR); nerve fibre layer (NFL); ganglion cell layer (GL); inner plexiform layer (IPL); inner nuclear layer (INL); outer plexiform layer (OPL); outer nuclear layer (ONL). H&E staining.
Figure 7.
Sections from the retina of Groups I (A), II (B), III (C), IV (D), and V (E) at incubation day 21. Blue stars (*): vacuole formation in the INL, IPL, and NFL layers. Retinal pigment epithelium (RPE); photoreceptor layer (PR); nerve fibre layer (NFL); ganglion cell layer (GL); inner plexiform layer (IPL); inner nuclear layer (INL); outer plexiform layer (OPL); outer nuclear layer (ONL). H&E staining.

Figure 8.
Comparison of retina layer thicknesses (μm) among Groups I, II, III, IV, and V on incubation day 21 in chicken embryos. Retinal thickness (A); thickness of RPE (B); thickness of PR (C); thickness of ONL (D); thickness of OPL (E); thickness of INL (F); thickness of IPL (G); thickness of GL (H); thickness of NFL (I). (a–c) Various superscript letters on the columns indicate a statistical difference (mean ± SD, p < 0.05).
Figure 8.
Comparison of retina layer thicknesses (μm) among Groups I, II, III, IV, and V on incubation day 21 in chicken embryos. Retinal thickness (A); thickness of RPE (B); thickness of PR (C); thickness of ONL (D); thickness of OPL (E); thickness of INL (F); thickness of IPL (G); thickness of GL (H); thickness of NFL (I). (a–c) Various superscript letters on the columns indicate a statistical difference (mean ± SD, p < 0.05).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Corneal epithelium thickness (µm) and total corneal thickness (µm) on days 15, 18, and 21 of incubation.
Table 1.
Corneal epithelium thickness (µm) and total corneal thickness (µm) on days 15, 18, and 21 of incubation.
| Groups n = 6 | Day 15 of Incubation | Day 18 of Incubation | Day 21 of Incubation | |||
|---|---|---|---|---|---|---|
| Thickness of epithelium | Total corneal thickness | Thickness of epithelium | Total corneal thickness | Thickness of epithelium | Total corneal thickness | |
| Group I | 25.22 ± 2.02 a | 328.42 ± 30.41 a | 29.68 ± 2.54 a | 416,70 ± 5.62 a | 33.12 ± 3.35 a | 417.85 ± 12.98 a |
| Group II | 24.99 ± 2.18 a | 318.59 ± 5.59 a | 29.24 ± 1.19 a | 413.51 ± 10.18 a | 33.33 ± 6.11 a | 413.05 ± 13.44 a |
| Group III | 15.96 ± 1.67 b | 249.22 ± 14.60 b | 24.72 ± 1.80 b | 308.73 ± 14.68 b | 27.16 ± 1.77 b | 377.58 ± 21.98 b |
| Group IV | 13.15 ± 1.71 c | 222.85 ± 21.31 c | 22.82 ± 1.15 c | 295.67 ± 7.16 c | 26.30 ± 1.45 b | 365.91 ± 13.43 bc |
| Group V | 13.71 ± 1.61 c | 210.04 ± 17.15 c | 22.22 ± 1.86 c | 291.36 ± 6.39 c | 24.92 ± 2.34 b | 359.04 ± 11.60 c |
(a–c) Various superscript letters on the columns indicate a statistical difference (mean ± SD, p < 0.05).
Table 2.
Ganglion cell numbers of the granular layer on days 15, 18, and 21 of incubation.
| Ganglion Cell Numbers | |||
|---|---|---|---|
| Groups n = 6 | Day 15 of incubation | Day 18 of incubation | Day 21 of incubation |
| Group I | 22.58 ± 2.91 a | 21.47 ± 3.21 a | 18.94 ± 4.86 a |
| Group II | 21.01 ± 3.28 a | 22,52 ± 4.32 a | 17.58 ± 3.07 a |
| Group III | 9.71 ± 3.56 b | 12.74 ± 2.93 b | 10.84 ± 4.36 b |
| Group IV | 7.66 ± 3.04 b | 9.11 ± 2.76 c | 10.33 ± 1.93 b |
| Group V | 7.44 ± 1.56 b | 8.86 ± 2.94 c | 8.27 ± 2.78 b |
(a–c) Various superscript letters on the columns indicate a statistical difference (mean ± SD, p < 0.05). Ganglion cell numbers were determined by counting the number on a 100 μm line length in three different fields per section.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Bölükbaş, F.; Öznurlu, Y. Investigation of the Effects of Monosodium Glutamate on the Embryonic Development of the Eye in Chickens. Vet. Sci. 2023, 10, 99. https://doi.org/10.3390/vetsci10020099
AMA Style
Bölükbaş F, Öznurlu Y. Investigation of the Effects of Monosodium Glutamate on the Embryonic Development of the Eye in Chickens. Veterinary Sciences. 2023; 10(2):99. https://doi.org/10.3390/vetsci10020099
Chicago/Turabian StyleBölükbaş, Ferhan, and Yasemin Öznurlu. 2023. "Investigation of the Effects of Monosodium Glutamate on the Embryonic Development of the Eye in Chickens" Veterinary Sciences 10, no. 2: 99. https://doi.org/10.3390/vetsci10020099
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.