
K-Ras Peptide Mimotope Induces Antigen Specific Th1 and B-Cell Immune Responses against G12A-Mutated K-Ras Antigen in Balb/c Mice


 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacteria Strains, Plasmids and Growth Conditions
2.2. Cloning and Expression of Recombinant L. lactis
2.3. Protein Extraction and Western Blotting
2.4. Ni-NTA-HRP ELISA Specific Quantification of K-Ras Mimotope
2.5. Mice Strains
2.6. Oral Immunization
2.7. Detection of Recombinant L. lactis from Mice Feces
2.8. Isolation of Blood, Sera, Intestinal Wash Samples and Splenocytes
2.9. Immunophenotyping of T and B-Cell Populations
2.10. Detection of K-Ras-Specific Serum IgG and Intestinal IgA
2.11. Ex Vivo Antigen Stimulation and Detection of IFN-γ Producing T-Cells
2.12. Ethics Approval and Consent to Participate
3. Results
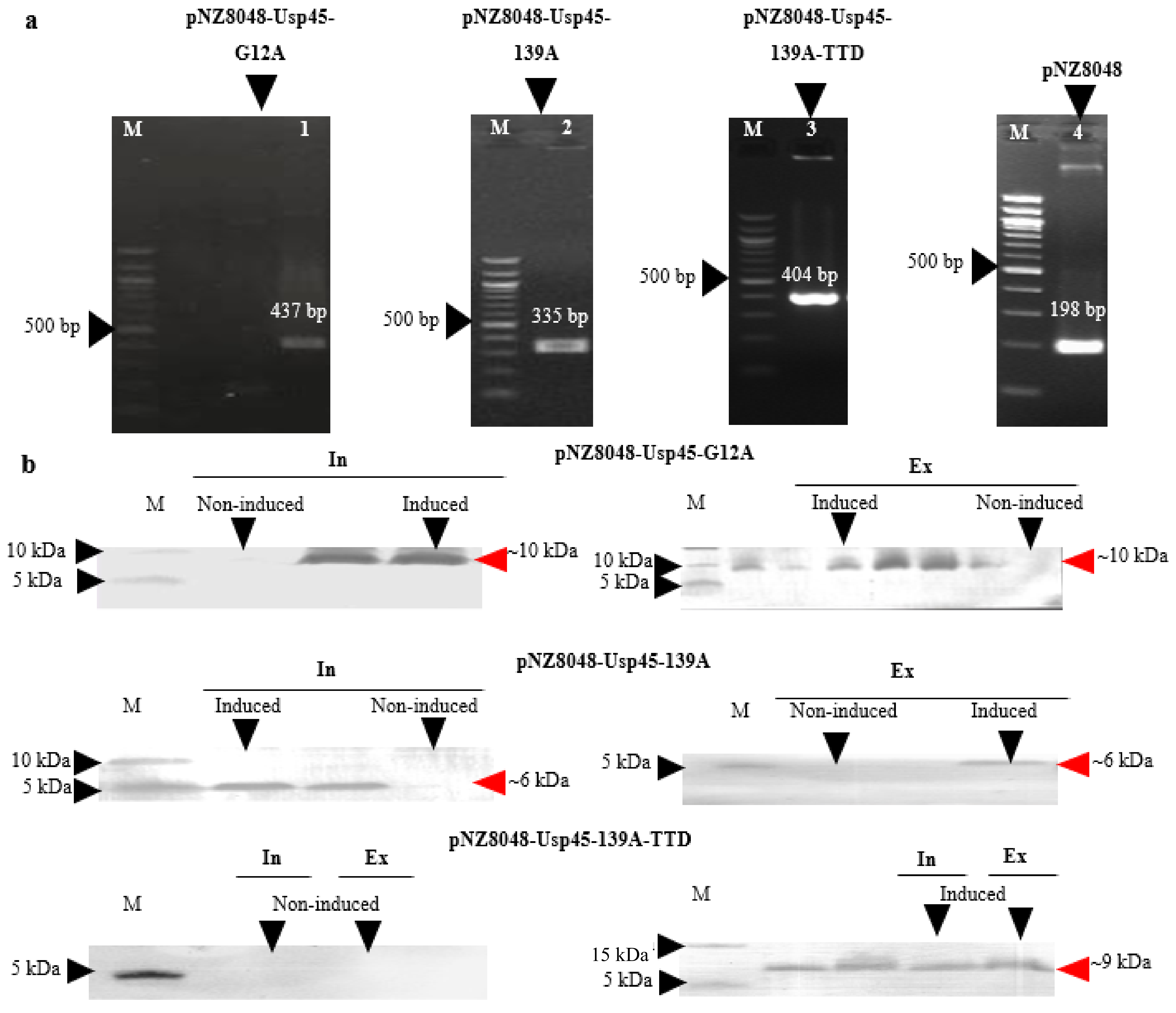
3.1. Expression and Extracellular Secretion of K-Ras Peptide Mimotopes by L. lactis NZ9000
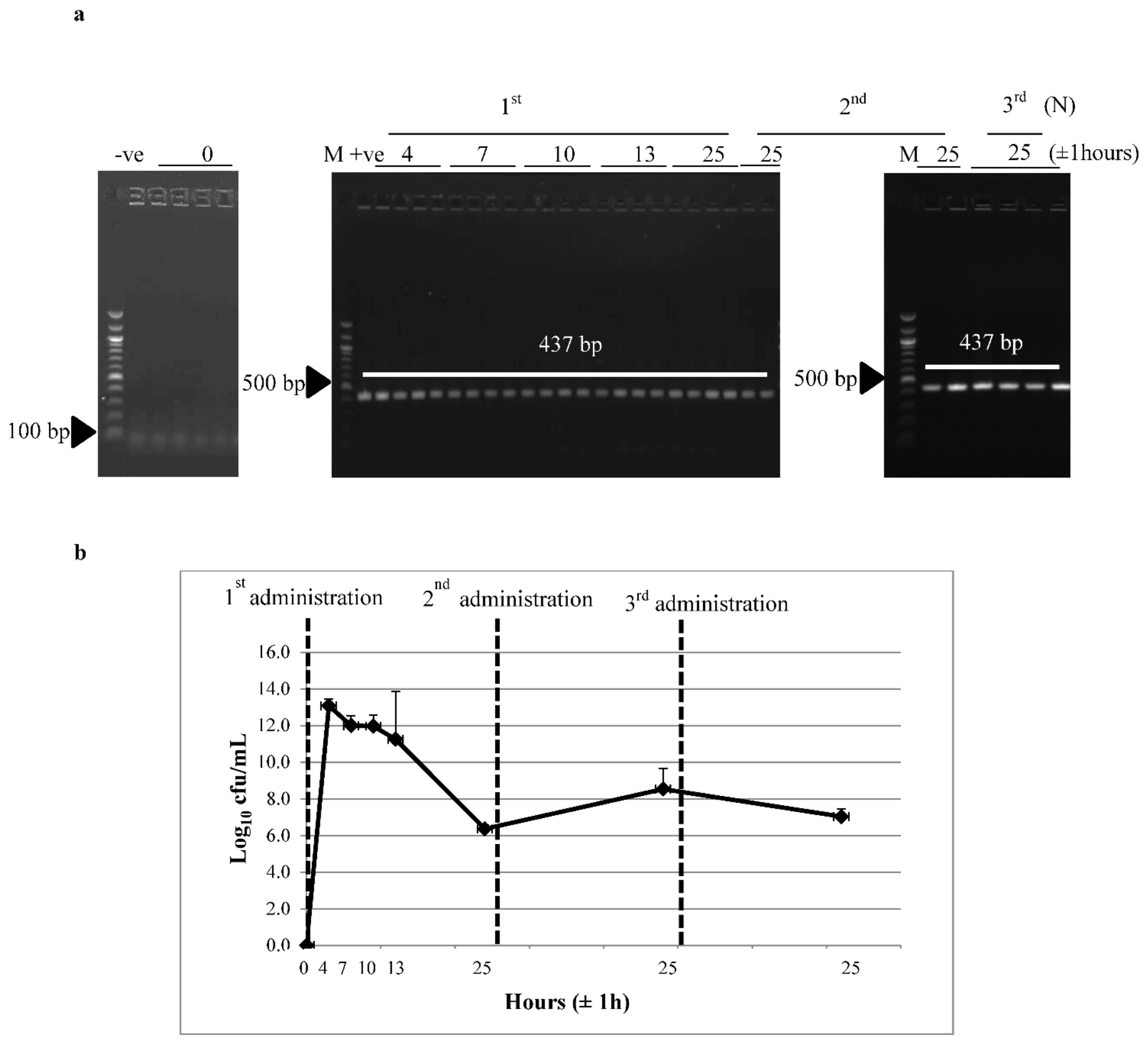
3.2. Survivability of Recombinant L. lactis through the GI Tract
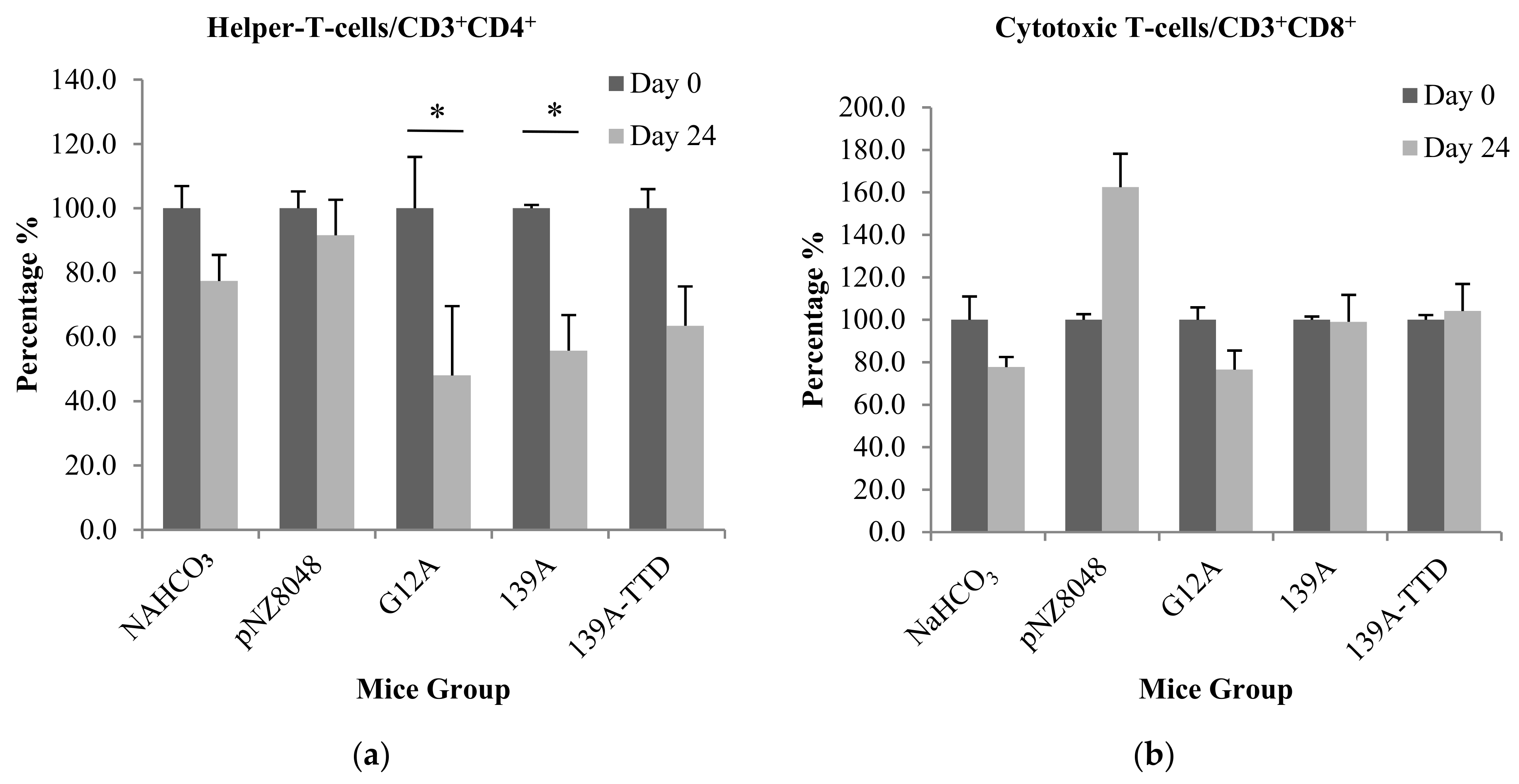
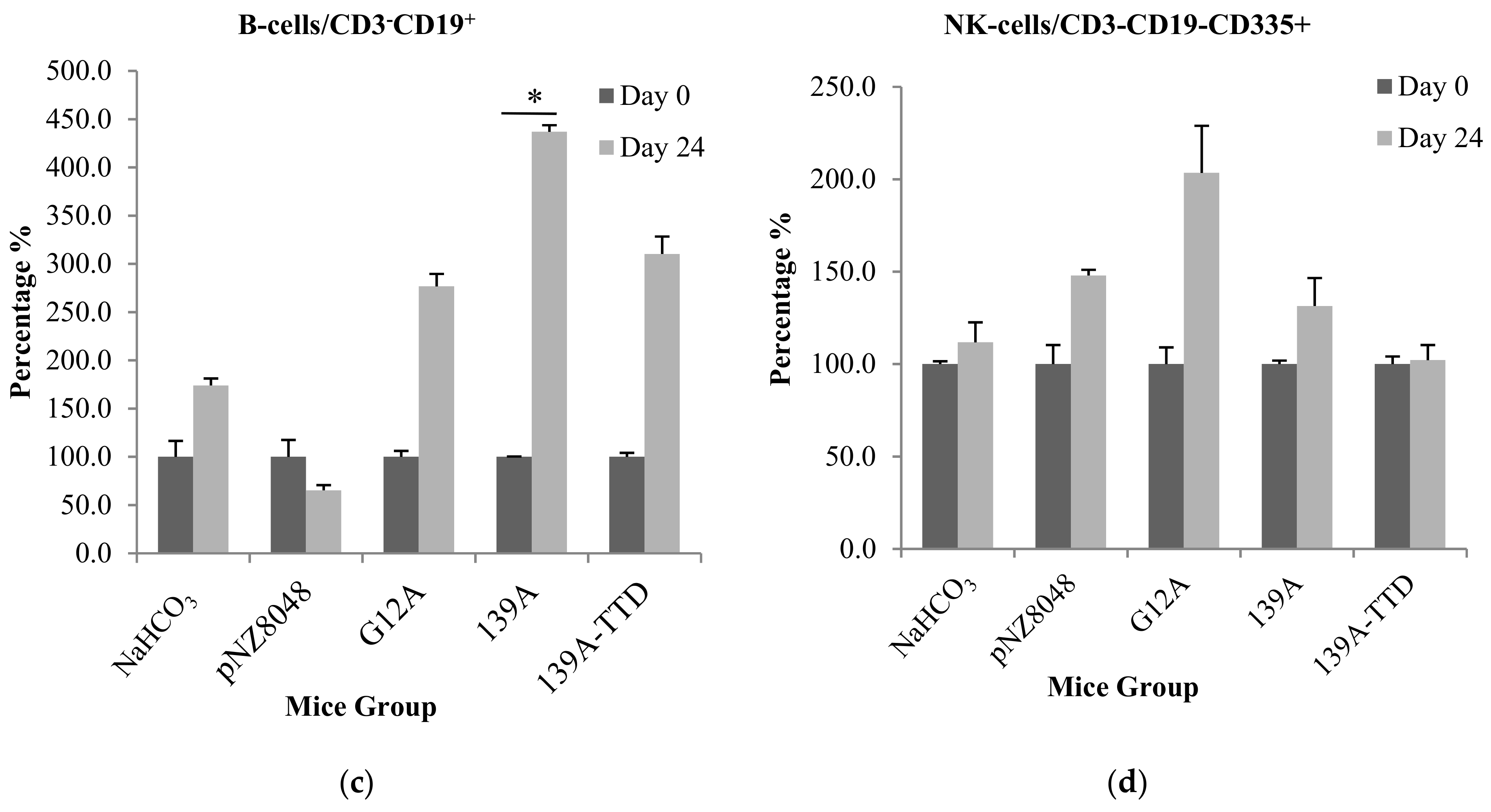
3.3. Oral Immunization of 139A-Secreting L. lactis Induces Elevation of B-Cell Population
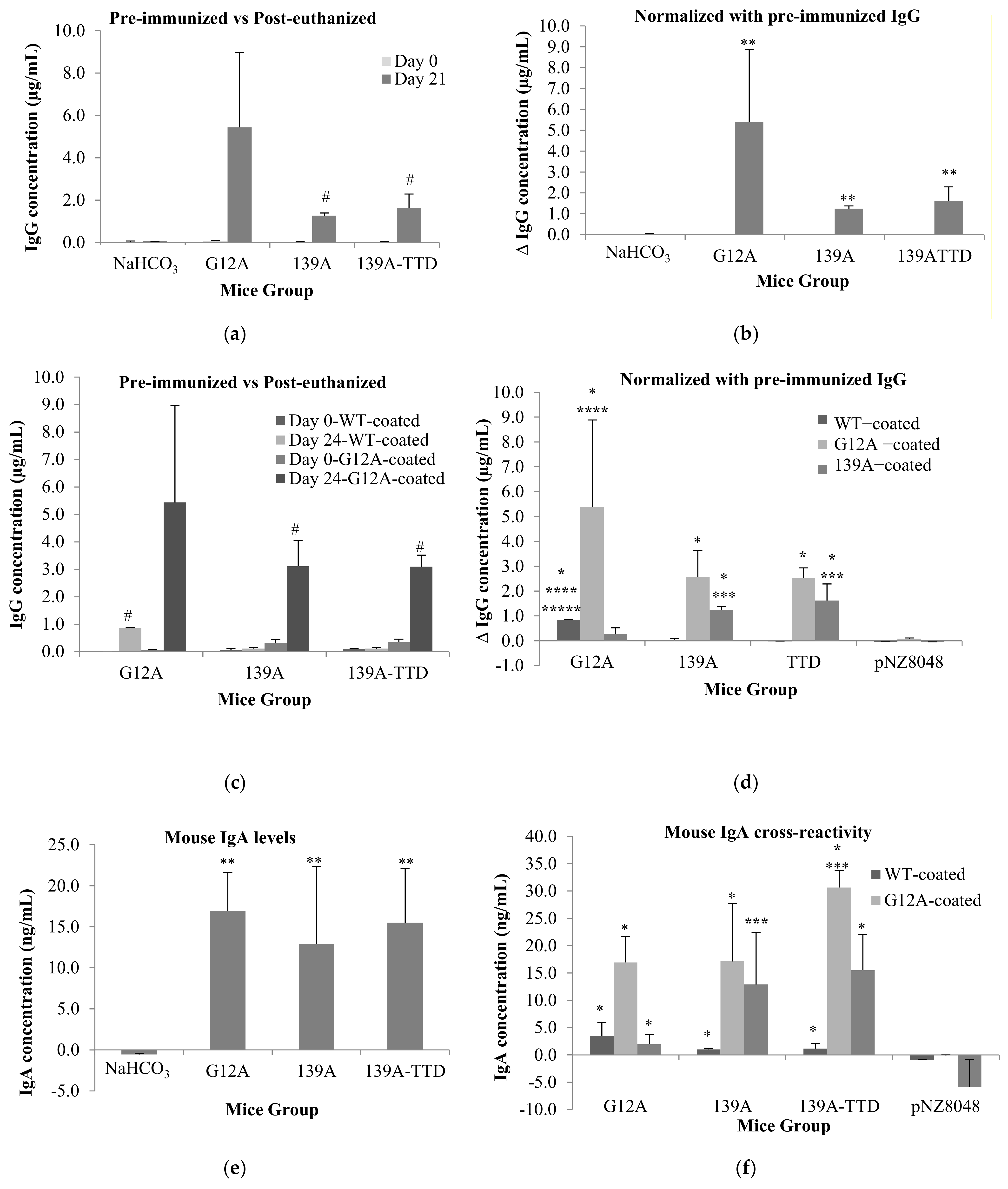
3.4. Recombinant L. lactis Secreting 139A-TTD Enhances G12A-Mutant K-Ras-Specific Humoral Response in BALB/c Mice
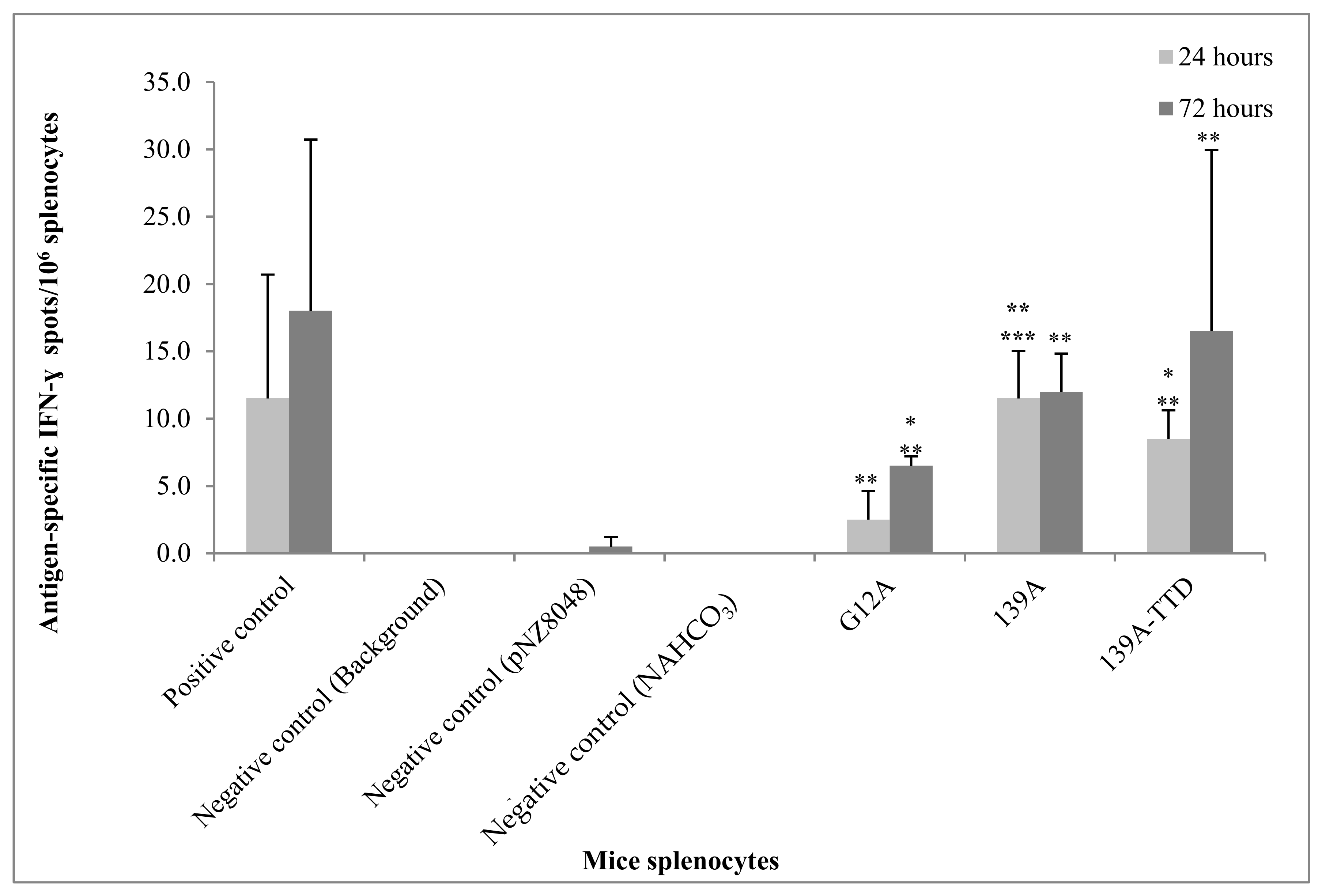
3.5. Re-Stimulation of 139A-TTD Immunized Mice Splenocytes Induces G12A-Mutant K-Ras-Specific T-Cell Activation
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Jančík, S.; Drabek, J.; Radzioch, D.; Hajduch, M. Clinical relevance of KRAS in human cancers. J. Biomed. Biotechnol. 2010, 2010, 150960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajasekharan, S.K.; Raman, T. Ras and Ras mutations in cancer. Cent. Eur. J. Biol. 2013, 8, 609–624. [Google Scholar] [CrossRef] [Green Version]
- Van Krieken, J.H.; Jung, A.; Kirchner, T.; Carneiro, F.; Seruca, R.; Bosman, F.T.; Quirke, P.; Fléjou, J.F.; Hansen, T.P.; De Hertogh, G.; et al. KRAS mutation testing for predicting response to anti-EGFR therapy for colorectal carcinoma: Proposal for an European quality assurance program. Virchows Arch. 2008, 453, 417–431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allegra, C.J.; Jessup, J.M.; Somerfield, M.R.; Hamilton, S.R.; Hammond, E.H.; Hayes, D.F.; McAllister, P.K.; Morton, R.F.; Schilsky, R.L. American Society of Clinical Oncology provisional clinical opinion: Testing for KRAS gene mutations in patients with metastatic colorectal carcinoma to predict response to anti-epidermal growth factor receptor monoclonal antibody therapy. J. Clin. Oncol. 2009, 27, 2091–2096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, C.; Du, X. KRAS mutation testing in metastatic colorectal cancer. World J. Gastroenterol. 2012, 18, 5171–5180. [Google Scholar]
- Tong, J.H.; Lung, R.W.; Sin, F.M.; Law, P.P.; Kang, W.; Chan, A.W.; Ma, B.B.; Mak, T.W.; Ng, S.S.; To, K.F. Characterization of rare transforming KRAS mutations in sporadic colorectal cancer. Cancer Biol. Ther. 2014, 15, 768–776. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Zhang, Q.; Li, K.; Yin, H.; Zheng, J.N. Composite peptide-based vaccines for cancer immunotherapy (Review). Int. J. Mol. Med. 2015, 35, 17–23. [Google Scholar] [CrossRef] [Green Version]
- Katsuda, M.; Yamaue, H. Cancer vaccine therapy based on peptides. Trends Immunother. 2017, 1, 10–18. [Google Scholar] [CrossRef] [Green Version]
- Wada, S.; Yada, E.; Ohtake, J.; Sasada, T. Personalized peptide vaccines for cancer therapy: Current progress and state of the art. Expert Rev. Precis. Med. Drug Dev. 2017, 2, 371–381. [Google Scholar] [CrossRef]
- Santos, P.M.; Butterfield, L.H. Dendritic cell-based cancer vaccines. J. Immunol. 2018, 200, 443–449. [Google Scholar] [CrossRef]
- Zhang, H.; Chen, J. Current status and future directions of cancer immunotherapy. J. Cancer 2018, 9, 1773–1781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knittelfelder, R.; Riemer, A.B.; Jensen-Jarolim, E. Mimotope vaccination—From allergy to cancer. Expert Opin. Biol. Ther. 2009, 9, 493–506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buhrman, J.D.; Slansky, J.E. Mimotope vaccine efficacy gets a “boost” from native tumor antigens. Oncoimmunology 2013, 2, e23492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riemer, A.B.; Jensen-Jarolim, E. Mimotope vaccines: Epitope mimics induce anti-cancer antibodies. Immunol. Lett. 2007, 113, 1–5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, L.; Liu, Z.; Fan, D. Overview of mimotopes and related strategies in tumor vaccine development. Expert Rev. Vaccines 2008, 7, 1547–1555. [Google Scholar] [CrossRef] [PubMed]
- Buhrman, J.D.; Slansky, J.E. Improving T cell responses to modified peptides in tumor vaccines. Immunol. Res. 2013, 55, 34–47. [Google Scholar] [CrossRef] [Green Version]
- Parkhurst, M.R.; Salgaller, M.L.; Southwood, S.; Robbins, P.F.; Sette, A.; Rosenberg, S.A.; Kawakami, Y. Improved induction of melanoma-reactive CTL with peptides from the melanoma antigen gp100 modified at HLA-A*0201-binding residues. J. Immunol. 1996, 157, 2539–2548. [Google Scholar]
- Yu, Z.; Theoret, M.R.; Touloukian, C.E.; Surman, D.R.; Garman, S.C.; Feigenbaum, L.; Baxter, T.K.; Baker, B.M.; Restifo, N.P. Poor immunogenicity of a self/tumor antigen derives from peptide-MHC-I instability and is independent of tolerance. J. Clin. Investig. 2004, 114, 551–559. [Google Scholar] [CrossRef] [Green Version]
- Ng, A.W.; Tan, P.J.; Hoo, W.P.; Liew, D.S.; Teo, M.Y.; Siak, P.Y.; Ng, S.M.; Tan, E.W.; Rahim, R.A.; Lim, R.L.; et al. In silico-guided sequence modifications of K-ras epitopes improve immunological outcome against G12V and G13D mutant KRAS antigens. PeerJ 2018, 6, e5056. [Google Scholar] [CrossRef] [Green Version]
- Diethelm-Okita, B.M.; Okita, D.K.; Banaszak, L.; Conti-Fine, B.M. Universal epitopes for human CD4+ cells on tetanus and diphtheria toxins. J. Infect. Dis. 2000, 181, 1001–1009. [Google Scholar] [CrossRef] [Green Version]
- Yano, A.; Ito, K.; Miwa, Y.; Kanazawa, Y.; Chiba, A.; Iigo, Y.; Kashimoto, Y.; Kanda, A.; Murata, S.; Makino, M. The Peptide Vaccine Combined with Prior Immunization of a Conventional Diphtheria-Tetanus Toxoid Vaccine Induced Amyloid β Binding Antibodies on Cynomolgus Monkeys and Guinea Pigs. J. Immunol. Res. 2015, 2015, 786501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Xing, Y.; Guo, L.; Lv, X.; Song, H.; Xi, T. Oral immunization with recombinant Lactococcus lactis delivering a multi-epitope antigen CTB-UE attenuates Helicobacter pylori infection in mice. Pathog. Dis. 2014, 72, 78–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morello, E.; Bermudez-Humaran, L.G.; Llull, D.; Solé, V.; Miraglio, N.; Langella, P.; Poquet, I. Lactococcus lactis, an Efficient Cell Factory for Recombinant Protein Production and Secretion. J. Mol. Microbiol. Biotechnol. 2008, 14, 48–58. [Google Scholar] [CrossRef] [PubMed]
- Pouwels, P.H.; Leer, R.J.; Shaw, M.; den Bak-Glashouwer, M.J.; Tielen, F.D.; Smit, E.; Martinez, B.; Jore, J.; Conway, P.L. Lactic acid bacteria as antigen delivery vehicles for oral immunization purposes. Int. J. Food Microbiol. 1998, 41, 155–167. [Google Scholar] [CrossRef]
- Seegers, J.F. Lactobacilli as live vaccine delivery vectors: Progress and prospects. Trends Biotechnol. 2002, 20, 508–515. [Google Scholar] [CrossRef]
- Rafter, J.J. The role of lactic acid bacteria in colon cancer prevention. Scand. J. Gastroenterol. 1995, 30, 497–502. [Google Scholar] [CrossRef]
- Hirayama, K.; Rafter, J. The role of lactic acid bacteria in colon cancer prevention: Mechanistic considerations. Antonie Van Leeuwenhoek 1999, 76, 391–394. [Google Scholar]
- Bermúdez-Humarán, L.G.; Cortes-Perez, N.G.; Lefèvre, F.; Guimarães, V.; Rabot, S.; Alcocer-Gonzalez, J.M.; Gratadoux, J.J.; Rodriguez-Padilla, C.; Tamez-Guerra, R.S.; Corthier, G.; et al. A novel mucosal vaccine based on live lactococci expressing E7 antigen and IL-12 induces systemic and mucosal immune responses and protects mice against human papillomavirus type 16-induced tumors. J. Immunol. 2005, 175, 7297–7302. [Google Scholar] [CrossRef]
- Ng, D.T.W.; Sarkar, C.A. Engineering signal peptides for enhanced protein secretion from Lactococcus lactis. Appl. Environ. Microbiol. 2013, 79, 347–356. [Google Scholar] [CrossRef] [Green Version]
- Miyoshi, A.; Jamet, E.; Commissaire, J.; Renault, P.; Langella, P.; Azevedo, V. A xylose-inducible expression system for Lactococcus lactis. FEMS Microbiol. Lett. 2004, 239, 205–212. [Google Scholar] [CrossRef] [Green Version]
- Hoo, W.P.Y.; Siak, P.Y.; Alias, N.A.R.; Wong, J.J.; In, L.L.A. K-ras peptide mimotope induces a humoral immune response against G12V K-ras antigen in BALB/c mice. Asia Pac. J. Mol. Biol. Biotechnol. 2020, 22–35. [Google Scholar] [CrossRef]
- Holo, H.; Nes, I.F. High-frequency transformation, by electroporation, of Lactococcus lactis subsp cremoris grown with glycine in osmotically-stabilized media. Appl. Environ. Microbiol. 1989, 55, 3119–3123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moodie, Z.; Price, L.; Gouttefangeas, C.; Mander, A.; Janetzki, S.; Löwer, M.; Welters, M.J.; Ottensmeier, C.; Van der Burg, S.H.; Britten, C.M. Response definition criteria for ELISPOT assays revisited. Cancer Immunol. Immunother. 2010, 59, 1489–1501. [Google Scholar] [CrossRef] [Green Version]
- Schägger, H. Tricine-SDS-PAGE. Nat. Protoc. 2006, 1, 16–22. [Google Scholar] [CrossRef] [PubMed]
- Haider, S.R.; Reid, H.J.; Sharp, B.L. Tricine-SDS-PAGE. In Protein Electrophoresis; Methods in Molecular Biology (Methods and, Protocols); Kurien, B., Scofield, R., Eds.; Humana Press: Totowa, NJ, USA, 2012; Volume 869. [Google Scholar]
- Sun, J.C.; Beilke, J.N.; Lanier, L.L. Adaptive immune features of natural killer cells. Nature 2009, 457, 557–561. [Google Scholar] [CrossRef]
- Donnet-Hughes, A.; Rochat, F.; Serrant, P.; Aeschlimann, J.M.; Schiffrin, E.J. Modulation of nonspecific mechanisms of defense by lactic acid bacteria: Effective dose. J. Dairy Sci. 1999, 82, 863–869. [Google Scholar] [CrossRef]
- Seaman, W.E. Natural killer cells and natural killer T cells. Arthritis Rheum. 2000, 43, 1204–1217. [Google Scholar] [CrossRef]
- Cooper, M.A.; Fehniger, T.A.; Caligiuri, M.A. The biology of human natural killer-cell subsets. Trends Immunol. 2001, 22, 633–640. [Google Scholar] [CrossRef]
- Parra, M.D.; Martinez de Morentin, B.E.; Cobo, J.M.; Mateos, A.; Martinez, J.A. Daily ingestion of fermented milk containing Lactobacillus casei DN114001 improves innate-defense capacity in healthy middle-aged people. J. Physiol. Biochem. 2004, 60, 85–91. [Google Scholar] [CrossRef]
- Seifert, S.; Bub, A.; Franz, C.; Watzl, B. Probiotic Lactobacillus casei Shirota supplementation does not modulate immunity in healthy men with reduced natural killer cell activity. J. Nutr. 2011, 141, 978–984. [Google Scholar] [CrossRef]
- Xin, K.Q.; Hoshino, Y.; Toda, Y.; Igimi, S.; Kojima, Y.; Jounai, N.; Ohba, K.; Kushiro, A.; Kiwaki, M.; Hamajima, K.; et al. Immunogenicity and protective efficacy of orally administered recombinant Lactococcus lactis expressing surface-bound HIV Env. Blood 2003, 102, 223–228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asensi, G.F.; de Sales, N.F.; Dutra, F.F.; Feijó, D.F.; Bozza, M.T.; Ulrich, R.G.; Miyoshi, A.; de Morais, K.; de Carvalho Azevedo, V.A.; Silva, J.T.; et al. Oral immunization with Lactococcus lactis secreting attenuated recombinant staphylococcal enterotoxin B induces a protective immune response in a murine model. Microb. Cell Fact. 2013, 12, 32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogra, P.L.; Mestecky, J.; Lamm, M.E.; Strober, W.; McGhee, J.R.; Bienenstock, J. Mucosal Immunology, 4th ed.; Academic Press: Cambridge, MA, USA, 2015. [Google Scholar]
- Shakya, A.K.; Chowdhury, M.Y.; Tao, W.; Gill, H.S. Mucosal vaccine delivery: Current state and a pediatric perspective. J. Control Release 2016, 240, 394–413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bermúdez-Humarán, L.G. Lactococcus lactis as a live vector for mucosal delivery of therapeutic proteins. Hum. Vaccin. 2009, 5, 264–267. [Google Scholar] [CrossRef] [Green Version]
- Adachi, K.; Kawana, K.; Yokoyama, T.; Fujii, T.; Tomio, A.; Miura, S.; Tomio, K.; Kojima, S.; Oda, K.; Sewaki, T.; et al. Oral immunization with a Lactobacillus casei vaccine expressing human papillomavirus (HPV) type 16 E7 is an effective strategy to induce mucosal cytotoxic lymphocytes against HPV16 E7. Vaccine 2010, 28, 2810–2817. [Google Scholar] [CrossRef]
- Lebeer, S.; Vanderleyden, J.; Keersmaecker, S.C. Host interactions of probiotic bacterial surface molecules: Comparison with commensals and pathogens. Nat. Rev. Microbiol. 2010, 8, 171–184. [Google Scholar] [CrossRef]
- Azizpour, M.; Hosseini, S.D.; Jafari, P.; Akbary, N. Lactococcus lactis: A new strategy for vaccination. Avicenna J. Med. Biotechnol. 2017, 9, 163–168. [Google Scholar]
- Hahn, H.P.; von Specht, B.U. Secretory delivery of recombinant proteins in attenuated Salmonella strains: Potential and limitations of Type I protein transporters. FEMS Immunol. Med. Microbiol. 2003, 37, 87–98. [Google Scholar] [CrossRef]
- Van Wely, K.H.; Swaving, J.; Freudl, R.; Driessen, A.J.M. Translocation of proteins across the cell envelope of Gram-positive bacteria. FEMS Microbiol. Rev. 2001, 25, 437–454. [Google Scholar] [CrossRef]
- Mathiesen, G.; Sveen, A.; Piard, J.C.; Axelsson, L.; Eijsink, V.G. Heterologous protein secretion by Lactobacillus plantarum using homologous Signal peptides. J. Appl. Microbiol. 2008, 105, 215–216. [Google Scholar] [CrossRef]
- Van Asseldonk, M.; de Vos, W.M.; Simons, G. Functional analysis of the Lactococcus lactis usp45 secretion signal in the secretion of a homologous proteinase and a heterologous α-amylase. Mol. Gen. Genet. 1993, 240, 428–434. [Google Scholar] [CrossRef] [PubMed]
- Dieye, Y.; Hoekman, A.J.; Clier, F.; Juillard, V.; Boot, H.J.; Piard, J.C. Ability of Lactococcus lactis to export viral capsid antigens: A crucial step for development of live vaccines. Appl. Environ. Microbiol. 2003, 69, 7281–7288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le Loir, Y.; Azevedo, V.; Oliveira, S.C.; Freitas, D.A.; Miyoshi, A.; Bermúdez-Humarán, L.G.; Nouaille, S.; Ribeiro, L.A.; Leclercq, S.; Gabriel, J.E.; et al. Protein secretion in Lactococcus lactis: An efficient way to increase the overall heterologous protein production. Microb. Cell Fact. 2005, 4, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hazebrouck, S.; Pothelune, L.; Azevedo, V.; Corthier, G.; Wal, J.M.; Langella, P. Efficient Production and Secretion of bovine β-lactoglobulin by Lactobacillus casei. Microb. Cell Fact. 2007, 6, 12. [Google Scholar] [CrossRef] [Green Version]
- Baradaran, A.; Sieo, C.C.; Foo, H.L.; Illias, R.M.; Yusoff, K.; Rahim, R.A. Cloning and in silico characterization of two Signal peptides from Pediococcus pentosaceus and their function for the secretion of heterologous protein in Lactococcus lactis. Biotechnol. Lett. 2013, 35, 235–238. [Google Scholar] [CrossRef]
- Joan, S.S.; Pui-Fong, J.; Song, A.A.; Chang, L.Y.; Yusoff, K.; AbuBakar, S.; Rahim, R.A. Oral vaccine of Lactococcus lactis harboring pandemic H1N1 2009 haemagglutinin1 and nisP anchor fusion protein elevates anti-HA1 sIgA levels in mice. Biotechnol. Lett. 2016, 38, 793–799. [Google Scholar] [CrossRef]
- Tayeb, I.; Jamel, B.; Essaid, L.; Nour-Eddine, K. Survival of Lactobacillus plantarum bj0021 and Pediococcus acidilactici in the digestive tract of rabbit. Int. J. Probiotics Prebiotics 2007, 2, 49–54. [Google Scholar]
- Klijn, N.; Weerkamp, A.H.; de Vos, W.M. Genetic marking of Lactococcus lactis shows its survival in the human gastrointestinal tract. Appl. Environ. Microbiol. 1995, 61, 2771–2774. [Google Scholar] [CrossRef] [Green Version]
- Kim, W.S.; Ren, J.; Dunn, N.W. Differentiation of Lactococcus lactis subspecies lactis and subspecies cremoris strains by their adaptive response to stresses. FEMS Microbiol. Lett. 1999, 171, 57–65. [Google Scholar] [CrossRef] [Green Version]
- Tan, E.W.; Tan, K.Y.; Phang, L.V.; Kumar, P.V.; In, L.L.A. Enhanced gastrointestinal survivability of recombinant Lactococcus lactis using a double coated mucoadhesive film approach. PLoS ONE 2019, 14, e0219912. [Google Scholar] [CrossRef]
- Speck, M.L. Acidophilus food products. Dev. Ind. Microbiol. 1978, 19, 95–101. [Google Scholar]
- Kim, H.S. Characterization of lactobacilli and bifidobacteria as applied to dietary adjuncts. Cult. Dairy Prod. J. 1988, 23, 6–9. [Google Scholar]
- Kang, S.H.; Hong, S.J.; Lee, Y.K.; Cho, S. Oral Vaccine Delivery for Intestinal Immunity-Biological Basis, Barriers, Delivery System, and M Cell Targeting. Polymers 2018, 10, 948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reilly, R.T.; Machiels, J.P.; Emens, L.A.; Ercolini, A.M.; Okoye, F.I.; Lei, R.Y.; Weintraub, D.; Jaffee, E.M. The collaboration of both humoral and cellular HER-2/neu-targeted immune responses is required for the complete eradication of HER-2/neu-expressing tumors. Cancer Res. 2001, 61, 880–883. [Google Scholar] [PubMed]
- Reilly, R.T.; Emens, L.A.; Jaffee, E.M. Humoral and cellular immune responses: Independent forces or collaborators in the fight against cancer? Curr. Opin. Investig. Drugs 2001, 2, 133–135. [Google Scholar]
- Emens, L.A. Cancer vaccines: On the threshold of success. Expert Opin. Emerg. Drugs 2008, 13, 295–308. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Siak, P.Y.; Wong, K.Y.; Song, A.A.-L.; Rahim, R.A.; In, L.L.A. K-Ras Peptide Mimotope Induces Antigen Specific Th1 and B-Cell Immune Responses against G12A-Mutated K-Ras Antigen in Balb/c Mice. Vaccines 2021, 9, 195. https://doi.org/10.3390/vaccines9030195
Siak PY, Wong KY, Song AA-L, Rahim RA, In LLA. K-Ras Peptide Mimotope Induces Antigen Specific Th1 and B-Cell Immune Responses against G12A-Mutated K-Ras Antigen in Balb/c Mice. Vaccines. 2021; 9(3):195. https://doi.org/10.3390/vaccines9030195
Chicago/Turabian StyleSiak, Pui Yan, Kuan Yee Wong, Adelene Ai-Lian Song, Raha Abdul Rahim, and Lionel Lian Aun In. 2021. "K-Ras Peptide Mimotope Induces Antigen Specific Th1 and B-Cell Immune Responses against G12A-Mutated K-Ras Antigen in Balb/c Mice" Vaccines 9, no. 3: 195. https://doi.org/10.3390/vaccines9030195