
Heterologous Booster Immunization Based on Inactivated SARS-CoV-2 Vaccine Enhances Humoral Immunity and Promotes BCR Repertoire Development
 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mouse Immunization and Ethics
2.2. ELISA
2.3. ELISpot Assay
2.4. Antibody-Secreting Cell ELISpot
2.5. Pseudovirus Neutralization Assay
2.6. Sorting of Antigen-Specific B-Cells and BCR Gene Amplification
2.7. Flow Cytometry
2.8. Serum Multiplex Cytokine Analysis
2.9. Statistical Analysis
3. Results
3.1. Booster Immunization Significantly Increases Antigen-Specific Antibodies in Mice
3.2. Heterologous Booster Immunization Induces Stronger Antibody-Secreting Cells and Induces IgG Antibody Class Switching
3.3. A Heterologous Boost Elicits Robust Cellular Immune Responses
3.4. Booster Immunization Affects the Levels of Cytokines in Serum
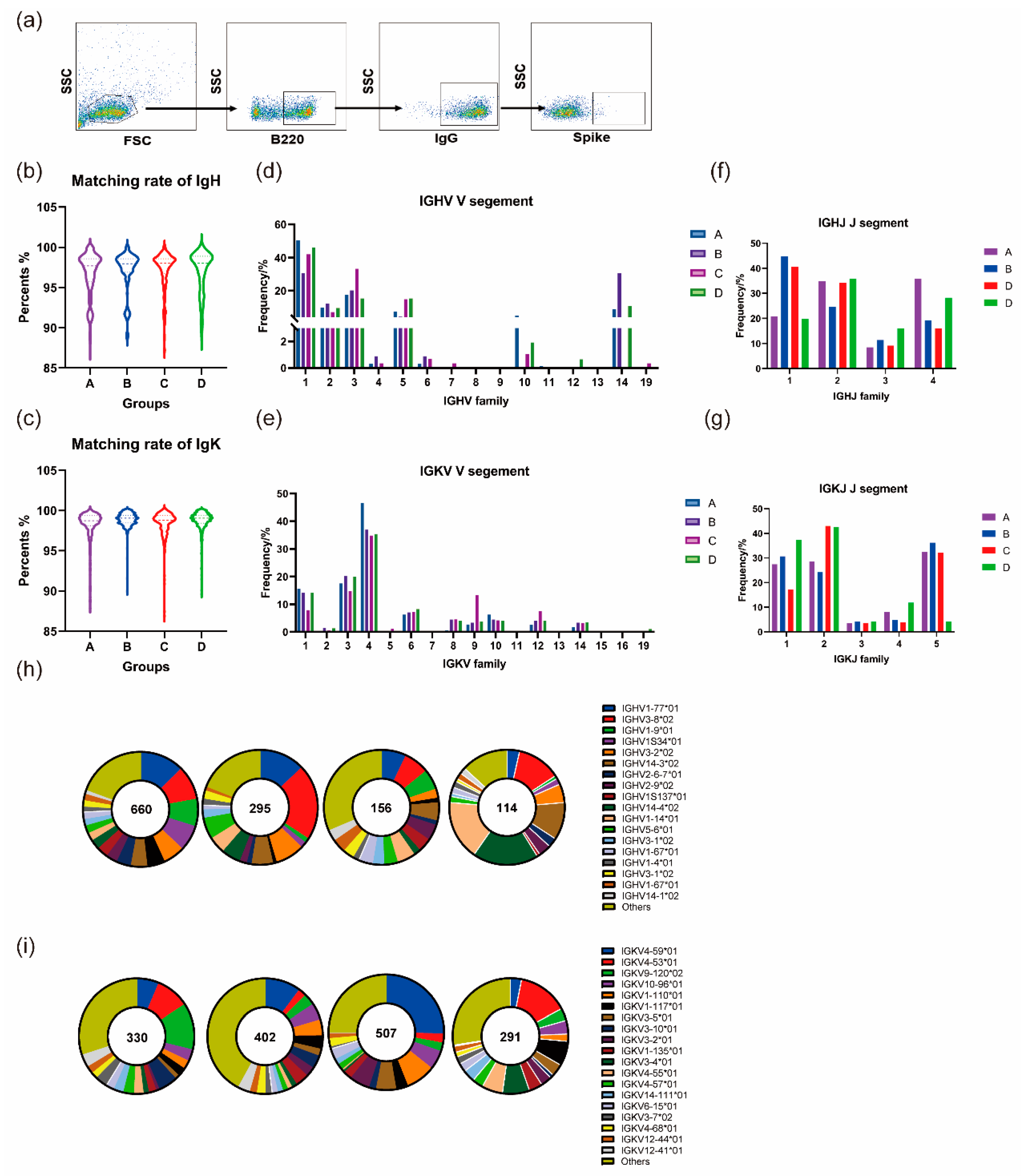
3.5. Homologous and Heterologous Boosters Promotes Continued Development of the BCR Lineage
3.5.1. Homologous and Heterologous Boosters Both Contribute to the Continuous Maturation of Germline Genes after the Prime Immunization
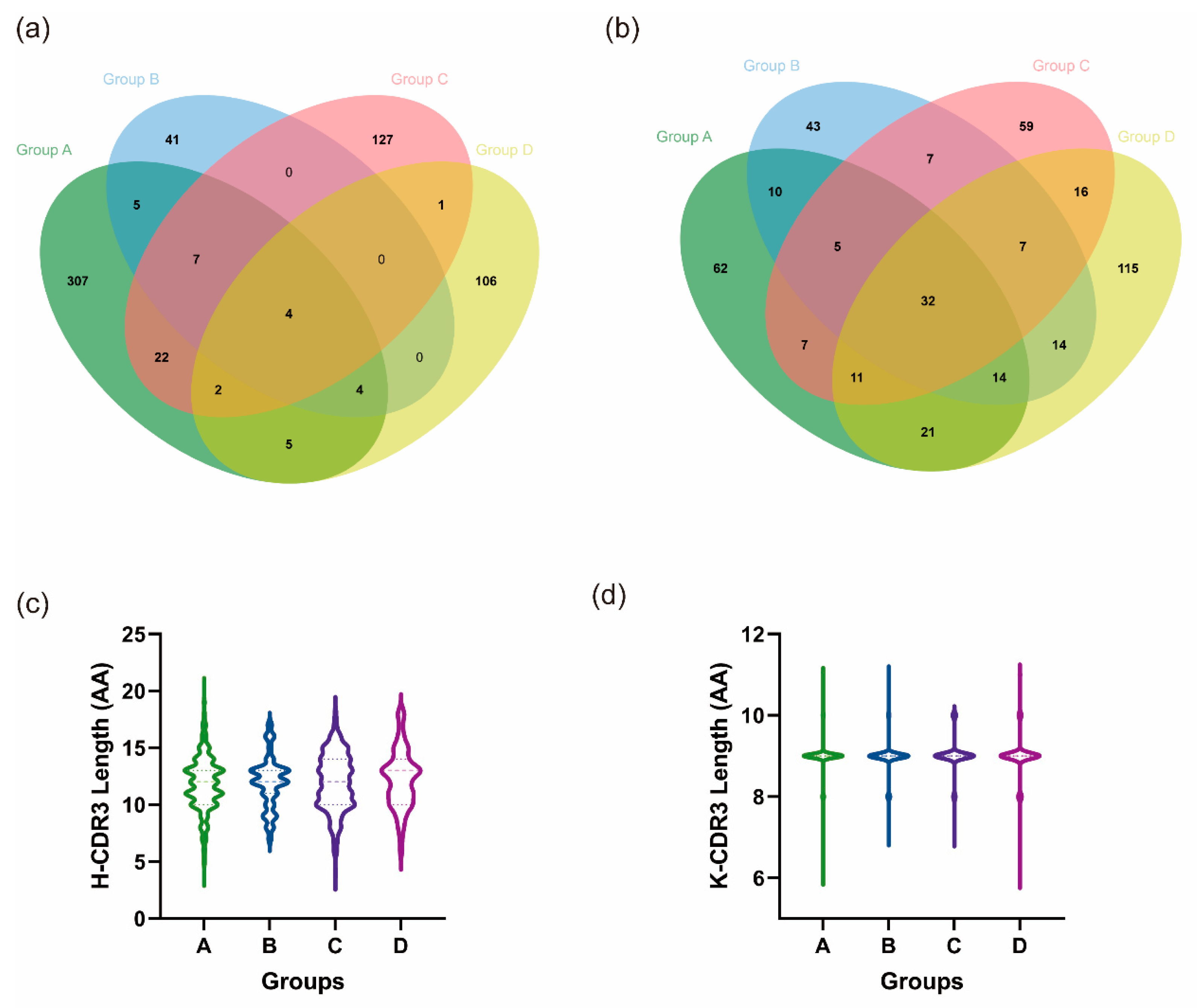
3.5.2. Heterologous Boosting Increases the Diversity of CDR3
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WHO Coronavirus (COVID-19) Dashboard. Available online: https://covid19.who.int (accessed on 31 October 2023).
- Jackson, C.B.; Farzan, M.; Chen, B.; Choe, H. Mechanisms of SARS-CoV-2 Entry into Cells. Nat. Rev. Mol. Cell Biol. 2022, 23, 3–20. [Google Scholar] [CrossRef] [PubMed]
- Buckner, C.M.; Kardava, L.; El Merhebi, O.; Narpala, S.R.; Serebryannyy, L.; Lin, B.C.; Wang, W.; Zhang, X.; de Assis, F.L.; Kelly, S.E.; et al. Interval between Prior SARS-CoV-2 Infection and Booster Vaccination Impacts Magnitude and Quality of Antibody and B Cell Responses. Cell 2022, 185, 4333–4346. [Google Scholar] [CrossRef] [PubMed]
- Röltgen, K.; Boyd, S.D. Antibody and B Cell Responses to SARS-CoV-2 Infection and Vaccination. Cell Host Microbe 2021, 29, 1063–1075. [Google Scholar] [CrossRef] [PubMed]
- Laidlaw, B.J.; Ellebedy, A.H. The Germinal Centre B Cell Response to SARS-CoV-2. Nat. Rev. Immunol. 2022, 22, 7–18. [Google Scholar] [CrossRef] [PubMed]
- Duan, K.; Liu, B.; Li, C.; Zhang, H.; Yu, T.; Qu, J.; Zhou, M.; Chen, L.; Meng, S.; Hu, Y.; et al. Effectiveness of Convalescent Plasma Therapy in Severe COVID-19 Patients. Proc. Natl. Acad. Sci. USA 2020, 117, 9490–9496. [Google Scholar] [CrossRef] [PubMed]
- Sharma, K.; Koirala, A.; Nicolopoulos, K.; Chiu, C.; Wood, N.; Britton, P.N. Vaccines for COVID-19: Where Do We Stand in 2021? Paediatr. Respir. Rev. 2021, 39, 22–31. [Google Scholar] [CrossRef] [PubMed]
- Muecksch, F.; Wang, Z.; Cho, A.; Gaebler, C.; Ben Tanfous, T.; DaSilva, J.; Bednarski, E.; Ramos, V.; Zong, S.; Johnson, B.; et al. Increased Memory B Cell Potency and Breadth after a SARS-CoV-2 mRNA Boost. Nature 2022, 607, 128–134. [Google Scholar] [CrossRef]
- Patone, M.; Mei, X.W.; Handunnetthi, L.; Dixon, S.; Zaccardi, F.; Shankar-Hari, M.; Watkinson, P.; Khunti, K.; Harnden, A.; Coupland, C.A.C.; et al. Risks of Myocarditis, Pericarditis, and Cardiac Arrhythmias Associated with COVID-19 Vaccination or SARS-CoV-2 Infection. Nat. Med. 2022, 28, 410–422. [Google Scholar] [CrossRef]
- Bernal, J.L.; Andrews, N.; Gower, C.; Robertson, C.; Stowe, J.; Tessier, E.; Simmons, R.; Cottrell, S.; Roberts, R.; O’doherty, M.; et al. Effectiveness of the Pfizer-BioNTech and Oxford-AstraZeneca Vaccines on COVID-19 Related Symptoms, Hospital Admissions, and Mortality in Older Adults in England: Test Negative Case-Control Study. BMJ 2021, 373, n1088. [Google Scholar] [CrossRef]
- Vanshylla, K.; Fan, C.; Wunsch, M.; Poopalasingam, N.; Meijers, M.; Kreer, C.; Kleipass, F.; Ruchnewitz, D.; Ercanoglu, M.S.; Gruell, H.; et al. Discovery of Ultrapotent Broadly Neutralizing Antibodies from SARS-CoV-2 Elite Neutralizers. Cell Host Microbe 2022, 30, 69–82.e10. [Google Scholar] [CrossRef]
- Yang, M.; Li, J.; Huang, Z.; Li, H.; Wang, Y.; Wang, X.; Kang, S.; Huang, X.; Wu, C.; Liu, T.; et al. Structural Basis of a Human Neutralizing Antibody Specific to the SARS-CoV-2 Spike Protein Receptor-Binding Domain. Microbiol. Spectr. 2021, 9, e0135221. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Patel, A.; Lai, L.; Chakravarthy, C.; Valanparambil, R.; Reddy, E.S.; Gottimukkala, K.; Davis-Gardner, M.E.; Edara, V.V.; Linderman, S.; et al. Structural Insights for Neutralization of Omicron Variants BA.1, BA.2, BA.4, and BA.5 by a Broadly Neutralizing SARS-CoV-2 Antibody. Sci. Adv. 2022, 8, eadd2032. [Google Scholar] [CrossRef] [PubMed]
- Huang, Q.; Han, X.; Yan, J. Structure-Based Neutralizing Mechanisms for SARS-CoV-2 Antibodies. Emerg. Microbes Infect. 2022, 11, 2412–2422. [Google Scholar] [CrossRef] [PubMed]
- Burki, T.K. Fourth Dose of COVID-19 Vaccines in Israel. Lancet Respir. Med. 2022, 10, e19. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Yang, Y.; Qiao, N.; Wang, X.; Ding, L.; Zhu, X.; Liang, Y.; Han, Z.; Liu, F.; Zhang, X.; et al. Early Assessment of the Safety and Immunogenicity of a Third Dose (Booster) of COVID-19 Immunization in Chinese adults. Front. Med. 2022, 16, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Nemet, I.; Kliker, L.; Lustig, Y.; Zuckerman, N.; Erster, O.; Cohen, C.; Kreiss, Y.; Alroy-Preis, S.; Regev-Yochay, G.; Mendelson, E.; et al. Third BNT162b2 Vaccination Neutralization of SARS-CoV-2 Omicron Infection. N. Engl. J. Med. 2021, 386, 492–494. [Google Scholar] [CrossRef] [PubMed]
- Barda, N.; Dagan, N.; Cohen, C.; A Hernán, M.; Lipsitch, M.; Kohane, I.S.; Reis, B.Y.; Balicer, R.D. Effectiveness of a Third Dose of the BNT162b2 mRNA COVID-19 Vaccine for Preventing Severe Outcomes in Israel: An observational study. Lancet 2021, 398, 2093–2100. [Google Scholar] [CrossRef] [PubMed]
- Abbasi, J. Fourth COVID-19 Vaccine Dose Increases Low Antibodies. JAMA 2022, 327, 517. [Google Scholar] [CrossRef]
- Duarte-Salles, T.; Prieto-Alhambra, D. Heterologous Vaccine Regimens against COVID-19. Lancet 2021, 398, 94–95. [Google Scholar] [CrossRef]
- Sapkota, B.; Saud, B.; Shrestha, R.; Al-Fahad, D.; Sah, R.; Shrestha, S.; Rodriguez-Morales, A.J. Heterologous Prime-Boost Strategies for COVID-19 Vaccines. J. Travel Med. 2022, 29, taab191. [Google Scholar] [CrossRef]
- He, Q.; Mao, Q.; An, C.; Zhang, J.; Gao, F.; Bian, L.; Li, C.; Liang, Z.; Xu, M.; Wang, J. Heterologous Prime-Boost: Breaking the Protective Immune Response Bottleneck of COVID-19 Vaccine Candidates. Emerg. Microbes Infect. 2021, 10, 629–637. [Google Scholar] [CrossRef] [PubMed]
- Limami, Y.; Khalki, L.; Zaid, N.; Khyatti, M.; El Turk, J.; Ammara, M.; Mtairag, E.M.; Oudghiri, M.; Naya, A.; Taberkant, M.; et al. Oxford-AstraZeneca ChAdOx1 COVID-19 Vaccine Does Not Alter Platelet Aggregation. Semin. Thromb. Hemost. 2021, 48, 109–111. [Google Scholar] [CrossRef]
- Scully, M.; Singh, D.; Lown, R.; Poles, A.; Solomon, T.; Levi, M.; Goldblatt, D.; Kotoucek, P.; Thomas, W.; Lester, W. Pathologic Antibodies to Platelet Factor 4 after ChAdOx1 nCoV-19 Vaccination. N. Engl. J. Med. 2021, 384, 2202–2211. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Shaw, R.H.; Stuart, A.S.V.; Greenland, M.; Aley, P.K.; Andrews, N.J.; Cameron, J.C.; Charlton, S.; A Clutterbuck, E.; Collins, A.M.; et al. Safety and Immunogenicity of Heterologous versus Homologous Prime-Boost Schedules with an Adenoviral Vectored and mRNA COVID-19 Vaccine (Com-COV): A Single-Blind, Randomised, Non-Inferiority Trial. Lancet 2021, 398, 856–869. [Google Scholar] [CrossRef] [PubMed]
- Groß, R.; Zanoni, M.; Seidel, A.; Conzelmann, C.; Gilg, A.; Krnavek, D.; Erdemci-Evin, S.; Mayer, B.; Hoffmann, M.; Pöhlmann, S.; et al. Heterologous ChAdOx1 nCoV-19 and BNT162b2 Prime-Boost Vaccination Elicits Potent Neutralizing Antibody Responses and T Cell Reactivity against Prevalent SARS-CoV-2 Variants. EBioMedicine 2022, 75, 103761. [Google Scholar] [CrossRef] [PubMed]
- Barros-Martins, J.; Hammerschmidt, S.I.; Cossmann, A.; Odak, I.; Stankov, M.V.; Ramos, G.M.; Dopfer-Jablonka, A.; Heidemann, A.; Ritter, C.; Friedrichsen, M.; et al. Immune Responses against SARS-CoV-2 Variants after Heterologous and Homologous ChAdOx1 nCoV-19/BNT162b2 Vaccination. Nat. Med. 2021, 27, 1525–1529. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Zhao, H.; Huang, B.; Zhao, T.; Wang, L.; Zhang, J.; Yang, Y.; Tang, X.; Tan, W.; Lin, A. Unravelling the Enhanced Vaccine Immunity by Heterologous KCONVAC/Ad5-nCoV COVID-19 Vaccination. Signal Transduct. Target. Ther. 2022, 7, 210. [Google Scholar] [CrossRef]
- Nie, J.; Li, Q.; Wu, J.; Zhao, C.; Hao, H.; Liu, H.; Zhang, L.; Nie, L.; Qin, H.; Wang, M.; et al. Establishment and Validation of a Pseudovirus Neutralization Assay for SARS-CoV-2. Emerg. Microbes Infect. 2020, 9, 680–686. [Google Scholar] [CrossRef]
- Tiller, T.; Busse, C.E.; Wardemann, H. Cloning and Expression of Murine Ig Genes from Single B Cells. J. Immunol. Methods 2009, 350, 183–193. [Google Scholar] [CrossRef]
- von Boehmer, L.; Liu, C.; Ackerman, S.; Gitlin, A.D.; Wang, Q.; Gazumyan, A.; Nussenzweig, M.C. Sequencing and Cloning of Antigen-Specific Antibodies from Mouse Memory B Cells. Nat. Protoc. 2016, 11, 1908–1923. [Google Scholar] [CrossRef]
- Vaccine Schedule for Children 6 Years or Younger|CDC. Available online: https://www.cdc.gov/vaccines/schedules/easy-to-read/child-easyread.html (accessed on 1 November 2023).
- Lu, S. Heterologous Prime–Boost Vaccination. Curr. Opin. Immunol. 2009, 21, 346–351. [Google Scholar] [CrossRef] [PubMed]
- Kardani, K.; Bolhassani, A.; Shahbazi, S. Prime-Boost Vaccine Strategy against Viral Infections: Mechanisms and Benefits. Vaccine 2016, 34, 413–423. [Google Scholar] [CrossRef] [PubMed]
- Ho, T.-C.; Chen, Y.-M.A.; Chan, H.-P.; Chang, C.-C.; Chuang, K.-P.; Lee, C.-H.; Yuan, C.-H.; Tyan, Y.-C.; Yang, M.-H. The Effects of Heterologous Immunization with Prime-Boost COVID-19 Vaccination against SARS-CoV-2. Vaccines 2021, 9, 1163. [Google Scholar] [CrossRef] [PubMed]
- Ciapponi, A.; Bardach, A.; Ares, L.R.; Glujovsky, D.; Cafferata, M.L.; Cesaroni, S.; Bhatti, A. Sequential Inactivated (IPV) and Live Oral (OPV) Poliovirus Vaccines for Preventing Poliomyelitis. Cochrane Database Syst. Rev. 2019, 12, CD011260. [Google Scholar] [CrossRef] [PubMed]
- Chiu, N.-C.; Chi, H.; Tu, Y.-K.; Huang, Y.-N.; Tai, Y.-L.; Weng, S.-L.; Chang, L.; Huang, D.T.-N.; Huang, F.-Y.; Lin, C.-Y. To Mix or Not to Mix? A Rapid Systematic Review of Heterologous Prime–Boost COVID-19 Vaccination. Expert Rev. Vaccines 2021, 20, 1211–1220. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Hou, L.; Guo, X.; Jin, P.; Wu, S.; Zhu, J.; Pan, H.; Wang, X.; Song, Z.; Wan, J.; et al. Heterologous AD5-nCOV Plus CoronaVac versus Homologous CoronaVac Vaccination: A Randomized Phase 4 Trial. Nat. Med. 2022, 28, 401–409. [Google Scholar] [CrossRef]
- Zuo, F.; Abolhassani, H.; Du, L.; Piralla, A.; Bertoglio, F.; de Campos-Mata, L.; Wan, H.; Schubert, M.; Wang, Y.; Sun, R.; et al. Heterologous Immunization with Inactivated Vaccine Followed by mRNA Booster Elicits Strong Humoral and Cellular Immune Responses against the SARS-CoV-2 Omicron Variant. Nat. Commun. 2022, 13, 2670. [Google Scholar] [CrossRef]
- Yang, J.; Liu, M.-Q.; Liu, L.; Li, X.; Xu, M.; Lin, H.; Liu, S.; Hu, Y.; Li, B.; Liu, B.; et al. A Triple-RBD-Based Mucosal Vaccine Provides Broad Protection against SARS-CoV-2 Variants of Concern. Cell. Mol. Immunol. 2022, 19, 1279–1289. [Google Scholar] [CrossRef]
- Wang, C.Y.; Hwang, K.-P.; Kuo, H.-K.; Peng, W.-J.; Shen, Y.-H.; Kuo, B.-S.; Huang, J.-H.; Liu, H.; Ho, Y.-H.; Lin, F.; et al. A Multitope SARS-CoV-2 Vaccine Provides Long-Lasting B Cell and T Cell Immunity against Delta and Omicron Variants. J. Clin. Investig. 2022, 132, e157707. [Google Scholar] [CrossRef]
- Cao, H.; Yang, S.; Wang, Y.; Luan, N.; Yin, X.; Lin, K.; Liu, C. An Established Th2-Oriented Response to an Alum-Adjuvanted SARS-CoV-2 Subunit Vaccine Is Not Reversible by Sequential Immunization with Nucleic Acid-Adjuvanted Th1-Oriented Subunit Vaccines. Vaccines 2021, 9, 1261. [Google Scholar] [CrossRef]
- Victora, G.D.; Nussenzweig, M.C. Germinal Centers. Annu. Rev. Immunol. 2022, 40, 413–442. [Google Scholar] [CrossRef] [PubMed]
- Mesin, L.; Ersching, J.; Victora, G.D. Germinal Center B Cell Dynamics. Immunity 2016, 45, 471–482. [Google Scholar] [CrossRef] [PubMed]
- Turner, J.S.; O’halloran, J.A.; Kalaidina, E.; Kim, W.; Schmitz, A.J.; Zhou, J.Q.; Lei, T.; Thapa, M.; Chen, R.E.; Case, J.B.; et al. SARS-CoV-2 mRNA Vaccines Induce Persistent Human Germinal Centre Responses. Nature 2021, 596, 109–113. [Google Scholar] [CrossRef] [PubMed]
- Mintz, M.A.; Cyster, J.G. T Follicular Helper Cells in Germinal Center B Cell Selection and Lymphomagenesis. Immunol. Rev. 2020, 296, 48–61. [Google Scholar] [CrossRef]
- Kak, G.; Raza, M.; Tiwari, B.K. Interferon-gamma (IFN-γ): Exploring Its Implications in Infectious Diseases. Biomol. Concepts 2018, 9, 64–79. [Google Scholar] [CrossRef]
- Tanaka, S.; Baba, Y. B Cell Receptor Signaling. Adv. Exp. Med. Biol. 2020, 1254, 23–36. [Google Scholar] [CrossRef]
- Treanor, B. B-cell Receptor: From Resting State to Activate. Immunology 2012, 136, 21–27. [Google Scholar] [CrossRef]
- Gao, H.; Yu, L.; Yan, F.; Zheng, Y.; Huang, H.; Zhuang, X.; Zeng, Y. Landscape of B Cell Receptor Repertoires in COVID-19 Patients Revealed Through CDR3 Sequencing of Immunoglobulin Heavy and Light Chains. Immunol. Investig. 2022, 51, 1994–2008. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Sequence Number of IgH | Species of H-CDR3 | Ratio | Sequence Number of IgK | Species of K-CDR3 | Ratio |
|---|---|---|---|---|---|---|
| A | 660 | 355 | 0.54 | 507 | 162 | 0.32 |
| B | 114 | 61 | 0.54 | 291 | 132 | 0.45 |
| C | 295 | 163 | 0.55 | 328 | 144 | 0.44 |
| D | 156 | 123 | 0.79 | 402 | 230 | 0.57 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.; Zeng, F.; Yue, R.; Ma, D.; Meng, Z.; Li, Q.; Zhang, Z.; Zhang, H.; Liao, Y.; Liao, Y.; et al. Heterologous Booster Immunization Based on Inactivated SARS-CoV-2 Vaccine Enhances Humoral Immunity and Promotes BCR Repertoire Development. Vaccines 2024, 12, 120. https://doi.org/10.3390/vaccines12020120
Li X, Zeng F, Yue R, Ma D, Meng Z, Li Q, Zhang Z, Zhang H, Liao Y, Liao Y, et al. Heterologous Booster Immunization Based on Inactivated SARS-CoV-2 Vaccine Enhances Humoral Immunity and Promotes BCR Repertoire Development. Vaccines. 2024; 12(2):120. https://doi.org/10.3390/vaccines12020120
Chicago/Turabian StyleLi, Xinghang, Fengyuan Zeng, Rong Yue, Danjing Ma, Ziyan Meng, Qi Li, Zhenxiao Zhang, Haobo Zhang, Yuansheng Liao, Yun Liao, and et al. 2024. "Heterologous Booster Immunization Based on Inactivated SARS-CoV-2 Vaccine Enhances Humoral Immunity and Promotes BCR Repertoire Development" Vaccines 12, no. 2: 120. https://doi.org/10.3390/vaccines12020120