
Chronic Fatigue and Dysautonomia following COVID-19 Vaccination Is Distinguished from Normal Vaccination Response by Altered Blood Markers

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Participants
2.2. Controls
2.3. Validation of SARS-CoV-2 Vaccination and Infection
2.4. Ethics
2.5. Laboratory Measurements
2.6. Statistical Methods
3. Results
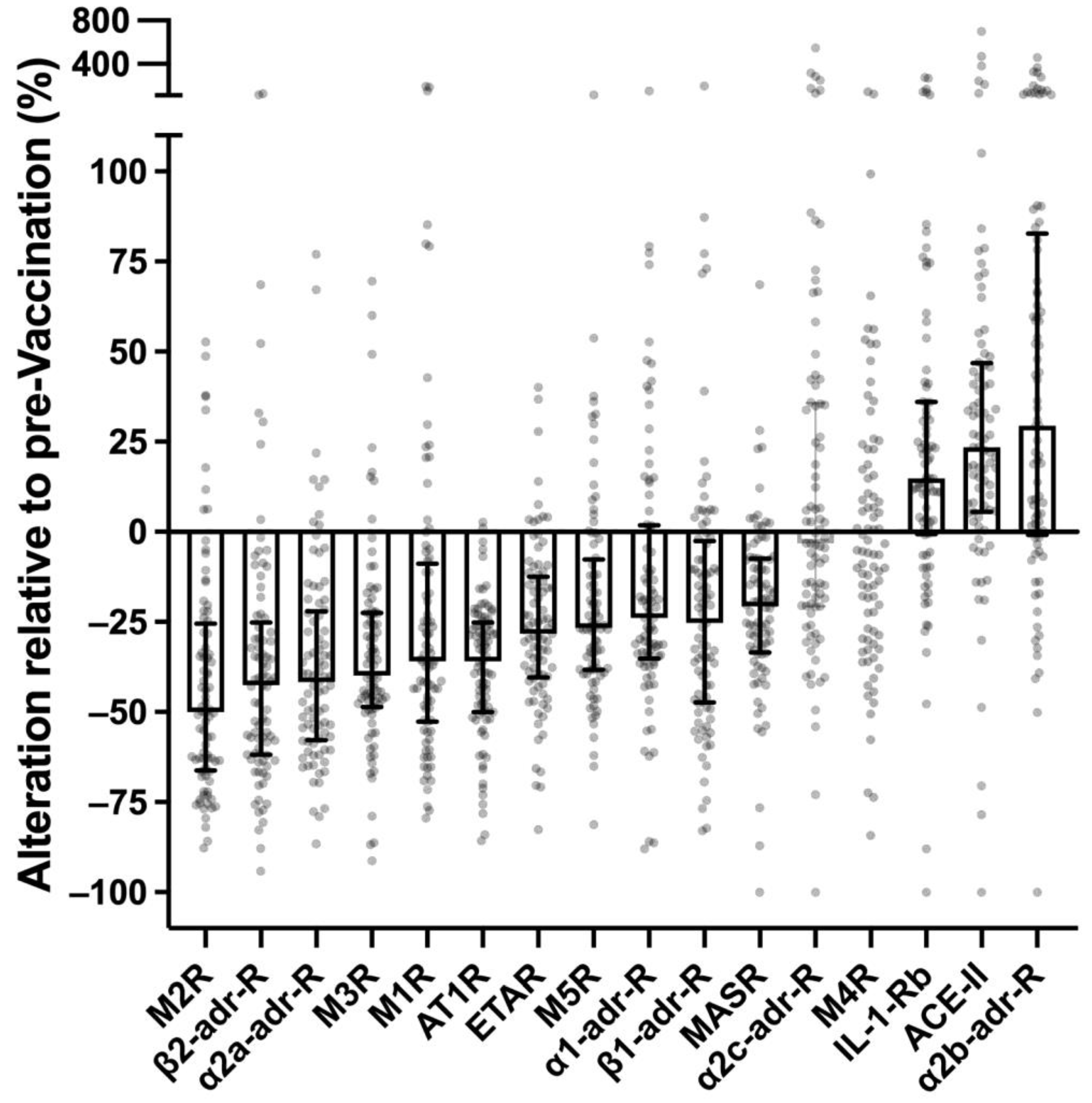
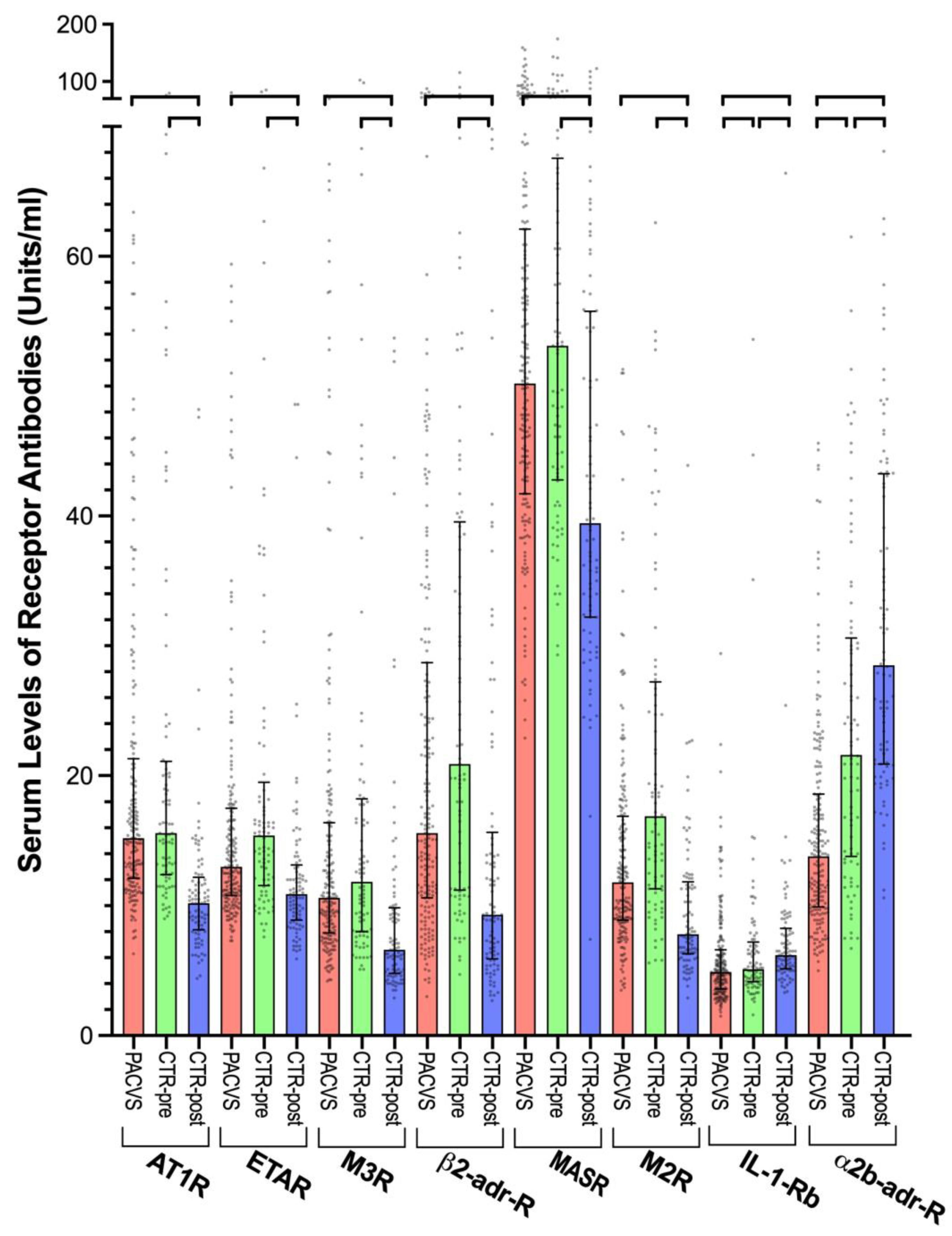
3.1. Impact of SARS-CoV-2 Vaccination on Receptor Antibodies in Healthy Controls
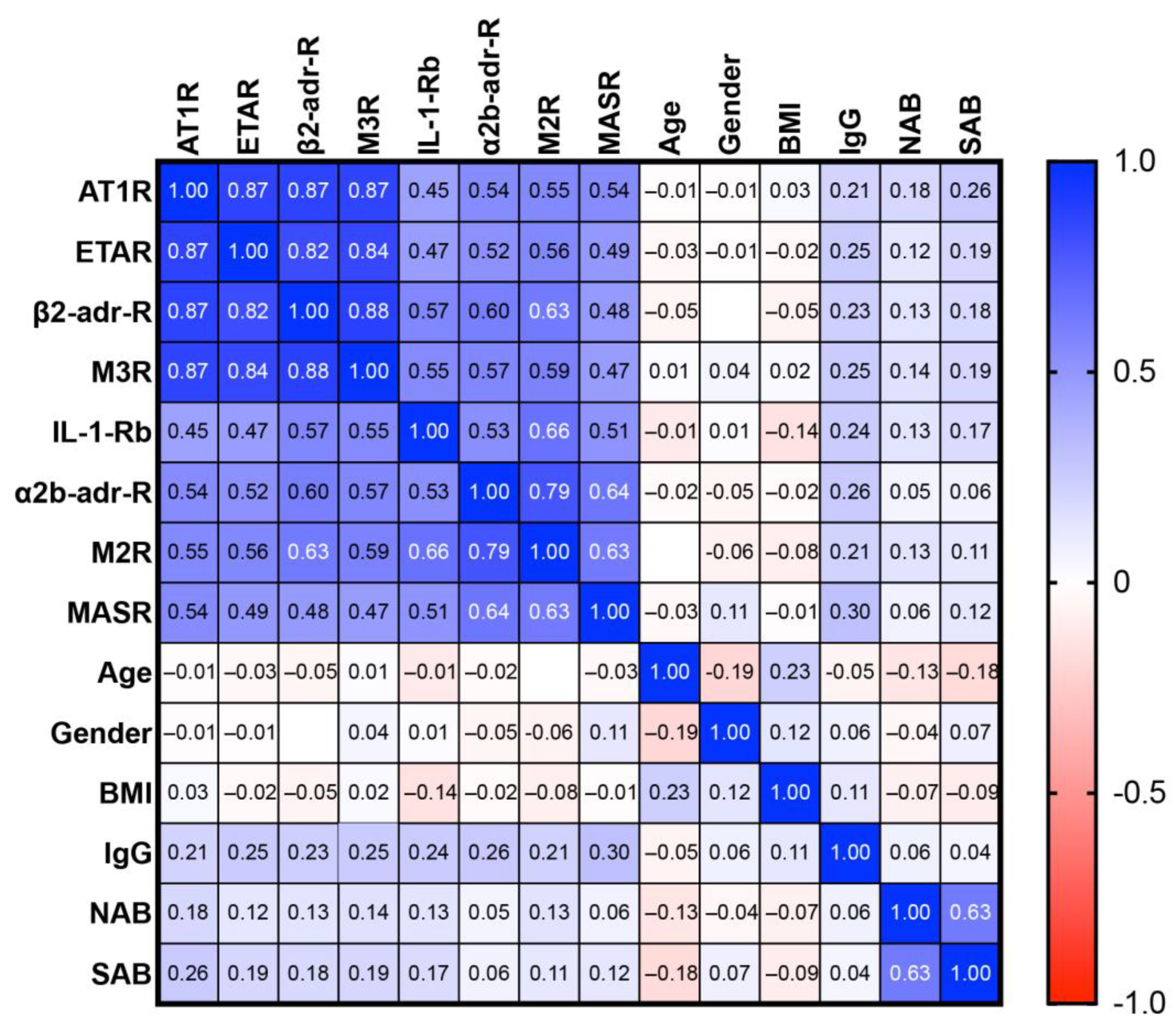
3.2. GPCR Antibodies in Post-Vaccination Controls and PACVS-Affected Subjects
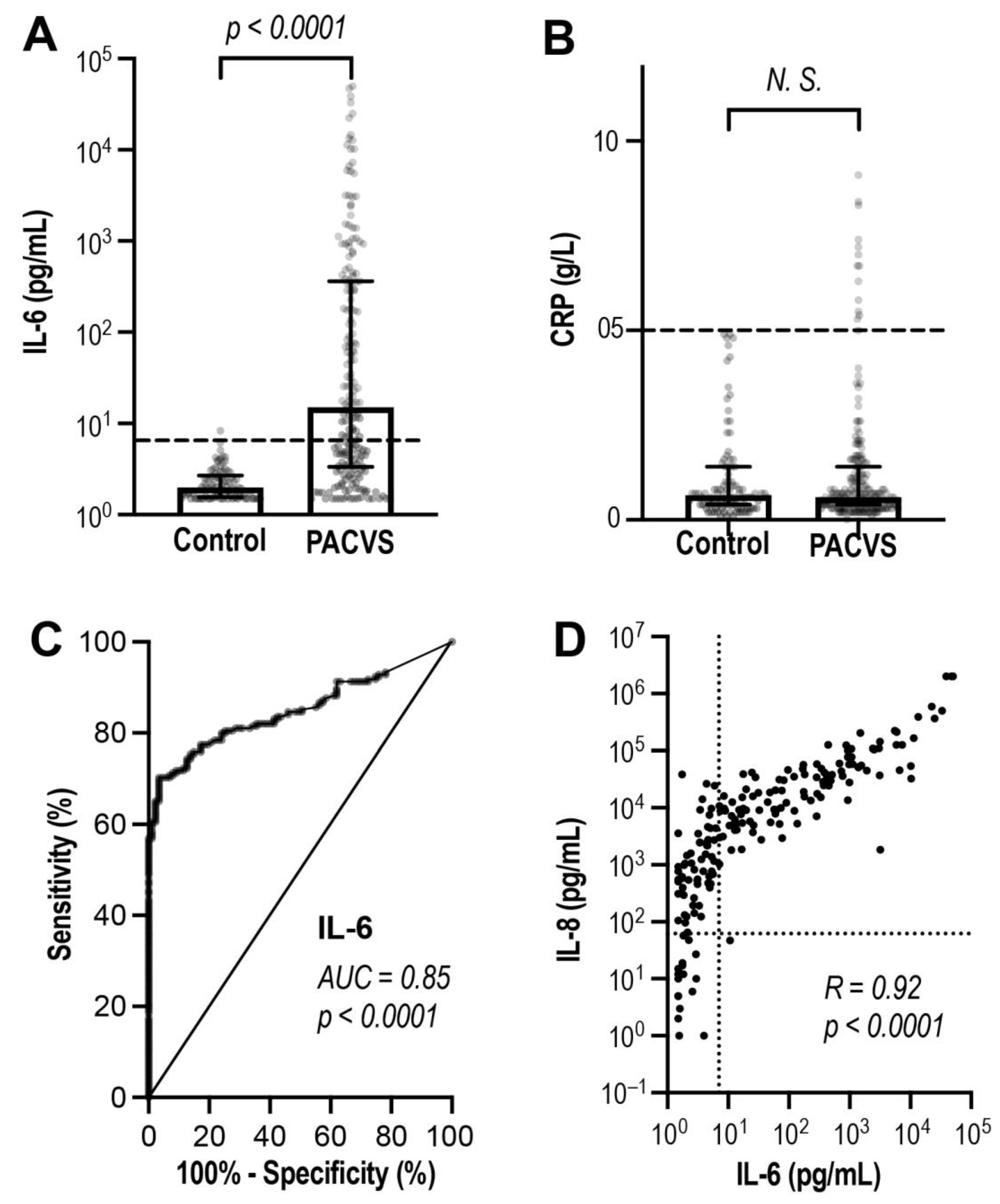
3.3. Discrimination of PACVS from Post-Vaccination Controls Based on Interleukins
3.4. Exclusion of SARS-CoV-2 Infection/COVID-19 Reconvalescence as Confounder of PACVS
4. Discussion
4.1. Salient Findings
- In healthy persons not affected by PACVS, the repertoire of receptor antibodies involved in cardiovascular regulation and immune homeostasis undergoes long-term adjustment following SARS-CoV-2 mRNA vaccination.
- The above adjustment seems blunted, absent or even inversed in persons who present clinical phenotypes of PACVS after SARS-CoV-2 mRNA vaccination.
- PACVS-afflicted persons can be distinguished from individuals subjected to SARS-CoV-2 mRNA vaccination without developing PACVS based on serum levels of IL-6/IL-8 and antibodies against AT1R and α2b-adr-R.
4.2. Limitations
- Our study is restricted to SARS-CoV-2 mRNA vaccines, for which we had an appropriate control cohort. Whether our findings apply to chronic sequelae following other types of SARS-CoV-2- vaccinations, or even vaccinations in general, remains to be investigated.
- The clinical PACVS phenotype studied here is based on a long list of symptoms. It is heterogeneous and possibly encompasses more than one clinical entity. Moreover, the selection of studied PACVS cases is biased by the exclusion of 71 applicants with potentially confounding co-morbidities or medications who could nevertheless suffer from PACVS.
- The PACVS cohort was recruited five or more months after vaccination. Matching pre-vaccination sera from these same persons could not be obtained. Consequently, vaccination-associated serological alterations in the PACVS cohort could not be determined intra-individually but had to be judged by comparing with a matched post-vaccination control cohort.
- Receptor antibodies were determined by IgG binding to the native receptors. We and others have previously demonstrated that such antibodies can modulate receptor function in several ways [38]; however, the functional properties of receptor antibodies were not directly assessed in this study.
- Our observation has been limited to a period of 5–6 months after vaccination. We do not know how long the observed effects last beyond this period.
4.3. The Physiological Response of Receptor Antibodies to SARS-CoV-2 mRNA Vaccination
- Downregulation of a cluster of receptor antibodies targeting the renin–angiotensin–aldosterone system and other components of cardiovascular regulation. Incidentally, some of these receptor antibodies are frequently increased in POTS [20,23,24], ME/CFS [18,22,25], severe COVID-19 [28,29,30,31,32], chronic heart failure [39,40] and allograft rejection [41]. The most distinctive candidate of this cluster is the AT1R antibody.
- Two receptor antibodies were upregulated. One of these, the IL-1-Rb antibody, is thought to play a role in immune homeostasis [35] and to have a protective effect against certain rheumatic diseases [42]. The α2b-adr-R receptor, on the other hand, plays a role in thrombogenesis and its inhibition by small molecule antagonists counteracts platelet aggregation induced by adenosine diphosphate, epinephrine or arachidonic acid in blood samples of healthy individuals [43].
4.4. Putative Pathogenic Role of Blunted Receptor Antibody Adaptation in PACVS
4.5. The Blood Marker Signature of PACVS
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| α1-adr-R-AB | Alpha-1 adrenergic receptor antibody |
| α2a-adr-R-AB | Alpha-2A adrenergic receptor antibody |
| α2b-adr-R-AB | Alpha-2B adrenergic receptor antibody |
| α2c-adr-R-AB | Alpha-2C adrenergic receptor antibody |
| ACE-II-AB | Angiotensin-converting enzyme 2 antibody |
| AT1R-AB | Angiotensin II type 1 receptor antibody |
| β1-adr-R-AB | Beta-1 adrenergic receptor antibody |
| β2-adr-R-AB | Beta-2 adrenergic receptor antibody |
| CRP | C-reactive protein |
| ETAR-AB | Endothelin-1 type A receptor antibody |
| IL-1-Rb-AB | Interleukin-1 receptor type 2 antibody |
| IL-6/-8 | Interleukin 6/8 |
| M1R-AB | muscarinic acetylcholine receptor M1 |
| M2R-AB | muscarinic acetylcholine receptor M2 |
| M3R-AB | muscarinic acetylcholine receptor M3 |
| M4R-AB | muscarinic acetylcholine receptor M4 |
| M5R-AB | muscarinic acetylcholine receptor M5 |
| MASR-AB | MAS 1 receptor antibody |
| MCAS | Mast cell activation syndrome |
| ME/CFS | Myalgic encephalomyelitis/chronic fatigue syndrome |
| NAB | PanIg reactivity against SARS-CoV-1 nucleocapsid protein |
| pBNP | pro-brain natriuretic peptide |
| PEM | Post exertional malaise |
| POTS | Postural tachycardia syndrome |
| PACVS | Post-acute COVID-19 vaccination syndrome |
| ROC | Receiver-operator characteristics |
| SAB | PanIg reactivity against SARS-CoV-1 spike S1 protein |
| SFN | Small fiber neuropathy |
References
- Scholkmann, F.; May, C.-A. COVID-19, post-acute COVID-19 syndrome (PACS, “long COVID”) and post-COVID-19 vaccination syndrome (PCVS, “post-COVIDvacsyndrome”): Similarities and differences. Pathol. Res. Pract. 2023, 246, 154497. [Google Scholar] [CrossRef] [PubMed]
- Almas, T.; Rehman, S.; Mansour, E.; Khedro, T.; Alansari, A.; Malik, J.; Alshareef, N.; Nagarajan, V.R.; Al-Awaid, A.H.; Alsufyani, R.; et al. Epidemiology, clinical ramifications, and cellular pathogenesis of COVID-19 mRNA-vaccination-induced adverse cardiovascular outcomes: A state-of-the-heart review. Biomed. Pharmacother. 2022, 149, 112843. [Google Scholar] [CrossRef] [PubMed]
- Finsterer, J. Neurological side effects of SARS-CoV-2 vaccinations. Acta Neurol. Scand. 2022, 145, 5–9. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Pan, J.; Zhang, C.; Sun, X. Cardiovascular Complications of COVID-19 Vaccines. Front. Cardiovasc. Med. 2022, 9, 840929. [Google Scholar] [CrossRef]
- Jara, L.J.; Vera-Lastra, O.; Mahroum, N.; Pineda, C.; Shoenfeld, Y. Autoimmune post-COVID vaccine syndromes: Does the spectrum of autoimmune/inflammatory syndrome expand? Clin. Rheumatol. 2022, 41, 1603–1609. [Google Scholar] [CrossRef]
- Lim, E.J.; Son, C.G. Review of case definitions for myalgic encephalomyelitis/chronic fatigue syndrome (ME/CFS). J. Transl. Med. 2020, 18, 289. [Google Scholar] [CrossRef]
- Carruthers, B.M.; van de Sande, M.I.; De Meirleir, K.L.; Klimas, N.G.; Broderick, G.; Mitchell, T.; Staines, D.; Powles, A.C.; Speight, N.; Vallings, R.; et al. Myalgic encephalomyelitis: International Consensus Criteria. J. Intern. Med. 2011, 270, 327–338. [Google Scholar] [CrossRef]
- Fedorowski, A. Postural orthostatic tachycardia syndrome: Clinical presentation, aetiology and management. J. Intern. Med. 2019, 285, 352–366. [Google Scholar] [CrossRef]
- Bair, M.J.; Krebs, E.E. Fibromyalgia. Ann. Intern. Med. 2020, 172, ITC33–ITC48. [Google Scholar] [CrossRef]
- Tavee, J.; Zhou, L. Small fiber neuropathy: A burning problem. Clevel. Clin. J. Med. 2009, 76, 297–305. [Google Scholar] [CrossRef]
- Valent, P.; Hartmann, K.; Bonadonna, P.; Niedoszytko, M.; Triggiani, M.; Arock, M.; Brockow, K. Mast Cell Activation Syndromes: Collegium Internationale Allergologicum Update 2022. Int. Arch. Allergy Immunol. 2022, 183, 693–705. [Google Scholar] [CrossRef] [PubMed]
- Blitshteyn, S.; Brook, J. Postural tachycardia syndrome (POTS) with anti-NMDA receptor antibodies after human papillomavirus vaccination. Immunol. Res. 2017, 65, 282–284. [Google Scholar] [CrossRef]
- Arana, J.; Mba-Jonas, A.; Jankosky, C.; Lewis, P.; Moro, P.L.; Shimabukuro, T.T.; Cano, M. Reports of Postural Orthostatic Tachycardia Syndrome After Human Papillomavirus Vaccination in the Vaccine Adverse Event Reporting System. J. Adolesc. Health 2017, 61, 577–582. [Google Scholar] [CrossRef]
- Martínez-Lavín, M. Hypothesis: Human papillomavirus vaccination syndrome—Small fiber neuropathy and dysautonomia could be its underlying pathogenesis. Clin. Rheumatol. 2015, 34, 1165–1169. [Google Scholar] [CrossRef] [PubMed]
- Hineno, A.; Ikeda, S.I. A Long-Term Observation on the Possible Adverse Effects in Japanese Adolescent Girls after Human Papillomavirus Vaccination. Vaccines 2021, 9, 856. [Google Scholar] [CrossRef] [PubMed]
- Afrin, L.B.; Dempsey, T.T.; Weinstock, L.B. Post-HPV-Vaccination Mast Cell Activation Syndrome: Possible Vaccine-Triggered Escalation of Undiagnosed Pre-Existing Mast Cell Disease? Vaccines 2022, 10, 127. [Google Scholar] [CrossRef]
- Agmon-Levin, N.; Zafrir, Y.; Kivity, S.; Balofsky, A.; Amital, H.; Shoenfeld, Y. Chronic fatigue syndrome and fibromyalgia following immunization with the hepatitis B vaccine: Another angle of the ‘autoimmune (auto-inflammatory) syndrome induced by adjuvants’ (ASIA). Immunol. Res. 2014, 60, 376–383. [Google Scholar] [CrossRef]
- Loebel, M.; Grabowski, P.; Heidecke, H.; Bauer, S.; Hanitsch, L.G.; Wittke, K.; Meisel, C.; Reinke, P.; Volk, H.D.; Fluge, O.; et al. Antibodies to beta adrenergic and muscarinic cholinergic receptors in patients with Chronic Fatigue Syndrome. Brain Behav. Immun. 2016, 52, 32–39. [Google Scholar] [CrossRef]
- Li, H.; Yu, X.; Liles, C.; Khan, M.; Vanderlinde-Wood, M.; Galloway, A.; Zillner, C.; Benbrook, A.; Reim, S.; Collier, D.; et al. Autoimmune basis for postural tachycardia syndrome. J. Am. Heart Assoc. 2014, 3, e000755. [Google Scholar] [CrossRef]
- Li, H.; Zhang, G.; Forsythe, E.; Okamoto, L.E.; Yu, X. Implications of Antimuscarinic Autoantibodies in Postural Tachycardia Syndrome. J. Cardiovasc. Transl. Res. 2022, 15, 438–440. [Google Scholar] [CrossRef]
- Kharraziha, I.; Axelsson, J.; Ricci, F.; Di Martino, G.; Persson, M.; Sutton, R.; Fedorowski, A.; Hamrefors, V. Serum Activity Against G Protein-Coupled Receptors and Severity of Orthostatic Symptoms in Postural Orthostatic Tachycardia Syndrome. J. Am. Heart Assoc. 2020, 9, e015989. [Google Scholar] [CrossRef] [PubMed]
- Freitag, H.; Szklarski, M.; Lorenz, S.; Sotzny, F.; Bauer, S.; Philippe, A.; Kedor, C.; Grabowski, P.; Lange, T.; Riemekasten, G.; et al. Autoantibodies to Vasoregulative G-Protein-Coupled Receptors Correlate with Symptom Severity, Autonomic Dysfunction and Disability in Myalgic Encephalomyelitis/Chronic Fatigue Syndrome. J. Clin. Med. 2021, 10, 3675. [Google Scholar] [CrossRef]
- Gunning, W.T., 3rd; Kvale, H.; Kramer, P.M.; Karabin, B.L.; Grubb, B.P. Postural Orthostatic Tachycardia Syndrome Is Associated With Elevated G-Protein Coupled Receptor Autoantibodies. J. Am. Heart Assoc. 2019, 8, e013602. [Google Scholar] [CrossRef]
- Gunning, W.T., 3rd; Stepkowski, S.M.; Kramer, P.M.; Karabin, B.L.; Grubb, B.P. Inflammatory Biomarkers in Postural Orthostatic Tachycardia Syndrome with Elevated G-Protein-Coupled Receptor Autoantibodies. J. Clin. Med. 2021, 10, 623. [Google Scholar] [CrossRef]
- Szklarski, M.; Freitag, H.; Lorenz, S.; Becker, S.C.; Sotzny, F.; Bauer, S.; Hartwig, J.; Heidecke, H.; Wittke, K.; Kedor, C.; et al. Delineating the Association Between Soluble CD26 and Autoantibodies Against G-Protein Coupled Receptors, Immunological and Cardiovascular Parameters Identifies Distinct Patterns in Post-Infectious vs. Non-Infection-Triggered Myalgic Encephalomyelitis/Chronic Fatigue Syndrome. Front. Immunol. 2021, 12, 644548. [Google Scholar] [CrossRef] [PubMed]
- Scheibenbogen, C.; Loebel, M.; Freitag, H.; Krueger, A.; Bauer, S.; Antelmann, M.; Doehner, W.; Scherbakov, N.; Heidecke, H.; Reinke, P.; et al. Immunoadsorption to remove ss2 adrenergic receptor antibodies in Chronic Fatigue Syndrome CFS/ME. PLoS ONE 2018, 13, e0193672. [Google Scholar] [CrossRef] [PubMed]
- Scheibenbogen, C.; Sotzny, F.; Hartwig, J.; Bauer, S.; Freitag, H.; Wittke, K.; Doehner, W.; Scherbakov, N.; Loebel, M.; Grabowski, P. Tolerability and Efficacy of s.c. IgG Self-Treatment in ME/CFS Patients with IgG/IgG Subclass Deficiency: A Proof-of-Concept Study. J. Clin. Med. 2021, 10, 2420. [Google Scholar] [CrossRef]
- Rodriguez-Perez, A.I.; Labandeira, C.M.; Pedrosa, M.A.; Valenzuela, R.; Suarez-Quintanilla, J.A.; Cortes-Ayaso, M.; Mayan-Conesa, P.; Labandeira-Garcia, J.L. Autoantibodies against ACE2 and angiotensin type-1 receptors increase severity of COVID-19. J. Autoimmun. 2021, 122, 102683. [Google Scholar] [CrossRef]
- Miedema, J.; Schreurs, M.; van der Sar-van der Brugge, S.; Paats, M.; Baart, S.; Bakker, M.; Hoek, R.; Dik, W.A.; Endeman, H.; Van Der Velden, V.; et al. Antibodies Against Angiotensin II Receptor Type 1 and Endothelin A Receptor Are Associated With an Unfavorable COVID19 Disease Course. Front. Immunol. 2021, 12, 684142. [Google Scholar] [CrossRef]
- Jiang, Y.; Duffy, F.; Hadlock, J.; Raappana, A.; Styrchak, S.; Beck, I.; Mast, F.D.; Miller, L.R.; Chour, W.; Houck, J.; et al. Angiotensin II receptor I auto-antibodies following SARS-CoV-2 infection. PLoS ONE 2021, 16, e0259902. [Google Scholar] [CrossRef]
- Cabral-Marques, O.; Halpert, G.; Schimke, L.F.; Ostrinski, Y.; Vojdani, A.; Baiocchi, G.C.; Freire, P.P.; Filgueiras, I.S.; Zyskind, I.; Lattin, M.T.; et al. Autoantibodies targeting GPCRs and RAS-related molecules associate with COVID-19 severity. Nat. Commun. 2022, 13, 1220. [Google Scholar] [CrossRef]
- Wallukat, G.; Hohberger, B.; Wenzel, K.; Furst, J.; Schulze-Rothe, S.; Wallukat, A.; Honicke, A.S.; Muller, J. Functional autoantibodies against G-protein coupled receptors in patients with persistent Long-COVID-19 symptoms. J. Transl. Autoimmun. 2021, 4, 100100. [Google Scholar] [CrossRef] [PubMed]
- Kedor, C.; Freitag, H.; Meyer-Arndt, L.; Wittke, K.; Hanitsch, L.G.; Zoller, T.; Steinbeis, F.; Haffke, M.; Rudolf, G.; Heidecker, B.; et al. A prospective observational study of post-COVID-19 chronic fatigue syndrome following the first pandemic wave in Germany and biomarkers associated with symptom severity. Nat. Commun. 2022, 13, 5104. [Google Scholar] [CrossRef] [PubMed]
- Bornstein, S.R.; Voit-Bak, K.; Donate, T.; Rodionov, R.N.; Gainetdinov, R.R.; Tselmin, S.; Kanczkowski, W.; Muller, G.M.; Achleitner, M.; Wang, J.; et al. Chronic post-COVID-19 syndrome and chronic fatigue syndrome: Is there a role for extracorporeal apheresis? Mol. Psychiatry 2021, 27, 34–37. [Google Scholar] [CrossRef] [PubMed]
- Cabral-Marques, O.; Marques, A.; Giil, L.M.; De Vito, R.; Rademacher, J.; Gunther, J.; Lange, T.; Humrich, J.Y.; Klapa, S.; Schinke, S.; et al. GPCR-specific autoantibody signatures are associated with physiological and pathological immune homeostasis. Nat. Commun. 2018, 9, 5224. [Google Scholar] [CrossRef]
- Davis, H.E.; Assaf, G.S.; McCorkell, L.; Wei, H.; Low, R.J.; Re’em, Y.; Redfield, S.; Austin, J.P.; Akrami, A. Characterizing long COVID in an international cohort: 7 months of symptoms and their impact. EClinicalMedicine 2021, 38, 101019. [Google Scholar] [CrossRef]
- Kuechler, A.S.; Weinhold, S.; Boege, F.; Adams, O.; Muller, L.; Babor, F.; Bennstein, S.B.; Pham, T.U.; Hejazi, M.; Reusing, S.B.; et al. A Diagnostic Strategy for Gauging Individual Humoral Ex Vivo Immune Responsiveness Following COVID-19 Vaccination. Vaccines 2022, 10, 1044. [Google Scholar] [CrossRef]
- Bornholz, B.; Weidtkamp-Peters, S.; Schmitmeier, S.; Seidel, C.A.; Herda, L.R.; Felix, S.B.; Lemoine, H.; Hescheler, J.; Nguemo, F.; Schafer, C.; et al. Impact of human autoantibodies on beta1-adrenergic receptor conformation, activity, and internalization. Cardiovasc. Res. 2013, 97, 472–480. [Google Scholar] [CrossRef]
- Boivin-Jahns, V.; Jahns, R. GPCR-autoantibodies in chronic heart failure. Front. Biosci. (Landmark Ed.) 2018, 23, 2065–2081. [Google Scholar] [CrossRef]
- Wallukat, G.; Schimke, I. Agonistic autoantibodies directed against G-protein-coupled receptors and their relationship to cardiovascular diseases. Semin. Immunopathol. 2014, 36, 351–363. [Google Scholar] [CrossRef]
- Dragun, D.; Muller, D.N.; Brasen, J.H.; Fritsche, L.; Nieminen-Kelha, M.; Dechend, R.; Kintscher, U.; Rudolph, B.; Hoebeke, J.; Eckert, D.; et al. Angiotensin II type 1-receptor activating antibodies in renal-allograft rejection. N. Engl. J. Med. 2005, 352, 558–569. [Google Scholar] [CrossRef]
- Cabral-Marques, O.; Riemekasten, G. Functional autoantibodies targeting G protein-coupled receptors in rheumatic diseases. Nat. Rev. Rheumatol. 2017, 13, 648–656. [Google Scholar] [CrossRef] [PubMed]
- Marketou, M.E.; Kintsurashvili, E.; Androulakis, N.E.; Kontaraki, J.; Alexandrakis, M.G.; Gavras, I.; Vardas, P.E.; Gavras, H. Blockade of platelet alpha2B-adrenergic receptors: A novel antiaggregant mechanism. Int. J. Cardiol. 2013, 168, 2561–2566. [Google Scholar] [CrossRef] [PubMed]
- Meyer, C.; Heidecke, H. Antibodies Against GPCR. Front. Biosci. Landmark Ed. 2018, 23, 2177–2194. [Google Scholar] [PubMed]
- Bester, J.; Pretorius, E. Effects of IL-1beta, IL-6 and IL-8 on erythrocytes, platelets and clot viscoelasticity. Sci. Rep. 2016, 6, 32188. [Google Scholar] [CrossRef] [PubMed]
- Seidel, F.; Scheibenbogen, C.; Heidecke, H.; Opgen-Rhein, B.; Pickardt, T.; Klingel, K.; Berger, F.; Messroghli, D.; Schubert, S. Compensatory Upregulation of Anti-Beta-Adrenergic Receptor Antibody Levels Might Prevent Heart Failure Presentation in Pediatric Myocarditis. Front. Pediatr. 2022, 10, 881208. [Google Scholar] [CrossRef]
- Fraser, C.M.; Venter, J.C. Anti-receptor antibodies in human disease. J. Allergy Clin. Immunol. 1984, 74, 661–673. [Google Scholar] [CrossRef]
- Yin, J.X.; Agbana, Y.L.; Sun, Z.S.; Fei, S.W.; Zhao, H.Q.; Zhou, X.N.; Chen, J.H.; Kassegne, K. Increased interleukin-6 is associated with long COVID-19: A systematic review and meta-analysis. Infect. Dis. Poverty 2023, 12, 43. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Median 1 | 25% Perc. | 75% Perc. | ∆ vs. PACVS (p, U-Test) | |
|---|---|---|---|---|
| AT1R | ||||
| PACVS 2 (n = 191) | 15.2 | 12.1 | 21.3 | - |
| Contr. pre 3 (n = 89) | 15.6 | 12.4 | 21.1 | N.S. 5 |
| Contr. post 4 (n = 89) | 10.4 | 8.1 | 12.4 | <0.0001 |
| ETAR | ||||
| PACVS (n = 191) | 13.5 | 10.8 | 18.5 | - |
| Contr. pre (n = 99) | 15.4 | 11.6 | 19.5 | N.S. |
| Contr. post (n = 89) | 11.0 | 8.8 | 13.8 | 0.0001 |
| IL-1-Rb | ||||
| PACVS (n = 191) | 4.9 | 3.8 | 6.9 | - |
| Contr. pre (n = 89) | 5.1 | 4.2 | 7.2 | N.S. |
| Contr. post (n = 89) | 6.2 | 5.3 | 8.2 | <0.0001 |
| M3R | ||||
| PACVS (n = 191) | 10.6 | 7.9 | 16.4 | - |
| Contr. pre (n = 89) | 11.9 | 8.0 | 18.2 | N.S. |
| Contr. post (n = 89) | 6.6 | 4.8 | 9.2 | <0.0001 |
| β2-adr-R | ||||
| PACVS (n = 191) | 12.8 | 8.9 | 16.6 | - |
| Contr. pre (n = 89) | 20.9 | 11.2 | 39.6 | N.S. |
| Contr. post (n = 89) | 9.3 | 5.8 | 14.4 | <0.0001 |
| MASR | ||||
| PACVS (n = 191) | 50.2 | 41.7 | 62.1 | - |
| Contr. pre (n = 89) | 53.1 | 42.8 | 67.6 | N.S. |
| Contr. post (n = 89) | 39.2 | 31.7 | 45.7 | <0.0001 |
| M2R | ||||
| PACVS (n = 191) | 11.8 | 8.9 | 16.9 | - |
| Contr. pre (n = 89) | 16.9 | 11.3 | 27.2 | <0.0001 |
| Contr. post (n = 89) | 7.7 | 6.2 | 11.7 | <0.0001 |
| α2b-adr-R | ||||
| PACVS (n = 191) | 13.8 | 9.9 | 18.6 | - |
| Contr. pre (n = 89) | 21.6 | 13.8 | 30.6 | <0.0001 |
| Contr. post (n = 89) | 27.9 | 20.9 | 43.2 | <0.0001 |
| ROC (AUC ± SE) | ROC (p) | Cut-off (U/mL) 1 | Sensitivity (%) 2 | |
|---|---|---|---|---|
| AT1R | 0.824 ± 0.027 | <0.0001 | ≤10.7 | 89.7 |
| ETAR | 0.681 ± 0.035 | <0.0001 | ≤11.5 | 64.9 |
| M3R | 0.741 ± 0.034 | <0.0001 | ≤12.4 | 40.3 |
| β2-adr-R | 0.681 ± 0.036 | <0.0001 | ≤11.6 | 66.5 |
| α2b-adr-R | 0.828 ± 0.025 | <0.0001 | ≥25.2 | 90.3 |
| M2R | 0.703 ± 0.034 | <0.0001 | ≥14.2 | 64.4 |
| MASR | 0.675 ± 0.037 | <0.0001 | ≤44.0 | 72.3 |
| IL-1-Rb | 0.913 ± 0.019 | <0.0001 | ≥5.8 | 66.5 |
| IL-6 | 0.850 ± 0.022 | <0.0001 | ≥2.3 | 82.0 |
| PACVS ± COVID 1 | PACVS w/o COVID vs. post-vacc. CTR 2 | |||
|---|---|---|---|---|
| Median Effect Size 3 (%) | Significance (p) 4 | Median Effect Size 3 (%) | Significance (p) 4 | |
| AT1R | +12.8 | 0.01 | +43 | <0.0001 |
| ETAR | +7.9 | 0.11 | ||
| β2-adr-R | +7.2 | 0.07 | ||
| M3R | +20.3 | 0.05 | +44.4 | <0.0001 |
| IL-1-Rb | +6.3 | 0.08 | ||
| α2b-adr-R | +4.8 | 0.50 | ||
| M2R | +9.5 | 0.06 | ||
| MASR | +4.1 | 0.40 | ||
| IL-6 | −1.3 | 0.33 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Semmler, A.; Mundorf, A.K.; Kuechler, A.S.; Schulze-Bosse, K.; Heidecke, H.; Schulze-Forster, K.; Schott, M.; Uhrberg, M.; Weinhold, S.; Lackner, K.J.; et al. Chronic Fatigue and Dysautonomia following COVID-19 Vaccination Is Distinguished from Normal Vaccination Response by Altered Blood Markers. Vaccines 2023, 11, 1642. https://doi.org/10.3390/vaccines11111642
Semmler A, Mundorf AK, Kuechler AS, Schulze-Bosse K, Heidecke H, Schulze-Forster K, Schott M, Uhrberg M, Weinhold S, Lackner KJ, et al. Chronic Fatigue and Dysautonomia following COVID-19 Vaccination Is Distinguished from Normal Vaccination Response by Altered Blood Markers. Vaccines. 2023; 11(11):1642. https://doi.org/10.3390/vaccines11111642
Chicago/Turabian StyleSemmler, Amelie, Anna Katharina Mundorf, Anna Sabrina Kuechler, Karin Schulze-Bosse, Harald Heidecke, Kai Schulze-Forster, Matthias Schott, Markus Uhrberg, Sandra Weinhold, Karl J. Lackner, and et al. 2023. "Chronic Fatigue and Dysautonomia following COVID-19 Vaccination Is Distinguished from Normal Vaccination Response by Altered Blood Markers" Vaccines 11, no. 11: 1642. https://doi.org/10.3390/vaccines11111642