
Lumpy Skin Disease—An Emerging Cattle Disease in Europe and Asia

Institute of Infectious Disease and Molecular Medicine and Division of Medical Virology, Department of Pathology, University of Cape Town, Cape Town 7700, South Africa
*
Author to whom correspondence should be addressed.
Vaccines 2023, 11(3), 578; https://doi.org/10.3390/vaccines11030578
Submission received: 25 January 2023
/
Revised: 20 February 2023
/
Accepted: 22 February 2023
/
Published: 2 March 2023
(This article belongs to the Special Issue Veterinary Vaccines and Diagnostic Tools 2.0)
Abstract
:Lumpy skin disease virus (LSDV) is a member of the Capripoxvirus genus, mainly infecting cattle and buffalo, which until relatively recently was only endemic in parts of Africa and then spread to the Middle East and lately Europe and Asia. Lumpy skin disease (LSD) is a notifiable disease with a serious impact on the beef industry as it causes mortality of up to 10% and has impacts on milk and meat production, as well as fertility. The close serological relationship between LSDV, goat poxvirus (GTPV) and sheep poxvirus (SPPV) has led to live attenuated GTPV and SPPV vaccines being used to protect against LSD in some countries. There is evidence that the SPPV vaccine does not protect from LSD as well as the GTPV and LSDV vaccines. One of the LSD vaccines used in Eastern Europe was found to be a combination of different Capripoxviruses, and a series of recombination events in the manufacturing process resulted in cattle being vaccinated with a range of recombinant LSDVs resulting in virulent LSDV which spread throughout Asia. It is likely that LSD will become endemic throughout Asia as it will be very challenging to control the spread of the virus without widespread vaccination.
1. Introduction
1.1. Background
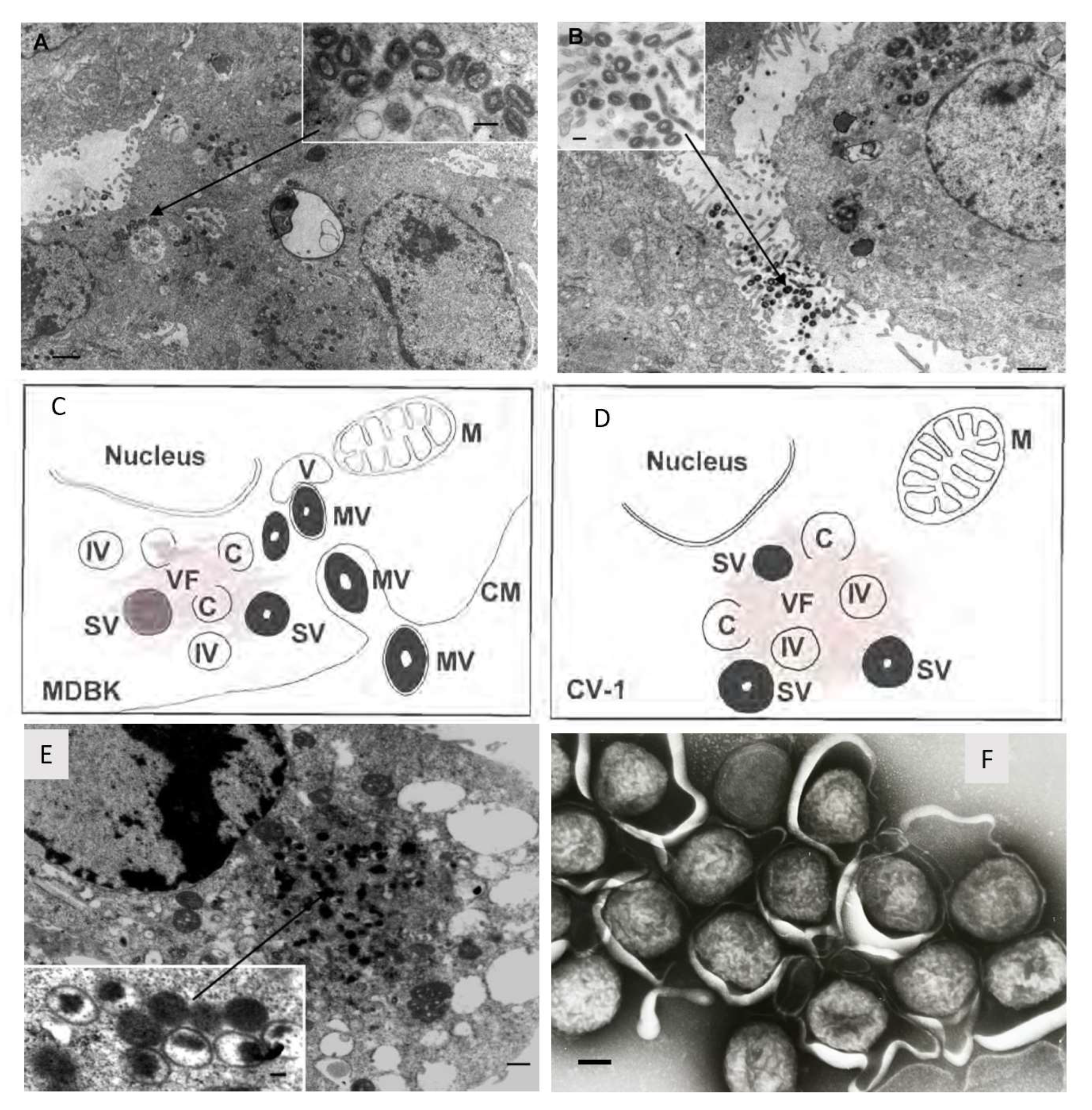
Lumpy skin disease virus (LSDV), which causes lumpy skin disease (LSD) in cattle, belongs to the Capripoxvirus genus of the Poxviridae family. Other members of the Capripoxvirus genus are sheeppox virus (SPPV) and goatpox virus (GTPV). Capripoxvirus particles are enveloped and brick-shaped measuring 294 ± 20 nm in length and 262 ± 22 nm in width [1]. Capripoxviruses have a large covalently linked double stranded DNA genome of 150 kb for SPPV and GTPV [2] and 151 kb for LSDV [3]. As seen with other poxviruses, LSDV replicates in the cytoplasm. The viral factory is established and then crescents are formed which will develop into immature virus and then intracellular mature virus. In the Golgi body or early endosome, the intracellular mature viruses are enveloped and then can be exported out of the cell to yield extracellular enveloped viruses [4,5,6]. In non-permissive cells, the life cycle is halted before maturity and although immature virus can be observed, no mature virus develops (refer to Figure 1) [6].
LSD is listed as a notifiable disease by the World Organization for Animal Health (OIE) due to the severe economic impact on the cattle industry. This is due to a number of factors including decreased milk and meat production, abortions, fertility problems, damaged hides and in some cases, death of the animals. LSD can also result in secondary bacterial infections. The resulting trade restrictions and response to outbreaks including vaccination and treatment further amplify the economic losses [7,8].
Figure 1.
Morphogenesis of recombinant LSDV-Rabies which is identical to LSDV. Permissive bovine Madin-Darby bovine kidney (MDBK) cells were infected with rLSDV-Rabies (1 f.f.u. per cell, 48 h; scale bar = 1 mm) with mature virions inside (A) and outside (B) the cell. The inserts show high-power virion structure (scale bar = 250 nm). A diagrammatic representation of rLSDV-RG replication in permissive (C) non-permissive (D) cells. M indicates mitochondria, C indicates crescent shaped membrane which precedes immature virion (IV) formation, (V) indicates vacuoles, (VF) indicates viral factory and electron dense areas where replication and maturation occurs, (IV) indicates immature virion, (MV) indicates mature virion, (CM) indicates cell membrane, and (SV) indicates semi-mature virion. (E) Non-permissive primate CV-1 cells were infected with rLSDV-Rabies (1 f.f.u. per cell, 48 h; scale bar = 0.5 mm). The insert is a higher magnification of the ‘viral factory’ (scale bar = 100 nm). Taken from [6,9]. (F) LSDV negatively stained(scale bar = 100 nm). Taken by Linda Stannard.
Figure 1.
Morphogenesis of recombinant LSDV-Rabies which is identical to LSDV. Permissive bovine Madin-Darby bovine kidney (MDBK) cells were infected with rLSDV-Rabies (1 f.f.u. per cell, 48 h; scale bar = 1 mm) with mature virions inside (A) and outside (B) the cell. The inserts show high-power virion structure (scale bar = 250 nm). A diagrammatic representation of rLSDV-RG replication in permissive (C) non-permissive (D) cells. M indicates mitochondria, C indicates crescent shaped membrane which precedes immature virion (IV) formation, (V) indicates vacuoles, (VF) indicates viral factory and electron dense areas where replication and maturation occurs, (IV) indicates immature virion, (MV) indicates mature virion, (CM) indicates cell membrane, and (SV) indicates semi-mature virion. (E) Non-permissive primate CV-1 cells were infected with rLSDV-Rabies (1 f.f.u. per cell, 48 h; scale bar = 0.5 mm). The insert is a higher magnification of the ‘viral factory’ (scale bar = 100 nm). Taken from [6,9]. (F) LSDV negatively stained(scale bar = 100 nm). Taken by Linda Stannard.

LSD manifests in cattle as fever, nasal and ocular discharge, and painful nodular lesions which form on the skin, muscles and mucosal tissue (Figure 2) [10,11]. Light and ultrastructural microscopy studies of infected cattle showed vasculitis and thrombosis to be central in the pathogenesis with replicating viral particles seen in the endothelial cells [5].
1.2. Host-Range
LSDV is host restricted; its natural hosts are cattle, buffalo and water buffalo [12,13,14]. Antibodies have been detected in black and blue wildebeest, eland, giraffe, greater kudu, African buffalo and other animal species [15]. Giraffe and impala can be experimentally infected and subsequently die from LSD [16]. Recently, LSDV was isolated from nodular lesions of a giraffe which died in a zoo in Vietnam [17]. The authors suggest that it was due to natural infection of the virus, although how this occurred is unknown. The role of wild animals in the epidemiology of LSDV is still not certain.
The host range is slightly broader in cell culture of LSDV. A number of primary cell types derived from lamb and bovine tissue enable the growth of LSDV to high titres and are commonly used for vaccine stock preparations [18,19]. LSDV also grows well in chick chorioallantoic membranes (CAMs) of embryonated eggs, although purification of the virus may need to be extensive which can cause a loss in yields [20]. Various cell lines have been investigated as more convenient alternatives. High titres of LSDV can be obtained when grown in Madin-Darby bovine kidney (MDBK) cells [21]. Unfortunately, this cell line is often contaminated with bovine viral diarrhoea virus (BVDV), and therefore cannot be used for the preparation of vaccine stocks. Removal of BVDV from LSDV cultured in MDBKs can however be achieved after two passages in CAMs [22]. Another is the baby hamster kidney 21 (BHK-21) cell line which was first investigated for LSDV culture in the 1950s–1960s, although it is not frequently used for this purpose [23]. Our laboratory very recently demonstrated that BHK-21 cells can be used for the construction of LSDV recombinants [24]. A recently developed embryonic skin of sheep (ESH-L) cell line was also shown to yield high titres of LSDV [19]. LSDV also replicates in Vero (African green monkey kidney) cells and the ovine testis (OA3.Ts) cell line [18,25].
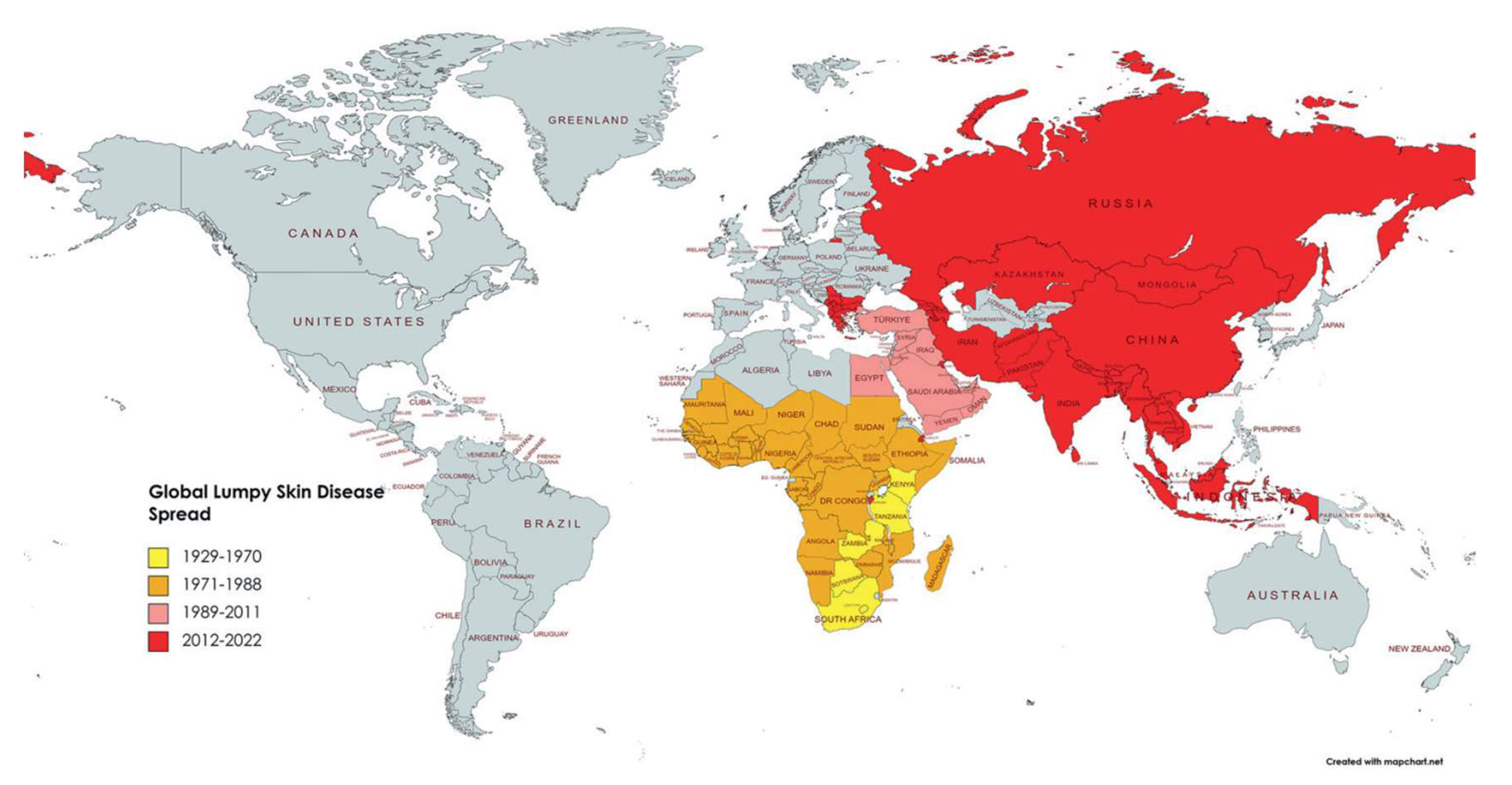
1.3. Distribution of LSD
Last century, LSD was regarded as a disease endemic to Africa with the first report from Northern Rhodesia (now Zambia) in 1929. In 1989, LSD spread to Israel [26] and then further to other countries in the Middle East [27]. In 2015, LSDV was found in the Balkans and in 2016, in Serbia [28]. In the last decade, LSDV spread to countries in Europe, Bangladesh, China, India and Russia [29,30,31,32,33]. Outbreaks of the disease also occurred more recently in Indonesia and Pakistan last year [34] (refer to Figure 3). Outbreaks in parts of Europe have been controlled [35], but it is likely that LSD will become endemic in most parts of Africa and Asia. The outbreak in the Balkans was controlled by vaccination with a live attenuated LSD vaccine based on the Neethling strain. This vaccine provided protection from infection within 14 days of vaccination [35]. However, there is always the concern of further spread of the disease. Vaccination is the best way to control the spread of LSD as subclinical infections are common and identification and removal of infected animals is not always possible [36].
Figure 3.
Lumpy skin disease prevalence worldwide and over time from 1929 to 2022. The impacted nations are shown in yellow between 1929 and 1970, orange between 1971 and 1988, pink between 1989 and 2011, and red between 2012 and 2022, with permission from A. Khalafalla, [37].
Figure 3.
Lumpy skin disease prevalence worldwide and over time from 1929 to 2022. The impacted nations are shown in yellow between 1929 and 1970, orange between 1971 and 1988, pink between 1989 and 2011, and red between 2012 and 2022, with permission from A. Khalafalla, [37].

1.4. Transmission
The main route of LSDV transmission is via a variety of arthropod vectors: ticks, mosquitoes and biting flies have all been implicated in transmission [38,39,40,41,42,43,44]. Climate change may impact on the spread of the arthropod vectors and as a result, the spread of LSDV [45]. To reduce the risk of insect transmission into areas with no LSDV, disinsectisation of vehicles transporting live animals is important [45].
LSDV is very stable in the environment, persisting for an extended time in necrotic lesions, desiccated crusts and airdried hides [46]. Direct contact between animals was initially deemed as inefficient for LSDV spread [47]. However, more recent work demonstrated the transmission of a virulent recombinant strain of LSDV (Saratov/2017) between infected bulls and in contact cattle in the absence of a vector [48]. There are also gaps in knowledge on how LSDV can persist in the environment, as there is a report in Russia where two almost identical isolates were identified with a two-year gap, with the implication that the virus circulated in cattle in the region or survived in fomites [49]. The presence of LSDV has been reported in semen after virulent challenge experiments [50] and insemination of heifers with infected semen can cause transmission of LSDV [51]. Of note, vaccination prevented LSDV being shed in semen after virulent challenge [50]. In natural outbreaks, LSDV can be transmitted to bovine foetuses and can result in abortion [52].
1.5. Current LSDV Vaccines Used in the Field
LSD is currently controlled using commercial vaccines which are mainly live attenuated LSDVs [53] which originated from the Neethling field strain of LSDV. This virus was attenuated by serial passage in primary lamb kidney cells and CAMs, carried out at the Onderstepoort Veterinary Research Institute (South Africa) in the 1950s–1960s [23,54]. This is known as the Neethling vaccine strain of LSDV (nLSDV). In South Africa, Neethling-like LSDV vaccine strains are marketed as Lumpyvax (MSD Animal Health), Herbivac LS (Deltamune) and Lumpy Skin Disease Vaccine (Onderstepoort Biological Products—OBP) and share 99.5% DNA sequence homology with each other [55]. nLSDV is also the most commonly used LSDV vaccine strain in Europe [56].
Due to the cross-reactivity between members of the Capripoxvirus genus, live attenuated GTPV and SPPV are also used to vaccinate cattle against LSDV [57]. GTPV-based vaccines have been reported to show the same protection against LSD as LSDV-based vaccines [58,59]. However, this is not the case with SPPV vaccines. In an experiment where groups of sheep, goats and cattle, vaccinated with Romania SPPV vaccine or nLSDV vaccine, were challenged with corresponding virulent strains, goats or sheep receiving the Romania SPPV vaccine were fully protected against challenge with virulent SPPV and GTPV strains, respectively. However, those cattle that were vaccinated with Romania SPPV vaccine showed only partial protection against LSDV challenge compared to full protection in cattle that received the nLSDV vaccine [60]. These results indicate that either LSDV or GTPV based vaccines could be used to protect from LSD in cattle.
1.6. Efficacy and Safety of Capripoxvirus Vaccines
A trial was conducted to test five LSDV-based live attenuated vaccines, namely: (1) Lumpy Skin Disease Vaccine [61] (Onderstepoort Biological Products OBP; South-Africa; batch 442); (2) Lumpyvax (MSD-Animal Health; South-Africa; batch BNDM07); (3) Kenyavac (Jordan Bioindustries Center Jovac; Jordan; batch 220115-04); (4) Herbivac LS (Deltamune; South-Africa); (5) Vaccin LSD Neethling O vivant (MCI Santé Animale; Morocco, batch 17BLSDN001). Animals were vaccinated and then challenged with virulent LSDV, 21 days after the last vaccination, and were all protected from challenge. They found that vaccination often resulted in a fever, which varied between the groups. Lumpyvax had a greater response whereas MCI- and Herbivac-vaccinated animals had less response. However, despite this, a Neethling response was seen in 43% of the animals vaccinated with Herbivac and 28% of the animals vaccinated with the MCI vaccine [53]. A Neethling response is the presence of superficial and smaller skin lesions which are distinct from those induced by a virulent field strain that disappear within 2–3 weeks without converting into necrotic scabs or ulcers [62]. Although some regard a Neethling response as negligible [62], these mild reactions have resulted in some hesitancy to use the live vaccines.
Adverse reactions in cattle vaccinated with a SPPV and an unverified LSDV vaccine were reported by farmers in Jordan; however, there is suspicion the LSDV vaccine was a smuggled virulent strain [63]. A randomised controlled field study on over 4000 cattle in Israel confirmed the efficacy of nLSDV vaccines compared to SPPV (x10RM65)-based vaccines. The relative risk of getting LSD in x10RM65 vs. nLSDV vaccinated animals was 2.635 (CI 95% = 1.44–4.82) and 11.2 (2.3–54.7) for severe morbidity. The relative risk of disease in x10RM65 vs. nLSDV vaccinated animals for laboratory confirmed cases was even higher at 4.28 (1.59–11.53), leading to the conclusion that nLSDV is significantly more effective at preventing LSD than x10RM65 [64].
1.7. Recombinant LSDV in the Field
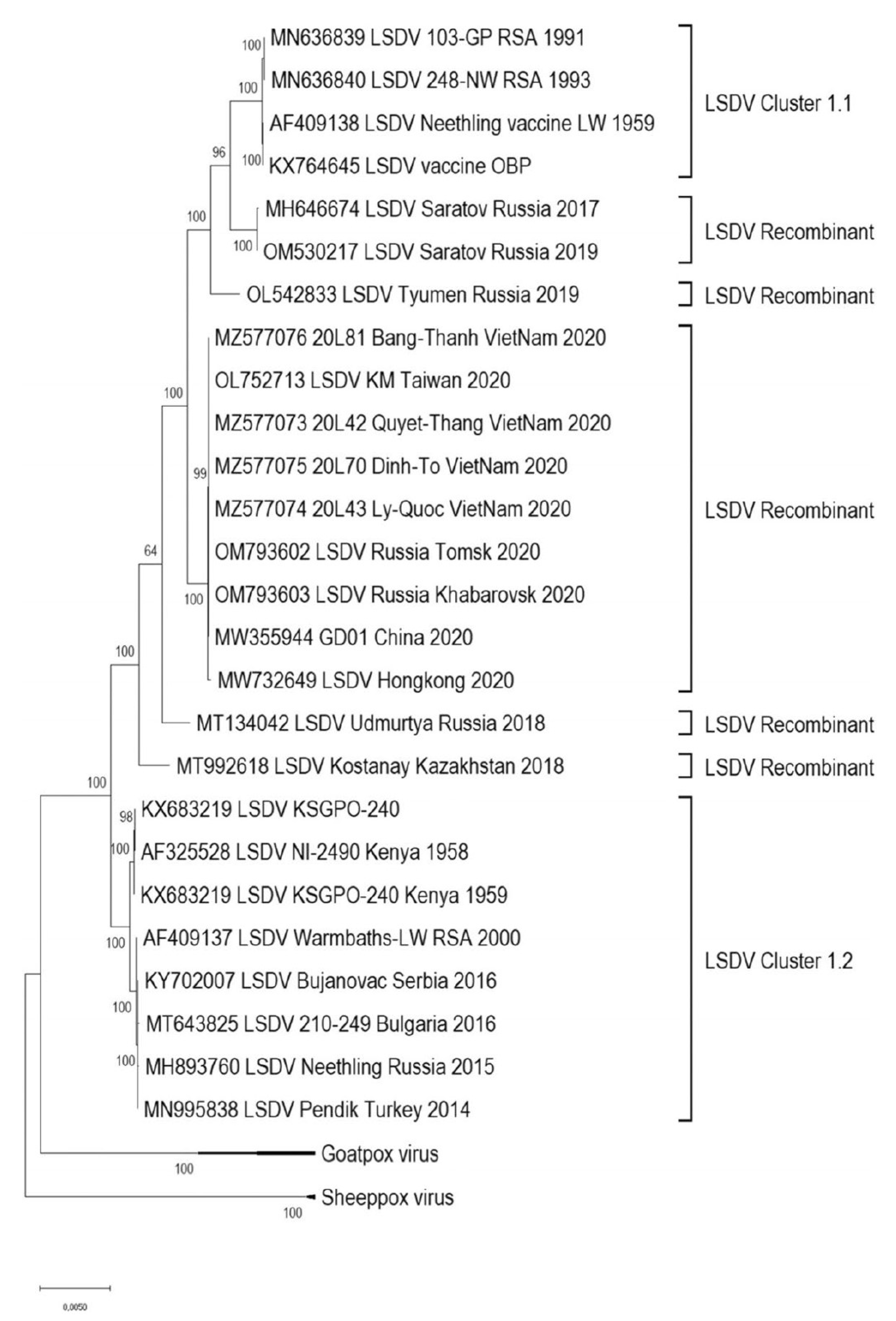
In the past, LSDV genomes were reported to be relatively stable. For example, when comparing a virulent LSDV field-strain from South Africa and a virulent Kenyan 2490 strain, there were only 38 amino acid differences in 29 of the 156 putative genes and mostly 1 to 3 amino acid changes in the variable terminal regions. There were also six deletions and eight insertions detected [66]. In a study comparing the prototype, the Neethling-WC/RSA/1957 strain from which the vaccine was derived, and the vaccine prepared in 1988 (LW-1959), only seven single-nucleotide polymorphisms were identified. In addition, the oldest available LSDV isolate LSDV/Haden/RSA/1954 was sequenced and compared with Neethling-type commercial vaccine viruses and had eight SNP differences. Both viruses formed part of Cluster 1.1 (refer to Figure 4). It is estimated that the substitution rate of LSDV is in the range 7.4 × 10−6 substitutions/site/year. Natural genetic drift is responsible for the differences between the phylogenetic clusters. In the natural population, there is a high frequency of synonymous mutations whereas highly cell passaged viruses have an increase in non-synonymous mutations. LSDV has high genetic stability and is well adapted to the cattle host [67]. This differs from the recent human outbreak of monkeypox virus, which is still adapting to the new human host, where point mutations continue to accumulate at an unexpected rate [68].
However, it is also well accepted that poxviruses can recombine under specific conditions to give chimeric viruses. The mechanisms include recombination, and horizontal gene transfer to acquire new genes [69]. The first experiments to show recombination between different orthopoxviruses were conducted in the 1960s [70,71,72]. Marker rescue studies were carried out to demonstrate how homologous recombination could be used to generate recombinant poxviruses with novel inserts [73] and this subsequently evolved into making recombinant poxvirus-based vaccines [74]. Poxviruses under specific stresses can duplicate genes to adapt to new conditions [69,75,76]. However, there was minimal evidence that Capripoxviruses would recombine in the field [77]. Prior to 2017, the LSDVs could be grouped into two clusters that were separate from GPPV and SPPV [78] (refer to Figure 4).
With this background, it was unexpected to identify recombinant LSDV in the field. Kazakhstan used a vaccine called Lumpivax (KEVEVAPI, Narobi, Kenya) to control an LSD outbreak (2017–2019) and following this, there was an emergence of vaccine-like LSDV recombinants in Kazakhstan and neighbouring China and Russia. One of the first reports of virulent vaccine-like strains of LSDV was from a LSD outbreak in Russia in 2017. An in-depth analysis of one of the recombinants, LSDV RUSSIA/Saratov/2017 identified 27 recombination events and included fragments from wild-type field strains of Capripoxviruses [79]. It was speculated that the vaccine being used in neighbouring Kazakhstan had reverted to virulence since, at that time, attenuated LSDV-based vaccines were not permitted in Russia [80]. It was further hypothesised that the recombinant LSDV had a fitness advantage in the colder weather conditions of the northern latitudes [78].
Further analysis of emerging recombinants showed a number of unique recombinants that emerged before 2020. From 2020 onwards, all circulating strains in Russia and South-Eastern Asia belonged to a single lineage radiating out in the region. After 2017, with the emergence of recombinant LSDVs from the Russian Federation, Kazakhstan, China and Vietnam, further distinct clusters were seen (refer to Figure 4) [78]. The recombination events were predicted to be between parental strains, a Kenyan KSGPO-like and LSDV/Vaccine/LW-1959-like strain. A comparison of recombination sites in five recombinant viruses demonstrated a unique mosaic pattern for each. However, there were some common break points at positions 80,000 bp, 84,000 bp, 124,000 bp and 141,000 bp (refer to Figure 5) [78]. These unique recombinant viruses illustrate the importance of whole genome sequencing to track poxviruses and conduct molecular epidemiology.
Further proof on the origin of the recombinant LSDVs came when the analysis of two batches of the vaccine Lumpivax found it to be a combination of different Capripoxviruses including a Neethling-like LSDV vaccine strain, a KSGP-like LSDV vaccine strain and a Sudan-like GTPV strain [27]. In this study, they directly analysed the genomes present in the vaccines and found that the vaccine-like recombinant strains could be divided into four groups. They each arose from multiple recombination events with a large number of genetic exchanges ranging between 126 and 146. Since the viruses were sequenced directly from the vaccine lots, it is likely the recombination occurred during the manufacturing process. Not all the recombinants would be expected to have equal fitness in animals and the most fit would be selected and transmitted. The emergence of recombinant vaccine-like LSDV strains in Asia is now thought to originate from spill-over from animals vaccinated with Lumpivax, highlighting the importance of vaccine quality control [27]. Whole genome sequencing of vaccine stocks should be part of the quality checks.
Figure 4.
Maximum likelihood phylogenetic tree representing genetic relationship of the complete genome sequences of 26 LSDVs, 2 GPVs and 2 SPPVs with permission from A. Sprygin [78].
Figure 4.
Maximum likelihood phylogenetic tree representing genetic relationship of the complete genome sequences of 26 LSDVs, 2 GPVs and 2 SPPVs with permission from A. Sprygin [78].

Figure 5.
Graphical representation of the position of the predicted recombination events across the five strains with the highest statistical significance. The events are indicated in black in the white backbone of the viral genome. Possible regions selected for by recombination are indicated by arrows (positions 80,000 bp, 124,000 bp and 141,000 bp). (A) LSDV/KZ-Konstanay/Kazakhstan/2018. (B) LSDV/Russia/Tyumen/2019. (C) LSDV/Russia/Udmurtya/2019. (D) LSDV/Russia/Saratov/2017. (E) LSDV/GD01/China/2020 with permission from A. Sprygin [78].
Figure 5.
Graphical representation of the position of the predicted recombination events across the five strains with the highest statistical significance. The events are indicated in black in the white backbone of the viral genome. Possible regions selected for by recombination are indicated by arrows (positions 80,000 bp, 124,000 bp and 141,000 bp). (A) LSDV/KZ-Konstanay/Kazakhstan/2018. (B) LSDV/Russia/Tyumen/2019. (C) LSDV/Russia/Udmurtya/2019. (D) LSDV/Russia/Saratov/2017. (E) LSDV/GD01/China/2020 with permission from A. Sprygin [78].

1.8. Next Generation Vaccines Being Developed for LSD including Modified LSDV, Inactivated LSDV and Subunit LSD Vaccines as well as mRNA Based Vaccines
When LSD first emerged in India, there was no commercial LSD vaccine and so the GTPV vaccine was used. Recently, a live-attenuated LSD vaccine, named Lumpi-ProVacInd, has been developed from a circulating LSDV isolate closely related to the Kenyan-type LSDV strain. The virus was attenuated by passaging fifty times in Vero cells. The vaccine protects from LSDV challenge and large clinical trials have taken place indicating that the vaccine is safe in cattle, buffalo and lactating pregnant animals. The vaccine is now being commercialised for use in Asia [81].
Targeted approaches to knock-out virulence genes have been used to create an attenuated LSDV for vaccine use. Kara et al. (2018) deleted two genes from a virulent LSDV in separate experiments to attempt to attenuate LSDV. These genes coded for homologues of an interleukin 10-like (LSDV005) and interferon- γ receptor-like genes (LSDV008), respectively. The resulting knockout LSDVs were still virulent and there were severe post-vaccinal reactions and febrile responses were observed for both constructs, with the interferon- γ receptor-like gene knock-out LSDV being more virulent than the interleukin 10-like knockout LSDV [82]. Chervyakova et al. (2022) knocked out four LSDV genes to attenuate the virulent LSDV Atyrau/KZ. These genes included the LSDV005 gene which encodes an interleukin-10-like protein; LSDV008, a homologue of the vaccinia virus B8R gene which encodes an interferon-γ receptor-like protein; LSDV142, a homologue of the N1L gene of the vaccinia virus, which encodes a protein of the Bcl-2-like protein family and inhibits both apoptosis and activation of the pro-inflammatory transcription factor NF-κB (nuclear factor kappa B); and LSDV066, a gene which encodes thymidine kinase. Bovine interleukin-18 mRNA was inserted into the thymidine kinase locus to knock this gene out, whereas the other three genes were deleted. The resulting candidate vaccine, LSDV Atyrau-5BJN (IL18), was safe and protected from virulent LSDV challenge [83]. In our laboratory, the superoxide dismutase gene (SOD) of the nLSDV vaccine was replaced with a synthetic, stabilised SOD gene to generate an nLSDV that potentially has further reduced virulence, as well as improved immunogenicity [84].
There is some hesitancy to introduce live attenuated LSDV vaccines into areas with no LSDV, not only for safety considerations, but also because of the impact on the “disease free status” of the country [85]. Therefore, there has been an effort to develop inactivated and subunit vaccines [86,87]. As a result, developments have been made to produce safer LSDV vaccines and to further improve their immunogenicity. Chemically inactivated nLSDV vaccines with adjuvants have been developed as safer alternatives to live nLSDV [85,86,88,89,90]. The combination of a low-molecular-weight copolymer adjuvant and inactivated LSDV, at titres similar to live vaccines, has given protection from LSDV challenge [85].
The state of New South Wales of Australia is working with the biotech company Tiba Biotech to develop LSDV mRNA-based vaccines [91]. This project is in the early stages of development. Since Australia is presently free of LSD, strategies based on other approaches than the live attenuated LSDV vaccine are desirable.
1.9. LSDV as a Vaccine Vector
Modern candidate recombinant vaccines are based on DNA or mRNA, subunit proteins, attenuated pathogens, inactivated pathogens or virus-vectored platforms. Live virus vectors encoding foreign antigens have many advantages compared to the other strategies: they can induce both humoral and cellular responses, as opposed to subunit protein-based vaccines that are often limited to eliciting antibody responses; by nature, they are highly immunogenic and therefore usually do not require adjuvants; cellular uptake is generally more effective due to active infection, compared to DNA vaccines that need to translocate to the host nucleus; and some may overcome logistical issues as they can be freeze-dried [91,92,93]. Poxviruses, particularly safe derivatives of vaccinia virus (VACV), have been used extensively as live vaccine vectors. Their genomes can tolerate very large insertions, enabling one virus to encode multiple antigens; VACV can hold the entire 27.3 kb genome of a human coronavirus [94]. Poxviruses complete their replication in the host cell’s cytoplasm, therefore there is less concern of host genomic integration of foreign genes [4].
Various methods exist to construct recombinant poxviruses. Modern, less frequently used methods include the generation of synthetic viruses by ligation of synthesised DNA fragments or the use of CRISPR/Cas9 [95,96]. Poxvirus genomes are not repaired well following Cas9 cleavage and so the use as an editing tool is not efficient. However, it works well as a selection agent to select recombinants and by speeding up isolation of recombinant poxviruses [97].
Traditionally, however, recombinant poxviruses are made via homologous recombination between a DNA plasmid encoding the foreign vaccine antigen(s) together with poxvirus promoters, and the parent poxvirus genome [98] (Figure 6).
Figure 6.
Schematic diagram of the generation of a recombinant poxvirus by homologous recombination of parental virus and a transfer vector. The plasmid encoding the transfer vector contains two antigen genes (purple arrows), an enhanced green fluorescent protein gene (egfp) as a marker gene (green arrow) as well as left and right flanks (49 and 50 flanks, respectively; blue arrows). Overall, 49, left flank of the transfer vector, ±300 bp of the 3′ end of 049 ORF of the LSDV genome; 50, right flank of the transfer vector, ±300 bp of the 3′ end of 050 ORF of the LSDV genome. All genes expressed by poxviruses are under the control of poxvirus promoters [99] (From student thesis where I am the superviso).
Figure 6.
Schematic diagram of the generation of a recombinant poxvirus by homologous recombination of parental virus and a transfer vector. The plasmid encoding the transfer vector contains two antigen genes (purple arrows), an enhanced green fluorescent protein gene (egfp) as a marker gene (green arrow) as well as left and right flanks (49 and 50 flanks, respectively; blue arrows). Overall, 49, left flank of the transfer vector, ±300 bp of the 3′ end of 049 ORF of the LSDV genome; 50, right flank of the transfer vector, ±300 bp of the 3′ end of 050 ORF of the LSDV genome. All genes expressed by poxviruses are under the control of poxvirus promoters [99] (From student thesis where I am the superviso).

An insertion site is chosen for the introduction of the foreign genes. The site should not be a mutational ‘hot-spot’ as this may cause the foreign genes to mutate or be deleted, and integration of the foreign genes should not disrupt sequences essential to LSDV. The LSDV thymidine kinase and ribonucleotide reductase genes have been used previously by the insertion of foreign sequences into these sites, resulting in disruption of these genes [100,101,102]. These sites are thought to further attenuate the vaccine and result in slower growth and so other insertion sites are more desirable. A putative non-essential gene encoding a G-protein-coupled chemokine receptor subfamily homologue has been tested as a site for homologous recombination [103]. The region between the convergent LSDV open reading frames (ORFs) 49 and 50 have also been used without gene disruption [20,104,105].
Inserted foreign genes require poxvirus promoters for the recognition of the poxvirus transcriptional machinery. Various promoters from other poxviruses have been shown to function in LSDV (Table 1) [24,106]. The timing of expression is also regulated by the promoter, such as whether it is expressed during the early stage of the poxvirus lifecycle, the late stage, or both. Choice of promoter may also affect the immunogenicity of the controlled antigen; for example, promoters that give early expression may enhance T-cell responses to the expressed antigen [107]. In addition, the stop codons of genes need to be followed by poxvirus transcription terminator sequences TTTTTAT, TTTTTCT or TTTTTGT [108]. Excluding the terminator can reduce transcription of the gene.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Summary of various poxvirus promoters that have been shown to be functional in LSDV.
| Promoter | Origin | Type | Sequence (5′ to 3′) | References |
|---|---|---|---|---|
| mH5 | VACV, modified | Early-late | AAAAATTGAAAATAAATACAAAGGTTCTTGAGGGTTGTGTTAAATTGAAAGCGAGAAATAATCATAAATAA | [109] |
| pLEO | VACV, modified | Early-late | TTTTATTTTTTTTTTTTGGAATATAAATATCCGGTAAAATTGAAAAAATATACACTAATTAGCGTCTCGTTTCAGACGCTAG | [110] |
| p7.5 | VACV, modified | Early-late | CCAAACCCACCCGCTTTTTATAGTAAGTTTTTCACCCATAAATAATAAATACAATAATTAATTTCTCGTAAAAGTAGAAAATATATTCTAATTTATTGCACGG | [111] |
| p11 | VACV | Late | TTTCATTTTGTTTTTTTCTATGCTATAA | [112] |
| pSS | Synthetic | Early-late | AAAATTGAAATTTTATTTTTTTTTTTTGGAATATAAATA | [113] |
| mFP | Fowlpox, modified | Early-late | AGAAAAATATCCTAAAATTGAATTGTAATTATCGATAATAA | [20] |
Reporter genes such as E. coli β-galactosidase (LacZ) or fluorescent markers such as GFP can be included to visualise cells infected with the recombinant LSDV [101,105]. Selection markers can also be used to enable the recombinant to grow in cell lines or conditions that the parent virus cannot. Mycophenolic acid tolerance induced by expression of the gpt gene of E. coli (ECOGPT) has been used in rLSDV-infected cells as a method of selection [9]. This method enriched for recombinants but did not exclude all parent virus. The VACV K1L gene, which rescues modified vaccinia Ankara (MVA) growth in rabbit kidney 13 (RK-13) cells, can also rescue LSDV growth in RK-13 cells, although growth tends to be slow [24,114].
LSDV-vectored cattle vaccines would be a major advantage, as these would provide dual or multivalent vaccines [115]. There are numerous recombinant Capripoxvirus vaccines that have been constructed (Table 2) and tested for immunogenicity as well as, in some cases, in challenge experiments. KS-1 recombinants have been successful in protecting against rinderpest virus [116], rift valley fever virus [117], Peste-des-petits-ruminants virus [118] and have provided partial protection from blue tongue virus challenge [119]. These experiments were performed in sheep and in many cases the recombinant KS vaccines also protected from SPPV challenge. KS-1 was originally thought to be of SPPV origin, but sequencing revealed that it was actually an LSDV derived vaccine which retained some virulence in cattle [120]. LSDV expressing rabies glycoprotein protects from rabies virus challenge in rodent models [6,121], as well as inducing protective levels of rabies virus antibodies in cattle [101]. LSDV expressing bovine ephemeral fever virus (BEFV) proteins induced good BEFV and LSDV neutralising antibody responses in rabbits and cattle. The cattle were protected from LSDV challenge [104]. Despite the success of recombinant LSDV vaccines, there are still none that have been commercialized. This is probably due to the poor profit margins in the affected communities where the diseases are prevalent; however, this may change with the expansion of LSD across Asia.
Table 2.
Experimental vaccines using Capripoxvirus as a live virus vector.
| Target Pathogen | Antigen Expressed | Capripoxvirus Backbone | Reference |
|---|---|---|---|
| Rindepest Virus | Rindepest Virus H and F gene | KS-1 * | [122] |
| Peste-des-petits-ruminants virus (PPRV) | hemagglutinin (H) or the fusion (F) protein gene of PPRV | KS-1 * | [118] |
| Blue Tongue Virus (BTV) | BTV VP7 | KS-1 * | [119] |
| Rift Valley Fever Virus (RVFV) | RVFV Gn and Gc glycoproteins | KS-1 * | [117] |
| HIV | Grttn polyprotein | nLSDV | [106,123] |
| Env and Gag | nLSDV with SODis | [105] | |
| Rabies virus (RABV) | RABV Glycoprotein | nLSDV | [6] |
| Mokola virus (MOKV), West Caucasian bat virus (WCBV), Rabies Virus (RABV) | RABV, MOKV and WCBV glycoprotein genes | nLSDV | [121] |
| Rift valley fever virus (RVFV) | RVFV Gn and Gc glycoproteins | nLSDV | [102,124,125] |
| Bovine ephemeral fever virus (BEFV) | BEFV Glycoprotein | nLSDV | [102,124,125] |
| Glycoprotein | nLSDV with either SODis or ΔSOD | [104] | |
| Glycoprotein and Matrix |
nLSDV: Neethling strain of LSDV; SODis: stabilised superoxide dismutase gene; ΔSOD: deleted superoxide dismutase gene. * KS-1 was originally thought to be of SPPV origin, but sequencing revealed that it was actually an LSDV derived vaccine [120].
LSDV is a host restricted virus and does not complete the replication cycle in non-ruminant hosts. LSDV expressing HIV proteins has been shown to be immunogenic in rhesus macaques and boost the responses primed by a MVA vectored vaccine. The combination regimen induced high-magnitude, broad and balanced CD4(+) and CD8(+) T-cell responses, and transient activation of the immune response. These studies support further development of LSDV as a vaccine vector [123] in non-ruminant hosts. LSDV expressing HIV Gag and Env and tested in combination with MVA induced good antibody responses in rabbits [105]. LSDV was injected into mice and the response after 24 h was compared with that of five host-restricted poxvirus species from three genera, namely Canarypox virus (CNPV), Fowlpox virus (FWPV), MVA and two novel South African avipoxviruses. These six viruses produced qualitatively and quantitatively distinct host responses with LSDV, followed by MVA, inducing the greatest interferon (IFN) response [126]. This illustrates that poxvirus vectors differ and may have different strengths as vaccine vectors.
2. Conclusions
Lumpy skin disease, an important notifiable disease of cattle, was originally an African disease and is now becoming established in large parts of the Middle East and Asia. There are available vaccines and the most effective are those based on attenuated LSDV. Vaccination remains the best way to control outbreaks. Given the spread of LSDV by biting insects, it is important to vaccinate to protect against LSD as well as control the insect population around infected animals. Next generation LSD vaccines are being developed which include further attenuation of LSDV and mRNA-based vaccines. In addition, LSDV is being investigated as a vaccine vector with the possibility of multivalent vaccines being developed.
3. Patents
The authors have filed patent applications on various nLSDV-based vaccines: CT/IB2019/054090, PA166012PCT; PCT/IB2022/056970, PA175637/PCT; PA179963/P, PA179694/P, PA176615/P.
Author Contributions
The review was drafted by L.W. and A.-L.W. with input from R.C. All authors have read and agreed to the published version of the manuscript.
Funding
This work is based on the research supported in part by the National Research Foundation of South Africa (Grant numbers 64815 and 138792) and the Poliomyelitis Research Foundation (Grant number 19/85).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The UCT group is discussing commercializing some of the LSDV recombinants made at the University of Cape Town. The funders and possible commercial partners had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Kitching, R.; Smale, C. Comparison of the external dimensions of capripoxvirus isolates. Res. Vet. Sci. 1986, 41, 425–427. [Google Scholar] [CrossRef] [PubMed]
- Tulman, E.R.; Afonso, C.L.; Lu, Z.; Zsak, L.; Sur, J.-H.; Sandybaev, N.T.; Kerembekova, U.Z.; Zaitsev, V.L.; Kutish, G.F.; Rock, D.L. The Genomes of Sheeppox and Goatpox Viruses. J. Virol. 2002, 76, 6054–6061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tulman, E.R.; Afonso, C.L.; Lu, Z.; Zsak, L.; Kutish, G.F.; Rock, D.L. Genome of Lumpy Skin Disease Virus. J. Virol. 2001, 75, 7122–7130. [Google Scholar] [CrossRef] [Green Version]
- McFadden, G. Poxvirus tropism. Nat. Rev. Genet. 2005, 3, 201–213. [Google Scholar] [CrossRef]
- Prozesky, L.; Barnard, B.J. A study of the pathology of lumpy skin disease in cattle. Onderstepoort J. Veter.-Res. 1982, 49, 167–175. [Google Scholar]
- Aspden, K.; Passmore, J.-A.; Tiedt, F.; Williamson, A.-L. Evaluation of lumpy skin disease virus, a capripoxvirus, as a replication-deficient vaccine vector. J. Gen. Virol. 2003, 84, 1985–1996. [Google Scholar] [CrossRef]
- Abutarbush, S.M.; Ababneh, M.M.; Al Zoubi, I.G.; Al Sheyab, O.M.; Alekish, M.O.; Al Gharabat, R.J. Lumpy Skin Disease in Jordan: Disease Emergence, Clinical Signs, Complications and Preliminary-associated Economic Losses. Transbound. Emerg. Dis. 2013, 62, 549–554. [Google Scholar] [CrossRef]
- Tuppurainen, E.; Dietze, K.; Wolff, J.; Bergmann, H.; Beltran-Alcrudo, D.; Fahrion, A.; Lamien, C.E.; Busch, F.; Sauter-Louis, C.; Conraths, F.J.; et al. Review: Vaccines and Vaccination against Lumpy Skin Disease. Vaccines 2021, 9, 1136. [Google Scholar] [CrossRef]
- Aspden, K.A. Study of the Host Restricted Lumpy Skin Disease Virus as a Vaccine Vector Using Rabies Virus as a Model; University of Cape Town: Cape Town, South Africa, 2002. [Google Scholar]
- Coetzer, J.A.W.; Tuppurainen, E. Lumpy skin disease. Infect. Dis. Livest. 2004, 2, 1268–1276. [Google Scholar]
- Möller, J.; Moritz, T.; Schlottau, K.; Krstevski, K.; Hoffmann, D.; Beer, M.; Hoffmann, B. Experimental lumpy skin disease virus infection of cattle: Comparison of a field strain and a vaccine strain. Arch. Virol. 2019, 164, 2931–2941. [Google Scholar] [CrossRef]
- Ahmed, E.M.; Eltarabilli, M.M.; Shahein, M.A.; Fawzy, M. Lumpy skin disease outbreaks investigation in Egyptian cattle and buffaloes: Serological evidence and molecular characterization of genome termini. Comp. Immunol. Microbiol. Infect. Dis. 2021, 76, 101639. [Google Scholar] [CrossRef] [PubMed]
- Fagbo, S.; Coetzer, J.A.; Venter, E.H. Seroprevalence of Rift Valley fever and lumpy skin disease in African buffalo (Syncerus caffer) in the Kruger National Park and Hluhluwe-iMfolozi Park, South Africa. J. S. Afr. Veter-Assoc. 2014, 85, 1075. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Kock, G. Lumpy skin disease (knopvelsiekte) of cattle in Southern Africa. J. Am. Vet. Med. Assoc. 1948, 112, 57. [Google Scholar]
- Hedger, R.; Hamblin, C. Neutralising antibodies to lumpy skin disease virus in African wildlife. Comp. Immunol. Microbiol. Infect. Dis. 1983, 6, 209–213. [Google Scholar] [CrossRef] [PubMed]
- Young, E.; Basson, P.A.; Weiss, K.E. Experimental infection of game animals with lumpy skin disease virus (prototype strain Neethling). Onderstepoort J. Vet.-Res. 1970, 37, 79–87. [Google Scholar]
- Dao, T.D.; Tran, L.H.; Nguyen, H.D.; Hoang, T.T.; Nguyen, G.H.; Tran, K.V.D.; Nguyen, H.X.; Van Dong, H.; Bui, A.N.; Bui, V.N. Characterization of Lumpy skin disease virus isolated from a giraffe in Vietnam. Transbound. Emerg. Dis. 2022, 69, e3268–e3272. [Google Scholar] [CrossRef]
- Rhazi, H.; Safini, N.; Mikou, K.; Alhyane, M.; Lenk, M.; Tadlaoui, K.O.; Elharrak, M. Comparative Sensitivity Study of Primary Cells, Vero, OA3.Tsand ESH-L cell lines to Lumpy Skin Disease, Sheeppox, and Goatpox viruses Detection and Growth. J. Virol. Methods 2021, 293, 114164. [Google Scholar] [CrossRef]
- Binepal, Y.S.; Ongadi, F.A.; Chepkwony, J.C. Alternative cell lines for the propagation of lumpy skin disease virus. Onderstepoort J. Veter.-Res. 2001, 68, 151–153. [Google Scholar]
- Omar, R. Comparison of the Two Lumpy Skin Disease Virus Vaccines, Neethling and Herbivac, and Construction of a Recombinant Herbivac-Rift Valley Fever Virus Vaccine; University of Cape Town: Sydney, Australia, 2015. [Google Scholar]
- Fay, P.; Cook, C.; Wijesiriwardana, N.; Tore, G.; Comtet, L.; Carpentier, A.; Shih, B.; Freimanis, G.; Haga, I.R.; Beard, P.M. Madin-Darby bovine kidney (MDBK) cells are a suitable cell line for the propagation and study of the bovine poxvirus lumpy skin disease virus. J. Virol. Methods 2020, 285, 113943. [Google Scholar] [CrossRef]
- Munyanduki, H.; Omar, R.; Douglass, N.; Williamson, A.-L. Removal of bovine viral diarrhea virus (BVDV) from lumpy skin disease virus (LSDV) vaccine stocks by passage on chorioallantoic membranes of fertilized hens’ eggs. J. Virol. Methods 2019, 275, 113752. [Google Scholar] [CrossRef]
- Weiss, K.E. Lumpy Skin Disease Virus. In Cytomegaloviruses. Rinderpest Virus. Lumpy Skin Disease Virus; Springer: Berlin/Heidelberg, Germany, 1968; pp. 111–131. [Google Scholar]
- van Diepen, M.; Chapman, R.; Douglass, N.; Whittle, L.; Chineka, N.; Galant, S.; Cotchobos, C.; Suzuki, A.; Williamson, A.-L. Advancements in the Growth and Construction of Recombinant Lumpy Skin Disease Virus (LSDV) for Use as a Vaccine Vector. Vaccines 2021, 9, 1131. [Google Scholar] [CrossRef] [PubMed]
- Babiuk, S.; Parkyn, G.; Copps, J.; Larence, J.E.; Sabara, M.I.; Bowden, T.R.; Boyle, D.B.; Kitching, R.P. Evaluation of an Ovine Testis Cell Line (OA3.Ts) for Propagation of Capripoxvirus Isolates and Development of an Immunostaining Technique for Viral Plaque Visualization. J. Vet. Diagn. Investig. 2007, 19, 486–491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeruham, I.; Nir, O.; Braverman, Y.; Davidson, M.; Grinstein, H.; Haymovitch, M.; Zamir, O. Spread of lumpy skin disease in Israeli dairy herds. Vet. Rec. 1995, 137, 91–93. [Google Scholar] [CrossRef] [PubMed]
- Vandenbussche, F.; Mathijs, E.; Philips, W.; Saduakassova, M.; De Leeuw, I.; Sultanov, A.; Haegeman, A.; De Clercq, K. Recombinant LSDV Strains in Asia: Vaccine Spillover or Natural Emergence? Viruses 2022, 14, 1429. [Google Scholar] [CrossRef]
- Manić, M.; Stojiljković, M.; Petrović, M.; Nišavić, J.; Bacić, D.; Petrović, T.; Vidanović, D.; Obrenović, S. Epizootic features and control measures for lumpy skin disease in south-east Serbia in 2016. Transbound. Emerg. Dis. 2019, 66, 2087–2099. [Google Scholar] [CrossRef] [PubMed]
- Lu, G.; Xie, J.; Luo, J.; Shao, R.; Jia, K.; Li, S. Lumpy skin disease outbreaks in China, since 3 August 2019. Transbound. Emerg. Dis. 2020, 68, 216–219. [Google Scholar] [CrossRef]
- Gupta, T.; Patial, V.; Bali, D.; Angaria, S.; Sharma, M.; Chahota, R. A review: Lumpy skin disease and its emergence in India. Vet. Res. Commun. 2020, 44, 111–118. [Google Scholar] [CrossRef]
- Kayesh, M.E.H.; Hussan, M.T.; Hashem, A.; Eliyas, M.; Anower, A.M. Lumpy Skin Disease Virus Infection: An Emerging Threat to Cattle Health in Bangladesh. Hosts Viruses 2020, 7, 97–108. [Google Scholar] [CrossRef]
- Sprygin, A.; Artyuchova, E.; Babin, Y.; Prutnikov, P.; Kostrova, E.; Byadovskaya, O.; Kononov, A. Epidemiological characterization of lumpy skin disease outbreaks in Russia in 2016. Transbound. Emerg. Dis. 2018, 65, 1514–1521. [Google Scholar] [CrossRef]
- Agianniotaki, E.I.; Mathijs, E.; Vandenbussche, F.; Tasioudi, K.E.; Haegeman, A.; Iliadou, P.; Chaintoutis, S.C.; Dovas, C.I.; Van Borm, S.; Chondrokouki, E.D.; et al. Complete Genome Sequence of the Lumpy Skin Disease Virus Isolated from the First Reported Case in Greece in 2015. Genome Announc. 2017, 5, e00550-17. [Google Scholar] [CrossRef] [Green Version]
- OIE-WAHIS. Events Management. Available online: https://wahis.woah.org/#/event-management (accessed on 1 December 2022).
- Klement, E.; Broglia, A.; Antoniou, S.E.; Tsiamadis, V.; Plevraki, E.; Petrovic, T.; Polacek, V.; Debeljak, Z.; Miteva, A.; Alexandrov, T.; et al. Neethling vaccine proved highly effective in controlling lumpy skin disease epidemics in the Balkans. Prev. Vet. Med. 2020, 181, 104595. [Google Scholar] [CrossRef] [PubMed]
- Kononov, A.; Prutnikov, P.; Shumilova, I.; Kononova, S.; Nesterov, A.; Byadovskaya, O.; Pestova, Y.; Diev, V.; Sprygin, A. Determination of lumpy skin disease virus in bovine meat and offal products following experimental infection. Transbound. Emerg. Dis. 2019, 66, 1332–1340. [Google Scholar] [CrossRef] [PubMed]
- Khalafalla, A. Lumpy Skin Disease: An Economically Significant Emerging Disease. Available online: https://www.intechopen.com/online-first/85014 (accessed on 10 January 2023).
- Tuppurainen, E.S.M.; Stoltsz, W.H.; Troskie, M.; Wallace, D.B.; Oura, C.A.L.; Mellor, P.S.; Coetzer, J.A.W.; Venter, E. A Potential Role for Ixodid (Hard) Tick Vectors in the Transmission of Lumpy Skin Disease Virus in Cattle. Transbound. Emerg. Dis. 2011, 58, 93–104. [Google Scholar] [CrossRef] [Green Version]
- Sohier, C.; Haegeman, A.; Mostin, L.; De Leeuw, I.; Van Campe, W.; De Vleeschauwer, A.; Tuppurainen, E.S.M.; van den Berg, T.; De Regge, N.; De Clercq, K. Experimental evidence of mechanical Lumpy Skin Disease virus transmission by Stomoxys calcitrans biting flies and Haematopota spp. horseflies. Sci. Rep. 2019, 9, 20076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paslaru, A.I.; Maurer, L.M.; Vögtlin, A.; Hoffmann, B.; Torgerson, P.R.; Mathis, A.; Veronesi, E. Putative roles of mosquitoes (Culicidae) and biting midges (Culicoides spp.) as mechanical or biological vectors of lumpy skin disease virus. Med. Vet. Entomol. 2022, 36, 381–389. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhao, L.; Yang, J.; Shi, M.; Nie, F.; Liu, S.; Wang, Z.; Huang, D.; Wu, H.; Li, D.; et al. Analysis of vaccine-like lumpy skin disease virus from flies near the western border of China. Transbound. Emerg. Dis. 2021, 69, 1813–1823. [Google Scholar] [CrossRef]
- Paslaru, A.I.; Verhulst, N.O.; Maurer, L.M.; Brendle, A.; Pauli, N.; Vögtlin, A.; Renzullo, S.; Ruedin, Y.; Hoffmann, B.; Torgerson, P.R.; et al. Potential mechanical transmission of Lumpy skin disease virus (LSDV) by the stable fly (Stomoxys calcitrans) through regurgitation and defecation. Curr. Res. Insect Sci. 2020, 1, 100007. [Google Scholar] [CrossRef]
- Lubinga, J.C.; Tuppurainen, E.S.M.; Mahlare, R.; Coetzer, J.A.W.; Stoltsz, W.H.; Venter, E.H. Evidence of Transstadial and Mechanical Transmission of Lumpy Skin Disease Virus by Amblyomma hebraeum Ticks. Transbound. Emerg. Dis. 2013, 62, 174–182. [Google Scholar] [CrossRef]
- Kitching, R.P.; Hammond, J.M.; Black, D.N. Studies on the Major Common Precipitating Antigen of Capripoxvirus. J. Gen. Virol. 1986, 67, 139–148. [Google Scholar] [CrossRef]
- Saegerman, C.; Bertagnoli, S.; Meyer, G.; Ganière, J.-P.; Caufour, P.; De Clercq, K.; Jacquiet, P.; Fournié, G.; Hautefeuille, C.; Etore, F.; et al. Risk of introduction of lumpy skin disease in France by the import of vectors in animal trucks. PLoS ONE 2018, 13, e0198506. [Google Scholar] [CrossRef] [Green Version]
- Namazi, F.; Tafti, A.K. Lumpy skin disease, an emerging transboundary viral disease: A review. Vet. Med. Sci. 2021, 7, 888–896. [Google Scholar] [CrossRef] [PubMed]
- Carn, V.M.; Kitching, R.P. An investigation of possible routes of transmission of lumpy skin disease virus (Neethling). Epidemiology Infect. 1995, 114, 219–226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aleksandr, K.; Olga, B.; David, W.B.; Pavel, P.; Yana, P.; Svetlana, K.; Alexander, N.; Vladimir, R.; Dmitriy, L.; Alexander, S. Non-vector-borne transmission of lumpy skin disease virus. Sci. Rep. 2020, 10, 7436. [Google Scholar] [CrossRef]
- Shumilova, I.; Krotova, A.; Nesterov, A.; Byadovskaya, O.; van Schalkwyk, A.; Sprygin, A. Overwintering of recombinant lumpy skin disease virus in northern latitudes, Russia. Transbound. Emerg. Dis. 2022, 69, e3239–e3243. [Google Scholar] [CrossRef] [PubMed]
- Osuagwuh, U.I.; Bagla, V.H.; Venter, E.H.; Annandale, C.H.; Irons, P. Absence of lumpy skin disease virus in semen of vaccinated bulls following vaccination and subsequent experimental infection. Vaccine 2007, 25, 2238–2243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Annandale, C.H.; Holm, D.E.; Ebersohn, K.; Venter, E.H. Seminal Transmission of Lumpy Skin Disease Virus in Heifers. Transbound. Emerg. Dis. 2014, 61, 443–448. [Google Scholar] [CrossRef] [Green Version]
- Şevik, M.; Doğan, M. Epidemiological and Molecular Studies on Lumpy Skin Disease Outbreaks in Turkey during 2014-2015. Transbound. Emerg. Dis. 2017, 64, 1268–1279. [Google Scholar] [CrossRef]
- Haegeman, A.; De Leeuw, I.; Mostin, L.; Van Campe, W.; Aerts, L.; Venter, E.; Tuppurainen, E.; Saegerman, C.; De Clercq, K. Comparative Evaluation of Lumpy Skin Disease Virus-Based Live Attenuated Vaccines. Vaccines 2021, 9, 473. [Google Scholar] [CrossRef]
- Van Rooyen, P.J.; Munz, E.K.; Weiss, K.E. The optimal conditions for the multiplication of Neethling-type lumpy skin disease virus in embryonated eggs. Onderstepoort J. Vet. Res. 1969, 36, 165–174. [Google Scholar]
- Mathijs, E.; Vandenbussche, F.; Haegeman, A.; King, A.; Nthangeni, B.; Potgieter, C.; Maartens, L.; Van Borm, S.; De Clercq, K. Complete Genome Sequences of the Neethling-Like Lumpy Skin Disease Virus Strains Obtained Directly from Three Commercial Live Attenuated Vaccines. Genome Announc. 2016, 4, e01255-16. [Google Scholar] [CrossRef] [Green Version]
- Calistri, P.; De Clercq, K.; Gubbins, S.; Klement, E.; Stegeman, A.; Abrahantes, J.C.; Marojevic, D.; Antoniou, S.; Broglia, A. Lumpy skin disease epidemiological report IV: Data collection and analysis. EFSA J. 2020, 18, e06010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Norian, R.; Ahangran, N.A.; Varshovi, H.R.; Azadmehr, A. Comparative efficacy of two heterologous capripox vaccines to control lumpy skin disease in cattle. Bulg. J. Vet. Med. 2019, 22, 171–179. [Google Scholar] [CrossRef]
- Gari, G.; Abie, G.; Gizaw, D.; Wubete, A.; Kidane, M.; Asgedom, H.; Bayissa, B.; Ayelet, G.; Oura, C.A.; Roger, F.; et al. Evaluation of the safety, immunogenicity and efficacy of three capripoxvirus vaccine strains against lumpy skin disease virus. Vaccine 2015, 33, 3256–3261. [Google Scholar] [CrossRef] [PubMed]
- Zhugunissov, K.; Bulatov, Y.; Orynbayev, M.; Kutumbetov, L.; Abduraimov, Y.; Shayakhmetov, Y.; Taranov, D.; Amanova, Z.; Mambetaliyev, M.; Absatova, Z.; et al. Goatpox virus (G20-LKV) vaccine strain elicits a protective response in cattle against lumpy skin disease at challenge with lumpy skin disease virulent field strain in a comparative study. Vet Microbiol. 2020, 245, 108695. [Google Scholar] [CrossRef] [PubMed]
- Hamdi, J.; Bamouh, Z.; Jazouli, M.; Boumart, Z.; Tadlaoui, K.O.; Fihri, O.F.; EL Harrak, M. Experimental evaluation of the cross-protection between Sheeppox and bovine Lumpy skin vaccines. Sci. Rep. 2020, 10, 8888. [Google Scholar] [CrossRef]
- Manual, O. Chapter 3.4.12 Lumpy Skin Disease. Available online: https://www.woah.org/fileadmin/Home/fr/Health_standards/tahm/3.04.12_LSD.pdf (accessed on 10 January 2023).
- Morgenstern, M.; Klement, E. The Effect of Vaccination with Live Attenuated Neethling Lumpy Skin Disease Vaccine on Milk Production and Mortality—An Analysis of 77 Dairy Farms in Israel. Vaccines 2020, 8, 324. [Google Scholar] [CrossRef]
- Abutarbush, S.; Hananeh, W.M.; Ramadan, W.; Al Sheyab, O.M.; Alnajjar, A.R.; Al Zoubi, I.G.; Knowles, N.J.; Tuppurainen, E.S.M.; Bachanek-Bankowska, K. Adverse Reactions to Field Vaccination Against Lumpy Skin Disease in Jordan. Transbound. Emerg. Dis. 2014, 63, e213–e219. [Google Scholar] [CrossRef]
- Ben-Gera, J.; Klement, E.; Khinich, E.; Stram, Y.; Shpigel, N.Y. Comparison of the efficacy of Neethling lumpy skin disease virus and x10RM65 sheep-pox live attenuated vaccines for the prevention of lumpy skin disease—The results of a randomized controlled field study. Vaccine 2015, 33, 4837–4842. [Google Scholar] [CrossRef]
- Tuppurainen, E.; Babiuk, S.; Klement, E. Lumpy Skin Disease; Springer: Berlin/Heidelberg, Germany, 2018. [Google Scholar]
- Kara, P.D.; Afonso, C.L.; Wallace, D.B.; Kutish, G.F.; Abolnik, C.; Lu, Z.; Vreede, F.T.; Taljaard, L.C.F.; Zsak, A.; Viljoen, G.J.; et al. Comparative sequence analysis of the South African vaccine strain and two virulent field isolates of Lumpy skin disease virus. Arch. Virol. 2003, 148, 1335–1356. [Google Scholar] [CrossRef]
- van Schalkwyk, A.; Kara, P.; Heath, L. Phylogenomic characterization of historic lumpy skin disease virus isolates from South Africa. Arch. Virol. 2022, 167, 2063–2070. [Google Scholar] [CrossRef]
- Desingu, P.A.; Rubeni, T.P.; Sundaresan, N.R. Evolution of monkeypox virus from 2017 to 2022: In the light of point mutations. Front. Microbiol. 2022, 13, 1037598. [Google Scholar] [CrossRef] [PubMed]
- Brennan, G.; Stoian, A.M.M.; Yu, H.; Rahman, M.J.; Banerjee, S.; Stroup, J.N.; Park, C.; Tazi, L.; Rothenburg, S. Molecular Mechanisms of Poxvirus Evolution. mBio 2022, e01526-22. [Google Scholar] [CrossRef] [PubMed]
- Bedson, H.S.; Dumbell, K.R. Hybrids derived from the viruses of alastrim and rabbit pox. Epidemiology Infect. 1964, 62, 141–146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dumbell, K.R.; Bedson, H.S. The use of ceiling temperature and reactivation in the isolation of pox virus hybrids. Epidemiology Infect. 1964, 62, 133–140. [Google Scholar] [CrossRef] [Green Version]
- Bedson, H.S.; Dumbell, K.R. Hybrids derived from the viruses of variola major and cowpox. Epidemiology Infect. 1964, 62, 147–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakano, E.; Panicali, D.; Paoletti, E. Molecular genetics of vaccinia virus: Demonstration of marker rescue. Proc. Natl. Acad. Sci. USA 1982, 79, 1593–1596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paoletti, E.; Weinberg, R.L.; Davis, S.W.; Davis, M. Genetically engineered poxviruses: A novel approach to the construction of live vaccines. Vaccine 1984, 2, 204–208. [Google Scholar] [CrossRef]
- Herbert, M.H.; Squire, C.J.; Mercer, A.A. Poxviral Ankyrin Proteins. Viruses 2015, 7, 709–738. [Google Scholar] [CrossRef]
- Erlandson, K.J.; Cotter, C.A.; Charity, J.C.; Martens, C.; Fischer, E.R.; Ricklefs, S.M.; Porcella, S.F.; Moss, B. Duplication of the A17L Locus of Vaccinia Virus Provides an Alternate Route to Rifampin Resistance. J. Virol. 2014, 88, 11576–11585. [Google Scholar] [CrossRef] [Green Version]
- Sprygin, A.; Mazloum, A.; van Schalkwyk, A.; Babiuk, S. Capripoxviruses, leporipoxviruses, and orthopoxviruses: Occurrences of recombination. Front. Microbiol. 2022, 13, 978829. [Google Scholar] [CrossRef]
- Krotova, A.; Byadovskaya, O.; Shumilova, I.; van Schalkwyk, A.; Sprygin, A. An in-depth bioinformatic analysis of the novel recombinant lumpy skin disease virus strains: From unique patterns to established lineage. BMC Genom. 2022, 23, 396. [Google Scholar] [CrossRef] [PubMed]
- Sprygin, A.; Babin, Y.; Pestova, Y.; Kononova, S.; Wallace, D.B.; Van Schalkwyk, A.; Byadovskaya, O.; Diev, V.; Lozovoy, D.; Kononov, A. Analysis and insights into recombination signals in lumpy skin disease virus recovered in the field. PLoS ONE 2018, 13, e0207480. [Google Scholar] [CrossRef]
- Kononov, A.; Byadovskaya, O.; Kononova, S.; Yashin, R.; Zinyakov, N.; Mischenko, V.; Perevozchikova, N.; Sprygin, A. Detection of vaccine-like strains of lumpy skin disease virus in outbreaks in Russia in 2017. Arch. Virol. 2019, 164, 1575–1585. [Google Scholar] [CrossRef] [PubMed]
- Kumar, N.; Tripathi, B.N. A serious skin virus epidemic sweeping through the Indian subcontinent is a threat to the livelihood of farmers. Virulence 2022, 13, 1943–1944. [Google Scholar] [CrossRef] [PubMed]
- Kara, P.D.; Mather, A.S.; Pretorius, A.; Chetty, T.; Babiuk, S.; Wallace, D.B. Characterisation of putative immunomodulatory gene knockouts of lumpy skin disease virus in cattle towards an improved vaccine. Vaccine 2018, 36, 4708–4715. [Google Scholar] [CrossRef] [PubMed]
- Chervyakova, O.; Issabek, A.; Sultankulova, K.; Bopi, A.; Kozhabergenov, N.; Omarova, Z.; Tulendibayev, A.; Aubakir, N.; Orynbayev, M. Lumpy Skin Disease Virus with Four Knocked Out Genes Was Attenuated In Vivo and Protects Cattle from Infection. Vaccines 2022, 10, 1705. [Google Scholar] [CrossRef] [PubMed]
- Douglass, N.; Munyanduki, H.; Omar, R.; Gers, S.; Mutowembwa, P.; Heath, L.; Williamson, A.-L. Influence of the Viral Superoxide Dismutase (SOD) Homologue on Lumpy Skin Disease Virus (LSDV) Growth, Histopathology and Pathogenicity. Vaccines 2020, 8, 664. [Google Scholar] [CrossRef]
- Wolff, J.; Beer, M.; Hoffmann, B. High Efficiency of Low Dose Preparations of an Inactivated Lumpy Skin Disease Virus Vaccine Candidate. Vaccines 2022, 10, 1029. [Google Scholar] [CrossRef]
- Wolff, J.; Moritz, T.; Schlottau, K.; Hoffmann, D.; Beer, M.; Hoffmann, B. Development of a Safe and Highly Efficient Inactivated Vaccine Candidate against Lumpy Skin Disease Virus. Vaccines 2020, 9, 4. [Google Scholar] [CrossRef]
- Kar, P.P.; Araveti, P.B.; Kuriakose, A.; Srivastava, A. Design of a multi-epitope protein as a subunit vaccine against lumpy skin disease using an immunoinformatics approach. Sci. Rep. 2022, 12, 19411. [Google Scholar] [CrossRef]
- Es-Sadeqy, Y.; Bamouh, Z.; Ennahli, A.; Safini, N.; El Mejdoub, S.; Tadlaoui, K.O.; Gavrilov, B.; El Harrak, M. Development of an inactivated combined vaccine for protection of cattle against lumpy skin disease and bluetongue viruses. Vet. Microbiol. 2021, 256, 109046. [Google Scholar] [CrossRef] [PubMed]
- Matsiela, M.S.; Naicker, L.; Dibakwane, V.S.; Ntombela, N.; Khoza, T.; Mokoena, N. Improved safety profile of inactivated Neethling strain of the Lumpy Skin Disease Vaccine. Vaccine X 2022, 12, 100209. [Google Scholar] [CrossRef]
- Malone, R.W. mRNA Vaccines in Livestock and Companion Animals Are Here Now. Available online: https://rwmalonemd.substack.com/p/mrna-vaccines-in-livestock-and-companion (accessed on 10 January 2023).
- Alcock, R.; Cottingham, M.G.; Rollier, C.S.; Furze, J.; De Costa, S.D.; Hanlon, M.; Spencer, A.J.; Honeycutt, J.D.; Wyllie, D.H.; Gilbert, S.C.; et al. Long-Term Thermostabilization of Live Poxviral and Adenoviral Vaccine Vectors at Supraphysiological Temperatures in Carbohydrate Glass. Sci. Transl. Med. 2010, 2, 19ra12. [Google Scholar] [CrossRef] [PubMed]
- Suschak, J.; Williams, J.A.; Schmaljohn, C.S. Advancements in DNA vaccine vectors, non-mechanical delivery methods, and molecular adjuvants to increase immunogenicity. Hum. Vaccines Immunother. 2017, 13, 2837–2848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rollier, C.S.; Reyes-Sandoval, A.; Cottingham, M.G.; Ewer, K.; Hill, A.V. Viral vectors as vaccine platforms: Deployment in sight. Curr. Opin. Immunol. 2011, 23, 377–382. [Google Scholar] [CrossRef]
- Thiel, V.; Herold, J.; Schelle, B.; Siddell, S.G. Infectious RNA transcribed in vitro from a cDNA copy of the human coronavirus genome cloned in vaccinia virus. J. Gen. Virol. 2001, 82, 1273–1281. [Google Scholar] [CrossRef]
- Chiuppesi, F.; Salazar, M.D.; Contreras, H.; Nguyen, V.H.; Martinez, J.; Park, Y.; Nguyen, J.; Kha, M.; Iniguez, A.; Zhou, Q.; et al. Development of a multi-antigenic SARS-CoV-2 vaccine candidate using a synthetic poxvirus platform. Nat. Commun. 2020, 11, 6121. [Google Scholar] [CrossRef]
- Laudermilch, E.; Chandran, K. MAVERICC: Marker-free Vaccinia Virus Engineering of Recombinants through in vitro CRISPR/Cas9 Cleavage. J. Mol. Biol. 2021, 433, 166896. [Google Scholar] [CrossRef]
- Gowripalan, A.; Smith, S.; Tscharke, D. Selection of Vaccinia Virus Recombinants Using CRISPR/Cas9. Bio-Protocol 2021, 11, e4270. [Google Scholar] [CrossRef]
- Wyatt, L.S.; Earl, P.L.; Moss, B. Generation of Recombinant Vaccinia Viruses. Curr. Protoc. Microbiol. 2015, 39, 14A.4.1–14A.4.18. [Google Scholar] [CrossRef]
- Suzuki, A. Development and Characterisation of Recombinant LSDV-Vectored Dual Vaccines Against Bovine Leukaemia Virus and Lumpy Skin Disease Virus; University of Cape Town: Cape Town, South Africa, 2019. [Google Scholar]
- Wallace, D.B.; Mather, A.; Kara, P.D.; Naicker, L.; Mokoena, N.; Pretorius, A.; Nefefe, T.; Thema, N.; Babiuk, S. Protection of Cattle Elicited Using a Bivalent Lumpy Skin Disease Virus-Vectored Recombinant Rift Valley Fever Vaccine. Front. Vet. Sci. 2020, 7, 256. [Google Scholar] [CrossRef] [PubMed]
- Aspden, K.; A van Dijk, A.; Bingham, J.; Cox, D.; Passmore, J.-A.; Williamson, A.-L. Immunogenicity of a recombinant lumpy skin disease virus (neethling vaccine strain) expressing the rabies virus glycoprotein in cattle. Vaccine 2002, 20, 2693–2701. [Google Scholar] [CrossRef] [PubMed]
- Wallace, D.B.; Viljoen, G.J. Immune responses to recombinants of the South African vaccine strain of lumpy skin disease virus generated by using thymidine kinase gene insertion. Vaccine 2005, 23, 3061–3067. [Google Scholar] [CrossRef] [PubMed]
- Cêtre-Sossah, C.; Dickmu, S.; Kwiatek, O.; Albina, E. A G-protein-coupled chemokine receptor: A putative insertion site for a multi-pathogen recombinant capripoxvirus vaccine strategy. J. Immunol. Methods 2017, 448, 112–115. [Google Scholar] [CrossRef] [Green Version]
- Douglass, N.; Omar, R.; Munyanduki, H.; Suzuki, A.; Mutowembwa, P.; Kara, P.; Heath, L.; Williamson, A.-L. The Development of Dual Vaccines against Lumpy Skin Disease (LSD) and Bovine Ephemeral Fever (BEF). Vaccines 2021, 9, 1512. [Google Scholar] [CrossRef]
- Chapman, R.; van Diepen, M.; Douglass, N.; Galant, S.; Jaffer, M.; Margolin, E.; Ximba, P.; Hermanus, T.; Moore, P.L.; Williamson, A.-L. Assessment of an LSDV-Vectored Vaccine for Heterologous Prime-Boost Immunizations against HIV. Vaccines 2021, 9, 1281. [Google Scholar] [CrossRef]
- Shen, Y. An Investigation into the Use of Lumpy Skin Disease Virus as a Vaccine Vector for a Potential HIV-1 Vaccine; University of Cape Town: Cape Town, South Africa, 2010. [Google Scholar]
- Alharbi, N.K. Poxviral promoters for improving the immunogenicity of MVA delivered vaccines. Hum. Vaccines Immunother. 2018, 15, 203–209. [Google Scholar] [CrossRef] [Green Version]
- Ink, B.S.; Pickup, D.J. Transcription of a poxvirus early gene is regulated both by a short promoter element and by a transcriptional termination signal controlling transcriptional interference. J. Virol. 1989, 63, 4632–4644. [Google Scholar] [CrossRef] [Green Version]
- Wyatt, L.S.; Shors, S.T.; Murphy, B.R.; Moss, B. Development of a replication-deficient recombinant vaccinia virus vaccine effective against parainfluenza virus 3 infection in an animal model. Vaccine 1996, 14, 1451–1458. [Google Scholar] [CrossRef]
- Di Pilato, M.; Mejías-Pérez, E.; Gomez, C.E.; Perdiguero, B.; Sorzano, C.O.S.; Esteban, M. New vaccinia virus promoter as a potential candidate for future vaccines. J. Gen. Virol. 2013, 94, 2771–2776. [Google Scholar] [CrossRef] [Green Version]
- Cochran, M.A.; Puckett, C.; Moss, B. In vitro mutagenesis of the promoter region for a vaccinia virus gene: Evidence for tandem early and late regulatory signals. J. Virol. 1985, 54, 30–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertholet, C.; Drillien, R.; Wittek, R. One hundred base pairs of 5’ flanking sequence of a vaccinia virus late gene are sufficient to temporally regulate late transcription. Proc. Natl. Acad. Sci. USA 1985, 82, 2096–2100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chakrabarti, S.; Sisler, J.R.; Moss, B. Compact, Synthetic, Vaccinia Virus Early/Late Promoter for Protein Expression. Biotechniques 1997, 23, 1094–1097. [Google Scholar] [CrossRef]
- Antoshkina, I.V.; Glazkova, D.V.; Urusov, F.A.; Bogoslovskaya, E.V.; Shipulin, G.A. Comparison of Recombinant MVA Selection Methods Based on F13L, D4R and K1L Genes. Viruses 2022, 14, 528. [Google Scholar] [CrossRef]
- Boshra, H.; Truong, T.; Nfon, C.; Gerdts, V.; Tikoo, S.; Babiuk, L.A.; Kara, P.; Mather, A.; Wallace, D.; Babiuk, S. Capripoxvirus-vectored vaccines against livestock diseases in Africa. Antivir. Res. 2013, 98, 217–227. [Google Scholar] [CrossRef] [PubMed]
- Romero, C.H.; Barrett, T.; Chamberlain, R.W.; Kitching, R.P.; Fleming, M.; Black, D.N. Recombinant Capripoxvirus Expressing the Hemagglutinin Protein Gene of Rinderpest Virus: Protection of Cattle against Rinderpest and Lumpy Skin Disease Viruses. Virology 1994, 204, 425–429. [Google Scholar] [CrossRef]
- Soi, R.K.; Rurangirwa, F.R.; McGuire, T.C.; Rwambo, P.M.; DeMartini, J.C.; Crawford, T.B. Protection of Sheep against Rift Valley Fever Virus and Sheep Poxvirus with a Recombinant Capripoxvirus Vaccine. Clin. Vaccine Immunol. 2010, 17, 1842–1849. [Google Scholar] [CrossRef] [Green Version]
- Caufour, P.; Rufael, T.; Lamien, C.E.; Lancelot, R.; Kidane, M.; Awel, D.; Sertse, T.; Kwiatek, O.; Libeau, G.; Sahle, M.; et al. Protective efficacy of a single immunization with capripoxvirus-vectored recombinant peste des petits ruminants vaccines in presence of pre-existing immunity. Vaccine 2014, 32, 3772–3779. [Google Scholar] [CrossRef]
- Wade-Evans, A.M.; Romero, C.H.; Mellor, P.; Takamatsu, H.; Anderson, J.; Thevasagayam, J.; Fleming, M.J.; Mertens, P.P.; Black, D.N. Expression of the major core structural protein (VP7) of bluetongue virus, by a recombinant capripox virus, provides partial protection of sheep against a virulent heterotypic bluetongue virus challenge. Virology 1996, 220, 227–231. [Google Scholar] [CrossRef] [Green Version]
- Boshra, H.; Truong, T.; Nfon, C.; Bowden, T.R.; Gerdts, V.; Tikoo, S.; Babiuk, L.A.; Kara, P.; Mather, A.; Wallace, D.B.; et al. A lumpy skin disease virus deficient of an IL-10 gene homologue provides protective immunity against virulent capripoxvirus challenge in sheep and goats. Antiviral Res. 2015, 123, 39–49. [Google Scholar] [CrossRef]
- Weyer, J.; Kuzmin, I.V.; Rupprecht, C.E.; Nel, L.H. Cross-protective and cross-reactive immune responses to recombinant vaccinia viruses expressing full-length lyssavirus glycoprotein genes. Epidemiol. Infect. 2007, 136, 670–678. [Google Scholar] [CrossRef] [PubMed]
- Ngichabe, C.K.; Wamwayi, H.M.; Ndungu, E.K.; Mirangi, P.K.; Bostock, C.J.; Black, D.N.; Barrett, T. Long term immunity in African cattle vaccinated with a recombinant capripox-rinderpest virus vaccine. Epidemiol. Infect. 2002, 128, 343–349. [Google Scholar] [CrossRef] [Green Version]
- Burgers, W.A.; Ginbot, Z.; Shen, Y.-J.; Chege, G.K.; Soares, A.P.; Müller, T.L.; Bunjun, R.; Kiravu, A.; Munyanduki, H.; Douglass, N.; et al. The novel capripoxvirus vector lumpy skin disease virus efficiently boosts modified vaccinia Ankara human immunodeficiency virus responses in rhesus macaques. J. Gen. Virol. 2014, 95, 2267–2272. [Google Scholar] [CrossRef] [PubMed]
- Cohen, A.D.C.; Van Dijk, A.A.; Korber, A.K.R.; Dumbell; Williamson, A.-L. Lumpy Skin Disease Virus as a Recombinant Vaccine Vector for Rift Valley Fever Virus and Bovine Ephemeral Fever Virus. In Proceedings of the European Society for Veterinary Virology. 4th International Congress on Veterinary Virology. "Virus survival and Vaccination", Moredun Research Institute, Edinburgh, Scotland, 24–27 August 1997; pp. 230–231. [Google Scholar]
- Wallace, D.B.; Ellis, C.E.; Espach, A.; Smith, S.J.; Greyling, R.R.; Viljoen, G.J. Protective immune responses induced by different recombinant vaccine regimes to Rift Valley fever. Vaccine 2006, 24, 7181–7189. [Google Scholar] [CrossRef] [PubMed]
- Offerman, K.; Deffur, A.; Carulei, O.; Wilkinson, R.; Douglass, N.; Williamson, A.-L. Six host-range restricted poxviruses from three genera induce distinct gene expression profiles in an in vivo mouse model. BMC Genom. 2015, 16, 510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 2.
Bonsmara cow with lumpy skin disease—photograph provided by P. B. Kloppers.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Whittle, L.; Chapman, R.; Williamson, A.-L. Lumpy Skin Disease—An Emerging Cattle Disease in Europe and Asia. Vaccines 2023, 11, 578. https://doi.org/10.3390/vaccines11030578
AMA Style
Whittle L, Chapman R, Williamson A-L. Lumpy Skin Disease—An Emerging Cattle Disease in Europe and Asia. Vaccines. 2023; 11(3):578. https://doi.org/10.3390/vaccines11030578
Chicago/Turabian StyleWhittle, Leah, Rosamund Chapman, and Anna-Lise Williamson. 2023. "Lumpy Skin Disease—An Emerging Cattle Disease in Europe and Asia" Vaccines 11, no. 3: 578. https://doi.org/10.3390/vaccines11030578
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.