
Specific Binding and Endocytosis of Liposomes to HEK293T Cells via Myrisoylated Pre-S1 Peptide Bound to Sodium Taurocholate Cotransporting Polypeptide


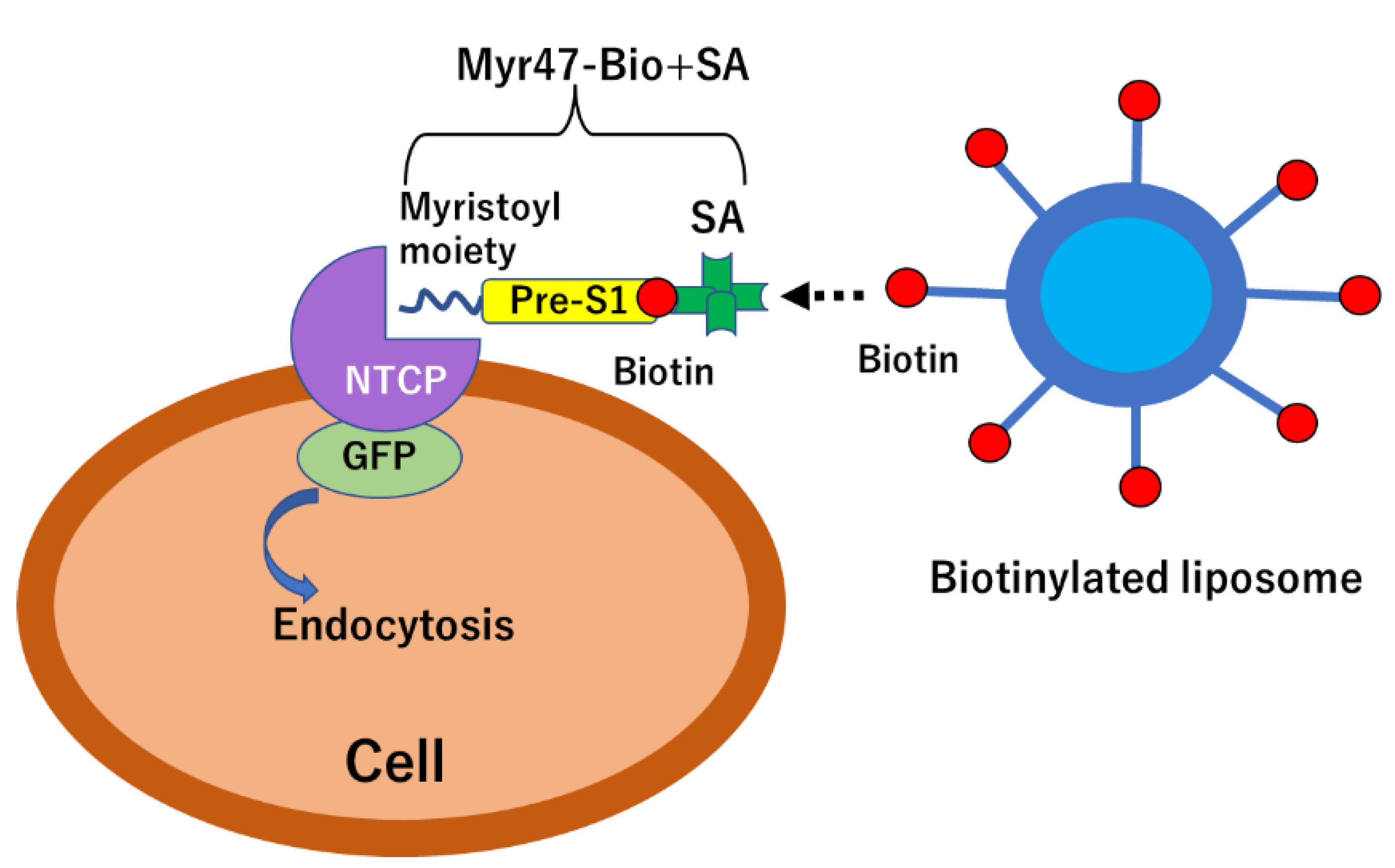{kind=link}
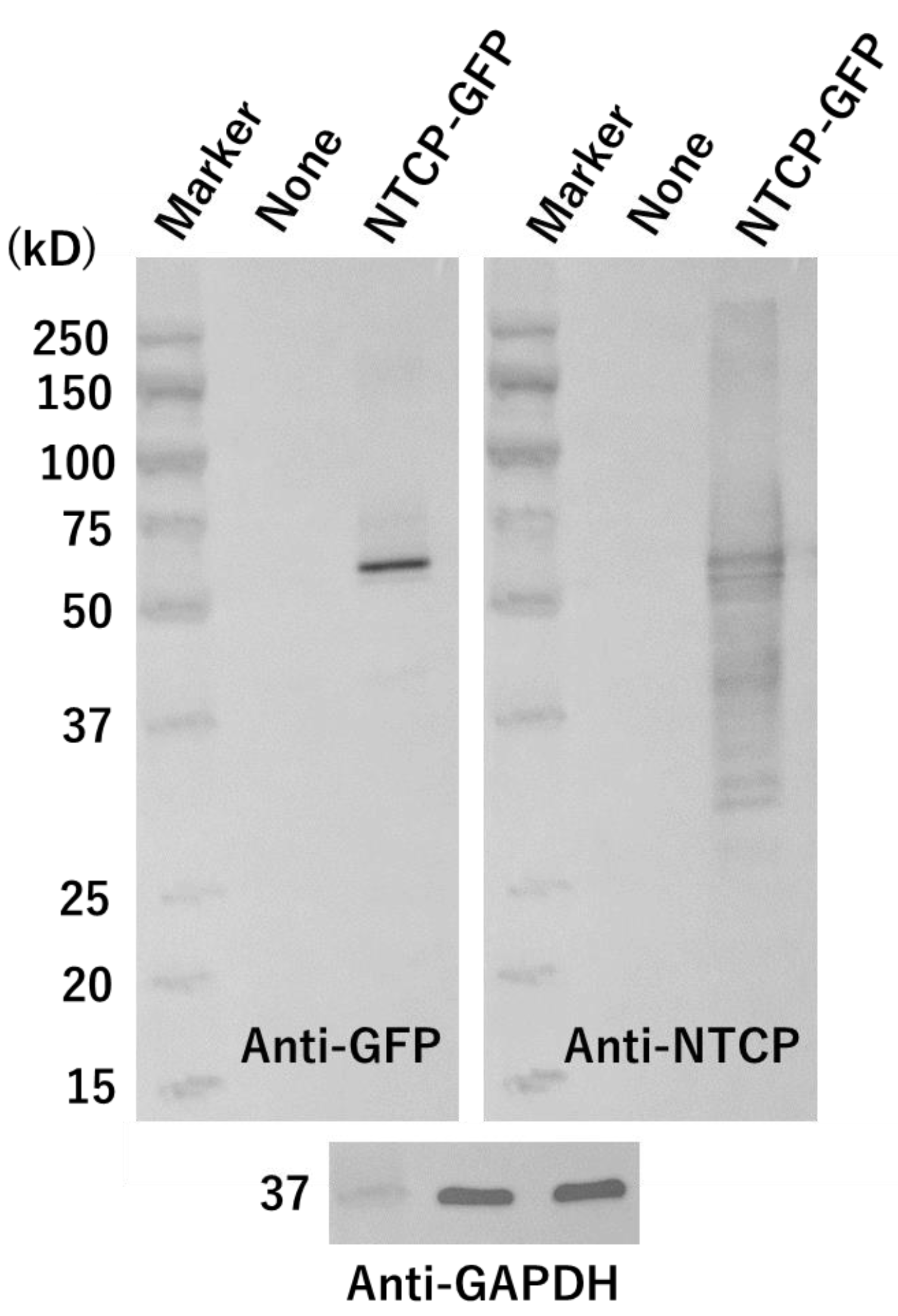{kind=link}
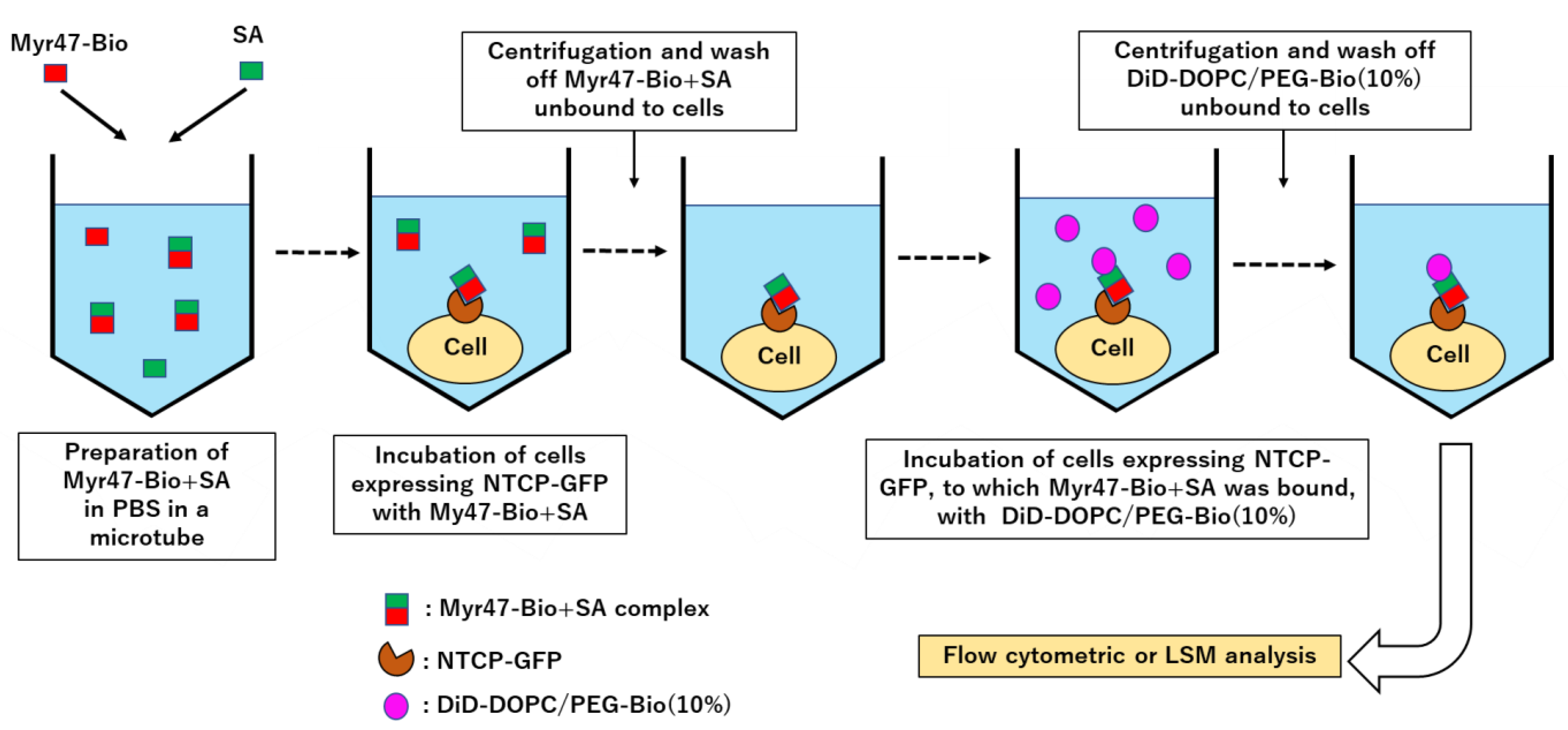{kind=link}
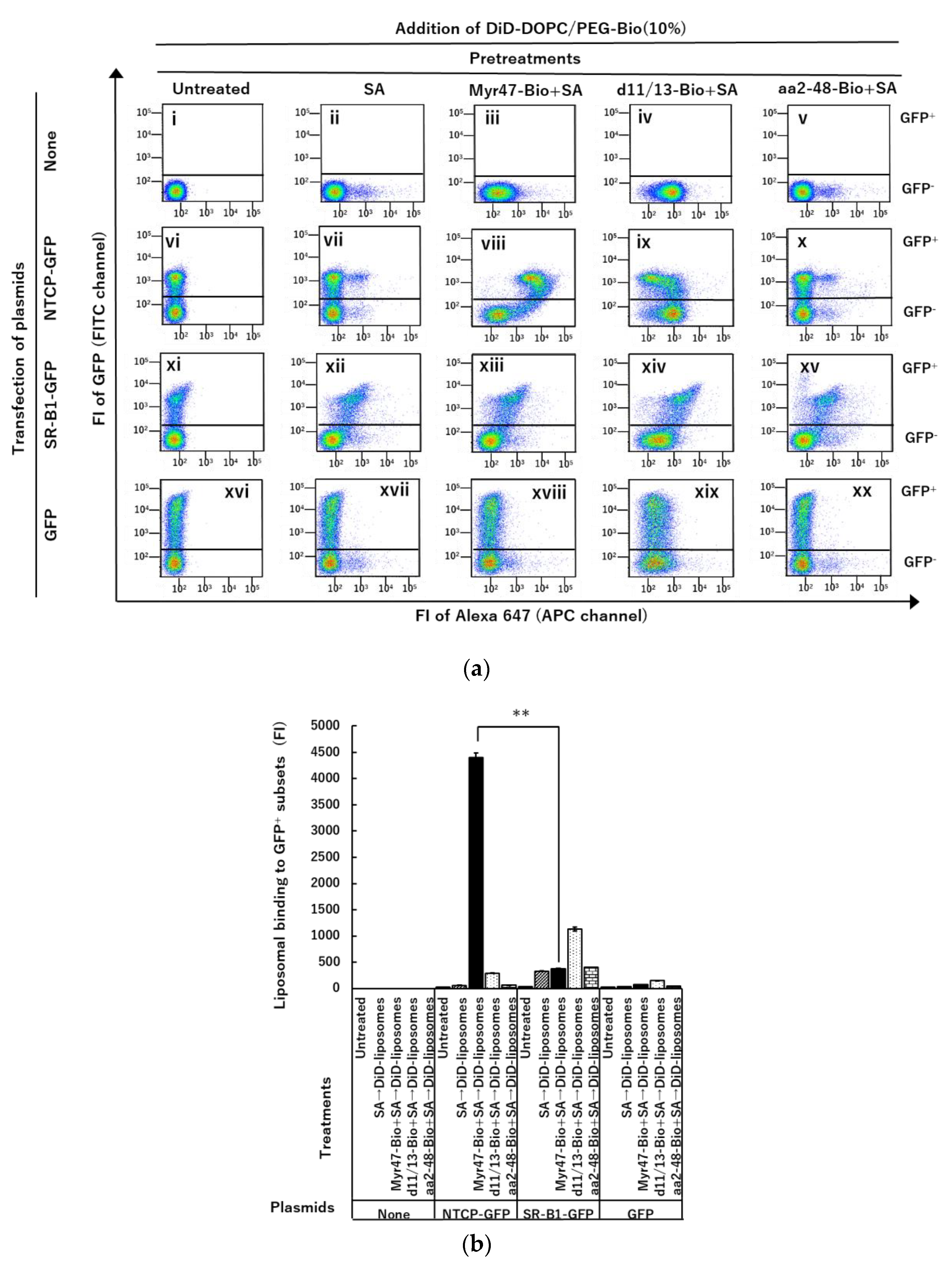{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of Liposomes
2.2. Biotinylated Pre-S1 Peptides and SA
2.3. Cell Culture
2.4. Transient Expression of NTCP-GFP and SR-B1-GFP
2.5. Western Blot Analysis
2.6. Liposomal Binding Assays Using Flow Cytometry
2.7. Laser Scanning Microscopy (LSM)
2.8. Quantification of Cells Endocytosing DiD-Liposomes Using Flow Cytometry and Trypan Blue
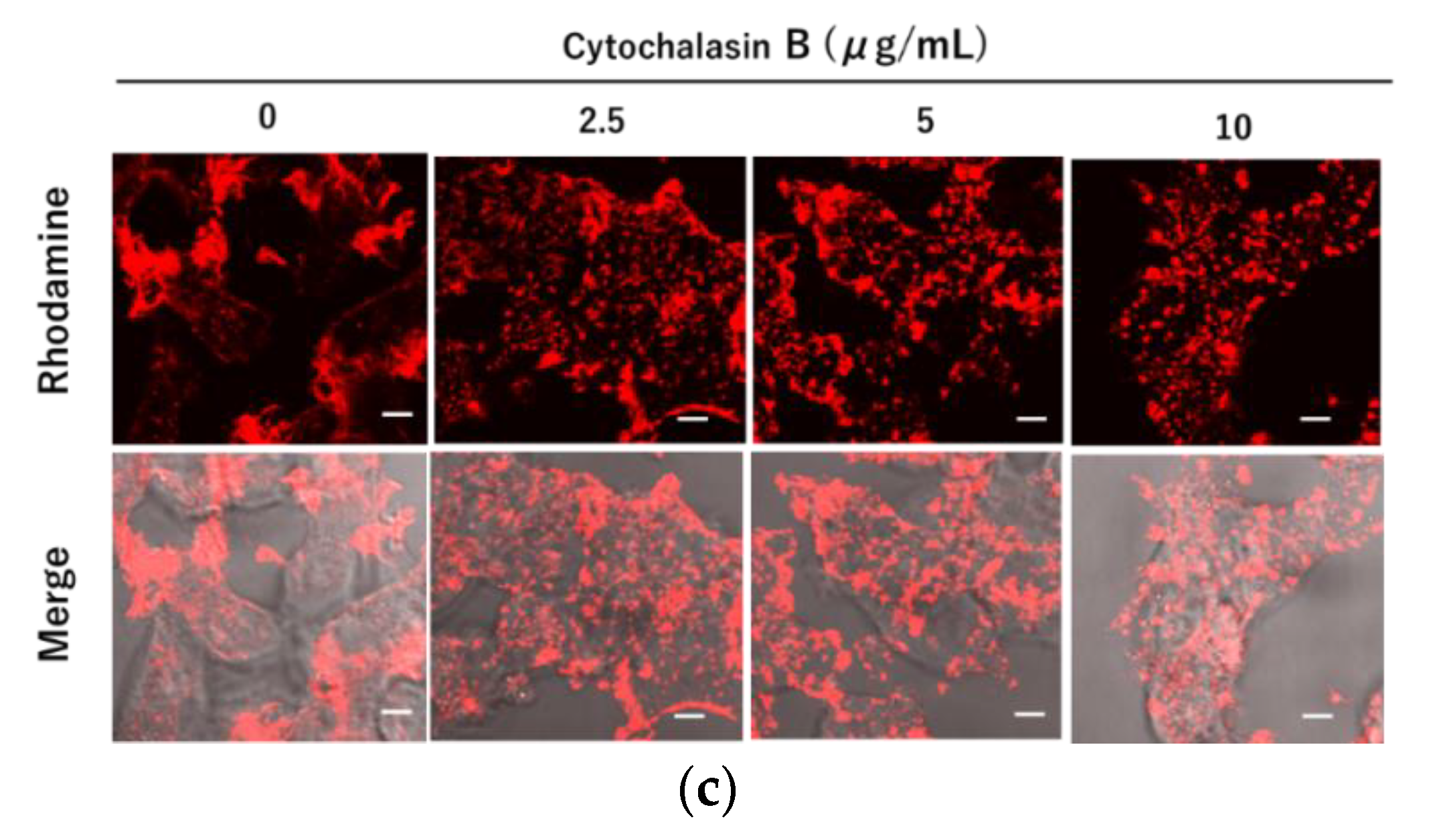
2.9. Actin Staining in HEK293T Cells Using Rhodamine Phalloidin
3. Results
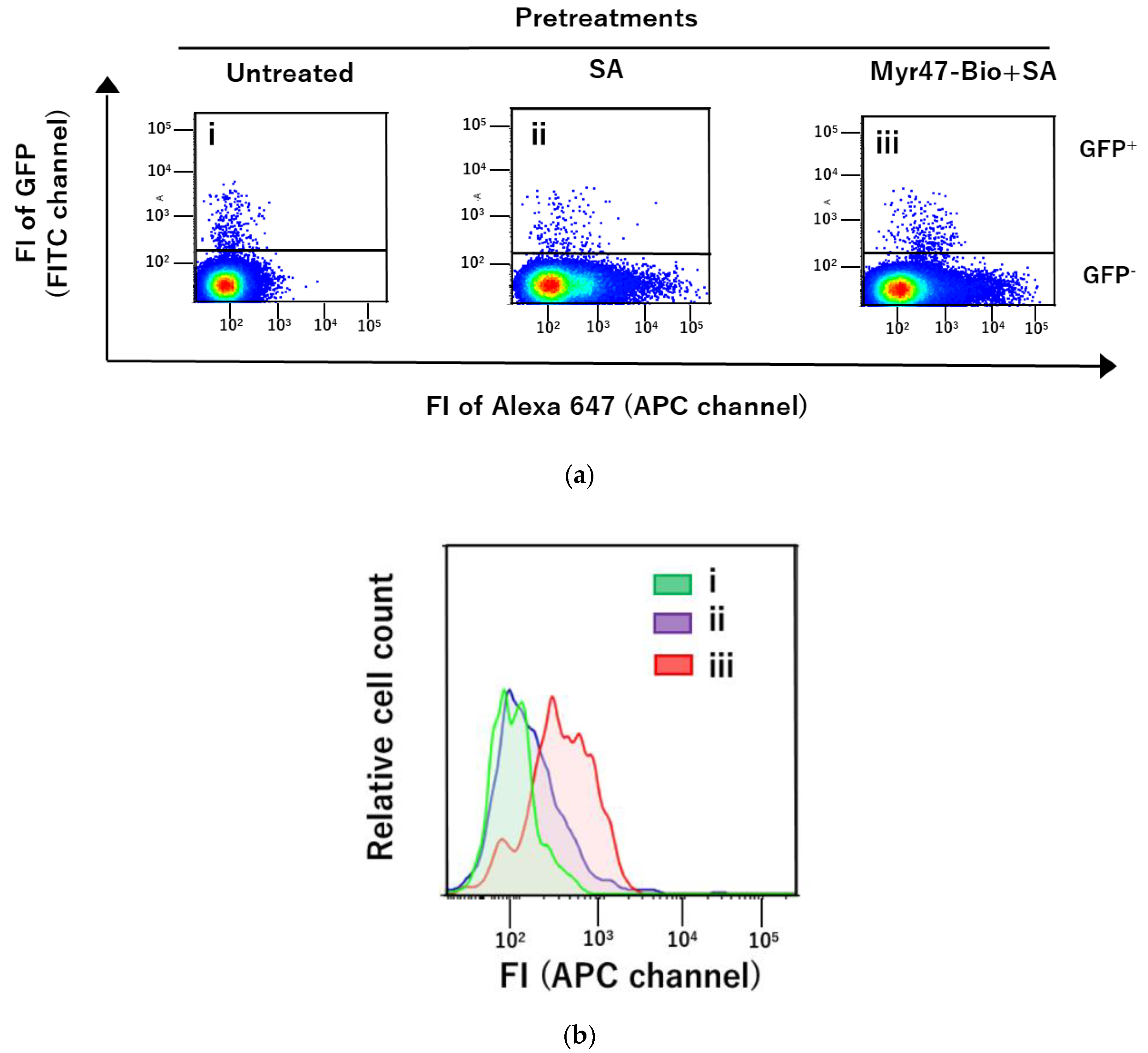
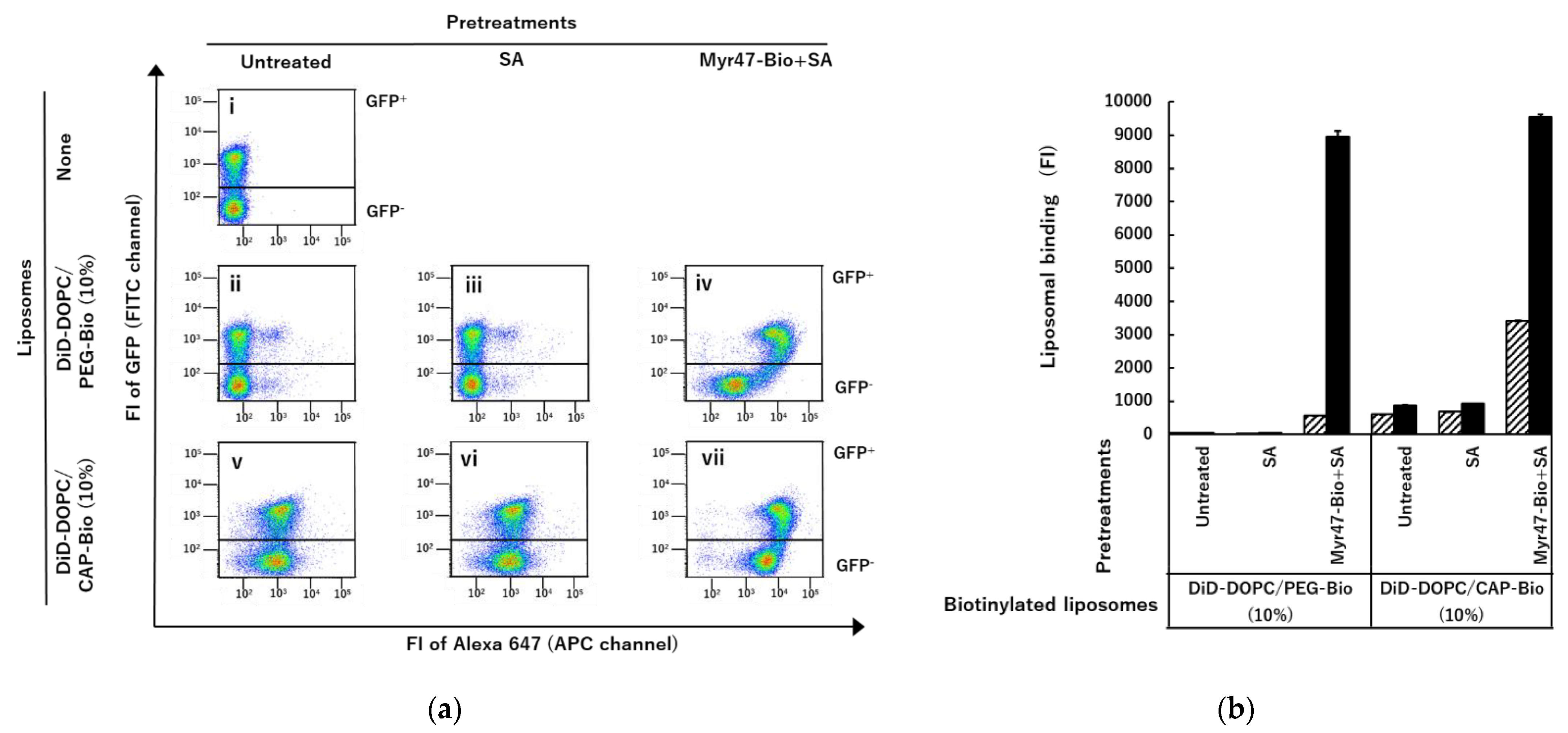
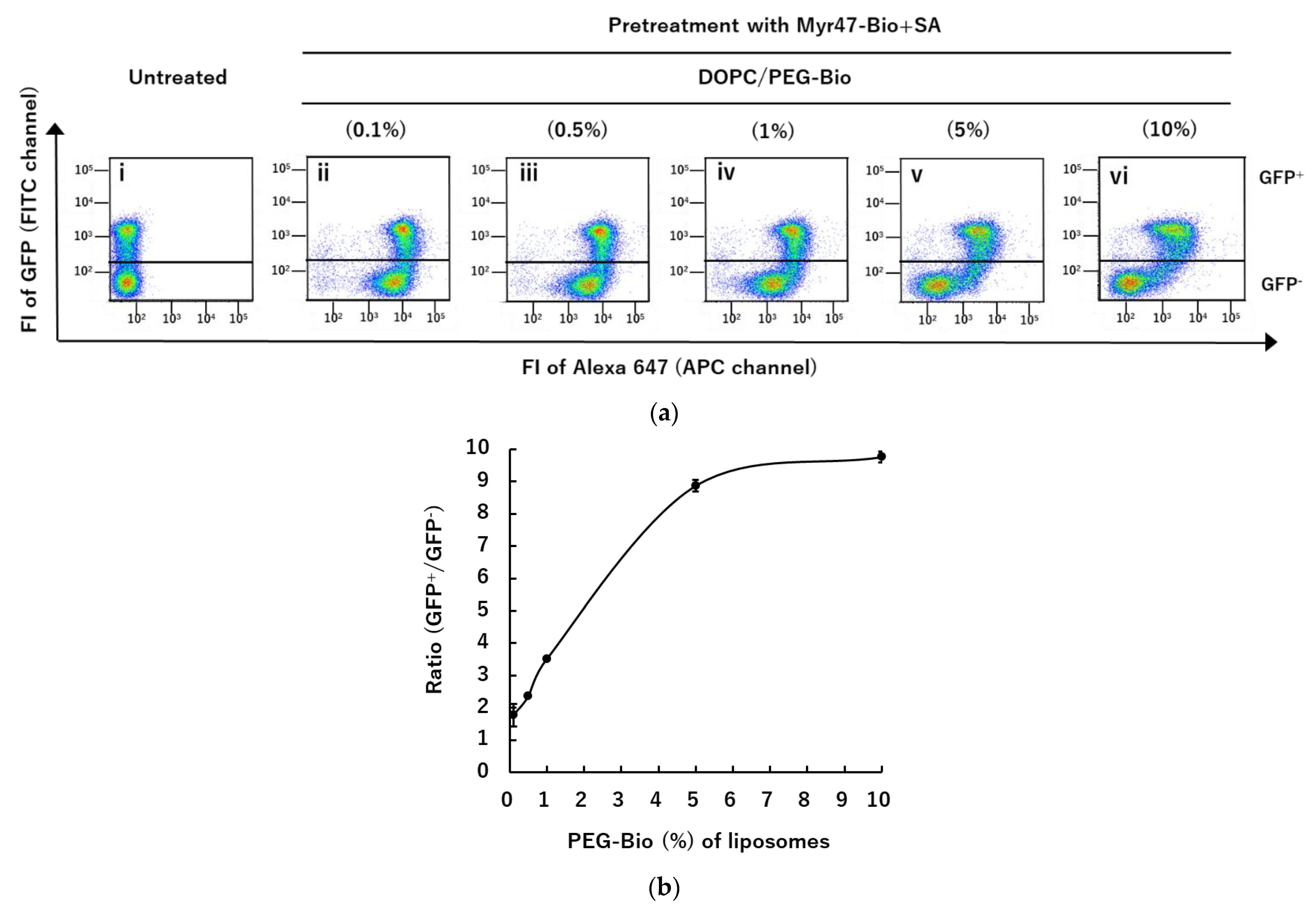
3.1. Specific Binding of DiD-DOPC/PEG-Bio (10%) to NTCP-GFP via Myr47-Bio+SA
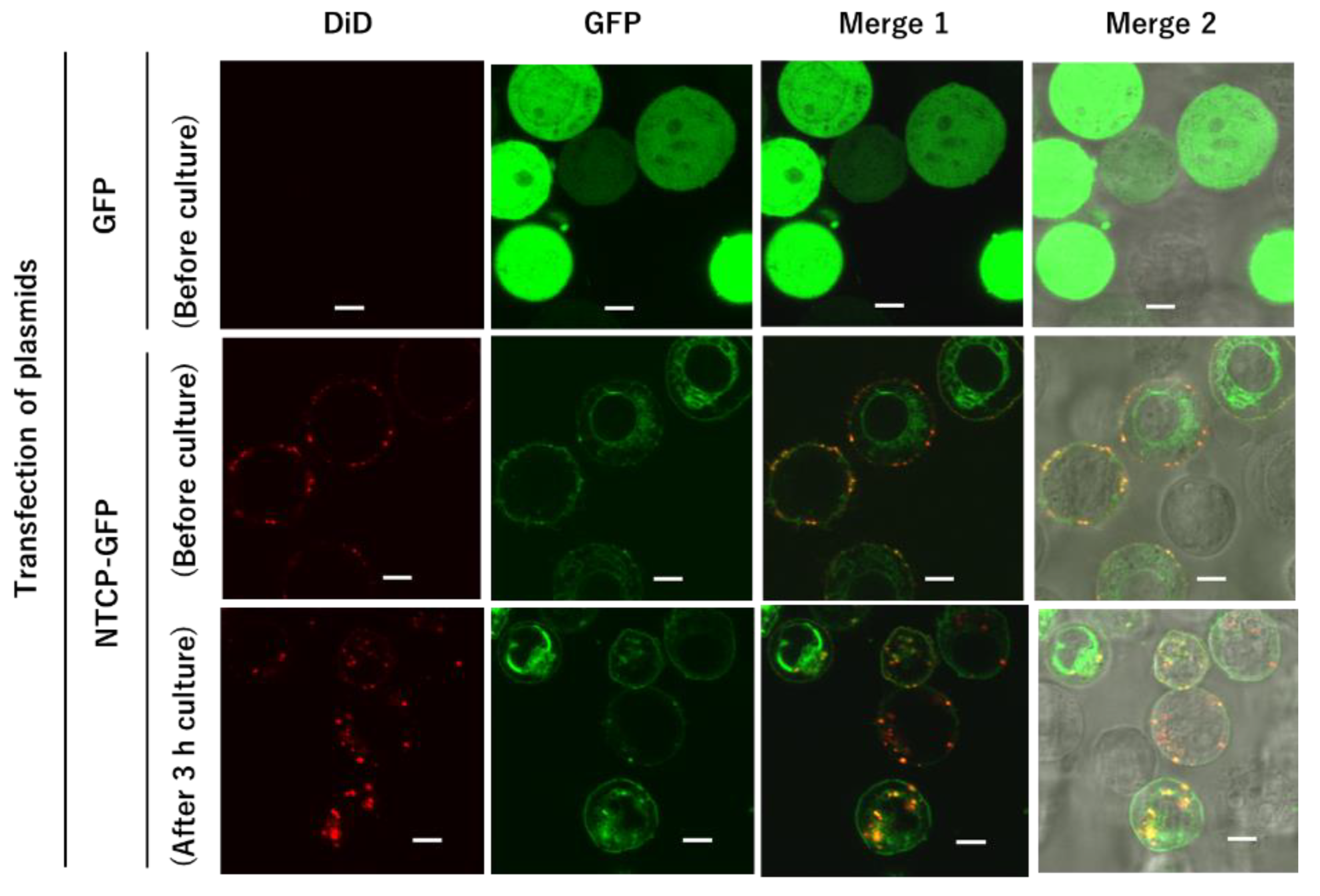
3.2. Liposomal Distribution in HEK293T Cells Expressing NTCP-GFP after Treatment with Myr47-Bio+SA Followed by DiD-DOPC/PEG-Bio (10%)
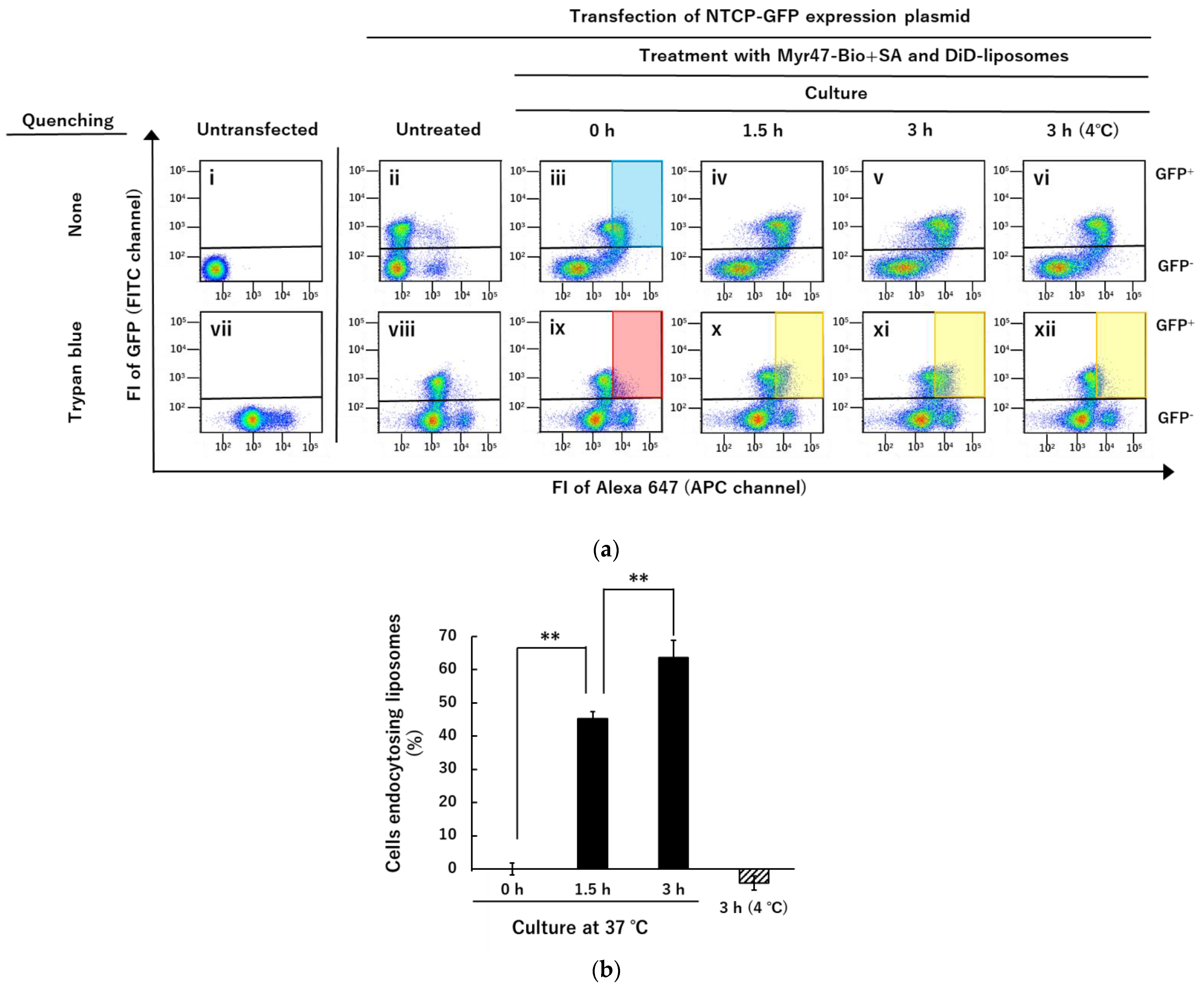
3.3. Quenching of DiD-Liposomes Bound on Cell Surface by Trypan Blue and Detection of Cells Endocytosing DiD-Liposomes in Flow Cytometry
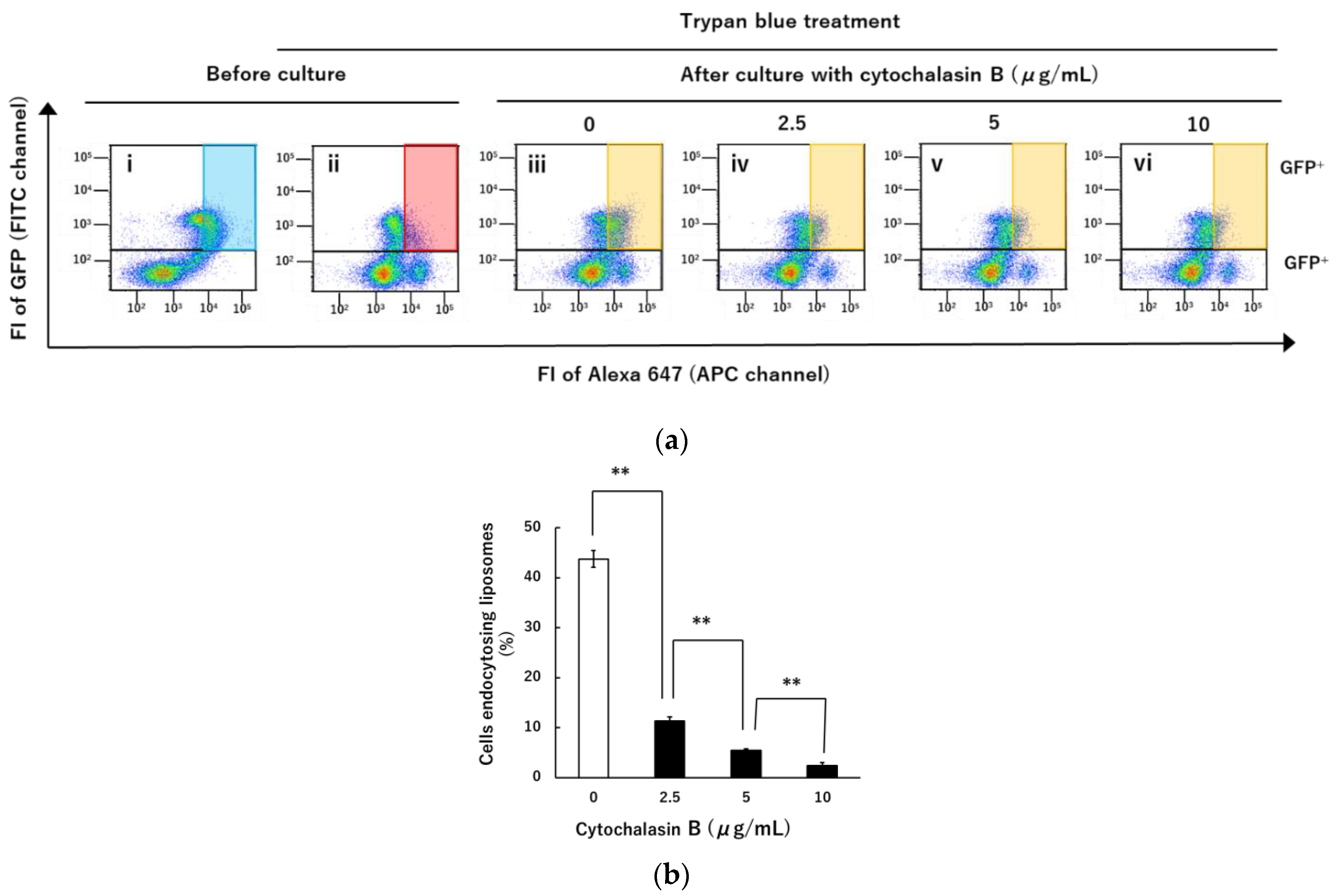
3.4. Inhibition of Liposomal Endocytosis via NTCP and Myr47 by Cytochalasin B
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Yardeni, D.; Ghany, M.G. Review article: Hepatitis B-current and emerging therapies. Aliment Pharm. Ther. 2022, 55, 805–819. [Google Scholar] [CrossRef]
- Herrscher, C.; Roingeard, P.; Blanchard, E. Hepatitis B Virus Entry into Cells. Cells 2020, 9, 1486. [Google Scholar] [CrossRef] [PubMed]
- Tsukuda, S.; Watashi, K. Hepatitis B virus biology and life cycle. Antivir. Res. 2020, 182, 104925. [Google Scholar] [CrossRef] [PubMed]
- Rehman, Z.; Fahim, A.; Sadia, H. Deciphering the mystery of hepatitis B virus receptors: A historical perspective. Virusdisease 2015, 26, 97–104. [Google Scholar] [CrossRef] [Green Version]
- Barrera, A.; Guerra, B.; Notvall, L.; Lanford, R.E. Mapping of the hepatitis B virus pre-S1 domain involved in receptor recognition. J. Virol. 2005, 79, 9786–9798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glebe, D.; Urban, S.; Knoop, E.V.; Cag, N.; Krass, P.; Grün, S.; Bulavaite, A.; Sasnauskas, K.; Gerlich, W.H. Mapping of the hepatitis B virus attachment site by use of infection-inhibiting preS1 lipopeptides and Tupaia hepatocytes. Gastroenterology 2005, 129, 234–245. [Google Scholar] [CrossRef]
- Gripon, P.; Cannie, I.; Urban, S. Efficient inhibition of hepatitis B virus infection by acylated peptides derived from the large viral surface protein. J. Virol. 2005, 79, 1613–1622. [Google Scholar] [CrossRef] [Green Version]
- Schulze, A.; Schieck, A.; Ni, Y.; Mier, W.; Urban, S. Fine mapping of pre-S sequence requirements for hepatitis B virus large envelope protein-mediated receptor interaction. J. Virol. 2010, 84, 1989–2000. [Google Scholar] [CrossRef] [Green Version]
- Yan, H.; Zhong, G.; Xu, G.; He, W.; Jing, Z.; Gao, Z.; Huang, Y.; Qi, Y.; Peng, B.; Wang, H.; et al. Sodium taurocholate cotransporting polypeptide is a functional receptor for human hepatitis B and D virus. Elife 2012, 1, e00049. [Google Scholar] [CrossRef]
- Kang, C.; Syed, Y.Y. Bulevirtide: First approval. Drugs 2020, 80, 1601–1605. [Google Scholar] [CrossRef]
- Koh, C.; Da, B.L.; Glenn, J.S. HBV/HDV coinfection: A challenge for therapeutics. Clin. Liver Dis. 2019, 23, 557–572. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Somiya, M.; Iijima, M.; Tatematsu, K.; Kuroda, S. A hepatitis B virus-derived human hepatic cell-specific heparin-binding peptide: Identification and application to a drug delivery system. Biomater. Sci. 2018, 7, 322–335. [Google Scholar] [CrossRef] [PubMed]
- Somiya, M.; Liu, Q.; Yoshimoto, N.; Iijima, M.; Tatematsu, K.; Nakai, T.; Okajima, T.; Kuroki, K.; Ueda, K.; Kuroda, S. Cellular uptake of hepatitis B virus envelope L particles is independent of sodium taurocholate cotransporting polypeptide, but dependent on heparan sulfate proteoglycan. Virology 2016, 497, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Fujita, K.; Somiya, M.; Kuroda, S.; Hinuma, S. Induction of lipid droplets in non-macrophage cells as well as macrophages by liposomes and exosomes. Biochem. Biophys. Res. Commun. 2019, 510, 184–190. [Google Scholar] [CrossRef]
- Fujita, K.; Koide, N.; Somiya, M.; Kuroda, S.; Hinuma, S. A regulatory role of scavenger receptor class B type 1 in endocytosis and lipid droplet formation induced by liposomes containing phosphatidylethanolamine in HEK293T cells. Biochim. Biophys. Acta Mol. Cell. Res. 2021, 1868, 118859. [Google Scholar] [CrossRef]
- Koide, N.; Fujita, K.; Kuroda, S.; Hinuma, S. Binding of liposomes composed of phosphatidylcholine to scavenger receptor class B type 1 and its modulation by phosphatidic acid in HEK293T cells. Biochim. Biophys. Acta-Mol. Cell Res. 2021, 1868, 119043. [Google Scholar] [CrossRef]
- Shen, W.J.; Asthana, S.; Kraemer, F.B.; Azhar, S. Scavenger receptor B type 1: Expression, molecular regulation, and cholesteroltransport function. J. Lipid Res. 2018, 59, 1114–1131. [Google Scholar] [CrossRef] [Green Version]
- Hinuma, S.; Fujita, K.; Kuroda, S. Binding of Nanoparticles Harboring Recombinant Large Surface Protein of Hepatitis B Virus to Scavenger Receptor Class B Type 1. Viruses 2021, 13, 1334. [Google Scholar] [CrossRef]
- Hinuma, S.; Kuroda, S. Binding of Hepatitis B Virus Pre-S1 Domain-Derived Synthetic Myristoylated Peptide to Scavenger Receptor Class B Type 1 with Differential Properties from Sodium Taurocholate Cotransporting Polypeptide. Viruses 2022, 14, 105. [Google Scholar] [CrossRef]
- Iwamoto, M.; Watashi, K.; Tsukuda, S.; Aly, H.H.; Fukasawa, M.; Fujimoto, A.; Suzuki, R.; Aizaki, H.; Ito, T.; Koiwai, O.; et al. Evaluation and identification of hepatitis B virus entry inhibitors using HepG2 cells overexpressing a membrane transporter NTCP. Biochem. Biophys. Res. Commun. 2014, 443, 808–813. [Google Scholar] [CrossRef]
- Sahlin, S.; Hed, J.; Runfquist, I. Differentiation between attached and ingested immune complexes by a fluorescence quenching cytofluorometric assay. J. Immunol. Methods 1983, 60, 115–124. [Google Scholar] [CrossRef] [PubMed]
- Loike, J.D.; Silverstein, S.C. A fluorescence quenching technique using trypan blue to differentiate between attached and ingested glutaraldehyde-fixed red blood cells in phagocytosing murine macrophages. J. Immunol. Methods 1983, 57, 373–379. [Google Scholar] [CrossRef] [PubMed]
- Avelar-Freitas, B.A.; Almeida, V.G.; Pinto, M.C.; Mourão, F.A.; Massensini, A.R.; Martins-Filho, O.A.; Rocha-Vieira, E.; Brito-Melo, G.E. Trypan blue exclusion assay by flow cytometry. Braz. J. Med. Biol. Res. 2014, 47, 307–315. [Google Scholar] [CrossRef] [PubMed]
- Sato, K.; Nagai, J.; Mitsui, N.; Yumoto, R.; Takano, M. Effects of endocytosis inhibitors on internalization of human IgG by Caco-2 human intestinal epithelial cells. Life Sci. 2009, 85, 800–807. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Zhang, Q.; Peng, Q.; Zhou, J.; Liao, L.; Sun, X.; Zhang, L.; Gong, T. Hepatitis B virus preS1-derived lipopeptide functionalized liposomes for targeting of hepatic cells. Biomaterials 2014, 35, 6130–6141. [Google Scholar] [CrossRef] [PubMed]
- Weinstock, J.V.; Kassab, J.T. Angiotensin II stimulation of granuloma macrophage phagocytosis and actin polymerization in murine schistosomiasis mansoni. Cell. Immunol. 1984, 89, 46–54. [Google Scholar] [CrossRef]











Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hinuma, S.; Fujita, K.; Kuroda, S. Specific Binding and Endocytosis of Liposomes to HEK293T Cells via Myrisoylated Pre-S1 Peptide Bound to Sodium Taurocholate Cotransporting Polypeptide. Vaccines 2022, 10, 2050. https://doi.org/10.3390/vaccines10122050
Hinuma S, Fujita K, Kuroda S. Specific Binding and Endocytosis of Liposomes to HEK293T Cells via Myrisoylated Pre-S1 Peptide Bound to Sodium Taurocholate Cotransporting Polypeptide. Vaccines. 2022; 10(12):2050. https://doi.org/10.3390/vaccines10122050
Chicago/Turabian StyleHinuma, Shuji, Kazuyo Fujita, and Shun’ichi Kuroda. 2022. "Specific Binding and Endocytosis of Liposomes to HEK293T Cells via Myrisoylated Pre-S1 Peptide Bound to Sodium Taurocholate Cotransporting Polypeptide" Vaccines 10, no. 12: 2050. https://doi.org/10.3390/vaccines10122050